EKSPLORASI DAN POTENSI JAMUR PELARUT FOSFAT
PADA EKOSISTEM LAHAN GAMBUT
DESA TELAGA SUKA KABUPATEN LABUHAN BATU
SKRIPSI
Oleh :
YULIA SANTIKA TELAUMBANUA 071202032
Budidaya Hutan
PROGRAM STUDI KEHUTANAN
FAKULTAS PERTANIAN
EKSPLORASI DAN POTENSI JAMUR PELARUT FOSFAT
PADA EKOSISTEM LAHAN GAMBUT
DESA TELAGA SUKA KABUPATEN LABUHAN BATU
SKRIPSI
Oleh :
YULIA SANTIKA TELAUMBANUA 071202032
Budidaya Hutan
Skripsi sebagai salah satu syarat untuk memperoleh gelar sarjana di Fakultas Pertanian
Universitas Sumatera Utara
PROGRAM STUDI KEHUTANAN
FAKULTAS PERTANIAN
LEMBAR PENGESAHAN
Judul Penelitian : Eksplorasi dan Potensi Jamur Pelarut Fosfat Pada Ekosistem Lahan Gambut Desa Telaga Suka Labuhan Batu
Nama Mahasiswa : Yulia Santika Telaumbanua
NIM : 071202032
Departemen : Kehutanan
Program Studi : Budidaya Hutan
Disetujui oleh : Komisi Pembimbing
Dr.Deni Elfiati, SP, MP Dr. Delvian, SP,MP
Ketua Anggota
Mengetahui
ABSTRAK
YULIA SANTIKA TELAUMBANUA: Eksplorasi dan Potensi Jamur Pelarut Fosfat pada Ekosistem Lahan Gambut Desa Telaga Suka Kabupaten Labuhan Batu. Dibawah bimbingan DENI ELFIATI dan DELVIAN.
Penelitian ini dilakukan untuk mengeksplorasi, menguji potensi serta mengindentifikasi jamur pelarut fosfat (JPF) unggul dari lahan gambut Desa Telaga Suka, Kabupaten Labuhan Batu. Sampel diambil dari 5 petak secara acak pada kedalaman 0-20 cm disekitar rhizosfir.Analisis sifat kimia tanah gambut dilakukan di Balai Penelitian Perbenihan Sumatera Utara sedangkan kegiatan isolasi, uji potensi serta identifikasi dilaksanakan di Laboratorium Biologi Tanah Program Studi Agroekoteknologi Fakultas Pertanian Universitas Sumatera Utara. Isolasi menggunakan media Pikovskaya dengan sumber fosfatCa3(PO4)2, sedangkan pada media uji potensi sumber fosfat dari AlPO4. Evaluasi potensi JPF secara kualitatif dengan mengukur luas diameter holozone menggunakan nilai indeks pelarutan. Pengukuran potensi JPF secara kuantitatif dengan mengukur kadar fosfat terlarut dengan metode Bray-II.
Hasil isolasi diperoleh 18 isolat JPF murni. Selanjutnya dilakukan uji potensi pada isolat yang diperoleh. Diameter holozone terbesar yaitu 1,955 cm dihasilkan isolat 17 dan diameter terkecil sebesar 0,887 cm dihasilkan isolat 14. Hasil pengukuran kadar fosfat tersedia paling besar adalah isolat 16 yaitu 19,668 ppm dan paling kecil oleh isolat 10 yaitu 8,348 ppm. Berdasarkan hasil uji potensi disimpulkan isolat 16 dan 17 memiliki kemampuan paling baik dalam melarutkan fosfat. Hasil indentifikasi baik secara makroskopis dan mikroskopis menunjukkan bahwa isolat 16 termasuk genus Penicillium dan isolat 17 termasuk genus
Aspergillus.
ABSTRACT
YULIA SANTIKA TELAUMBANUA. Exploration and Potential Mushroom Phosphates Solvents on Peatland Ecosystems Suka Lake District village of Labuhan Batu, under the guidance of DENI ELFIATI and DELVIAN.
The research was conducted to explore, examine and identify potential phosphate solubilizing fungi (PSF) to solubilize insoluble phosphate from peat of Telaga Suka Village, Labuhan Batu regency. Samples taken from five random plots at 0-20 cm depth around rhizosfir. The chemical analyze properties of peat soils conducted in North Sumatra Seed Research Center while the activities of isolation, and identification of potential trials conducted at the Laboratory of Soil Biology Agroekoteknologi Studies Program Faculty of Agriculture, University of North Sumatra. Isolation using Pikovskaya media with the source fosfat from Ca3(PO4)2, whereas in the potential test media using AlPO4 as source of insoluble
phosphate. Evaluate the potential JPF qualitatively by measuring the diameter of holozone extensive use of the dilution index. JPF potential measurements quantitatively by measuring levels of dissolved phosphate with Bray-II method.
The results obtained 18 isolates of isolation of pure PSF. The next test conducted on the potential of isolates obtained. Largest diameter of 1,955 cm holozone generated isolates 17 and the smallest diameter of 0,887 cm isolates produced 14. The measurement results are available most of the phosphate levels were 16 isolates of 19,668 ppm and the smallest by 10 isolates of 8,348 ppm. Based on test results concluded the potential for isolates 16 and 17 have the best ability in dissolving phosphate. The results both of macroscopic and microscopically identification showed that isolate number 16 including Penicillium genus and isolate number 17 including of Aspergillus genus.
RIWAYAT HIDUP
Yulia Santika Telaumbanua, dilahirkan di Medan pada tanggal 30 Juli
1989 dari Ayah Samaeli Telaumbanua dan Ibu Tineke Puspawati Linanda. Penulis merupakan putri kedua dari tiga bersaudara.
Tahun 2007 penulis lulus dari SMA Negri 4 Medan dan pada tahun yang
sama lulus seleksi masuk Universitas Sumatera Utara melalui jalur SPMB. Penulis memilih program studi Budidaya Hutan, Departemen Kehutanan, Fakultas
Pertanian.
Penulis mengikuti Praktek Pengenalan dan Pengelolaan Hutan (P3H) di hutan dataran rendah Aras Napal Kabupaten Langkat dan hutan mangrove Pulau
Sembilan, Pangkalan Susu. Selama mengikuti perkuliahan, penulis pernah menjadi asisten pratikum mata kuliah Pemanenan Hasil Hutan pada tahun 2010
dan asisten lapangan kegiatan Pengenalan Ekosistem Hutan di Lau Kawar Kabupaten Karo pada tahun 2010. Penulis melakukan Praktek Kerja Lapangan di PT. Andalas Merapi Timber, Sumatera Barat pada bulan Januari-Februari 2011.
Penulis melaksanakan penelitian yang berjudul Eksplorasi dan Potensi Jamur Pelarut Fosfat Pada Ekosistem Lahan Gambut Desa Telaga Suka Labuhan
KATA PENGANTAR
Puji syukur penulis panjatkan ke hadirat Tuhan Yang Maha Esa, atas segala berkat dan rahmatNya sehingga penulis dapat menyelesaikan skripsi yang
berjudul “Eksplorasi dan Potensi Jamur Pelarut Fosfat Pada Ekosistem Lahan Gambut Desa Telaga Suka Labuhan Batu”.
Dengan segala kerendahan hati, penulis mengucapkan terima kasih kepada banyak pihak yang turut membantu penulis menyelesaikan skripsi ini.
1. Ayah Samaeli Telaumbanua dan Ibu Tineke Puspawati Linanda serta keluarga besar yang telah membesarkan, mendidik serta selalu mendukung
penulis lewat doanya yang tulus,
2. Komisi pembimbing saya, Dr. Deni Elfiati, SP, MP selaku ketua dan Dr. Delvian, SP. MP selaku anggota yang membimbing, memberi masukan
yang sangat bermanfaat selama penelitian hingga penulisan skripsi ini selesai, 3. Teman-teman Budidaya Hutan stambuk 2007 dan seluruh pegawai di
Program Studi Kehutanan yang memberi dukungan hingga skripsi ini selesai. Penulis menyadari bahwa dalam penulisan skripsi ini tidak luput dari kekurangan, oleh sebab itu penulis mengharapkan saran dan kritik yang
membangun demi penyempurnaan skripsi ini. Penulis mengharapkan skripsi ini dapat bermanfaat bagi pengembangan ilmu pengetahuan khususnya di bidang
kehutanan. Akhir kata penulis mengucapkan terima kasih.
Medan, Juli 2011
DAFTAR ISI
Indentifikasi jamur pelarut fosfat yang potensial melarutkan fosfat ... 15
HASIL DAN PEMBAHASAN Karakteristik Sifat Kimia Sampel Tanah Gambut ………… ... 16
Isolasi Jamur Pelarut Fosfat dari Bahan Tanah Gambut ……… 19
Kemampuan JPF Melarutkan P pada Media Pikovskaya Padat. ………... 22
Kemampuan JPF Melarutkan P pada Media Pikovskaya Cair ………….. 26
Indentifikasi Jamur Pelarut Fosfat dari Bahan Tanah Gambut …….……. 30
DAFTAR TABEL
No Hal
DAFTAR GAMBAR
No Hal
1 Ilustrasi petak contoh pengambilan ... 12
2 Morfologi Aspergillus sp ... 31
3 Morfologi Penicillium sp ……… 31
4 Aspergillus sp ... 31
DAFTAR LAMPIRAN
No Hal
1 Dokumentasi tahap penelitian ………... 39 2 Kriteria penilaian sifat kimia tanahStaf Pusat Penelitian Tanah-
ABSTRAK
YULIA SANTIKA TELAUMBANUA: Eksplorasi dan Potensi Jamur Pelarut Fosfat pada Ekosistem Lahan Gambut Desa Telaga Suka Kabupaten Labuhan Batu. Dibawah bimbingan DENI ELFIATI dan DELVIAN.
Penelitian ini dilakukan untuk mengeksplorasi, menguji potensi serta mengindentifikasi jamur pelarut fosfat (JPF) unggul dari lahan gambut Desa Telaga Suka, Kabupaten Labuhan Batu. Sampel diambil dari 5 petak secara acak pada kedalaman 0-20 cm disekitar rhizosfir.Analisis sifat kimia tanah gambut dilakukan di Balai Penelitian Perbenihan Sumatera Utara sedangkan kegiatan isolasi, uji potensi serta identifikasi dilaksanakan di Laboratorium Biologi Tanah Program Studi Agroekoteknologi Fakultas Pertanian Universitas Sumatera Utara. Isolasi menggunakan media Pikovskaya dengan sumber fosfatCa3(PO4)2, sedangkan pada media uji potensi sumber fosfat dari AlPO4. Evaluasi potensi JPF secara kualitatif dengan mengukur luas diameter holozone menggunakan nilai indeks pelarutan. Pengukuran potensi JPF secara kuantitatif dengan mengukur kadar fosfat terlarut dengan metode Bray-II.
Hasil isolasi diperoleh 18 isolat JPF murni. Selanjutnya dilakukan uji potensi pada isolat yang diperoleh. Diameter holozone terbesar yaitu 1,955 cm dihasilkan isolat 17 dan diameter terkecil sebesar 0,887 cm dihasilkan isolat 14. Hasil pengukuran kadar fosfat tersedia paling besar adalah isolat 16 yaitu 19,668 ppm dan paling kecil oleh isolat 10 yaitu 8,348 ppm. Berdasarkan hasil uji potensi disimpulkan isolat 16 dan 17 memiliki kemampuan paling baik dalam melarutkan fosfat. Hasil indentifikasi baik secara makroskopis dan mikroskopis menunjukkan bahwa isolat 16 termasuk genus Penicillium dan isolat 17 termasuk genus
Aspergillus.
ABSTRACT
YULIA SANTIKA TELAUMBANUA. Exploration and Potential Mushroom Phosphates Solvents on Peatland Ecosystems Suka Lake District village of Labuhan Batu, under the guidance of DENI ELFIATI and DELVIAN.
The research was conducted to explore, examine and identify potential phosphate solubilizing fungi (PSF) to solubilize insoluble phosphate from peat of Telaga Suka Village, Labuhan Batu regency. Samples taken from five random plots at 0-20 cm depth around rhizosfir. The chemical analyze properties of peat soils conducted in North Sumatra Seed Research Center while the activities of isolation, and identification of potential trials conducted at the Laboratory of Soil Biology Agroekoteknologi Studies Program Faculty of Agriculture, University of North Sumatra. Isolation using Pikovskaya media with the source fosfat from Ca3(PO4)2, whereas in the potential test media using AlPO4 as source of insoluble
phosphate. Evaluate the potential JPF qualitatively by measuring the diameter of holozone extensive use of the dilution index. JPF potential measurements quantitatively by measuring levels of dissolved phosphate with Bray-II method.
The results obtained 18 isolates of isolation of pure PSF. The next test conducted on the potential of isolates obtained. Largest diameter of 1,955 cm holozone generated isolates 17 and the smallest diameter of 0,887 cm isolates produced 14. The measurement results are available most of the phosphate levels were 16 isolates of 19,668 ppm and the smallest by 10 isolates of 8,348 ppm. Based on test results concluded the potential for isolates 16 and 17 have the best ability in dissolving phosphate. The results both of macroscopic and microscopically identification showed that isolate number 16 including Penicillium genus and isolate number 17 including of Aspergillus genus.
PENDAHULUAN
Latar Belakang
Lahan gambut merupakan tanah organik yang tersebar luas di Indonesia.
Gambut merupakan areal potensial dan banyak diperhatikan, terutama dalam ekstensifikasi pertanian. Melihat dari segi budidaya tanaman, gambut dikategorikan tidak produktif karena memiliki tingkat kemasaman tanah yang tinggi dan umumnya ketersediaan unsur-unsur hara didalamnya terikat kation
organik.
Gambut terbentuk akibat tingginya penumpukan bahan organik mati yang
telah mengalami humifikasi namun proses mineralisasinya berjalan lambat. Dekomposisi berjalan lambat karena pH yang rendah serta kondisi anaerob gambut. Tingkat kemasaman gambut relatif tinggi dengan kisaran pH 3-5.
Semakin tebal gambut menyebabkan basa-basa yang dikandungnya semakin rendah sehingga reaksi tanah menjadi semakin masam. Kondisi ini mengakibatkan
aktivitas mikroba tanah terhambat (Darmawijaya, 1992).
Kandungan bahan organik gambut yang tinggi merupakan sumber hara makro dan mikro yang bermanfaat bagi organisme gambut. Kondisi tanah gambut
yang masam menyebabkan hara berada pada kondisi tidak tersedia bagi tanaman maupun mikroba tanah. Gambut juga mengandung beragam asam organik
bermanfaat serta derivate fenolat yang bersifat racun bagi tanaman.
Fosfat merupakan hara makro essensial bagi tanaman. Fosfat berperan sebagai sumber energi utama dalam proses fisiologis tanaman serta dalam reaksi
Pada lahan yang mengalami retensi fosfat (P), pemupukan dianggap
sebagai jalan keluar karena cepat menyediakan P bagi tanaman. Pemupukan P sering tidak efisien karena hara P cepat tersedia dalam jumlah besar namun tidak dibarengi penyerapan yang besar oleh akar. Semakin lama P tersedia bersentuhan
dengan tanah menyebabkan semakin banyak P yang terikat sehingga terjadi penumpukan unsur P dalam tanah. Pada lahan gambut pemupukan tidak efesien
karena gambut mengandung sejumlah besar kation organik pengikat hara. Penumpukan hara dalam tanah dapat merusak kondisi tanah sehingga tidak dapat digunakan untuk musim tanam selanjutnya.
Status hara P dapat ditingkatkan secara efektif dan efisiensi dengan memanfaatkan mikroba pelarut fosfat yang mampu membebaskan P-terikat dari
tanah dan meningkatkan serapan P oleh akar tanaman. Mikroba pelarut fosfat adalah mikroba yang mampu melarutkan ikatan fosfat menjadi bentuk tersedia. Mikroba pelarut fosfat dapat berupa bakteri (BPF), jamur (JPF), aktinomisetes
atau khamir (Premono, 1998).
Keberadaan mikroba pelarut fosfat dipengaruhi pH tanah. Pada kondisi
masam jamur pelarut fosfat (JPF) dapat tumbuh optimum dibanding bakteri dan aktinomisetes. Pertumbuhan bakteri pelarut fosfat (BPF) optimum pada pH netral dan meningkat seiring dengan meningkatnya pH tanah (Ginting, 2006).
Pada kondisi masam aluminium dan besi banyak terlarut dalam tanah. Dalam jumlah yang berlebihan, logam-logam tersebut akan membentuk ikatan
Lingkungan gambut yang sesuai untuk pertumbuhan JPF serta potensinya
dalam melarutkan ikatan fosfat pada kondisi masam, menjadi peluang untuk mengembangkan JPF pada areal gambut. Jamur pelarut fosfat dapat juga dijadikan starter pupuk hayati untuk merehabilitasi lahan gambut. Penggunaan biofertilizer
seperti JPF selain murah juga tidak menimbulkan dampak negatif terhadap tanah dan lingkungan.
Selama ini eksplorasi jamur pelarut fosfat umum dilakukan pada tanah mineral dan masih sedikit eksplorasi serupa pada tanah gambut. Atas dasar inilah penelitian ini dilakukan, untuk dapat melihat keberadaan jamur pelarut fosfat dan
menguji potensinya dalam melarutkan ikatan fosfat pada lahan gambut.
Tujuan Penelitian
Tujuan penelitian ini adalah mengeksplorasi jamur pelarut fosfat pada lahan gambut dan mengkaji kemampuannya melarutkan fosfat serta mengindentifikasi jamur pelarut fosfat paling potensial.
Kegunaan Penelitian
Penelitian ini diharapkan, dapat memberi informasi jamur pelarut fosfat
TINJAUAN PUSTAKA
Tanah Gambut
Gambut dibentuk oleh lingkungan yang khas dengan suasana tergenang
yang terjadi hampir sepanjang tahun. Keadaan hidro-topografi berupa genangan menciptakan kondisi anaerob yang memperlambat aktivitas dekomposer. Hanya sebagian kecil mikroba tanah yang mampu beradaptasi pada kondisi demikian. Hal inilah yang menyebabkan laju penimbunan bahan organik lebih besar dari
mineralisasinya (Noor, 2001).
Dalam klasifikasi tanah (soil taxonomy), tanah gambut dikenal sebagai organosol atau histosols yaitu tanah yang memiliki lapisan bahan organik dengan berat jenis (BD) dalam keadaan lembab < 0,1 g cm-3 dengan tebal > 60 cm atau lapisan organik dengan BD > 0,1 g cm-3 dengan tebal > 40 cm. Gambut dapat pula diklasifikasikan berdasarkan tingkat kematangan, kedalaman, kesuburan dan posisi pembentukannya (ICRAF, 2008).
Gambut kaya bahan organik yang menjadi sumber hara makro dan mikro bagi tanaman. Ketebalan gambut berbanding lurus dengan kapasitas tukar kation dan berbanding terbalik terhadap kejenuhan basa. Hal ini berarti semakin tebal
gambut maka kapasitas kation meningkat sehingga basa-basa yang dikandungnya semakin rendah dan menyebabkan reaksi tanah semakin masam. Kondisi ini
berpengaruh pada ketersediaan hara yang rendah (ICRAF, 2008).
Peranan dan Sumber Fosfat
Setiap tanaman sedikitnya membutuhkan 16 unsur hara agar
Salah satu hara yang berperan penting bagi pertumbuhan dan perkembangan
tumbuhan adalah fosfor karena termasuk hara makro esensial. Konsentrasi P dalam tanaman umumnya antara 0,1% sampai 0,4%. Unsur P terdapat di seluruh sel hidup tanaman yang menyusun jaringan tanaman seperti asam nukleat,
fosfolipida dan fitin (Tisdale, 1990).
Unsur P diperlukan dalam pembentukan primordial bunga serta organ
reproduksi, dalam pemasakan buah, pembentukan biji terutama pada tanaman serealia. Pada proses fisiologis tanaman, fosfat berperan sebagai sumber energi utama reaksi metabolisme dan biosintesis. Dalam proses glikolisis, pernafasan
atau fotosintesis, energi ini dilepaskan dan digunakan untuk menyusun ikatan pirofosfat yang kaya energi. Fosfat berfungsi sebagai aktifator enzim yang
mengatur proses-proses enzimatik dalam tanaman. Fosfat berperan dalam pembentukan asam nukleat (RNA dan DNA), menyimpan serta memindahkan energi ATP dan ADP, merangsang pembelahan sel dan membantu proses
asimilasi dan respirasi (Poerwowidodo, 2000).
Kekurangan fosfat dapat mengganggu sistem fisiologis tanaman seperti
pertumbuhan akar. Terhambatnya pertumbuhan akar turut mengganggu absorpsi unsur hara lain sehingga pertumbuhan tanaman menjadi terhambat. Namun kelebihan fosfat juga akan memberi dampak negatif karena membuat umur
tumbuhan terlihat lebih pendek dibanding tumbuhan dengan kadar fosfat normal (Rosmarkam dan Yuwono, 2002).
anorganik berasal dari mineral tanah yang mengandung fosfat. Pada tanah gambut
hara P umumnya berbentuk P-organik. Tanaman tidak dapat langsung memanfaatkan P-organik sehingga memerlukan proses mineralisasi untuk dapat diserap tanaman (Rosmarkam dan Yuwono, 2002).
Senyawa P-organik dalam tanah antara lain fosfolipida, asam suksinat, fitin dan inositol fosfat yang dapat didekomposisi dengan baik oleh mikroba
tanah. Unsur P-anorganik mudah bersenyawa dengan berbagai ikatan seperti Al, Fe, Ca, dan Mn. Senyawa P-anorganik dapat diklasifikasikan menjadi 4 bagian yaitu besi fosfat (FePO4), aluminium fosfat (AlPO4), kalsium fosfat (Ca3(PO4)2) dan reductant soluble. Bentuk FePO4 dan AlPO4 dominan ditemukan pada tanah masam (Rosmarkam dan Yuwono, 2002).
Derajat pengikatan P yang tinggi terjadi bila kondisi pH sangat rendah atau sangat tinggi. Kelarutan Fe dan Al tinggi pada kondisi pH rendah dan berakibat peningkatan jumlah P-terikat. Pada kondisi pH yang tinggi, ion Ca aktif terlarut
dalam tanah. Kadar P relatif tinggi pada lapisan atas dan terus menurun secara bertahap mulai lapisan 40 cm (Noor, 2001).
Mikroba Pelarut Fosfat
Siklus hara tanah dipengaruhi mikroba tanah dan tumbuhan. Beberapa mikroba mampu melarutkan fosfat yang tidak larut dalam air serta dapat
mencegahnya terfiksasi dengan mengubah bentuk P dalam tanah. Mikroba yang memiliki kemampuan demikian disebut mikroba pelarut fosfat. Mikroba pelarut
(GA3), siderofor, antibiotika, vitamin, non vitamin, substansi pemacu
pertumbuhan seperti indole acetic acid (IAA) serta sifat mikrob yang mampu mengkolonisasi akar dengan cepat dan luas (Ponmurugan dan Gopi, 2006).
Mikroba pelarut fosfat hidup di sekitar perakaran tanaman, mulai
permukaan tanah sampai kedalaman 25 cm. Keberadaannya berkaitan dengan jumlah bahan organik yang akan mempengaruhi populasi serta aktivitasnya dalam
tanah. Mikroba yang hidup dekat daerah perakaran secara fisiologis lebih aktif dibanding mikroba yang hidup jauh dari daerah perakaran. Keberadaan mikroba pelarut fosfat beragam dari satu tempat ke tempat lainnya karena perbedaan sifat
biologis mikroba itu sendiri. Terdapat mikroba yang hidup pada kondisi masam dan ada pula yang hidup pada kondisi netral dan basa, ada yang hipofilik,
mesofilik dan termofilik ada yang hidup aerob maupun anaerob (Ginting, 2006).
Pelarutan P oleh mikroba berkaitan dengan produksi asam organik hasil
metabolisme serta mineralisasi P-organik menjadi bentuk P-anorganik. Mikroba pelarut fosfat dapat melarutkan fosfat baik dari ikatan organik maupun ikatan
P-anorganik. Pelarutan dilakukan dengan mekanisme kimia dengan bantuan asam organik dan mekanisme biologi dengan bantuan enzim. Pada umumnya mekanisme pelarutan dilakukan secara kimia (Rahmawati, 2005).
Mekanisme kimia pelarutan fosfat dimulai saat mikroba pelarut fosfat mengekresikan sejumlah asam organik berbobot molekul rendah hasil
penurunan pH. Penurunan pH dapat pula disebabkan oleh pembebasan asam sulfat
dan nitrat pada oksidasi kemoautotrofik sulfur dan amonium. Perubahan pH berperan penting dalam peningkatan kelarutan fosfat. Asam-asam organik tersebut akan bereaksi dengan bahan pengikat fosfat seperti Al3+, Fe3+, Ca2+ atau Mg2+ membentuk khelat organik yang stabil yang mampu membebaskan ion fosfat terikat sehingga dapat dimanfaatkan oleh tumbuhan (Setiawati, 1998).
Asam organik dapat meningkatkan ketersediaan P di dalam tanah melalui beberapa mekanisme diantaranya adalah: (1) anion organik bersaing dengan ortofosfat pada permukaan tapak jerapan koloid tanah yang bermuatan positif,
sehingga memperbesar peluang ortofosfat dapat diserap oleh tanaman; (2) pelepasan ortofosfat dari ikatan logam P melalui pembentukan kompleks logam
organik dan (3) modifikasi muatan permukaan jerapan oleh ligan organik (Ginting, 2006).
Mikroba pelarut fosfat melakukan mekanisme biologis pelarutan fosfat
dengan memproduksi enzim fosfatase dan enzim fitase. Hasil penelitian Rao (1994), menunjukkan bahwa mikroba pelarut fosfat mampu meningkatkan
ketersediaan unsur P melalui aktivitas enzim. Enzim fosfatase dihasilkan saat ketersediaan fosfat rendah yang diekskresikan oleh akar tanaman dan mikroba. Dalam tanah enzim tersebut dominan dihasilkan oleh mikroba (Joner, 2000 dalam
Ginting, 2006). Senyawa fosfat organik diuraikan menjadi bentuk fosfat anorganik pada proses mineralisasi dengan bantuan enzim fosfatase. Enzim ini dapat
Mikroba membutuhkan fosfat dalam bentuk tersedia untuk aktivitas
metabolisme dan sintesa protoplasma. Mikroba pelarut fosfat memiliki mekanisme khusus yang mampu memanfaatkan fosfat terikat dalam tanah. Fosfat yang telah berhasil dilarutkan dimanfaatkan kembali oleh mikroba pelarut fosfat
atau mikroba lainnya. Selain mengasimilasi fosfat yang dibebaskannya, mikroba tersebut melepaskan sejumlah besar fosfat terlarut yang merupakan kelebihan dari
pasokan nutrisinya ke dalam larutan tanah. Hara fosfat yang larut akan masuk kedalam akar tanaman secara difusi. Kondisi ini akan meningkatkan fosfat tersedia yang dapat diserap akar tanaman. Jika mikroba mati maka P-organik yang
terdapat dalam jaringan mikroba akan lepas kembali dalam bentuk P-anorganik (Ginting, 2006).
Jamur Pelarut Fosfat
Jamur pelarut fosfat merupakan salah satu anggota mikroba tanah yang dapat meningkatkan ketersediaan dan pengambilan P oleh tumbuhan. Bentuk
ikatan P yang umum ditemui pada kondisi masam adalah AlPO4 dan FePO4. Jamur pelarut fosfat mampu melarutkan P dalam bentuk AlPO4 lebih baik dibanding BPF pada kondisi masam. Penelitian Lestari dan Saraswati (1997) melaporkan bahwa jamur pelarut P mampu meningkatkan kadar fosfat terlarut sebesar 27-47% di tanah masam. Penelitian Goenadi (1994), menunjukkan JPF
Jamur pelarut fosfat memiliki 3 mekanisme dalam meningkatkan
penyerapan P yaitu: (1) secara fisik dimana infeksi jamur pada akar tanaman dapat membantu pengambilan fosfor dengan memperluas permukaan sampai akar; (2) secara kimia jamur diduga mendorong perubahan pH perakaran. Jamur juga
menghasilkan asam sitrat dan asam oksalat yang menggantikan posisi ion fosfat yang terfikasasi; (3) secara fisiologi, jamur menghasilkan hormon auksin,
sitokinin dan giberelin yang mampu memperlambat proses penuaan akar sehingga memperpanjang masa penyerapan unsur hara (Premono, 1998).
Prinsip dasar isolasi mikroba pelarut fosfat ialah menyeleksi mikroba
dalam media pertumbuhan spesifik yang mengandung sumber P terikat. Kemampuan mikroba pelarut fosfat dalam melarutkan fosfat terikat dapat
diketahui dengan mengembangkan biakan murni pada media Pikovskaya yang berwarna putih keruh, karena mengandung P tidak larut air seperti kalsium fosfat Ca3(PO4)2. Pertumbuhan mikroba pelarut fosfat dicirikan dengan zona bening (holozone) di sekeliling koloni mikroba. Mikroba pelarut fosfat yang potensial dapat diseleksi dengan melihat luas zona bening paling besar pada media padat.
Pengukuran potensi pelarutan fosfat secara kualitatif ini menggunakan nilai indeks pelarutan (dissolving index), yaitu nisbah antara diameter zona jernih terhadap diameter koloni. Kemampuan pelarut fosfat terikat secara kuantitatif dapat diukur
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan pada bulan Maret sampai Juni 2011. Pengambilan
sampel tanah dilakukan di lahan gambut Desa Telaga Suka, Kabupaten Labuhan Batu. Analisis tanah dilaksanakan di Balai Penelitian Teknologi Perbenihan Sumatera Utara. Isolasi jamur, uji potensi dan identifikasi jamur pelarut fosfat dilakukan di Laboratorium Biologi Tanah Program Studi Agroekoteknologi
Fakultas Pertanian Universitas Sumatera Utara.
Bahan dan Alat
Bahan yang digunakan adalah contoh tanah gambut di bawah tegakan tanaman perkebunan karet dan sawit, kapas, akuades, kantung plastik, label, alkohol 96%, plastik kraf, aluminium foil, kaca preparat dan kaca objek. Media
padat Pikovskaya untuk komposisi per liter akuades: (glukosa 10 g; Ca3(PO4)2 5 g; (NH4)2SO4 0,5 g; KCl 0,2 g; MgSO4.7H2O 0,1 g; MnSO4 0,002 g; FeSO4 0,002 g; ekstrak khamir 0,5 g; agar 20 g; akuades), larutan fisiologis (8,5 g NaCl per liter akuades), AlPO4 5 gr.
Alat yang digunakan adalah cangkul, Erlenmeyer 250 ml, pipet tetes,
cawan petri, tabung reaksi, timbangan, inkubator, laminar air flow, gelas ukur volume 100 ml, autoklaf, rotarimixer, sentrifuse 7000 rpm, shaker, jarum ose,
sprayer, kamera digital, masker, sarung tangan, bunsen, kotak es (cool box) dan mikroskop.
Prosedur Penelitian 1. Pembuatan petak
Petak pengambilan sampel yang dibuat berdasarkan metode ICRAF (Ervayenri et al., 1999). Ukuran petak pengambilan sampel adalah 20 m × 20 m. Penetapan petak contoh dilakukan secara acak sebanyak lima petak contoh.
Gambar 1. Ilustrasi petak contoh pengambilan sampel tanah Keterangan :
: tempat pengambilan sampel tanah
2. Pengambilan contoh tanah
Pengambilan sampel dilakukan pada lima petak contoh dimana dari tiap petak diambil sampel tanah dari 6 titik pada kedalaman 0-20 cm di sekitar rhizosfer tanaman. Berat tanah yang diambil pada tiap titik adalah 500 g sehingga
total berat sampel tanah adalah 3000 g. Sampel tanah dari tiap titik dimasukkan dalam kantung plastik yang terpisah. Sampel tanah selanjutnya dibawa ke
3. Isolasi jamur pelarut fosfat
Sepuluh (10) g tanah dimasukkan ke dalam Erlenmeyer 250 ml yang berisi 90 ml larutan fisiologis steril (pengenceran 10-1), kemudian dikocok selama 30 menit pada shaker. Dibuat pengenceran secara serial, dari pengenceran 10-1 diambil 1 ml dan dimasukkan ke dalam tabung reaksi yang telah berisi 9 ml larutan fisiologis steril (pengenceran 10-2) selanjutnya dikocok di atas rotarimixer sampai homogen. Dari pengenceran 10-2 dipipet sebanyak 1 ml dan dimasukkan ke dalam tabung reaksi yang berisi 9 ml larutan fisiologis (disebut pengenceran 10-3) dilakukan hal serupa berturut-turut sampai pengenceran 10-5. Dari pengenceran 10-3 dipipet sebanyak 1 ml, masukkan ke dalam cawan petri yang telah steril dan dilakukan hal yang sama pada pengenceran 10-4 dan 10-5. Dipakai suspensi tanah dari 3 pengenceran sebagai antisipasi bila pada pengenceran tersebut tidak diperoleh jamur pelarut fosfat. Selanjutnya tuangkan 12 ml media Pikovskaya (suhu sekitar 45-50ºC) ke dalam cawan petri yang telah berisi 1 ml
suspensi tanah, lalu putar cawan petri kearah kanan 3 kali dan ke arah kiri 3 kali agar media bercampur dengan suspensi tanah merata, biarkan sampai media
mengeras (padat). Setelah media mengeras, cawan petri diinkubasi pada inkubator dalam keadaan terbalik selama 3 hari dengan suhu 28-30ºC. Setelah diinkubasi selama 3 hari dilakukan pengamatan pada jamur yang tumbuh pada media.
4. Uji potensi pada media padat
Jamur pelarut fosfat yang murni selanjutnya diuji kemampuannya melarutkan fosfat dalam cawan petri berisi media Pikovskaya padat steril. Bahan yang digunakan dalam pembuatan media uji ini sama dengan bahan media
Pikovskaya pada tahap isolasi, namun Ca3(PO4)2 g/L pada media isolasi diganti dengan AlPO4 dengan dosis 5 g/L media. Media uji dimasukkan dalam cawan petri dan dibiarkan mengeras. Selanjutnya biakan murni ditumbuhkan pada media uji. Tiap biakan murni diberi 3 ulangan untuk mendapatkan rataan hasil yang valid. Inkubasi dilaksanakan selama 7 hari. Jamur pelarut fosfat yang membentuk
holozone paling cepat dengan diameter paling besar secara kualitatif di sekitar koloni menunjukkan besar kecilnya potensi jamur pelarut fosfat dalam melarutkan
unsur P dari bentuk yang tidak terlarut. Dihitung potensi jamur dengan menggunakan nilai indeks pelarutan yaitu nisbah antara diameter zona jernih terhadap diameter koloni (Premono, 1998).
5. Uji potensi pada media cair
Sebanyak 50 ml media Pikovskaya cair (sumber P AlPO4) ditempatkan dalam Erlenmeyer 250 ml yang disterilkan dalam autoklaf pada suhu 121˚C dengan tekanan 1,5 atm. Sebanyak 1 jarum ose spora jamur pelarut fosfat diinokulasikan pada media cair tersebut. Tiap biakan murni diberi 3 ulangan
untuk mendapatkan rataan hasil yang valid. Inkubasi secara diam dilakukan selama 7 hari pada suhu kamar. Setelah proses inokulasi selesai, kultur
ditentukan kadar P-tersedianya dengan metode kolorimetri dan dihitung dengan
Bray-II. Hal serupa dilakukan juga pada kontrol yaitu media tanpa inokulum dan dibandingkan hasil yang diperoleh. Jamur yang paling besar meningkatkan P terlarut secara kuantitatif pada media (dibandingkan dengan hasil kontrol)
merupakan jamur yang efektif dan potensial. Setelah itu pH medium diukur dengan pH meter untuk mengetahui pengaruh pelarutan fosfat oleh jamur
terhadap pH media.
6. Identifikasi jamur pelarut fosfat yang potensial melarutkan fosfat
Setelah diperoleh jamur pelarut fosfat paling potensial selanjutnya
dilakukan indentifikasi pada jamur tersebut. Biakan murni jamur diremajakan pada media potato dextrose agar (PDA) dan diinkubasi selama 3 hari. Jamur yang telah tumbuh pada media, diamati ciri-ciri makroskpisnya, yaitu ciri koloni seperti sifat tumbuh hifa, warna koloni dan diameter koloni. Jamur juga ditumbuhkan pada kaca objek yang diberi potongan PDA yang dioles tipis dengan spora JPF
potensial. Potongan agar kemudian ditutup dengan kaca objek. Biakan pada kaca objek ditempatkan dalam cawan petri yang telah diberi pelembab berupa kapas
basah. Biakan pada kaca diinkubasi selama 3 hari pada kondisi ruangan. Setelah masa inkubasi, jamur yang tumbuh pada kaca preparat diamati ciri mikroskopisnya yaitu ciri hifa, tipe percabangan hifa, serta ciri-ciri konidia
dibawah mikroskop. Ciri yang ditemukan dari masing-masing jamur kemudian dideskripsikan dan dicocokkan dengan buku indentifikasi jamur (Gilman, 1971).
HASIL DAN PEMBAHASAN
Karakteristik Sifat Kimia Sampel Tanah Gambut
Keberadaan mikroba di dalam tanah terutama dipengaruhi oleh sifat kimia
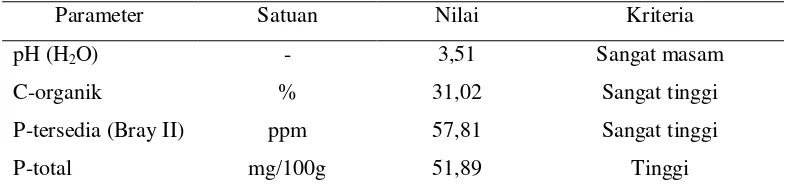
tanah. Hasil analisis sifat kimia sampel tanah dapat dilihat pada Tabel 1.
Tabel 1. Hasil analisis sifat kimia sampel tanah gambut
Parameter Satuan Nilai Kriteria
pH (H2O) - 3,51 Sangat masam
C-organik % 31,02 Sangat tinggi
P-tersedia (Bray II) ppm 57,81 Sangat tinggi
P-total mg/100g 51,89 Tinggi
Sumber kriteria: Staf Pusat Penelitian Tanah-Bogor dan BPP-Medan (Lampiran 2)
Hasil analisis menunjukkan pH tanah termasuk dalam kriteria sangat
masam. Menurut Barchia (2006) tingginya kemasaman tanah gambut disebabkan tingginya asam fenolat dan asam-asam organik lain hasil dekomposisi bahan
organik yang banyak mengandung lignin. Tapak pertukaran tanah gambut didominasi ion hidrogen yang merupakan kation yang berperan dalam pertukaran kation (KTK). Kation gambut akan mengikat basa-basa yang umumnya adalah
hara yang dibutuhkan tanaman. Jumlah ion H+ yang tinggi menyebabkan hara terikat semakin tinggi serta diikuti pula dengan penurunan pH. Penurunan pH
akan diikuti kejenuhan basa yang rendah. Hal ini juga berarti ketersediaan hara yang semakin kecil.
Ketersedian fosfat dalam tanah sangat dipengaruhi oleh pH karena P
sangat rentan diikat pada kondisi masam maupun alkalin. Ketersediaan fosfat akan menurun pada pH <5,5 atau >7,0. Pada kondisi masam aktivitas besi dan
aktivitas fiksasi atau jerapan dilakukan oleh kalsium dan magnesium yang banyak
tersedia dan larut. Menurut Poerwowidodo (2000), umumnya ketersediaan fosfat dalam tanah maksimum dijumpai pada kisaran pH 5,5–7,0.
Hasil pengukuran C-organik sampel tanah gambut adalah 31,02 %. Nilai
tersebut menggambarkan pada lahan gambut memiliki simpanan karbon yang sangat tinggi. Kadar C-organik dapat memberi gambaran kandungan bahan
organik dalam tanah. Bila nilai C-organik dikonversikan akan diperoleh kadar bahan organik sebesar 53,35. Bahan organik sebagai sumber hara makro dan mikro tanaman juga menjadi sumber nutrisi bagi kehidupan mikroba tanah yang
akan mempengaruhi populasi dan aktivitasnya. Selain sebagai sumber nutrisi, bahan organik berperan dalam menentukan nilai kapasitas tukar kation (KTK).
Kapasitas tukar kation tanah berhubungan dengan basa-basanya yang merupakan hasil dekomposisi bahan organik. Hal ini dikarenakan bahan organik yang terdekomposisi menghasilkan CO2 serta asam-asam organik yang akan melepaskan ion H+ yang mempengaruhi pH tanah. Nilai pH menunjukkan banyaknya konsentrasi ion hidrogen (H+) di dalam tanah. Semakin banyak H+ yang dilepaskan maka pH tanah akan semakin masam. Penurunan pH ini akan diikuti peningkatan nilai KTK tanah.
Kandungan bahan organik berhubungan dengan keadaan P-total serta
hubungan antara bahan organik dengan pH tanah. Bahan organik mengandung berbagai hara, termasuk fosfat yang akan terlepas selama dekomposisi baik dalam
pendekomposer. Jika pH mendukung, jumlah dan aktivitas dekomposer akan
meningkat sehingga semakin besar hara yang dilepaskan dalam tanah.
Mikroba tanah mampu menghasilkan enzim ekstraseluler yaitu kelompok enzim fosfatase dan fitase yang berperan dalam mekanisme pelarutan P-organik
menjadi P-anorganik secara biologis. Hal ini didukung oleh hasil penelitian Rao (1994) yang menunjukkan bahwa mikroba pelarut fosfat mampu meningkatkan
ketersediaan unsur P melalui aktivitas enzim. Enzim ini diproduksi selama proses dekomposisi bahan organik berlangsung. Dengan kata lain enzim fosfatase yang dihasilkan berbanding lurus dengan besar bahan organik yang didekomposisi.
Aktivitas fosfatase dalam tanah meningkat dengan meningkatnya C-organik, temperatur dan kelembaban. Selain faktor diatas, kemasaman atau pH turut
mempengaruhi aktivitas fosfatase. Hasil penelitian Fitriatin et al., (2008) menunjukkan bahwa pH media mempengaruhi aktivitas fosfatase. Jamur lebih dominan aktivitas fosfatasenya pada pH masam karena merupakan habitat yang
baik untuk pertumbuhannya yang optimum.
Pada kebanyakan tanah, P-organik berkorelasi dengan C-organik tanah
sehingga mineralisasi P meningkat dengan meningkatnya total C-organik. Semakin tinggi C-organik dan semakin rendah P-organik menggambarkan semakin meningkatnya imobilisasi P. Fosfat anorganik dapat diimobilisasi
menjadi P-organik oleh mikroba pelarut fosfat.
Berdasarkan penelitian Lestari dan Saraswati (1997), jamur pelarut fosfat
dikarenakan pertumbuhan optimum jamur berada pada kondisi masam.
Sebaliknya, kondisi masam kurang sesuai bagi pertumbuhan kelompok bakteri dan aktinomisetes. Pertumbuhan kelompok bakteri optimum pada pH netral dan meningkat seiring dengan meningkatnya pH.
Isolasi Jamur Pelarut Fosfat dari Bahan Tanah Gambut
Biakan campuran yang tumbuh di media isolasi diamati dan dihitung
jumlah mikroba yang mampu membentuk holozone. Populasi mikroba pelarut fosfat yang diperoleh tergolong tinggi yaitu 36,461×106 SPK/ml. Berdasarkan penampakan struktur tumbuhnya, mikroba pelarut fosfat hasil isolasi dibedakan
atas bakteri pelarut fosfat dan jamur pelarut fosfat. Bakteri pelarut fosfat tampak tumbuh pada 68 cawan petri dengan jumlah koloni 34,132×106 SPK/ml sedangkan JPF tampak tumbuh pada 34 cawan petri dengan jumlah koloni 2,329×106 SPK/ml. Dominannya keberadaan BPF dibanding JPF diperkirakan karena populasi mikroba pelarut fosfat dari kelompok bakteri di dalam tanah jauh
lebih besar dibanding kelompok jamur. Hal ini menyebabkan walau telah dilakukan pengenceran, suspensi tetap mengandung populasi BPF yang tinggi.
Hal ini sesuai pernyataan Ginting (2006) yang menyatakan bahwa populasi bakteri pelarut fosfat dapat mencapai 12×106 organisme per gram tanah sedangkan populasi jamur berkisar 2×104 - 1×106 organisme per gram tanah. Menurut Rao (1994) bakteri merupakan kelompok mikroba tanah paling dominan dan dapat melingkupi separuh dari biomassa mikroba dalam tanah.
Tingginya populasi BPF dibanding JPF yang diisolasi dari bahan gambut
tidak sesuai dengan pernyataan Hardjowigeno (1992) yang menyatakan bakteri berkembang baik pada pH 5,5 atau lebih, sedangkan pada pH kurang dari 5,5 perkembangannya sangat terhambat. Hal sebaliknya terjadi pada JPF yang
seharusnya tumbuh optimum pada kondisi masam, namun pada media isolasi memiliki jumlah populasi yang lebih rendah dibanding BPF. Sampel tanah yang
digunakan memiliki pH sangat masam yaitu 3,51. Dengan keadaan pH demikian seharusnya pertumbuhan BPF terhambat dan pertumbuhan JPF meningkat.
Tingginya populasi BPF kemungkinan karena media isolasi yang
digunakan telah diatur besar pH nya sekitar netral serta kandungan nutrisi media di dalamnya sesuai untuk pertumbuhan mikroba. Hal inilah yang menyebabkan
BPF dapat berkembang dengan pesat karena kondisi pH yang netral tidak menghambat pertumbuhannya serta nutrisi yang tercukupi membuat BPF mampu berkembang maksimal. Populasi BPF yang tinggi selain didukung media tumbuh
yang sesuai juga didukung sifat genetiknya yang berkembang lebih cepat dibanding jamur. Kondisi ini memungkinkan terjadinya penghambatan
pertumbuhan JPF oleh BPF pada media isolasi. Menurut Foth (1994), bakteri dapat berbiak sangat cepat dengan pembelahan biner yang dapat berlangsung tiap menit bahkan lebih cepat bila kondisi lingkungan dan nutrisi mendukung. Media
isolasi yang digunakan mengandung nutrisi lengkap untuk pertumbuhan mikroba. Kelengkapan nutrisi inilah yang kemungkinan turut mendukung tingginya
perkembangbiakan BPF.
Umumnya populasi mikroba di sekitar perakaran lebih tinggi dibanding daerah
yang lebih dalam. Hal ini disebabkan adanya eksudat akar tanaman yang menyediakan berbagai bahan organik yang dapat menstimulir pertumbuhan mikroba. Bahan organik yang dikeluarkan oleh akar dapat berupa eksudat akar,
sekresi akar, lisat akar maupun musigel yang secara langsung mempengaruhi hidupnya. Selain eksudat akar yang dihasilkan, akar tanaman mempengaruhi
kehidupan mikroba secara fisiologis. Mikroba yang berada dekat daerah perakaran akan lebih aktif daripada mikroba yang jauh dari perakaran. Kemungkinan hal inilah yang membuat pada tahap isolasi diperoleh banyak mikroba tanah termasuk
mikroba pelarut fosfat.
Pengambilan sampel dari daerah top soil didukung pernyataan Rao (1994) yang menyatakan bahwa jamur umumnya dijumpai pada tanah yang lebih dangkal dan jarang ditemukan di bagian tanah yang lebih dalam. Dijelaskan hal ini berdasarkan ketersediaan bahan organik dan rasio antara oksigen dan karbon
dioksida yang berbeda antara daerah permukaan dengan bagian tanah yang lebih dalam. Sehingga pemilihan sampel dari bagian top soil sesuai dengan tujuan penelitian yaitu untuk memperoleh jamur pelarut fosfat.
Hasil pemurnian isolasi 30 sampel tanah yang diambil dari 5 petak pengamatan pada kedalaman 0-20 cm diperoleh 18 isolat JPF. Isolasi
dan mendapatkan hasil pengujian yang valid dari aktivitas 1 jenis mikroba saja
(Fitter, 1991).
Kemampuan JPF Melarutkan P dalam Media Pikovskaya Padat
Jamur pelarut fosfat yang diperoleh selanjutnya diukur kemampuannya
melarutkan P-terikat pada media Pikovskaya padat. Sebagai sumber P media padat adalah AlPO4. Penggantian sumber fosfat ini mengacu pada penelitian yang telah dilakukan Premono (1994) dengan tujuan untuk menjaring mikroba dari tanah masam yang diduga mampu melarutkan aluminium fosfat maupun besi fosfat. Setiap perlakuan isolat dilakukan dengan 3 ulangan untuk mendapat hasil rataan
diameter holozone yang valid.
Jamur yang tumbuh pada media akan melarutkan P yang ditandai dengan
terbentuknya holozone yang mengelilingi JPF. Holozone terbentuk sebagai akibat terjadinya pelarutan butiran aluminium fosfat dari media. Evaluasi kemampuan JPF dilakukan dengan mengukur lebar sempitnya diameter holozone yang di sekeliling koloni. Cara ini umum dilakukan, namun karena tidak memperhitungkan faktor pertumbuhan koloni, sering menghasilkan hubungan
korelasi yang rendah antara lebar holozone dengan jumlah P-terlarut secara kualitatif. Menurut Premono (1994), hal tersebut dapat diatasi dengan menggunakan nilai indeks pelarutan (dissolving indeks) yaitu nisbah antara diameter holozone terhadap diameter koloni. Hasil indeks tersebut terbukti berkorelasi tinggi terhadap jumlah P yang dapat dilarutkan secara kualitatif. Maka
Hasil pengukuran diameter holozone menunjukkan dari 18 JPF hanya 12 JPF diantaranya yang mampu melarutkan fosfat dari persenyawaan aluminium fosfat. Hal ini menunjukkan bahwa JPF yang mampu melarutkan ikatan Ca3(PO4)2 pada media isolasi, belum tentu memiliki kemampuan untuk melarutkan fosfat
yang terikat aluminium. Hal ini sesuai dengan pernyataan Elfiati (2005) yang menyatakan setiap jamur memiliki kemampuan yang berbeda-beda dalam
melarutkan ikatan fosfat Ca3(PO4)2, AlPO4 dan FePO4. Pernyataan tersebut terbukti oleh isolat yang mampu melarutkan Ca3(PO4)2 pada media isolasi, namun tidak mampu melarutkan ikatan AlPO4 pada media uji potensi. Hasil pengukuran diameter holozone pada media uji padat dapat dilihat pada Tabel 2.
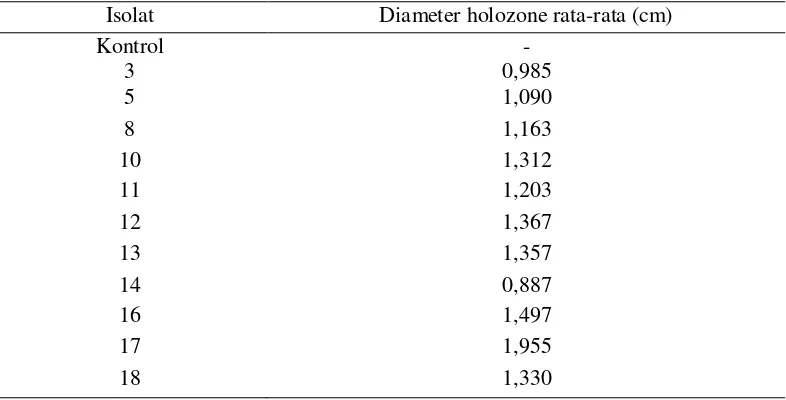
Tabel 2. Hasil pengukuran diameter holozone rata-ratapada media padat
Isolat Diameter holozone rata-rata (cm)
Kontrol
Di antara JPF yang mampu melarutkan P dari AlPO4 didapatkan 5 isolat yang paling besar membentuk holozone yaitu isolat 10, 12, 16, 17 dan 18. Diameter holozone terbesar ditunjukkan oleh isolat 17 dengan diameter holozone
Hal ini menunjukkan bahwa seluruh isolat JPF yang diuji memiliki efektivitas
yang berbeda dalam melarutkan fosfat pada media Pikovskaya padat.
Dilihat dari luasan holozone yang dihasilkan diketahui bahwa kemampuan jamur dalam melarutkan fosfat bervariasi. Perbedaan kemampuan tiap isolat JPF
membentuk holozone, diduga karena isolat JPF tidak berasal dari titik pengambilan sampel tanah yang sama. Titik pengambilan sampel yang berbeda
tentunya memiliki kondisi lingkungan, kelembaban, keadaan nutrisi dari bahan organik dan eksudat akar tegakan yang berbeda pula. Spesies jamur yang berbeda baik yang berasal dari tanah yang sama, maupun yang berasal dari tanah yang
berbeda, bisa sama ataupun berbeda kemampuannya dalan melarutkan fosfat. Sebaliknya, spesies jamur yang sama tetapi berasal dari tanah yang berbeda,
kemampuannya pun dapat berbeda. Menurut Goenadi (1994), kemampuan bakteri dan jamur pelarut fosfat dalam melarutkan fosfat berbeda-beda tergantung jenis strain.
Efektivitas JPF dalam melarutkan P terikat berkaitan erat dengan cara adaptasi JPF terhadap lingkungannya. Lahan gambut yang masam sesuai untuk
pertumbuhan JPF karena umumnya jamur tumbuh optimal pada kondisi masam. Hal ini memungkinkan efektifitas JPF melarutkan fosfat pada gambut menjadi tinggi. Dijelaskan Rao (1994), bahwa lingkungan yang sesuai akan meningkatkan
aktivitas JPF dalam mengeluarkan asam-asam organik dan enzim untuk melarutkan unsur P tanah serta beberapa hormon tumbuh.
molekul rendah hasil metabolismenya ke dalam tanah. Asam-asam organik
tersebut dapat membentuk kompleks stabil dengan kation-kation pengikat P di dalam tanah seperti Al dan Fe yang merupakan pengikat P pada tanah masam. Setiap JPF memiliki kemampuan yang berbeda secara genetik dalam
mengekskresikan jenis dan jumlah asam organik. Sifat asam organik lebih penting dari jumlah yang dihasilkan. Hal ini terlihat dari perbedaan kemampuan tiap jenis
JPF dalam melarutkan P. Efektivitas asam-asam organik yang dihasilkan tergantung pada kondisi lingkungan mikro di dalam tanah.
Menurut Ginting (2006), kemampuan tiap mikroba pelarut fosfat tumbuh
dan melarutkan fosfat berbeda-beda yang diidentifikasi dari luas holozone dan waktu terbentuknya. Mikroba pelarut fosfat yang unggul akan menghasikan
diameter holozone yang paling besar dan lebih cepat dibanding koloni lain.
Menurut Noor (2001), luas holozone yang terbentuk di sekitar koloni JPF secara kualitatif dapat menunjukkan besar kecilnya kemampuan JPF melarutkan
unsur P dari bentuk yang sukar larut. Inilah yang mendasari pemilihan 5 isolat diatas untuk dapat diuji kembali pada media cair. Hal ini diperkuat Tatiek (1991)
yang menyatakan daerah bening pada media padat tidak dapat menunjukkan banyak sedikitnya jumlah P terlarut yang dapat dihasilkan JPF, namun luas sempitnya daerah bening pada media padat dapat menunjukkan besar kecilnya
jamur dalam melarutkan P. Untuk itulah perlu dilakukan uji lebih lanjut pada media cair untuk mengetahui kemampuan isolat melarutkan fosfat secara
Pada pengujian di media padat tampak pertumbuhan tiap JPF
berbeda-beda. Menurut Goenadi (1994), hal ini dapat disebabkan beberapa hal:
1. konsentrasi fraksi AlPO4 dalam media yang tidak merata dalam petri mempengaruhi holozone yang terbentuk,
2. ketebalan media yang tidak seragam di dalam cawan akan mempengaruhi
holozone yang terbentuk. Fraksi aluminium fosfat pada media yang lebih tebal tentunya lebih sulit untuk dilarutkan daripada media yang tipis,
3. mikroba pelarut fosfat ada yang mampu tumbuh dengan cepat dan ada mikroba yang tumbuh lambat. Misalnya, Penicillium sp umumnya memiliki diamater koloni yang lebih kecil daripada Aspergillus sp. Nilai indeks pelarutan Penicillium sp lebih besar dari Aspergillus sp, tetapi kemampuannya melarutkan fosfat in vitro Penicillium sp lebih kecil daripada Aspergillus sp.
Kemampuan JPF Melarutkan P dalam Media Pikovskaya Cair
Lima isolat unggul dari uji media padat selanjutnya diuji pada media
Pikovskaya cair. Inkubasi dilakukan selama 7 hari pada suhu ruang (25-27˚C) karena pelarutan fosfat optimum pada kisaran 25-35˚C. Sebagai sumber P terikat
digunakan AlPO4. Berdasarkan hasil penelitian Goenadi (1994), uji in vitro lebih sensiftif daripada uji indeks pelarutan fosfat. Uji in vitro menggunakan media Pikovskaya tanpa agar. Metode ini tergolong lebih rumit dan mahal dalam
menganalisis besar P terbebas dalam media. Oleh karena itu umumnya uji ini dilakukan setelah uji indeks pelarutan fosfat pada media padat dengan jumlah
Besar fosfat terlarut yang dihasilkan tiap isolat JPF bervariasi satu dengan
yang lainnya. Pada kontrol ditemukan P terlarut sebesar 0,516 ppm. Pelarutan fosfat pada kontrol ini kemungkinan disebabkan pemanasan saat sterilisasi media. Sedangkan pada media yang diberikan isolat, besar P terlarut bervariasi antar
isolat. Pembebasan ikatan fosfat terjadi akibat reaksi antara AlPO4 terhadap asam organik yang dilepaskan JPF. Hasil pengukuran pH dan kadar P-tersedia media uji
cair dapat dilihat pada Tabel 3.
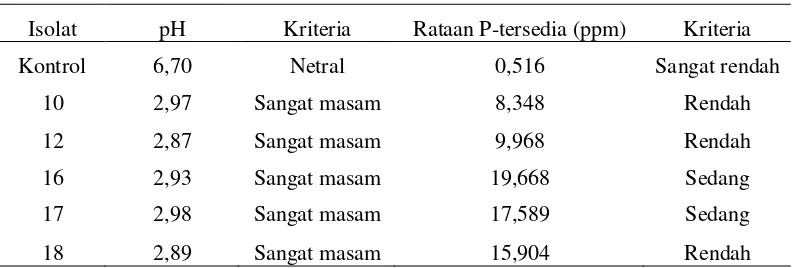
Tabel 3. Hasil pengukuran pH dan P tersedia
Isolat pH Kriteria Rataan P-tersedia (ppm) Kriteria
Kontrol 6,70 Netral 0,516 Sangat rendah
10 2,97 Sangat masam 8,348 Rendah
12 2,87 Sangat masam 9,968 Rendah
16 2,93 Sangat masam 19,668 Sedang
17 2,98 Sangat masam 17,589 Sedang
18 2,89 Sangat masam 15,904 Rendah
Sumber kriteria: Staf Pusat Penelitian Tanah-Bogor dan BPP-Medan (Lampiran 2)
Isolat yang menghasilkan P tersedia paling besar adalah isolat 16 dengan P
tersedia dari AlPO4 sebesar 19,668 ppm. Isolat yang menghasilkan P tersedia paling kecil adalah isolat 10 dengan dengan jumlah P terbebas dari AlPO4 sebesar 8,348 ppm. Besar P tersedia pada media diukur menggunakan metode Bray-II, dan pembacaannya menggunakan spektrometer dengan panjang gelombang 660 nm.
Selain mengukur kadar P-tersedia, dilakukan juga pengukuran pH media untuk mengetahui perubahan yang terjadi. Pada media yang tidak diberi isolat
yang diberi isolat JPF, nilai pH media cenderung menurun menjadi sangat masam
(Tabel 3).
Perubahan pH berperan penting dalam peningkatan kelarutan fosfat. Kisaran pH media setelah inkubasi adalah 2,87-2,98. Penurunan pH ini diduga
akibat pembebasan sejumlah asam-asam organik oleh JPF. Hal ini merupakan bentuk adaptasi JPF terhadap media yang mengandung P terikat yang lebih tinggi
dari P terlarut. Terlihat dari Tabel 3 bahwa peningkatan P tersedia diikuti penurunan pH media. Jamur pelarut fosfat sama halnya seperti tumbuhan, memerlukan fosfat dalam metabolismenya. Fosfat merupakan sumber energi
primer bagi oksidasi mikroba. Jamur pelarut fosfat dengan asam organik yang dihasilkannya mampu melepaskan P dari ikatannya menjadi bentuk terbebas.
Pembebasan fosfat dapat terjadi karena asam-asam organik ini akan membentuk khelat (kompleks stabil) dengan kation Al, Fe dan Ca yang mengikat P, sehingga ion H2PO4- terbebas (Rao, 1994). Jamur pelarut fosfat menggunakan sebagian kecil dari fosfat terbebas untuk metabolismenya dan sisanya dapat dimanfaatkan organisme lain seperti tumbuhan. Jika JPF mati maka P-organik yang terdapat
dalam jaringan mikroba akan lepas kembali dalam bentuk P-anorganik
Hal ini sesuai dengan pernyataan Rao (1994), yang menyatakan penurunan pH pada media disebabkan oleh asam-asam organik yang dihasilkan JPF sebagai
hasil metabolismenya diantaranya asam sitrat, glutamat, suksinat, laktat, oksalat, glioksalat, malat, fumarat, tartarat, dan ά-ketobutirat. Menurut Premono (1994)
proses anabolisme dalam sintesis asam amino dan makromolekul lain yang
dibutuhkan JPF untuk kelangsungan metabolismenya. Akibat refleks genetik, JPF menghasilkan asam-asam organik ini dalam jumlah berlebih, sehingga sebagian dari asam organik tersebut berdifusi keluar sel sebagai reaksi keseimbangan
osmosis (Premono, 1994).
Asam-asam organik melarutkan P pada media dan dalam tanah melalui
mekanisme antara lain: kompetisi anion ortofosfat pada tapak jerapan, perubahan pH media, pengikatan logam membentuk logam organik dan khelat oleh ligan organik. Terdapatnya asam-asam organik ini dalam tanah sangat penting artinya
dalam mengurangi ikatan P oleh unsur penjerapannya dan mengurangi daya racun logam seperti aluminium pada tanah masam. Menurut Premono (1994), kecepatan
pelarutan P dari mineral P oleh asam organik ditentukan oleh: (1) kecepatan difusi asam organik dari larutan tanah, (2) waktu kontak antara asam organik dan permukaan mineral, (3) tingkat dissosiasi asam organik, (4) tipe dan letak gugus
fungsi asam organik, (5) affinitas kimia agen pengkhelat terhadap logam dan (6) kadar asam organik dalam larutan tanah.
Urutan kemampuan asam organik dalam melarutkan fosfat adalah: asam sitrat > asam oksalat = asam tartarat = asam malat > asam laktat = asam fumarat = asam asetat. Asam organik yang mampu membentuk komplek yang lebih mantap
dengan kation logam akan lebih efektif dalam melepas Al dan Fe mineral tanah sehingga akan melepas P yang lebih besar. Urutan kemudahan fosfat terlepas
mengikuti urutan Ca3(PO4)2 > AlPO4 > FePO4 (Premono, 1994).
dikeluarkan sebagai bentuk toleransi tumbuhan terhadap jumlah logam yang
berlebihan sehingga bersifat racun dalam tanah. Asam organik yang dihasilkan akar tanaman di tanah masam berperan dalam penolakan Al melalui pelepasannya dari akar dan detoksifikasi Al dalam simpals. Asam organik seperti asam sitrat
dapat mengkhelat Al dan mereduksi pengaruh racunnya pada tumbuhan (Prasetiyono dan Tasliah, 2003).
Identifikasi Jamur Pelarut Fosfat dari Bahan Tanah Gambut
Identifikasi dilakukan pada JPF yang mampu membentuk holozone paling cepat, berwarna bening dengan diameter paling tinggi pada media padat serta
mampu melarutkan P-terikat paling besar pada media cair. Hal ini dilakukan karena banyaknya JPF yang diperoleh dari hasil isolasi namun tidak semuanya
mampu melarutkan bentuk AlPO4 dengan baik. Alasan lain, karena penelitian ini bertujuan untuk mencari JPF paling potensial. Sehingga hanya JPF yang menunjukkan potensi paling baik selama pengujian saja yang diidentifikasi.
Berdasarkan pengamatan secara makroskopis dan mikroskopis dapat disimpulkan isolat 10 dan 16 termasuk dalam genus Penicillium sp sedangkan isolat 12, 17 dan 18 termasuk dalam genus Aspergillus sp. Secara makroskopis, genus Penicilium sp tumbuh menebal seperti kapas putih. Pada bagian tengah koloni Penicillium sp yang berumur 7 hari berwarna hijau keabu-abuan yang diduga sebagai spora. Sedangkan pada Aspergillus sp yang berumur 7 hari, koloni bersporulasi lebat dan pada awal pertumbuhan membentuk lapisan padat yang
a
b
c
d e Untuk mengetahui gambaran morfologi sel Aspergillus sp dan Penicillium sp
dapat dilihat pada gambar di bawah ini.
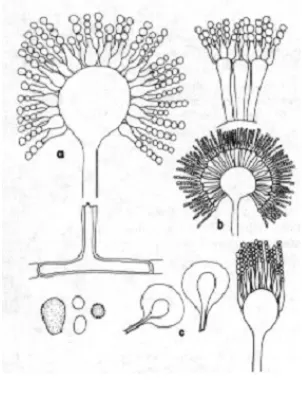
Gambar 2. Morfologi Aspergillus sp Gambar 3. Morfologi Penicillium sp Keterangan:
Gambar 2. Morfologi sel Aspergillus sp. a. Vesikel, b. Metulae, c. Spora (Malloch, 1997). Gambar 3. Morfologi sel Penicillium sp. (Madjid, 2010).
Hasil pengamatan langsung secara mikroskopis di bawah mikroskop dapat dilihat pada gambar di bawah ini.
Gambar 4. Aspergillus sp Gambar 5. Penicillium sp Keterangan:
Gambar 4. Penampakan Aspergillus sp. dibawah mikroskop (perbesaran 10 kali) (a. spora, b. tangkai konidia)
Ciri penampakan mikroskopis kedua genus JPF diuraikan sebagai berikut:
Aspergillus sp.
Tangkai konidiofor bening, dan umumnya berdinding tebal dan menyolok. Kepala konidia berbentuk bulat, kemudian merekah menjadi kolom-kolom yang
terpisah. Vesikula berbentuk bulat hingga semibulat, dan berdiameter 25-50 μm. Fialid terbentuk langsung pada vesikula atau pada metula (pada kepala konidia
yang besar), dan berukuran (10-15) × (4-8) μm. Metula berukuran (7-10) × (4-6) μm. Konidia berbentuk bulat hingga semibulat, berdiameter 5-6,5 μm, hitam.
Aspergillus sp tergolong mikroba mesofilik dengan pertumbuhan pada suhu 35ºC-37ºC (optimum), 6ºC-8ºC (minimum), 45ºC-47ºC (maksimum). Derajat keasaman untuk pertumbuhannya adalah 2 - 8,5 tetapi pertumbuhan akan lebih baik pada
kondisi keasaman atau pH yang rendah (Gilman, 1971). Taksonomi fungi Aspergillus sp:
Kingdom : Myceteae (Fungi) Divisio : Ascomycota Kelas : Eurotiomycetes Ordo : Eurotiales Famili : Trichocomaceae Genus : Aspergillus Spesies : Aspergillus sp.
Penicillium sp.
Konidiofor berukuran (400-500) × (3,0-4,0) μm, khususnya tepi koloni, berdinding tipis, berwarna bening, vertisil tidak teratur dan terdiri atas 3-4 tingkat
serta mempunyai cabang yang berkumpul. Fialid berbentuk agak silindris dengan leher pendek yang tidak mencolok dan berukuran (4,5-6,5) × (2,2-2,5) μm.
Taksonomi fungi Penicillium sp:
Spesies : Penicillium sp.
Jenis JPF yang paling banyak diteliti adalah Aspergillus sp dan Penicillium sp. Hal ini berdasarkan penelitian-penelitian yang telah dilakukan tentang JPF memberikan hasil banyak diantara spesies dari genus tersebut yang berkemampuan tinggi dalam melarutkan fosfat. Jamur pelarut fosfat memiliki kemampuan yang jauh melebihi BPF dalam melarutkan Ca3(PO4)2, AlPO4, dan FePO4. Indikasi tersebut menunjukkan bahwa kemampuan jamur yang mempunyai spektrum lebar dalam melarutkan beberapa bentuk ikatan P yang ada
di dalam tanah. Berdasarkan penelitian yang pernah dilakukan diketahui bahwa
Aspergillus niger dapat meningkatkan kelarutan P dari AlPO4 sebesar 135% dan dapat meningkatkan P larut pada tanah ultisol sebesar 30,4% dibandingkan
kontrol. Kelompok Penicillium sp mampu melarutkan 26-40% Ca3(PO4)2, sedangkan Aspergillus sp mampu melarutkan 18% (Anas, 1993).
Penelitian sejenis mendapatkan kombinasi Aspergillus sp dan Penicillium sp mampu meningkatkan ketersediaan P 26-40% sedangkan Aspergilus sp sekitar 18%. Aspergillus sp dapat dijumpai pada berbagai habitat dan kondisi yang berbeda serta tahan terhadap kondisi kelembaban yang rendah dan suhu ekstrim.
Penicillium sp dapat tumbuh pada temperatur 22-27˚ C, tumbuh optimal pada pH netral sampai agak masam (Gilman, 1971).
kemampuan yang berbeda dalam menghasilkan jenis dan jumlah asam organik.
Dengan melihat hasil pengujian JPF pada media padat dan cair dapat disimpulkan bahwa isolat 16 dan 17 memiliki kemampuan paling baik dalam melarutkan P dari AlPO4 baik pada media padat maupun media cair.
Berdasarkan penelitian yang telah dilakukan diketahui beberapa asam organik yang mampu dihasilkan Aspergillus sp seperti asam sitrat sedangkan
Penicillium sp mampu menghasilkan asam sitrat dan oksalat. Asam sitrat dan oksalat tergolong asam organik yang paling kuat dalam melepaskan ikatan P. Urutan kemampuan asam organik dalam melarutkan fosfat adalah: asam sitrat >
asam oksalat = asam tartarat = asam malat > asam laktat = asam fumarat = asam asetat. Karena kemampuannya menghasilkan asam organik tersebutlah
KESIMPULAN DAN SARAN
Kesimpulan1. Populasi mikroba pelarut fosfat yang diperoleh adalah 36,461×106 SPK/ml yang dibedakan atas bakteri pelarut fosfat sejumlah 34,132×106 SPK/ml dan jamur pelarut fosfat sejumlah 2,329×106 SPK/ml.
2. Hasil uji diperoleh diameter holozone terbesar 1,955 cm oleh isolat 17 dan diameter terkecil sebesar 0,887 cm oleh isolat 14. Hasil pengukuran P-tersedia
kontrol sebesar 0,516 ppm. Isolat yang menghasilkan P-tersedia paling besar adalah isolat 16 yaitu 19,668 ppm. Isolat yang menghasilkan P-tersedia paling
kecil adalah isolat 10 yaitu 8,348 ppm.
3. Hasil indentifikasi baik secara makroskopis maupun mikroskopis menunjukkan bahwa isolat 10 dan 16 termasuk genus Penicillium dan isolat 12, 17 dan 18 termasuk genus Aspergillus.
Saran
Perlu dilakukan uji potensi lanjutan dengan sumber P terikat FePO4 yang juga merupakan ikatan P pada gambut. Sehingga dapat diketahui JPF yang sangat potensial dalam membebaskan ikatan AlPO4 dan FePO4 pada lahan gambut.
DAFTAR PUSTAKA
Anas, I., E. Premono dan R. Widyastuti. 1993. Peningkatan Efisiensi Pemupukan P dengan Menggunakan Mikroorganisme Pelarut P. Pusat antar Universitas IPB. Bogor.
Balai Penelitian Bioteknologi Tanaman Pangan. 2001. BioPhos: Teknologi Maju untuk Meningkatkan Efisiensi Pemupukan Fosfat. Bogor.
Barchia, M.F. 2006. Gambut Agroekosistem dan Transformasi Karbon. UGM Press, Yogyakarta.
Beauchamp, E.G. dan D.J. Hume. 1997. Agricultural Soil Manipulation: The Use of Bacteris, Manuring and Plowing. p. 643-664. In J.D. Van Elsas, J.T. Trevors, and E.M.H. Wellington (Eds.), Modern Soil Microbiology. Marcel Dekker, New York.
Darmawijaya, M.I. 1992. Klasifikasi Tanah dan Teori Bagi Penelitian Tanah dan Pelaksanaan Pertanian di Indonesia. UGM Press, Yogyakarta.
Elfiati, D. 2005. Peranan Mikroba Pelarut Fosfat Terhadap Pertumbuhan Tanaman. USU e-Repository. Medan.
Ellis, M.B. 1993. Dematiaceous Hyphomycetes. CAB International. England.
Ervayenri, S., Sukarno. N., dan Kusmana. C. 1999. Arbuskula Mycorrhiza Fungi (AMF) Diversity in Peat Soil Influenced by Vegetation: Types Procedings of International Conference on Mycorrhiza in Sustainable Tropical Agriculture and Forest Ecosystem 27-30 Oktober 1997 Bogor, Indonesia. Fitriatin, B. N, B. Joy dan Subroto, T. The Influence of Organic Phosphorous
Substrate on Phosphatase activity of Soil Microbes. 2008. Procedding International Seminar of Chemistry. 30-31 October. Indonesia.
Fitriatin, B. N., A.Yuniarti., O.Mulyani., F.S.Fauziah, dan M.D Tiara. 2009. Pengaruh Mikroorganisme Pelarut Fosfat Dan Pupuk P terhadap P Tersedia, Aktivitas Fosfatase, Populasi Mikroorganisme Pelarut Fosfat, Konsentrasi P Tanaman dan Hasil Padi Gogo (Oryza sativa. L.) pada Ultisols. Jurnal Agrikultura, Vol. 20, No 3.
Fitter, A.H. 1991. Fisiologi Lingkungan Tanaman. UGM Press, Yogyakarta. Foth, H.D. 1994. Dasar-dasar Ilmu Tanah. Terjemahan Soemartono Adisoemarto.
Airlangga. Jakarta.
Ginting, R.C.B., R. Saraswati, dan E. Husen. 2006. Mikroorganisme Pelarut Fosfat. Pupuk Organik dan Pupuk Hayati. Balai Besar Litbang Sumber Daya Lahan Pertanian. Badan Penelitian dan Pengembangan Pertanian, Bogor.
Hal. 144-146
Goenadi, D.H., dan R. Saraswati. 1994. Kemampuan Melarutkan Fosfat dari Beberapa Isolat Fungi Pelarut Fosfat. Menara Perkebunan 61(3):61-66.
Hanafiah, A.S. 1994. Mikroorganisme Pelarut P Sebagai Suatu Alternatif Pengganti Fungsi Pupuk TSP dan Kapur dalam Upaya Mengatasi Ketersediaan Fosfat bagi Tanaman. Makalah dibawakan pada seminar hasil-hasil pertanian RATA XV. BKS-PTN Barat banda Lampung.
Hardjowigeno, S. 1992. Ilmu Tanah. Mediyatama Sarana Perkasa, Jakarta.
ICRAF. 2008. Lahan Gambut: Potensi untuk Pertanian dan Aspek Lingkungan. Bogor.
Joner, E.J., I.M. Aarle dan M. Vosatka. 2000. Phosphatase Activity of Extraradical Arbuscular Mycorrhiza Hyphae: a review. Plant Soil 226:199-210
Lakitan. B. 1993. Dasar-dasar Fisiologi Tumbuhan. PT. RajaGrafindo Persada. Jakarta.
Lestari, Y. dan R. Saraswati. 1997. Aktivitas Enzim Fosfatase Jamur Pelarut Fosfat pada Tanah Podzolik Merah Kuning dalam Prosiding Seminar Pembangunan Pertanian Berkelanjutan Menyongsong Era Globalisasi, Banjarmasin, 13-14 Maret 1997.
Madjid, 2010.
Malloch, D. 1997. Moulds, Isolation, Cultivation and Identivication Methods. Departement of Biology University of Toronto. Toronto. http://http://www.botany.utoronto.ca/ResearchLabs/MallochLab/Malloch /Moulds/Moulds.html
Marlina, M. 1997. Keragaman Bakteri Pelarut Fosfat pada Tanah di Lahan Hutan Primer, Hutan Sekunder, Pertanaman Kopi dan Lahan Kritis di Sumber Jaya Lampung Barat. Skripsi. Universitas Lampung. Bandar Lampung. Hlm 67
Nasution, R. R. 2005. Isolasi dan Uji Potensi Mikroorganisme Pelarut Fosfat yang Berasal dari Bahan Tanah Histosol. Skripsi. USU e-Repositiry. Medan. Noor. M. 2001. Pertanian Lahan Gambut: Potensi dan Kendala. Penerbit
Novizan. 2002. Petunjuk Pemupukan yang Efektif. Agromedia Pustaka, Jakarta.
Poerwowidodo. 2000. Telaah Kesuburan Tanah. Angkasa, Bandung.
Ponmurugan, P dan C. Gopi. 2006. In Vitro Production of Growth Regulators and Phospatase Activity by Phosphate Solubilizing Bacteria. African Journal of Biotechnology 5 (4): 348.
Prasetiyono, J dan Tasliah. 2003. Strategi Pendekatan Bioteknologi untuk Pemuliaan Tanaman Toleran Keracunan Aluminium. Jurnal Ilmu Pertanian Vol.10 No. 1: 64-67
Premono, E.M. 1994. Jasad Renik Pelarut Fosfat, Pengaruhnya Terhadap P tanah dan Efisiensi Pemupukan P Tanaman Tebu. Disertasi Program Pascasarjana IPB
Premono, E.M. 1998. Ulas Balik: Mikrob Pelarut Fosfat untuk Mengefisienkan Pupuk Fosfat dan Prospeknya di Indonesia (Enhancement of Phosphate-Fertilizer Efficiency by Phosphate Solubilizing Microbes and Its Prospect in Indonesia). Hayati 5(4):89-94.
Rahmawati, N. 2005. Pemanfaatan Biofertilizer Pada Pertanian Organik. Tesis. USU e-Repository. Medan.
Rao, N.S.S. 1994. Mikroorganisme Tanah dan Pertumbuhan Tanaman. UI Press, Jakarta.
Rosmarkam, A., dan N.W.Yuwono. 2002. Ilmu Kesuburan Tanah. Kanisius, Yogyakarta.
Setiawati, T. C. 1998. Efektifitas Mikroba Pelarut P dalam Meningkatkan Ketersediaan P dan Pertumbuhan Tembakau Besuki Na-Oogst (Nicotiana tabacum L.). Tesis. Program Pascasarjana. IPB. Bogor.
Sutanto, N. 2002. Penerapan Pertanian Organik. Kanisius, Jakarta.
Tatiek, H. 1991. Bakteri Pelarut Fosfat Asal Beberapa Jenis Tanah dan Efeknya terhadap Pertumbuhan dan Hasil Jagung (Zea mays L.). Disertasi Universitas Padjajaran. Bandung.
Tisdale, S.L, W.L. Nelson dan J.D. Beaton. 1990. Soil Fertility and Fertilizer. McMillan Publ.Co., New York. 4th ed.
Lampiran 1. Dokumentasi tahap penelitian
Gambar 1. Pengambilan sampel Gambar 2. Isolasi mikroba pelarut fosfat
Gambar 3. Pemurnian Gambar 4. Uji potensi media padat
Lampiran 2. Kriteria penilaian sifat kimia tanah Staf Pusat Penelitian Tanah Bogor (1983) dan BPP-Medan (1982)
Sifat tanah Satuan Sangat rendah
Rendah Sedang Tinggi Sanga
t
Netral Agak alkalis Alkalis
Lampiran 3. Prosedur penetapan P-tersedia tanah dengan metode Bray-II
Alat: shaker; pipet skala dan volumetrik; corong; spetrometer; tabung reaksi; Erlenmeyer.
Bahan:
1. Larutan Amonium fluoride: larutkan 3,7 g NH4F dengan H2O menjadi 100ml 2. Larutan HCl 5 N: larutkan 20,2 mL HCl pekat dengan H2O menjadi 500 ml 3. Larutan Bray-II: larutkan 30 ml larutan NH4F 1 N dengan 20 ml HCl 5 N
8. Pereaksi fosfat A: campurkan bahan nomor 4,5 dan 6, jadikan 2 L dengan menambahkan H2O
9. Pereaksi fosfat B: campurkan 1 g asam ascorbat ke dalam 200 ml pereaksi fosfat A
10.Larutan standar P 50 ppm: larutkan 0,275 g K2HPO4. 3H2O dengan H2O hingga 1 L
11.Larutan standar 0- 0,5- 1,0- 2,0- 3,0- 4,0- 5,0 ppm P: pipet larutan standar 50 ppm P masing-masing sebanyak 0-1-2-4-6-8 dan 10 ml ke dalam labu ukur 100 ml dan penuhkan dengan H2O
Cara kerja:
1. Timbang 2 g contoh tanah dan tempatkan pada gelas Erlenmeyer 250 cc 2. Tambahkan larutan Bray-II sebanyak 20 ml dan goncang selama 30 menit 3. Caring dengan kertas whatman No. 42
4. Pipet filtrat sebanyak 5 ml dan tempatkan pada tabung reaksi 5. Tembahkan pereaksi fosfat B sebanyak 10 ml, biarkan 5 menit
6. Ukur tarnsmitan pada spektrometer dengan panjang gelombang 660 nm 7. Pada saat bersamaan pipet masing-masing 5 ml larutan standar P 0- 0,5- 1,0-
2,0- 3,0- 4,0 dan 5,0 ppm P ke tabung reaksi, kemudian tambahkan 10 ml pereaksi fosfat B
8. Ukur transmitan standar pada spektrometer dengan panjang gelombang 660 nm.
Perhitungan:
Nilai absorben = - log transmitan/100