PENDAHULUAN
Kelapa sawit (Elaeis guineensis) merupakan salah satu komoditas perkebunan andalan bagi perekonomian Indonesia. Kualitas dan tingkat produksi Crude Palm Oil
(CPO) yang tinggi merupakan keunggulan dari komiditas ini. Data Ditjen Perkebunan menyebutkan pada tahun 2005 luas perkebunan kelapa sawit di Indonesia mencapai 5.358.423 hektar dengan total produksi minyak hingga 10.683.756 ton. Dalam proses produksi CPO dihasilkan banyak limbah yang masih belum dimanfaatkan secara maksimal sehungga berpotensi mencemari lingkungan. Tandan Kosong Kelapa Sawit (TKKS) merupakan limbah padat terbesar dari pabrik minyak CPO. Komponen utama TKKS adalah lignin, selulosa, dan hemiselulosa yang umum disebut lignoselulosa (Darnoko 1992).
TKKS bukan satu-satunya limbah yang tergolong lignoselulosa. Bagas tebu (ampas tebu) juga mengandung serat lignoselulosa yang sukar terdegradasi. Kandungan lignin yang tinggi pada TKKS dan bagas tebu menyebabkan kendala dalam pemanfaatan selulosa murni bagi industri yang berbahan baku selulosa seperti industri pulp dan kertas ataupun industri pembuatan papan serat. Salah satu cara untuk mengatasi masalah ini menggunakan mikroorganisme dalam rangka biodelignifikasi. Mikroorganisme yang berperan penting dalam biodelignifikasi adalah Fungi Pelapuk Putih (FPP) dari kelas
Basidiomycetes. Kemampuan FPP melakukan biodelignifikasi disebabkan oleh enzim ligninolitik ekstraselular yang dihasilkannya (Kirk & Chang 1990). Enzim ligninolitik ekstraselular tersebut adalah lakase, lignin peroksidase (LiP), dan mangan peroksidase (MnP) (Gold & Alic 1993). Produksi enzim ligninolitik dari limbah lignoselulosa mempunyai arti penting karena selain dapat mengatasi masalah limbah organik lignoselulosa, juga dapat memberi nilai tambah dari limbah tersebut.
Palmieri et al (1999) disebutkan bahwa Cu (tembaga) merupakan induser yang efisien dalam menaikkan aktivitas lakase dari
P.ostreatus. Dedak dapat digunakan sebagai substrat alami dan dapat meningkatkan pertumbuhan miselium menjadi lebih tebal dan padat (Silverio et al 1981). Dedak telah banyak digunakan sebagai induser untuk pertumbuhan miselium dalam budidaya jamur tiram karena kandungan nitrogen, karbohidrat,
dan karbonnya serta vitamin B1 (tiamin) (Okwujiako 1990).
Penelitian bertujuan mengoptimumkan produksi enzim ligninolitik oleh Omphalina
sp. dan P.ostreatus pada TKKS dan bagas tebu dengan pemberian suplemen CuSO4,
dedak, dan kombinasi keduanya. Selain itu akan dilakukan karakterisasi pH optimum enzim ligninolitik yang dihasilkan. Hipotesis penelitian ini adalah produksi enzim ligninolitik oleh Omphalina sp. dan
P.ostreatus pada TKKS dan bagas tebu akan meningkat dengan pemberian suplemen CuSO4 dan dedak. Manfaat penelitian ini
adalah memberikan informasi tentang suplemen yang paling baik untuk optimasi produksi enzim ligninolitik oleh Omphalina
sp. dan P.ostreatus pada limbah organik lignoselulosa TKKS dan bagas tebu.
TINJAUAN PUSTAKA
Limbah Padat Organik LignoselulosaLignoselulosa merupakan hasil samping atau residu dari berbagai industri seperti industri pulp kertas, industri kayu, dan industri pertanian. Lignoselulosa sendiri merupakan bahan biopolimer dan komponen penyusun utama kayu dan tanaman bukan kayu yang berlimpah jumlahnya di alam. Lignoselulosa merupakan komponen yang sukar terdegradasi dibandingkan dengan jenis polisakarida lainnya (Reeves & Schmidt 1994).
Limbah padat organik lignoselulosa yang paling umum dijumpai adalah TKKS dari industri pengolahan kelapa sawit menjadi minyak sawit mentah (CPO). Tandan kosong adalah tandan yang keluar dari mesin penebah dan tidak mengandung buah. Komponen dalam TKKS adalah selulosa 32,55%, hemiselulosa 31,70%, dan lignin 28,54% (Santosa 1993). Selain itu TKKS juga mengandung lemak 5,35% dan protein 4,45% yang dihitung berdasarkan berat kering oven. Komponen lain yang terdapat dalam TKKS adalah ekstrak yang berupa minyak dan lemak (Fitriasari 2001 dalam Yuniawati 2006). Limbah lignoselulosa yang lain adalah bagas tebu yang merupakan residu penggilingan tebu. Bagas tebu mengandung SiO2 3,01%,
abu 3,82%, lignin 22,09%, selulosa 37,65%, pentosan 27,97%, dan nira 1,81% (Santosa 1993).
PENDAHULUAN
Kelapa sawit (Elaeis guineensis) merupakan salah satu komoditas perkebunan andalan bagi perekonomian Indonesia. Kualitas dan tingkat produksi Crude Palm Oil
(CPO) yang tinggi merupakan keunggulan dari komiditas ini. Data Ditjen Perkebunan menyebutkan pada tahun 2005 luas perkebunan kelapa sawit di Indonesia mencapai 5.358.423 hektar dengan total produksi minyak hingga 10.683.756 ton. Dalam proses produksi CPO dihasilkan banyak limbah yang masih belum dimanfaatkan secara maksimal sehungga berpotensi mencemari lingkungan. Tandan Kosong Kelapa Sawit (TKKS) merupakan limbah padat terbesar dari pabrik minyak CPO. Komponen utama TKKS adalah lignin, selulosa, dan hemiselulosa yang umum disebut lignoselulosa (Darnoko 1992).
TKKS bukan satu-satunya limbah yang tergolong lignoselulosa. Bagas tebu (ampas tebu) juga mengandung serat lignoselulosa yang sukar terdegradasi. Kandungan lignin yang tinggi pada TKKS dan bagas tebu menyebabkan kendala dalam pemanfaatan selulosa murni bagi industri yang berbahan baku selulosa seperti industri pulp dan kertas ataupun industri pembuatan papan serat. Salah satu cara untuk mengatasi masalah ini menggunakan mikroorganisme dalam rangka biodelignifikasi. Mikroorganisme yang berperan penting dalam biodelignifikasi adalah Fungi Pelapuk Putih (FPP) dari kelas
Basidiomycetes. Kemampuan FPP melakukan biodelignifikasi disebabkan oleh enzim ligninolitik ekstraselular yang dihasilkannya (Kirk & Chang 1990). Enzim ligninolitik ekstraselular tersebut adalah lakase, lignin peroksidase (LiP), dan mangan peroksidase (MnP) (Gold & Alic 1993). Produksi enzim ligninolitik dari limbah lignoselulosa mempunyai arti penting karena selain dapat mengatasi masalah limbah organik lignoselulosa, juga dapat memberi nilai tambah dari limbah tersebut.
Palmieri et al (1999) disebutkan bahwa Cu (tembaga) merupakan induser yang efisien dalam menaikkan aktivitas lakase dari
P.ostreatus. Dedak dapat digunakan sebagai substrat alami dan dapat meningkatkan pertumbuhan miselium menjadi lebih tebal dan padat (Silverio et al 1981). Dedak telah banyak digunakan sebagai induser untuk pertumbuhan miselium dalam budidaya jamur tiram karena kandungan nitrogen, karbohidrat,
dan karbonnya serta vitamin B1 (tiamin) (Okwujiako 1990).
Penelitian bertujuan mengoptimumkan produksi enzim ligninolitik oleh Omphalina
sp. dan P.ostreatus pada TKKS dan bagas tebu dengan pemberian suplemen CuSO4,
dedak, dan kombinasi keduanya. Selain itu akan dilakukan karakterisasi pH optimum enzim ligninolitik yang dihasilkan. Hipotesis penelitian ini adalah produksi enzim ligninolitik oleh Omphalina sp. dan
P.ostreatus pada TKKS dan bagas tebu akan meningkat dengan pemberian suplemen CuSO4 dan dedak. Manfaat penelitian ini
adalah memberikan informasi tentang suplemen yang paling baik untuk optimasi produksi enzim ligninolitik oleh Omphalina
sp. dan P.ostreatus pada limbah organik lignoselulosa TKKS dan bagas tebu.
TINJAUAN PUSTAKA
Limbah Padat Organik LignoselulosaLignoselulosa merupakan hasil samping atau residu dari berbagai industri seperti industri pulp kertas, industri kayu, dan industri pertanian. Lignoselulosa sendiri merupakan bahan biopolimer dan komponen penyusun utama kayu dan tanaman bukan kayu yang berlimpah jumlahnya di alam. Lignoselulosa merupakan komponen yang sukar terdegradasi dibandingkan dengan jenis polisakarida lainnya (Reeves & Schmidt 1994).
Limbah padat organik lignoselulosa yang paling umum dijumpai adalah TKKS dari industri pengolahan kelapa sawit menjadi minyak sawit mentah (CPO). Tandan kosong adalah tandan yang keluar dari mesin penebah dan tidak mengandung buah. Komponen dalam TKKS adalah selulosa 32,55%, hemiselulosa 31,70%, dan lignin 28,54% (Santosa 1993). Selain itu TKKS juga mengandung lemak 5,35% dan protein 4,45% yang dihitung berdasarkan berat kering oven. Komponen lain yang terdapat dalam TKKS adalah ekstrak yang berupa minyak dan lemak (Fitriasari 2001 dalam Yuniawati 2006). Limbah lignoselulosa yang lain adalah bagas tebu yang merupakan residu penggilingan tebu. Bagas tebu mengandung SiO2 3,01%,
abu 3,82%, lignin 22,09%, selulosa 37,65%, pentosan 27,97%, dan nira 1,81% (Santosa 1993).
adalah kandungan lignin yang sulit terdegradasi baik secara enzimatik maupun kimia (Ohkuma et al 2001). Komponen lignoselulosa lainnya, selulosa dan hemiselulosa, merupakan polisakarida yang dapat dihidrolisis menjadi monomernya yang dapat diubah kembali secara kimia menjadi etanol, butanol, aseton, dan gliserol (Wyman 2002).
Lignin
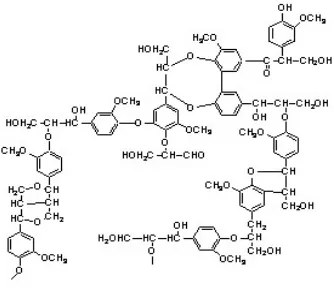
Lignin merupakan senyawa kimia yang umumnya diperoleh pada kayu dan merupakan bagian integral dari dinding sel tumbuhan. Lignin adalah bahan polimer alam terbanyak kedua setelah selulosa (Sarkanen & Ludwig 1971). Lignin merupakan polimer yang sukar larut dalam asam dan basa kuat dan sulit terdegradasi secara kimiawi maupun secara enzimatis. Lignin pada kayu terdapat pada lamela tengah antara selulosa, hemiselulosa, dan pektin yang berfungsi sebagai perekat atau penguat dinding sel. Lignin berperan sangat penting bagi tumbuhan sebagai sarana pengangkut air, nutrisi, dan metabolit dalam sel tumbuhan.
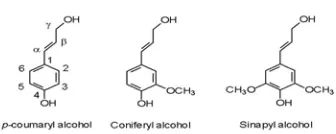
Biosintesis lignin dimulai dengan sintesis monolignol. Monolignol disintesis di sitosol dalam bentuk glukosida. Glukosida kemudian ditranspor melalui membran sel menuju apoplas. Glukosa kemudian dihilangkan dan monolignol kemudian dipolimerisasi membentuk lignin (Aust dan Benson 1993). Tiga jenis monolignol (tergolong dalam unit fenilpropana) yaitu koniferil alkohol, sinapil alkohol, dan parakuramil alkohol (Gambar 1). Molekul lignin tersusun dalam bentuk tiga dimensi dan berikatan silang pada setiap molekul penyusunnya dengan jenis ikatan kimia yang berbeda. Unit fenil propana terikat satu sama lain dengan ikatan eter dan ikatan C-C, dengan persentasi ikatan eter lebih banyak. Gambar 2 memperlihatkan struktur lignin.
Kayu dengan kandungan lignin yang tinggi bersifat tahan lama (durable) dan merupakan bahan baku yang baik bagi industri furnitur karena lignin menghasilkan lebih banyak energi saat dibakar dibandingkan dengan selulosa. Kandungan lignin yang tinggi justru menjadi halangan pada industri kertas, lignin dalam kandungan tinggi membuat pulp menjadi kaku dan kekuatannya rendah sehingga lignin harus dihilangkan dari pulp terlebih dahulu sebelum diolah menjadi kertas.
Gambar 1 Struktur tiga jenis monolignol
Gambar 2 Struktur lignin
Fungi Pelapuk Putih (White-rot Fungus)
Fungi Pelapuk Putih (FPP) dari kelas
Basidiomycetes merupakan organisme yang bekerja efisien dan efektif dalam proses biodelignifikasi. Ada jenis jamur lain yang juga mampu mendegradasi lignin, seperti fungi pelapuk coklat (Brown-rot fungus) namun enzim yang dihasilkan oleh jenis jamur ini tidak bekerja se-efektif enzim yang dihasilkan FPP. Proses biodelignifikasi ini dimulai saat FPP menembus dan membentuk koloni dalam sel kayu lalu mengeluarkan enzim yang berdifusi melalui lumen dan dinding sel. Hal ini menyebabkan terjadinya penurunan kekuatan fisik kayu dan pembengkakan jaringan kayu. Intinya FPP, yang menggunakan selulosa sebagai sumber karbon, memiliki kemampuan yang unik untuk mendegradasi lignin secara keseluruhan membentuk karbon dioksida untuk memperoleh molekul selulosa. Jenis FPP yang banyak ditemui di alam adalah Armillariella mellea, Heterobasidion annosum, Coriolus versicolor, Xylaria polymorpha, Ganoderma lucidum, Agraylie sp., dan Pholiota sp. Beberapa yang digunakan dalam industri yang terkait dengan degradasi lignin seperti industri
Gambar 3 Jenis FPP C. versicolor (Deacon 2005)
(Hattaka 2005), C. versicolor, Trametes versicolor, Phanerochaete chrysosporium, dan G. Lucidum (Deacon 2005).
FPP tergolong jamur yang sangat efisien dalam peggunaan nitrogen . Sebagai contoh kandungan nitrogen C. versicolcor saat ditumbuhkan pada media C:N dengan rasio 32:1 kira-kira sebesar 4%, namun saat ditumbuhkan pada media C:N rasio 1600:1 diperoleh 0,2% nitrogen. Hal ini menunjukkan dalam kondisi sedikit nitrogen, jamur lebih mengalokasikan nitrogen untuk memproduksi enzim ekstraselular dan komponen esensial sel, dan jamur ini juga secara efisien mendaur ulang nitrogen di dalam miseliumnya (Deacon 2005)
Ciri-ciri serangan FPP pada kayu yaitu semua komponen utama kayu terdekomposisi dan digunakan oleh jamur sehingga terjadi penyusutan kayu, walaupun tidak terlihat jelas dan bentuk kayu relatif tidak berubah bahkan pada tahap pelapukan lebih lanjut (Hatakka 1994). Ciri serangan lainnya yaitu tidak ada retak melintang serat, kayu menjadi pucat/warna kayu pudar, dan meninggalkan warna putih pada kayu (Gold & Alic 1993). Kemampuan FPP mendegradasi lignin tidak lepas dari peran enzim ekstraselular yang dihasilkannya yang biasa disebut enzim ligninolitik.
Enzim Ligninolitik Ekstraselular
Enzim adalah biokatalisator yang sangat efektif karena enzim dapat mempercepat perubahan kimia (Koolman & Rohm 2000). Kerja kebanyakan enzim sangat spesifik terkait dengan tipe reaksi dan jenis reaktan yang dikatalisis oleh enzim. Menurut Pelczar
et al (1988) enzim tergolong dalam dua tipe berdasarkan lokasi kerjanya yaitu enzim intraselular dan ekstraselular. Enzim intraselular mensintesis bahan-bahan selular dan menjalankan proses metabolisme untuk menyediakan energi bagi sel. Enzim ekstraselular berfungsi melakukan
perubahan-perubahan seperlunya pada nutrien yang terdapat di sekitarnya yang memungkinkan nutrien tersebut memasuki sel. Proses yang tejadi pada jamur yang tumbuh pada bahan lignoselulosa mengeluarkan enzim ekstraselular yang akan mendegradasi bahan tersebut sebagai sumber nutrisinya. Bahan lignoselulosa yang terdiri dari selulosa, hemiselulosa, dan lignin merupakan bahan polimer sehingga enzim yang disekresi oleh jamur akan mengubah bahan lignoselulosa menjadi monomernya sehingga lebih mudah masuk ke dalam sel.
FPP yang hidup pada bahan organik lignoselulosa mengeluarkan enzim ekstraselular yang dapat mendegradasi bahan tersebut sebagai nutrisinya, terutama lignin, sehingga disebut enzim ligninolitik. Terdapat tiga jenis enzim ligninolitik dalam proses degradasi lignin yaitu fenol oksidase (lakase), lignin peroksidase (LiP), dan mangan peroksidase (MnP). Lakase merupakan enzim yang mengandung tembaga dan biasa pula disebut 1,4-benzendiol oksidase (EC 1.10.3.2). Lakase umumnya ditemukan pada tanaman tingkat tinggi dan berbagai mikroorganisme. Reaksi enzimatik pada lakase merupakan reaksi oksidasi yang menghasilkan satu elektron hasil oksidasi senyawa fenol dan mereduksi oksigen menjadi air(www.enzymeindia.com/enzymes/laccase.a sp2006). Pemanfaatan lakase dapat diaplikasikan pada berbagai bidang industri antara lain pada proses bioremediasi dan biodegradasi polutan organik pada tanah seperti klorofenol (Ahn et al 2002), proses dekolorinasi dan detoksifikasi pada pewarna tekstil (Abadulla et al 2000), dan proses bleaching pada biodelignifikasi pulp industri kertas (Bourbonnais & Paice 1992).
LiP (EC 1.11.1.14) tidak diproduksi oleh semua jenis FPP. LiP merupakan enzim yang mengandung heme dengan potensial redoks yang tinggi yang membutuhkan dua metabolit utama agar dapat bekerja. Metabolit tersebut adalah hidrogen peroksida (H2O2) dan veratril
alkohol yang digunakan sebagai mediator untuk reaksi redoks (mekanisme ping-pong). MnP (EC 1.11.1.13) juga merupakan enzim yang mengandung heme dan menggunakan H2O2 untuk mengkatalisis oksidasi Mn2+
BAHAN DAN METODE
Bahan dan AlatBahan yang digunakan dalam penelitian ini adalah isolat Omphalina sp., isolat
Pleurotus ostreatus, kentang, gula pasir, bubuk agar, akuades, biji jewawut, tandan kosong kelapa sawit (TKKS), bagas tebu, dedak padi, CuSO4 150μM, bufer asetat 0,5 M
pH 5,0, bufer fosfat pH 6,5, bufer Na-laktat 50mM, 2,2-azinobis(3-etilbenztiazoline-6-sulfonat) (ABTS) 1mM, guaiakol 4mM, H2O2
1mM dan 5mM, MnSO4 1mM, bufer asetat
0,05M pH 5,0, dan veratril alkohol 8mM. Alat yang digunakan yaitu laminar air flow cabinet, sentrifus Eppendorf 5417R, autoklaf, spektrofotometer UV-Vis, tabung Eppendorf, pH meter, neraca analitik, cawan Petri, botol media, pipet volumetrik, mikropipet, mortar, pemanas, pengaduk magnetik, dan peralatan gelas.
Metode
Rancangan Percobaan
Penelitian yang dilakukan menggunakan model Rancangan Acak Kelompok Lengkap atau Faktorial RAKL (Mattjik & Sumertajaya 2002). Model linier rancangan ini dituliskan sebagai berikut:
Yijk = µ + αi + ßj + (αβ)ij + ρk + εijk
Keterangan: i = 1,2,3,4 j = 1,2 k = 1,2
Yijk : pengamatan pertumbuhan hifa jamur
pada media ke-j yang diberi suplemen ke-i dari isolat ke-k
µ : rataan umum
αi : pengaruh perlakuan suplemen ke-i
ßj : pengaruh media ke-j
(αβ)ij : interaksi antara perlakuan suplemen
ke-i dan media ke-j ρk : pengaruh isolat ke-k
εijk : pengaruh galat percobaan pada media
ke-j yang diberi suplemen ke-i dari isolat ke-k
Pembuatan Media Potatoes Dextrose Agar (PDA) dan Biji Jewawut
Sebanyak 100g kentang dikupas dan dipotong dadu. Kentang dimasukkan ke dalam wadah berisi 500mL air kemudian dididihkan. Ekstrak kentang disaring ke dalam labu
Erlenmeyer kemudian ditambahkan agar 7,5g, gula pasir 10g, dan air hingga tepat 500mL. Labu Erlenmeyer ditutup dengan sumbat kapas dan aluminium foil lalu diautoklaf pada suhu 1210C selama 15 menit. Media PDA yang telah disterilisasi dituang ke cawan petri steril. Penuangan dilakukan di laminar air flow cabinet secara aseptik. Setelah dituang ke cawan petri, media dibiarkan hingga padat. Sebanyak 1kg biji jewawut dididihkan hingga biji jewawut pecah. Setelah mendidih biji jewawut ditiriskan dan diisi ke dalam botol media hingga setengah penuh. Botol ditutup dengan kertas lalu diautoklaf pada suhu 1210C selama 15 menit.
Peremajaan dan Pembuatan Inokulum
Isolat Omphalina sp. dan P. ostreatus yang telah tersedia di media PDA dipotong dadu dan dipindahkan 2 potong ke dalam media PDA secara aseptik. Masing-masing isolat dikulturkan pada 7 media PDA. Setelah selesai, cawan petri ditutup dengan parafilm
dan diinkubasi selama satu minggu.
Fungi yang telah tumbuh setelah diinkubasi kemudian dipindahkan lagi ke dalam media biji jewawut steril. Isolat fungi pada PDA dipotong dadu, kemudian dipindahkan 2-3 potong ke dalam media biji jewawut secara aseptik. Setiap isolat fungi dikulturkan dalam 24 media biji jewawut. Setelah selesai, media biji jewawut lalu diinkubasi selama 1-2 minggu untuk menumbuhkan miselium fungi.
Pembuatan Media TKKS dan Bagas Tebu
Sebanyak 500 gram TKKS dan 500 gram bagas tebu yang telah dipotong ditimbang pada wadah yang berbeda, kemudian dikondisikan sesuai perlakuan yang akan diberikan. Perlakuan yang diberikan yaitu penambahan suplemen CuSO4, dedak, CuSO4
dan dedak, serta tanpa suplemen CuSO4 dan
dedak (kontrol). Sebanyak 0,0119 gram CuSO4 dilarutkan dalam 500 mL air kemudian
dicampurkan dengan media TKKS dan bagas tebu pada wadah yang berbeda lalu diaduk rata. Media yang telah bercampur dengan CuSO4 kemudian dimasukkan ke dalam
plastik tahan panas lalu ditutup dengan cincin pipa, diikat dengan karet gelang, disumbat dengan kapas, dan ditutup dengan kertas.
rata. Media kemudian dimasukkan ke dalam plastik tahan panas lalu ditutup dengan cincin pipa, diikat dengan karet gelang, disumbat dengan kapas, dan ditutup dengan kertas.
Perlakuan ketiga adalah TKKS dan bagas tebu diberi penambahan CuSO4 dan dedak.
Sebanyak 0,0119 gram CuSO4 dilarutkan
dalam 500 mL air, sedangkan untuk dedak sebanyak 150 gram dicampur ke dalam media TKKS dan bagas tebu lalu ditambahkan 500 mL air dan diaduk rata bersama larutan CuSO4 tadi. Setelah tercampur rata media
kemudian dimasukkan ke dalam plastik tahan panas lalu ditutup dengan cincin pipa, diikat dengan karet gelang, disumbat dengan kapas, dan ditutup dengan kertas.
Sebagai kontrol pertumbuhan perlakuan yang diberikan yaitu media tidak diberi penambahan suplemen. Media TKKS dan bagas tebu dicampur dengan 500 mL air, diaduk rata, kemudian dimasukkan ke dalam plastik tahan panas lalu ditutup dengan cincin pipa, diikat dengan karet gelang, disumbat dengan kapas, dan ditutup dengan kertas. Seluruh media kemudian disterilisasi di autoklaf 30 menit 121ºC. Untuk setiap perlakuan pada dua jenis media yang digunakan (TKKS dan bagas tebu) dilakukan 3 kali ulangan.
Penanaman Isolat Fungi pada Media TKKS dan Bagas Tebu
Dua biakan fungi yang telah tumbuh pada media biji jewawut diaduk lalu diambil 10 gram dan diinokulasi secara aseptik ke dalam media TKKS dan bagas tebu untuk setiap perlakuan. Setelah dilakukan penanaman kedua jenis fungi pada seluruh media akan diperoleh 48 bag log, 24 media untuk
P.ostreatus dan 24 lainnya untuk Omphalina
sp. Biakan jamur dalam bag log kemudian diinkubasi selama 3 bulan hingga miselium tumbuh merata. Pertumbuhan miselium diamati setiap bulan selama periode inkubasi.
Ekstraksi Enzim
Biakan fungi yang telah tumbuh pada media bagas dan TKKS setelah diinkubasi 2-3 bulan diambil 20 gram, kemudian digerus di dalam mortar bersama 10 mL bufer fosfat pH 7. Setelah halus, dimasukkan ke tabung Eppendorf dan disentrifus dengan kecepatan 5000 rpm selama 10 menit dengan suhu 0-4ºC. Supernatan dipisahkan dari pelet lalu dimasukkan ke tabung Eppendorf yang lain. Jika supernatan yang diperoleh masih keruh dilakukan sentrifus lagi hingga diperoleh
supernatan yang jernih/bebas kotoran. Supernatan yang diperoleh merupakan ekstrak kasar enzim yang kemudian akan dianalisis aktivitas enzimnya dengan variasi pH (5, 6, dan 7) untuk setiap pengukuran.
Analisis Aktivitas Enzim
Aktivitas Lakase (Buswell et al 1995 dalam Fitria 2005)
Larutan sampel yang akan dianalisis dibuat dengan 0,4 mL filtrat enzim dicampur dengan 0,5 mL bufer asetat 0,5 M pH 5/pH 6/pH 7 dan 0,1 mL ABTS 1 mM. Campuran ini dimasukkan ke dalam kuvet kemudian dikocok. Setelah dikocok larutan diukur absorbannya pada panjang gelombang 420 nm dengan interval waktu 0-30 menit. Aktivitas enzim diukur berdasarkan persamaan berikut (Fitria 2005)
Aktivitas Enzim (U/mL) = (At - Ao) x Vtotal (mL) x 10
9
εmaks x d x Venzim (mL) x t
εmaks = absorptivitas molar ABTS (36000
M-1 cm-1)
d = tebal kuvet (cm)
Satu unit aktivitas lakase didefinisikan sebagai jumlah enzim yang dibutuhkan untuk mengoksidasi 1 nmol ABTS per menit.
Aktivitas Lignin Peroksidase (LiP) (Tien & Kirk 1984 dalam Fitria 2005)
Sebanyak 0,2 mL filtrat enzim, 0,05 mL H2O2 5 mM, 0,1 mL veratril alkohol 8
mM, 0,2 mL bufer asetat 0,05 M pH 5/pH 6/pH 7, dan 0,45 mL akuades dimasukkan ke dalam kuvet kemudian dikocok. Larutan tersebut dibaca absorbannya pada panjang gelombang 310 nm pada interval waktu 0-30 menit. Aktivitas enzim diukur berdasarkan persamaan berikut (Fitria 2005).
Aktivitas Enzim (U/mL) = (At - Ao) x Vtotal (mL) x 109 εmaks x d x Venzim (mL) x t
εmaks = absorptivitas molar veratril alkohol
(9300 M-1 cm-1) d = tebal kuvet (cm)
Satu unit LiP didefinisikan sebagai banyak enzim yang mengoksidasi 1 nmol substrat per menit.
Aktivitas Mangan Peroksidase (MnP) (Kofujita et al 1992 dalam Fitria 2005)
0,2 mL MnSO4 1 mM, 0,1 mL guaiakol 4
mM, dan 0,3 mL akuades (campuran A) dimasukkan ke dalam kuvet berisi 0,2 mL filtrat enzim. Larutan ini dikocok lalu diukur absorbannya pada panjang gelombang 465 nm dengan interval waktu 0-30 menit. Pengukuran aktivitas MnP diperoleh dengan melakukan reaksi dengan komposisi pereaksi yang sama dengan campuran A hanya saja tanpa penambahan MnSO4, sehingga akuades
yang ditambahkan pada reaksi ini sebanyak 0,5 mL (campuran B). Larutan ini kemudian diukur absorbannya pada panjang gelombang 465 nm dengan interval waktu 0-30 menit. Aktivitas enzim diukur berdasarkan persamaan berikut (Fitria 2005).
Aktivitas Enzim (U/mL) = (At - Ao) x Vtotal (mL) x 109 εmaks x d x Venzim (mL) x t
εmaks = absorptivitas molar guaiakol
(12100 M-1 cm-1) d = tebal kuvet (cm)
Aktivitas MnP setiap unit = Aktivitas enzim A – Aktivitas enzim B
Satu unit MnP sebanding dengan 1 nmol produk yang dihasilkan per menit.
HASIL DAN PEMBAHASAN
Pertumbuhan Miselium FPPTahap awal penelitian ini adalah peremajaan isolat murni FPP dari stok yang tersedia pada media PDA. Peremajaan isolat dilakukan dengan menumbuhkan pada media PDA. Selanjutnya dilakukan pembuatan inokulum dengan menumbuhkan pada media biji jewawut. Peremajaan pada media PDA dilakukan selama 1 minggu hingga miselium tumbuh menutupi seluruh permukaan media agar. Hasil peremajaan kedua isolat jamur pada media PDA (Gambar 4) menunjukkan isolat Omphalina sp. tumbuh lebih cepat dengan produksi miselium yang lebih banyak dibandingkan dengan isolat P.ostreatus. Setelah miselium tumbuh menutupi media agar, isolat kemudian diinokulasi ke dalam media biji jewawut.
Hasil pertumbuhan isolat pada media biji jewawut (Gambar 5) menunjukkan miselium tumbuh banyak dan sangat kompak. Kecepatan pertumbuhan miselium kedua jamur pada media biji jewawut berbeda. Berdasarkan pengamatan miselium isolat
Omphalina sp. tumbuh lebih cepat dan
menyelimuti seluruh media biji jewawut dalam waktu inkubasi 1 minggu sedangkan miselium P.ostreatus tumbuh lebih lambat, membutuhkan waktu inkubasi 2 minggu hingga miselium menyelimuti seluruh media biji jewawut. Berdasarkan pengamatan pertumbuhan miselia fungi di media PDA dan biji jewawut dapat dibedakan juga struktur miselia. Miselium tumbuh lebih kompak pada media biji jewawut dibandingkan dengan PDA. Hal ini dapat disebabkan dalam biji jewawut terkandung cadangan lemak yang berguna untuk pertumbuhan biji. Cadangan lemak ini dapat digunakan oleh fungi sebagai sumber nutrisi, terutama sumber karbon, untuk mendukung pertumbuhannya yang akan mendorong semakin banyaknya miselium yang tumbuh pada media biji jewawut dibandingkan dengan PDA.
Setelah miselium tumbuh merata pada media biji jewawut, isolat diinokulasi ke media pertumbuhan dengan substrat utama berupa TKKS dan bagas tebu. Bag log yang digunakan tingginya 18cm dan berat bersih media yang digunakan sebesar 500 gram. Media pertumbuhan yang digunakan telah dikondisikan sesuai perlakuan yang diberikan yaitu penambahan CuSO4, dedak, atau
campuran CuSO4 dan dedak, serta perlakuan
kontrol. Isolat kemudian diinkubasi selama tiga bulan. Selama periode inkubasi dilakukan pengamatan pertumbuhan miselium pada media TKKS dan bagas tebu. Pengamatan dilakukan setiap bulan selama tiga bulan masa inkubasi. Parameter yang diamati yaitu tinggi pertumbuhan miselium pada setiap bag log. Pengukuran tinggi pertumbuhan miselium dimulai pada batas atas media pertumbuhan hingga batas akhir pertumbuhan miselium pada media.
(a)
(b)
Gambar 4 Pertumbuhan miselium pada PDA isolat Omphalina sp. (a) dan
0,2 mL MnSO4 1 mM, 0,1 mL guaiakol 4
mM, dan 0,3 mL akuades (campuran A) dimasukkan ke dalam kuvet berisi 0,2 mL filtrat enzim. Larutan ini dikocok lalu diukur absorbannya pada panjang gelombang 465 nm dengan interval waktu 0-30 menit. Pengukuran aktivitas MnP diperoleh dengan melakukan reaksi dengan komposisi pereaksi yang sama dengan campuran A hanya saja tanpa penambahan MnSO4, sehingga akuades
yang ditambahkan pada reaksi ini sebanyak 0,5 mL (campuran B). Larutan ini kemudian diukur absorbannya pada panjang gelombang 465 nm dengan interval waktu 0-30 menit. Aktivitas enzim diukur berdasarkan persamaan berikut (Fitria 2005).
Aktivitas Enzim (U/mL) = (At - Ao) x Vtotal (mL) x 109 εmaks x d x Venzim (mL) x t
εmaks = absorptivitas molar guaiakol
(12100 M-1 cm-1) d = tebal kuvet (cm)
Aktivitas MnP setiap unit = Aktivitas enzim A – Aktivitas enzim B
Satu unit MnP sebanding dengan 1 nmol produk yang dihasilkan per menit.
HASIL DAN PEMBAHASAN
Pertumbuhan Miselium FPPTahap awal penelitian ini adalah peremajaan isolat murni FPP dari stok yang tersedia pada media PDA. Peremajaan isolat dilakukan dengan menumbuhkan pada media PDA. Selanjutnya dilakukan pembuatan inokulum dengan menumbuhkan pada media biji jewawut. Peremajaan pada media PDA dilakukan selama 1 minggu hingga miselium tumbuh menutupi seluruh permukaan media agar. Hasil peremajaan kedua isolat jamur pada media PDA (Gambar 4) menunjukkan isolat Omphalina sp. tumbuh lebih cepat dengan produksi miselium yang lebih banyak dibandingkan dengan isolat P.ostreatus. Setelah miselium tumbuh menutupi media agar, isolat kemudian diinokulasi ke dalam media biji jewawut.
Hasil pertumbuhan isolat pada media biji jewawut (Gambar 5) menunjukkan miselium tumbuh banyak dan sangat kompak. Kecepatan pertumbuhan miselium kedua jamur pada media biji jewawut berbeda. Berdasarkan pengamatan miselium isolat
Omphalina sp. tumbuh lebih cepat dan
menyelimuti seluruh media biji jewawut dalam waktu inkubasi 1 minggu sedangkan miselium P.ostreatus tumbuh lebih lambat, membutuhkan waktu inkubasi 2 minggu hingga miselium menyelimuti seluruh media biji jewawut. Berdasarkan pengamatan pertumbuhan miselia fungi di media PDA dan biji jewawut dapat dibedakan juga struktur miselia. Miselium tumbuh lebih kompak pada media biji jewawut dibandingkan dengan PDA. Hal ini dapat disebabkan dalam biji jewawut terkandung cadangan lemak yang berguna untuk pertumbuhan biji. Cadangan lemak ini dapat digunakan oleh fungi sebagai sumber nutrisi, terutama sumber karbon, untuk mendukung pertumbuhannya yang akan mendorong semakin banyaknya miselium yang tumbuh pada media biji jewawut dibandingkan dengan PDA.
Setelah miselium tumbuh merata pada media biji jewawut, isolat diinokulasi ke media pertumbuhan dengan substrat utama berupa TKKS dan bagas tebu. Bag log yang digunakan tingginya 18cm dan berat bersih media yang digunakan sebesar 500 gram. Media pertumbuhan yang digunakan telah dikondisikan sesuai perlakuan yang diberikan yaitu penambahan CuSO4, dedak, atau
campuran CuSO4 dan dedak, serta perlakuan
kontrol. Isolat kemudian diinkubasi selama tiga bulan. Selama periode inkubasi dilakukan pengamatan pertumbuhan miselium pada media TKKS dan bagas tebu. Pengamatan dilakukan setiap bulan selama tiga bulan masa inkubasi. Parameter yang diamati yaitu tinggi pertumbuhan miselium pada setiap bag log. Pengukuran tinggi pertumbuhan miselium dimulai pada batas atas media pertumbuhan hingga batas akhir pertumbuhan miselium pada media.
(a)
(b)
Gambar 4 Pertumbuhan miselium pada PDA isolat Omphalina sp. (a) dan
(a)
(b)
Gambar 5 Pertumbuhan miselium pada media biji jewawut isolat Omphalina sp. (a) dan P.ostreatus (b)
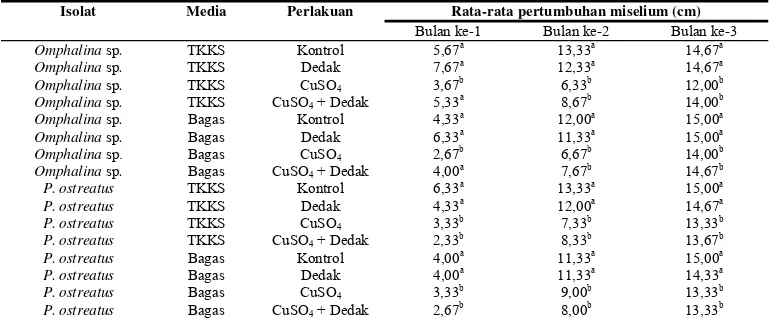
Berdasarkan data terlihat isolat Omphalina sp. baik pada TKKS dan bagas tebu. Pertimbuhannya cenderung lebih baik pada bulan pertama hingga bulan ketiga masa inkubasi dibandingkan dengan P.ostreatus.
Data tersebut menunjukkan Omphalina sp. dapat memanfaatkan sumber nitrogen dan karbon lebih efektif dibanding P.ostreatus
sehingga pertumbuhan miselium lebih baik (Tabel 1).
Bulan pertama masa inkubasi menunjukkan miselium tumbuh lebih baik pada media yang diberi dedak dan perlakuan kontrol, sedangkan pada perlakuan CuSO4 dan
campuran CuSO4 dan dedak, miselium
cenderung tumbuh lebih lambat dibandingkan dengan dua perlakuan lainnya. Hasil ini sama dengan penelitian lainnya yang menyebutkan miselium tumbuh dengan baik pada media yang diberi dedak, yang disebabkan dedak memiliki kandungan karbon dan nitrogen yang tinggi (Yuniawati 2006). Selain merupakan sumber nitrogen dedak juga mengandung tiamin yang merupakan salah satu vitamin yang dibutuhkan dalam pertumbuhan miselium (Tominaga 1978).
Pertumbuhan miselium pada bulan kedua dan bulan ketiga terus menunjukkan peningkatan pada kedua isolat fungi di setiap media tumbuh. Pada perlakuan penambahan CuSO4 dan campuran CuSO4 dan dedak
pertumbuhan miselium lebih rendah dibanding perlakuan dedak. Hal ini berbeda dengan penelitian lain dari Palmieri et al
(1999) menyebutkan pada isolat P.ostreatus, Cu adalah penyokong aktivitas lakase.
Hasil uji Duncan pada data pertumbuhan miselium di bulan ketiga menunjukkan tidak ada perbedaan nyata antara perlakuan kontrol dan dedak terhadap pertumbuhan miselium,
namun kelompok ini menunjukkan perbedaan nyata dengan perlakuan CuSO4 dan campuran
CuSO4 dan dedak. Ditinjau berdasarkan data
pertumbuhan miselium dan analisis statistika dapat dikatakan bahwa perlakuan kontrol menunjukkan pertumbuhan miselium lebih baik, terutama pada isolat P.ostreatus yang menunjukkan rata-rata pertumbuhan miselium pada bag log kontrol lebih baik dibandingkan dengan perlakuan lain, sehingga tidak diperlukan dedak, CuSO4 maupun campuran
CuSO4 dan dedak untuk menunjang
pertumbuhan miselium FPP. Ditilik dari segi ekonomi, hal ini menguntungkan sebab fungi dapat tumbuh dengan baik namun biaya produksi dapat ditekan.
Data yang diperoleh juga menunjukkan bahwa perlakuan CuSO4 dan campuran
CuSO4 dan dedak cenderung menghambat
pertumbuhan fungi. Hal ini terlihat dengan rendahnya pertumbuhan miselium di bulan pertama dan kedua dibandingkan dengan perlakuan kontrol. Pada bulan ketiga pertumbuhan miselium pada perlakuan CuSO4
dan campuran CuSO4 dan dedak menunjukkan
peningkatan yang signifikan dari bulan sebelumnya. Hal ini dapat disebabkan daya hambat suplemen yang diberikan telah berkurang atau sudah tidak menghambat sama sekali sehingga pertumbuhan miselium lebih baik.
Apabila media pertumbuhan dibandingkan dari data Tabel 1, terlihat bahwa media pertumbuhan bagas tebu lebih baik dalam mendukung pertumbuhan miselium dibandingkan dengan TKKS. Hal ini terlihat dari pertumbuhan miselium Omphalina sp.
pada perlakuan kontrol TKKS dan bagas tebu masing-masing 14,67 cm dan 15,00 cm, begitu pula pada perlakuan lainnya. Berbeda dengan isolat Omphalina sp., isolat
P.ostreatus menunjukkan pertumbuhan miseliumnya lebih baik pada media TKKS dibandingkan dengan media bagas tebu. Hal ini ditunjukkan pada media yang diberi dedak dan campuran CuSO4 dan dedak. Ini berarti
sumber karbon dan nitrogen yang diperlukan fungi untuk aktivitas metabolisme dan pertumbuhan miselium lebih banyak terdapat pada media TKKS dengan kandungan lignin yang lebih tinggi yaitu 28,54% (Santosa 1993) dibanding bagas tebu sebesar 22,09%.
Tabel 1 Pertumbuhan miselium FPP
Isolat Media Perlakuan Rata-rata pertumbuhan miselium (cm)
Bulan ke-1 Bulan ke-2 Bulan ke-3
Omphalina sp. TKKS Kontrol 5,67a
13,33a
14,67a
Omphalina sp. TKKS Dedak 7,67a
12,33a
14,67a
Omphalina sp. TKKS CuSO4 3,67b 6,33b 12,00b
Omphalina sp. TKKS CuSO4 + Dedak 5,33a 8,67b 14,00b Omphalina sp. Bagas Kontrol 4,33a
12,00a
15,00a
Omphalina sp. Bagas Dedak 6,33a
11,33a
15,00a
Omphalina sp. Bagas CuSO4 2,67b 6,67b 14,00b
Omphalina sp. Bagas CuSO4 + Dedak 4,00a 7,67b 14,67b
P. ostreatus TKKS Kontrol 6,33a
13,33a
15,00a
P. ostreatus TKKS Dedak 4,33a
12,00a
14,67a
P. ostreatus TKKS CuSO4 3,33b 7,33b 13,33b
P. ostreatus TKKS CuSO4 + Dedak 2,33b 8,33b 13,67b
P. ostreatus Bagas Kontrol 4,00a
11,33a
15,00a
P. ostreatus Bagas Dedak 4,00a
11,33a
14,33a
P. ostreatus Bagas CuSO4 3,33b 9,00b 13,33b
P. ostreatus Bagas CuSO4 + Dedak 2,67b 8,00b 13,33b
Keterangan: Angka yang diikuti huruf yang sama tidak berbeda nyata berdasarkan uji Duncan (P<0,05)
37,65% dibandingkan dengan TKKS sebesar 32,55% (Santosa 1993). Perbedaan kecepatan tumbuh Omphalina sp. dan P.ostreatus pada kedua media menunjukkan adanya spesifisitas tiap fungi pada media pertumbuhan. Diduga
P.ostreatus lebih sesuai untuk media yang kandungan ligninnya lebih tinggi seperti pada TKKS sedangkan Omphalina sp. sesuai pada media yang mempunyai kandungan selulosa yang lebih tinggi, dalam hal ini yaitu bagas tebu.
Analisis Aktivitas Enzim Ligninolitik
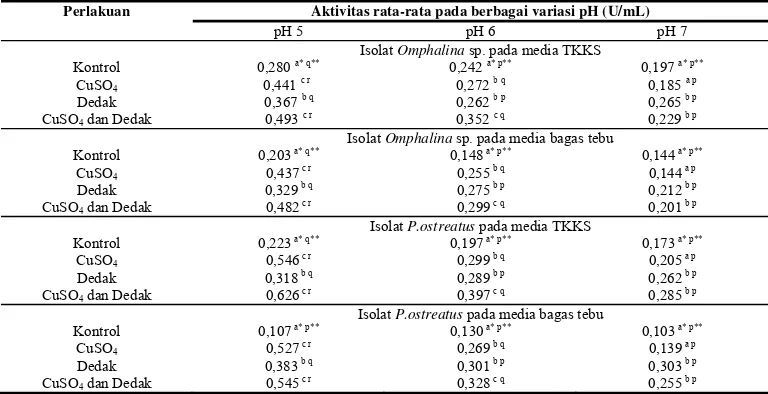
Analisis aktivitas enzim ligninolitik dari kedua isolat jamur dilakukan setelah masa inkubasi tiga bulan pada media TKKS dan bagas tebu. Tahap awal dilakukan ekstraksi enzim ligninolitik dari media TKKS dan bagas tebu. Ekstraksi dilakukan dengan bufer fosfat pH 7 karena FPP umumnya hidup pada pH netral (Panji et al 1996). Pada tahap analisis aktivitas enzim ligninolitik dilakukan karakterisasi pH untuk setiap enzim ligninolitik yaitu pH 5, 6, dan 7 terhadap bufer yang digunakan. Hasil uji Duncan terhadap aktivitas lakase (Tabel 2) menunjukkan terdapat perbedaan nyata antara aktivitas lakase pada pH 5 dengan pH 6 dan 7. Analisis pada pH 5 menunjukkan aktivitas optimum pada seluruh perlakuan dibandingkan dengan pH 6 dan 7. Nilai pH optimum yang diperoleh sesuai dengan metode analisis aktivitas lakase yang dikembangkan Buswell et al (1995) yang menggunakan bufer asetat pH 5 dalam analisisnya. Hasil penelitian lainnya terhadap aktivitas lakase pada P.ostreatus
menunjukkan lakase optimum pada pH 6,5 dengan nilai aktivitas lakase rata-rata mencapai 2,4 U/mL (Mansur et al 2003).
Aktivitas lakase dengan penambahan suplemen CuSO4, dedak, dan campuran
CuSO4 dan dedak memperlihatkan pengaruh
positif yaitu meningkatnya aktivitas lakase dibandingkan dengan perlakuan kontrol pada setiap pH. Isolat Omphalina sp. pada pH 5, mensekresikan lakase di media TKKS dan bagas tebu perlakuan kontrol masing-masing sebesar 0,280U/mL dan 0,203U/mL. Terdapat pengaruh positif terhadap penambahan suplemen CuSO4, dedak, dan campuran
CuSO4 dan dedak terhadap aktivitas lakase
yaitu aktivitas enzim meningkat hingga 0,441U/mL, 0,367U/mL, dan 0, 493U/mL pada TKKS (Tabel 2) dan pada bagas tebu 0,437U/mL, 0,329U/mL, dan 0,482U/mL. Jadi peningkatan tertinggi terlihat pada pemberian suplemen campuran CuSO4 dan
dedak baik pada TKKS maupun bagas. Aktivitas lakase dari fungi P.ostreatus
pada media TKKS juga meningkat dengan pemberian suplemen. Aktivitas lakase pada perlakuan kontrol sebesar 0,223U/mL, sedangkan aktivitas lakase pada TKKS yang diberi suplemen CuSO4 sebesar 0,546U/mL,
pada penambahan dedak sebesar 0,318U/mL, dan pada perlakuan campuran CuSO4 dan
dedak aktivitas lakase sebesar 0,626 U/mL. Aktivitas lakase pada pH optimum 5, pada media bagas tebu isolat P.ostreatus
menunjukkan pada kontrol aktivitas lakase sebesar 0,107U/mL, sedangkan pada pemberian suplemen aktivitas lakase lebih tinggi yaitu 0,527U/mL pada penambahan suplemen CuSO4, 0,383U/mL pada pemberian
suplemen dedak, dan aktivitas tertinggi diperoleh sebesar 0,545U/mL pada pemberian suplemen campuran CuSO4 dan dedak.
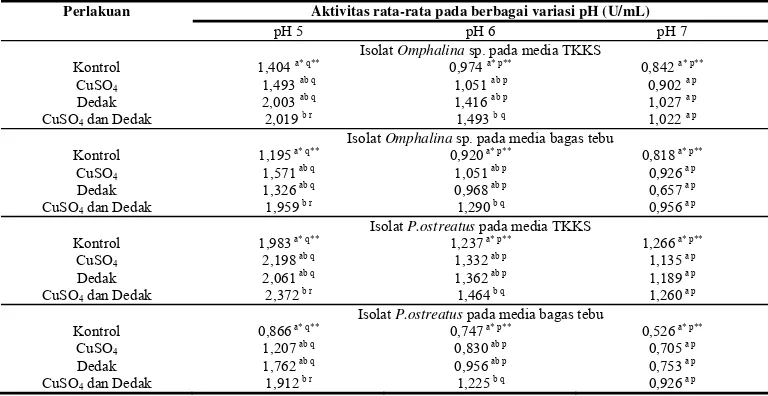
Tabel 2 Aktivitas lakase dari FPP dalam media lignoselulosa
Perlakuan Aktivitas rata-rata pada berbagai variasi pH (U/mL)
pH 5 pH 6 pH 7
Isolat Omphalina sp. pada media TKKS Kontrol 0,280 a* q**
0,242 a* p**
0,197 a* p**
CuSO4 0,441 c r 0,272 b q 0,185 a p
Dedak 0,367 b q
0,262 b p
0,265 b p
CuSO4 dan Dedak 0,493 c r 0,352 c q 0,229 b p
Isolat Omphalina sp. pada media bagas tebu Kontrol 0,203a* q**
0,148a* p**
0,144 a* p**
CuSO4 0,437c r 0,255b q 0,144 a p
Dedak 0,329b q
0,275b p
0,212 b p
CuSO4 dan Dedak 0,482c r 0,299c q 0,201 b p
Isolat P.ostreatus pada media TKKS Kontrol 0,223a* q**
0,197a* p**
0,173 a* p**
CuSO4 0,546c r 0,299b q 0,205 a p
Dedak 0,318b q
0,289b p
0,262 b p
CuSO4 dan Dedak 0,626c r 0,397c q 0,285 b p
Isolat P.ostreatus pada media bagas tebu Kontrol 0,107a* p**
0,130a* p**
0,103 a* p**
CuSO4 0,527c r 0,269b q 0,139 a p
Dedak 0,383b q
0,301b p
0,303 b p
CuSO4 dan Dedak 0,545c r 0,328c q 0,255 b p
Keterangan: Analisis statistik dilakukan terpisah pada tiap kategori isolat dan media
* = Angka yang diikuti huruf yang sama (vertikal) tidak berbeda nyata berdasarkan uji Duncan (P<0,05)
**= Angka yang diikuti huruf yang sama (horizontal) tidak berbeda nyata berdasarkan uji Duncan (P<0,05)
Aktivitas rata-rata enzim ligninolitik optimum pada pH 5 juga ditunjukkan dari hasil analisis aktivitas lignin peroksidase pada berbagai perlakuan dengan pH 5, 6, dan 7. (Tabel 3). Hasil uji Duncan memperlihatkan aktivitas enzim lignin peroksidase optimum pada pH 5 untuk setiap perlakuan, berbeda nyata dibandingkan dengan pH 6 dan 7. Hasil ini berbeda dengan yang dilaporkan Bockle et al (1999) yang dalam penelitiannya menunjukkan lignin peroksidase optimum pada pH 3,0.
Sekresi lignin peroksidase oleh Omphalina
sp. pada media TKKS dan bagas tebu pada perlakuan kontrol sebesar 1,404U/mL dan 1,195U/mL. Aktivitas lignin peroksidase meningkat dengan pemberian suplemen, baik pada bagas tebu maupun TKKS. Penambahan suplemen pada media TKKS menunjukkan aktivitas lignin peroksidase pada pH optimum 5 sebesar 1,493U/mL pada penambahan CuSO4, 2,043U/mL pada penambahan dedak,
serta 2,019U/mL pada penambahan campuran CuSO4 dan dedak. Sedangkan penambahan
suplemen CuSO4, dedak, dan campuran
CuSO4 dan dedak pada media bagas tebu
menunjukkan aktivitas sebesar 1,571U/mL, 1,326U/mL, dan 1,959U/mL. Hasil ini menunjukkan bahwa pemberian campuran CuSO4 dan dedak meningkatkan aktivitas
lignin peroksidase.
Aktivitas lignin peroksidase yang disekresikan oleh P.ostreatus pada media
TKKS juga menunjukkan pengaruh positif yaitu meningkatnya aktivitas lignin peroksidase dengan penambahan suplemen sama seperti pada isolat Omphalina sp. Aktivitas lignin peroksidase pada pH 5 pada kontrol sebesar 1,983U/mL, penambahan CuSO4 sebesar 2,198U/mL, penambahan
dedak sebesar 2,061U/mL, dan penambahan campuran CuSO4 dan dedak sebesar
2,372U/mL. Aktivitas lignin peroksidase pada bagas tebu diperoleh aktivitas pada kontrol sebesar 0,866U/mL, penambahan CuSO4
sebesar 1,207U/mL, penambahan dedak sebesar 1,762U/mL, dan pada penambahan CuSO4 dan dedak sebesar 1,912U/mL. Data
menunjukkan pada pH optimum yang menunjukkan perbedaan nyata hanya pada penambahan suplemen campuran CuSO4 dan
Tabel 3 Aktivitas lignin peroksidase dari FPP dalam media lignoselulosa
Perlakuan Aktivitas rata-rata pada berbagai variasi pH (U/mL)
pH 5 pH 6 pH 7
Isolat Omphalina sp. pada media TKKS Kontrol 1,404 a* q**
0,974 a* p**
0,842 a* p**
CuSO4 1,493 ab q 1,051 ab p 0,902 a p
Dedak 2,003 ab q
1,416 ab p
1,027 a p
CuSO4 dan Dedak 2,019 b r 1,493 b q 1,022 a p
Isolat Omphalina sp. pada media bagas tebu Kontrol 1,195a* q**
0,920a* p**
0,818 a* p**
CuSO4 1,571ab q 1,051ab p 0,926 a p
Dedak 1,326ab q
0,968ab p
0,657 a p
CuSO4 dan Dedak 1,959b r 1,290b q 0,956 a p
Isolat P.ostreatus pada media TKKS Kontrol 1,983a* q**
1,237a* p**
1,266 a* p**
CuSO4 2,198ab q 1,332ab p 1,135 a p
Dedak 2,061ab q
1,362ab p
1,189 a p
CuSO4 dan Dedak 2,372b r 1,464b q 1,260 a p
Isolat P.ostreatus pada media bagas tebu Kontrol 0,866a* q**
0,747a* p**
0,526 a* p**
CuSO4 1,207ab q 0,830ab p 0,705 a p
Dedak 1,762ab q
0,956ab p
0,753 a p
CuSO4 dan Dedak 1,912b r 1,225b q 0,926 a p
Keterangan: Analisis statistik dilakukan terpisah pada tiap kategori isolat dan media
* = Angka yang diikuti huruf yang sama (vertikal) tidak berbeda nyata berdasarkan uji Duncan (P<0,05)
**= Angka yang diikuti huruf yang sama (horizontal) tidak berbeda nyata berdasarkan uji Duncan (P<0,05)
ligninolitik meningkat dibanding perlakuan kontrol.
Aktivitas lignin peroksidase optimum dengan penambahan suplemen campuran CuSO4 dan dedak, sama seperti lakase.
Namun hal ini tidak tergambar pada hasil analisis aktivitas lignin peroksidase pada media TKKS dengan isolat Omphalina sp.
yang menunjukkan aktivitas tertinggi terjadi pada penambahan dedak. Uji Duncan menunjukan ada perbedaan nyata antara setiap perlakuan terhadap aktivitas enzim lignin peroksidase. Hal ini berarti penambahan suplemen untuk meningkatkan aktivitas enzim ligninolitik berpengaruh terhadap produksi enzim lignin peroksidase.
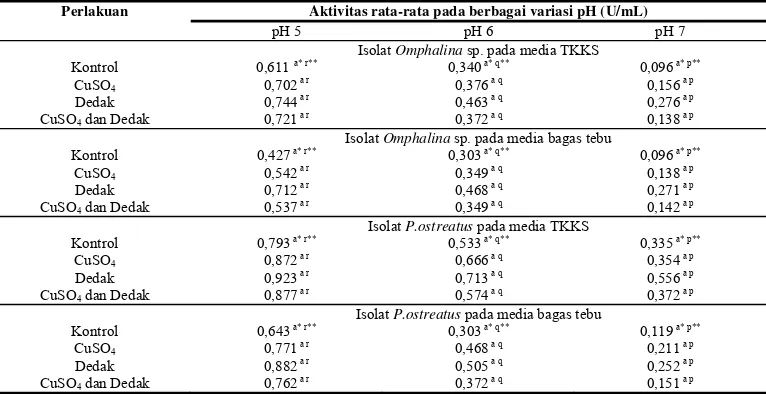
Sama seperti lakase dan lignin peroksidase, analisis aktivitas enzim mangan peroksidase juga menunjukkan aktivitas yang optimum pada pH 5. Sebagai perbandingan terhadap hasil yang diperoleh, Bockle et al
(1999) menyebutkan pH optimum untuk MnP yang disekresikan P.pulmonarius yaitu pH 4,5. Data aktivitas mangan peroksidase terlihat pada Tabel 4. Aktivitas mangan peroksidase yang disekresikan oleh
Omphalina sp. pada media TKKS pada pH optimum, yaitu pH 5 menunjukkan aktivitas kontrol sebesar 0,611U/mL. Penambahan suplemen menunjukkan peningkatan aktivitas mangan peroksidase dibanding kontrol,
dengan aktivitas sebesar 0,702U/mL pada penambahan CuSO4, pada penambahan dedak
sebesar 0,744U/mL, dan campuran CuSO4 dan
dedak sebesar 0,721U/mL. Pada pH 5, isolat
Omphalina sp. pada media bagas tebu untuk kontrol menunjukkan aktivitas mangan peroksidase sebesar 0,427U/mL. Sama seperti TKKS terjadi peningkatan aktivitas enzim mangan peroksidase dengan penambahan suplemen dengan aktivitas sebesar 0,542U/mL pada penambahan CuSO4, 0,712U/mL pada
penambahan dedak, sedangkan penambahan campuran CuSO4 dan dedak sebesar
0,537U/mL. Berdasarkan hasil ini baik pada TKKS maupun bagas penambahan suplemen CuSO4, dedak, dan campuran CuSO4 dan dedak tidak nyata meningkatkan aktivitas mangan peroksidase.
Hal yang sama juga terlihat pada isolat
P.ostreatus pada kedua media tumbuh. Data yang diperoleh pada perlakuan kontrol pada media TKKS pada pH optimum, pH 5, menunjukkan aktivitas mangan peroksidase sebesar 0,793U/mL, dan pada perlakuan pemberian suplemen CuSO4, dedak, dan
campuran CuSO4 dan dedak sebesar
Tabel 4 Aktivitas mangan peroksidase dari FPP dalam media lignoselulosa
Perlakuan Aktivitas rata-rata pada berbagai variasi pH (U/mL)
pH 5 pH 6 pH 7
Isolat Omphalina sp. pada media TKKS Kontrol 0,611 a* r**
0,340a* q**
0,096 a* p**
CuSO4 0,702a r 0,376a q 0,156 a p
Dedak 0,744a r
0,463a q
0,276 a p
CuSO4 dan Dedak 0,721a r 0,372a q 0,138 a p
Isolat Omphalina sp. pada media bagas tebu Kontrol 0,427a* r**
0,303a* q**
0,096 a* p**
CuSO4 0,542a r 0,349a q 0,138 a p
Dedak 0,712a r
0,468a q
0,271 a p
CuSO4 dan Dedak 0,537a r 0,349a q 0,142 a p
Isolat P.ostreatus pada media TKKS Kontrol 0,793a* r**
0,533a* q**
0,335 a* p**
CuSO4 0,872a r 0,666a q 0,354 a p
Dedak 0,923a r
0,713a q
0,556 a p
CuSO4 dan Dedak 0,877a r 0,574a q 0,372 a p
Isolat P.ostreatus pada media bagas tebu Kontrol 0,643a* r**
0,303a* q**
0,119 a* p**
CuSO4 0,771a r 0,468a q 0,211 a p
Dedak 0,882a r
0,505a q
0,252 a p
CuSO4 dan Dedak 0,762a r 0,372a q 0,151 a p
Keterangan: Analisis statistik dilakukan terpisah pada tiap kategori isolat dan media
* = Angka yang diikuti huruf yang sama (vertikal) tidak berbeda nyata berdasarkan uji Duncan (P<0,05)
**= Angka yang diikuti huruf yang sama (horizontal) tidak berbeda nyata berdasarkan uji Duncan (P<0,05)
campuran CuSO4 dan dedak masing-masing
sebesar 0,771U/mL, 0,882U/mL, dan 0,762U/mL. Sama dengan pada Omphalina
sp. pemberian suplemen tidak berbeda nyata dengan kontrol.
Berdasarkan hasil ini terlihat bahwa lakase dan lignin peroksidase memiliki kecenderungan yang sama, menunjukkan peningkatan aktivitas dengan penambahan suplemen, namun tidak untuk mangan peroksidase. Hal ini diduga disebabkan oleh kandungan mangan peroksidase dalam enzim ligninolitik yang disekresikan baik
P.ostreatusmaupun Omphalina sp. sangat sedikit dibanding lakase ataupun lignin peroksidase. Hasil juga menunjukkan bahwa pertumbuhan miselium yang baik akan membuat lebih banyak enzim ligninolitik yang disekresikan sehingga aktivitasnya dalam mendegradasi lignin lebih besar. Namun pada perlakuan kontrol tidak terlihat demikian, meskipun miselium pada perlakuan kontrol tumbuh lebih baik dibandingkan dengan perlakuan CuSO4 dan campuran CuSO4 dan
dedak, aktivitas enzim ligninolitik perlakuan kontrol lebih rendah. Hal ini sesuai dengan penelitian yang dilaporkan Panji et al (1996) yang menyatakan banyaknya miselium tidak menunjukkan aktivitas enzim ligninolitik dalam mendegradasi lignin ataupun selulosa. Perlakuan dedak juga menunjukkan aktivitas enzim ligninolitik yang baik. Dedak merupakan suplemen yang telah banyak
digunakan dalam budi daya jamur. Silverio et al (1981) menyebutkan dedak adalah substrat alami yang dapat mendukung miselium
Auricularia menjadi tebal dan kompak. Walaupun begitu penambahan suplemen ini tidak berperan banyak dalam peningkatan aktivitas enzim mangan peroksidase.
Disebutkan Fitria (2005), isolat FPP
Phanerochaete chrysosporium banyak menjadi objek penelitian karena menghasilkan aktivitas lignin peroksidase dan mangan peroksidase yang tinggi. Hasil yang diperoleh pada penelitian ini juga menunjukkan aktivitas lignin peroksidase yang tinggi dihasilkan
Omphalina sp. dan P.ostreatus. Hal ini berbeda dengan beberapa penelitian sebelumnya yang menyatakan Pleurotus spp. yang ditumbuhkan pada media glucose-malt-yeast menunjukkan aktivitas enzim lignin peroksidase yang rendah (Fukushima & Kirk 1995 dalam Mansur et al 2003). Perlakuan dedak menunjukkan aktivitas enzim mangan peroksidase tertinggi untuk kedua isolat walaupun tidak berbeda nyata. Hal ini berbeda dengan dua enzim sebelumnya yang menunjukkan aktivitas tertinggi pada penambahan campuran CuSO4 dan dedak.
Meskipun begitu pada penambahan CuSO4
serta campuran CuSO4 dan dedak, aktivitas
lainnya. Hal ini berarti seluruh suplemen dapat digunakan sebagai suplemen untuk meningkatkan sekresi enzim ligninolitik, namun tanpa suplemen sekalipun tidak akan berpengaruh terhadap aktivitas mangan peroksidase. Hasil yang diperoleh juga menunjukkan perbedaan antara P.ostreatus
dan Omphalina sp. dalam hal aktivitas enzim ligninolitik. P.ostreatus walaupun pertumbuhannya tidak secepat Omphalina sp. namun aktivitas enzim ligninolitik yang dihasilkannya lebih tinggi. Hal ini sesuai dengan penelitian yang menyebutkan enzim ligninolitik pada Pleurotus spp. lebih aktif dan lebih baik dalam mendegradasi lignin dibandingkan C.versicolor dan Ceriporiopsis subvermispora (Fukushima & Kirk 1995, Marzullo et al 1995). Menarik juga untuk ditinjau dari hasil yang diperoleh adalah penambahan suplemen CuSO4 dan campuran
CuSO4 dan dedak yang menghambat
pertumbuhan miselium pada dua bulan pertama sehingga miselium tidak tumbuh dengan lebih baik dibandingkan dengan pada perlakuan dedak dan kontrol. Namun pada analisis aktivitas enzim ligninolitik pemberian CuSO4 dan campuran CuSO4 dan dedak
menghasilkan aktivitas lakase dan lignin peroksidase yang tinggi dibandingkan dengan perlakuan kontrol dan penambahan dedak.
SIMPULAN DAN SARAN
SimpulanBerdasarkan hasil penelitian, diketahui
P.ostreatus lebih baik dibandingkan dengan
Omphalina sp. dalam memproduksi enzim ligninolitik, terkait dengan interaksi dengan media dan suplemen yang diberikan. Kedua media pertumbuhan yang digunakan, TKKS dan bagas tebu, juga terbukti sama baiknya dalam mendukung pertumbuhan FPP, namun bagas tebu diduga lebih spesifik dengan isolat Omphalina sp., sedangkan P.ostreatus tumbuh lebih baik pada TKKS Hasil penelitian juga menunjukkan suplemen CuSO4, dedak, dan
campuran CuSO4 dan dedak yang diberikan
untuk mendukung pertumbuhan jamur dapat meningkatkan produksi enzim ligninolitik. Namun, terkait dengan pertumbuhan miselium jamur, suplemen dedak lebih baik dibanding lainnya, tetapi dalam hal produksi enzim ligninolitik, media yang diberi suplemen campuran CuSO4 dan dedak lebih baik dalam
mendukung produksi enzim ligninolitik dari isolat FPP. Hasil ini hanya berlaku pada
aktivitas lakase dan lignin peroksidase, sedangkan untuk mangan peroksidase tidak ada perbedaan nyata anatra tiap suplemen. Suplemen yang memberikan aktivitas tertinggi pada enzim lakase adalah campuran CuSO4 dan dedak, begitu pula dengan enzim lignin peroksidase. Suplemen dedak memberian peningkatan aktivitas enzim mangan peroksidase. Selain mengetahui suplemen yang baik, hasil penelitian juga menunjukkan pH optimum untuk semua tipe enzim ligninolitik baik dari isolat Omphalina
sp. maupun P.ostreatus pada pH 5.
Saran
Perlu dilakukan analisis yang berkesinambungan selama masa pertumbuhan jamur untuk mengetahui enzim yang dominan disekresikan oleh jamur dalam masa pertumbuhannya, baik pada fase log maupun fase stasioner. Terkait dengan pemanfaatan di bidang industri, untuk produksi enzim dalam skala yang lebih besar, penggunaan suplemen ini dapat diterapkan, terutama campuran CuSO4 dan dedak yang menghasilkan
aktivitas ketiga enzim ligninolitik yang tinggi. Cu telah terbukti sebagai induser yang baik untuk pertumbuhan FPP serta peningkatan produksi enzim ligninolitik, maka sebaiknya dilakukan pengujian terhadap unsur anorganik lainnya seperti Mg dan Ca yang juga mampu berperan sebaga kofaktor dalam kerja enzim.
DAFTAR PUSTAKA
Abadulla E, Tzanov T, Costa S, Robra KH, Cavoca-Paulo A, Gubitz GM. 2000. Decolorization and detoxification of textile dyes with a laccase from
Trametes hirsute. Appl Environ Microbiol 66: 3357-3362.
Ahn MY, Dec J, Kim JE, Bollog JM. 2002. Treatment of 2,4-dichlorophenol polluted soil with free and immobilized laccase. J Environ Qual. 31:1509-1515.
Aust SD, Benson JT. 1993. The fungus among us: use of white rot fungi to biodegrade environmental pollutants.
AKTIVITAS ENZIM LIGNINOLITIK FUNGI PELAPUK
PUTIH
Omphalina sp
. DAN
Pleurotus ostreatus
PADA LIMBAH
LIGNOSELULOSA
RIO ICHSAN SYAFRIZAL
PROGRAM STUDI BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
lainnya. Hal ini berarti seluruh suplemen dapat digunakan sebagai suplemen untuk meningkatkan sekresi enzim ligninolitik, namun tanpa suplemen sekalipun tidak akan berpengaruh terhadap aktivitas mangan peroksidase. Hasil yang diperoleh juga menunjukkan perbedaan antara P.ostreatus
dan Omphalina sp. dalam hal aktivitas enzim ligninolitik. P.ostreatus walaupun pertumbuhannya tidak secepat Omphalina sp. namun aktivitas enzim ligninolitik yang dihasilkannya lebih tinggi. Hal ini sesuai dengan penelitian yang menyebutkan enzim ligninolitik pada Pleurotus spp. lebih aktif dan lebih baik dalam mendegradasi lignin dibandingkan C.versicolor dan Ceriporiopsis subvermispora (Fukushima & Kirk 1995, Marzullo et al 1995). Menarik juga untuk ditinjau dari hasil yang diperoleh adalah penambahan suplemen CuSO4 dan campuran
CuSO4 dan dedak yang menghambat
pertumbuhan miselium pada dua bulan pertama sehingga miselium tidak tumbuh dengan lebih baik dibandingkan dengan pada perlakuan dedak dan kontrol. Namun pada analisis aktivitas enzim ligninolitik pemberian CuSO4 dan campuran CuSO4 dan dedak
menghasilkan aktivitas lakase dan lignin peroksidase yang tinggi dibandingkan dengan perlakuan kontrol dan penambahan dedak.
SIMPULAN DAN SARAN
SimpulanBerdasarkan hasil penelitian, diketahui
P.ostreatus lebih baik dibandingkan dengan
Omphalina sp. dalam memproduksi enzim ligninolitik, terkait dengan interaksi dengan media dan suplemen yang diberikan. Kedua media pertumbuhan yang digunakan, TKKS dan bagas tebu, juga terbukti sama baiknya dalam mendukung pertumbuhan FPP, namun bagas tebu diduga lebih spesifik dengan isolat Omphalina sp., sedangkan P.ostreatus tumbuh lebih baik pada TKKS Hasil penelitian juga menunjukkan suplemen CuSO4, dedak, dan
campuran CuSO4 dan dedak yang diberikan
untuk mendukung pertumbuhan jamur dapat meningkatkan produksi enzim ligninolitik. Namun, terkait dengan pertumbuhan miselium jamur, suplemen dedak lebih baik dibanding lainnya, tetapi dalam hal produksi enzim ligninolitik, media yang diberi suplemen campuran CuSO4 dan dedak lebih baik dalam
mendukung produksi enzim ligninolitik dari isolat FPP. Hasil ini hanya berlaku pada
aktivitas lakase dan lignin peroksidase, sedangkan untuk mangan peroksidase tidak ada perbedaan nyata anatra tiap suplemen. Suplemen yang memberikan aktivitas tertinggi pada enzim lakase adalah campuran CuSO4 dan dedak, begitu pula dengan enzim lignin peroksidase. Suplemen dedak memberian peningkatan aktivitas enzim mangan peroksidase. Selain mengetahui suplemen yang baik, hasil penelitian juga menunjukkan pH optimum untuk semua tipe enzim ligninolitik baik dari isolat Omphalina
sp. maupun P.ostreatus pada pH 5.
Saran
Perlu dilakukan analisis yang berkesinambungan selama masa pertumbuhan jamur untuk mengetahui enzim yang dominan disekresikan oleh jamur dalam masa pertumbuhannya, baik pada fase log maupun fase stasioner. Terkait dengan pemanfaatan di bidang industri, untuk produksi enzim dalam skala yang lebih besar, penggunaan suplemen ini dapat diterapkan, terutama campuran CuSO4 dan dedak yang menghasilkan
aktivitas ketiga enzim ligninolitik yang tinggi. Cu telah terbukti sebagai induser yang baik untuk pertumbuhan FPP serta peningkatan produksi enzim ligninolitik, maka sebaiknya dilakukan pengujian terhadap unsur anorganik lainnya seperti Mg dan Ca yang juga mampu berperan sebaga kofaktor dalam kerja enzim.
DAFTAR PUSTAKA
Abadulla E, Tzanov T, Costa S, Robra KH, Cavoca-Paulo A, Gubitz GM. 2000. Decolorization and detoxification of textile dyes with a laccase from
Trametes hirsute. Appl Environ Microbiol 66: 3357-3362.
Ahn MY, Dec J, Kim JE, Bollog JM. 2002. Treatment of 2,4-dichlorophenol polluted soil with free and immobilized laccase. J Environ Qual. 31:1509-1515.
Aust SD, Benson JT. 1993. The fungus among us: use of white rot fungi to biodegrade environmental pollutants.
Bockle B, Martinez MJ, Guillen F, Martinez AT. 1999. Mechanism of peroxidase inactivation in liquid cultures of the ligninolytic fungus Pleurotus pulmonarius. Appl Environ Microbiol. 65(3): 923-928.
Bourbonnais R, Paice MG. 1992. Demethylation and delignification of kraft pulp by Trametes versicolor
laccase in the presence of 2,2-azinobis (3-ethylbenzthiazoline–6-sulphonate).
Appl Microbiol Biotechnol. 36:823-827.
Darnoko. 1992. Potensi limbah lignoselulosa kelapa sawit melalui biokonversi.
Berita Penelitian Perkebunan 2:85-95.
Deacon J. 2005. White Rot Fungi. helios.bto.ed.ac.uk/bto/fungal/ biology/ woodrots. htm [17 Oktober 2006].
Fitria R. 2005. Optimasi produksi enzim ligninolitik oleh isolat A1 dan
G.lucidum serta pemurnian parsial dan karakterisasi lakase. Depok: Program Studi Ekstensi Kimia, Universitas Indonesia.
Fukushima Y, Kirk KT. 1995. Laccase component of the Ceriporiopsis subvermispora lignin-degrading system.
Appl Environ Microbiol. 61:872-876.
Gold MH, Alic M. 1993. Molecular biology of the lignin - degrading Basidiomycete Phanerochaete chrysosporium.
Microbiol Rev 57: 605-622.
Hatakka A. 1994. Lignin modifying enzyme from selected white-rot fungi: production and role in lignin degradation. FEMS Microbiol Rev 13: 125-135.
Hatakka A et al. 2005. Environmental biotechnology and biotechnology of natural resources. Proceedings of the Scanbalt Meeting; Helsinki, 31 Okt 2005. Helsinki: Microbiology Society. Hlm 1078-1092.
Kirk KT, Chang HM. 1990. Biotechnology in Pulp and Paper Manufacture. New York: Butterworth-Heinemann.
Kirk KT, Tien M, Croan S, Murtagh KE, Farrell RL.1986. Production of multiple
ligninases by Phanerochaete chrysosporium, effect of selected growth conditions and the use of a mutant strain. Enzyme Microbiol Technol. 8:27-32.
Koolman J, Rohm KH. 2000. Atlas Berwarna dan Teks Biokimia. Wanandi SI, penerjemah; Jakarta: Erlangga. Terjemahan dari: Color Atlas of Biochemistry.
Mansur M, Arias ME, Copa-Patino JL, Flardh M, Gonzalez AE. 2003. The white-rot fungus Pleurotus ostreatus secretes laccase isozymes with different substrete specificities. Mycologia. 95(6): 1013-1020.
Marzullo L, Cannio R, Giardina P, Santini MT, Sannia G., 1995 Veratryl alcohol oxidase from Pleurotus ostreatus
participates in lignin biodegradation and prevents polymerization of laccase-oxidized substrates. J Biol Chem. 270:3823-3827.
Mattjik AA. Sumertajaya IM. 2002.
Perancangan Percobaan. Jilid ke-1. Ed ke-2. Bogor: IPB Press.
Ohkuma M, Yoshima M, Toru J, Toshiaki K. 2001. Lignin degradation and roles of white rot fungi: study of an efficient symbiotic system in fungus-growing termitesand its application to bioremediation. RIKEN Rev. 42: 39-42.
Okwujiako IA. 1990. The effect of vitamin on the vegetative growth and fruitbody of
Pleurotus sajorcaju. J Mush. Tropics. p.10.
Palmieri G, Giardina P, Bianco C, Fontanella B, Sannia G. 1999. Copper induction of laccase isoenzymes in the ligninolytic fungus Pleurotus ostreatus. Applied and Environ Microbiol. P.920-924.
Panji TH, Tahang H, Yusuf H, Goenadi DH. 1996. Optimasi pH kadar air dan suhu pada biodelignifikasi in-vitro tandan kosong kelapa sawit. Menara Perkebunan Jurnal Penelitian Bioteknologi Perkebunan. 64(2): 79-91.
Tjitrosomo SS, Angka SL, penerjemah; Jakarta:UI Press. Terjemahan dari:
Elements of Microbiology.
Reeves JB, Schmidt WF. 1994. Solid-state fermentation 13NMR analysis of forage and product-derived fiber and lignin residues, resolution of some discrepancies among chemical, infrared, and phyrolysis gas chromatography mass spectroscopic analysis. J Agric Food Chem. 42:1462-1468.
Santosa I. 1993. Biodegradasi lignoselulosa dalam pemanfaatan limbah kelapa sawit. Menara Perkebunan. 64 (3): 80-85.
Sarkanen KV, Ludwig CH.1971. Lignin, Occurrence, Formation, Structure, and Reaction. Toronto: John Wiley & Sons. p 81-83.
Silverio CM, Viela LC, Guilatco FL, Hernandez NB. 1981. Mushroom culture on enriched composed sawdust.
J Technol. 6(4): 23-40.
Tien M, Kirk KT. 1984. Lignin degrading enzyme from Phanerochaeate
chrysosporium: purification, characterization, and catalytic properties of a unique H2O2-requiring
oxygenase. Proc Natl Acad Sci. 81: 2280-2284.
Tominaga Y. 1978. Tricholoma matsutake, In the Biology and Cultivation of Edible Mushrooms. New York: Academic Press.
Wyman CE. 2002. Potensial synergies and challenges in refining cellulosic biomass to fuels, chemicals, and power.
Biotechnol Progress 9.
Yuniawati S. 2006. Optimasi media dan inokulum jamur pelapuk putih untuk pengomposan tandan kosong kelapa sawit. Bogor: Universitas Pakuan.
AKTIVITAS ENZIM LIGNINOLITIK FUNGI PELAPUK
PUTIH
Omphalina sp
. DAN
Pleurotus ostreatus
PADA LIMBAH
LIGNOSELULOSA
RIO ICHSAN SYAFRIZAL
PROGRAM STUDI BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
AKTIVITAS ENZIM LIGNINOLITIK FUNGI PELAPUK
PUTIH
Omphalina sp
. DAN
Pleurotus ostreatus
PADA LIMBAH
LIGNOSELULOSA
RIO ICHSAN SYAFRIZAL
PROGRAM STUDI BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
RIO ICHSAN SYAFRIZAL. Aktivitas Enzim Ligninolitik Fungi Pelapuk Putih
Omphalina sp.
dan
Pleurotus ostreatus
pada Limbah Lignoselulosa. Dibimbing
oleh EDY DJAUHARI PURWAKUSUMAH, SISWANTO, dan HAPPY
WIDIASTUTI.
Fungi Pelapuk Putih (FPP)
Omphalina sp.
dan
Pleurotus ostreatus
mampu
mendegradasi lignin pada limbah lignoselulosa seperti Tandan Kosong Kelapa
Sawit (TKKS) dan bagas tebu dengan memproduksi enzim ligninolitik. Penelitian
bertujuan mengoptimumkan produksi enzim ligninolitik oleh Fungi Pelapuk Putih
(FPP) pada TKKS dan bagas tebu dengan pemberian suplemen CuSO
4, dedak, dan
kombinasi CuSO
4dan dedak, serta mengkarakterisasi pH optimum enzim
ligninolitik.
Aktivitas rata-rata tertinggi enzim lakase ditunjukkan oleh isolat
P.ostreatus
pada media TKKS, pada perlakuan kontrol sebesar 0,223 U/mL,
CuSO
40,546 U/mL, dan kombinasi CuSO
4dan dedak sebesar 0,626 U/mL,
berbeda dengan tiga perlakuan lainnya, pada perlakuan dedak aktivitas rata-rata
tertinggi enzim lakase sebesar 0,383 U/mL ditunjukkan oleh isolat
P.ostreatus
pada media bagas tebu. Aktivitas rata-rata tertinggi enzim lignin peroksidase
ditunjukkan oleh isolat
P.ostreatus
pada media TKKS, perlakuan kontrol sebesar
1,983 U/mL, perlakuan CuSO
4, dedak, dan kombinasi CuSO
4dan dedak sebesar
2,198 U/mL, 2,061 U/mL, dan 2,372 U/mL. Aktivitas rata-rata tertinggi enzim
mangan peroksidase ditunjukkan oleh isolat
P.ostreatus
pada media TKKS,
ABSTRACT
RIO ICHSAN SYAFRIZAL. Ligninolytic Enzyme Activity of White-Rot Fungus
Omphalina sp.
and
Pleurotus ostreatus
at Lignocellulose Wastes. Under the
direction of EDY DJAUHARI PURWAKUSUMAH, SISWANTO, and HAPPY
WIDIASTUTI.
White-rot
fungus
Omphalina sp.
and
Pleurotus ostreatus
were able to held
biodegradation on lignocellulose waste such as Empty Bunches of Oil Palm
(EBOP) and bagasse by produced ligninolytic enzyme. The aim of this research
were not only to optimized ligninolytic enzyme production by white-rot fungus in
EBOP and bagasse supplemented by CuSO
4, rice bran, and the mixture of CuSO
4and rice bran but also determined optimum pH of ligninolytic enzyme activity.
Judul Skripsi : Aktivitas Enzim Ligninolitik Fungi Pelapuk Putih
Omphalina sp
.
dan
Pleurotus ostreatus
pada Limbah Lignoselulosa
Nama
: Rio Ichsan Syafrizal
NIM
:
G44103023
Disetujui
Komisi Pembimbing
Drs. H. Edy Djauhari PK., M.Si.
Pembimbing Utama
Dr. Siswanto, DEA, APU
Pembimbing Anggota
Dr. Ir. Happy Widiastuti, M.Si.
Pembimbing Anggota
Diketahui
Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
Prof. Dr. Ir. Yonny Koesmaryono, M.S.
NIP 131 473 999
PRAKATA
Alhamdulillah. Penulis memanjatkan puji syukur kepada Allah SWT, Sang
Mahadaya atas berkah dan petunjuk-Nya sehingga penulis mampu menyelesaikan
penelitian ini. Penelitian yang dilakukan berjudul Aktivitas Enzim Ligninolitik
Fungi Pelapuk Putih
Omphalina sp
. dan
Pleurotus ostreatus
pada Limbah
Lignoselulosa. Penelitian dimulai bulan September 2006 hingga Mei 2007,
bertempat di Laboratorium Biologi Molekular dan Rekayasa Genetika, Balai
Penelitian Bioteknologi Perkebunan Indonesia, Jalan Taman Kencana No.1,
Bogor.
Penulis mengucapkan terima kasih terutama kepada Drs. H. Edy Djauhari
PK, M.Si. selaku pembimbing utama, Dr. Siswanto, DEA, APU dan Dr. Ir. Happy
Widiastuti, M.Si. selaku pembimbing lapangan yang telah banyak memberikan
arahan selama proses penelitian dan penyusunan skripsi. Penulis juga
mengucapkan terima kasih kepada Dr. Darmono Taniwiryono selaku Kepala Balai
Penelitian Bioteknologi Perkebunan Indonesia, Dr. Djoko Santoso, M.Sc. selaku
Ketua Kelompok Peneliti Biologi Molekular dan Rekayasa Genetika, Kak Ahmad
Topani, A.Ma. dan Mbak Riana selaku teknisi pembimbing penelitian, seluruh
peneliti, teknisi, dan staf di Laboratorium Biologi Molekular dan Rekayasa
Genetika, Balai Penelitian Bioteknologi Perkebunan Indonesia, serta seluruh
dosen dan staf Biokimia. Terima kasih kepada orang tua, saudara dan keluarga,
serta Rizki, Dwi, Adi, Ayu, Sekar, Tisha, Erlank, Hurri, Rama, Metty, dan semua
sahabat biokimia angkatan 40 atas doa, dukungan, dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2007
RIWAYAT HIDUP
Penulis dilahirkan di Manado pada tanggal 13 Januari 1985 dari ayah
Djamaluddin dan ibu Eva. Penulis merupakan anak pertama dari tiga besaudara.
Tahun 2003 penulis menyelesaikan pendidikan di SMU Negeri 5
Makassar dan pada tahun yang sama lulus seleksi masuk IPB melalui jalur
Undangan Seleksi Masuk IPB (USMI). Penulis memilih Program Studi Biokimia,
Fakultas Matematika dan Ilmu Pengetahuan Alam.
PENDAHULUAN
Kelapa sawit (Elaeis guineensis) merupakan salah satu komoditas perkebunan andalan bagi perekonomian Indonesia. Kualitas dan tingkat produksi Crude Palm Oil
(CPO) yang tinggi merupakan keunggulan dari komiditas ini. Data Ditjen Perkebunan menyebutkan pada tahun 2005 luas perkebunan kelapa sawit di Indonesia mencapai 5.358.423 hektar dengan total produksi minyak hingga 10.683.756 ton. Dalam proses produksi CPO dihasilkan banyak limbah yang masih belum dimanfaatkan secara maksimal sehungga berpotensi mencemari lingkungan. Tandan Kosong Kelapa Sawit (TKKS) merupakan limbah padat terbesar dari pabrik minyak CPO. Komponen utama TKKS adalah lignin, selulosa, dan hemiselulosa yang umum disebut lignoselulosa (Darnoko 1992).
TKKS bukan satu-satunya limbah yang tergolong lignoselulosa. Bagas tebu (ampas tebu) juga mengandung serat lignoselulosa yang sukar terdegradasi. Kandungan lignin yang tinggi pada TKKS dan bagas tebu menyebabkan kendala dalam pemanfaatan selulosa murni bagi industri yang berbahan baku selulosa seperti industri pulp dan kertas ataupun industri pembuatan papan serat. Salah satu cara untuk mengatasi masalah ini menggunakan mikroorganisme dalam rangka biodelignifikasi. Mikroorganisme yang berperan penting dalam biodelignifikasi adalah Fungi Pelapuk Putih (FPP) dari kelas
Basidiomycetes. Kemampuan FPP melakukan biodelignifikasi disebabkan oleh enzim ligninolitik ekstraselular yang dihasilkannya (Kirk & Chang 1990). Enzim ligninolitik ekstraselular tersebut adalah lakase, lignin peroksidase (LiP), dan mangan peroksidase (MnP) (Gold & Alic 1993). Produksi enzim ligninolitik dari limbah lignoselulosa mempunyai arti penting karena selain dapat mengatasi masalah limbah organik lignoselulosa, juga dapat memberi nilai tambah dari limbah tersebut.
Palmieri et al (1999) disebutkan bahwa Cu (tembaga) merupakan induser yang efisien dalam menaikkan aktivitas lakase dari
P.ostreatus. Dedak dapat digunakan sebagai substrat alami dan dapat meningkatkan pertumbuhan miselium menjadi lebih tebal dan padat (Silverio et al 1981). Dedak telah banyak digunakan sebagai induser untuk pertumbuhan miselium dalam budidaya jamur t