i
Dengan ini saya menyatakan bahwa tesis Struktur Genetika Populasi Ikan Malalugis (Decapterus macarellus) di Perairan Sekitar Pulau Sulawesi Berdasarkan Mt-DNA Marker adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Januari 2012
iii
ACHMAD ZAMRONI. Population Genetic Structure of Malalugis Fish (Decapterus macarellus) in The Waters Around The Island of Sulawesi Based on Mt-DNA Marker. Under direction of NEVIATY P. ZAMANI and ESTU NUGROHO.
Studies on genetic diversity of Malalugis (Decapterus macarellus) has been undertaken around Sulawesi to obtain population genetic structure of the study area. The study was based on RFLP analysis of mitochondrial genome DNA extracted from fish tissue (meat, fins). Fish samples collected from several populations of small-scale fisheries captured in several locations around Sulawesi. The results showed that the genetic diversity obtained by including low, ie between 0 to 0.3698. Provided two major groups in the population structure Malalugis fish in the waters around Sulawesi Island, namely: the first group is represented by a population of Makassar Strait,Bone Bay, Flores Sea, Banda Sea, Tolo Bay, Molucca Sea and Tomini Bay and the second group is represented by the Celebes Sea population. The population of the Bone Bay, Flores Sea, Banda Sea, Tolo Bay and Molucca Sea has a very close kinship, so that probably originated from the same stock. In the population there Tomini Bay locals, because there is little difference with adjacent populations (Maluku Sea and Banda Sea).
v
ACHMAD ZAMRONI! Struktur Genetika Populasi Ikan Malalugis (Decapterus macarellus) di Perairan Sekitar Pulau Sulawesi Berdasarkan Mt-DNA Marker. Dibimbing oleh NEVIATY P. ZAMANI dan ESTU NUGROHO.
Ikan Malalugis merupakan ikan pelagis yang bersifat oseanik dan tereksploitasi tinggi. Tujuan dari penelitian ini adalah untuk mengkaji struktur genetika populasi ikan Malalugis (Decapterus macarellus) di perairan sekitar pulau Sulawesi yang meliputi Laut Sulawesi, Selat Makassar, Teluk Bone, Laut Flores, Teluk Tolo, Teluk Tomini, Laut Banda, dan Laut Maluku. Informasi yang diperoleh berupa struktur populasi/unit stok ikan Malalugis berdasarkan perbedaan struktur genetika populasi. Lokasi penelitian meliputi Laut Sulawesi, Selat Makassar, Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo, Laut Maluku dan Teluk Tomini. Waktu penelitian dilaksanakan selama bulan Maret 2010 – Oktober 2011. Analisis sampel dilaksanakan di laboratorium Genetika, Balai Penelitian Perikanan Laut, Muara Baru, Jakarta. Analisis untuk mendapatkan data polimorfisme fragmen dan situs restriksi dilaksanakan dengan metode RFLP (Restriction Fragment Length Polymorphism) dengan menggunakan 6 jenis enzim restriksi (restriction endonuclease). Dalam amplifikasi digunakan 2 macam ‘primer’, yaitu primer HN20 dan LN20; sedang enzim restriksi yang dipakai 6 jenis, yaitu Mbo I, Alu I, Hind III, Taq I, Rsa I dan Xba I. Untuk analisis data dibantu dengan menggunakan perangkat lunak TFPGA (Tools For Population Genetic Analyses).
Hasil penelitian menunjukkan dari enam enzim restriksi (Alu I, Hind III, Mbo I, Rsa I, Taq I dan Xba I) yang digunakan untuk memotong sekuens mtDNA ikan malalugis, semuanya dapat memberikan situs pemotongan. Tipe-tipe haplotipe yang diperoleh dari hasil restriksi dapat teridentifikasi 6 jenis alel atau composite haplotype, yaitu AAAAAA, AAAABA, AAAAAB, BAABAC, BABCAC dan BAABCC. Tipe komposit haplotipe AAAAAA terdapat pada semua populasi, hal ini mengindikasikan bahwa ikan malalugis yang mempunyai tipe komposit haplotipe AAAAAA bersifat lebih adaptif. Nilai keragaman haplotipe (haplotype diversity) ikan malalugis yang diperoleh berkisar antara 0 – 0,3698, dengan nilai terendah (0) pada populasi Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo dan Laut Maluku. Nilai tertinggi keragaman haplotipe ikan Malalugis pada penelitian ini terdapat pada populasi Laut Sulawesi, yaitu sebesar 0,3698. Tingginya nilai keragaman haplotipe diduga disebabkan oleh adanya pengaruh dari populasi yang berasal dari Samudera Pasifik. Berdasarkan analisis berpasangan Fst menunjukkan bahwa terdapat perbedaan genetika yang cukup signifikan antara populasi Laut Sulawesi dengan ketujuh populasi lainnya. Jarak genetika terdekat adalah antara Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo dengan Laut Maluku (0,0000). Hal ini mengindikasikan bahwa kelima populasi tersebut berasal dari asal atau stok populasi yang sama.
Maluku dan Teluk Tomini berasal dari satu unit stok (mempunyai stok yang sama). Hal ini terjadi karena kelima populasi tersebut diduga merupakan jalur migrasi dari ikan Malalugis. Teluk Tomini sedikit berbeda dengan lima populasi lainnya (Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo dan Laut Maluku), sedikit perbedaan tersebut diduga di perairan Teluk Tomini terdapat populasi ikan malalugis yang bersifat lokal.
vii
© Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
ix Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
xi
Judul Penelitian : Struktur Genetika Populasi Ikan Malalugis (Decapterus macarellus) di Perairan Sekitar Pulau Sulawesi Berdasarkan Mt-DNA Marker
Nama : Achmad Zamroni
NRP : C551090151
Program Studi : Ilmu Kelautan
Disetujui Komisi Pembimbing
Dr. Ir. Neviaty P. Zamani, M.Sc. Dr. Ir. Estu Nugroho, M.Sc. Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana
Ilmu Kelautan
Dr. Ir. Neviaty P. Zamani, M.Sc. Dr. Ir. Dahrul Syah, M.Sc. Agr.
xiii
Syukur Alhamdulillah kami panjatkan ke hadirat Al-Khaliq Azza wa Jalla yang telah melimpahkan rahmat serta karunia-Nya sehingga kami dapat menyelesaikan penulisan hasil penelitian yang berjudul “ (#%+(%# '-'(.+)
$&%/)0. +)- )/)/%".0 ,. '#).#)- '+.()# %/)%
%/)1'0. '#,)0)#+)- ( )#+'#”. Penulisan hasil penelitian ini sebagai salah satu persyaratan untuk memperolah gelar Magister Sains pada Program Studi Ilmu Kelautan, Program Magister Pascasarjana Institut Pertanian Bogor.
Ucapan terimakasih yang sebesar-besarnya kami sampaikan kepada Ibu Dr. Ir. Neviaty P. Zamani, M.Sc., selaku Ketua Komisi Pembimbing dan Bapak Dr. Ir. Estu Nugroho, M.Sc., selaku Anggota Komisi Pembimbing, atas bimbingan, motivasi, diskusi dan arahannya serta ilmu pengetahuan yang telah banyak diberikan kepada kami. Tidak lupa kepada Bapak Drs. Suwarso, M.Si. dan teman-teman peneliti BRPL dari Kelti Pelagis Kecil atas bantuannya dalam pengumpulan data/sampel. Serta kepada Istri dan anak tercinta, atas dorongan semangat, kesabaran serta doa yang tulus.
Penulisan hasil penelitian ini telah kami upayakan dengan sebaik mungkin, akan tetapi kemungkinan masih banyak kekurangan, oleh karena itu kami mengharap masukan dan saran yang membangun demi kesempurnaan serta tambahan pengetahuan bagi kami. Atas semua masukan dan sarannya kami ucapkan banyak terimakasih.
Bogor, Januari 2012
xv
Penulis dilahirkan di Mojokerto pada tanggal 1 September 1979 sebagai putra pertama dari pasangan H. Imam Hanafi dan Hj. Siti Maisyaroh. Penulis lulus dari SMU Negeri 1 Sooko, Mojokerto pada tahun 1998. Pendidikan sarjana ditempuh di Program Studi Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Teknologi Sepuluh November, lulus pada tahun 2003. Pada tahun 2009 penulis mendapat kesempatan melanjutkan pendidikan pada Sekolah Pascasarjana IPB dengan memilih mayor Ilmu Kelautan. Beasiswa pendidikan pascasarjana diperoleh dari Kementrian Kelautan dan Perikanan Republik Indonesia.
xvii
2.5. Hukum Keseimbangan Hardy-Weinberg ... 19
3.4. Analisis Sampel ... 31
3.5. Interpretasi dan Analisis Data ... 34
4. HASIL DAN PEMBAHASAN ... 35
4.1. Keragaman Haplotipe Ikan Malalugis ... 35
4.2. Analisis Berpasangan Fst ... 39
4.3. Jarak Genetika ... 40
4.4. Struktur Genetika Populasi dan Hubungan Kekerabatan (Filogenetik) ... 42
5. SIMPULAN DAN SARAN ... 51
6. DAFTAR PUSTAKA ... 53
7. LAMPIRAN ... 57
xix
1. Pembentukan zigot pada kawin acak ... 20 2. Frekuensi haplotipe ikan malalugis (D. macarellus) hasil
restriksi dengan menggunakan enzim Alu I, Hind III, Mbo I,
Rsa I, Taq I dan Xba I ... 36 3. Keragaman genetik (diversitas haplotype, h) sekuen mtDNA
D-loop dari tujuh populasi ikan layang biru (D. macarellus) hasil restriksi oleh enzim restriksi Alu I, Hind III, Mbo I
(Nde II), Rsa I, Taq I dan Xba I ... 38 4. Hasil analisis antar populais ikan malalugis berdasarkan
xxi
1. Kerangka pemikiran penelitian genetika populasi
ikan malalugis ... 5 2. Ikan malalugis ... 8 3. Bentuk maxilla dan supramaxilla A) ikan malalugis;
B) ikan layang deles (FAO) ... 9 4. Peta Wilayah Wallacea ... 10 5. Thermohaline Circulation ... 12 6. Zimogram esterase dari ikan sidat (Anguilla sp) di
kawasan Segara Anakan, Cilacap ... 17 7. Peta lokasi penelitian dan lokasi sampling ... 29 8. Contoh visualisasi hasil pemotongan sekuen mtDNA D-loop
ikan layang biru (D.macarellus) oleh enzim Mbo I
dan Rsa I ... 35 9. Dendrogram hubungan kekerabatan (filogeni) dari
8 populasi ikan malalugis ... 43 10. Pola arus air permukaan laut pada musim barat,
a. Rizal et al. (2009) dan b. Wrytki (1961) ... 44 11. Pola arus air permukaan laut pada musim timur,
a. Rizal et al. (2009) dan b. Wrytki (1961) ... 44 12. Kesuburan perairan laut rata-rata musiman tahun
2004 – 2006 ... 47 13. Pola arus permukaan laut di Teluk Tomini pada saat
musim timur ... 48 14. Struktur genetika populasi ikan malalugis di perairan sekitar
xxiii
1. Contoh visualisasi hasil pemotongan sekuen mtDNA D-loop
ikan layang biru (D.macarellus) oleh enzim Alu I dan Hind III ... 56
2. Contoh visualisasi hasil pemotongan sekuen mtDNA D-loop
ikan layang biru (D.macarellus) oleh enzim Mbo I dan Rsa I ... 57
3. Contoh visualisasi hasil pemotongan sekuen mtDNA D-loop
ikan layang biru (D.macarellus) oleh enzim Taq I dan Xba I ... 58
4. Hasil penghitungan keragaman genetika dengan TFPGA ... 59
5. Hasil penghitungan Analisis Berpasangan Fst ... 70
!
! ! )()# '/)+)-"
Spesies ikan malalugis atau juga disebut layang biru (Decapterus macarellus) merupakan salah satu jenis ikan pelagis kecil yang tersebar luas di perairan Indonesia. Habitatnya terutama di perairan laut dalam (oseanik), membentuk gerombolan (schooling) yang besar di lapisan permukaan dan dalam siklus hidupnya memiliki kebiasaan sebagai ikan peruaya (migrasi). Daerah penyebarannya meliputi perairan Samudera Hindia (lepas pantai barat Sumatra Utara, perairan Mentawai dan selatan Jawa) (Hariati, 2005), juga di perairan Indonesia bagian timur (Suwarso, et al., 2000). Pola distribusi dan sifat migrasi dari ikan layang biru tersebut diduga terdapat penstrukturan genetika dari populasi ikan. Borsa (2003) menerangkan bahwa kemampuan persebaran yang luas dari suatu organisme tidak berarti terjadi homogenitas secara geografis.
Menurut Suwarso et al. (1998) ikan tersebut mempunyai nilai ekonomis tinggi dan bersifat exportable (untuk ikan umpan pada perikanan Tuna long-line) serta dikonsumsi secara lokal. Eksploitasi terhadap ikan layang biru telah berlangsung sejak lama, baik oleh perikanan industri (legal maupun illegal) maupun oleh perikanan yang bersifat skala kecil. Peningkatan eksploitasi diperkirakan terus berlangsung di berbagai perairan akibat permintaan pasar yang semakin tinggi, sehingga di beberapa wilayah terlihat tendensi penurunan hasil tangkapan dan ukuran ikan. Tekanan terhadap lingkungan berupa eksploitasi yang semakin meningkat menyebabkan terjadi penurunan kelimpahan stok dan rata-rata ukuran ikan; seleksi genetika yang merugikan terhadap fekunditas yang potensial; mengurangi rata-rata ukuran memijah; mengubah rasio jenis kelamin dan keseimbangan interspesifik; serta hilangnya diversitas genetika (Wilson & Clarke, 1996).
Pulau Sulawesi berada di Indonesia bagian tengah yang dibatasi oleh beberapa perairan yaitu Laut Sulawesi (utara), Selat Makassar (barat), Laut Flores (selatan), Laut Banda dan Maluku (timur). Perairan tersebut termasuk dalam kawasan Segitiga Karang/Coral Triangle yang mempunyai biodiversitas spesies laut yang sangat tinggi (Allen, 2000). Kawasan Segitiga Karang juga merupakan nursery ground dan rute migrasi bagi ikan dan mamalia laut. Arus Lintas Indonesia (Arlindo) atau disebut juga dengan Indonesian Through Flow oleh para ahli oseanografi merupakan arus massa air antar samudera yang melewati Indonesia. Arus ini mengalir dari Samudera Pasifik menuju Samudera Hindia melalui perairan di sekitar pulau Sulawesi. Menurut Hasanudin (1998), Arlindo secara dominan masuk dari Laut Sulawesi sebelah selatan Mindanao, kemudian menuju ke Selat Makassar. Ujung dari jalur ini kemudian bercabang menjadi dua, sebagian langsung menuju Samudera Hindia melalui Selat Lombok, dan sebagian lagi berbelok ke timur melewati Laut Flores menuju Laut Banda, kemudian menuju Samudera Hindia melalui Selat Ombai dan Laut Timor. Hasanudin (1998) menerangkan lebih lanjut bahwa jalur lain (kedua) dari Arlindo adalah masuk dari Laut Maluku kemudian menuju Laut Seram dan mengalir ke laut Banda melalui selat Manipa. Pada saat terjadi fenomena El-Nino, pergerakan sebagian dari massa air tadi berbalik arah dari wilayah perairan Indonesia menuju Samudra Pasifik. Ditinjau dari sejarah geologinya, pulau Sulawesi diduga di masa lampau, pulau ini tidak pernah bersatu dengan daratan manapun (Hall 2001). Berbeda halnya dengan Sumatra, Jawa, Bali, dan Kalimantan yang pernah bersatu dengan daratan Asia (Sundaland), serta Papua yang pernah bersatu dengan daratan Australia (Sahulland) sebelum kala Pleistosen (Pleistocene) berakhir (Shekelle dan Leksono, 2004).
yang dapat mempengaruhi fisiologi organisme untuk selanjutnya mempengaruhi struktur genetikanya. Rute pertukaran gen antara organisme tropis di Samudera Hindia dengan Samudera Pasifik terus berlangsung hingga saat ini. Pertukaran tersebut secara garis besar melalui perantara Arlindo (Gordon dan Fine, 1996). Beberapa peneliti menyebutkan bahwa dalam skala geografis variasi kondisi lingkungan diduga dapat menimbulkan variabilitas genetika pada ikan laut (Saunders et. al., 1986; Renno et. al., 1990; Effenberger dan Suchentrunk, 1999). Borsa (2003) menjelaskan bahwa perubahan tinggi muka laut pada kala Pleistosen mempengaruhi struktur genetika.
Informasi mengenai struktur populasi ikan layang biru berbasis genetika di perairan sekitar pulau Sulawesi belum tersedia. Struktur genetika populasi sebagai unit biologi sangat penting dipahami untuk menentukan keputusan manajemen perikanan yang tepat. Struktur genetika populasi dipelajari berdasarkan karakter polimorfisme yang terdapat dalam genom DNA mitochondria (mtDNA) melalui pemotongan (restriksi) sekuen teramplifikasi dengan menggunakan ezim restriksi. Dalam penelitian ini akan dipelajari variasi genetika populasi ikan malalugis (D. macarellus) di perairan sekitar pulau Sulawesi yang selanjutnya data tersebut dapat digunakan sebagai landasan yang kuat dalam pengelolaan stok ikan.
! ! '#%*%0)- )0)/)2
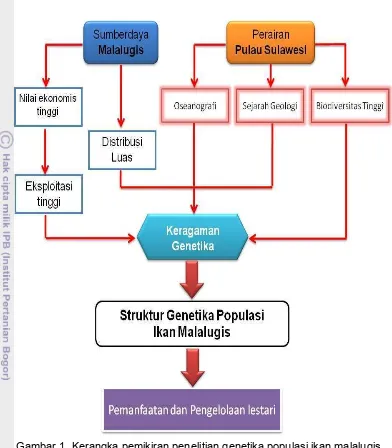
Nilai ekonomis yang tinggi pada ikan malalugis menyebabkan eksploitasi ikan tersebut meningkat. Peningkatan eksploitasi dapat berpengaruh terhadap kelimpahan stok, rata-rata ukuran ikan dan struktur genetika. Disamping itu biodiversitas yang tinggi serta kondisi oseanografi berupa pola arus laut dan variasi tinggi muka air laut di kawasan perairan di pulau Sulawesi dapat mempengaruhi struktur genetika populasi dari suatu organisme.
Gambar 1. Kerangka pemikiran penelitian genetika populasi ikan malalugis.
!4! %5%)- ,)- )-6))(
!7! .&$('0.0
Berdasarkan sifat biologi ikan malalugis dan kondisi geografis serta oseanografi perairan di sekitar pulau Sulawesi, penelitian dilakukan dengan landasan hipotesis sebagai berikut:
a) Struktur populasi ikan malalugis di Laut Sulawesi, Selat Makassar, Laut Maluku dan Teluk Tomini cenderung mempunyai persamaan.
b) Struktur populasi ikan malalugis di Teluk Tolo, Laut Banda, Teluk Bone dan Laut Flores cenderung berbeda dengan keempat populasi yang lain, karena pengaruh dari Laut Arafura.
!
! ! +)- )/)/%".0
Jenis ikan layang di perairan Indonesia terdapat lima jenis spesies, yaitu: layang biasa (Decapterus russelli), layang deles (D. macrosoma), layang ekor merah (D. kurroides dan D. tabl) dan malalugis/layang biru (D. macarellus). Spesies ikan malalugis umumnya tertangkap di perairan laut dalam (deep water species) dengan kadar garam paling rendah 34 per mil (Hariati, 2005). Secara morfologis ikan malalugis hampir sama dengan ikan layang lain, sedikit perbedaan dengan spesies layang lain adalah pada warna yang lebih biru (gelap) (Gambar 2). Klasifikasi ikan malalugis adalah sebagai berikut:
$*).-8Eukaryota - Whittaker & Margulis,1978 - eukaryotes
.-",$*8Animalia - C. Linnaeus, 1758 - animals
%9+.-",$*8Bilateria - (Hatschek, 1888) Cavalier-Smith, 1983
#)-:28Deuterostomia - Grobben, 1908
-6#)+.-",$*8 Chordonia - (Haeckel, 1874) Cavalier-Smith, 1998
2;/%*8Chordata - Bateson, 1885 - Chordates
%9&2;/%*8Vertebrata - Cuvier, 1812 - Vertebrates
-6#)&2;/%*8Gnathostomata - auct. - Jawed Vertebrates
%&'#:/)008Osteichthyes - Huxley, 1880 - Bony Fishes
/)008Actinopterygii - Huxley, 1880 - Ray-Finned Fishes
%9:/)008Actinopterygii - Ray-Finned Fishes
-6#):/)008Actinopteri
$2$#(8Clupeocephala
%&'#$#,'#8Acanthopterygii
#,'#8Perciformes
%9$#,'#8Percoidei
)*./;8Carangidae - Jacks and pompanos
'-%08Decapterus - Berry, 1968
&':.6.: -)*'8macarellus - (Cuvier, 1833)
:.'-(.6.: -)*'8- Decapterus macarellus (Cuvier, 1833)
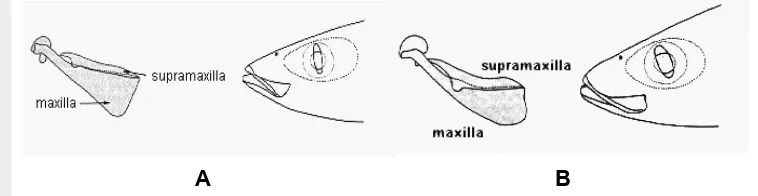
Jenis ikan layang tersebut tertangkap di perairan dengan kedalaman di atas 100 m, antara lain di perairan Selat Malaka bagian utara, Samudera Hindia, Teluk Tomini, Laut Sulawesi dan Laut Banda. Ikan malalugis dan layang deles yang berukuran relatif besar (± 25 cm) sukar dibedakan, karena keduanya mempunyai bentuk dan penampang badan yang hampir sama. Salah satu ciri yang membedakan adalah ikan malalugis tidak bergigi sedangkan layang deles mempunyai gigi-gigi kecil pada rahang bawah (Tarp dan Kailola, 1985). Menurut FAO untuk membedakan antara malalugis dan layang deles bisa dilihat dari bentuk maxilla dan supramaxilla (Gambar 3).
Gambar 3. Bentuk maxilla dan supramaxilla A) ikan malalugis; B) ikan layang deles (FAO).
Analisis contoh isi lambung ikan malalugis di perairan Teluk Tomini diperoleh hasil bahwa makanan ikan malalugis didominasi oleh jenis-jenis fitoplankton terutama Diatomae dan Dinoflagellata serta zooplankton dari kelas Crustacea, Mollusca dan Copepoda (Hariati, 2005). Sesuai dengan sifat hidupnya yang merupakan jenis ikan pelagis, ikan malalugis utamanya tertangkap dengan pukat cincin. Umumnya di setiap daerah penangkapan, ikan malalugis tertangkap sepanjang tahun dengan beberapa puncak hasil tangkapan. Hasil tangkapan tertinggi di perairan Banda Aceh terjadi pada bulan Maret – Mei dan bulan Oktober, sedangkan di perairan ZEE Selat Malaka terjadi pada bulan Juni – Oktober. Perairan Barat Sumatera puncak hasil tangkapa ikan malalugis terjadi pada Maret dan bulan September. Penangkapan ikan pelagis dengan pukat cincin di perairan Teluk Tomini berlangsung sepanjang tahun dan pada bulan Desember – Februari produksi sangat menurun. Pada perairan Parigi dan sekitarnya musim penangkapan berlangsung antara bulan Mei – Oktober, sedangkan di perairan Poso antara bulan Maret – September (Hariati, 2005).
! ! %/)% %/)1'0.
sebelah Barat dan garis Lydekker di sebelah Timur (Gambar 4). Ditinjau dari sejarah geologinya, pulau Sulawesi sangat menarik, karena diduga, di masa lampau, pulau ini tidak pernah bersatu dengan daratan manapun (Hall 2001). Berbeda halnya dengan Sumatra, Jawa, Bali, dan Kalimantan yang pernah bersatu dengan daratan Asia (Sundaland), serta Papua yang pernah bersatu dengan daratan Australia (Sahulland) sebelum kala Pleistosen (Pleistocene) berakhir.
Gambar 4. Peta Wilayah Wallacea. Wallacea merupakan daerah peralihan antara daratan Sunda (Asia) dan daratan Sahul (Australia). (putih = daratan, abu-abu tua = daratan sebelum kala Pleistosen berakhir, abu-abu muda = lautan dalam) ( Whitten et al. 2002)
terdapat diSulawesi, 61% di antaranya bersifat endemik. Sebagai perbandingan, pulau Kalimantan yang mempunyai endemisitas paling tinggi di Sundaland, hanya mempunyai 18% mamallia endemik (Whitten et al. 2002).
Menurut Myers et al. (2000) daerah Wallacea termasuk dalam 25 “hotspot” paling penting untuk konservasi. Daerah ini mempunyai 529 spesies vertebrata endemik (1,9% dari jumlah di dunia). Spesies-spesies tersebut mengalami ancaman yang serius, sebab hanya 15% habitat alami yang masih tersisa. Dari habitat alami yang masih tersisa tersebut, 39,2% di antaranya terdapat dalam kawasan konservasi. Habitat alami yang masih tersisa tersebut hanya akan efektif untuk melindungi biodiversitas di Sulawesi, jika tersebar sesuai dengan distribusi biodiversitas tersebut. Oleh sebab itu upaya konservasi di Sulawesi harus dirancang secara komprehensif.
Profesor John A. Katili, ahli geologi Indonesia yang merumuskan geomorfologi Pulau Sulawesi bahwa terjadinya Sulawesi akibat tabrakan dua pulau (Sulawesi bagian Timur dan Sulawesi bagian Barat) antara 19 sampai 13 juta tahun yang lalu, terdorong oleh tabrakan antara lempeng benua yang merupakan fundasi Sulawesi Timur bersama Pulau-Pulau Banggai dan Sula, yang pada gilirannya merupakan bagian dari lempeng Australia, dengan Sulawesi Barat yang selempeng dengan pulau-pulau Kalimantan, Jawa dan Sumatra, Sulawesi menjadi salah satu wilayah geologis paling rumit di dunia.
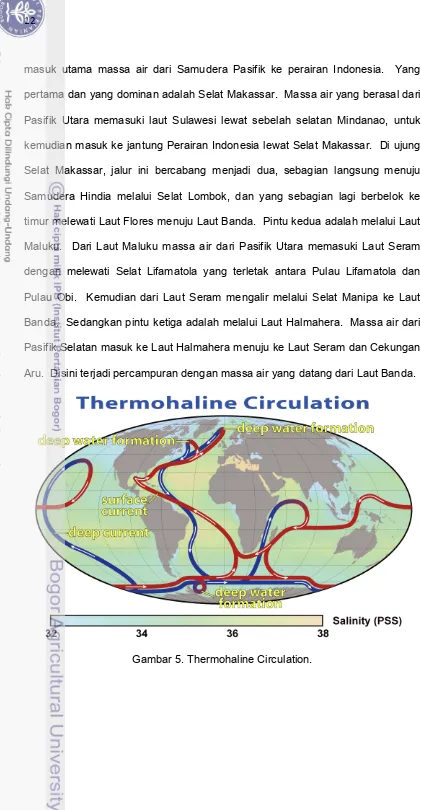
masuk utama massa air dari Samudera Pasifik ke perairan Indonesia. Yang pertama dan yang dominan adalah Selat Makassar. Massa air yang berasal dari Pasifik Utara memasuki laut Sulawesi lewat sebelah selatan Mindanao, untuk kemudian masuk ke jantung Perairan Indonesia lewat Selat Makassar. Di ujung Selat Makassar, jalur ini bercabang menjadi dua, sebagian langsung menuju Samudera Hindia melalui Selat Lombok, dan yang sebagian lagi berbelok ke timur melewati Laut Flores menuju Laut Banda. Pintu kedua adalah melalui Laut Maluku. Dari Laut Maluku massa air dari Pasifik Utara memasuki Laut Seram dengan melewati Selat Lifamatola yang terletak antara Pulau Lifamatola dan Pulau Obi. Kemudian dari Laut Seram mengalir melalui Selat Manipa ke Laut Banda. Sedangkan pintu ketiga adalah melalui Laut Halmahera. Massa air dari Pasifik Selatan masuk ke Laut Halmahera menuju ke Laut Seram dan Cekungan Aru. Disini terjadi percampuran dengan massa air yang datang dari Laut Banda.
!4! (#%+(%# '-'(.+) $&%/)0.
Genetika (dari bahasa Yunani γέννω atau genno yang berarti “melahirkan”) merupakan ilmu dari cabang biologi yang mempelajari berbagai aspek yang menyangkut pewarisan sifat dan variasi sifat ada organisme maupun suborganisme (Griffiths et al., 2000). Genetika populasi menghabiskan sebagian besar waktunya untuk melakukan salah satu dari dua hal: menggambarkan struktur genetik populasi atau berteori pada tekanan evolusi yang bekerja pada populasi (Gillespie, 1998). Windelspecht (2007) menerangkan bahwa seorang ahli genetika populasi mempelajari bagaimana frekuensi dari perubahan alel dalam suatu populasi dari waktu ke waktu, biasanya dalam menanggapi atau merespon suatu tekanan yang selektif. Dalam hal ini, ahli genetika populasi umumnya kurang tertarik dalam mempelajari mekanisme molekuler regulasi gen, melainkan menggunakan matematika dan statistik untuk menggambarkan perubahan populasi.
Struktur populasi, sekelompok individu atau sub kelompok dari suatu spesies yang memiliki kesamaan struktur atau pola genetika (genetic pool), dapat dipelajari berdasarkan frekuensi genetika dari setiap gen yang terlibat dalam ekspresi fenotipik. Pada tingkat molekuler (DNA) ikan laut menunjukkan variabilitas genetik walaupun dalam derajat yang lebih rendah disbanding ikan air tawar baik pada level supraspesifik maupun taksa kelompok individu (populasi dan sub-populasi) dimana pada tingkat protein (studi allozyme) tidak terlihat. Menurut Graves et al. in Saunders et al. (1986), terdapat variabilitas genetika yang disebabkan oleh aliran gen (gene flow) inter-oseanik serta menimbulkan diferensiasi genetika.
populasi (variabilitas intrapopulasi) dan variabilitas antar populasi (divergensi interpopulasi). Variabilitas intrapopulasi dinyatakan dengan parameter diversitas haplotipe atau diversitas nucleon (h), banyaknya neukleomorf (unit polimorfisme pada nucleon yang terdapat dalam bentuk pola situs restriksi), jumlah rata-rata perbedaan situs restriksi, jumlah segregasi situs restriksi atau jumlah situs restriksi polimorfis dalam sejumlah sampel nukleon. Nukleon merupakan suatu segmen DNA, identik dengan gen dalam DNA ini (nuclear DNA), yang dicirikan oleh peta situs restriksi, atau jumlah dan ukuran fragmen DNA. Divergensi interpopulasi dipelajari berdasarkan parameter jarak genetika (δ) dan analisis terhadap perbedaan situs restriksi (Nei dan Tajima, 1981). Nei dan Tajima (1981) menambahkan bahwa variabilitas genetika nucleon berhubungan dengan laju mutasi per-nukleon dimana perubahan situs restriksi terjadi secara evolusioner dan disebabkan oleh substitusi, insersi (insertion) atau penghapusan (deletion) nukleotida. Jenis dan jumlah enzim restriksi yang sama digunakan dalam analisis sampel untuk membandingkan nilai parameter diantara nukleon atau organisme yang berbeda.
!7! $&%/)0. )/)* #(. '-'(.+)
Untuk mempelajari pola pewarisan sifat pada tingkat populasi terlebih dahulu perlu difahami pengertian populasi dalam arti genetika atau lazim disebut juga &$&%/)0. '-,'/.)-. Populasi mendelian ialah sekelompok individu suatu spesies yang bereproduksi secara seksual, hidup di tempat tertentu pada saat yang sama, dan di antara mereka terjadi perkawinan (interbreeding) sehingga masing-masing akan memberikan kontribusi genetik ke dalam /%-"+)-"
"'-, yaitu sekumpulan informasi genetik yang dibawa oleh semua individu di dalam populasi.
Deskripsi susunan genetik suatu populasi mendelian dapat diperoleh apabila kita mengetahui macam genotipe yang ada dan juga banyaknya masing-masing genotipe tersebut. Sebagai contoh, di dalam populasi tertentu terdapat tiga macam genotipe, yaitu AA, Aa, dan aa. Maka, proporsi atau persentase genotipe AA, Aa, dan aa akan menggambarkan susunan genetik populasi tempat mereka berada. Adapun nilai proporsi atau persentase genotipe tersebut dikenal dengan istilah 6#'+%'-0. "'-$(.&'. Jadi, frekuensi genotipe dapat dikatakan sebagai proporsi atau persentase genotipe tertentu di dalam suatu populasi! Dengan perkataan lain, dapat juga didefinisikan bahwa frekuensi genotipe adalah proporsi atau persentase individu di dalam suatu populasi yang tergolong ke dalam genotipe tertentu. Pada contoh di atas jika banyaknya genotipe AA, Aa, dan aa masing-masing 30, 50, dan 20 individu, maka frekuensi genotipe AA = 0,30 (30%), Aa = 0,50 (50%), dan aa = 0,20 (20%).
melangsungkan perkawinan sehingga terjadi transmisi gen dari generasi ke generasi. Dalam proses transmisi ini, genotipe tetua (parental) akan dibongkar dan dirakit kembali menjadi genotipe keturunannya melalui segregasi dan rekombinasi gen-gen yang dibawa oleh tiap gamet yang terbentuk, sementara gen-gen itu sendiri akan mengalami kesinambungan (kontinyuitas). Dengan demikian, deskripsi susunan genetik populasi dilihat dari gen-gen yang terdapat di dalamnya sebenarnya justru lebih bermakna bila dibandingkan dengan tinjauan dari genotipenya.
Susunan genetik suatu populasi ditinjau dari gen-gen yang ada dinyatakan sebagai 6#'+%'-0. "'-, atau disebut juga 6#'+%'-0. )/'/, yaitu proporsi atau persentase alel tertentu pada suatu lokus. Jika kita gunakan contoh perhitungan frekuensi genotipe tersebut di atas, maka frekuensi alelnya dapat dihitung sebagai berikut.
AA Aa aa Total
Banyaknya individu 30 50 20 100
Banyaknya alel A 60 50 - 110
Banyaknya alel a - 50 40 90
Karena di dalam tiap individu AA terdapat dua buah alel A, maka di dalam populasi yang mempunyai 30 individu AA terdapat 60 alel A. Demikian juga, karena tiap individu Aa membawa sebuah alel A, maka populasi yang mempunyai 50 individu Aa akan membawa 50 alel A. Sementara itu, pada individu aa dengan sendirinya tidak terdapat alel A, sehingga secara keseluruhan banyaknya alel A di dalam populasi tersebut adalah 60 + 50 + 0 = 110. Dengan cara yang sama dapat dihitung banyaknya alel a di dalam populasi, yaitu 0 + 50 + 40 = 90. Oleh karena itu, frekuensi alel A = 110/200 = 0,55 (55%), sedang frekuensi a = 90/200 = 0,45 (45%).
Frekuensi alel berkisar dari 0 hingga 1. Suatu populasi yang mempunyai alel dengan frekuensi = 1 dikatakan mengalami fiksasi untuk alel tersebut.
Frekuensi alel pada suatu populasi spesies organisme dapat dihitung atas dasar data elektroforesis protein/enzim atau zimogram yang menampilkan pita-pita sebagai gambaran mobililitas masing-masing polipeptida penyusun protein (Gambar 6). Elektroforesis merupakan teknik pemisahan molekul yang berbeda-beda ukuran dan muatan listriknya. Oleh karena itu, molekul-molekul yang akan dipisahkan tersebut harus bermuatan listrik seperti halnya protein dan DNA.
Jarak migrasi (cm)
4 3 2 1
Individu 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Genotipe CL LL LL CL CL CL LL CL CL CL LL CL LL LL CL Gambar 6. Zimogram esterase dari ikan sidat (Anguilla sp) di kawasan Segara
Anakan, Cilacap.
yang sama molekul berukuran besar akan menempuh jarak migrasi yang lebih pendek daripada jarak migrasi molekul berukuran kecil.
Pola pita seperti pada zimogram esterase di atas sebenarnya merupakan gambaran fenotipe, bukan genotipe. Namun, analisis variasi fenotipe terhadap kebanyakan enzim pada berbagai macam organisme sering kali dapat memberikan dasar genetik secara sederhana. Seperti diketahui, tiap enzim dapat mengandung sebuah polipeptida atau lebih dengan susunan asam amino yang berbeda sehingga menghasilkan fenotipe berupa pita-pita dengan mobilitas yang berbeda. Variasi fenotipe ini disebabkan oleh perbedaan alel yang menyusun genotipe.
Jika alel-alel yang menyebabkan perbedaan polipeptida pada enzim tertentu terletak pada suatu lokus, maka bentuk alternatif enzim yang diekspresikannya dikenal sebagai )/$<.*. Alel yang mengatur alozim biasanya bersifat kodominan, yang berarti dalam keadaan heterozigot kedua-duanya akan diekspresikan. Dengan demikian, individu pada Gambar 15.1 yang menampilkan pita lambat dan pita cepat (nomor 1, 4, 5, 6, 8, 9, 10, 12, dan 15) memiliki genotipe heterozigot, yaitu CL (C=cepat; L=lambat). Sementara itu, individu yang hanya menampilkan pita lambat (nomor 2, 3, 7, 11, 13, dan 14) adalah homozigot LL. Begitu pula individu dengan hanya satu pita cepat (kebetulan pada zimogram tersebut tidak ada) dikatakan mempunyai genotipe homozigot CC.
! ! %+%* '0'.*9)-")- )#,; '.-9'#"
Populasi mendelian yang berukuran besar sangat memungkinkan terjadinya kawin acak (panmiksia) di antara individu-individu anggotanya. Artinya, tiap individu memiliki peluang yang sama untuk bertemu dengan individu lain, baik dengan genotipe yang sama maupun berbeda dengannya. Dengan adanya sistem kawin acak ini, frekuensi alel akan senantiasa konstan dari generasi ke generasi. Prinsip ini dirumuskan oleh G.H. Hardy, ahli matematika dari Inggris, dan W.Weinberg, dokter dari Jerman,. sehingga selanjutnya dikenal sebagai 2%+%* +'0'.*9)-")- )#,; '.-9'#".
Di samping kawin acak, ada persyaratan lain yang harus dipenuhi bagi berlakunya hukum keseimbangan Hardy-Weinberg, yaitu tidak terjadi migrasi, mutasi, dan seleksi. Dengan perkatan lain, terjadinya peristiwa-peristiwa ini serta sistem kawin yang tidak acak akan mengakibatkan perubahan frekuensi alel.
Deduksi terhadap hukum keseimbangan Hardy-Weinberg meliputi tiga langkah, yaitu (1) dari tetua kepada gamet-gamet yang dihasilkannya, (2) dari penggabungan gamet-gamet kepada genotipe zigot yang dibentuk, dan (3) dari genotipe zigot kepada frekuensi alel pada generasi keturunan. Secara lebih rinci ketiga langkah ini dapat dijelaskan sebagai berikut.
Kembali kita misalkan bahwa pada generasi tetua terdapat genotipe AA, Aa, dan aa, masing-masing dengan frekuensi P, H, dan Q. Sementara itu, frekuensi alel A adalah p, sedang frekuensi alel a adalah q. Dari populasi generasi tetua ini akan dihasilkan dua macam gamet, yaitu A dan a. Frekuensi gamet A sama dengan frekuensi alel A (p). Begitu juga, frekuensi gamet a sama dengan frekuensi alel a (q).
frekuensi genotipe sebagai hasil kali frekuensi gamet yang bergabung. Pada Tabel 1 terlihat bahwa tiga macam genotipe zigot akan terbentuk, yakni AA, Aa, dan aa, masing-masing dengan frekuensi p2, 2pq, dan q2.
Tabel 1. Pembentukan zigot pada kawin acak Gamet-gamet Ε
Oleh karena frekuensi genotipe zigot telah didapatkan, maka frekuensi alel pada populasi zigot atau populasi generasi keturunan dapat dihitung. Fekuensi alel A = p2 + ½ (2pq) = p2 + pq = p (p + q) = p. Frekuensi alel a = q2 + ½ (2pq) = q2 + pq = q (p + q) = q. Dengan demikian, dapat dilihat bahwa frekuensi alel pada generasi keturunan sama dengan frekuensi alel pada generasi tetua.
perbedaan frekuensi golongan darah sistem ABO yang terlihat sangat nyata antara ras yang satu dan lainnya.
Laju migrasi dapat didefinisikan sebagai proporsi atau persentase alel tertentu di dalam suatu populasi yang digantikan oleh alel migran pada tiap generasi. Sebagai contoh, jika pada tiap generasi sebanyak 80 dari 1000 ekor ikan normal digantikan oleh ikan albino, maka dikatakan bahwa laju migrasinya 0,08 atau 8%.
Secara matematika, hubungan antara perubahan frekuensi alel dan laju migrasi dapat dilihat sebagai persamaan berikut ini.
pn - P = (po - P)(1 - m)n
pn = frekuensi alel pada populasi yang diamati setelah n generasi migrasi
P = frekuensi alel pada populasi migran
po = frekuensi alel pada populasi awal (sebelum terjadi migrasi)
m = laju migrasi n = jumlah generasi
Faktor lain yang dapat menyebabkan terjadinya perubahan frekuensi alel adalah mutasi. Namun, peristiwa yang sangat mendasari proses evolusi ini sebenarnya tidak begitu nyata pengaruhnya dalam perubahan frekuensi alel. Hal ini terutama karena laju mutasi yang umumnya terlalu rendah untuk dapat menyebabkan terjadinya perubahan frekuensi alel. Selain itu, individu-individu mutan biasanya mempunyai daya hidup (viabilitas), dan juga tingkat kesuburan (fertilitas), yang rendah.
Hubungan matematika antara laju mutasi dan perubahan frekuensi alel dapat dirumuskan seperti pada contoh berikut ini. Misalnya, di dalam suatu populasi terdapat alel A dan a, masing-masing dengan frekuensi awal po dan qo.
Mutasi berlangsung dari A ke a dengan laju mutasi sebesar u. Sebaliknya, laju mutasi alel a menjadi A adalah v. Dengan demikian, perubahan frekuensi alel A akibat mutasi adalah ∆p = vqo - upo, sedang perubahan frekuensi alel a akibat
mutasi adalah ∆q = upo - vqo.
Ketika dicapai keseimbangan di antara kedua arah mutasi tersebut nilai ∆p dan ∆q adalah 0. Oleh karena itu, vqo = upo, atau secara umum vq = up. Jika
persamaan ini dielaborasi, maka akan didapatkan p = v/(u + v) dan q = u/(u + v).
!=! *.($:2$-,#.) *(
Beberapa teknik eksplorasi dapat dimanfaatkan oleh para biologiwan untuk menggali informasi yang terkandung dalam genom mitochondria. Teknik eksplorasi yang pertama didasarkan pada sekuen, basa-basa penyusun genom mitochondria. Hal ini memberikan informasi yang sangat lengkap mengenai urutan basa-basanya, namun cara ini membutuhkan tenaga dan biaya yang sangat besar. Alternatif lain yang Iebih efisien dari teknik pertama tadi ialah analisis hanya bagian tertentu dari genom mitochondria. Berkat penemuan Saiki et al. (1985, 1988) mengenai amplifikasi DNA dengan PCR (Polymerase Chain Reaction), analisis genom mitochondria secara partial dapat dilakukan dengan mudah. Teknik eksplorasi yang kedua didasarkan pada pnggunaan enzim restriksi (restriction endonucleases) untuk membandingkan genom mitochondria antar individu maupun takson. Pemotongan dengan enzim restriksi yang berbada akan menghasilkan potongan DNA yang berbeda dari DNA yang sama. Perbedaan genom mitochondria dapat dibandingkan hanya dangan perbandingan jumlah dan ukuran fragmen-fragmen yang dipotong oleh enzim restriksi tersebut. Variasi yang dihasilkan oleh perbedaan panjang fragmen yang dipotong oleh enzim restriksi ini dikenal sebagai Restriction Fragment Length Polymorphism (RFLP). Pendekatan ini relatif sederhana dan mampu memberikan informasi dari semua bagian genom mitochondria. Setiap situs restriksi dapat dipetakan pada molekul DNA mitochondria sehingga menghasilkan peta situs restriksi DNA mitochondria (Solihin, 1994).
dari genom mitochondria berevolusi dengan kecepatan yang berbeda. Diketahui bahwa tingkat evolusi dari suatu gen atau bagian dari DNA merupakan faktor penting yang menentukan penggunaan penanda DNA dalam studi sistematika dan biogeografi. Gen-gen yang terkonservasi dengan baik dapat dijadikan sebagai dasar penelusuran kesamaan asal muasal (ancient taxa), sedangkan gen-gen yang tak terkonservasi dengan baik yaitu gen-gen yang berevolusi dengan cepat dapat digunakan untuk perbandingan galur-galur baru. Secara umum evolusi sekuen DNA mitochondria lebih cepat 5 sampai 10 kali dari genom inti (Brown et al., 1979) dan bahkan gen tRNA seratus kali lebih cepat dari DNA inti (Brown et al., 1982); (iv) DNA mitochondria hewan tidak memiliki intron ataupun spacer yang berukuran besar antar gennya. Hal inilah yang mebyebabkan ukuran genom mitochondria hewan lebih kecil dibandingkan dengan genom mitochondria tanaman; (v) DNA mitochondria bersifat khusus karena diturunkan melalui induk betinanya tanpa mengalami rekombinasi (strict maternal inheritance). Akibatnya afinitas genetik yang diatur oleh genom mitochondria merupakan refleksi dari Phylogeni matriarcale; vi) DNA mitochondria sangat polimorf, baik untuk intrapopulasi maupun untuk interspesies (Solihin, 1994).
!>! -)/.0.0
dianalisis menurut cara RFLP (Restriction Fragment Length Polymorphism) atau RAPD (Random Amplified Polymorphic DNA) sesuai dengan tujuannya. Metode RFLP melihat perbedaan profil dan panjang fragmen DNA dari individu yang berbeda berdasarkan hasil pemotongan enzim restriksi yang sama; sedangkan metode fingerprinting RAPD memperhatikan perbedaan hasil amplifikasi dari individu yang berbeda dengan menggunakan ‘primer’ tertentu.
Tahap ekstraksi dan isolasi merupakan dua tahapan penting yang sangat menentukan keberhasilan analisis DNA. Jenis jaringan yang biasa digunakan dapat berupa hati, otot, sirip, darah, sel kultur dan jaringan lain, baik dalam kondisi segar, telah difiksasi atau beku. Penggunaan jaringan dalam keadaan beku atau diawet biasanya lebih umum dipilih sehubungan dengan prosedur analisa yang harus dilaksankan di tempat yang bersih dengan menggunakan peralatan khusus.
!?! $/;*'#)0' 2).-
'):(.$-Teknik PCR ditemukan tahun 1985 oleh Dr. Kary Mullis di Cetus Corporation, dan telah berkembang pesat dalam menunjang revolusi besar dalam bidang biologi molekuler (Zyskind dan Bernstein, 1993). Awalnya Mullis menggunakan enzim polymerase DNA I (fragmen Klenov) pada tahap ekstensi polinukleotida primer, tetapi karena fragmen Klenov tidak stabil pada suhu tinggi, maka diganti dengan Taq DNA-polymerase, suatu enzim yang dihasilkan oleh bakteri thermofilik yang tahan terhadap suhu tinggi tanpa kehilangan aktivitasnya dan tetap aktif selama siklus PCR, suhu optimum 75oC. Teknik ini dikembangkan untuk menghasilkan sekuen DNA tertentu dalam jumlah besar tanpa melalui cloning pada sel hidup.
perubahan sifat fisik DNA terhadap suhu. Replikasi terjadi jika terdapat untai tunggal DNA yang bertindak sebagai template (cetakan) dan energi pembangun basa (dNTP). Enzim DNA polymerase akan membantu katalis pembuatan DNA untai lainnya yang merupakan komplemen dari template DAN. Reaksi ini harus dimulai dengan ‘primer’ (pemula), yaitu suatu potongan pendek DNA (oligonukleotida) yang umumnya terdiri dari 20 nukleotida dan disintesa dengan susunan nukleotida spesifik. Masing-masing primer akan berpasangan dengan sekuen tertentu yang mengapit daerah DNA target amplifikasi pada tiap pita DNA.
Siklus pokok PCR berlangsung dalam tiga tahap, yaitu denaturasi template pada suhu tinggi (94 – 97oC), annealing oligonukleutida primer pada suhu 55 – 72oC dan ekstensi DNA-polymerase primer pada ujung 3’ pada suhu 72oC. Siklus diulang sebanyak 25 – 30 kali. Pada tahap denaturasi, untai DNA pilin ganda dibuka melalui pemanasan hingga tiap pita DNA terpisah; annealing adalah pelekatan primaer pada masing-masing untai pita DNA; yang terakhir pada tahap ekstensi, ezim DNA polymerase aktif memperpanjang primer hingga terbentuk untaian pasangan basa sepanjang sekuen DNA target. Dari keseluruhan proses jumlah DNA target yang dihasilkan meningkat secara eksponensial karena template yang baru akan terbentuk pada setiap siklus.
polymerase direduksi melalui denaturasi sekuen pada suhu tinggi. Dalam perkembangan selanjutnya PCR sangat bermanfaat dalam aplikasi sebagai berikut:
Ø Amplifikasi RNA untuk kepentingan deteksi atau cloning
Ø Amplifikasi sekuen yang diapit (flanking) melalui PCR yang dibalik (inverse PCR)
Ø Template bagi sequencing DNA
Ø Aplikasi dalam teknik standard, yaitu riset pada koloni bakteri bagi keperluan screening, penyisipan (insertion) atau penghapusan (deletion) sekuen nukleotida ke dalam sekuen nukleotida, serta untuk menentukan orientasi dan lokasi dari fragmen restriksi.
! ! -<.* '0(#.+0.
Enzim restriksi adalah enzim yang dapat memotong DNA pada sekuesn spesifik sehinggak dihasilkan fragmen-fragmen nukleotida yang lebih pendek dengan ukuran tertentu (enzim pemotong). Enzim restriksi dapat mengenal sekuen 4-basa, 5-basa atau 6-basa, dan akan memotong molekul DNA di tempat manapun yang dikenal. Pemotong 4-basa biasanya menghasilkan 3 – 6 kali lebih banyak fragmen disbanding pemotong 6-basa (Ferris dan Berg, 1987). Dapat dikatakan enzim restriksi merupakan ‘tool’ prinsip dalam analisis genom mtDNA.
(Gyllensten dan Wilson 1987). Pada genus Salmo enzim Mbo I (^GATC) dapat mengenal dan menghasilkan fragmen 25 bp.
4!
4! ! $+)0. ,)- )+(%
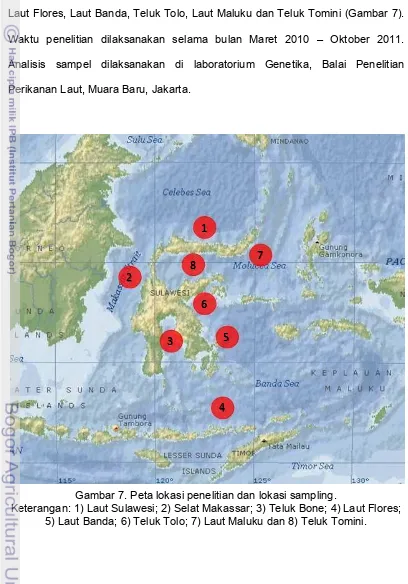
'-'/.(.)-Lokasi penelitian meliputi Laut Sulawesi, Selat Makassar, Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo, Laut Maluku dan Teluk Tomini (Gambar 7). Waktu penelitian dilaksanakan selama bulan Maret 2010 – Oktober 2011. Analisis sampel dilaksanakan di laboratorium Genetika, Balai Penelitian Perikanan Laut, Muara Baru, Jakarta.
Gambar 7. Peta lokasi penelitian dan lokasi sampling.
Keterangan: 1) Laut Sulawesi; 2) Selat Makassar; 3) Teluk Bone; 4) Laut Flores; 5) Laut Banda; 6) Teluk Tolo; 7) Laut Maluku dan 8) Teluk Tomini.
1
2
8
4
3 5
6
7
4! ! )2)- ,)- /)(
'-'/.(.)-Data genetika yang digunakan berupa data polimorfisme fragmen dan situs restriksi yang diperoleh dari hasil analisis RFLP terhadap sekuen teramplifikasi dari genom mtDNA. Genom mtDNA diekstrak dari contoh jaringan daging bagian punggung ikan. Bahan-bahan kimia yang dipakai dalam analisis sampel adalah sebagai berikut:
Pengambilan dan pengawetan sampel: • Ethanol 70%
Ekstraksi dan purifikasi:
• PureLinkTM Genomic DNA Kits - Invitrogen Isolasi dan amplifikasi (PCR):
• Primer TDKD dan PRO yaitu:
primer HN20: 5’ – GTG TTA TGC TTT AGT – TAA GC – 3’, dan primer LN20: 5’ – ACC ACT AGC ACC CAA AGC TA – 3’. • H2O
• Pure taq ready to-go PCR beads • Bromophenol
Restriksi:
• 6 jenis enzim restriksi, terdiri dari: Mbo I, Alu I, Hind III, Taq I, Rsa I dan
Xba I.
• Buffer dari masing-masing enzim Elektroforesis:
• Bubuk agarose • SYBR SafeTM
• Loading bromophenol • Marker DNA
Visualisasi: • H2O
Peralatan yang digunakan dalam analisis DNA dengan metode RFLP adalah: gelas beker, pinset, pipet mikro, tube mikro, erlenmeyer, hot plate stirrer, gelas ukur, inkubator, sentrifuge, mesin PCR, Vortex, elektroforesis sistem, Blue Light transilluminator dan kamera.
4!4! '-")*9./)- )*&'/
Sampel yang diambil berupa jaringan ikan D. macarellus hasil tangkapan nelayan one day fishing (1 hari operasi) yang beroperasi/melakukan penangkapan pada lokasi penelitian. Sampel diambil secara acak pada setiap lokasi dengan perincian sebagai berikut: Laut Sulawesi 8 ekor, Selat Makassar 13 ekor, Teluk Bone 10 ekor, Laut Flores 10 ekor, Laut Banda 9 ekor, Teluk Tolo 8 ekor, Laut Maluku 8 ekor dan Teluk Tomini 8 ekor. Pada setiap ikan diambil jaringan daging bagian punggung. Spesimen (sampel jaringan ikan) disimpan dalam tabung mikro (micro tube) berukuran 1.500 µl dan diawet dengan ethanol 70% kemudian dibawa ke laboratorium untuk dianalisis lebih lanjut.
4!7! -)/.0.0 )*&'/
ekstraksi jaringan daging bagian punggung ikan. Analisis dilaksanakan melalui tahapan sebagai berikut :
Ekstraksi dan purifikasi Amplifikasi (PCR) Digesti (restriksi) Elektroforesis Visualisasi.
Dalam amplifikasi digunakan 2 macam ‘primer’, yaitu primer TDKD dan PRO; sedang enzim restriksi yang dipakai 6 jenis, yaitu Mbo I, Alu I, Hind III, Taq I, Rsa I
dan Xba I.
DNA ikan diekstrak dari 5-10 mg daging bagian punggung dengan menggunakan metode mini column, sebagai berikut: potongan daging ikan dimasukkan kedalam tabung 1,5 ml yang telah berisi 180 µl larutan PureLink Genomic Digestion Buffer dan 20 µl Proteinase K, kemudian diaduk dengan menggunakan vortex dan diinkubasi pada suhu 55oC sampai potongan daging/sirip hancur (1 – 4 jam). Setelah potongan daging/sirip hancur dilakukan proses sentrifuge dengan kecepatan maksimum (15.000 rpm) selama 3 menit sehingga diperoleh supernatan (lysate). Kemudian lysate tersebut dipindahkan ke dalam tabung mikrosentrifuge yang baru.
Lysate (~640 µl) dipindahkan ke dalam PureLink Spin Column dan disentrifuge pada kecepatan 10.000 rpm selama 1 menit pada suhu ruangan. Collection tube (tube bagian bawah) diganti dengan PureLink Collection Tube yang baru (memindahkan mini column ke collection tube yang baru). Kemudian ditambahkan 500 µl larutan Wash Buffer 1 dan disentrifuge dengan 10.000 rpm selama 1 menit. Collection tube diganti lagi dengan yang baru lalu ditambahkan 500 µl larutan Wash Buffer 2 dan disentrifuge dengan kecepatan maksimum selama 3 menit pada suhu ruangan.
Spin column dipindahkan ke dalam mikrotube 1,5 ml baru dan steril lalu ditambahkan 25 – 200 µl larutan PureLink Genomic Elution Buffer ke dalam colomn. Kemudian diinkubasi dalam suhu ruangan selama 1 menit lalu disentrifuge dengan kecepatan maksimum selama 1 menit. Cairan yang ada dalam mikrotube 1,5 ml tersebut telah mengandung DNA yang telah dipurifikasi dan akan dilakukan proses selanjutnya yaitu Amplifikasi. Kemudian column dibuang.
Primer yang digunakan untuk mengamplifikasi daerah control region mtDNA D-loop adalah:
- primer HN20: 5’ – GTG TTA TGC TTT AGT – TAA GC – 3’, dan - primer LN20: 5’ – ACC ACT AGC ACC CAA AGC TA – 3’.
Produk DNA yang diperoleh kemudian direstriksi dengan menggunakan enzim restriksi (restriction endonuklease) sesuai dengan prosedur standar perusahaan. Hasil restriksi kemudian dipisahkan secara elektroforesis dengan menggunakan gel agarose 1,5% dalam TBE-buffer (Tris-Boric-EDTA) dan diamati dengan blue-light illuminator serta di cetak gambarnya dengan kamera SLR.
4! ! -('#&#'()0. ,)- -)/.0.0 )()
7!
7! ! '#)")*)- )
Panjang sekue dari hasil amplifikasi (P LN20 adalah sekitar 1 kasi (PCR) dengan menggunakan pasangan prim
itar 1.000 bp (base pairs). Panjang sekuens ini yang diperoleh pada penelitian ikan malalugis y l. (1999) pada perairan Laut Maluku dan Laut B sangan primer yang sama. Penelitian yang dilak terhadap spesies D. russelli dan D. macro asangan primer HN20 dan LN20 juga menghas
.000 bp. Hal ini mengindikasikan bahwa pasanga akan primer spesifik untuk ikan genus Decapterus
oh visualisasi hasil pemotongan sekuen mtDNA biru (D.macarellus) oleh enzim Mbo I dan Rsa
enzim restriksi (Alu I, Hind III, Mbo I, Rsa I, Ta untuk memotong sekuens mtDNA ikan malalug
dapat teridentifikasi 6 jenis alel atau composite haplotype, yaitu AAAAAA, AAAABA, AAAAAB, BAABAC, BABCAC dan BAABCC (Tabel 2.).
Tabel 2. Frekuensi haplotipe ikan malalugis (D. macarellus) hasil restriksi
Nilai keragaman haplotipe (haplotype diversity) ikan malalugis yang diperoleh berkisar antara 0 – 0,3698, dengan nilai terendah (0) pada populasi Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo dan Laut Maluku (Tabel 3.). Tingkat keragaman haplotipe ikan malalugis ini lebih rendah jika dibandingkan dengan nilai keragaman ikan laut pada umumnya yaitu berkisar antara 0,600 – 0,900 (Nugroho et al., 2001). Menurut Avise et al. (1989) dalam Tabata et al. (1997) menyebutkan bahwa keragaman haplotipe keseluruhan mtDNA untuk beberapa ikan berada dalam kisaran 0,473 – 0,998. Rendahnya nilai keragaman haplotipe pada ikan malalugis menunjukkan keragaman genetika yang rendah pula. Rendahnya keragaman genetika mengindikasikan bahwa sumberdaya dari ikan malalugis terjadi perubahan struktur genetika menjadi lebih seragam. Diduga ada beberapa faktor yang menyebabkan terjadinya perubahan ini. Faktor pertama adalah bahwa ikan malalugis merupakan ikan layang yang bersifat oseanik dan peruaya sehingga mempunyai jalur migrasi yang cukup luas. Hal ini dapat menyebabkan terjadinya silang dan percampuran genetika antar populasi, sehingga seiring berjalannya waktu, variasi/keberagaman ikan malalugis menjadi berkurang.
merugikan terhadap fekunditas yang potensial; mengurangi rata-rata ukuran memijah; mengubah rasio jenis kelamin dan keseimbangan interspesifik; serta hilangnya diversitas genetika. Terutama pada unit-unit populasi yang mempunyai keragaman genetika sangat rendah (h=0) seperti pada populasi Laut Flores, Laut Banda dan Laut Sulawesi, unit-unit populasi tersebut sangat rentan terhadap perubahan-perubahan (penangkapan, alam).
Menurut Zein (2007) menyatakan bahwa, eksploitasi dapat menyebabkan peningkatan laju genetic drift, selain itu, populasi yang kecil cenderung akan terjadi kawin silang dalam (inbreeding), sehingga hal tersebut dapat berdampak buruk terhadap kelangsungan hidup dari populasi yang ada. Indikasinya adalah turunnya keragaman genetik dalam populasi, yaitu turunnya keragaman haplotipe dan keragaman nukleotida (Zein, 2007). Adanya penurunan variabilitas genetik ini dapat membahayakan kelangsungan hidup populasi karena dapat mengurangi kemampuan individual dalam menghadapi tekanan seleksi alamiah, terutama akibat perubahan lingkungan (Hedrick, 2000).
Nilai tertinggi keragaman haplotipe ikan Malalugis pada penelitian ini terdapat pada populasi Laut Sulawesi, yaitu sebesar 0,3698 (Tabel 3). Tingginya nilai keragaman haplotipe diduga disebabkan oleh adanya pengaruh dari populasi yang berasal dari Samudera Pasifik, atau bisa juga karena spesies ikan Malalugis di Laut Sulawesi mempunyai sifat dapat beradaptasi secara plasticity (kelenturan). Pengaruh dari populasi yang berasal dari Samudera Pasifik diduga disebabkan oleh adanya arus yang mengalir dari Samudera Pasifik menuju Laut Sulawesi melalui percabangan Selatan Mindanao ke arah Baratdaya. Arus tersebut dibelokkan ke selatan kemudian ketika sampai di bagian tengah laut dibelokkan ke Timur dan kembali mengalir ke Timur di sepanjang pantai Utara Sulawesi (Hasanudin, 1998). Walaupun pengaruh arus terhadap ikan dewasa masih diperdebatkan, akan tetapi arus tersebut berpengaruh terhadap larva ikan. Suatu spesies yang mampu beradaptasi secara plasticity akan menghasilkan variasi baik fenotip maupun genotip sebagai respon terhadap kondisi lingkungan tertentu sehingga dapat meningkatkan kemampuan individu untuk tetap bertahan hidup dan berkembang biak (Sultan, 1987; Taylor dan Aarsen, 1989).
7! ! -)/.0.0
secara umum terdapat perbedaan genetika antara spesies Samudera Pasifik dengan Samudera Hindia.
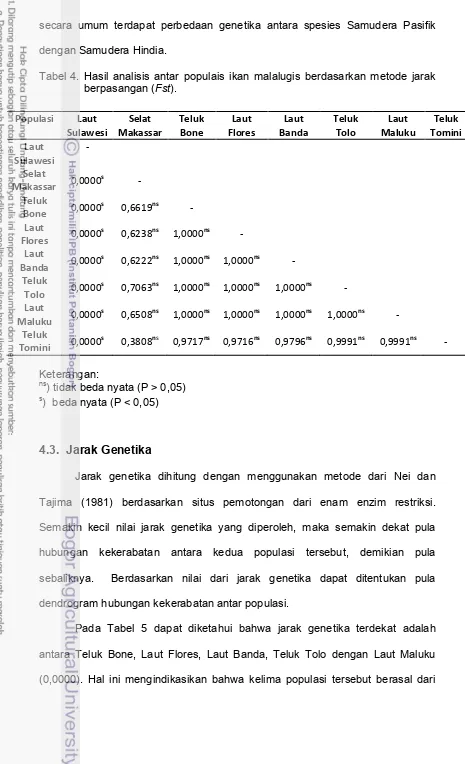
Tabel 4. Hasil analisis antar populais ikan malalugis berdasarkan metode jarak berpasangan (Fst).
Jarak genetika dihitung dengan menggunakan metode dari Nei dan Tajima (1981) berdasarkan situs pemotongan dari enam enzim restriksi. Semakin kecil nilai jarak genetika yang diperoleh, maka semakin dekat pula hubungan kekerabatan antara kedua populasi tersebut, demikian pula sebaliknya. Berdasarkan nilai dari jarak genetika dapat ditentukan pula dendrogram hubungan kekerabatan antar populasi.
asal atau stok populasi yang sama, disamping itu secara geografis ketiga populasi tersebut saling berhubungan, sehingga memungkinkan terjadinya gene flow (aliran gen). Arnaud et al. (1999) menyebutkan bahwa pada populasi Laut Banda dengan Laut Maluku terjadi gene flow terhadap spesies ikan malalugis.
Populasi Teluk Tomini, Selat Makassar dan Laut Sulawesi mempunyai nilai jarak genetika dengan populasi lainnya. Akan tetapi populasi Teluk Tomini dan Selat Makassar jarak genetiknya sangat dekat dengan populasi Laut Maluku, Teluk Tolo, Laut Banda, Laut Flores dan Teluk Bone hal ini menunjukkan bahwa populasi dari Teluk Tomini dan Selat Makassar mempunyai kekerabatan yang dekat dengan kelima populasi tersebut. Jarak terjauh diketahui berasal dari populasi Laut Sulawesi dengan ketujuh populasi lainnya (0,1405 – 0,1733), sehingga hal ini semakin memperkuat indikasi bahwa populasi Laut Sulawesi berbeda dengan tujuh populasi yang lain.
Tabel 5. Jarak genetika Nei antar populasi ikan malalugis
7!7! (#%+(%# '-'(.+) $&%/)0. ,)- %9%-")- ./$"'-'(.+
Berdasarkan dendrogram hubungan kekerabatan dari kedelapan populasi ikan malalugis dapat dipisahkan menjadi dua group populasi yang berasal dari dua garis keturunan mtDNA, yaitu "#$%& &'#()*) (clade 1) terdiri dari populasi Selat Makassar, Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo, Laut Maluku dan Teluk Tomini; sedangkan "#$%& +' ,%) (clade 2) terdiri dari populasi Laut Sulawesi (Gambar 9). Diduga populasi group pertama berasal atau terpengaruh dari populasi Samudera Hindia, sedangkan populasi group kedua berasal atau terpengaruh dari populasi Samudera Pasifik. Gordon dan Fine (1996) menyebutkan bahwa sampai saat ini terjadi pertukaran gen antara organisme tropis di Samudera Hindia dengan Samudera Pasifik. Dalam populasi group pertama terbagi menjadi dua sub populasi, yaitu sub populasi Selat Makassar dengan sub populasi Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo, Laut Maluku dan Teluk Tomini. Sub populasi dari Selat Makassar secara statistik (analisis berpasangan) dan jarak genetika termasuk dalam populasi group pertama, akan tetapi ada sedikit perbedaan pada sub populasi Selat Makassar tersebut. Perbedaan tersebut diduga karena masuknya ikan dari populasi group ke-dua (Laut Sulawesi) ke Selat Makassar melalui perantaraan arus permukaan laut yang bergerak dari Laut Sulawesi ke Selat Makassar sepanjang tahun (Gambar 10 dan Gambar 11). Hal tersebut menyebabkan dua kemungkinan, yang pertama adalah ikan dari populasi Laut Sulawesi bercampur dengan ikan berhaplotipe AAAAAA di Selat Makassar, sehingga memunculkan haplotipe baru.
haplotipe baru. Gaylord dan Gaines (2000) menjelaskan bahwa arus laut dapat mempengaruhi distribusi populasi, perubahan atau perbedaan karakteristik air yang dapat mempengaruhi fisiologi organisme untuk selanjutnya mempengaruhi struktur genetikanya.
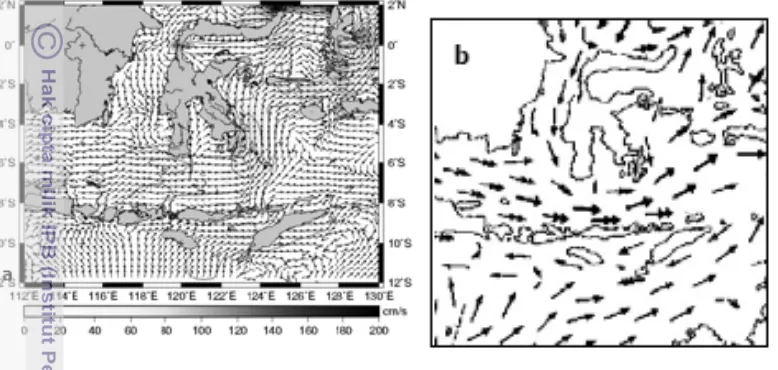
Gambar 10. Pola arus air permukaan laut pada musim barat, a. Rizal et al. (2009) dan b. Wrytki (1961)
Gambar 11. Pola arus air permukaan laut pada musim timur, a. Rizal et al. (2009) dan b. Wrytki (1961)
Tomini sedikit berbeda dengan lima populasi lainnya (Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo dan Laut Maluku) tapi masih mempunyai kekerabatan yang dekat. Adanya sedikit perbedaan tersebut diduga di perairan Teluk Tomini terdapat populasi ikan malalugis yang bersifat lokal. Populasi di perairan Teluk Tomini yang bersifat lokal tersebut disebabkan perairan Teluk Tomini bersifat semi tertutup (semi enclosed), sehingga memungkinkan spesies dalam perairan tersebut terisolasi. Disamping itu, dalam sejarah geologi Pulau Sulawesi di masa lampau (zaman Pleistosen) tidak pernah bersatu dengan daratan manapun (Hall, 2001), Pulau Sulawesi merupakan pulau yang berdiri sendiri dan bukan merupakan pecahan dari pulau lain, seperti Pulau Kalimantan dan Sumatera.
peralihan I perairan Selat Makassar cenderung lebih subur dari pada perairan timur Sulawesi (Laut Maluku, Laut Banda dan Laut Flores) (Gambar 12). Faktor eksternal adalah arus laut, terutama arus permukaan laut, karena arus laut mempunyai peranan penting dalam penyebaran larva-larva ikan (Fahmi, 2010). Seperti pada keterangan sebelumnya, masuknya populasi ikan Laut Sulawesi ke Selat Makassar diduga karena arus. Sub populasi Teluk Tomini juga didominasi ikan malalugis berhaplotipe AAAAAA (populasi group pertama), hal ini karena masuknya sub populasi ikan Laut Maluku melalui perantaraan arus yang mengalir menuju Teluk Tomini pada musim timur (Burhanuddin et al., 2004) (Gambar 13), selain itu pada daerah sekitar mulut teluk mempunyai kandungan klorofil yang tinggi pada musim timur (BRPL 2005). Persebaran ikan malalugis berhaplotipe AAAAAA pada perairan Laut Flores dan Laut Maluku juga disebabkan oleh faktor mencari makan (internal) dan juga faktor arus (eksternal). Pada Gambar 12 dapat diketahui bahwa kandungan klorofil-a di perairan timur Pulau Sulawesi lebih besar/subur daripada perairan barat Pulau Sulawesi. Sedangkan pola arus di Laut Flores menurut Gambar 10 dan 11 menunjukkan bahwa pada musim barat dan musim timur arus mengalir menuju ke arah timur (Laut Banda), walaupun pada musim timur arus tersebut melemah karena ada pengaruh arus ke arah barat di selatan Pulau Sulawesi.
Gambar 13. Pola arus permukaan laut di Teluk Tomini pada saat musim timur (Burhanuddin et al., 2004).
Gambar 14. Struktur genetika populasi ikan malalugis di perairan sekitar Pulau Sulawesi.
Keterangan:
!
! !
.*&%/)-Terdapat dua kelompok utama pada struktur genetika populasi ikan malalugis di perairan sekitar Pulau Sulawesi, yaitu: kelompok pertama diwakili oleh populasi Selat Makassar, Teluk Bone, Laut Flores, Laut Banda, Teluk Tolo, Laut Maluku dan Teluk Tomini, dan kelompok kedua diwakili oleh populasi Laut Sulawesi. Terdapat perbedaan genetika yang cukup signifikan antara populasi Laut Sulawesi dengan ketujuh populasi lainnya. Keragaman haplotipe (genetika) ikan malalugis di perairan sekitar Pulau Sulawesi termasuk rendah yaitu antara 0 – 0,3698
! !
)#)-1. Teluk Tomini diduga merupakan perairan yang endemik, sehingga diperlukan penelitian lanjutan mengenai konservasi di perairan Teluk Tomini.
2. Ikan malalugis berhaplotipe AAAAAA tersebar luas, sehingga diperlukan penelitian lanjutan mengenai kelimpahan ikan tersebut dan sejauh mana penyebarannya.
Allen GR. 2000. Indo-Pacific coral-reef fishes as indicators of conservation hotspots. Proceedings 9th International Coral Reef Symposium; Bali, Indonesia 23 – 27 October 2000. Vol. 2.
Arnaud S, Bonhomme F, Borsa P. 1999. Mitochondrial DNA analysis of the genetic relationships among population of scad mackerel (Decapterus macarellus, D. macrosoma, and D. russelli) in South-East Asia. Marine Biology 135: 699 – 707.
Avise JC, Bowen BW, Lamb T. 1989. DNA fingerprints from hypervariable mitochondrial genotypes. Molecular Biology Evolution 6:258-269. Beacham TD, Withler RE, Wood CC. 1995. Stock identification of Sockeye
Salmon by means of minisatellite DNA variation. North American J. Fish Management. 15(2): 249 – 265.
Bermingham E. 1990. Mitochondrial DNA and the analysis of fish population structure. In: D. H. Whitmore, Ph.D. (Ed.). Electrophoretic and Isoelectric Focusing Techniques in Fisheries management. CRC Press. Inc. Boca Raton. Florida. 107 – 129.
Bermingham E, Avise JC. 1986. Molecular zoogeography of freshwater fishes in the Southeastern United States. Genetics. 113: 939 – 965.
Borsa P. 2003. Genetic structure of round scad mackerel Decapterus macrosoma (Carangidae) in the Indo-Malay archipelago. Marine Biology 142: 575 - 581.
[BRPL DKP] Balai Riset Perikanan Laut, Departemen Kelautan dan Perikanan. 2005. Teluk Tomini: Ekologi, Potensi Sumberdaya, Profil Perikanan dan Biologi Beberapa Jenis Ikan Ekonomis Penting. Jakarta: BRPL DKP.
Burhanuddin, Supangat A, Sulistiyo B, Rameyo T, Kepel CR. 2004. Profil sumberdaya kelautan dan perikanan Teluk Tomini. Jakarta. BRKP-DKP.
Cross TF, Mills CPR, Courcy Williams M de. 1992. An intensive studi of allozyme variation in freshwater resident and anadromous trout, Salmo trutta L. in western Ireland. Journal of Fish Biology. 40: 25 – 32. Effenberger S, Suchentrunk F. 1999. RFLP analysis of the mitochondrial DNA of
otters (Lutra lutra) from Europe-implication for conservation of flagship spesies. Biological Conservation. 90: 229 – 234.
Fahmi MR. 2010. Phenotypic plastisity kunci sukses adaptasi ikan migrasi: studi kasus ikan sidat. Prosiding Forum Inovasi Teknologi Akuakultur; Bandar Lampung Indonesia 20 – 23 April 2010.