KAJIAN KOMPATIBILITAS ISOLAT
CENDAWAN MIK0RIZ;A ARBUSKULA (CMA) TERHADAP PERTUMBUHAN DUA KLON JATI (Tectona grandis L.f.)
HASIL PERBANYAKAN KULTUR JARINGAN
OLEH :
S U R A Y A
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
Sesungguhnya dalam penciptaan langit dan bumi,
Dan silih bergantinya malam dan siang terdapat tanda-tanda bagi orang-orang yang berfikir, Yakni orang-orang yang mengingat Allah
sambil berdiri, atau duduk atau dalam keadaan berbaring, dan mereka memikirkan tentang penciptaan langit dan bumi,
seraya berkata : Ya Tuhan Kami, Tidaklah Engkau ciptakan ini dengan sia-sia, Maha Suci Engkau,
maka peliharalah kami dari siksa api neraka.
(QS.
AliImran
:190- 191)
Tuntutlah ilmu, sesungguhnya menuntnt ilmu adalah
pendekatan dM kepada
Allah
Azza Wajalla, dan mengajarkannya kepada nrang yang tidak mengetahuhya adalah Sodaqoh. Sesunggahnya itmu pengetahuan menempatkan orangnyadalam kedudukan terhorsmat dan mulia (tinggi). Ilmu pengetahuan addah keindahan bagi ahlinya
di dunk dan di alrhirat. (
MR.
Ar-rabZY )ABSTRAK
SURAYA. Kajian kompatibilitas isolat cendawan mikoriza arbuskula (CMA)
terhadap pertwnbuhan dua klon jati (Tectona gradis L.E) hasil perbanyakan kultur
jaringan. Dibawah bimbingan Bapak Dr. YADI SETIADI, MSc. sebagai Ketua dan
Ibu Ir. CORRY TWN, MSi. sebagai Anggota.
Penelitian telah dilaksanakan sejak bulan Pebruari sampai dengan bulan
September 2001, dengan tujuan untuk (1) Mengetahui interaksi terbaik antara 2 jenis
klon jati (Tectona g r d s L . f ) hasil perbanyakan kultur jaringan dengan beberapa
isolat CMA yang dicobakan, dan (2) Mengetahui jenis isolat yang kompatibel
terhadap klon jati tertentu.
Kegiatan penelitian terdiri dari tahap di persemaian dan analisa. Bahan
tanaman yang digunakan terdiri dari klon jati nomor 03 dan klon jati nomor 21,
sedangkan isolat CMA yang dipakai terdiri dari isolat Glomus etunicatum, Glomus
agregatum, Acaulospora tuberculata dm Myco fer.
Tahapan pelaksanaan kegiatan meliputi tahap induksi perakaran
+
inokulasiisolat -t aklimatisasi, tahap penyapihan, pemeiiharaan dan pernanenan dilakukan di
Pusbanghut Sumberdaya Hutan Cepu; analisa infeksi CMA pada akar tanaman
dilakukan di Laboratorium Bioteknologi Kehutanan Pusat Antar Universitas (PAU),
Institut Pertanian Bogor @B); analisa kadar klorofil dilakukan di Laboratorium Pusat Studi Pemuliaan Tanaman (PSPT),
PB;
sedangkan analisa tanah clan analisa jaringan tanaman dilakukan di Laboratorium Tanah, IPB dengan menggunakansesudah bibit dipindah ke persemaian. Pengamatan dilakukan terhadap tinggi,
diameter, berat kering total, rasio pucuk akar, peningkatan pertumbuhan tinggi,
kandungan klorofil, mycorrhizal dependency dan infeksi aka.
Hasil penelitian menunjukkan bahwa bibit jati dapat bersimbiosis dengan
jenis CMA, hal ini ditunjukkan dengan adanya infeksi pada akar bibit klon jati yang diinokulasikan. Penambahan isolat Glornus agregahm pada klon jati nomor 3 dan
nomor 21, akan memberikan pertumbuhan yang lebih baik dibandingkan isolat
KAJIAN KOMPATIBILITAS ISOLAT
CENDAWAN MZKORIZA ARBUSKULA (CMA) TERHADAP
PERTUMBUHAN
DUA KLON JATI (Tectona grandis L.f.)HASIL PERBANYAKAN KULTUR JARUVGAN
OLEH :
S U R A Y A
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister
Sains pada
Program
Studi Ilmu
Pengetahuan KehutananPROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judui Tesb : Kajian Kompatibiiitas Isolat Cendawan Mikoriza Arbuskula (CMA) terhadap Pertumbuhan Dua KIon Jati (Tectmra gran#is L.f.) Hasil Perbanyakan Kultur Jaringan
Nama Mahasiswa : S U R A Y A Nomor Pokok : 98230
Program Studi : Ilmu Pengetahuan Kehutanan
Menyetuj ui,
1. Komisi Pembimbing
Dr. Ir. Yadi Setiodi, MSc
Ketua
Mengetahui,
Ir, Corry
TWN,
MSiAnggota
Ketua Program Studi
Xlmu Pengetahuan Kehutanrrn
31!
~?&vafrida Manuwoto. MScSURAT PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudul :
KAJIAN KQMPATIBILITAS ISOLAT CENDAWAN MIKORTZA
ARBUSKULA (CMA) TEaaADAP PERTUMBUHAN DUA KLON
JATI (Tectona grandis L.f.) HASIL PERBANYAKAN KULTUR
JArnGAN
Adalah benar merupakan hasil karya saya sendiri dan belum pernah dipublikasikan.
Semua sumber data dan informasi yang digunakan telah dinyatakan secara jelas dan
dapat diperiksa kebenarannya.
Bogor,
September
2002RIWAYAT HIDUP
Penulis dilahirkan di kota Pontianak pada tanggal 23 Oktober 1973 sebagai
anak sulung dari dua bersaudara dari pasangan H. Hamzah Ahmad dan Dra.
Kamisah.
Pendidikan Sarjana ditempuh di Jurusan Kehutanan, Fakultas Pertanian
Universitas Tanjungpura Pontianak (UNTAN) dan lulus pada tahun 1997. Pada tahun
1998 penulis mendapat kesempatan untuk melanjutkan pendidikan ke program
Magister pada program studi Ilmu Pengetahuan Kehutanan, Program Pasca Sarjana
PRAKATA
Syukur Alhamdulillah penulis panjatkan kehadirat Allah Subhanahuwata'ala,
akhirnya penulis dapat menyelesaikan penulisan tesis ini yang merupakan salah satu
syarat dalam penyelesaian program Strata dua (S2) di Program Pascasarjana IPB.
Pada kesempatan ini, ucapan terima kasih yang tulus dan penghargaan yang
tinggi penulis sampaikan kepada bapak Dr. Ir. Yadi Setiadi, MSc. selaku ketua
komisi pembimbing dan Ibu Ir. Corry TWN, MSi, selaku anggota komisi
pembimbing atas segala bantuan, bimbingan, arahan dan dorongan sehingga penulis
dapat menyelesaikan penelitian dan penulisan tesis ini. Banyak ha1 yang penulis
peroleh melebihi dari sekedar bimbingan akademik.
Rasa terima kasih juga disampaikan kepada Pimpinan Institut Pertanian
Bogor, Direktur Program Pascasarjana IPB dan Ketua Program Studi
IPK
yang telahmemberikan kesempatan kepada penulis untuk mengikuti pendidikan Pascasarjana di
IPB .
Ungkapan terima kasih secara khusus penulis sampaikan kepada Bapak Dr.
Ir. Irdika Mansur, M.For.Sc, selaku dosen penguji luar komisi atas bimbingan,
arahan dan saran-sarannya baik secara langsung maupun tidak langsung terutarna
dalam perbaikan penulisan tesis.
Ucapan terima kasih juga disampaikan kepada semua Staf Laboran di
Laboratorium PSPT dan Laboratorium Bioteknologi PAU IPB terutama Mas
penelitian ini. Kepada rekan-rekan Mahasiswa Program Studi IPK Program
Pascasarjana IPB, yang telah memberikan bantuan, dorongan semangat dan
kerjasama yang baik selama pendidikan d m penelitian. Demikian pula tidak dapat
dilupakan jasa saudari Luki, Teti dan Susi yang telah banyak memberikan bantuan
tenaga dan dorongan semangat .
Rasa kagum dan hormat, serta penghargaan yang setinggi-tingginya ditujukan
kepada kedua orangtua H. Hamzah Ahmad dan Dra. Kamisah yang dengan ketulusan
hati telah mencurahkan kasih sayang, membimbing, dan mendo'akan penulis hingga
dapat menyelesaikan tugas akhir ini .
Akhirulkalam, penulis mohon maaf atas kekhilafan selama penelitian dan
penulisan tesis ini. Dengan diiringi do's yang tulus, penulis berharap kiianya seluruh
proses kegiatan studi ini mudah-mudahan dapat bernilai ibadah dihadapan Allah
Subhanahuwata'ala. Semoga hasil penelitian ini dapat bermanfaat demi untuk
kemajuan umat manusia dan ilmu pengetahuan, khususnya bagi pengembangan jati
klon hasil perbanyakan kultur jaringan.
Bogor, September 2002
DAFTAR IS1
Halaman
DAFTAR TABEL ...
DAFTAR GAMBAR . . .
DAFTAR LAMPIRAN ...
PENDAHULUAN ...
Latar Belakang ...
Tujuan Penelitian . . . Hipotesis ...
[image:100.611.82.513.150.712.2]TIN JAUAN PUSTAKA . . . . ...
Gambaran Umum Jati (Tectona grandzs L f )
. . .
Nama Daerah
...
Penyebaran
Habitus . . .
Kelas Kayu dan Penggunaan ...
Cendawan Mikoriza Arbuskula (CMA) . . . ...
Struktur Umum Cendawan Mikoriza Arbuskula
Peranan CMA dalam Meningkatkan Penyerapan Unsur
...
Hara dan Pertumbuhan Tanaman
. . .
Kompatibilitas CMA dengan Tanaman Inang
METODOLOGI PENELITIAN ...
Tempat dan Waktu Penelitian ...
Bahan dan Alat ...
Metode Penelitian ...
Pengamatan dan Pengumpulan Data ...
Analisis Data ... ... HAS& DAN PEMBAHASAN
...
Hasil
Tinggi Bibit . . .
Diameter Batang ...
Berat Kering Total (BKT) ...
Rasio Pucuk Akar (RPA) ...
Kandungan Klorofil ...
Peningkatan Pertumbuhan Tinggi @GI) ...
Mycorrhizal Dependency ...
Infeksi Akar ...
Pembahasan 46
Hubungan Serapan Hara dan Pertumbuhan Tanaman ... 46 Kompatibilitas antara Isolat Mikoriza dengan Klon Jati
(Tectona grandls L . f ) ... 54
KESIMPULAN 5 6
...
Kesimpulan 56
. . .
Saran 56
DAFTAR PUSTAKA ... 57
12. Rata-rata kandungan hara C, N, P, K, Ca, Mg, Cu dan Zn jaringan
DAFTAR
GAMBAR
Halaman
Histogram Pengaruh Interaksi Klon Jati dan Jenis Isolat CMA terhadap Pertambahan rata-rata Tinggi bibit jati pada Umur 12
[image:104.614.85.517.109.722.2]Minggu setelah dipindah ke Persemaian.. ... 24
Grafik Pertumbuhan rata-rata Tinggi Bibit Jati (Tectona grandis
L. f) sampai Umur 12 minggu setelah dipindeh ke Persemaian.. ... 26
Histogram Pengaruh Jenis Klon terhadap Diameter Batang
Tanaman pada Umur 12 Minggu setelah dipindah ke Persemaian.. . 28
Histogram Pengaruh Jenis Isolat CMA terhadap Diameter Batang .
Tanaman pada Umur 12 Minggu setelah dipindah ke Persemaian.. 28
Grafik Pertumbuhan rata-rata Diameter Batang Bibit Jati (Tectona
grandis
L . 9
sampai Umur 12 minggu setelah dipindah ke...
Persemaian.. 3 0
Histogram Pengaruh Interaksi Klon Jati dan Jenis Isolat CMA terhadap Berat Kering Total Bibit Jati pada Umur 12 Minggu
setelah dipindah ke Persemaian.. ... 32
Histogram Pengaruh Interaksi Klon Jati dan Jenis Isolat CMA terhadap Rasio Pucuk Akar Bibit Jati pada Umur 12 Minggu
setelah dipindah ke Persemaian. ... 34
Histogram Pengamh Interaksi Klon Jati dan Jenis Isolat CMA terhadap Kandungan Klorofil Daun Bibit Jati pada Umur 12
...
Minggu setelah dipindah ke Persemaian.. 36
Histogram Penganth Jenis Isolat CMA terhadap Peningkatan Pertumbuhan Tinggi @GI) Bibit Jati pada Umur 12 Minggu
...
setelah dipindah ke Persemaian.. 38
Histogram Pengaruh Interaksi Klon Jati dan Jenis Isolat CMA terhadap Nilai Mycorrhizal Dependency bibit jati pada Umur 12
Minggu setelah dipindah ke Persemaian.. ... 40
12 . Histogram Pengaruh Jenis Isolat CMA terhadap Infeksi Akar .
(Awal dan Akhir) Bibit Jati (Tectom g r d s L f) ... 43
13 . Penampakan visual bibit jati yang bermikoriza (KIM2 dan K2M2) ... dengan yang tidak bermikoriza (KIM0 dan K2MO) 47
PENDAHULUAN
Latar Belakang
Pengelolaan hutan tanaman di Pulau Jawa khususnya hutan tanaman jati sampai
sekarang masih menjadi perhatian yang sangat besar terutama oleh pihak Perum
Perhutani karena luasnya yang lebih dari 1 juta ha, juga mengingat tanaman ini sangat
potensial dan merupakan salah satu komoditas andalan karena termasuk jenis kayu
mewah dan bernilai ekonomi tinggi. Setiap tahunnya tidak kurang dari 500.000 m3
kayu pertukangan jati telah dihasilkan (Cordes, 1997). Luas areal hutan yang tidak
mungkin bertambah luas menyebabkan Perum Perhutani berusaha untuk meningkatkan
produktivitas, bahkan bila mungkin akan meningkatkannya demi mempertahankan
pasokan terhadap pennintaan kayu jati yang tentunya akan meningkat dengan makin
bertambahnya jumlah penduduk
Untuk mangantisipasi ha1 tersebut saat ini Pusbang SDH Perum Perhutani telah
mengembangkan jati berkualitas tinggi dengan memanfaatkan teknologi kultur jaringan
yang diharapkan dapat menjamin kualitas genetik bibit yang dihasilkan. Teknik ini
merupakan suatu cara perbanyakan secara cepat klon-klon terpilih dengan
menggunakan jaringan tanaman yang ditumbuhkan secara mikro untuk menghasilkan
tanaman yang sempurna dalam jumlah yang lebih besar. Pada akhirnya diharapkan
tanaman hasil kultur jaringan ini akan tumbuh sama dengan induk atau tetuanya.
Akan tetapi tanaman kultur jaringan memiliki beberapa permasalahan
lingkungan aseptik, unsur hara menjadi prioritas utama sehingga tanaman tidak akan
mengalami kekurangan hara tetapi pada saat tanaman sudah dipindah ke lapangan
dirnana ketersediaan hara di dalam tanah yang kadang tidak mencukupi kebutuhan
tanaman dapat saja menyebabkan tanaman tidak tumbuh dengan baik.
Untuk itu peranan mikoriza sangatlah penting karena kngsi mikoriza yang dapat
membantu proses penyerapan unsur hara di dalam tanah dari yang awalnya tidak
tersedia menjadi dapat tersedia bagi tanaman sehingga pemakaian mikoriza khususnya
pada tanaman kultur jaringan masih sangat perlu dilakukan. Hal inilah yang mendasari
penelitian ini karena pemakaian mikoriza pada awal masa pertumbuhan setelah tahap
inisiasi (perbanyakan) sangatlah penting karena sekali suatu tanaman terinfeksi oleh
cendawan mikoriza maka manfaat akan diperoleh selama hidupnya.
Untuk proses induksi akar, saat ini Pusbang SDH Perum Perhutani tidak lagi
menggunakan teknik aseptik tetapi sudah dilakukan pada kondisi non aseptik dimana
eksplant ditanam langsung pada media aklimatisasi sehingga proses induksi akar dan
aklimatisasi dapat dilakukan dalam satu waktu. Adanya penambahan isolat mikoriza
pada tahap induksi akar dan aklimatisasi ini adalah agar nantinya tanaman diharapkan
sudah dapat berasosiasi (terinfeksi) dengan mikoriza sebelum di pindah ke persemaian.
Walaupun jati (Tectona grandis L.f ) merupakan salah satu jenis tanaman hutan
yang telah diketahui dapat berasosiasi dengan CMA (Coster, 1921 ddam Manan,
1994), akan tetapi tingkat kompatibilitas tanaman tersebut terhadap jenis isolat CMA
dapat berbeda antara spesies tanaman atau bahkan varietas (kultivar) dalam spesies
(Azcon dan Ocampo, 198 1) sehingga perbedaan nomor ldon juga dapat memberikan
sangat tergantung pada kesesuaian antara faktor-faktor jenis CMA, tanarnan dan tanah
serta interaksi antara ketiga faktor tersebut. Akibat ketidaksesuaian antara isolat CMA
dengan tanaman inang diantaranya tidak akan dapat membantu pertumbuhan tanaman
dan bahkan sebdiknya dapat menirnbulkan gejala-gejda pada daun tanaman yang
terlihat hijau kekufiing-kuningan (Santoso, 1994). Untuk itu perlu dilakukan pengujian
kompatibilitas dari beberapa jeflis isolat CMA terhadap jenis Mon jati has3
perbanyakan h l t u r jaringan.
Penelitian ini bertujuan untuk :
1. Mengetahui interaksi terbaik antara Mon jati (Tectona grandis L.f) nomor 3 dan nomor 21 hasii perbanyakan kultur jaringan dengan beberapa isolat CMA yang
dicobakan.
2. Mengetahui kompatibilitas antara jenis isolat dengan Mon jati tertentu.
Hipotesis
Hipotesis dalarn penelitian ini adatah :
1. Terdapat interaksi yang terbaik antara isolat CMA dengan jenis Mon jati hasil
perbanyakan kultur jaringan.
TINJAUAN
PUSTAKA
Gambaran Umum Jati (Tectona grmdis L.f.)
Nama Daerah
Jati (Tectona grandis L.f.) termasuk famili Verbenaceae, di beberapa daerah di
Indonesia dikenal dengan narna yang berbeda-beda, diantaranya deleg, dodokan, jate,
jatih, jatos, kiati dan kulidawa. Sedangkan di negara lain dikenal dengm gzati
(Venezuala), teak ( B i m , India, Thailand, USA., Jeman), teck (Perancis) dan teca
(Brazilia) (Martawijaya et al., 198 1).
Penyebaran
Penyebaran jati di Indonesia terutama di Pulau Jawa, selain itu terdapat juga di
Pulau Muna, Maluku (Wetar) dan Nusa Tenggara. Sedangkan di luar Indonesia
terdapat di India, Burma, Thailand d m Vietnam.
Pohon jati jarang tumbuh lurus memanjang, tetapi tumbuh melebar, tinggi dapat
mencapai 45 m dengan tinggi batang bebas cabang 15
-
20 m, sedangkan diameterumumnya 50 cm dengan bentuk batang tidak teratur dan beralur (Martawijaya et al.,
1981). Pada musim kemarau, pohon jati akan menggumrkan daumya. Hal ini
dipengaruhi oleh cepat atau lambat datmgnya musim kering, semakin tinggi kadar
kelembaban di atrnosfir maka semakin lama pula pohon jati ini dapat mernpertahankan
selain karena cuaca pengguguran daun ini juga dipengaruhi oleh keadaan setempat dan
umur dari pohon itu sendiri (Cordes, 1992).
Jati merupakan jenis tanaman yang membutuhkan cahaya dalam masa
pertumbuhannya, permudaan jati tidak tahan terhadap naungan dan sangat sensitif
terhadap tekanan dari gulma yang dapat menyebabkan kematian (Cordes, 1992).
Perakaran tanaman jati pada waktu muda merupakan akar tunggal yang tumbuh cepat
dengan akar-akar permukaan yang banyak. Akar tunggal tersebut segera bercabang
banyak sehingga merupakan percabangan lanjut dari akar yang mendalam, tetapi tidak
lebih dalam dari 1,5 m pada tanah berat dan 3 m pada tanah yang ringan (Martawijaya
et al., 1981).
Jati tumbuh baik di daerah dengan musim kering yang nyata, tipe curah hujan C
sampai F, jumlah hujan rata-rata 1200 - 2000 mrnlth dan ketinggian tempat sampai
700 m dpl. Jati dapat tumbuh pada berbagai macam formasi geologi dan tidak terikat
pada satu jenis tanah tertentu, tetapi memerlukan tanah yang berdrainase dan beraerasi
cukup baik Pada tanah-tanah yang dangkal, padat serta becek pertumbuhannya kurang
baik dan mudah diserang oleh hama dan penyakit. Jati menyukai tanah sarang yang
mengandung kapur (Martawijaya ef al., 198 1).
Kelas Kayu dan Penggunaan
Jati mempunyai berat jenis kering udara 0,59 - 0,82 dengan rata-rata 0,7, kelas
awet I
-
I1 dan kelas kuat 11. Kayu jati baik digunakan untuk venir dan kayu lapislapis biasanya untuk venir muka karena mempunyai lingkaran tumbuh yang tampak
jelas baik pada bidang transversal maupun pada bidang radial, sehingga sering
menimbulkan gambar yang indah. Selain itu jati juga baik digunakan untuk meubelair,
bahan konstruksi, bahan pembuat kapal, tong dan pipa. Namun untuk bahan yang
memerlukan kekenydan tinggi jati tidak baik digunakan karena sifatnya yang agak
rapuh (Martawijaya ef al., 1981).
Cendawan Mikoriza Arbuskula (CMA)
Struktur Umum Cendawan Mikoriza Arbuskula
Mikoriza berasal dari bahasa Yunani, mykes (cendawan) dan rhiza (akar).
Mikoriza adalah struktur yang terbentuk karena asosiasi antara fungi dengan akar
tumbuhan tingkat tinggi (Tjitrosono, 1983 dalm Santoso, 1994), atau menurut
Supriyanto (1994) adalah suatu struktur yang menggambarkan hubungan simbiotik
mutualisme antara akar tanaman dengan cendawan.
Berdasarkan bentuk dan cara infeksi cendawan terhadap tanaman inang,
mikoriza dikelompokkan menjadi 2 tipe, yaitu endomikoriza dan ektomikoriza (EM).
Menurut karakteristiknya endomikoriza dibedakan menjadi 6 tipe, yaitu mikoriza
vesikula arbuskula, ektendomikoriza, mikoriza arbutoid, mikoriza monotropoid,
mikoriza ericoid dan mikoriza anggrek (orchid mycorrhizae)(Smith dan Read, 1997).
Mikoriza arbuskula terbentuk hampir pada semua tumbuhan berklorofil, yaitu
Bryophita, Pteridophita, Gymnospermae dan Angospermae. Coster (1 92 1) dalam
Manan (1994), mengatakan bahwa endomikoriza terdapat pada jati (Tectona grandis
L.f.). Selain itu, Coster juga melihat adanya struktur tersebut pada jenis lain,
diantaranya Podmarpus spp, Araucaria excedsa, Agathis alba, Cupressus festigiata,
Parmponia parv~jlora, Altingia excelsa, dan lain-lain.
Adapun karakteristik yang mencirikan bahwa tanaman terinfeksi dengan
endomikoriza yaitu : a). perakaran yang terinfeksi tidak membesar, b). cendawan tidak
membentuk struktur lapisan hifa pada permukaan akar, c). hifa terinfeksi sel korteks
secara intra dan interseluler, dl. adanya struktur khusus sistem percabangan hifa yang
disebut arbuskula dan pada sub ordo tertentu juga membentuk struktur oval yang
disebut vesikula (Harley dan Smith, 1983).
Secara taxonomi CMA termasuk ke dalam kelas Zygomycetes dan ordo
Glomales. Sampai sekarang telah diketahui lebih dari 126 species cendawan rnikoriza
arbuskula. Berdasarkan struktur arbushla dan atau vesikula yang dibentuknya ordo
Glomales digolongkan ke dalam 2 sub ordo, yaitu Gigqorineae dan Glomineae. Sub
ordo Gigasporineae terdiri atas satu famili, yaitu Gigasporaceae yang mempunyai 2
genus : Gigaspora dan Scuteldospora. Kedua genus ini tidak membentuk vesikula
dalam asosiasinya dengan akar tanaman. Sub ordo Glomineae terdiri atas 4 famili yaitu
Glomaceae, Acaulosporaceae, Archaeoyoraceae dan Paraglomaceae. Famili
Gdomaceae terdiri atas 2 genus yaitu Glomus dan Sclerocystes, famili
dari famili Archaeosporaceae adalah Archaeospora dan genus dari famili
Paraglomaceae adalah Paraglomus. Ke-4 genus ini dapat membentuk struktur
arbuskula dan vesikula pada akar tanaman (Morton, 2002).
Mikoriza arbuskula merupakan struktur yang terdiri atas hifa eksternal, hifa
internal, hifa gelung, arbuskula dan atau vesikula. Ada 2 tipe hifa eksternal pada
cendawan mikoriza arbuskula, yaitu absorber & h e (hifa-hifa yang bercabang halus)
dan runner hyphae (diameter berukurm iebih besar). Absorber hyphae merupakan
kunci dalam absorpsi nutrient, sedangkan runner hyphae merupakan jalur utarna
translokasi nutrient di dalam rniselium akar (Smith dan Read, 1997).
Hifa cendawan mikoriza arbuskula (CMA) merupakan hifa tidak bersekat yang
tumbuh diantara sel-sel korteks akar dan bercabang-cabang didalamnya. Hifa dapat
menembus ke dalam sel-sel korteks tetapi jarang sekali dapat menembus sel-sel
endodermis ke silinder pusat (stele). Pada sel-sel yang terinfeksi
akan
dibentuk gelung-gelung hifa atau cabang-cabang hifa kompleks yang dinamakan arbuskula. Arbuskula
ini diduga berperan sebagai pemindah unsur h a yang diserap oleh cendawan untuk
tanaman (Harley dan Smith, 1983). Selain arbuskula, sering juga dijumpai struktur
menggelembung yang dibentuk secara interkaler atau apikal pada hifa-hifa utama,
struktur ini dinamakan vesikula. Vesikula mengandung banyak lemak dm fbngsi
Peranan CMA dalam Meningkatkan Penyerapan Ussur Hara dan Pertumbuhan
Tanaman
Penyerapan unsur hara oleh tanarnan dalam bentuk ion dilakukan melalui ujung-
ujung akar. Bagian akar yang paling ujung dinamakan kaliptra yang menutupi jaringan
meristem. Melalui kaliptra dan daerah meristem inilah terjadi absorpsi air dan garam-
gararn mineral, tetapi dalam jumlah yang kecil. Penyerapan terbesar dilakukan oleh
bulu-bulu akar (Dwijoseputro, 1990).
Infeksi rnikoriza pada akar tanaman dapat meningkatkan kemampuan tanaman
untuk menyerap unsur hara yang terikat dan tidak tersedia bagi tanaman serta
meningkatkan kemampuan tanaman untuk menyerap air, sehingga tanaman masih
dapat hidup dengan baik pada kondisi tanah kering (Jeffiies, 1987). Mekanisme
penyerapan unsur hara pada tanaman yang terinfeksi CMA adalah bertambah luasnya
permukaan absorpsi d m meningkatkan volume daerah penyerapan oleh adanya hifa
diluar permukaan akar (hifa eksternal), serta kemarnpuan hifa yang lebih tinggi dalam
mengabsorpsi zat makanan dibanding bulu-bulu akar (Abbott et al., 1992). Hal inilah
yang menyebabkan tanaman bermikoriza mampu menyerap unsur hara lebih banyak
dan lebih baik dibandingkan dengan tanaman tidak bermikoriza.
Secara umum, besarnya pengaruh peningkatan pertumbuhan oleh infeksi akar
melalui mikoriza terutama disebabkan oleh peningkatan P khususnya dari sumber P
tersedia. Hifa eksternal CMA dapat memperoleh P larut relatif lebih besar
dibandingkan dengan akar karena daya jelajah hifa dapat mencapai 10 cm lebih dari
kecil(2 - 4 pm) menyebabkan volume tanah yang dapat diterobosi menjadi lebih besar
(Bolan, 1991). Adanya pmingkatan penyerapan P oleh tanaman yang terinfeksi
mikoriza arbuskula diduga karena adanya peningkatan aktivitas enzim fosfatase pada
rhizosfer dan akar tanaman tersebut. Unsur P bagi tanaman berguna untuk : a)
memacu pertumbuhan akar dan pembentukan sistem perakaran yang baik dari benih
dan tanaman muda, b). mempercepat pembungaan dan pemasakan buah atau biji dan
c). sebagai penyusun inti sel, lemak dan protein (Setyamidjaya, 1986). Lebih lanjut
Lingga (1991), menambahkan bahwa fungsi phospor bagi tanaman adalah sebagai
bahan mentah untuk pembentukan sejumlah protein clan juga mernbentuk asimilasi dan
pernapasan dalam bentuk energi ATP dan ADP.
Simbiosis antara tanaman dan CMA adalah menguntungkan dan penting untuk
kelangsungan hidup cendawan karena cendawan mengambil fotosintesis dari tanaman
(Sieverding, 199 1). Proses fotosintesis sangat terkait dengan jumlah klorofil yang
terkandung dalam tanaman. Meningkatnya kadar Morofil daun kemungkinan
disebabkan oleh meningkatnya serapan unsur Mg dan N pada tanaman bermikoriza.
Kedua unsur tersebut berperan penting dalam pembentukan klorofil daun. Infeksi
jamur mikoriza mempunyai kontribusi cukup penting terhadap serapan Mg oleh
tanaman meskipun ha1 ini belum benyak diteliti (Marschner dan Dell, 1994). Hal yang
sama terjadi pada unsur N bahwa pada tanaman yang bermikoriza serapan unsur N
lebih tinggi dibandingkan dengan tanaman yang tidak bermikoriza Amet et al., dalarn
Marschner dan Dell (1 994) telah mengukur secara langsung terhadap pengambilan d m
bahwa setelah 30 hari, 2,5% dan 3,5% dari N total masing-masing pada tanaman
berrnikoriza dan tidak bennikoriza berasal dari
'%
yang dihasilkan oleh hifa jamurmikoriza. Tingginya kadar N pada tanaman berrnikoriza adalah akibat serapan N oleh
hifa mikoriza.
Kompatibilitas CMA dengan Tanaman Inang
Kompatibilitas CMA dengan tanaman inang ini tidak terlepas dari efektivitas dan
infektivitasnya. Efektivitas CMA sangat tergantung pada kesesuaian antara faktor-
faktor jenis CMA, tanaman dan tanah, serta interaksi ketiga faktor tersebut. Jenis
tanaman berpengaruh dalam hal perbedaan tingkat ketergantungan pada mikoriza
karena ada tanaman tertentu yang sangat membutuhkan keberadaan mikoriza, seperti
ubi kayu (Sieverding, 1991), sedangkan tanaman lainnya malah tidak
membutuhkannya, seperti lobak (Dodd et al., 1987). Oleh karena itu, tidak mudah
untuk menentukan efektivitasnya dalam meningkatkan pertumbuhan dan serapan unsur
hara oleh tanaman tertentu. Perbedaan kompatibilitas ini ditunjukkan dari hasil
penelitian Rohayati (1999) yang menggunakan beberapa isolat CMA pada 2 jenis klon
jati (hasil perbanyakan kultur jaringan) pada saat penyapihan yaitu klon 07 dan klon
03 Hasilnya menunjukkan bahwa pembhan isolat CMA Glomus aggregatum pada
bibit jati nomor kion 07 asal KPH Cepu dapat menghasilkan persentase infeksi
tertinggi (89,29%) dan penarnbahan isolat CMA Glomus manzhobis pada klon yang
sama menunjukkan persentase infeksi terendah.. Sedangkan pada bibit jati nomor Mon
isolat CMA Glomus agregatum (9137%) dan persentase infeksi terendah adalah isolat
CMA Mix : G i g a p r a sp.
+
Glomus sp. yaitu 4737%.Kemarnpuan satu cendawan dapat berbeda dalam membentuk mikoriza dengan
CMA lainnya. Masing-masing species CMA mempunyai kemampuan menghasilkan
hifa yang berbeda yang mana hal ini dipengaruhi oleh pengikatan kualitas inokulum
rnaupun kemampuan infeksi. Mekanisme dapat terjadi karena masing-masing species
CMA mernpunyai kemampuan yang berbeda sehingga satu species dapat berkembang
sedangkan species lainnya tidak. Karakteristik fungi yang menentukan keefektifannya
adalah kemampuan untuk menginfeksi akar secara cepat agar mikoriza sudah
terbentuk ketika umur tanaman masih relatif muda (Hayman, 1983) dan kemampuan
untuk membentuk rniselium eksternal secara ekstensif (Mosse, 1986). Abbott et a/.
(1992) mendapatkan korelasi yang erat antara keefektivan spesies fungi dan persentase
panjang akar yang terinfeksi pada masa awal pertumbuhan tanaman.
Dengan melihat keadaan di atas maka kesesuaian dan efektivitas fingi mikoriza
dengan jenis tanaman adalah sangat penting untuk membantu penyerapan unsur hara
dari dalam tanah sehingga tanaman dapat tumbuh dengan baik sejak awal dan untuk
METODOLOGI PENELITIAN
Tempat Dan Waktu Penelitian
Penelitian ini dilaksanakan di Pusat Pengembangan Sumberdaya Hutan Cepu,
Analisis Infeksi Cendawan Mikoriza Arbuskula (CMA) pada akar tanaman
dilaksanakan di laboratorium Bioteknologi Kehutanan Pusat Antar Universitas IPB,
Analisa kadar klorofil dilaksanakan di Laboratorium PSPT IPB, Analisa tmah dan
jaringan tanarnan dilaksanakan di Laboratorium Tanah IPB. Waktu Penelitian selarna
*
8 bulan yaitu mulai dari bulan Pebruari sampai dengan September 200 1.
Bahan Dan Alat
Bahan-bahan yang digunakan dalam penelitian ini adalah klon eksplant hasil
perbanyakan kultur jaringan jati berasal dari Pusat Pengembangan Sumberdaya Hutan
Cepu, isolat cendawan mikoriza arbuskula yang terdiri dari jenis Glornus agregatum
dari Pusbanghut Cepu, Glomus ebunicaturn, Acaulospora tuberculata dan Mycofer
dari PAU Biotek IPB, media perakaran berupa pasir dan tanah yang telah disterilkan,
zat pengatur tumbuh (IBA), media top soil, bahan kimia (untuk analisa struktur akar
dan analisa klorofil).
Alat-alat yang digunakan adalah bak plastik ukuran 33 x 26 x 11 cm, polibag
ukuran 250 gr, neraca analitik, erlenmeyer, labu ukur, botol semprot, pengaduk,
gunting, pinset, cawan petri, objek glass dan cover glass, rnistar ukur, kaliper, oven,
Metode Penelitian
Kegiatan yang dilakukan dalam penelitian ini terbagi atas tahap induksi
perakaran
+
inokulasi spora i- aklimatisasi, tahap penyapihan dan pemeliharaafl.Induksi Perakaran
+
Inokulasi Spora+
AklimatisasiEksplant dikeluarkan dengan hati-hati dari botol kultur lalu dipotong
+
3 cmdengan menyertakan 3
-
4 helai daun. Selanjutnya dicelupkan ke dalam 3 rnl larutanIBA selama
+
15 - 30 menit.Eksplant yang telah dicelupkan dalam IBA tadi selanjutnya ditanam pada bak
plastik ukuran 33 x 26 x 11 cm yang telah diisi media pasir dan tanah yang telah
disterilkan dengan perbandingan 3 : 1 dengan tinggi media sekitar 3 cm. Tiap bak
berisi 25 eksplant. Bermmaan dengan itu dilakukan inokuiasi cendawan mikoriza
arbuskula (CMA) disekitar tanaman dengan dosis inokulum sebanyak
+
120 flak.Pemberian isolat dilakukan dengan membuat jalur-jalur yang jaraknya sekitar 1 cm dari
eksplant lalu isolat ditanam di dalam jalur tadi. Selanjutnya wadah tanam ditutup
dengan kaca sampai kondisi tanaman siap dipindah ke polibag, kira-kira sekitar 30 -
40 hari.
Penyapihan
Media yang digunakan untuk penyapihan adalah lapisan tanah top soil.
Penyapihan dilakukan dengan memindahkan bibit yang sudah berumur 1 bulan secara
inokulasinya untuk menghindari patah atau putusnya akar saat dipindahkan. Dalam
tahap penyapihan ini, diambil 10 bibit dalarn kondisi yang homogen.
Pem berian Nutrisi
Pemberian Nutrisi dilakukan dengan menyemprotkan pupuk daun (Suburi)
dengan dosis 1 ml pada seluruh permukaan tanaman setelah penyapihan. Dilakukan 2
rninggu sekali dengan dosis yang sama.
Pemeliharaan Tanaman
Pemeliharaan tanaman meliputi kegiatan penyiraman menggunakan botol sem-
prot yang dilakukan pada pagi dan sore hari secara teratur sesuai kebutuhan sarnpai
kapasitas lapang, pencabutan rumput dan pemeliharaan semai dari gangguan hama.
Rancangan Percobaan
Rancangan percobaan yang digunakan dalam penelitian ini adalah Percobaan
Faktorial2 x 5 dalam Pola Acak Lengkap (RAL), yaitu Faktor Jenis Klon (K) terdiri
dari 2 taraf dan Faktor Jenis Isolat CMA (M) terdiri dari 5 taraf, sehingga terdapat 10
perlakuan dengan 5 ulangan. Model percobaan yang dilakukan adalah model tetap
(taraf faktor K dan M tetap) karena hanya tertuju pada taraf-taraf faktor yang
Perincian dari masing-masing faktor perlakuan adalah sebagai berikut :
1. Faktor perlakuan jenis klon terdiri dari dua taraf, yaitu :
K1
= Klon nomor 03K2 = Klon nomor 2 1
2. Faktor perlakuan jenis rnikoriza (isolat CMA) terdiri dari lima taraf, yaitu :
Mo
= tanpa mikoriza (kontrol)MI = Glomus etunicatum
M2 = Glomus agregaturn
M3 = Acaulospora fuberculuta
N
= MycoferPengamatan dan Pengumpulan Data
Pengamatan dan pengukuran peubah yang dilakukan meliputi
Pengukuran Tinggi dan Diameter Bibit
Dalam pengukuran tinggi dan diameter bibit, pengukuran pertama dilakukan
pada saat bibit dipindahkan ke dalam polibag dan pengukuran selanjutnya dilakukan
seminggu sekali.
Pengukuran Bobot Kering Total
Pengukuran bobot kering total dilakukan pada akbir penelitian, dimana tiap
tanaman dibagi menjadi bagian akar dan pucuk, lalu kedua bagian tersebut dioven
dalam bungkus terpisah selama 24 jam pada suhu 70-80'~. Setelah pengovenan lalu
dilakukan penimbangan. Berat kering total diperoleh dengan menambahkan berat akar
Rasio Pucuk Akar
Rasio pucuk akar ditentukan dengan membandingkan berat kering pucuk dengan
berat kering akar tanaman.
Peningkatan Pertumbuhan Tinggi (High Growth IncrementlHGI)
Menurut Supriyanto (1994), peningkatan perturnbuhan tinggi (HGI) ini
dinyatakan dengan persen untuk membandingkan selisih dari tinggi bibit bermikoriza
dan tinggi bibit tanpa mikoriza dengan tinggi bibit tanpa mikoriza dikalikan 100 %.
HGI ( O h ) = H t - H c x 100%
U
Dimana :
Ht = tinggi bibit bermikoriza
Hc = tinggi bibit tanpa mikoriza (kontrol)
Mycorrhizal Dependency
Pengukuran Mycorrhizal Dependency ini dinyatakan dalam persen, yaitu dengan
membandingkan selisih berat kering bibit diinokulasi (perlakuan) dengan berat kering
bibit tanpa dinokulasi mikoriza dikalikan 100 %. Nilai ini penting untuk mengetahui
tingkat ketergantungan tanaman terhadap mikoriza.
Dw x 100%
Mycorrhizd Dependency(%) =
-
DWC
Dimana : DWt = berat kering bibit yang dinokulasi mikoriza
Analisa Klorofd
Kandungan Morofil diamati pada akhir penelitian. Analisa dilakukan
Laboratorium PSPT
IPB.
Caranya dengan menghancurkan daun jati menggunakanmesin pencacah dalam larutan aseton 85%, selanjutnya larutan disaring untuk
mernisahkan supernatan. Supernatan diencerkan kemudian dilakukan pengukuran
klorofil daun dengan menggunakan spektrofotometer. Nilai konsentrasi dalam mgAt
kemudian diubah menjadi mg/gr berat basah dengan menggunakan rumus berikut :
konsentrasi (mg/#) x volume aseton yang digunakan (It) Konsentrasi (mglgr bemt basah) =
gr contoh
Analisa Media
Analisa media dimaksudkan untuk mengetahui kandungan hara yang terdapat
pada media. Diamati pada awaI dan akhir penelitian. Analisa media meliputi pH, persen
C d m N, P-tersedia, Cu, Zn, K, Ca, Mg dan nilai KTK. Analisa dilakukan di
Laboratorium Tanah IPB.
Analisis Infeksi Akar
Analisis infeksi CMA dilakukan untuk melihat adanya simbiosis antara CMA
dengan akar tanaman yang dinyatakan dalam perstntase infeksi. Pengukuran
persentase infeksi akar dilakukan pada awal (saat penyapihan) dan akhir penelitian
Akar direndam dalam Latutan KOH dibuang setelah akar larutan KOH 10% dan betwarna kuning bersih dan dibilas dibiarkan selama 24 jam dengan air biasa sebanyak 4 kali
Larutan HCI dibuang dan akar diberi
larutan staining (glicerol, HCI 1% dan Akar direndam dalam
aquadest
-
75% : 5% : 20%) + trypan larutan tlCl2% sampai blue 0.05 mg), dibiarkan selama 24 jam jaringan akar memutih (tampak bening)Y
Larutan staining diganti dengan larutan destaining (glicerol, HCI 1 %
dan aquadest
-
50% : 5% : 45%) tanpa trypan blue, dibiarkanselama 24 jam
lalu dibuat preparat (1 0 potonganlpreparat)
Perhitungan persentase akar yang terinfeksi mikoriza dilakukan di bawah mikroskop
L
Preparat diamati dengan menggunakan rnikroskop fase kontras dan dihitung persen
infeksi akarnya dengan menggunakan rumus :
% Akar yang terinfeksi = C bidang pandangpositif x 100%
Analisa Jaringan Tanaman
Analisa jaringan tanaman dimaksudkan untuk mengetahui kandungan unsur hara
yang dibutuhkan tanaman. Diarnati pada akhir penelitian. Analisa jaringan meliputi
unsur N, P, K, Ca, Mg, Cu d m Zn. Analisa jaringan dilakukan di Laboratorium Tanah
IPB .
Analisis Data
Model linier yang digunakan dalam RAL Faktorial adalah :
Dimana :
Yijk = Nilai pertumbuhan klon jati pada petak percobaan ke-k yang memperoleh
kombinasi perlakuan ij (taraf ke-i dari faktor jenis klon dan taraf ke-j dari
faktor jenis isolat)
P = Rata-rata pertumbuhan klon jati yang sesungguhnya
Ki
= Pengaruh aditif dari jenis klon jati ke-iMj = Pengaruh aditif dari jenis isolat CMA ke-j
(KM)ij = Pengaruh interaksi antara klon jati ke-i, dan jenis isolat CMA ke-j
wjk = Pengaruh galat percobaan pada petak percobaan ke-k yang memperoleh
Pengolahan data dilakukan secara statistik terhadap data-data peubah
pertumbuhan. Untuk mengetahui pengaruh perlakuan dan untuk memudahkan
pengolahan data, maka semua data perhihrngan statistik yang didapat dari hasil
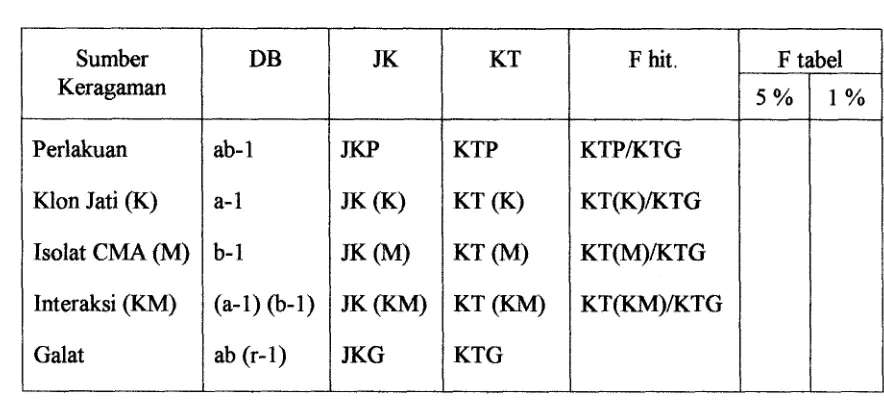
pengamatan dibuat dalam tabel analisis keragaman RAZ, Faktorial berikut :
Tabel 1. Analisis Ragam Percobaan Faktorial yang terdiri dari h a Faktor dengan
RAL
Untuk mengetahui pengaruh perlakuan yang diberikan, maka digunakan uji-F. Adapun
hipotesis yang akan diuji untuk model tetap adalah :
1. &, : (ap)ij = 0, yang berarti tidak ada pengaruh interaksi antara jenis klon jati dan jenis isolat CMA terhadap peubah yang diamati.
HI : Ada pngaruh interaksi antara jenis klon jati d m jenis isolat CMA terhadap
peubah yang diamati.
2 . &, : ai = 0, yang berarti jenis Mon jati yang dicobakan tidak mempengaruhi peubah
yang diamati.
HI : jenis Mon jati yang dicobakan mempengaruhi peubah yang diamati. Sumber
Keragaman
Perlakuan
Klon Jati (K)
Isolat CMA (M)
Interaksi (KM)
Galat
DB
ab- 1
a-1
b- 1
(a- 1
-
1 ab (r-1)JK
JKP
JK (K)
JK(A4)
JK (KM)
JKG
KT
KTP
KT (K)
KT(M)
KT (KM)
KTG
F hit.
KTPKTG
KT(K)/KTG
KT(M)KTG
KT(KM)KTG
F tabel
[image:126.611.91.533.243.443.2]3.
&
:pj
= 0, yang berarti jenis isolat CMA yang diberikan tidak mempengaruhipeubah yang diamati.
HI : jenis isolat CMA yang diberikan mempengaruhi peubah yang diamati.
Kriteria Pengambilan keputusan terhadap uji - F adalah :
*
F hitung < F tabel, rnaka terima &% F hitung 2 F tabel, maka terima HI
Menurut Gaspersz (1994), hipotesis tentang pengaruh interaksi ditempatkan
pada bagian pertama, hal ini menunjukkan bahwa dalam percobaan faktorial perlu
terlebih dahulu menguji hipotesis interaksi karena jika pengaruh interaksi berbeda
nyata (& ditolak), maka tidak perlu lagi melakukan pengujian hipotesis pengaruh
utama (hipotesis 2 dan 3) tetapi yang penting adalah mengusut lebih jauh bagaimana
bentuk hubungan ketergantungan diantara faktor K dan M. Tapi jika pengujian
terhadap hipotesis mengenai pengaruh interaksi (KM) tidak berbeda nyata (6
diterima), maka pengujian terhadap hipotesis pengaruh utama faktor K dan pengaruh
utama faktor M dilakukan. Apabila ddarn sidik ragam terdapat pengaruh perlakuan,
analisis dilanjutkan dengan Uji Wilayah Berganda Duncan untuk menguji pengaruh
antar dua perlakuan. Pengolahan data dilakukan dengan menggunakan Program
HASIL DAN PEMBAHASAN
Hasil
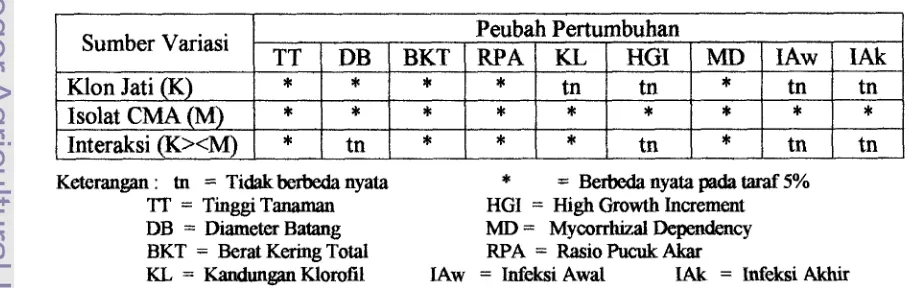
Hasil analisis sidik ragam terhadap berbagai peubah pertumbuhan menunjukkan
bahwa interaksi (K><M) berpengaruh nyata terhadap tinggi tanaman, berat kering
total, rasio pucuk akar, dan kandungan klorofil daun dan mycorrhizal dependency;
serta tidak berpengaruh nyata terhadap diameter batang, peningkatan pertumbuhan
tinggi (HGI) dan infeksi akar baik pada awal maupun akhir. Sedangkan klon jati (K)
berpengaruh nyata terhadap tinggi tanaman, diameter batang, berat kering total, rasio
pucuk akar dan mycorrhizal dependency; serta tidak berpengaruh nyata terhadap
kandungan klorofil, HGI serta infeksi baik pada awal (saat tanaman siap dipindah ke
persemaian) maupun akhir (setelah bibit berumur 12 minggu di persemaian). Untuk
Isolat CMA (M) semua variabel pengarnatan menunjukkan adanya pengaruh yang
nyata pada taraf 5 %. Adapun daftar rekapitulasi analisa sidik ragam pengaruh isolat
CMA terhadap klon jati pada berbagai peubah pertumbuhan secara rinci dapat dilihat
pada Tabel 2.
Tabel 2. Rekapitulasi Hasil Sidik Ragam Pengaruh Isolat CMA terhadap Klon Jati pada Berbagai Peubah Pertumbuhan.
Keterangan : tn = Tidak berbeda nyata
*
= Berbeda nyata pada taraf 5%'IT = Tinggi Tanaman HGI = High Growth Increment DB = Diameter Batang MD = Mycorrhizal Dependency BKT = Berat Kering Total RPA = Rasio Pucuk Akar
KL = Kandungan Klorofil 1Aw = Infeksi Awal IAk = Infeksi iAkhir Sumber Variasi
Klon Jati (K)
Isolat CMA (M)
[image:128.612.80.534.562.706.2]Tinggi Bibit
Hasil sidik ragam menunjukkan bahwa interaksi antara kedua faktor tersebut
(K><M) memberikan pengaruh yang nyata pada taraf 5 %, sehingga tidak perlu lagi
melakukan pengujian dari pengaruh utarna faktor jenis klon (K) dan jenis isolat CMA
(M). Bentuk interaksi antara faktor jenis klon (K) dan faktor isolat CMA (M)
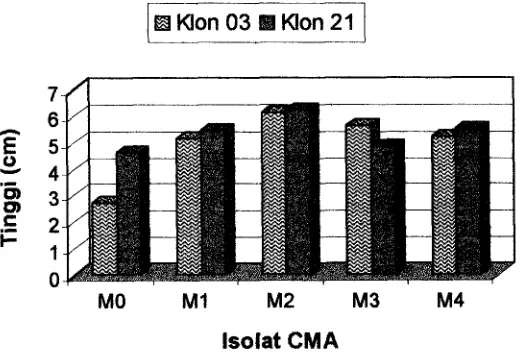
terhadap respon tinggi tanaman ditunjukkan pada Gambar 1.
Klon 03 El Klon 21
lsolat CMA
Gambar 1. Histogram Pengaruh Interaksi Klon Jati dan Jenis Isolat CMA terhadap Pertambahan rata-rata Tinggi bibit jati pada Umur 12 Minggu setelah
dipindah ke Persemaian.
Keterangan : MO : Tanpa mikoriza (Kontrol) M3 : Acaulospora tuberculata
M1 : Glomus etunicatum M4 : Mycofer
M2 : Glomus agregatum
Pada Gambar 1 menunjukkan bahwa klon nomor 21 dengan isolat CMA
Glomus agregatrum (M2) menunjukkan hasil yang lebih tinggi dibandingkan dengan
jenis isolat lainnya. Selanjutnya karena pengaruh sederhana dari masing-masing
faktor tidak sama maka dilakukan uji lanjut dengan menggunakan uji pembandingan
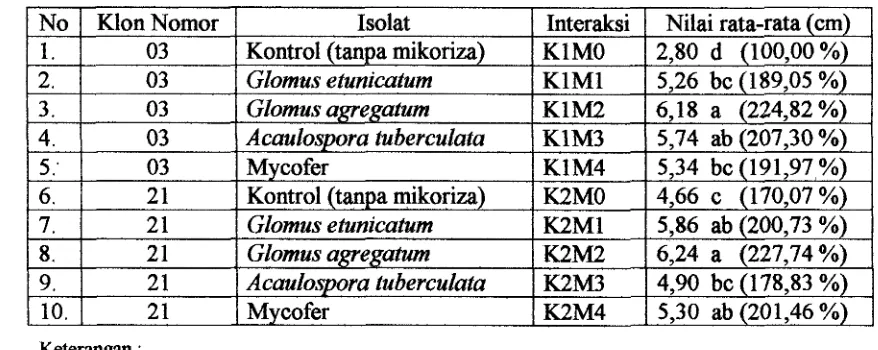
[image:129.609.188.450.267.444.2]Tabel 3. Uji Beda Nyata Pengaruh Interaksi Klon Jati dan Jenis Isolat CMA terhadap Tinggi Tanaman pada Umur 12 Minggu setelah dipindah ke Persemaian.
Keterangan :
-
Nilai yang diikuti huruf yang sama tidak berbeda nyata pada taraf nyata 5 %-
Angka di dalam kurung menunjuk;kan nilai relatif (pmentase) terhadry, kontrol5: 6. 7. 8. 9. 10.
Tabel 3 menunjukkan bahwa pada klon 03, interaksi klon dengan penambahan
isolat G. agregatum (KlM2) memberikan hasil yang tidak berbeda dengan
penambahan isolat Acaulospora tuberculata, sedangkan penambahan isolat A.
tuberculata tidak berbeda dengan isolat
G.
etunicatum dan Mycofer. Akan tetapipenambahan isolat G. agregatum berbeda nyata dengan isolat G. etunicatum dan
Mycofer. Adapun pada klon 21, interaksi klon dengan penambahan isolat CMA G.
agreratum (K2M2) tidak berbeda dengan isolat G. etunicatum dan Mycofer 03
2 1 2 1 2 1 2 1 2 1
sedangkan penambahan isolat G. agregatum berbeda nyata dengan isolat A.
tuberculata. Sedangkan untuk interaksi antara Mon 21 dengan isolat A. tuberculata
tidak berbeda dengan tanpa mikoriza (kontrol).
Tabel 3 juga menunjukkan bahwa dari hasil pengujian pengaruh sederhana Mycofer
Kontrol (tanpa mikoriza) Glomus etunicatum Glomus agregatum A caulospora tubercu lata Mycofer
faktor Mon pada jenis isolat dapat disimpulkan, apabila kita melihat pada jenis isolat
maka dari kedua klon yang digunakan penambahan isolat G. agregatum tidak berbeda KIM4 K2MO K2Ml K2M2 K2M3 K2M4
5,34 bc (191,97,%) 4,66 c (170,07 %)
5,86 ab (200,73 %)
6,24 a (227,74 %)
4,90 bc (178,83 %)
dengan penambahan isolat G. etunicaferrn, A. tuberculuta dan Mycofer, tetapi berbeda
dengan isolat kontrol (tanpa mikoriza).
Hasil pengukuran dari nilai rata-rata pertambahan tinggi bibit yang diukur
setiap minggu untuk setiap taraf perlalcuan jenis isolat CMA ditunjukkan dalam grafik
pada Gambar 2.
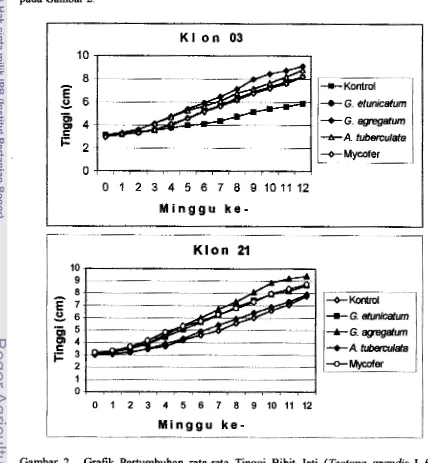
I
-+
MycoferM i n g g u k e -
K l o n
21/ [image:131.603.79.507.206.669.2]M i n g g u k e -
I
Pengaruh inokulasi CMA terhadap pertambahan tinggi kedua klon jati setelah
dipindah ke persemaian menunjukkan bahwa peningkatan tinggi sudah terlihat rata-
rata pada minggu ke-3 setelah tanaman dipindah ke persemaian. Awalnya pemberian
masing-masing isolat C;. agregatzsm dan A. tzcbercuZata pada klon 03 menunjukkan adanya peningkatan pertambahan tinggi yang sama dibandingkan dengan isolat
lainnya, tapi perubahan mulai tampak pada minggu ke-6 dimana tanaman yang
diinokulasi isolat G. agregadlwn m l a i mengungguli tanaman yang diinokulasi isolat
lainnya. Sementara masing-masing isolat G. eiunicufarm dan Mycofer menunjukkan
peningkatan pertambahan tinggi yang sama sarnpai pada akhir pengamatan.
Berikutnya pada klon 21, perbedaan peningkatan pertambahan tinggi mulai tampak
pada minggu ke-5 yaitu Mon dengan pemberian jenis isolat G. ugregafarm. Sedanglcan
untuk jenis isolat mycofer dan G. eturzicuhrm masing-masing menunjukkan
peningkatan pertambahan tinggi yang harnpir sama. Peningkatan pertambahan tinggi
terendah sebelum kontrol adalah pada penggunaan isolat A. tuberalafa.
Diameter Brmtamg
Hasil sidik ragam menunjukkan bahwa interaksi antara kedua faktor tersebut
(K><M) tidak memberikan pengaruh yang nyata pada taraf 5 %, sedangkan pengaruh
utama faktor klon jati
(K)
dan pengaruh utama faktor jenis isolat CMA (M)memberikan pengaruh yang nyata pada t a d 5 % terhadap pertambahan diameter
bibit jati (Tecfona grandis L.Q. EMini menjelaskan bahwa terdapat perbedaan respon
pertarnbahan diameter batang dari perlakuan yang diberikan. Untuk melihat p e w h
Klon 21 Won 03 Klon Jati
Gambar 3 Histogram Pengaruh Jenis Klon terhadap Diameter Batang Tanaman pada Umur 12 Minggu setelah dipindah ke Persemaian.
Pada histogram terlihat bahwa bibit jati nomor klon 21 memperlihatkan
pertarnbahan diameter yang lebih baik dibanding klon nomor 03. Selanjutnya untuk
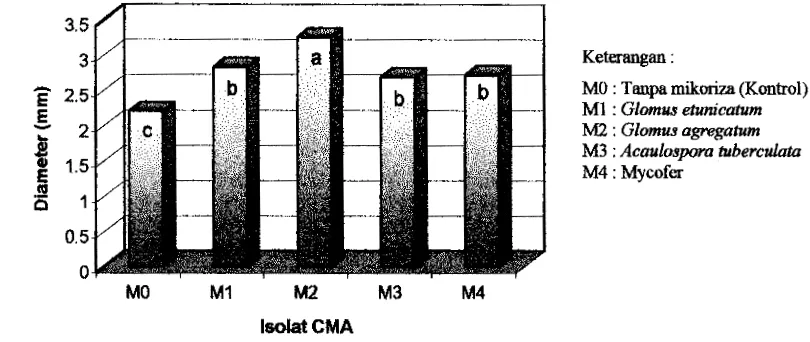
mengetahui pengaruh utama faktw jenis isolat CMA secara lebih jelas ditunjukkan
dalam bentuk histogram pada Gambar 4.
MO M I M2 M3 M4
lsdat CMA
Keterangan :
MO : Tanpa m i k d (Kontrol)
M1 : Glomus ehcnicatum
M2 : Glomus agregatum M3 : Acaulospora tuberculata M4 : Mycofer
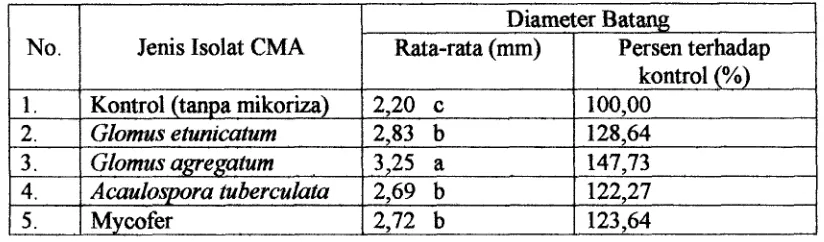
Tabel 4 . Uji Beda Nyata Pengaruh Jenis Isolat CMA terhadap Diameter Batang Bibit Jati pada Umur 12 Minggu setelah dipindah ke Persemaian.
*) Nilai yang W t i dengan huruf sama tidak berbeda nyata pada tingkat kepercayaan 95%
Hasil uji Duncan (Tabel 4 ) terhadap pengaruh utama faktor isolat CMA
menunjukkan bahwa pertambahan diameter terbaik ditunjukkan oleh bibit jati yang
diinokulasi dengan isolat CMA G. agregatum dengan rata-rata diameter sebesar 3,25
mm (147,73 %) diikuti dengan isolat G . etunzcatum. dengan rata-rata tinggi sebesar
2,83 mm (128,64 %). Isolat G . agregatzim menunjukkan perbedaan yang nyata
dengan ke-tiga isolat lainnya dan kontrol, sedangkan antara isolat G. etunicatum, A.
tuberculata dan Mycofer tidak menunjukkan adanya perbedaan yang nyata.
Hasil pengukuran dari nilai rata-rata pertambahan diameter batang bibit yang
diukur setiap minggu untuk setiap taraf perlakuan jenis isolat CMA ditunjukkan
dalam grafik pada Gambar 5.
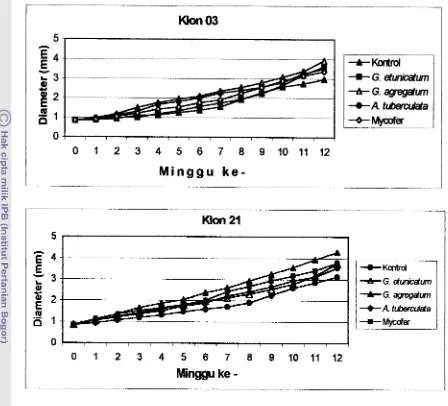
Pengaruh inokulasi CMA terhadap pertambahan diameter kedua klon jati
setelah dipindah ke persemaian menunjukkan bahwa peningkatan diameter sudah
terlihat rata-rata pada minggu ke-3 setelah tanaman dipindah ke persemaian. No. 1 . 2. 3. 4 . 5 .
Jenis Isolat CMA
Kontrol (tanpa mikoriza)
Glomus etunicatum Glomus agregatune , ,
AcauIospora tuberculata
Mycofer
Diameter Batang Rata-rata (mm)
2,20 c
2,83 b
3,25 a
2,69 b
2,72 b
Persen terhadap kontrol (%) 100,OO
[image:134.609.102.514.133.254.2]0
0 1 2 3 4 5 6 7 8 9 1 0 1 1 1 2
M i n g g u k e -
[image:135.603.75.523.81.487.2]4 5 6 7 8 9 1 0 1 1 1 2 Minggu ke
-
Gambar 5. Grafik Pertumbuhan rata-rata Diameter Batang Bibit Jati (Tectona gradis
L.0
sampai Umur 12 minggu setelah dipindah ke PersemaianAwalnya pemberian masing-masing isolat G. agregatum dan A . tuberculata pada
klon 03 menunjukkan adanya peningkatan diameter yang hampir sama dibandingkan
dengan isolat lainnya, tapi perubahan mulai tampak pada minggu ke-7 dimana
tanaman yang diinokulasi isolat G. agregatum mulai mengungguli tanaman yang
diinokulasi isolat lainnya. Sementara masing-masing isolat Mycofer menunjukkan
akhir pengamatan. Berikutnya pada klon 2 1, perbedaan peningkatan pertambahan
tinggi mulai tampak pada minggu ke-2 yaitu klon dengan pemberian jenis isolat G.
agregatum. Sedangkan untuk jenis isolat mycofer, G, etzrnicatum dan A, tuberculata
masing-masing menunjukkan peningkatan pertambahan diameter yang hampir sama
mulai dari minggu pertama sampai dengan minggu ke-6. Untuk selanjutnya isolat
Mycofer mengungguli kedua isolat lainnya.
Berat Kering Total (BKT)
Berdasarkan hasil sidik ragam menunjukkan bahwa interaksi antara kedua
faktor (K><M) memberikan penganrh yang nyata pada taraf 5 % terhadap berat
kering total bibit jati (T. grandis L.f) sehingga tidak perlu lagi melakukan pengujian
dari pengaruh utama faktor jenis klon (K) dan jenis isolat CMA (M). Adapun bentuk
interaksi antara faktor jenis klon
(K)
dan faktor isolat CMA (M) terhadap responberat kering total bibit tanaman ditunjukkan pada Gambar 6.
Pada Gambar 6 terlihat bahwa klon nomor 21 dengan isolat CMA G.
agregatum (M2) menunjukn hasil yang lebih tinggi dibandingkan dengan jenis
isolat lainnya. Selanjutnya karma pengaruh sederhana dari masing-masing faktor
tidak sama maka dilakukan uji lanjut dengan menggunakan uji pembandingan nilai
lsolat
C M AGambar 6. Histogram Pengaruh Interaksi Klon Jati dan Jenis Isolat CMA terhadap Berat Kering Total bibit jati pada Umur 12 Minggu setelah dipindah ke Persemaian.
Keterangan : MO : Tanpa mikoriza (Kontrol) M3 : Acaulospora tuberculata
M1 : Glomus etunicatum M4 : Mywfer
M2 : GIomus agregatum
Tabel 5 menunjukkan bahwa pada klon 03, penambahan isolat G. agregaturn
menghasilkan perbedaan yang nyata terhadap nilai berat kering total tanaman dengan
ke-tiga isolat yang dicobakan (G. etunicaturn, A. tuberculata dm Mycofer) d m kontrol (tanpa rnikoriza). Sedmgkan penarnbahan isolat G. etunicatum tidak berbeda
nyata dengan isolat A. iuberculata. Penambahan isolat Mycofer pada klon 03 juga
tidak menunjukkan perbedaan yang nyata dengan ke-tiga isolat yang lainnya. Adapun
pada klon 2 1, penambahan isolat Ch4A G. agreraturn (K2M2) berbeda nyata dengan
isolat G. etunicatum, A. tuberculata, Mycofer dan kontrol, sedangkan penambahan
isolat G. etunicatum tidak berbeda nyata dengan isolat A. tuberculata dan Mycofer. Tabel 5 juga menunjukkan bahwa dari hasil pengujian pengaruh sederhana
maka dari kedua klon yang digunakan penambahan isolat G, agregafarm menunjukkan
perbedm yang nyata dengan ke-3 isolat yang lain (G. efunicatm, A. tubercztlafa dan Mycofer) j u g berbeda dengan kontrol (tanpa mikoriza).
Tabel 5. Uji Beda Nyata Pengaruh Interaksi Klon Jati dan Jenis Isolat CMA terhadap Berat Kering Total Tanaman pada Umur 12 Minggu setelah dipindah ke Persemaian.
Ketmngan :
-
Nilai yang *ti huruf yang sama ti& berbed;t nyata pada taraf nyata 5 %-
An& di datam kurung m e n u n . niiai relatif (presentase) terhadap iconmiRasio Pucuk A h r (RPA)
Berdasarkan hasil sidik ragam menunjukkan bahwa interaksi antara kedua
faktor tersebut (K><MJ memberikan penganlh yang nyata pada taraf 5 % terhadap
rasio pucuk akar bibit jati (T. grandis
L.0.
Nilai rasio pucuk akar ini berbandingterbalik dengan nilai berat kering total tanaman.
Hasil uji Duncan terhadap pengaruh interaksi antara kedua faktor (KXM)
secara lebih jelas ditunjukkan dalam hntuk histogram pada Gambar 7.
No 1. 2. 3. 4. 5. 6. 7. 8. 9. 10. KlonNomor 03 03 03 03 03 21 21 21 2 1 2 1
Isolat
Kontrol (tanpa mikoriza)
Glornus ehmicahcm Glomus agregatum Acaulospora tubercuhta
My cofer
Kontrol (tanpa mikoriza)
G l o m s etunicatum GZomars agregatum
-
-
-
-Acmlospara ttlbermlata Mycofer Interaksi KIM0 K l M l KIM2 KIM3 KIM4 K2MO K2M1 K2M2 K2M3 K2M4Nilai rata-rata (gr) 0,85d (100,00%) 1,89 b (222,35 %)
2,37 a (278,82 %) 2,04 b (240,OO %)
[image:138.614.115.538.246.416.2]I
Klon 03 Klon 21/
Gambar 7. Histogram Pengaruh Interaksi Klon Jati d m Jenis Isolat CMA terhadap Rasio Pucuk Akar bibit jati pada Umur 12 Minggu setelah dipindah ke Persemaian.
Gambar 7 menunjukkan bahwa perlakuan tanpa mikoriza pada klon nomor 03
menunjukkan nilai yang paling besar dibandingkan dengan perlakuan lainnya. Selanjutnya karena pengaruh sederhana dari masing-masing faktor tidak sama maka
dilakukan uji lanjut dengan menggunakan uji pembandingan nilai tengah perlakuan
(Tabel 6).
Hasil uji Duncan terhadap penganlh utama faktor isolat CMA menunjukkan
bahwa nilai rasio pucuk akar terbesar ditunjukkan oleh perlakuan kontrol pada klon
nomor 03 sebesar 3,276 sedangkan yang terkecil adalak bibit jati yang diinokulasi
dengan isolat CMA A. tubercarlata yaitu sebesar 1,582. Tampak pada tabel bahwa
bibit jati klon 21 yang dinokulasi dengan sernua isolat mikoriza tidak menunjukkan
perbedaan yang nyata, tetapi untuk Mon 03 jika dibandingkan dengan kontrol maka
Tabel 6. Uji Beda Nyata Pengaruh interaksi Klon Jati dan Jenis Isolat CMA terhadap Rasio Pucuk Akar Tanaman pada Umur 12 Minggu setelah dipindah ke Persemaian.
Keterangan :
-
Nilai yang d i W huruf yang sama tidak berbeda nyata pada taraf nyata 5 %-
Angka di dalam