PRASELEKSI SIFAT RESISTENSI TANAMAN
HANDEULEUM [Graptophyllum pictum (L.) Griff.] TERHADAP
Doleschallia bisaltide Cramer. (LEPIDOPTERA:
NYMPHALIDAE)
PENI LESTARI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul Praseleksi Sifat Resistensi Tanaman Handeuleum [Graptophyllum pictum (L.) Griff.] terhadap Doleschallia bisaltide Cramer (Lepidoptera: Nymphalidae) adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk karya apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Juli 2010
ABSTRACT
PENI LESTARI. Preselection of Caricature Plant [Graptophyllum pictum (L.) Griff.] Resistance Trait to Doleschallia bisaltide Cramer (Lepidoptera: Nymphalidae). Supervised by NURUL KHUMAIDA, DEWI SARTIAMI, and GUSTAAF ADOLF WATTIMENA.
Caricature plant is one of the medicinal plants developed in Indonesia. The leaves of this plant can be used for treating hemorrhoids, cleaning blood especially after uttering and menstruation, rheumatism, and preventing teeth plaque. The biggest problem in caricature plant development is Doleschallia bisaltide (Lepidoptera: Nymphalidae) attack. Resistance variety is one of the ways to solve this problem. Generally, plants have resistance mechanism to prevent herbivore attack. Therefore, it is needed to study caricature plant-D. bisaltide interaction to find varietal control for this insect. The objective of this research is to do early selection on 13 accessions of caricature plant for resistance to D. bisaltide. Moreover, we also interest to identify resistance level of 13 caricature plant accessions to D.bisaltide. This research was divided into three studies, including a) morphology and phytochemical identification on thirteen accessions of caricature plant; b) oviposition preference of adult D. bisaltide on thirteen accessions of caricature plant; and c) D. bisaltide larvae consumption activity preference of on thirteen accessions of caricature plant. All studies were done at Indonesian for Medicinal and Aromatic Crops Research Institute (BALITTRO) from May 2009 until February 2010. Thirteen accessions of caricature plant were come from exploration result of Khumaida et.al in year 2008. Based on qualitative analysis, there were changes in each kind of secondary metabolites after D.bisaltide larvae feeding, there were no change in tannin and alkaloid contains, but there decreases in flavonoid, glycoside, and steroid. It was found increasing of triterpenoid and saponin contain after attack caricature plant leaves. Moreover, there was an increased in calcium and C/N ratio on almost caricature plant accessions. Caricature plant colors tend to influenced adult oviposition preference. Based on all observation, it can be concluded that accession 12 was resistant to D. bisaltide attack with antixenosis mechanism. Accession 1, 2, 5, and 9 were susceptible to D. bisaltide. Chlorophyll might be used as selection character. By this research, we haven’t identified about antibiosis mechanism yet.
RINGKASAN PENELITIAN
PENI LESTARI. Praseleksi Sifat Resistensi Tanaman Handeuleum [Graptophyllum pictum (L.) Griff.] Terhadap Doleschallia bisaltide Cramer (Lepidoptera: Nymphalidae). Di bimbing oleh NURUL KHUMAIDA, DEWI SARTIAMI, dan GUSTAAF ADOLF WATTIMENA.
Tanaman handeuleum merupakan salah satu tanaman obat yang dikembangkan di Indonesia. Daun tanaman ini dapat digunakan untuk mengobati wasir, memperlancar pembuangan urin, membersihkan darah, rematik, dan mencegah pembentukan plak pada gigi. Masalah utama dalam pengembangan tanaman ini adalah serangan larva Doleschallia bisaltide (Lepidoptera :Nymphalidae). Varietas resisten merupakan salah satu pertahanan garis depan dalam menangkal serangan hama. Secara umum, tanaman, termasuk handeuleum, memiliki mekanisme resistensi untuk mencegah serangan herbivora. Karena itu, diperlukan kajian mengenai interaksi handeuleum-D.bisaltide untuk menentukan aplikasi pengendalian hama secara biologi terhadap serangga ini. Tujuan penelitian ini adalah untuk melakukan seleksi awal sifat resistensi pada 13 aksesi tanaman handeuleum terhadap serangan D. bisaltide. Lebih lanjut, kami juga tertarik untuk melakukan identifikasi awal mengenai tingkat dan mekanisme resistensi 13 aksesi handeuleum terhadap D. bisaltide.
Penelitian ini terbagi atas tiga subpenelitian, yakni a) identifikasi karakter morfologi dan kandungan fitokimia 13 aksesi handeuleum; b) studi preferensi peletakan telur oleh imago betina D. bisaltide pada 13 aksesi handeuleum; dan c) studi aktivitas makan larva D. bisaltide pada 13 aksesi handeuleum. Semua subpenelitian dilakukan di Balai Penelitian Tanaman Obat dan Aromatik (BALITTRO) sejak Mei 2008 hingga Februari 2010. Tiga belas aksesi tanaman handeuleum merupakan hasil eksplorasi Khumaida et al. pada tahun 2008.
Berdasarkan hasil analisis kualitatif, terdapat perbedaan kombinasi konsentrasi masing-masing jenis metabolit sekunder setelah terserang larva D.bisaltide, diantaranya tidak terdapat perubahan konsentrasi pada tanin dan alkaloid, penurunan pada flavonoid, glikosida, dan steroid. Pada penelitian ini juga terdapat indikasi adanya peningkatan konsentrasi triterpenoid dan saponin setelah tanaman terserang larva. Lebih lanjut, ditemukan adanya peningkatan penyerapan kalsium dan C/N rasio pada daun hampir semua aksesi handeuleum yang diujikan. Warna daun tanaman handeuleum diduga berpengaruh terhadap preferensi peletakan telur imago D. bisaltide.
handeuleum memiliki mekanisme antibiosis. Berdasarkan percobaan aktivitas peletakan telur dan aktivitas makan, dapat disimpulkan bahwa aksesi 12 merupakan aksesi resisten terhadap D. bisaltide dengan mekanisme antixenosis. Aksesi 1, 2, 5, dan 9 merupakan aksesi rentan.
Klorofil dapat dijadikan kandidat karakter seleksi untuk sifat resistensi tanaman handeuleum terhadap D. bisaltide. Analisis korelasi menunjukkan bahwa klorofil tidak berkorelasi dengan flavonoid, namun senyawa ini berkorelasi erat dengan salah satu senyawa golongan flavonoid, yakni antosianin. Karenanya, untuk mencegah kehilangan aksesi-aksesi dengan kandungan flavonoid tinggi akibat seleksi pada karakter klorofil, maka tipe seleksi yang disarankan adalah seleksi indeks atau seleksi independent cooling. Seleksi dapat dilakukan pada generasi lanjut.
© Hak Cipta Milik IPB, tahun 2010 Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan sebagian pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
PRASELEKSI SIFAT RESISTENSI TANAMAN
HANDEULEUM [Graptophyllum pictum (L.) Griff.] TERHADAP
Doleschallia bisaltide (LEPIDOPTERA: NYMPHALIDAE)
PENI LESTARI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains Pada Mayor Pemuliaan dan Bioteknologi Tanaman
Departemen Agronomi dan Hortikultura
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dosen Penguji dari Luar Komisi Pembimbing:
Judul Tesis : Praseleksi Sifat Resistensi Tanaman Handeuleum [Graptophyllum pictum (L.) Griff.] terhadap Doleschallia bisaltide Cramer.
(Lepidoptera: Nymphalidae) Nama : Peni Lestari
NRP : A253080201
Disetujui Komisi Pembimbing
Dr. Ir. Nurul Khumaida, M.Si. Ketua
Dra. Dewi Sartiami, M.Si. Prof. Dr. Ir. Gustaaf Adolf Wattimena, M.Sc.
Anggota Anggota
Diketahui
Koordintor Mayor Dekan Sekolah Pascasarjana Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, M.Sc. Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
Kupersembahkan karya ini untuk para motivator yang selalu menemaniku melangkah, manjalani hidup
PRAKATA
Alhamdulillah, puji syukur penulis panjatkan ke hadirat Allah SWT atas segala karunia-Nya, sehingga penyusunan tesis ini berhasil diselesaikan. Tesis ini berjudul Praseleksi Sifat Resistensi Tanaman Handeuleum [Graptophyllum pictum (L.) Griff.] terhadap Doleschallia bisaltide Cramer. (Lepidoptera: Nymphalidae).
Terima kasih penulis sampaikan kepada Dr. Ir. Nurul Khumaida, M.Si, Dra. Dewi Sartiami, M.Si, dan Prof. Dr. Ir. Gustaaf Adolf Wattimena, M.Sc. atas bantuan serta bimbingannya selama penelitian dan penyusunan tesis. Kepada program KKP3T atas fasilitas yang diberikan selama penelitian berlangsung. Kepada bapak Cucu Sukmana, ibu Tri Lestari Mardiningsih dan staf kebun BALITTRO, penulis sangat menghargai bantuannya selama pelaksanaan penelitian. Penulis juga menyampaikan ucapan terima kasih yang tak terhingga kepada Bruce Ochieng Obura dan Wage Ratna Rohaeni yang selalu memberi inspirasi dan motivasi terus menerus, Sumiyati, Tri Handayani, dan rekan-rekan di Pondok Rizki untuk bantuan dan semangat yang tak kunjung berhenti diberikan. Kepada Umarul Anshori Bin Asyar sebagai kakak dan teman yang baik, juga kepada teman-teman di Mayor Pemuliaan dan Bioteknologi Tanaman 2008 atas kerjasamanya selama ini. Ungkapan terimakasih juga disampaikan kepada ayah, ibu, adik, kakak beserta seluruh keluarga atas restu, doa, dan kasih sayang yang terus mengalir.
Semoga karya ilmiah ini bermanfaat untuk banyak pihak.
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta Selatan tanggal 26 November 1985 dari ayah Sunarmanto dan ibu Aida Purpriyani. Penulis merupakan anak keempat dari tujuh bersaudara. Penulis banyak menghabiskan masa kecil dan remaja di daerah Depok, Jawa Barat dan DKI Jakarta. Pendidikan sarjana ditempuh di Program Studi Hortikultura, Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor melalui jalur Undangan Seleksi Masuk (USMI) IPB, dan lulus tahun 2008. Selama menempuh jenjang S1, sebagian biaya pendidikan penulis berasal dari Goodwill International Foundation dan Beasiswa BPPOM IPB. Pada tahun yang sama, penulis sempat bekerja sebagai Breeder Assistant di PT. EASTWEST SEED INDONESIA selama 3 bulan sebelum melanjutkan studi ke jenjang pascasarjana.
DAFTAR TABEL ... xiii
2.2 Khasiat Handeuleum Sebagai Tanaman Obat ... 8
2.3 Karakteristik Doleschallia bisaltide ... 9
2.4 Interaksi Tanaman dengan Serangga ... 12
2.5 Mekanisme Resistensi Tanaman Terhadap Hama ... 14
2.5.1 Resistensi Langsung ... 15
2.5.2 Resistensi Tidak Langsung ... 16
2.6 Metabolit sekunder Tanaman yang Berhubungan dengan Resistensi 16
III IDENTIFIKASI KARAKTER MORFOLOGI DAN KANDUNGAN FITOKIMIA TIGA BELAS AKSESI HANDEULEUM
3.5.4 Pengamatan Karakter Morfologi Daun ... 23
3.5.5 Analisa Kandungan Fitokimia Daun Sebelum dan Setelah Terserang Larva D. bisaltide ... 24
3.5.6 Analisa Data ... 25
3.6 Hasil dan Pembahasan ... 26
3.6.1 Karakter Morfologi ... 26
Halaman
3.6.3 Korelasi antar karakter morfologi dan fitokimia tanaman ... 41
3.7 Kesimpulan ... 44
IV STUDI PREFERENSI PELETAKAN TELUR OLEH IMAGO BETINA D. bisaltide PADA TIGA BELAS AKSESI HANDEULEUM
4.5.5 Uji preferensi peletakan telur imago D. bisaltide ... 50
4.5.6 Analisis data ... 50
4.6 Hasil dan Pembahasan ... 51
4.6.1 Aktivitas peletakan telur D. bisaltide ... 51
4.6.2 Analisis hubungan pengaruh langsung dan tidak langsung karakteristik handeuleum dengan aktivitas peletakan telur D. bisaltide ... 57
4.7 Kesimpulan ... 62
V STUDI AKTIVITAS MAKAN LARVA D. bisaltide PADA TIGA BELAS AKSESI HANDEULEUM
5.5.5 Uji Aktivitas Makan Larva D. bisaltide pada 13 Aksesi Tanaman Handeuleum ... 67
5.5.6 Analisa Data ... 68
5.6 Hasil dan Pembahasan ... 69
5.6.1 Aktivitas Makan Larva D. bisaltide ... 69
5.6.2 Analisis hubungan pengaruh langsung dan tidak langsung karakteristik handeuleum dengan aktivitas makan D. bisaltide ... 73
Halaman
VI PEMBAHASAN UMUM ... 78
VII KESIMPULAN 7.1 Kesimpulan ... 84
7.2 Saran ... 84
VIII DAFTAR PUSTAKA ... 86
DAFTAR TABEL
Halaman 3.1 Asal dan deskripsi 13 aksesi handeuleum yang diuji ... 23 3.2 Analisis sidik ragam ... 25 3.3 Luas area daun total dan kerapatan trikoma 13 aksesi handeuleum . 28 3.4 Kriteria heritabilitas ... 28 3.5 Rekapitulasi sidik ragam, ragam genetik, ragam fenotipe, ragam
galat, serta nilai heritabilitas karakter luas area daun total dan
kerapatan trikoma tanaman handeuleum 29
3.6 Kandungan metabolit sekunder daun tanaman handeuleum
sebelum dan setelah terserang larva D. bisaltide ... 30 3.7 Persentase perubahan kandungan metabolit sekunder pada daun
13 aksesi handeuleum ... 34 3.8 Perbandingan kandungan pigmen 13 aksesi handeuleum sebelum dan
setelah terserang larva D. bisaltide ... 35 3.9 Persentase pertambahan konsentrasi fitokimia 13 aksesi
handeuleum ... 40 3.10 Korelasi antar karakter morfologi dan fitokimia pada tanaman
handeuleum ... 42 4.1 Jumlah telur yang diletakkan D. bisaltide pada 13 aksesi handeuleum
dalam tiga periode pengujian ... 52 4.2 Persentase jumlah telur yang diletakkan D. bisaltide pada 13
aksesi handeuleum dalam tiga periode pengujian ... 53 4.3 Persentase pigmen antosianin, klorofil, dan karotenoid pada 13 aksesi
tanaman handeuleum ... 55 4.4 Korelasi antar karakter morfologi dan fitokimia daun
handeuleum dengan jumlah telur yang diletakkan ... 58 4.5 Pengaruh langsung dan pengaruh tidak langsung karakter terhadap
peletakan telur imago D. bisaltide ... 60 5.1 Pengelompokan kriteria resistensi tanaman berdasarkan
Halaman 5.2 Luas area konsumsi setiap fase instar D. bisaltide pada 13 aksesi
tanaman handeuleum ... 70 5.3 Luas area makan, kriteria kerusakan relatif, dan kategori tingkat
resistensi 13 aksesi handeuleum terhadap larva D. bisaltide ... 71 5.4 Korelasi luas area konsumsi D. Bisaltide dengan
kandungan fitokimia daun 13 aksesi tanaman handeuleum ... 73 5.5 Pengaruh langsung dan tidak langsung peubah terhadap
DAFTAR GAMBAR
Halaman
1.1 Bagan Alir Penelitian ... 5
2.1 Keragaan hadeuleum ... 7
2.2 Doleschallia bisaltide ... 9
2.3 Telur dan larva D. bisaltide ... 10
2.4 Gejala serangan larva D. bisaltide pada tanaman handeuleum ... 11
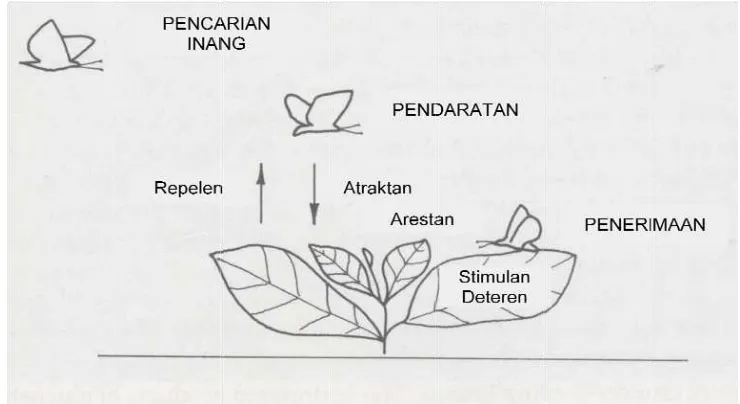
2.5 Proses penerimaan serangga terhadap tanaman inang ... 16
3.1 Keragaan trikoma 13 aksesi handeuleum ... 27
3.2 Grafik persentase perubahan kandungan antosianin, klorofil, dan karotenoid pada 13 aksesi tanaman handeuleum setelah tanaman terserang larva D. bisaltide ... 35
3.3 Grafik perbandingan kandungan C/N rasio pada daun 13 aksesi handeuleum sebelum dan setelah terserang D. bisaltide ... 37
3.4 Besarnya perubahan kandungan unsur C dan N pada tanaman handeuleum setelah terserang larva D. bisaltide ... 38
3.5 Grafik perbandingan kandungan kalsium pada daun 13 aksesi handeuleum sebelum dan setelah terserang larva D. bisaltide ... 38
3.6 Grafik perbandingan kandungan serat pada daun 13 aksesi handeuleum sebelum dan setelah terserang larva D. bisaltide ... 39
3.7 Lintasan biosintesis metabolit sekunder ... 43
4.1 Morfologi daun tanaman handeuleum ... 54
4.2 Diagram lintas fenotipik antar 4 karakter dengan aktivitas peletakan telur oleh imago D. bisaltide ... 60
5.1 Lama hidup setiap stadia instar larva D. bisaltide ... 72
5.2 Diagram lintas fenotipik anar 3 karakter dengan aktivitas makan larva D. bisaltide ... 74
DAFTAR LAMPIRAN
Halaman 1 Formulasi obat berbahan handeuleum ... 99 2 Kurva kenormalan kerapatan trikoma tanaman handeuleum ... 103 3 Kurva kenormalan karakter luas area daun total berdasarkan
Saphiro-Wilk ... 103 4 Korelasi antar karakter morfologi dan fitokimia pada tanaman
handeuleum dengan aktivitas peletakan telur dan aktivitas makan D.
1.1 Latar Balakang
Indonesia adalah negara tropis dengan biodiversitas terbesar kedua di dunia setelah negara Brazil. Kekayaan ini tidak hanya dari segi jumlah, tetapi juga dari tingkat keragaman jenis flora yang tumbuh di hampir seluruh wilayah Indonesia. Di antara sejumlah keragaman hayati tersebut, cukup banyak spesies tumbuhan dan atau tanaman yang dimanfaatkan masyarakat sebagai bahan baku obat herbal. Tercatat terdapat 1300 spesies tanaman obat yang telah digunakan di Indonesia. Sembilan puluh enam spesies di antaranya digunakan suku Sunda untuk pengobatan (Roosita et al. 2008). Banyaknya keragaman tanaman obat yang ada di Indonesia telah diketahui masyarakat dunia, di antaranya negara Jepang. Subagyo (2001) menyatakan bahwa sebagian resep obat herbal yang dipatenkan pemerintah Jepang merupakan resep obat tradisional Indonesia. Bahan baku resep tersebut juga merupakan tanaman Indonesia. Hal ini tentu patut menjadi bahan perenungan bangsa Indonesia agar kekayaan alam yang dimiliki bangsa ini tidak berpindah tangan ke negara lain.
Obat herbal tidak hanya digunakan masyarakat Indonesia, tetapi juga masyarakat dunia. Hal ini tercermin dari data penjualan obat herbal di pasaran dunia. Tercatat total nilai dagang di pasaran internasional untuk obat berbahan baku alami mencapai 5 triliun Dollar AS pada tahun 2005. Nilai ini diperkirakan akan terus mengalami peningkatan. Di lain pihak, omset penjualan Indonesia baru mencapai 100 juta Dollar AS per tahun (Kintoko 2006). Mengacu pada hal tersebut, terbuka peluang bagi Indonesia untuk meningkatkan ekspor di bidang ini. Seiring dengan meningkatnya tuntutan konsumen tentang keamanan obat herbal, saat ini kandungan suatu tanaman obat harus dianalisis sehingga dapat dipertanggungjawabkan khasiatnya secara ilmiah. Hal ini mengacu pada standar keamanan obat berbahan alami yang dikeluarkan oleh Badan Pengawas Obat dan Makanan (BPOM).
diketahui mengandung flavonoid, alkaloid, steroid, saponin, glikosida, serta emolien dan lendir (Willaman 1995; Isnawati 2003). Kondisi ini menunjukkan bahwa handeuleum memiliki potensi yang cukup besar untuk dibudidayakan secara komersial dan dalam skala besar sebagai komoditas fitofarmaka. Selama ini, handeuleum telah digunakan secara luas sebagai bahan obat wasir, melancarkan air seni, membersihkan darah menstruasi dan nifas, rematik, bisul, dan penyakit lainnya (Wibowo 2000).
Telah banyak perusahaan jamu berskala nasional yang memanfaatkan bagian tanaman handeuleum sebagai bahan baku obat racikan mereka, di antaranya PT Fimedco Surabaya dan PT Sidomuncul Semarang (Wibowo 2000). Pusat Studi Biofarmaka IPB (2008) menambahkan satu lagi perusahaan, yaitu PT. Indo Farma. Ketiga perusahaan ini menggunakan handeuleum sebagai salah satu bahan untuk mengobati wasir. PT Sidomuncul Semarang dan PT Indo Farma memerlukan 1-2 ton daun handeuleum/bulan (Pusat Studi Biofarmaka IPB 2008). Selain formula yang digunakan kedua perusahaan tersebut, terdapat beberapa formula yang dikembangkan menggunakan daun handeuleum (Lampiran 1).
1.2 Tujuan
Secara umum penelitian ini bertujuan untuk melakukan seleksi awal untuk sifat resistensi beberapa aksesi handeuleum terhadap D. bisaltide. Penelitian ini juga bertujuan mengidentifikasi mekanisme resistensi tanaman handeuleum terhadap serangan D. bisaltide.
1.3 Hipotesis
1. Terdapat aksesi handeuleum yang resisten terhadap D. bisaltide.
2. Terdapat informasi mengenai mekanisme resistensi 13 aksesi handeuleum terhadap serangan D. bisaltide.
1.4 Manfaat Penelitian
Penelitian ini memberikan informasi tentang mekanisme dan tingkat resistensi 13 aksesi tanaman handeuleum. Informasi mengenai tingkat resistensi dan kandungan fitokimia pada handeuleum dapat digunakan sebagai sumber genetik pada program perakitan varietas. Hasil penelitian ini juga dapat digunakan sebagai salah satu program pengendalian hama secara terpadu.
1.5
Kerangka PemikiranPenggunaan insektisida untuk mengendalikan serangga ini bukanlah langkah yang tepat, karena kekhawatiran adanya residu insektisida pada daun. Sejauh ini musuh alami D. bisaltide belum diteliti, sehingga salah satu cara yang mungkin ditempuh untuk menanggulangi serangan hama adalah dengan membudidayakan tanaman yang resisten terhadap D. bisaltide. Untuk keperluan tersebut, dibutuhkan perakitan varietas yang resisten terhadap serangan hama tersebut.
Gam
b
aar 1.1 Ba
g
an A
llir Pene
liti
TINJAUAN PUSTAKA
2.1 Tanaman Handeuleum
Selain dikenal dengan nama handeuleum, Graptophyllum pictum (L.) Griff. (Acanthaceae) juga sering disebut daun ungu, demung, tulak, wungu (Jawa); daun temen-temen (Sunda), temen (Bali); karotong (Madura), daun putri, dongora (Ambon); dan kobi-kobi (Ternate) (Wibowo 2000). Tanaman ini termasuk perdu yang memiliki batang tegak berukuran kecil dengan tinggi tanaman mencapai 3 m. Daun handeuleum memiliki panjang 10-15 cm dengan bentuk bulat telur memanjang atau elips. Daun handeuleum juga memiliki permukaan yang tipis dan mengkilap serta bagian tepi daun yang cenderung rata. Duduk daun berhadapan-bersilangan. Batang handeuleum berbentuk segitiga tumpul berwarna ungu atau ungu kehijauan, dan berkayu ketika tua. Ciri khas handeuleum adalah kandungan senyawa vomifoliol pada daunnya (BPOM 2004; Kristina dan Mardiningsih 2008).
Bunga handeuleum berbentuk tabung berwarna ungu kemerahan. Bunga tersebut tersusun pada tandan bunga yang terdapat pada bagian terminal tanaman (Sjamsudihajat et al. 1991). Buah handeuleum berbentuk kotak, lonjong, berwarna ungu kecokelatan, sedangkan bijinya berbentuk bulat dan berwarna putih (Anonim 2008). Taksonomi tanaman handeuleum adalah sebagai berikut: Kingdom : Plantae
Subkingdom : Tracheobionta Superdivisio : Spermatophyta Divisio : Magnoliophyta Class : Magnoliopsida Subclass : Asteridae
Ordo : Scrophulariales Family : Acanthaceae Genus : Graptophyllum
Berdasarkan hasil observasi yang dilakukan Khumaida et al. (2008) terhadap masyarakat Papua, Kepulauan Maluku, dan Jawa Barat, tanaman ini ditanam di pekarangan sebagai tanaman obat. Keragaan tanaman handeuleum ditampilkan pada Gambar 2.1.
Gambar 2.1 Keragaan handeuleum. Daun (kiri) dan bunga (kanan) aksesi 1. (Sumber: Novita 2008)
Handeuleum dapat tumbuh di tempat terbuka, beriklim kering dan lembab (Kristina dan Mardiningsih 2008). Meskipun tanaman ini termasuk golongan xeriscape (Wong 2008), hasil penelitian Sakya (2006) menunjukkan bahwa pemberian air 30% di bawah kapasitas lapang dapat menurunkan pertumbuhannya. Selain air, Djazuli dan Fathan (2000) melaporkan bahwa produktivitas tanaman tertinggi dicapai bila tanaman handeuleum diberi pupuk kandang dan dipangkas pada umur 2-4 Bulan Setelah Tanam (BST). Tanaman handeuleum umumnya dikembangbiakkan menggunakan stek batang karena buah dan biji sulit terbentuk. Wibowo (2000) menyebutkan bahwa handeuleum mampu hidup pada ketinggian di bawah 800 mdpl. Semakin tinggi dataran tersebut, semakin ungu warna daun handeuleum. Hal ini dapat terjadi karena adanya peningkatan senyawa flavonoid yang dikandungnya.
(2000) hama yang menyerang handeuleum meliputi Coccus Sp., Pseudococcus longispinus, Planococcus lilacinus, Valanga Sp., dan D. bisaltide. Serangga Doleschallia bisaltidae memiliki tingkat penyerangan paling tinggi dibandingkan kelima jenis serangga lainnya. Penurunan kualitas dan kuantitas hasil akibat serangan hama ini dapat mencapai 70%. Oleh karena itu, pengetahuan tentang cara mengelola hama tersebut diperlukan untuk menghindari penurunan kualitas dan kuantitas hasil yang terjadi.
2.2 Khasiat Handeuleum Sebagai Tanaman Obat
Handeuleum diketahui memiliki kandungan unsur kalsium pada bagian batang, Kalium, Natrium, Magnesium, minyak atsiri, tanin, alkaloid nontoksik, asam folat, lemak, fenol, polifenol, saponin, flavonoid, steroid, glikosida dan lendir, serta emolien (Perry 1980; Willaman 1995). Kandungan flavonoid pada tanaman handeuleum merupakan zat yang memiliki efek katartik dan dapat mematikan bakteri , sedangkan senyawa steroid berperan sebagai antiinflamasi (Willaman 1995). Selain steroid, Wiart (2006) menyatakan kandungan glikosida pada tanaman juga dapat berefek antiinflamasi.
Berdasarkan pengujian pada Pithyum ultinum, Rhizoctonia solani, Sclerotium rolsfii, dan Phytophtora parasitica di laboratorium, Goun et al. (2003) menyimpulkan bahwa ekstrak tanaman handeuleum dapat berfungsi sebagai anticendawan. Lebih lanjut Goun et al. (2003) menyatakan bahwa ekstrak G. pictum 100 mg/ml juga dapat menurunkan pertumbuhan Xanthomonas campestris. Nendra (2008) melaporkan bahwa handeuleum juga dapat menghambat pertumbuhan bakteri Streptococcus mutans. Aktivitas bakteri ini akan menyebabkan plak pada gigi sehingga handeuleum juga dapat berfungsi sebagai penghambat pembentukan plak gigi. Berdasarkan hasil penelitian Isnawati (2003), tanaman handeuleum juga diketahui mengandung antosianin dan leukoantosianin. Pemeriksaan lebih lanjut menggunakan UV iluminator diketahui bahwa handeuleum juga mengandung asam protokatekuat, flavon, dan flavonol (3-hidroksi tersubtitusi).
batu ampedu, dan batuk darah. Hasil penelitian Mu’minah (2007) menyebutkan bahwa ekstrak etanol pada daun handeuleum dapat menurunkan kadar total lipid dan kolesterol LDL dalam serum darah. Selain menurunkan kolesterol, ekstrak etanol yang diaplikasikan pada mencit juga menurunkan berat badan. Selain bagian daun, bunga handeuleum juga dapat digunakan untuk mengatasi gangguan menstruasi. Sumastuti (2000) menyatakan bahwa tanaman ini diduga mengandung senyawa yang dapat merangsang pergerakan dinding usus. Saat ini, handeuleum telah terdaftar di BPOM sebagai salah satu bahan baku obat herbal. Pengakuan handeuleum sebagai tanaman obat juga terlihat dari adanya tanaman ini sebagai salah satu tanaman obat yang dipamerkan pada ulang tahun Taman Lindung Cibodas ke-156 (Darmawan 2008).
2.3 Krakteristik Doleschallia bisaltide
Serangga Doleschallia bisaltide termasuk ke dalam ordo Lepidoptera famili Nymphalidae (Braby 2000; Darrow 2007). Deskripsi James Cook University (2009) menyebutkan imago dari serangga ini berukuran kecil sampai sedang. Sebagian besar tubuhnya tertutup sisik halus. Imago D. bisaltide memiliki sayap berwarna cokelat keemasan dengan warna cokelat tua kehitaman yang terletak di bagian ujung atas sayap (Gambar 2.2 a). Berdasarkan pengamatan, imago jantan memiliki warna sayap bagian dalam yang lebih atraktif dibandingkan imago betina (Gambar 2.2b).
tanaman berupa bekas gerigitan pada daun tanaman tersebut. Serangan umumnya berawal pada bagian pucuk tanaman, kemudian meluas ke daun yang lebih tua. Larva D. bisaltide termasuk defoliator, sehingga serangannya dapat menghabiskan seluruh daun tanaman. Gejala serangan larva D. bisaltide ditampilkan pada Gambar 2.4.
Gambar 2.4 Gejala serangan larva Doleschallia bisaltide pada tanaman handeuleum. (A) daun, (B) pucuk, (C) tajuk
2.4 Interaksi Tanaman dengan Serangga
Interaksi antara tanaman dengan herbivora mendominasi lingkungan terestrial di bumi. Interaksi antara keduanya tidak selalu merugikan. Beberapa serangga berperan sebagai penyerbuk, terutama imago ordo Lepidoptera dan imago Hymenoptera dari famili Apidae. Akan tetapi, sebagian besar herbivora umumnya menimbulkan kerugian pada tanaman karena menyebabkan penurunan produktivitas tanaman. Hal ini terjadi akibat hilangnya bagian tanaman karena dikonsumsi serangga.
Dalam interaksi antara tanaman dan serangga, dikenal serangga yang bersifat monofag, oligofag, dan polifag (Schoonhoven et al. 1998; Schoonhoven et al. 2005). Price (1997) menyatakan bahwa suatu serangga bersifat monofag karena dalam genomnya tidak menghasilkan kombinasi enzim yang dapat mematahkan mekanisme pertahanan tanaman yang bukan inangnya. Serangga ini umumnya memproduksi sejumlah besar enzim untuk menetralkan racun pada sumber makanan mereka. Smith (1989) dan Price (1997) menyatakan serangga polifag umumnya memproduksi enzim nonspesifik yang dikenal dengan nama microsomal mixed-function oxidaces (MFO) untuk menyesuaikan diri dengan banyak senyawa yang berpotensi menjadi sumber makanan mereka. Jermy (1984) mengklasifikasikan monofag dan oligofag dalam golongan spesialis, sedangkan polifag digolongkan ke dalam generalis.
Salah satu contoh serangga dengan sifat makan spesialis adalah D. bisaltide. Interaksi tanaman-herbivora, seperti yang terjadi pada handeuleum dengan D. bisaltide dapat terjadi akibat kombinasi antara faktor morfologi tanaman, kimia, dan fenologi (Chew dan Rodman 1979). Beberapa hasil penelitian menunjukkan terdapat faktor-faktor yang mempengaruhi imago dalam memilih inang untuk meletakkan telur. Faktor tersebut di antaranya kerapatan tanaman inang pada suatu populasi, keberadaan nektar sebagai bahan makanan imago (Wiklund dan Hrberg 1978; Murphy et al. 1984; Brommer dan Fred 1999), jenis dan konsentrasi senyawa kimia (Mitchell dan Heath 1987; Pereyra dan Bowers 1988), seperti konsentrasi nitrogen (Wolfson 1980; Letoumeau dan Fox 1989).
2008). Price (1997) menyatakan bahwa kontak kemoreseptor antara serangga dan tanaman inangnya merupakan penciri utama suatu serangga untuk menentukan tanaman inang yang cocok baginya. Rosenthal dan Janzen (1979) menyatakan bahwa serangga juga memilih tanaman inang berdasarkan adanya senyawa yang diperlukan serangga untuk melangsungkan hidupnya serta rendahnya kadar senyawa toksik pada tanaman tersebut.
Selain penanda kimia, bentuk dan warna daun suatu tanaman juga dapat menarik perhatian imago untuk meletakkan telur mereka pada tanaman tersebut (Gilbert 1975; Rausher 1978; Stanton 1982). Hasil penelitian Kristina dan Mardiningsih (2008) menunjukkan bahwa imago D. bisaltide memilih warna daun untuk meletakkan telurnya. Serangga ini lebih menyukai jenis handeuleum yang berwarna ungu dibandingkan yang berwarna hijau. Lebih lanjut Kristina dan Mardiningsih (2008) menyatakan bahwa warna ungu pada daun handeuleum disebabkan oleh tingginya kandungan flavonoid. Panda dan Kush (1995) menyatakan bahwa flavonoid dapat menarik serangga. Wink (2006) mengemukakan bahwa flavonoid, senyawa volatil, atau terpenoid dapat menarik serangga untuk melakukan polinasi atau peletakan telur pada tanaman tersebut. Hal ini mengindikasikan bahwa aksesi handeuleum yang berwarna lebih ungu, dengan asumsi kandungan flavonoid yang tinggi, akan lebih rentan terhadap serangan D. bisaltide dibandingkan aksesi yang sedikit mengandung senyawa flavonoid. Lev-Yadun et al. (2004) menyatakan bahwa serangga akan memilih tanaman yang cocok dengan warna tubuhnya, sehingga dapat membantu kamuflase. Hal ini bermanfaat sebagai tempat berlindung dari musuh alaminya.
al. (1989) menyatakan bahwa peranan metabolit sekunder dalam interaksi tanaman inang dengan serangga fitofag juga bergantung pada fisiologi dan populasi serangga. Selain menggunakan metabolit sekunder, terdapat beberapa mekanisme lain yang digunakan tanaman untuk mempertahankan diri, di antaranya morfologi dan keragaman genetik tanaman.
Wahlberg et al. (2005) mengemukakan bahwa geografi mempengaruhi fenotipe dan fisiologi tanaman dan herbivora. Inang di tempat tertentu akan memiliki jenis hama tertentu yang berbeda dengan daerah lainnya, walaupun mereka satu spesies. Janz et al. (2006) mengemukakan, semakin beragam genetik suatu spesies tumbuhan, semakin beragam pula spesies serangga yang menyerang tumbuhan tersebut. Hal yang sama juga berlaku pada handeuleum sehingga varietas handeuleum yang memiliki tingkat resistensi yang lebih tinggi dapat dirakit dengan memanfaatkan keragaman genetik dari setiap aksesi handeuleum dari tempat yang berbeda. Selain usaha tanaman untuk mengurangi serangan herbivora, lingkungan juga memiliki pengaruh yang besar terhadap serangan herbivora pada tanaman. Lingkungan dengan kadar CO2 yang tinggi dalam udara
dapat meningkatkan serangan hama (Srivastava, et al. 2002).
2.5 Mekanisme Resistensi Tanaman Terhadap Hama
2.5.1 Resistensi langsung
Resistensi langsung merupakan sistem resistensi yang langsung berpengaruh terhadap herbivora. Sistem resistensi ini dilakukan tanaman dengan memproduksi senyawa yang dapat mengganggu pencernaan harbivora. Sifat resistensi langsung dapat berupa morfologi, seperti lapisan lilin, duri, atau trikoma sehingga mempersulit herbivora untuk mengkonsumsi bagian tertentu. Metode lainnya adalah dengan adanya getah pada batang atau daun sehingga hama terperangkap di dalamnya (Price 1997).
Mekanisme resistensi lainnya adalah dengan memproduksi metabolit sekunder. Umumnya senyawa metabolit sekunder nonvolatil bersifat deterrent bagi serangga. Produksi senyawa tersebut dapat bersifat konstitutif, namun umumnya bersifat inducible, sehingga agar senyawa tersebut diproduksi diperlukan elicitor, seperti saliva herbivora (D’Auria et al. 2002). Berdasarkan penelitian, saliva herbivora banyak mengandung senyawa golongan konjugat asam lemak dan asam amino. Salah satu komponen saliva herbivora adalah glucose oksidase. Glucose oksidase berperan mengubah gula glukosa sederhana menjadi asam gluconat dan hidrogen peroksida (www.farmingfirst.org 2002; GMO Compass 2008). Senyawa ini memberikan informasi kepada gen untuk mensintesis asam jasmonat. Asam jasmonat merupakan penanda bagi gen dalam genom tanaman untuk mensintesis senyawa yang dapat menghidrolisis glucose oxidase, sehingga tanaman dapat bertahan dari serangan herbivora.
Whittaker dan Feeny (1971) menyatakan bahwa metabolit sekunder dapat digunakan tanaman untuk mempertahankan diri dari serangan herbivora atau kompetitornya. Lebih jauh mereka memasukkan senyawa tersebut ke dalam golongan allelopati. Jermy (1984), begitu juga Panda dan Kush (1995) menjelaskan bahwa adanya senyawa metabolit sekunder tertentu dapat bersifat seperti sidik jari bagi serangga untuk menemukan inangnya. Arbain (1995) menyatakan senyawa ini dapat mempengaruhi tingkah laku, pertumbuhan, dan reproduksi serangga. Akan tetapi, Price (1997) menyatakan senyawa allelopati hanya akan diproduksi tanaman bila tanaman tersebut telah tumbuh dengan baik. Beberapa jenis metabolit sekunder yang dikeluarkan tanaman untuk mempertahankan diri antara lain tanin, alkaloid, fenolik, terpenoid, dan flavonoid.
Pada tanaman, umumnya tanin diproduksi pada daun yang telah berkembang sempurna (Janzen 1979). Hal ini dikarenakan, produksi tanin pada pucuk dapat mempengaruhi pertumbuhan bagian tersebut. Kemungkinan hal inilah yang menyebabkan imago serangga meletakkan telur pada bagian pucuk tanaman. Larva instar pertama lepidoptera juga umumnya mengkonsumsi daun muda.
Panda dan Kush (1995) menyatakan bahwa tanin dapat menurunkan daya cerna protein pada serangga. Hasil penelitian Arnold dan Schultz (2002) pada Populus menunjukkan bahwa produksi senyawa ini bersifat inducible. Lebih jauh diungkapkan terdapat indikasi bahwa luka akibat aktivitas makan serangga mengakibatkan aktivitas biosintesis tanin pada bagian tersebut meningkat. Crozier et al. (2006) mengemukakan bahwa tanin dapat mengikat protein dalam pencernaan serangga dengan menonaktifkan fungsi enzim pencerna protein pada sistem pencernaan serangga. Proses ini akan berdampak negatif bagi nutrisi serangga.
nikotin. Dalam interaksi tumbuhan-herbivora, Adler et al. (2001) menyatakan senyawa ini merupakan salah satu mekanisme resistensi tumbuhan berbasis kimiawi. Alkaloid dengan konsentrasi tertentu dalam tubuh tumbuhan dapat mengurangi serangan herbivora. Flavonoid merupakan senyawa turunan fenol yang bertanggung jawab terhadap zat warna merah, ungu, biru, dan sebagian zat warna kuning dalam tumbuhan (Ferdian 2008). Salah satu senyawa yang termasuk flavonoid adalah antosianin. Selain sebagai pigmen tanaman, antosianin juga diketahui bersifat antibiosis (Panda dan Kush 1995).
BAB III
IDENTIFIKASI KARAKTER MORFOLOGI DAN KANDUNGAN
FITOKIMIA TIGA BELAS AKSESI HANDEULEUM
3.1 Abstrak
Secara teori, penerimaan serangga terhadap tanaman inangnya bergantung pada kombinasi komposisi senyawa kimia dan morfologi tanaman yang bersifat stimulan dan deterrent bagi serangga. Pada penelitian ini dilakukan analisis lebih lanjut terhadap beberapa karakter morfologi serta kandungan fitokimia sebagai langkah identifikasi awal mekanisme resistensi 13 aksesi handeuleum terhadap Doleschallia bisaltide (Lepidoptera: Nymphalidae). Penelitian ini bertujuan untuk mengetahui perubahan komposisi fitokimia 13 aksesi handeuleum setelah terserang larva D. bisaltide. Penelitian dilakukan pada bulan November 2009- Februari 2010 bertempat di BALITTRO dan IPB. Masing-masing aksesi dibagi menjadi dua kelompok. Kelompok pertama langsung dipanen tanpa perlakukan (sampel daun sebelum terserang), kelompok kedua diinfestasikan larva instar III D. bisaltide selama sehari kemudian dipanen (sampel daun setelah terserang). Hasil penelitian menunjukkan terdapat variasi pada karakter luas area daun total, keragaan dan kerapatan trikoma, serta kandungan fitokimia 13 aksesi tanaman handeuleum. Serangan D. bisaltide mempengaruhi komposisi pigmen tanaman, senyawa alkaloid, tanin, triterpenoid, flavonoid, steroid, glikosida, saponin; rasio C/N, kalsium, dan serat 13 aksesi tanaman handeuleum. Produksi senyawa tanin, alkaloid, dan glikosida diduga bersifat konstitutif, sedangkan triterpenoid, flavonoid, saponin, dan steroid diduga diproduksi tanaman secara inducible. Korelasi yang erat terjadi antara antosianin, klorofil, dan karotenoid; N dan C/N rasio; serta antara triterpenoid dan steroid.
Kata Kunci: komposisi fitokimia, morfologi, handeuleum
Abstract
caricature plant accessions. D. bisaltide attack influences composition of plant pigments, alkaloids, tannin, triterpenoid, flavonoids, steroid, glycosides, saponin; C/N ratio, calcium, and cellulose of 13 accessions. Tannin, alkaloids, and glycosides were produced constitutively, while triterpenoid, flavonoids, saponin, and steroid were produced inducible. There are significant correlation value between anthocyanin, chlorophyll, and carotenoid; N and C/N ratio; and also between triterpenoids and steroid.
Key words: phytochemical composition, morphology, caricature plant
3.2 Pendahuluan
Interaksi herbivora-tumbuhan umumnya bersifat antagonis, yakni tumbuhan berupaya mencegah herbivora datang dan mengkonsumsi bagian tubuh tumbuhan. Schoonhoven et al. (1998) menyatakan mekanisme tersebut berupa morfologi dan kimiawi tumbuhan, atau dikenal dengan istilah resistensi. Van Zandt dan Agrawal (2004) menyatakan bahwa resistensi tanaman terhadap herbivora meliputi perubahan komposisi senyawa yang bersifat toksik dan antinutrisi bagi herbivora, serta penampakan morfologi, seperti kekerasan daun, kerapatan trikoma, duri, dan bulu pada tumbuhan.
Senyawa toksik pada tumbuhan umumnya terkait dengan komposisi metabolit sekunder. Menurut Fraenkel (1969) metabolit sekunder merupakan mekanisme resistensi utama pada tumbuhan. Lebih lanjut Fraenkel menyatakan bahwa komposisi kandungan metabolit sekunder menentukan tingkat penerimaan spesies herbivora tertentu terhadap tanaman tertentu, termasuk serangga. Schwachtje dan Baldwin (2008) menyatakan bahwa metabolit primer juga berperan sebagai antinutrisi bagi serangga. Oleh karena itu, identifikasi kandungan fitokimia tumbuhan yang berperan sebagai mekanisme resistensi terhadap serangan herbivora sangat diperlukan. Banyak penelitian yang telah mengungkapkan beberapa jenis metabolit sekunder yang umum ditemui pada berbagai tanaman budidaya, di antaranya alkaloid (Adler et al. 2001; Anizewski 2007), saponin (Papadopoulou et al. 1999; Saha et al. 2010), triterpenoid (Tholl et al. 2004), steroid, flavonoid, tanin, dan glikosida (Papadopoulou et al. 1999).
dikarenakan adanya dual peranan metabolit sekunder tanaman, baik deterrent ataupun stimulan, bagi serangga. Peranan tersebut bergantung pada spesialisasi serangganya. Panda dan Kush (1995); Schoonhoven et al. (2005); serta Alotaiba dan Elsayed (2007) menyatakan bahwa metabolit sekunder umumnya bersifat deterrent bagi serangga generalis, namun justru merupakan stimulan bagi serangga spesialis. Sebagai contoh, iridoid glikosida bersifat deterrent bagi Locusta migratoria (Orthophtera: Acridae), tetapi senyawa ini merupakan stimulan bagi Euphydryas editha (Lepidoptera: Nymphalidae). Panda dan Kush (1995) menyatakan hal ini dikarenakan serangga spesialis umumnya dapat mendetoksifikasi senyawa yang bersifat toksik.
Hasil penelitian Khumaida et al. (2008) pada 38 aksesi tanaman handeuleum menunjukkan bahwa 38 aksesi tersebut memiliki karakter morfologi yang serupa, sedangkan kandungan saponin, tanin, alkaloid, glikosida, flavonoid, steroid, dan triterpenoid bervariasi. Tiga belas aksesi di antaranya memiliki kandungan fitokimia tinggi. Secara teori, penerimaan serangga terhadap tanaman inangnya bergantung pada perbandingan komposisi senyawa kimia yang bersifat stimulan dan deterrent; serta morfologi tanaman tersebut (Panda dan Kush 1995; Schoonhoven et al. 2005). Oleh karena itu, pada penelitian ini dilakukan analisis lebih lanjut terhadap beberapa karakter morfologi serta kandungan fitokimia sebagai langkah identifikasi awal mekanisme resistensi 13 aksesi handeuleum terhadap Doleschallia bisaltide (Lepidoptera: Nymphalidae).
3.3 Tujuan
3.4 Hipotesis
Terdapat perbedaan karakter morfologi pada 13 aksesi handeuleum. Terdapat perubahan komposisi fitokimia pada 13 aksesi handeuleum setelah terserang larva D. bisaltide.
3.5 Metodologi Penelitian
Metodologi penelitian yang digunakan berupa pengamatan pada beberapa karakter morfologi dan fitokimia 13 aksesi handeuleum untuk mengidentifikasi mekanisme resistensi tanaman handeuleum.
3.5.1 Waktu dan Tempat
Pengamatan karakter morfologi dan pengumpulan sampel untuk pengujian fitokimia 13 aksesi tanaman handeuleum dilakukan pada bulan November sampai Desember 2009. Pengambilan sampel fitokimia, karakter morfologi, serta pengujian kandungan metabolit sekunder dan metabolit primer dilakukan di Balai Penelitian Tanaman Obat dan Aromatik (BALITTRO), Bogor. Analisis kandungan pigmen 13 aksesi tanaman handeuleum dilakukan di Laboratorium Research Group Crop Improvement (RGCI), Departemen Agronomi dan Hortikultura, IPB. Pengamatan keragaan dan kerapatan trikoma dilakukan di Laboratorium Ekofisiologi, Departemen Agronomi dan Hortikultura, IPB.
3.5.2 Persiapan Tanaman Uji
kualitatif minimal 12 jam setelah tanaman tersebut dikonsumsi herbivora. Kedua
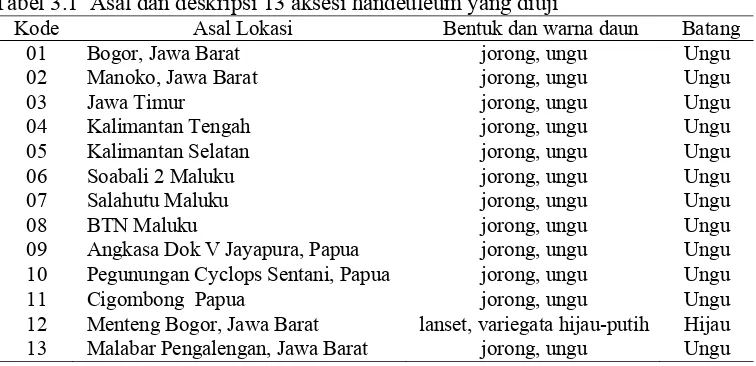
kelompok sampel tersebut dikeringkan pada suhu 40oC dan diekstrak sebelum dianalisis. Asal daerah ketigabelas aksesi yang diuji dtampilkan pada Tabel 3.1.
Tabel 3.1 Asal dan deskripsi 13 aksesi handeuleum yang diuji
Kode Asal Lokasi Bentuk dan warna daun Batang
01 Bogor, Jawa Barat jorong, ungu Ungu
02 Manoko, Jawa Barat jorong, ungu Ungu
03 Jawa Timur jorong, ungu Ungu
04 Kalimantan Tengah jorong, ungu Ungu
05 Kalimantan Selatan jorong, ungu Ungu
06 Soabali 2 Maluku jorong, ungu Ungu
07 Salahutu Maluku jorong, ungu Ungu
08 BTN Maluku jorong, ungu Ungu
09 Angkasa Dok V Jayapura, Papua jorong, ungu Ungu 10 Pegunungan Cyclops Sentani, Papua jorong, ungu Ungu
11 Cigombong Papua jorong, ungu Ungu
12 Menteng Bogor, Jawa Barat lanset, variegata hijau-putih Hijau 13 Malabar Pengalengan, Jawa Barat jorong, ungu Ungu
3.5.3 Persiapan serangga uji
Larva D. bisaltide yang digunakan untuk pengujian merupakan hasil rearing di rumah kaca. Larva serangga ini dipelihara hingga instar III di dalam kotak perbanyakan dan diberi pakan daun handeuleum. Larva D. bisaltide instar III kemudian diletakkan pada tanaman uji untuk memperoleh sampel pada kelompok daun setelah terserang larva D. bisaltide.
3.5.4 Pengamatan Karakter Morfologi Daun
1. Seluruh daun satu tanaman digambar pada kertas milimeter blok 2. Gambar daun dipotong, kemudian ditimbang
3. Sebagai kontrol, kertas dengan luas 580 cm2 ditimbang
4. Berat potongan gambar daun dibandingkan dengan berat kertas dengan menggunakan persamaan berikut:
3.5.5 Analisis Kandungan Fitokimia Daun Sebelum dan Setelah Terserang
Larva D.bisaltide
Analisis kandungan fitokimia daun handeuleum dilakukan terhadap beberapa senyawa yang diduga berhubungan dengan mekanisme resistensi tanaman handeuleum terhadap D. bisaltide. Senyawa tersebut terdiri atas alkaloid, saponin, triterpenoid, steroid, flavonoid, tanin, dan glikosida; serat, nitrogen, unsur kalsium, dan C organik; serta kandungan pigmen yakni pigmen klorofil total, karotenoid total, dan antosianin total. Analisis yang dilakukan meliputi :
• Analisis kandungan pigmen tanaman
Kandungan pigmen tanaman dianalisis dengan metode Sims dan Gamon (2002) menggunakan alat UV spektrofotometer. Konsentrasi Setiap pigmen diidentifikasi pada panjang gelombang yang berbeda-beda, dimana panjang gelombang 663 nm untuk pigmen klorofil a, 647 nm untuk klorofil b, 537 nm untuk antosianin, dan 470 nm untuk pigmen karotenoid. Data hasil pembacaan menggunakan alat UV spektrofotometer kemudian dikonversi ke dalam satuan mol/m2 dengan tahapan perhitungan sebagai berikut:
Antosianin = 0.01373*A537 - 0.00697*A647 – 0.002228*A663 Klorofil a = 0.01373*A663 – 0.000897*A537 – 0.003046*A663 Klorofil b = 0.02405*A647 – 0.004305*A537 – 0.005507*A663
Sehingga klorofil total dapat dihitung menggunakan persamaan berikut: Klorofil total = 7.15*A663-18.71*A647
Untuk mengkonversi konsentrasi pigmen per satuan luas digunakan perhitungan berikut:
Pigmen/area = (pigmen*6/1000) / (Luas area daun total dalam m2)
• Analisis metabolit sekunder
Analisis yang digunakan merupakan analisis visual berdasarkan kepekatan warna larutan. Data berupa skoring, berdasarkan standar Laboratorium Pengujian Balai Penelitian Tanaman Obat dan Aromatik (BALITTRO).
• Analisis selulosa, kalsium, nitrogen, dan C organik
Analisis yang dilakukan merupakan analisis kuantitatif berdasarkan metode standar yang digunakan pada Laboratorium Pengujian Balai Penelitian Tanaman Obat dan Aromatik (BALITTRO).
3.5.6 Analisis Data
Data fitokimia
Data fitokimia daun handeuleum dianalisis dengan membandingkan secara visual antara aksesi yang satu dengan aksesi lainnya.
Data morfologi
Data morfologi diuji menggunakan analisis sidik ragam. Apabila berbeda nyata, dilanjutkan dengan uji Tukey dengan bantuan program SAS v 9.0. Uji kenormalan data menggunakan metode Saphiro-Wilk. Uji korelasi antar karakter morfologi dan fitokimi dilakukan berdasarkan persamaan Pearson. Uji kenormalan dan korelasi dilakukan dengan program MINITAB 14. Hasil sidik ragam (Tabel 3.2) digunakan untuk menduga nilai ragam fenotipe, ragam genetik, dan ragam lingkungan.
Tabel 3.2. Analisis sidik ragam (Annichiarico 2002)
Sumber keragaman Db KT E(KT)
Ulangan r-1
-Aksesi g-1 M2 σ2e + r(σ2g + σ2gl + σ2gm + σ2glm)
Galat (r-1)(g-1) M1 σ2e
Pendugaan ragam genetik adalah sebagai berikut:
σ2
e = ragam lingkungan = Kuadrat Tengah Galat (KTG) = M1
σ2
g =ragam genetik = (M2 – M1) / r
σ2
p = σ2g + σ2e
Pendugaan nilai heritabilitas arti luas dihitung berdasarkan analisis ragam (Allard 1966) dengan persamaan berikut:
h2bs = (σ2G / σ2P) x 100%
dimana:
h2bs = heritabilitas arti luas
σ2
G = ragam genetik
σ2
P =ragam fenotipe
3.6 Hasil dan Pembahasan
3.6.1 Karakter Morfologi
Morfologi tanaman merupakan karakter yang berperan dalam proses pencarian inang oleh serangga (Schoonhoven et al. 2005). Hal tersebut diduga juga berlaku pada interaksi antara tanaman handeuleum dengan D. bisaltide sebagai hama. Bentuk tajuk tanaman dan warna bagian tertentu dari tanaman dapat menjadi stimulan atau deterrent bagi serangga. Karakter morfologi yang diduga berperan dalam proses penerimaan D. bisaltide terhadap tanaman handeuleum adalah luas area daun total, warna tanaman, serta kerapatan trikoma pada daun tanaman handeuleum.
Hasil uji kenormalan sebaran data menunjukkan bahwa karakter kerapatan trikoma dan luas area daun total menyebar normal (Lampiran 2 dan Lampiran 3). Berdasarkan pengamatan, kerapatan trikoma pada 13 aksesi bervariasi. Luas area daun total 13 aksesi tidak berbeda nyata, tetapi, bila diurutkan, aksesi 3 (1 020.17 cm2) dan aksesi 8 (1 027.86 cm2) merupakan aksesi dengan luas area daun total terluas, sedangkan aksesi 1 (403.42 cm2) dan aksesi 12 (514.10 cm2) merupakan aksesi dengan luas area daun total tersempit. Luas area daun total dan kerapatan trikoma masing-masing aksesi ditampilkan pada Tabel 3.3.
Tabel 3.3 Luas area daun total dan kerapatan trikoma 13 aksesi handeuleum Aksesi Luas area daun total (cm2) Kerapatan trikoma/ 0.038 mm2
1 403.42 ±147.12 1.5 ±0.71
Heritabilitas merupakan suatu parameter yang menentukan seberapa besar suatu karakter dapat diturunkan dari induk kepada keturunannya. Nilai heritabilitas juga mencerminkan seberapa besar peran lingkungan dalam mempengaruhi ekspresi karakter tertentu. Semakin besar nilai heritabilitas, semakin besar potensi suatu karakter diturunkan pada keturunannya. Hal ini menandakan semakin kecil pengaruh lingkungan pada karakter tersebut. Kriteria heritabilitas (%) menurut Stansfield (1983 dalam Komariah et al. 2004) serta Zen dan Bahar (1996) adalah sebagai berikut:
Tabel 3.4 Kriteria Heritabilitas
Nilai duga heritabilitas (%) Kriteria heritabilitas 0 < x < 20 Rendah 20 ≤ x < 50 Sedang
Nilai rekapitulasi sidik ragam pada karakter luas area daun total dan kerapatan trikoma ditampilkan pada Tabel 3.5. Nilai heritabilitas karakter kerapatan trikoma sebesar 21.69%. Nilai heritabilitas untuk karakter luas area daun total adalah 20.89%. Berdasarkan klasifikasi Zen dan Bahar (1996), maka nilai heritabilitas kedua karakter termasuk kategori sedang. Hal ini berarti, kedua karakter ini dapat diturunkan, namun sedikit dipengaruhi lingkungan. Pendugaan ragam genetik (σ2g),
ragam fenotipe (σ2p), ragam lingkungannya (σ2e), serta nilai heritabilitas ditampilkan
pada Tabel 3.5.
Tabel 3.5 Rekapitulasi sidik ragam, ragam genetik, ragam fenotipe, ragam galat, serta nilai heritabilitas pada karakter luas area daun total dan kerapatan trikoma tanaman handeuleum
Rekapitulasi komponen sidik ragam No Sumber keragaman KT Luas area
daun total
KT Kerapatan trikoma/0.038 mm2
1 Ulangan 46286.176 58.500
2 Aksesi 138910.867 23.955
3 Galat 90898.128 15.417
Komponen Pendugaan Parameter Genetik
No Parameter genetik Luas area daun total Kerapatan trikoma/0.038 mm2
1 Ragam genetik (σ2g) 24006.367 4.269
2 Ragam fenotipe (σ2p) 114904.498 19.686
3 Ragam galat (σ2e) 90898.128 15.417
4 Nilai heritabilitas 20.893S 21.687S
Keterangan : S = sedang; nilai ragam genetik, ragam fenotipe, dan ragam lingkungan diduga berdasarkan sidik ragam Tabel 3.2.
3.6.2 Kandungan Fitokimia
aksesi 12 lebih rendah dibandingkan aksesi lainnya. Perbandingan kandungan metabolit sekunder sebelum dan setelah tanaman terserang larva D. bisaltide ditampilkan pada Tabel 3.6.
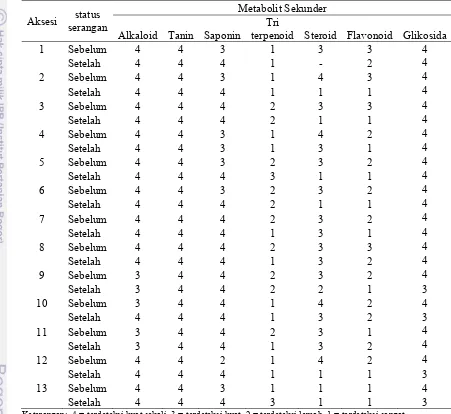
Tabel 3.6 Kandungan metabolit sekunder daun tanaman handeuleum sebelum dan setelah terserang larva D. bisaltide
Aksesi status
Keterangan: 4 = terdeteksi kuat sekali, 3 = terdeteksi kuat, 2 = terdeteksi lemah, 1 = terdeteksi sangat lemah, - = tidak terdeteksi
pada ada atau tidaknya stres lingkungan. Senyawa golongan inducible adalah senyawa yang diproduksi atau berhenti diproduksi bila tanaman mengalami stres lingkungan.
Dalam proses pelepasannya, senyawa dibagi menjadi dua kelompok, yakni senyawa yang dilepas secara konstitutif dan inducible. Senyawa yang dilepas secara konstitutif adalah senyawa yang selalu dikeluarkan tanaman baik ada atau tidaknya stres lingkungan, sedangkan senyawa yang dilepas secara inducible adalah kelompok senyawa yang dilepaskan ke lingkungan hanya pada kondisi tertentu. Senyawa yang dilepas secara konstitutif adalah senyawa yang diproduksi secara konstitutif, sedangkan senyawa yang dilepas secara inducible dapat merupakan senyawa yang diproduksi secara konstitutif maupun inducible. Pada penelitian ini, kandungan fitokimia digolongkan berdasarkan proses produksi, bukan regulasi pelepasannya.
Hasil analisa kandungan sampel daun handeuleum setelah terserang larva D. bisaltide menunjukkan konsentrasi alkaloid, tanin, dan glikosida cenderung tidak mengalami perubahan. Panda dan Kush (1995) menyatakan bahwa alkaloid umumnya diproduksi, kemudian dialokasikan dalam jaringan tanaman yang rentan terhadap serangan harbivora. Aniszewski (2007) menyatakan alkaloid diproduksi tanaman dan dikompartementasi di dalam vakuola, sehingga Vilarino dan Ravetta (2007) dapat menyatakan bahwa alkaloid diproduksi tanaman secara konstitutif. Namun Vilarino dan Ravetta (2007) menambahkan bila tanaman mengalami stress, seperti akibat serangan herbivora, konsentrasi alkaloid dapat meningkat.
Hasil penelitian Vilarino dan Ravetta (2007) diperkuat oleh hasil penelitian Vazquez-Flota et al. (2004) yang menunjukkan bahwa aplikasi etilen dan asam jasmonat dapat menstimulasi peningkatan konsentrasi alkaloid pada Catharanthus roseus. Hasil penelitian ini juga dipertegas oleh Gaines (2004) pada kultur jaringan. Berdasarkan penjabaran tersebut, dapat disimpulkan bahwa senyawa alkaloid pada tanaman handeuleum diproduksi secara konstitutif, namun dikonjugasikan atau disimpan pada bagian tertentu untuk mencegah autotoksisitas pada tanaman. Karenanya, tidak terdapat perubahan konsentrasi setelah tanaman terserang larva D. bisaltide.
konstitutif. Hal ini didukung pernyataan Hogedal dan Molgaard (2000 dalam Beninger dan Cloutier 2008) bahwa umumnya produksi glikosida dalam tanaman bersifat konstitutif. Glikosida merupakan salah satu senyawa yang diketahui bersifat deterrent bagi banyak serangga. Akan tetapi, berdasarkan studi literatur, senyawa ini dapat disequisterasi oleh sejumlah serangga spesialis (Vickery dan Vickery 1981). Hasil penelitian Pereyra dan Bowers (1988), begitu juga dengan Bowers dan Stamp (1997) menunjukkan hal tersebut juga berlaku bagi famili Nymphalidae. Berdasarkan hal tersebut, diduga D. bisaltide menggunakan kandungan glikosida yang terkandung dalam tanaman sebagai salah satu sidik jari untuk menemukan inangnya.
Kandungan saponin, triterpenoid, steroid, dan flavonoid mengalami perubahan konsentrasi. Pada 13 aksesi handeuleum, kandungan flavonoid cenderung mengalami penurunan konsentrasi. Kandungan saponin cenderung mengalami peningkatan konsentrasi. Konsentrasi triterpenoid pada daun setelah terserang tidak memiliki pola tertentu. Pada aksesi 1, 2, 4, 6, 9, 10, dan 12 konsentrasi triterpenoid tidak mengalami perubahan, pada aksesi 5 dan 13 konsentrasi triterpenoid meningkat, pada aksesi 7, 8, dan 11 konsentrasi triterpenoid dalam daun menurun. Konsentrasi steroid umumnya mengalami penurunan setelah tanaman terserang. Pada aksesi 1, penurunan yang terjadi sangat drastis. Senyawa ini tidak terdeteksi terdapat dalam jaringan kelompok daun setelah tanaman terserang. Diduga konsentrasi steroid menjadi sangat rendah pada sampel daun setelah terserang, sehingga tidak terdeteksi pada pengujian secara kualitatif.
setelah daun terserang kandungan senyawa ini menurun, tetapi konsentrasinya masih lebih tinggi dibandingkan aksesi 12 (Tabel 3.7).
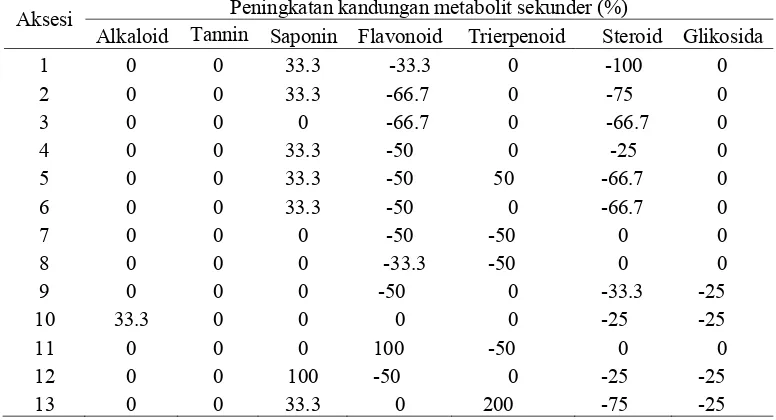
Perubahan komposisi senyawa steroid, flavonoid, saponin, dan triterpenoid setelah tanaman terserang mengindikasikan bahwa senyawa tersebut diproduksi tanaman secara inducible, yang berarti konsentrasi senyawa tersebut akan berubah setelah tanaman terserang herbivora. Perubahan ini terjadi akibat herbivora mengeluarkan elicitor, berupa saliva herbivora (Southwood 1996; Felton 2008; State 2009), Saliva serangga mengandung glucose oxidase. Senyawa inilah yang merupakan elicitor yang dikenali tanaman sebagai sinyal adanya bahaya. State (2009) menambahkan bahwa senyawa ini pula yang menyebabkan respon yang berbeda antara tanaman yang terserang herbivora dengan kerusakan mekanis lainnya, seperti dirusak menggunakan gunting. Sinyal tersebut akan mengaktifkan lintasan transduksi pada tanaman yang kemudian mengaktifkan gen-gen untuk memproduksi senyawa pertahanan lebih banyak atau lebih sedikit (Memelink 2009). Persentase perubahan kandungan metabolit sekunder dihitung berdasarkan persamaan berikut:
Persen perubahan metabolit skeunder = [(skor akhir-skor awal)/skor awal] x 100%
Tabel 3.7 Persentase perubahan kandungan metabolit sekunder pada daun 13 aksesi handeuleum
Aksesi Peningkatan kandungan metabolit sekunder (%)
Alkaloid Tannin Saponin Flavonoid Trierpenoid Steroid Glikosida
1 0 0 33.3 -33.3 0 -100 0
2 0 0 33.3 -66.7 0 -75 0
3 0 0 0 -66.7 0 -66.7 0
4 0 0 33.3 -50 0 -25 0
5 0 0 33.3 -50 50 -66.7 0
6 0 0 33.3 -50 0 -66.7 0
7 0 0 0 -50 -50 0 0
8 0 0 0 -33.3 -50 0 0
9 0 0 0 -50 0 -33.3 -25
10 33.3 0 0 0 0 -25 -25
11 0 0 0 100 -50 0 0
12 0 0 100 -50 0 -25 -25
13 0 0 33.3 0 200 -75 -25
Tabel 3.8 Perbandingan kandungan pigmen 13 aksesi handeuleum sebelum dan setelah terserang larva D. bisaltide.
Aksesi
Pigmen (mol/m2)
Antosianin Klorofil Karotenoid
Sebelum Setelah Sebelum Setelah Sebelum Setelah
1 0.620 0.708 0.342 0.424 0.225 0.277
2 0.476 0.617 0.391 0.233 0.223 0.232
3 0.391 0.509 0.391 0.440 0.224 0.268
4 0.259 0.494 0.242 0.292 0.154 0.218
5 0.373 0.817 0.414 0.687 0.225 0.407
6 0.312 0.423 0.355 0.331 0.197 0.208
7 0.313 0.429 0.307 0.387 0.196 0.229
8 0.600 0.673 0.502 0.402 0.301 0.302
9 0.453 0.496 0.512 0.390 0.278 0.240
10 0.375 0.532 0.437 0.458 0.244 0.272
11 0.130 0.076 0.566 0.559 0.246 0.245
12 0.457 0.272 0.520 0.274 0.285 0.175
13 0.620 0.708 0.342 0.424 0.225 0.277
Serangan larva D. bisaltide menyebabkan penurunan kandungan klorofil pada aksesi 1-11, tetapi menyebabkan hal sebaliknya pada aksesi 12 dan 13. Hasil analisis menunjukkan adanya peningkatan pigmen karotenoid pada daun handeuleum pada aksesi 1-9, dan 11, sedangkan pada aksesi 10, 12, dan 13 justru sebaliknya (Gambar 3.2).
Gambar 3.2 Grafik persentase perubahan kandungan antosianin, klorofil, dan karotenoid pada 13 aksesi tanaman handeuleum setelah tanaman terserang larva D. bisaltide.
Hasil analisis pigmen pada 13 aksesi menunjukkan kandungan antosianin dan karotenoid pada daun setelah terserang larva D. bisaltide secara umum mengalami peningkatan, kecuali pada aksesi 3, 9, 10, 12, dan 13 (Gambar 3.2) yang justru mengalami penurunan. Peningkatan konsentrasi pigmen antosianin berkisar 9-118 %, sedangkan peningkatan karotenoid berkisar 4-80 %. Peningkatan kedua jenis pigmen mengindikasikan bahwa keduanya tidak hanya berfungsi sebagai pewarna, tetapi juga berperan dalam sistem resistensi tanaman handeuleum terhadap D. bisaltide. Hal ini didukung oleh pernyataan Guo et al. (2008) dan Currtiss et al. (2008) bahwa selain sebagai pigmen, antosianin dan karotenoid juga bersifat antibiosis. Lebih lanjut Guo et al. (2008) menyatakan bahwa biosintesis kedua pigmen ini bersifat inducible. Salah satu faktor yang mempengaruhinya adalah pelukaan akibat aktivitas herbivora.
Kandungan klorofil total secara umum meningkat pada daun tanaman setelah terserang larva kecuali pada aksesi 10 dan 13. Peningkatan terbesar terjadi pada aksesi 6, yakni sebesar 80.95%, sedangkan penurunan kandungan klorofil terbesar terjadi pada aksesi 13, yaitu sebesar 38.72% (Tabel 3.9 dan Gambar 3.2). Hasil analisis menunjukkan adanya indikasi bahwa kandungan klorofil berkorelasi dengan nitrogen (Tabel 3.10). Hal ini sesuai dengan pernyataan Hirota dan Kato (2001) menyatakan bahwa kandungan klorofil berkorelasi positif dengan kandungan nitrogen tanaman. Semakin tinggi kandungan nitrogen tanaman, semakin tinggi pula kandungan klorofil tanaman tersebut.
Gambar 3.3 Grafik perbandingan kandungan C/N rasio pada pada daun 13 aksesi handeuleum sebelum dan setelah terserang larva D. bisaltide.
Keterangan: Data merupakan rataan ± SE.
Gambar 3.3 memperlihatkan bahwa serangan larva D. bisaltide pada daun handeuleum menyebabkan penurunan konsentrasi baik pada C organik maupun nitrogen. Akan tetapi, karena besarnya persentase penurunan nitrogen lebih tinggi dibandingkan besarnya penurunan konsentrasi C organik, maka C/N rasio pada tanaman meningkat. Judkins dan Wander (1949) menyatakan bahwa serangan hama mempengaruhi status nitrogen dalam tubuh tanaman. Hasil penelitian Newingham et al. (2007) menunjukkan bahwa aktivitas makan herbivora meningkatkan alokasi nitrogen dalam tubuh tanaman.
Gambar 3.4 Besarnya perubahan kandungan unsur C dan N pada tanaman handeuleum setelah terserang larva D. bisaltide.
Gambar 3.5 menunjukkan adanya perbandingan kandungan kalsium sebelum dan setelah tanaman terserang. Peningkatan kandungan kalsium setelah tanaman terserang mengindikasikan bahwa unsur ini diserap tanaman secara inducible. Hal ini bertolak belakang dengan hasil penelitian Ruiz et al. (2002) pada tanaman Pancratium sickenbergeri penyerapan kalsium cenderung bersifat konstitutif dibandingkan inducible. Hasil penelitian ini serupa dengan hasil penelitian Molano-Flores (2001) pada Sida rhombifolia yang menunjukkan aktivitas makan herbivora pada tanaman akan meningkatkan pembentukan kristal kalsium. Schwachtje dan Baldwin (2008) menyatakan bahwa kalsium merupakan bahan baku untuk membentuk CaOx (Kalsium oksalat). Senyawa ini berupa kristal yang dapat membuat tumpul mandibel serangga menggigit mengunyah. Karenanya, unsur kalsium merupakan salah satu unsur yang berperan penting dalam sistem resistensi tanaman terhadap herbivora.
Gambar 3.6 Grafik perbandingan kandungan serat pada daun 13 aksesi handeuleum sebelum dan setelah terserang larva D. bisaltide.
Tabel 3.9 Persentase pertambahan konsentrasi fitokimia 13 aksesi handeuleum
Aksesi
Persentase perubahan kandungan fitokimia tanaman handeuleum (%)
Anto Kloro Karo Alka Tannin Sapo Flavo Terpen Stero Gliko C N C/N Ca Serat
1 14.21 24.06 23.3 0 0 50 -50 0 -150 0 -2.45 -29.08 37.55 34.62 2.64
2 - - - 0 0 50 -100 0 -150 0 -1.54 -23.66 28.98 29.76 6.42
3 29.57 -40.5 4.21 0 0 0 -100 0 -100 0 7.93 -20.41 35.61 57.32 0.21
4 29.97 12.52 19.57 0 0 50 -50 0 -50 0 -3.40 -16.62 15.86 24.30 -9.26
5 90.7 20.89 41.23 0 0 50 -50 50 -100 0 -6.98 -21.33 18.25 24.00 -6.86
6 118.8 66.17 80.95 0 0 50 -50 0 -100 0 -8.59 -15.51 8.19 82.54 0.28
7 35.61 -6.77 5.64 0 0 0 -50 -50 0 0 5.43 -25.89 42.25 138.00 2.56
8 37.18 26.13 16.77 0 0 0 -50 -50 0 0 -4.41 -30.73 38.00 73.77 -25.73
9 12.1 -19.9 0.5 0 0 0 -50 0 -50 -50 -5.28 -23.04 23.07 94.44 -1.45
10 9.38 -23.83 -13.61 50 0 0 0 0 -50 -50 -9.23 -30.48 30.56 91.94 -3.07
11 41.87 4.77 11.51 0 0 0 50 -50 0 0 0.95 -20.89 27.60 79.66 1.88
12 -41.73 -1.27 -0.25 0 0 100 -50 0 -50 -50 -1.44 -28.90 38.63 120.34 7.57 13 -40.41 -47.32 -38.72 0 0 50 0 100 -150 -50 -3.68 -32.84 43.41 87.67 -16.62
3.6.3 Korelasi antar karakter morfologi dan fitokimia tanaman
Analisis korelasi menunjukkan bahwa klorofil dan karotenoid berkorelasi negatif nyata dengan antosianin, sedangkan korelasi antara keduanya bersifat positif. Nitrogen berkorelasi negatif sangat nyata dengan nilai C/N, luas area daun total dan kerapatan trikoma. Nilai C/N berkorelasi nyata positif dengan triterpenoid, luas area daun total, dan kerapatan trikoma. Saponin berkorelasi nyata positif dengan luas area daun total. Triterpenoid berkorelasi negatif sangat nyata dengan kandungan steroid. Hasil analisis korelasi antar karakter morfologi dan fitokimia tanaman handeuleum ditampilkan pada Tabel 3.10.
Roy (2000) menyatakan bahwa korelasi menggambarkan keeratan hubungan antara satu karakter dengan karakter lainnya. Lebih jauh Roy (2000) yang dipertegas oleh Mattjik dan Sumertajaya (2002) menyatakan bahwa bila nilai korelasi antara dua peubah mendekati -1 atau 1 menunjukkan bahwa hubungan antara kedua peubah tersebut sangat erat. Berdasarkan analisis korelasi, nilai yang mendekati -1 atau 1 terdapat pada korelasi antara ketiga jenis pigmen tanaman yang dianalisis, N dan C/N rasio, serta antara triterpenoid dan steroid.
Pigmen tanaman saling mendominasi untuk menimbulkan warna tertentu pada bagian tubuh tumbuhan, karenanya wajar bila hubungan antara ketiga jenis pigmen tanaman yang diuji menunjukkan korelasi yang sangat kuat. Klorofil dan karotenoid berkorelasi negatif nyata dengan antosianin, sedangkan korelasi antara keduanya bersifat positif. Produksi antosianin bertolak belakang dengan klorofil, dan penentu di antara keduanya adalah suhu.
Tabel 3.10 Korelasi antar karakter morfologi dan fitokimia pada tanaman handeuleum
Antosianin Klorofil Karotenoid C organik Nitrogen C/N Kalsium Serat Alkaloid Saponin Flavo Tterpen Steroid Luas daun Klorofil -0.99**
Karotenoid -0.89** 0.83** C organik -0.19 0.14 0.33
Nitrogen -0.29 0.30 0.25 0.53
C/N 0.25 -0.27 -0.13 -0.13 -0.91**
Ca 0.30 -0.32 -0.21 -0.52 -0.43 0.25
Serat 0.06 -0.15 0.31 0.00 0.01 -0.03 0.23
Alkaloid -0.05 0.04 0.07 -0.57 -0.15 -0.11 0.41 0.36
Saponin 0.48 -0.50 -0.33 0.12 -0.29 0.38 -0.33 0.14 -0.53
Flavonoid 0.38 -0.38 -0.36 -0.40 0.02 -0.21 0.14 0.28 0.38 0.12
Triterpenoid 0.20 -0.25 0.02 0.11 -0.47 0.60* -0.25 0.13 -0.10 0.55 0.11
Steroid -0.52 0.54 0.35 0.01 0.27 -0.30 0.19 -0.06 0.00 -0.46 -0.37 -0.84** luas daun 0.10 -0.16 0.16 -0.14 -0.57* 0.58* 0.15 0.52 0.05 0.58* -0.02 0.54 -0.19
Trikoma -0.05 0.05 -0.12 -0.22 -0.65* 0.66* -0.00 0.23 0.12 0.44 0.33 0.32 -0.11 0.42 Keterangan : Korelasi menggunakan metode Pearson. * nyata pada taraf 5%, ** sangat nyata pada taraf 1%.
Flavo = flavonoid; tterpen = triterpenoid
Pada tabel 3.10 nampak bahwa triterpenoid memiliki nilai korelasi yang nyata, tinggi, dan bertolak belakang dengan steroid, yakni -0.84. Apabila dikaitkan dengan jalur lintasan metabolismenya (Gambar 3.7), steroid merupakan senyawa turunan dari triterpenoid, sehingga dalam prosesnya tanaman dapat memilih untuk menghentikan proses hanya sampai terbentuk triterpenoid atau merombak triterpenoid yang telah terbentuk untuk memproduksi steroid. Hasil analisis menunjukkan bahwa konsentrasi steroid cenderung mengalami penurunan setelah tanaman terserang larva D. bisaltide. Hal ini mengindikasikan bahwa sebagian besar triterpenoid yang dihasilkan tidak digunakan tanaman untuk membentuk steroid.
3.7 Kesimpulan
Berdasarkan penjabaran sebelumnya, kesimpulan yang dapat ditarik adalah 1. Terdapat variasi pada karakter luas area daun total, keragaan dan kerapatan
trikoma, serta kandungan fitokimia 13 aksesi tanaman handeuleum.
2. Serangan D. bisaltide mempengaruhi komposisi pigmen tanaman, senyawa alkaloid, tanin, triterpenoid, flavonoid, steroid, glikosida, dan saponin 13 aksesi tanaman handeuleum. Begitu pula dengan konsentrasi rasio C/N, kalsium dan serat pada tanaman.
3. Produksi senyawa tanin, alkaloid, dan glikosida diduga bersifat konstitutif, sedangkan triterpenoid, flavonoid, saponin, dan steroid diduga diproduksi tanaman secara inducible.
4. Serangan D. bisaltide cenderung meningkatkan produksi pigmen dalam tubuh tanaman. Demikian juga dengan C/N rasio dan penyerapan kalsium.