IDENTIFIKASI KERAGAMAN GEN HORMON
PERTUMBUHAN (GH-
Msp
I) PADA KERBAU LOKAL
(
Bubalus bubalis
)
SKRIPSI
LIDIA PUSPA AGUSTIANI
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
RINGKASAN
LIDIA PUSPA AGUSTIANI. 2009. Identifikasi Keragaman Gen Hormon Pertumbuhan (GH-MspI) pada Kerbau Lokal (Bubalus bubalis). Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Dr. Ir. Cece Sumantri, M.Agr.Sc. Pembimbing Anggota : Dr. Ir. Achmad Farajallah, M.Si.
Kerbau lokal (Bubalus bubalis) merupakan sumber plasma nutfah yang dapat dimanfaatkan dalam rangka meningkatkan ketersediaan pangan karena memiliki potensi besar sebagai penghasil daging. Perbaikan secara genetik merupakan usaha yang dapat dilakukan untuk meningkatkan produktivitas kerbau yaitu dengan mencari keragaman gen yang memiliki kaitan erat dengan pertumbuhan. Salah satunya adalah gen hormon pertumbuhan (GH-MspI).
Penelitian ini bertujuan untuk mengidentifikasi keragaman gen hormon pertumbuhan (GH-MspI) intron 3 pada kerbau lokal dengan metode PCR-RFLP yang dapat dijadikan sebagai dasar informasi untuk melakukan seleksi. Penelitian ini dilaksanakan selama empat bulan di Laboratorium Zoologi Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Sampel darah kerbau yang digunakan berjumlah 320 sampel yang diambil dari empat daerah, yaitu 75 sampel dari Semarang, 103 sampel dari Mataram, 65 sampel dari Medan dan 77 sampel dari Banten. Pengidentifikasian keragaman gen hormon pertumbuhan (GH-MspI) dilakukan menggunakan metode PCR-RFLP. Metode ini mendeteksi keragaman gen dengan memanfaatkan perbedaan runutan DNA yang bisa dikenali oleh enzim restriksi MspI. Total sampel yang berhasil diamplifikasi sebanyak 261 sampel dengan persentase keberhasilan sebesar 81,56%. Panjang fragmen hasil amplifikasi gen GH-MspI intron 3 adalah 327 pb dengan situs pemotongan pada posisi 104. Lokus GH-MspI pada kerbau lokal bersifat polimorfik dengan 3 genotipe, yaitu genotipe +/+ (223 pb dan 104 pb), genotipe +/- (327 pb, 223 pb dan 104 pb) dan genotipe -/- (327 pb). Frekuensi genotipe total yaitu +/+ (0,805), +/- (0,146) dan -/- (0,05) serta frekuensi alel total yaitu + (0,878) dan – (0,122). Genotipe +/+ dan tipe alel + memiliki frekuensi kemunculan yang paling besar pada tiap populasi. Nilai heterozigositas tertinggi terdapat pada populasi kerbau asal Semarang (0,562) dan terendah pada populasi asal Banten (0,093). Rata-rata heterozigositas dari semua populasi adalah 0,370. Nilai indeks fiksasi gen GH menunjukkan bahwa tidak terjadi fiksasi gen GH karena nilainya menyebar dan tidak sama dengan nol.
ABSTRACT
Identification of Growth Hormone Gene (GH-MspI) in Local Buffalo (Bubalus bubalis)
Agustiani, L. P., C. Sumantri and A. Farajallah
The aim of this study was to identify polymorphism of buffalo’s growth hormone (GH-MspI) gene that localized in intron 3 using Polymerase Chain Reaction (PCR) – Restriction Fragment Length Polymorphism (RFLP) method. Total of the blood samples were 320 which is from Semarang, Medan, Mataram and Banten. Polymorphism in 261 samples were identified and revealed three genotypies that are 223 bp and 104 bp for the +/+ genotype; 327 bp, 222 bp and 104 bp for the +/- genotype and 327 bp (no digestion) for the -/- genotype. Frequencies of the +/+ genotype and + allele were the highest in whole populations. Frequencies were 0.805 for +/+ genotype, 0.146 for +/- genotype, and 0.05 for -/- genotype. The frequency of + allele was 0.878. The frecuencies of + allele and +/+ genotype were the highest. The highest heterozygocities was 0.562 for Semarang population and the lowest was 0.093 for Banten population and the average heterozygocities for all populations detected was 0.370. Index fixation value of GH gene showed there was no gene fixation in all populations.
IDENTIFIKASI KERAGAMAN GEN HORMON PERTUMBUHAN
(GH-
Msp
I) PADA KERBAU LOKAL
(
Bubalus bubalis
)
LIDIA PUSPA AGUSTIANI D14052429
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
IDENTIFIKASI KERAGAMAN GEN HORMON
PERTUMBUHAN (GH-
Msp
I) PADA KERBAU LOKAL
(
Bubalus bubalis
)
Oleh
LIDIA PUSPA AGUSTIANI D14052429
Skripsi ini telah disetujui dan disidangkan di hadapan Komisi Ujian Lisan pada tanggal 14 April 2009
Pembimbing Utama Pembimbing Anggota
Dr. Ir. Cece Sumantri, M. Agr. Sc Dr. Ir. Achmad Farajallah, M. Si
Dekan Ketua Departemen
Fakultas Peternakan Ilmu Produksi dan Teknologi Peternakan Institut Pertanian Bogor Fakultas Peternakan
Institut Pertanian Bogor
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 15 Agustus 1987 di Bogor. Penulis
merupakan anak pertama dari tiga bersaudara dari pasangan (Alm.) Bapak N. Walidi
dan Ibu Lilis Djubaedah.
Penulis menyelesaikan sekolah dasar pada tahun 1999 dari SD Negeri
Ciriung 02, Cibinong. Pendidikan lanjutan menengah pertama diselesaikan pada
tahun 2002 di SLTP Negeri 1 Cibinong dan pendidikan lanjutan menengah atas
diselesaikan pada tahun 2005 di SMA Negeri 3 Bogor.
Penulis diterima sebagai mahasiswa pada Departemen Ilmu Produksi dan
Teknologi Peternakan, Fakultas Peternakan Institut Pertanian Bogor melalui jalur
Undangan Seleksi Masuk IPB (USMI) pada tahun 2005.
Selama mengikuti pendidikan, penulis aktif di organisasi dan kegiatan
kemahasiswaan yaitu HIMAPROTER 2006/2007 dan 2007/2008, Uni Konservasi
Fauna (UKF) 2005/2006 dan 2006/2007, dan Paduan Suara Fakultas Peternakan
Graziono Symponia. Selain itu, penulis juga berperan aktif dalam berbagai
KATA PENGANTAR
Puji syukur Penulis panjatkan ke hadirat Allah SWT atas limpahan rahmat,
taufik dan hidayah-Nya sehingga Penulis dapat menyelesaikan studi, penelitian,
seminar dan skripsi ini. Shalawat dan salam semoga senantiasa dicurahkan kepada
Nabi Muhammad SAW.
Skripsi dengan judul “Identifikasi Keragaman Gen GH-MspI pada Kerbau
Lokal (Bubalus bubalis)” ini disusun sebagai salah satu syarat untuk memperoleh
gelar sarjana Peternakan, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian
ini bertujuan untuk mengidentifikasi keragaman gen hormon pertumbuhan
(GH-MspI) intron 3 pada kerbau lokal dengan metode PCR-RFLP yang dapat dijadikan
sebagai dasar informasi untuk melakukan seleksi.
Penulis menyadari sepenuhnya bahwa dalam penulisan skripsi ini masih jauh
dari sempurna, oleh sebab itu Penulis memohon maaf apabila terdapat kekurangan.
Tak lupa ucapan terima kasih Penulis sampaikan kepada semua pihak yang turut
membantu penyusunan skripsi ini, hanya Allah Yang Maha Pemurah dan Penyayang
yang akan membalasnya. Semoga skripsi ini bermanfaat bagi dunia pendidikan dan
peternakan serta menjadi catatan amal saleh. Amin.
Bogor, April 2009
DAFTAR ISI
Restriction Fragment Length Polymorphism (RFLP) .... 6
Single-Strand Conformation Polymorphism (SSCP) ... 6
Denaturing Gradient Gel Electrophoresis (DGGE) ... 7
Halaman
Pewarnaan Perak... 11
Pendeteksian Keragaman DNA ... 11
HASIL DAN PEMBAHASAN ... 13
Amplifikasi Gen Hormon Pertumbuhan (GH-MspI) ... 13
Pendeteksian Keragaman Gen GH-MspI dengan PCR ... 14
Keragaman Gen GH-MspI Kerbau Lokal ... 16
Nilai Heterozigositas ... 17
Indeks Fiksasi ... 18
KESIMPULAN DAN SARAN ... 19
Kesimpulan ... 19
Saran ... 19
UCAPAN TERIMAKASIH ... 20
DAFTAR PUSTAKA ... 21
DAFTAR TABEL
Nomor Halaman
1. Populasi Kerbau Terbanyak di 10 Propinsi Indonesia ... 4
2. Hasil Perhitungan Frekuensi Genotipe dan Frekuensi Alel
Gen GH-MspI ... 16
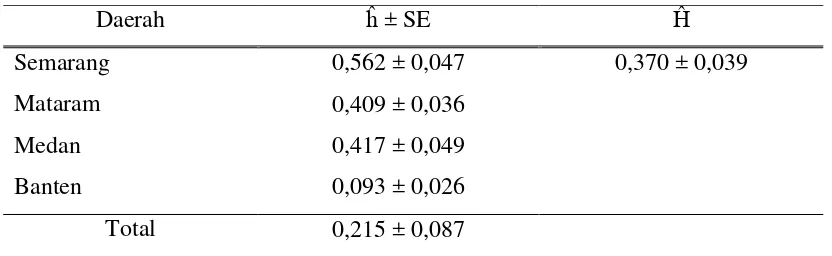
3. Nilai heterozigositas (ĥ) dan Rataan Heterozigositas (Ĥ) Gen
GH- MspI Kerbau Lokal... 17
IDENTIFIKASI KERAGAMAN GEN HORMON
PERTUMBUHAN (GH-
Msp
I) PADA KERBAU LOKAL
(
Bubalus bubalis
)
SKRIPSI
LIDIA PUSPA AGUSTIANI
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
RINGKASAN
LIDIA PUSPA AGUSTIANI. 2009. Identifikasi Keragaman Gen Hormon Pertumbuhan (GH-MspI) pada Kerbau Lokal (Bubalus bubalis). Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Dr. Ir. Cece Sumantri, M.Agr.Sc. Pembimbing Anggota : Dr. Ir. Achmad Farajallah, M.Si.
Kerbau lokal (Bubalus bubalis) merupakan sumber plasma nutfah yang dapat dimanfaatkan dalam rangka meningkatkan ketersediaan pangan karena memiliki potensi besar sebagai penghasil daging. Perbaikan secara genetik merupakan usaha yang dapat dilakukan untuk meningkatkan produktivitas kerbau yaitu dengan mencari keragaman gen yang memiliki kaitan erat dengan pertumbuhan. Salah satunya adalah gen hormon pertumbuhan (GH-MspI).
Penelitian ini bertujuan untuk mengidentifikasi keragaman gen hormon pertumbuhan (GH-MspI) intron 3 pada kerbau lokal dengan metode PCR-RFLP yang dapat dijadikan sebagai dasar informasi untuk melakukan seleksi. Penelitian ini dilaksanakan selama empat bulan di Laboratorium Zoologi Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Sampel darah kerbau yang digunakan berjumlah 320 sampel yang diambil dari empat daerah, yaitu 75 sampel dari Semarang, 103 sampel dari Mataram, 65 sampel dari Medan dan 77 sampel dari Banten. Pengidentifikasian keragaman gen hormon pertumbuhan (GH-MspI) dilakukan menggunakan metode PCR-RFLP. Metode ini mendeteksi keragaman gen dengan memanfaatkan perbedaan runutan DNA yang bisa dikenali oleh enzim restriksi MspI. Total sampel yang berhasil diamplifikasi sebanyak 261 sampel dengan persentase keberhasilan sebesar 81,56%. Panjang fragmen hasil amplifikasi gen GH-MspI intron 3 adalah 327 pb dengan situs pemotongan pada posisi 104. Lokus GH-MspI pada kerbau lokal bersifat polimorfik dengan 3 genotipe, yaitu genotipe +/+ (223 pb dan 104 pb), genotipe +/- (327 pb, 223 pb dan 104 pb) dan genotipe -/- (327 pb). Frekuensi genotipe total yaitu +/+ (0,805), +/- (0,146) dan -/- (0,05) serta frekuensi alel total yaitu + (0,878) dan – (0,122). Genotipe +/+ dan tipe alel + memiliki frekuensi kemunculan yang paling besar pada tiap populasi. Nilai heterozigositas tertinggi terdapat pada populasi kerbau asal Semarang (0,562) dan terendah pada populasi asal Banten (0,093). Rata-rata heterozigositas dari semua populasi adalah 0,370. Nilai indeks fiksasi gen GH menunjukkan bahwa tidak terjadi fiksasi gen GH karena nilainya menyebar dan tidak sama dengan nol.
ABSTRACT
Identification of Growth Hormone Gene (GH-MspI) in Local Buffalo (Bubalus bubalis)
Agustiani, L. P., C. Sumantri and A. Farajallah
The aim of this study was to identify polymorphism of buffalo’s growth hormone (GH-MspI) gene that localized in intron 3 using Polymerase Chain Reaction (PCR) – Restriction Fragment Length Polymorphism (RFLP) method. Total of the blood samples were 320 which is from Semarang, Medan, Mataram and Banten. Polymorphism in 261 samples were identified and revealed three genotypies that are 223 bp and 104 bp for the +/+ genotype; 327 bp, 222 bp and 104 bp for the +/- genotype and 327 bp (no digestion) for the -/- genotype. Frequencies of the +/+ genotype and + allele were the highest in whole populations. Frequencies were 0.805 for +/+ genotype, 0.146 for +/- genotype, and 0.05 for -/- genotype. The frequency of + allele was 0.878. The frecuencies of + allele and +/+ genotype were the highest. The highest heterozygocities was 0.562 for Semarang population and the lowest was 0.093 for Banten population and the average heterozygocities for all populations detected was 0.370. Index fixation value of GH gene showed there was no gene fixation in all populations.
IDENTIFIKASI KERAGAMAN GEN HORMON PERTUMBUHAN
(GH-
Msp
I) PADA KERBAU LOKAL
(
Bubalus bubalis
)
LIDIA PUSPA AGUSTIANI D14052429
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
IDENTIFIKASI KERAGAMAN GEN HORMON
PERTUMBUHAN (GH-
Msp
I) PADA KERBAU LOKAL
(
Bubalus bubalis
)
Oleh
LIDIA PUSPA AGUSTIANI D14052429
Skripsi ini telah disetujui dan disidangkan di hadapan Komisi Ujian Lisan pada tanggal 14 April 2009
Pembimbing Utama Pembimbing Anggota
Dr. Ir. Cece Sumantri, M. Agr. Sc Dr. Ir. Achmad Farajallah, M. Si
Dekan Ketua Departemen
Fakultas Peternakan Ilmu Produksi dan Teknologi Peternakan Institut Pertanian Bogor Fakultas Peternakan
Institut Pertanian Bogor
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 15 Agustus 1987 di Bogor. Penulis
merupakan anak pertama dari tiga bersaudara dari pasangan (Alm.) Bapak N. Walidi
dan Ibu Lilis Djubaedah.
Penulis menyelesaikan sekolah dasar pada tahun 1999 dari SD Negeri
Ciriung 02, Cibinong. Pendidikan lanjutan menengah pertama diselesaikan pada
tahun 2002 di SLTP Negeri 1 Cibinong dan pendidikan lanjutan menengah atas
diselesaikan pada tahun 2005 di SMA Negeri 3 Bogor.
Penulis diterima sebagai mahasiswa pada Departemen Ilmu Produksi dan
Teknologi Peternakan, Fakultas Peternakan Institut Pertanian Bogor melalui jalur
Undangan Seleksi Masuk IPB (USMI) pada tahun 2005.
Selama mengikuti pendidikan, penulis aktif di organisasi dan kegiatan
kemahasiswaan yaitu HIMAPROTER 2006/2007 dan 2007/2008, Uni Konservasi
Fauna (UKF) 2005/2006 dan 2006/2007, dan Paduan Suara Fakultas Peternakan
Graziono Symponia. Selain itu, penulis juga berperan aktif dalam berbagai
KATA PENGANTAR
Puji syukur Penulis panjatkan ke hadirat Allah SWT atas limpahan rahmat,
taufik dan hidayah-Nya sehingga Penulis dapat menyelesaikan studi, penelitian,
seminar dan skripsi ini. Shalawat dan salam semoga senantiasa dicurahkan kepada
Nabi Muhammad SAW.
Skripsi dengan judul “Identifikasi Keragaman Gen GH-MspI pada Kerbau
Lokal (Bubalus bubalis)” ini disusun sebagai salah satu syarat untuk memperoleh
gelar sarjana Peternakan, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian
ini bertujuan untuk mengidentifikasi keragaman gen hormon pertumbuhan
(GH-MspI) intron 3 pada kerbau lokal dengan metode PCR-RFLP yang dapat dijadikan
sebagai dasar informasi untuk melakukan seleksi.
Penulis menyadari sepenuhnya bahwa dalam penulisan skripsi ini masih jauh
dari sempurna, oleh sebab itu Penulis memohon maaf apabila terdapat kekurangan.
Tak lupa ucapan terima kasih Penulis sampaikan kepada semua pihak yang turut
membantu penyusunan skripsi ini, hanya Allah Yang Maha Pemurah dan Penyayang
yang akan membalasnya. Semoga skripsi ini bermanfaat bagi dunia pendidikan dan
peternakan serta menjadi catatan amal saleh. Amin.
Bogor, April 2009
DAFTAR ISI
Restriction Fragment Length Polymorphism (RFLP) .... 6
Single-Strand Conformation Polymorphism (SSCP) ... 6
Denaturing Gradient Gel Electrophoresis (DGGE) ... 7
Halaman
Pewarnaan Perak... 11
Pendeteksian Keragaman DNA ... 11
HASIL DAN PEMBAHASAN ... 13
Amplifikasi Gen Hormon Pertumbuhan (GH-MspI) ... 13
Pendeteksian Keragaman Gen GH-MspI dengan PCR ... 14
Keragaman Gen GH-MspI Kerbau Lokal ... 16
Nilai Heterozigositas ... 17
Indeks Fiksasi ... 18
KESIMPULAN DAN SARAN ... 19
Kesimpulan ... 19
Saran ... 19
UCAPAN TERIMAKASIH ... 20
DAFTAR PUSTAKA ... 21
DAFTAR TABEL
Nomor Halaman
1. Populasi Kerbau Terbanyak di 10 Propinsi Indonesia ... 4
2. Hasil Perhitungan Frekuensi Genotipe dan Frekuensi Alel
Gen GH-MspI ... 16
3. Nilai heterozigositas (ĥ) dan Rataan Heterozigositas (Ĥ) Gen
GH- MspI Kerbau Lokal... 17
DAFTAR GAMBAR
Nomor Halaman
1. Pita DNA hasil elektroforesis setelah pewarnaan perak. Fragmen
hasil amplifikasi PCR ditambah dengan enzim pemotong MspI. ... 12
2. Posisi Penempelan Primer (cetak tebal) pada sekuen Gen GH-MspI. Terjadi Mutasi pada Situs Pemotongan C|CGG pada Posisi 104
(C menjadi T)(Gordon et al., 1983)... 14
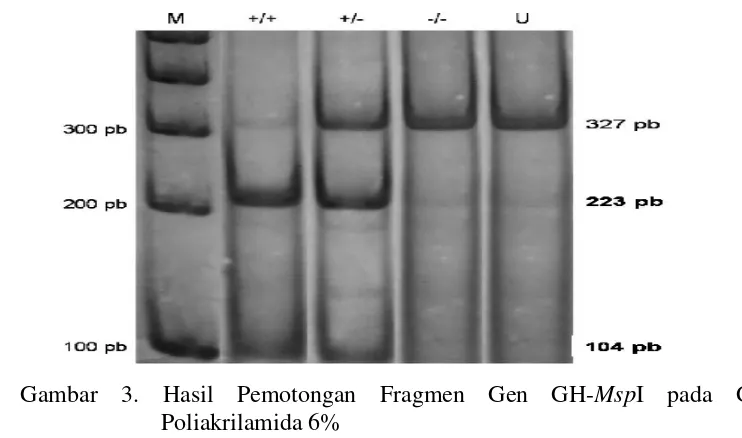
3. Hasil Pemotongan Fragmen Gen GH-MspI pada Gel
Poliakrilamida 6% ... 15
DAFTAR LAMPIRAN
Nomor Halaman
1. Modifikasi Metode Ekstraksi DNA Menggunakan Genomic DNA
Mini Kit Geneaid ... 26
1 PENDAHULUAN
Latar Belakang
Kerbau lokal (Bubalus bubalis) merupakan sumber plasma nutfah yang dapat
dimanfaatkan dalam rangka meningkatkan ketersediaan pangan. Kerbau memiliki
potensi besar sebagai penghasil daging. Tercatat bahwa pertambahan bobot badan
kerbau yang digemukkan bisa mencapai 0,8 sampai 1 kg per hari yang berarti tidak
kalah dengan pengemukkan sapi (Bahri dan Talib, 2007). Akan tetapi di Indonesia
pemeliharaan ternak kerbau masih dilakukan secara tradisional serta hanya ditujukan
untuk ternak kerja dan tabungan hidup. Hal ini mengakibatkan rendahnya
pertumbuhan kerbau dan menurunnya populasi kerbau di Indonesia. Pertumbuhan
populasi kerbau dari data statistik peternakan adalah sekitar 3,41% dengan jumlah
total populasi 2,201 juta ekor. Perkembangan ternak kerbau di Indonesia selama 5
tahun terakhir menunjukkan terjadinya penurunan populasi pada tahun 2004 dan
2005 masing-masing sebesar 2,28% dan 11,43% dibandingkan dengan tahun
sebelumnya (Direktorat Jenderal Peternakan, 2006).
Upaya perbaikan pertumbuhan dapat dilakukan dengan seleksi untuk
meningkatkan kualitas ternak kerbau. Kemajuan teknologi sekarang ini
memungkinkan seleksi langsung pada DNA yang mengontrol sifat-sifat ekonomis.
Keterkaitan antara keragaman DNA dengan sifat-sifat ekonomis ini dapat dijadikan
dasar untuk melakukan metode MAS (Marker Assisted Selection). Metode MAS
merupakan cara yang lebih unggul untuk melakukan seleksi karena dapat
mempercepat pengambilan keputusan dalam manajemen ternak.
Gen hormon pertumbuhan (GH) merupakan salah satu gen yang berkaitan
erat dengan pertumbuhan. Berdasarkan penelitian, gen hormon pertumbuhan (GH)
berpengaruh terhadap produksi ternak. Oleh karena itu, hormon pertumbuhan
merupakan kandidat gen yang dapat digunakan untuk penanda genetik dalam
program seleksi kerbau.
Tujuan
Penelitian ini bertujuan untuk mengidentifikasi keragaman gen hormon
pertumbuhan (GH-MspI) intron 3 pada kerbau lokal (Bubalus bubalis) pada populasi
2 TINJAUAN PUSTAKA
Kerbau
Klasifikasi kerbau menurut Bhattacharya (1993) adalah kelas Mamalia, ordo
Ungulata, famili Bovidae, sub familia Bovinae, genus Bubalus, dan spesies bubalis.
Kerbau termasuk dalam spesies Bubalus bubalis diduga hasil domestikasi dari
Bubalis arnee, kerbau liar dari India yang dijumpai pada daerah Assam. Ternak
kerbau adalah ternak semi akuatik. Indonesia adalah habitat yang baik untuk ternak
kerbau dimana 40% dari wilayah Indonesia beriklim tropis basah (Praharani dan
Triwulanningsih, 2007).
Kerbau diketahui memiliki beberapa kelebihan dibandingkan dengan sapi.
Diwyanto dan Handiwirawan (2006) menyatakan bahwa kerbau dapat hidup dalam
kondisi pakan yang berkualitas rendah. Kemampuan ini menjadikan peternakan
kerbau sangat baik untuk dikembangkan karena dengan input yang sedikit, kerbau
masih dapat menghasilkan output yang tinggi. Selain itu, pemeliharaan kerbau yang
masih ditujukan untuk ternak kerja sedikit demi sedikit harus diubah karena sudah
dapat digantikan dengan mesin atau traktor. Dengan demikian potensi kerbau sebagai
penghasil daging dapat lebih dioptimalkan dengan melakukan pemeliharaan yang
sesuai.
Berdasarkan karakteristiknya, kerbau dibagi menjadi dua, yaitu kerbau sungai
dan kerbau lumpur (rawa). Kedua bangsa kerbau ini merupakan kerbau Asia yang
telah didomestikasi dan memiliki jumlah pasangan kromosom yang berbeda, yaitu
kerbau lumpur memiliki 48 kromoson dan kerbau sungai memiliki 50 kromosom.
Perkawinan keduanya akan menghasilkan keturunan yang juga fertil baik pada ternak
jantan maupun betina, hanya diduga bahwa daya reproduksi crossbred tersebut lebih
rendah dibandingkan masing-masing tetuanya (Bahri dan Talib, 2007).
Kerbau sungai menunjukkan kesenangan terhadap air mengalir yang bersih,
sedangkan kerbau lumpur suka berkubang dalam lumpur, rawa-rawa dan air
menggenang. Kerbau tipe lumpur biasa digunakan sebagai ternak kerja dan penghasil
daging sedangkan kerbau sungai merupakan tipe penghasil susu (Hasinah dan
Handiwirawan, 2006). Kerbau lumpur memiliki dua bentuk tanduk, yaitu bertanduk
normal yang memanjang ke belakang lalu melengkung ke atas dan tanduk tergantung
3 sungai (Murrah) memiliki dua tipe tanduk yaitu tanduk normal melingkar ke atas
membentuk spiral dan tanduk tergantung yaitu tanduk jatuh ke bawah lalu
melengkung ke dalam (Asoen, 2008). Kerbau rawa memiliki beberapa ciri-ciri yang
spesifik, diantaranya 1) warna kulit kerbau rawa pada umumnya abu-abu, 2)
memiliki garis kalung (chevron), 3) memiliki unyeng-unyeng (whorls) yang terdapat
di pinggang, dan 4) memiliki warna kaki putih dan hitam (Erdiansyah, 2008).
Populasi Kerbau di Indonesia
Ternak kerbau di Indonesia sebagian besar merupakan kerbau rawa (swamp
buffalo) yang mencapai 95% dengan keragaman warna, ukuran dan tingkah laku
yang besar pula. Sisanya sebanyak 5% termasuk kerbau sungai (river buffalo) yang
banyak dipelihara di Sumatera Utara. Puslitbang Peternakan (2006) menyatakan
bahwa ada kecenderungan populasi dan mutu genetik kerbau nasional menurun dari
tahun ke tahun, karena sistem perkawinannya tidak menentu.
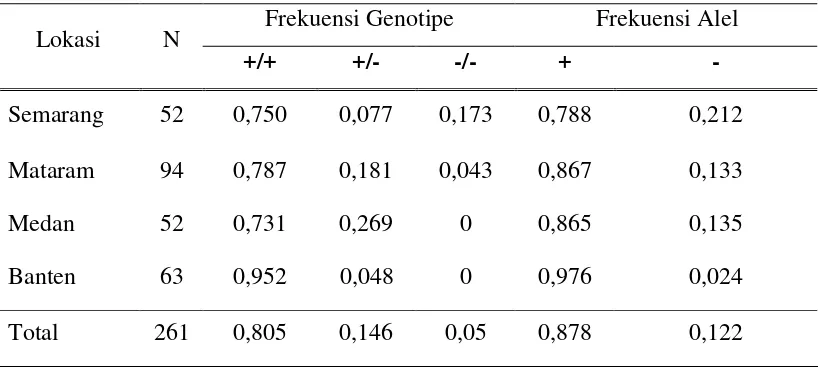
Tabel 1. Populasi Kerbau Terbanyak di 10 Propinsi Indonesia
Lokasi Jumlah (ekor)
Data populasi kerbau di Indonesia yang diperoleh dari Direktorat Jenderal
Peternakan (2006) menunjukkan bahwa rataan pertumbuhan populasi kerbau di
Indonesia adalah sekitar 3,41% per tahun. Jumlah populasi kerbau di Indonesia
adalah sebanyak 2,201 juta ekor yang menyebar hampir di seluruh propinsi tetapi
4 terakhir menunjukkan terjadinya penurunan populasi pada tahun 2004 dan 2005
masing-masing sebesar 2,28% dan 11,43% dibandingkan dengan tahun sebelumnya.
Sementara pada tahun 2003 dan 2006 terjadi peningkatan masing-masing sebesar
2,35% dan 3,41%.
Karakteristik Pertumbuhan Kerbau
Kerbau merupakan ternak yang lambat dewasa secara seksual, umur dewasa
kelamin antara 2 sampai 3 tahun bergantung dari tipe, kondisi manajemen,
perkawinan dan pakan. Kerbau dapat hidup sampai 15-20 tahun. Kerbau sungai
memiliki bobot jantan dewasa antara 300–700 kg dan 250-650 kg pada betina,
sedangkan kerbau lumpur jantan memiliki bobot dewasa 500 kg dan kerbau betina
400 kg dengan tinggi pundak jantan dan betina adalah 135 dan 130 cm (Fahimuddin,
1975). BPTP (2002) menyebutkan bahwa kerbau lumpur pada umur 1 tahun dapat
mencapai bobot 195-200 kg, panjang badan 95,4-97,6 cm dan lingkar dada
135,7-138,4 cm. Kerbau lumpur dewasa berumur 3 tahun mencapai berat badan 400-500 kg
dengan panjang badan 128-138 cm dan lingkar dada 174,6-177,0 cm. Kerbau
lumpur memiliki bentuk tubuh yang gempal, padat dan berisi yang membuktikan
bahwa kerbau ini mampu mengubah pakan yang berkualitas rendah menjadi daging.
Laju pertumbuhan kerbau terbilang lebih rendah dibandingkan dengan sapi, hal ini
terkait dengan manajemen pemeliharaannya (Fahimuddin, 1975).
Peternakan Kerbau di Indonesia
Di Indonesia, pemeliharaan kerbau masih dilakukan secara tradisional, baik
dalam pemberian pakan, perkandangan, pemeliharaan kesehatan, penggunaan tenaga
ternak dan pemerahan serta pengolahan susu. Keterbatasan peternakan kerbau
diantaranya adalah kualitas pakan yang rendah, terbatasnya pengetahuan peternak
tentang reproduksi dan belum diterapkannya teknologi tepat guna. Beternak kerbau
secara umum merupakan usaha turun temurun sebagai tabungan. Pertimbangan
ekonomi masih jauh dari pertimbangan peternak kerbau, yang ada hanya kerbau
dipelihara dan setelah besar akan dijual. Sistem pemeliharaan kerbau secara umum
hanya dengan melepaskan kerbau di padang pengembalaan untuk memperoleh
5 Pemeliharaan dan pemberian pakan anak kerbau, kerbau dara, kerbau laktasi, kerbau
kering dan pejantan tidak dibedakan (Ibrahim, 2008).
Gen Hormon Pertumbuhan (GH)
Hormon pertumbuhan merupakan salah satu faktor yang paling penting dalam
pertumbuhan dan perkembangan sel hewan (Pierzchała et al., 2004). Pada hewan
yang sedang tumbuh, hormon pertumbuhan dapat meningkatkan efisiensi produksi,
pengurangan deposisi lemak, merangsang pertumbuhan otot, meningkatkan efisiensi
pertumbuhan pakan, meningkatkan pertumbuhan organ, dan meningkatkan
pertumbuhan tulang (Etherton dan Bauman, 1998). Hormon pertumbuhan
dibutuhkan untuk pertumbuhan jaringan dan metabolisme lemak yang berperan
penting untuk reproduksi, laktasi, dan pertumbuhan tubuh (Burton et al., 1994;
Ohlsson et al., 1998). Hubungan tersebut menjadikan hormon pertumbuhan sebagai
salah satu kandidat gen yang dapat digunakan sebagai penanda genetik dalam
program seleksi ternak.
Walis et al. (1973) menyatakan bahwa bovine growth hormone (bGH)
merupakan sebuah peptida tunggal dengan berat molekul 22 KDa. Lingappa et al.
(1977) dan Wallis et al. (1973) menyatakan bahwa bGH disusun oleh 190 atau 191
asam amino dengan panjang sekuen nukleotida 2856 pb (Gordon et al., 1983) dan
menurut Hediger et al., (1990) gen bGH terletak pada kromosom nomor 19. Gen
bGH terdiri atas 5 ekson ( Gordon et al., 1983 dan Woychick et al., 1982).
Cowan et al. (1989) dan Hilbert et al. (1989) mendeteksi situs keragaman
untuk enzim restriksi MspI, situs keragaman GH-MspI terletak pada intron 3 dari gen
hormon pertumbuhan pada posisi 1547 (Zhang et al., 1993). Dilaporkan bahwa
keragaman gen GH-MspI pada babi berpengaruh positif terhadap laju pertumbuhan
dan kualitas karkas (Pierzchała et al., 2004). Thomas et al. (2006) menyatakan
bahwa pada sapi Brangus bergenotipe +/- (heterozigot) fragmen GH-MspI memiliki
pengaruh positif terhadap pertambahan bobot badan harian dan karkas. Selain itu,
genotip +/+ dan +/- framen GH-MspI berpengaruh positif pada sifat bobot badan dan
kualitas daging (Unanian et al., 2000)Keragaman gen GH-MspI juga dilaporkan pada
berbagai ternak meliputi sapi Holstein Beijing (Zhou et al., 2005); sapi Brahman
muda (Beauchemin et al., 2006); sapi Pesisir, sapi Bali, sapi Limousin dan sapi
6 Analisis Keragaman DNA
Polimerase chain reaction (PCR) adalah suatu reaksi in vitro untuk
menggandakan molekul DNA pada target tertentu dengan cara mensintesa molekul
DNA baru yang berkomplemen dengan molekul DNA tersebut dengan enzim
polymerase dan oligonukleotida pendek sebagai primer dalam mesin thermocycler
Metode ini berjalan secara enzimatik melalui mekanisme perubahan suhu. Proses
yang terjadi dalam mesin PCR melipti tiga tahap utama yaitu denaturasi (pemisahan
untai ganda DNA), annealing (penempelan primer) dan ekstensi (pemanjangan
primer). Proses dari mulai denaturasi, penempelan dan ektensi disebut sebagai satu
siklus. Produk PCR dapat langsung divisualisasikan melalui proses elektroforesis dan
dapat digunakan untuk analisis lebih lanjut (Muladno, 2002). Keragaman DNA
amplikon atau produk PCR bisa dilakukan dengan berbagai cara, antara lain RFLP,
SSCP, DGGE dan sequensing.
Restriction Fragment Length Polymorphism (RFLP)
PCR-RFLP merupakan metode analisis lanjutan dari produk PCR. Metode
PCR memanfaatkan perbedaan pola pemotongan enzim restriksi atau enzim
pemotong yang berbeda pada tiap-tiap mikroorganisme. Analisis RFLP sering
digunakan untuk mendeteksi lokasi genetik dalam kromosom yang menyandikan
penyakit yang diturunkan (Orita et al., 1989) ataupun untuk mendeteksi adanya
keragaman pada gen yang berhubungan dengan sifat ekonomis, seperti produksi dan
kualitas susu (Sumantri et al., 2004; Sumantri et al., 2005).
Single-StrandConformationPolymorphism (SSCP)
PCR-SSCP merupakan salah satu metode analisis lebih lanjut yang
memanfaatkan produk PCR. Metode PCR-SSCP merupakan metode yang handal
dalam mendeteksi adanya mutasi secara cepat. Dalam analisis ini, sekuen mutasi
dapat dideteksi dengan perubahan pola migrasi dari bentuk ikatan utas tunggal DNA
pada gel poliakrilamida (Hayasi, 1991), yang disebut sebagai perubahan konformasi
atau bentuk molekul. Pendeteksian dalam SSCP dipengaruhi oleh matriks gel,
7
DenaturingGradient Gel Electrophoresis (DGGE)
Elektroforesis yang dilakukan menggunakan gel poliakrilamida dengan
gradien urea yang ditambah dengan formamida. Pemisahan dilakukan tanpa enzim
restriksi dan sekuen bukan berdasarkan berat molekul. Teknik ini menggunakan
dasar perbedaan stabilitas produk PCR. Dengan demikian sangat tergantung dari
jumlah ikatan hidrogen yang ada dalam DNA tersebut (Suryanto, 2008).
Analisis Sekuen (Sequencing)
Analisis sekuen merupakan suatu teknik yang dianggap paling baik untuk
melihat keanekaragaman hayati suatu kelompok organisme. Teknik ini berkembang
setelah orang menciptakan mesin DNA sequencer. Pada prinsipnya polimorfisme
dilihat dari urutan atau sekuen DNA dari fragmen tertentu dari suatu genom
organisme (Suryanto, 2008). Sequencing merupakan proses penentuan urutan
nukleotida pada suatu fragmen DNA atau RNA. Sequencing menghasilkan
penggambaran linear simbolik yang disebut sekuens yang meringkas sebagian besar
struktur tingkat atom atas molekul yang disekuensing. Sequencing DNA akan
menghasilkan sekuens DNA yang digambarkan sebagai untaian abjad lambang
8 METODE
Lokasi dan Waktu
Penelitian ini dilakukan selama empat bulan, dimulai dari bulan September
2008 – Desember 2008. Lokasi penelitian ini adalah Laboratorium Zoologi
Departemen Biologi, dan Laboratorium Pemuliaan dan Genetika Departemen Ilmu
Produksi dan Teknologi Peternakan, Institut Pertanian Bogor.
Materi
Sampel Darah dan Ekstraksi DNA
Sampel darah kerbau yang digunakan berjumlah 320 sampel yang diambil
dari empat daerah, yaitu 75 sampel dari Semarang, 103 sampel dari Mataram, 65
sampel dari Medan dan 77 sampel dari Banten. Ekstraksi DNA dilakukan
menggunakan Genomic DNA mini kit (Geneaid).
Primer
unit/µl dan buffernya, , dNTP 10 nmol/µl, air destilata dan enzim restriksi (RE) MspI
masing – masing dengan buffernya.
Alat – alat yang digunakan antara lain tabung PCR, mesin thermocycler
(TaKaRa PCR Thermal Cycler MP4), alat sentrifugasi, pipet mikro dengan tipsnya,
vortex, dan power supply 500vA.
Elektroforesis
Bahan – bahan yang digunakan adalah air destilata steril, akrilamida 30%, 5 x
TBE, TEMED (tetramethylendiamine) dan APS (ammonium persulfat) 10%, loading
9 Alat – alat yang digunakan antara lain dua lempeng kaca (20 x 16 cm) untuk cetakan
gel, gelas ukur, tabung reaksi, sisir pencetak sumur, pipet mikro 2 µl dengan tipsnya
dan power supply 500vA.
Keragaman genotip pada masing – masing individu ternak dapat ditentukan
melalui pita – pita DNA yang ditemukan. Frekuensi alel dihitung dengan rumus Nei
(1987), sebagai berikut :
xi =
Dimana :
xi = Frekuensi alel i
nii = Jumlah individu bergenotip ii
nij = Jumlah individu bergenotip ij
n = Jumlah individu sampel
Derajat heterozigositas (ĥ) dihitung berdasarkan frekuensi alel pada tiap lokus
DNA menggunakan rumus Nei (1987) :
ĥ =
Keterangan :
ĥ = Nilai heterozigositas lokus xi = Frekuensi alel i
n = Jumlah individu sampel
Ragam heterozigositas (Vsl( ĥ )) diantara individu dalam satu kesatuan
frekuensi alel populasi dapat dihitung dengan rumus sebagai berikut : 2nii + ∑nij
2n
2n (1 - ∑xi2)
10 Vsl( ĥ ) = {} { 2(2n – 2)( ∑xi3 – (∑xi2)2 ) + ∑xi2 – (∑xi2)2 }
dan SE diperoleh dari akar ragam heterozigositas (Nei, 1987).
Rataan heterozigositas (Ĥ) dihitung dengan rumus sebagai berikut :
Keterangan :
ĥj= derajat heterozigositas untuk lokus ke-j
r = jumlah lokus yang diuji
Ĥ = rataan heterozigositas
Indeks fiksasi pada masing – masing populasi diperoleh dari persamaan :
Keterangan :
Xkii = Frekuensi genotipe homozigot alel i pada populasi ke-k
Xki = Frekuensi alel i
Prosedur
Pengambilan dan Penanganan Sampel
Sampel darah diambil melalui vena jugularis menggunakan jarum dan tabung
vaccutainer berheparin.
Ekstraksi DNA dari Sampel Darah
Ekstraksi DNA dilakukan menggunakan Genomic DNA mini kit Geneaid
11 Amplifikasi DNA dengan Teknik PCR-RFLP
Proses amplifikasi DNA secara umum menggunakan metode sebagai berikut:
2 µl sampel DNA dimasukkan kedalam tabung PCR, kemudian ditambah 0,2 µl
primer. Larutan dalam tabung kemudian ditambah dengan premix yang terdiri dari
18,45 µl air destilata, 2,5 µl 10 x bufer, 2 µl 2,5 mM MgCl2, 0,2 µl dNTP 2mM, dan
0,15 µl enzim taq polymerase. Tabung kemudian diInkubasi pada mesin
thermocycler dengan suhu annealing 620C.
Produk PCR sebanyak 2 µl dipindahkan ke dalam tabung baru 0,2 ml dan
ditambahkan dengan enzim restriksi MspI dan buffernya. Campuran tersebut
diinkubasikan dalam inkubator pada suhu 37°C overnight.
Elektroforesis
Potongan DNA produk PCR dipisahkan dengan teknik elektroforesis gel
polyakrilamida 6%. Gel dibuat dengan cara mencampurkan 12 ml air destilata, 4 ml
5 x TBE, 4 ml akrilamida 30%, 15 µl TEMED, dan 160 µl APS 10%. Sebanyak 2 µl
produk PCR dilarutkan dalam loading dye. Elektroforesis dilakukan selama 40 menit
pada tagangan konstan 220 v atau sampai pewarna bromtimol blue mencapai bagian
bawah gel. Setelah elektroforesis selesai, gel diambil untuk dilakukan pewarnaan
perak.
Pewarnaan perak
Pewarnaan perak (silver staining) dilakukan dengan langkah sebagai berikut:
gel dimasukan kedalam larutan CTAB 0,2 g/200 ml air destilata selama 8 menit
sambil digoyang, kemudian dicuci dengan air destilata selama 2 x 2 menit. Air
tersebut dibuang dan ditambahkan larutan NH4OH selama 6 menit sambil digoyang.
Kemudian dilanjutkan dengan larutan AgNO3 selama 10 menit sambil digoyang.
Kemudian gel dicuci kembali dengan air destilata 2 x 2 menit. untuk memunculkan
pita, gel direndam dalam larutan Na2CO3. Setelah pita muncul, ditambahkan larutan
asam asetat.
Pendeteksian keragaman DNA
Setelah pewarnaan, akan muncul pita – pita DNA. Setiap pita DNA
dibandingkan dengan marker untuk mengetahui panjangnya. Setiap pita DNA dari
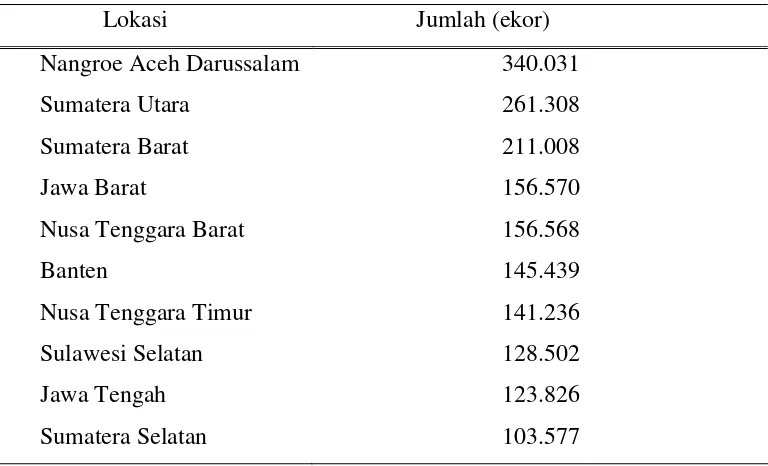
12 merupakan gambar pita-pita DNA yang menunjukkan genotipe kerbau. Kerbau
memiliki genotipe +/+ apabila terdapat fragmen terpotong di panjang 223 pb dan 104
pb. Genotipe +/- apabila terdapat fragmen gabungan yaitu terdapat 3 pita di panjang
327, 223, dan 104 pb. Genotipe -/- apabila fragmen tidak terpotong, hanya terdapat 1
pita di panjang 327 pb.
Gambar 1. Pita DNA hasil elektroforesis setelah pewarnaan perak. Fragmen hasil amplifikasi PCR ditambah dengan enzim pemotong MspI.
M +/+ +/-
-/-327 pb
223 pb
13 HASIL DAN PEMBAHASAN
Amplifikasi Gen Hormon Pertumbuhan (GH-MspI)
Gen hormon pertumbuhan pada kerbau lokal berhasil diamplifikasi dengan
menggunakan metode PCR (Polymerase Chain Reaction). Primer yang digunakan
yaitu pasangan primer berdasarkan penelitian Mitra et al. (1995) yang digunakan
pada ternak sapi. Primer ini berhasil mengamplifikasi gen target sepanjang 327 pb di
intron 3 dan ekson 4. Hasil amplifikasi disajikan pada Gambar 1.
Gambar 1. Hasil Amplifikasi Gen Hormon Pertumbuhan dengan PCR dalam Gel 6% Poliakrilamida
Fragmen gen GH-MspI intron 3 dan ekson 4 sepanjang 327 pb dapat
diketahui dengan mencocokkan pasangan primer pada sekuen gen GH pada GenBank
(nomor akses M57764). Panjang fragmen ini berbeda 2 pb dengan hasil yang
didapatkan oleh Zhou et al. (2005) yaitu 329 pb. Posisi penempelan primer dan titik
mutasi pada intron 3 pada sekuen gen GH-MspI disajikan pada Gambar 2.
Suhu annealing merupakan suhu yang memungkinkan terjadinya penempelan
primer pada sampel. Suhu annealing sangat penting dalam proses amplifikasi karena
proses pemanjangan DNA baru dimulai dari primer. Suhu annealing yang digunakan
dalam penelitian ini adalah 620C, suhu ini merupakan suhu optimal untuk
penempelan primer dan berbeda dengan suhu yang digunakan oleh Zhou et al. (2005)
dalam penelitiannya yang menggunakan suhu annealing 600C. Hal ini mungkin saja 327 pb 400 pb
300 pb
200 pb
14
1441 CCCCCACGGG CAAGAATGAG GCCCAGCAGA AATCAGTGAG TGGCAACCTC GGACCGAGGA
1501 GCAGGGGACC TCCTTCATCC TAAGTAGGCT GCCCCAGCTC CCGCAC|CGGC CTGGGGCGGC 1561 CTTCTCCCCG AGGTGGCGGA GGTTGTTGGA TGGCAGTGGA GGATGATGGT GGGCGGTGGT 1621 GGCAGGAGGT CCTCGGGCAG AGGCCGACCT TGCAGGGCTG CCCCAGACCC GCGGCACCCA 1681 CCGACCACCC ACCTGCCAGC AGGACTTGGA GCTGCTTCGC ATCTCACTGC TCCTCATCCA 1741 GTCGTGGCTT GGGCCCCTGC AGTTCCTCAG CAGAGTCTTC ACCAACAGCT TGGTGTTTGG
terjadi namun selama perbedaan suhu menghasilkan produk amplifikasi yang sama,
maka kisaran suhu tersebut dapat digunakan.
forward
reverse
Gambar 2. Posisi Penempelan Primer (cetak tebal) pada sekuen Gen
GH-MspI. Terjadi Mutasi pada Situs Pemotongan C|CGG pada Posisi 104 (C menjadi T)(Gordon et al., 1983)
Menurut Viljoen et al. (2005), keberhasilan dalam mengamplifikasi DNA
bergantung pada interaksi komponen PCR dalam kosentrasi yang tepat. Beberapa hal
yang umum dilakukan untuk optimasi PCR diantaranya adalah suhu penempelan
primer, kosentrasi Mg2+, kosentrasi primer, dan kosentrasi DNA target. Selain itu,
terdapatnya inhibitor yang dapat menghambat proses PCR harus dieliminasi
semaksimal mungkin. Inhibitor yang biasa ditemukan mencemari PCR diantaranya
adalah material organik, logam berat, lipid, hemoglobin dan urea.
Pendeteksian Keragaman Gen GH-MspI dengan PCR-RFLP
Metode restriction fragment length polymorphisms (RFLP) digunakan untuk
mengidentifikasi keragaman gen GH-MspI pada kerbau lokal. Analisis RFLP
mendeteksi keragaman dengan memanfaatkan perbedaan panjang fragmen DNA
yang dipotong oleh enzim restriksi MspI. Enzim restriksi MspI mengenali situs
pemotongan empat basa C│CGG. Cowan et al. (1989) and Hilbert et al. (1989)
mendeteksi situs keragaman untuk enzim restriksi MspI, keragaman berlokasi di
intron 3 dari gen GH pada posisi 1547 (Zhang et al., 1993). Terjadinya mutasi
transisi yang merubah basa C – T menyebabkan situs pemotongan untuk enzim
restriksi MspI berubah. Mutasi transisi yang terjadi karena adanya subtitusi antara
basa Adenin dengan Guanin (Purin) atau antara basa Sitosin dengan Timin
(Pirimidin) (Paolella, 1998).
Diketahui bahwa titik mutasi gen GH-MspI berada pada daerah intron 3,
15 akan hilang (splicing). Mutasi yang terjadi pada sekuen yang tidak ditranskripsikan
(intron) akan mengakibatkan silent mutation yaitu mutasi yang tidak mengakibatkan
perubahan fenotipik. Selain itu, silent mutation juga dapat terjadi karena 1) mutasi
yang terjadi tidak menyebabkan perubahan asam amino karena beberapa asam amino
yang sama dikodekan oleh kodon yang berbeda, 2) mutasi tidak terjadi pada situs
aktif protein (Paolella, 1998). Namun Funk (2001) menyatakan bahwa tingkat
polimorfisme dan mutasi yang tinggi pada daerah non-coding diduga dapat
mempengaruhi ekspresi gen secara tidak langsung. Mutasi pada intron dapat
mengganggu proses spilicing, seperti tidak terpotongnya intron atau timbulnya splice
site baru, hal ini yang akan menyebabkan terganggunya pengaturan ekspresi gen.
Sebagai contoh, menurut Gunadi (2006) hampir 50% penyakit genetik pada manusia
diakibatkan oleh mutasi yang mengganggu splicing dan salah satu contoh
penyakitnya adalah ß-thalasemia.
Terdeteksinya keragaman gen GH-MspI karena ditemukan 2 alel + dan -
dengan genotipe yaitu +/+, +/-, dan -/-. Genotipe homozigot (+/+ dan -/-) berarti
bahwa kedua tetua masing-masing menyumbangkan gen (alel) yang sama dan
genotipe heterozigot (+/-) merupakan kombinasi gen yang berbeda dari kedua tetua
atau kromosom yang berpasangan. Ketiga jenis genotipe dapat dilihat pada Gambar
3.
16 Keragaman Gen GH-MspI Kerbau Lokal
Nei (1987) menyatakan bahwa suatu alel dikatakan polimorfik jika memiliki
frekuensi alel sama dengan atau kurang dari 0,99(99%) atau dengan kata lain suatu
alel dikatakan monomorfik jika frekuensi alel sama dengan atau kurang dari
0,01(1%). Frekuensi alel yaitu frekuensi relatif dari suatu alel dalam populasi atau
jumlah suatu alel terhadap jumlah total alel dalam suatu populasi (Nei dan Kumar,
2000). Selain dengan mengetahui frekuensi alel, keragaman genetik juga dapat
diukur secara akurat dengan nilai heterozigositas (Nei, 1987). Pendugaan nilai
heterozigositas memiliki arti penting untuk diketahui karena dapat memberikan
informasi mengenai tingkat polimorfisme suatu alel, serta prospek populasi dimasa
yang akan datang (Falconer dan Macay, 1996).
Berdasarkan hasil gen GH-MspI pada kerbau lokal bersifat polimorfik.
Sejumlah 261 sampel dari 320 sampel berhasil diidentifikasi keragaman gen
GH-MspI dengan metode PCR-RFLP yang berarti persentase keberhasilannya sebesar
81,56%. Dari total sampel, kemunculan alel + sebesar 0,878 dan alel – sebesar 0,122.
Frekuensi genotipe +/+ sebesar 0,805, genotipe +/- sebesar 0,146 dan genotipe -/-
sebesar 0,05. Frekuensi genotipe dan frekuensi alel gen GH-MspI secara lengkap
disajikan pada Tabel 2.
Tabel 2. Hasil Perhitungan Frekuensi Genotipe dan Frekuensi Alel Gen
17 populasi Medan (0,731). Frekuensi genotipe +/- tertinggi terdapat di populasi Medan
(0,269) dan terendah di populasi Banten (0,048), sedangkan frekuensi genotipe -/-
tertinggi terdapat di populasi Semarang (0,173) dan terendah terdapat di populasi
Medan dan Banten (tidak ditemukan genotipe -/-).
Alel + juga memiliki frekuensi yang paling tinggi di setiap populasi, dengan
frekuensi tertinggi terdapat pada populasi Banten (0,976) dan terendah pada populasi
Semarang (0,788). Sebaliknya untuk alel -, frekuensi yang tertinggi terdapat pada
populasi Semarang (0,212) dan terendah pada populasi Banten (0,024).
Berdasarkan hasil perhitungan ini dapat diartikan bahwa kerbau lokal
Indonesia berpotensi untuk dijadikan ternak pedaging karena berdasarkan beberapa
penelitian menyebutkan bahwa genotip +/+ dan +/- framen GH-MspI berpengaruh
positif pada sifat bobot badan dan kualitas daging (Unanian et al., 2000).
Nilai Heterozigositas
Nilai heterozigositas dari masing-masing populasi berkisar 0,093 – 0,562.
Populasi Semarang memiliki nilai heterozigositas tertinggi dan populasi Banten
terendah. Nilai heterozigositas total adalah 21,5 % dan nilai rataan heterozigositas
dari seluruh populasi adalah 37%. Dapat dikatakan bahwa nilai heterozigositas di
keseluruhan populasi rendah, karena menurut Javanmard et al., (2005) nilai
heterozigositas di bawah 0,5 (50%) mengindikasikan rendahnya variasi suatu gen
dalam populasi.
Tabel 3. Nilai heterozigositas (ĥ) dan Rataan Heterozigositas (Ĥ) Gen
GH-MspIKerbau Lokal
Berdasarkan penelitian Primasari (2009), nilai heterozigositas dari gen
GHRH pada populasi kerbau lokal yang sama didapatkan hasil nilai heterozigositas
18 diperoleh pada penelitian ini yang berarti pelaksanaan usaha peternakan tradisional
ternyata sejalan dengan upaya seleksi gen GH.
Indeks Fiksasi
Indeks fiksasi dapat digunakan untuk mengetahui pola perkawinan dan
seleksi yang dilakukan dalam populasi. Indeks fiksasi bisa bernilai positif maupun
negatif, hal tersebut dipengaruhi oleh adanya inbreeding, seleksi dan kawin tidak
acak. Nilai indeks fiksasi gen GH-MspI kerbau lokal dari empat populasi di
Indonesia disajikan pada Tabel 5.
Tabel 5. Nilai Indeks Fiksasi Gen GH-MspI Kerbau Lokal
Daerah Alel FISki
Semarang + 0,7725
- 0,7665
Mataram + 0,3062
- 0,2195
Medan + -0,1475
- -0,1561
Banten + -0,0245
- -0,0246
Nilai indeks fiksasi tertinggi adalah 0,7725 (alel +) yaitu pada populasi
kerbau lokal Semarang. Sedangkan nilai indeks fiksasi terendah terdapat pada
populasi Medan yaitu -0,1561 (alel -). Nilai indeks fiksasi menyebar dan tidak
bernilai nol, hal ini menandakan bahwa tidak terjadinya fiksasi gen GH atau tidak
19 KESIMPULAN DAN SARAN
Kesimpulan
Gen GH-MspI pada kerbau lokal bersifat polimorfik. Penelitian menemukan
2 tipe alel yaitu alel + (0,878) dan alel – (0,122) serta genotipe yaitu +/+ (0,805),
genotipe +/- (0,146) dan genotipe -/- (0,05). Tipe alel + dan genotipe +/+ memiliki
frekeunsi kemunculan yang paling besar pada tiap populasi. Tingginya frekuensi
genotip +/+ menunjukan bahwa kerbau lokal diduga berpotensi untuk menjadi ternak
pedaging karena genotip ini berpengaruh positif terhadap pertumbuhan bobot badan
dan kualitas daging. Nilai heterozigositas total sebesar 21,5% dengan nilai terendah
pada populasi Banten (9,3%) dan tertinggi pada populasi Semarang (56,2%). Nilai
indeks fiksasi menyebar dan tidak bernilai nol yang berarti gen GH tidak terfiksasi
tau masih terjadinya kawin acak.
Saran
Diperlukan penelitian lanjutan untuk mengetahui hubungan antara keragaman
gen GH-MspI dengan sifat-sifat kuantitatif sehingga selanjutnya dapat dijadikan
20 UCAPAN TERIMAKASIH
Puji dan syukur penulis panjatkan kepada Allah SWT atas limpahan rahmat
dan karunia-Nya sehingga penulis dapat menyelesaikan penulisan skripsi ini. Pada
kesempatan ini pula penulis ingin mengucapkan terima kasih yang sebesar-besarnya
kepada :
1. Mama dan Alm. Bapak tercinta yang telah memberikan kasih sayang, nasehat,
bimbingan dan pengorbanan yang begitu besar, kedua adik tersayang, Aditya
Prabowo dan Fahmi Ramadhan serta beserta seluruh keluarga lainnya yang telah
membantu hingga penulis dapat menyelesaikan studi di Fakultas Peternakan.
2. Dr. Ir. Cece Sumantri, M.Agr.Sc dan Dr. Ir. Achmad Farajallah, M.Si sebagai
dosen pembimbing atas segala bimbingan, perhatian, motivasi dan arahan bagi
penulis dalam penelitian dan skripsi ini.
3. Dr. Ir. Heni Nuraeni, MSi dan Ir. Lilis Khotijah, MS sebagai dosen penguji
sidang yang telah banyak memberikan saran dalam penyelesaian tugas akhir ini.
4. Terima kasih Penulis ucapkan kepada teman-teman satu tim (Mira, Ires dan
Wiwin), Grup Zoo (Pak Khoirul, Bu Bibah, Bu Ria, Kak Wildan, Kak Ogi, Mba
Muli) dan Kak Eryk Andreas atas bimbingannya.
5. Sahabat-sahabat tercinta “3G” (Asty, Tristy, Thata, Mpit dan Nenk) atas semua
kebersamaan dan keceriaan selama berada Fakultas Peternakan, persahabatan
yang telah terjalin semoga tak akan pernah terlupakan
6. Personil maupun alumni Alcatraz (Aan, Othel, Windy, Trimi, Mbo, Asty,
Mamah, Baqi, Mpus, Vina, Yuni dan Putri) yang telah menjadi kelurga selama
di IPB.
7. Seluruh teman-teman di IPTP 42 yang tidak bisa disebutkan satu per satu.
8. Kak Vamy yang telah memberikan semangat serta kasih sayangnya selama ini.
Semoga skripsi ini dapat bermanfaat bagi dunia peternakan Indonesia. Amin.
Bogor, April 2009
21 DAFTAR PUSTAKA
Asoen, N. J. F. 2008. Studi craniometrics dan pendugaan jarak genetik kerbau sungai, rawa dan silangannya di Sumatera Utara. Skripsi. Fakultas Peternakan. Institut Pertanian Bogor, Bogor.
Bahri, S dan C. Thalib. 2007. Strategi pengembangan pembibitan ternak kerbau. Prosiding. Seminar dan Lokakarya Nasional Usaha Ternak Kerbau 2008. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Balai Pengkajian Teknologi Peternakan. 2002. Dalam : Lendhanie, U. U. 2005. Karakteristik reproduksi kerbau rawa dalam kondisi lingkungan peternakan rakyat. J. of Bioscientiae vol. 2 (1):43-48.
Beauchemin, V. R., M. G. Thomas, D. E. Franked dan G. A. Silver. 2006. Evaluation of DNA polymorphisms involving growth hormone relative to growth and carcass characteristics in Brahman steers. Genet. Mol. Res. 5:438-447.
Bhattacharya. 1993. Dalam : W.G.A. Williamson dan W. J. A. Payne (Editor). Pengantar Peternakan di Daerah Tropis. Gadjah Mada University Press. Yogyakarta.
Burton, J. L., B. W. Mc Bride, E. Block dan D. R. Glimm. 1994. A review bovine growth hormone. Can. J. Anim. Sci. 74 : 167-201
Cowan, C. M., M. R. Dentine, R. L. Ax dan L. A. Schuler. 1989. Restriction fragment length polymorphism associated with growth hormone and prolactin genes in Holstein bulls: evidence for a novel growth hormone allele. Anim. Genet. 20 157–165
Direktorat Jenderal Peternakan. 2006. Statistik Peternakan 2006. CV Arena Seni, Jakarta.
Diwyanto, K. dan H. Handiwirawan. 2006. Strategi pengembangan ternak kerbau: aspek penjaringan dan distribusi. Prosiding. Lokakarya Nasional Usaha Ternak Kerbau Mendukung Program Kecukupan Daging Sapi. 2006. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Erdiansyah, E. 2008. Studi keragaman fenotipe dan pendugaan jarak genetik antara kerbau lokal di Kabupaten Dompu NTB. Skripsi. Fakultas Peternakan. Institut Pertanian Bogor, Bogor.
Etherton, T.D. dan D.E. Bauman. 1998. Biology of somatotropin in growth and lactation of domestic animals. Physical Rev., 78: 745-61.
22 Falconer, D. S. Dan T. F. C. Mackay. 1996. Introduction to Quantitative Genetics.
Fourth Ed. Longman Inc., New York.
Funk, D. 2001. Genetics technologies in the 1992. http//www.inform.umd.edu. [02-03-2009].
Gunadi. 2006. Antisense oligonukleotide : potensial terapi penyakit genetik akibat gangguan splicing. http//www.inovasionline.com. [20-04-2009].
Gordon, D. F., D. P. Quick, C. R. Ewin, J. E. Donelson dan R. A. Maurer. 1983. Nucleotide sequence of the bovine growth hormone chromosomal gene. Mol. Cell. Endocrinol.33:81–95
Hasinah, H. dan E. Handiwirawan. 2006. Keragaman genetik ternak kerbau di Indonesia. Prosiding. Lokakarya Nasional Usaha Ternak Kerbau Mendukung Program Kecukupan Daging Sapi 2006. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Hayashi, K. 1991. PCR-SSCP: A Simple and Sensitive Method for Detection of Mutations in the Genomic DNA. PCR Methods Appl. 1:34-38.
Hediger, R, S. E. Johnson, W. Barendse, R. D. Drinkwater, S. S. Moore dan J. Hetzel. 1990. Assignment of the growth hormone gene locus to 19q26-qter in cattle and to 11q25-qter in sheep by in situ hybridization. Genomics 8:171– 174.
Hilbert, P., A. Marcotte, A. Schwers, R. Hanset, G. Vassart dan M. Georgens. 1989. Analysis of genetic variation in the Belgian Blue cattle breed using DNA sequence polymorphism at the growth hormone, low density lipoprotein receptor, a- subunit of glycoprotein hormones and thyroglobin loci. Anim. Genet.20383–394.
Ibrahim, L. 2008. Produksi susu, reproduksi dan manajemen kerbau perah di Sumatera Barat. J. Peternakan vol. 5 (1) : 1-9.
Jakaria. 2008. Keragaman genetik gen hormon pertumbuhan pada sapi Pesisir Sumatera Barat. Disertasi. Fakultas Peternakan. Institut Pertanian Bogor, Bogor.
Javanmard, A., N. Asadzadeh, M. H. Banabazi dan J. Tavakolian. 2005. The allele and genotype frequencies of bovine pituitary specific transcription factor and leptin genes in Iranian cattle and buffalo populations using PCR-RFLP. Iranian journal of biotechnology vol. 39(2).
23 Mitra, A., P. Schlee, C. R. Balakrishnan dan F. Pirchner. 1995. Polymorphisms at growth hormone and prolactin loci in Indian cattle and buffalo. J. Anim. Breed. Genet. 112: 71–74.
Muladno. 2002. Seputar Teknologi Rekayasa Genetik. Pustaka Wira Usaha Muda. Bogor.
Nataraj, A. J., I. O. Glander, N. Kusukawa dan W. E. Highsmith Jr. 1999. Single-Strand Conformation Polymorphism and Heteroduplex Analysis for Gel-Based Mutation Detection. Electrophoresis 20:1177-1185.
Nei, M. 1987. Molecular Evalutionery Genetics. Columbia University Press. New York.
Nei, M and S. Kumar. 2000. Molecular Evolution and Phylogenetics. Oxford University Press, Inc., New York.
Ohlsson, C., B. A. Bengtsson, O. G. P. Isaksson, T. T. Andreassen dan M. C. Slootweg. 1998. Growth hormone and bone. Endocr. Rev. 19 : 55-79.
Orita, M., H. Iwahana, H. Kanazawa, K. Hayashi, dan T. Sekiya. 1989. Detection of polymorphisms of human DNA by gel electrophoresis as single-strand conformation polymorphisms. Proc. Natl. Acad. Sci. 86:2766-2770.
Paolella, P. 1998. Introduction to Molecular Biology. Mc Graw Hill Companies, Inc., Boston, Massachusetts.
Pierzchala, M., B. Tadeusz dan K. Jolanta. 2004. Growth rate and carcass quality in relation to GH/MspI and GH/HaeII PCR-RFLP polymorhism in pigs. Animal Science Papers and Reports vol. 22(1):57-64.
Praharani, L dan E. Triwulanningsih. 2007. Karakterisasi bibit kerbau pada agroekosistem dataran tinggi. Prosiding. Seminar dan lokakarya nasional usaha ternak kerbau. 2008. Pusat Penelitian dan Pengembangan Peternakan, Bogor.
Primasari, A. 2009. Identifikasi keragaman gen growth hormone-releasing hormone (GHRH) pada kerbau lokal (Bubalus bubalis) dengan metode PCR-RFLP. Skripsi. Fakultas Peternakan, Institut Pertanian Bogor, Bogor.
Pusat Penelitian dan Pengembangan Peternakan. 2006. Studi karakterisasi kerbau sungai, kerbau lumpur dan persilangannya di Sumatera Utara, Jawa Tengah dan Jawa Timur. Badan penelitian dan pengembangan peternakan Departemen Pertanian, Bogor. http://www.deptan.com. [02-03-2009]
24 Sumantri, C., A. Anggraeni, R. R. A. Maheswari, K. Dwiyanto, dan A. Farajallah. 2005. Pengaruh genotipe kappa-kasein terhadap kualitas susu pada sapi perah FH di BPTU Baturaden. Prosiding. Seminar Nasional Teknologi Peternakan dan Veteriner 2005.
Suryanto, D. 2008. Melihat keanekaragaman organism melalui beberapa teknik genetika molekuler. Program Studi Biologi. Fakultas Matematika dan Ilmu Pengetahuan Alam. Universitas Sumatera Utara. http://library.usu.ac.id. [04-03-2009]
Thomas, M. G., G. A. Silver dan R. M. Enns. 2006. Relationships of DNA polymorphisms in growth hormone (GH) to growth and carcass traits observed in a population of Brangus bulls with a larger number of sires. Int. Plant and Animal Genome XIV: P526 (Abstract).
Unanian, M. M., C. C. Borreto, A. R. de Freitas dan C. M. T. Cordeiro. Association between GH gene polymorphism and weight traits in Nellore Novines. Rev. Bras zootec. 29 : 1380-1386.
Viljoen, G. J., L., H. Nel, dan J. R. Crowther. 2005. Molecular Diagnostic PCR Handbook. Springer, Dordrecht, Netherland.
Wallis, M. 1973. The primary structure of bovine growth hormone; FEBS Lett. 35: 11–14
Woychick, R. P., S. A. Camper, R. H. Lyons, S. Horowtts, E. C. Goodwin dan F. M. Rottman. 1982. Cloning and nucleotide sequencing of the bovine growth hormone gene. Nucleic Acids Res. 10:7197–7210.
Zhang, H. M., K. C. Maddock, D. R. Brown, S. K. Denise dan R. L. Ax. 1993. A novel allele of the bovine somatotropin gene detected by PCR-RFLP analysis. J. Anim. Sci.712276.
26 Lampiran 1. Modifikasi Metode Isolasi DNA Menggunakan Genomic DNA Mini
Kit (Geneaid)
Sampel darah
↓
Sentrifugasi 3500 rpm, 10 menit Sel darah putih dipindahkan ke tabung 1,5 ml
↓ Pindahkan GD Column ke tabung 1,5 ml
↓
+ Bufer pengelusi 100 μl
↓
Sentrifugasi 10000 rpm, 1 menit
27 Lampiran 2. Sekuen Gen GH-MspI yang diakses di GenBank (no. M57764)
LOCUS BOVGHGH 2856 bp DNA linear MAM 27-APR-1993 DEFINITION Bovine growth hormone gene, complete cds.
ACCESSION M57764 M28453 VERSION M57764.1 GI:163091 KEYWORDS growth hormone. SOURCE Bos taurus (cattle) ORGANISM Bos taurus
Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Laurasiatheria; Cetartiodactyla; Ruminantia; Pecora; Bovidae; Bovinae; Bos.
REFERENCE 1 (bases 1 to 2856)
AUTHORS Gordon,D.F., Quick,D.P., Erwin,C.R., Donelson,J.E. and Maurer,R.A. TITLE Nucleotide sequence of the bovine growth hormone chromosomal gene JOURNAL Mol. Cell. Endocrinol. 33 (1), 81-95 (1983)
PUBMED 6357899
28
1 gtactggggt gggttgcctt tctcttctcc aggggattta tctgacccag ggattgaacc 61 tgagtctcct gcatttgcag ctagattctt tacggctgag ccacctggga agcccattcg 121 cttctgctgc tgctgctgct gctaagttgc ttcagtcgtg tccgacctgt gcgacgccat 181 agacagcagc ccaccaggtc cccgtccctg ggattctcca ggcaagaaca ttggagtggg 241 ttgccatttc ctcctccaat gcatgaaagt gaaaagtgaa agtgaagtca ctcagttgtg 301 tccgaccctc agcgacccca tggactgcag ccttccagaa tggggtgcca ttgccttctc 361 ctcgcttctg ctacctcccc tttaaaaaga aaacctatgg ggtgggctct caagctgaga 421 ccctgtgtgc acagccctct ggctggtggc agtggagacg ggatgatgac aagcctgggg 481 gacatgaccc cagagaagga acgggaacag gatgagtgag aggaggttct aaattatcca 541 ttagcacagg ctgccagtgg tccttgcata aatgtataga gcacacaggt ggggggaaag 601 ggagagagag aagaagccag ggtataaaaa tggcccagca gggaccaatt ccaggatccc 661 aggacccagt tcaccagacg actcagggtc ctgtggacag ctcaccagct atgatggctg 721 caggtaagct cgctaaaatc ccctccattc gcgtgtccta aaggggtaat gcggggggcc 781 ctgccgatgg atgtgttcag agctttgggc tttagggctt ccgaatgtga acataggtat 841 ctacacccag acatttggcc aagtttgaaa tgttctcagt ccctggaggg aagggtaggt 901 ggggctggca ggagatcagg cgtctagctc cctggggccc tccgtcgcgg ccctcctggt 961 ctctccctag gcccccggac ctccctgctc ctggctttcg ccctgctctg cctgccctgg 1021 actcaggtgg tgggcgcctt cccagccatg tccttgtccg gcctgtttgc caacgctgtg 1081 ctccgggctc agcacctgca tcagctggct gctgacacct tcaaagagtt tgtaagctcc 1141 cgagggatgc gtcctagggg tggggaggca ggaaggggtg aatccacacc ccctccacac 1201 agtgggagga aactgaggag ttcagccgta ttttatccaa gtagggatgt ggttagggga 1261 gcagaaacgg gggtgtgtgg ggtggggagg gttccgaata aggcggggag gggaaccgcg 1321 caccagctta gacctgggtg ggtgtgttct tcccccagga gcgcacctac atcccggagg 1381 gacagagata ctccatccag aacacccagg ttgccttctg cttctctgaa accatcccgg 1441 cccccacggg caagaatgag gcccagcaga aatcagtgag tggcaacctc ggaccgagga 1501 gcaggggacc tccttcatcc taagtaggct gccccagctc ccgcaccggc ctggggcggc 1561 cttctccccg aggtggcgga ggttgttgga tggcagtgga ggatgatggt gggcggtggt 1621 ggcaggaggt cctcgggcag aggccgacct tgcagggctg ccccagaccc gcggcaccca 1681 ccgaccaccc acctgccagc aggacttgga gctgcttcgc atctcactgc tcctcatcca 1741 gtcgtggctt gggcccctgc agttcctcag cagagtcttc accaacagct tggtgtttgg 1801 cacctcggac cgtgtctatg agaagctgaa ggacctggag gaaggcatcc tggccctgat 1861 gcgggtgggg atggcgttgt gggtcccttc catgtggggg ccatgcccgc cctctcctgg 1921 cttagccagg agaatgcacg tgggcttggg gagacagatc cctgctctct ccctctttct 1981 agcagtccag ccttgaccca ggggaaacct tttccccttt tgaaacctcc ttcctcgccc 2041 ttctccaagc ctgtagggga gggtggaaaa tggagcgggc aggagggagc tgctcctgag 2101 ggcccttcgg cctctctgtc tctccctccc ttggcaggag ctggaagatg gcaccccccg 2161 ggctgggcag atcctcaagc agacctatga caaatttgac acaaacatgc gcagtgacga 2221 cgcgctgctc aagaactacg gtctgctctc ctgcttccgg aaggacctgc ataagacgga 2281 gacgtacctg agggtcatga agtgccgccg cttcggggag gccagctgtg ccttctagtt 2341 gccagccatc tgttgtttgc ccctcccccg tgccttcctt gaccctggaa ggtgccactc 2401 ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca ttgtctgagt aggtgtcatt 2461 ctattctggg gggtggggtg gggcaggaca gcaaggggga ggattgggaa gacaatagca 2521 ggcatgctgg ggatgcggtg ggctctatgg gtacccaggt gctgaagaat tgacccggtt 2581 cctcctgggc cagaaagaag caggcacatc cccttctctg tgacacaccc tgtccacgcc 2641 cctggttctt agttccagcc ccactcatag gacactcata gctcaggagg gctccgcctt 2701 caatcccacc cgctaaagta cttggagcgg tctctccctc cctcatcagc ccaccaaacc 2761 aaacctagcc tccaagagtg ggaagaaatt aaagcaagat aggctattaa gtgcagaggg 2821 agagaaaatg cctccaacat gtgaggaagt aatgag