INHIBISI SENYAWA BAHAN ALAM PADA PROTEIN
ANTIAPOPTOSIS SEL KANKER PANKREAS
MELALUI PENDEKATAN
IN SILICO
MUHAMAD SHOLEHUDDIN MALIK IBROHIM
DEPARTEMEN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Inhibisi Senyawa Bahan Alam pada Protein Antiapoptosis Sel Kanker Pankreas melalui Pendekatan In Silico adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
MUHAMAD SHOLEHUDDIN MALIK IBROHIM. Inhibisi Senyawa Bahan Alam sebagai Inhibitor Protein Antiapoptosis pada Sel Kanker Pankreas Melalui Pendekatan In Silico. Dibimbing oleh ARMI WULANAWATI dan HARRY NOVIARDI.
Kanker pankreas merupakan jenis tumor ganas yang sangat mematikan. Penderita kanker pankreas umumnya ditangani dengan kemoradioterapi dan obat gemsitabin, namun masih belum memberikan hasil klinis yang baik. Penelitian ini bertujuan menentukan potensi senyawa bahan alam sebagai inhibitor protein antiapoptosis sel kanker pankreas berdasarkan energi bebas Gibbs (∆G), konstanta inhibisi, interaksi hidrogen, dan toksisitas senyawa bahan alam secara in silico. Jumlah senyawa bahan alam yang berpotensi menjadi obat kanker sangat banyak, sehingga penelitian di laboratorium secara satu-per-satu terhadap senyawa bahan alam tersebut sangat tidak efisien dari segi waktu dan biaya. Oleh karena itu, untuk mengefisienkan hal tersebut digunakan metode komputasi. Hasil dari inhibisi beberapa senyawa bahan alam dengan standar gemsitabin terhadap protein antiapoptosis menyatakan bahwa brusein D merupakan senyawa yang sangat berpotensi menjadi obat kanker pankreas ditinjau dari segi energi bebas Gibbs, jumlah interaksi hidrogen, dan toksisitasnya.
Kata kunci: in silico, kanker pankreas, protein antiapoptoss, senyawa bahan alam
ABSTRACT
MUHAMAD SHOLEHUDDIN MALIK IBROHIM. Inhibition of Natural Products as Inhibitor of Antiapoptotic Protein in Pancreatic Cancer by In Silico Approach. Dibimbing oleh ARMI WULANAWATI dan HARRY NOVIARDI.
Pancreatic cancer is a deadliest cancer. Patients with pancreatic cancer are usually treated by chemoradiotherapy and gemsitabin, but the treatment does not always give good clinical outcomes. This study is aimed to determine the potential of natural products as inhibitors of antiapoptotic pancreatic cancer protein cells based on the Gibbs free energy (ΔG), inhibition constants, hydrogen interactions, and toxicity of natural products by in silico method. Natural products that are potential to become drugs are numerous. Thus, laboratory experiments on each compound would be inefficient in terms of time and costs. Therefore, for efficiency computational method is offered. The results of the inhibition of some natural products usng gemsitabin as a standard against antiapoptosis proteins is brusein D that is the most potential compound to become a pancreatic cancer drug in terms of Gibbs free energy, the amount of hydrogen interactions, and toxicity.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Kimia
INHIBISI SENYAWA BAHAN ALAM PADA PROTEIN
ANTIAPOPTOSIS SEL KANKER PANKREAS
MELALUI PENDEKATAN
IN SILICO
DEPARTEMEN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul Skripsi : Inhibisi Senyawa Bahan Alam pada Protein Antiapoptosis Sel Kanker Pankreas melalui Pendekatan In Silico
Nama : Muhamad Sholehuddin Malik Ibrohim NIM : G44100104
Disetujui oleh
Armi Wulanawati, SSi, MSi
Pembimbing I Harry Noviardi, SSi, MSi Pembimbing II
Diketahui oleh
Prof Dr Dra Purwantiningsih Sugita, MS Ketua Departemen
PRAKATA
Puji syukur penulis ucapkan kepada Allah SWT atas limpahan rahmat dan karunia-Nya sehingga penulis dapat menyelesaikan karya ilmiah dengan judul Inhibisi Senyawa Bahan Alam pada Protein Antiapoptosis Sel Kanker Pankreas melalui Pendekatan In Silico.
Terima kasih penulis ucapkan kepada Ibu Armi Wulanawati, SSi, MSi dan Bapak Harry Noviardi, SSi, MSi selaku pembimbing atas ilmu, arahan dan bimbingannya kepada penulis selama melaksanakan penelitian ini. Penulis juga mengucapkan terima kasih kepada Ayah, Mama, Siti, Nene, Bibi dan seluruh keluarga atas dorongan semangat, doa dan kasih sayangnya. Ucapan terima kasih juga penulis ucapkan kepada Lona Mahdriani Puspita, Muhamad Adam Nurfalah, serta teman-teman AKAPELA yang selalu memberi semangat dan masukan.
Penulis berharap karya ilmiah ini dapat bermanfaat bagi perkembangan ilmu pengetahuan.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
PENDAHULUAN 1
BAHAN DAN METODE 2
Alat dan Bahan 2
Metode 2
HASIL DAN PEMBAHASAN 4
Tapak aktif Protein 5
Interaksi Protein-Ligan 7
Toksisitas Ligan berdasarkan LD50 14
SIMPULAN DAN SARAN 16
Simpulan 16
Saran 16
DAFTAR PUSTAKA 16
LAMPIRAN 19
DAFTAR TABEL
1 Grid tapak aktif berbagai jenis protein antiapoptosis 7 2 Nilai ∆G terbaik dari interaksi Hsp90 dan Epigalokatekin-3-galat 9 3 Residu dan jarak interaksi hidrogen MAPK-brusein D secara
fleksibel dan kaku 13
4 Tingkat toksisitas ligan berdasarkan nilai LD50 15
DAFTAR GAMBAR
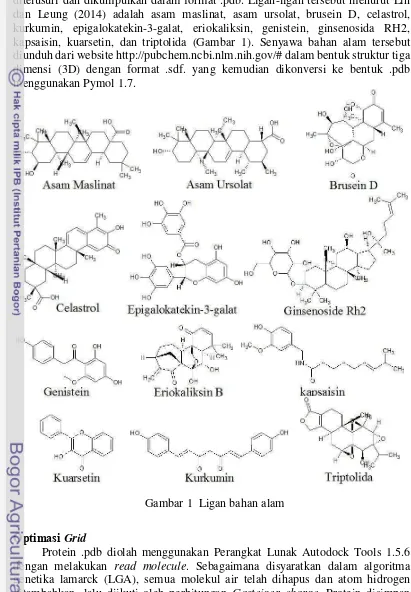
1 Ligan bahan alam 3
2 Gemsitabin 5
3 Grid protein Hsp27 x, y, z berturut-turut merah, hijau, dan biru. 6 4 Interaksi Hsp27 dengan ligan brusein D 6
5 Grid tapak aktif protein Hsp90 8
6 Interaksi hidrogen Hsp90 dengan Epigalokatekin-3-galat 9 7 Nilai ΔG interaksi protein-ligan dan jumlah interaksi hidrogen
secara fleksibel 10
8 Ikatan σ gemsitabin yang dapat berotasi 11 9 Nilai ΔG interaksi protein-ligan dan jumlah interaksi hidrogen
secara kaku 11
10 Perbandingan nilai ΔG dan jumlah interaksi hidrogen pada
docking ligan brusein D terhadap protein MAPK 12 11 Interaksi hidrogen protein MAPK dengan ligan brusein D yang
di-docking secara fleksibel 12
12 Interaksi hidrogen protein MAPK dengan ligan brusein D yang
di-docking secara kaku 13
13 Tanaman Brucea javanica (L.) Merr. 14
DAFTAR LAMPIRAN
1 Diagram alir penelitian 19
PENDAHULUAN
Kanker pankreas merupakan salah satu tumor ganas paling mematikan di dunia. Kasus kanker pankreas terus meningkat dalam 2 dekade terakhir dengan tingkat morbiditas dan mortalitas penderita yang sangat tinggi (Nakamura et al. 2013). Penderita kanker pankreas yang dapat bertahan hidup selama 5 tahun berjumlah kurang dari 5% (Zhao et al. 2014).
Kanker pankreas terbagi menjadi dua tingkatan, yaitu kanker yang hanya terdapat pada pankreas (lokal) dan kanker yang telah menjalar ke bagian lainnya (metastasis). Kanker pankreas lokal memiliki 3 tingkatan, yaitu stadium I, II, dan III. Penderita kanker pankreas dalam tingkatan tersebut biasanya mendapatkan penanganan penyembuhan mulai dari kemoterapi hingga radioterapi (Wolfgang et al. 2013). Operasi juga memungkinkan untuk dilakukan, walaupun tingkat kesembuhannya masih rendah, karena sekitar 80% penderita sudah dalam keadaan stadium III atau metastasis (Lin dan Leung 2014). Sedikitnya gejala awal tertentu menyebabkan penyakit ini kebanyakan ditemukan setelah sel kanker pankreas bermetastasis (Oettle 2014). Pada keadaan metastasis, sel kanker telah menyebar dari pusat tumor utama, lalu berpindah mengikuti sirkulasi aliran darah dan tumbuh di jaringan lain terutama lambung dan hati (Chiang dan Massagua 2008). Penderita yang sudah mencapai tingkatan ini biasanya diberi obat gemsitabin dengan dosis yang tinggi, walaupun jumlah penderita yang dapat bertahan hidup dalam 1 tahun dengan penggunaan obat ini masih kurang dari 20% (Wolfgang et al. 2013).
Gemsitabin merupakan analog dari pirimidina yang bekerja merusak susunan dari asam nukleat sehingga menghambat proses pembelahan sel. Penghambatan tersebut mendorong sel untuk mengaktifkan mekanisme apoptosis atau sel melakukan bunuh diri (Cerqueira et al. 2007). Namun, dalam dosis yang tinggi, obat ini dapat menyebabkan keracunan pada hati dan ginjal, anemia, dan trombositopenia (Moysan et al. 2013). Oleh karena itu, telah banyak diteliti obat kanker yang berbasis bahan alam. Obat kanker yang banyak dikembangkan saat ini berasal dari bahan alam yang digabung dengan kemoterapi dan radioterapi konvensional (Lin dan Leung 2014).
Bahan alam biasanya dikemas menjadi obat-obatan herbal dan sebagai sebagai obat pelengkap dan obat alternatif. Senyawa bahan alam yang banyak diteliti dan telah diketahui memiliki aktivitas antikanker pada sel kanker payudara dan prostat antara lain kurkumin, kapsaisin, triptolida dan celastrol, asam ursolik dan asam maslinat, ginsenosida, brusein D, dan eriokaliksin B. Senyawa bahan alam tersebut sebelumnya (Lin dan Leung 2014). Senyawa bahan alam tersebut berperan sebagai ligan yang mengganggu inisiasi dan perkembangan sel kanker melalui berbagai mekanisme seperti proliferasi, diferensiasi, apoptosis, dan metastasis (Pratheeshkumar et al. 2012).
2
(Yan et al. 2012). Beberapa protein antiapoptosis yang akan dihambat pada penelitian ini adalah K-Ras, keluarga protein NFκB, MAPK, JAK/STAT, Hsps, Bcl, COX-2, Nrf-2, SHH, dan Wnt/β-katenin (Lin dan Leung 2014). Analisis penghambatan menggunakan metode komputasi atau in silico yang merupakan metode docking atau penambatan ligan pada suatu protein. Metode ini lazim digunakan dalam tahap pencarian suatu kandidat molekul obat, karena metode ini sangat cepat, ekonomis, andal, dan ramah lingkungan (Ördög dan Grolmusz 2008). Penelitian ini bertujuan menentukan potensi senyawa bahan alam sebagai inhibitor protein antiapoptosis sel kanker pankreas berdasarkan energi bebas Gibbs (∆G), tetapan inhibisi, interaksi hidrogen, dan toksisitas senyawa bahan alam secara in silico menggunakan perangkat lunak Autodock Tools 1.5.6, Autogrid 4.2, Autodock 4.2, Pymol 1.7, dan Toxicity Estimation Software Tool (TEST) 4.1. Senyawa bahan alam yang terbaik dapat diteliti lebih lanjut secara in vivo dan in vitro, untuk menggantikan obat kanker kovensional gemsitabin yang memiliki hasil klinis yang rendah serta beberapa efek samping.
BAHAN DAN METODE
Alat dan Bahan
Alat-alat yang digunakan adalah seperangkat komputer dengan chip processor AMD A10-6800K quadcore 4.1GHz, random access memory (RAM) 8 gigabit, dan video graphics array AMD Radeon HD 8670D, ditunjang dengan akses internet 4G LTE menggunakan modem ZTE mf825a untuk mengunduh data-data protein dan substrat.
Bahan-bahan yang digunakan pada penelitian ini berupa perangkat lunak Autodock Tools 1.5.6, Autogrid 4.2, Autodock 4.2, Pymol 1.7, dan Toxicity Estimation Software Tool (TEST) 4.1.
Metode Lingkup Kerja
Penelitian ini terdiri atas beberapa tahapan. Bagan alir penelitian dapat dilihat pada Lampiran 1.
Preparasi Fail Protein dan Ligan
3 (Lampiran 2). Semua residu seperti air atau ligan lain yang tidak diinginkan dihilangkan menggunakan perangkat lunak Pymol 1.7.
Ligan yang akan digunakan untuk menginhibisi protein antiapoptosis juga ditelusuri dan dikumpulkan dalam format .pdb. Ligan-ligan tersebut menurut Lin dan Leung (2014) adalah asam maslinat, asam ursolat, brusein D, celastrol, kurkumin, epigalokatekin-3-galat, eriokaliksin, genistein, ginsenosida RH2, kapsaisin, kuarsetin, dan triptolida (Gambar 1). Senyawa bahan alam tersebut diunduh dari website http://pubchem.ncbi.nlm.nih.gov/# dalam bentuk struktur tiga dimensi (3D) dengan format .sdf. yang kemudian dikonversi ke bentuk .pdb menggunakan Pymol 1.7.
Optimasi Grid
Protein .pdb diolah menggunakan Perangkat Lunak Autodock Tools 1.5.6 dengan melakukan read molecule. Sebagaimana disyaratkan dalam algoritma genetika lamarck (LGA), semua molekul air telah dihapus dan atom hidrogen ditambahkan, lalu diikuti oleh perhitungan Gasteiger charge. Protein disimpan
4
dengan ekstensi .pdbqt. Kemudian ligan dibuka dengan input ligand, diatur agar sehingga memiliki torsi yang bebas dan disimpan juga dengan ekstensi .pdbqt.
Ukuran grid ditentukan pada setiap protein dengan nilai sumbu x, y, dan z yang berbeda-beda. Jarak antargrid juga ditetapkan berbeda-beda pada setiap protein dengan satuan angstrom (Å). Grid dibuat sebesar mungkin sampai tepat menutupi seluruh permukaan protein. Fail grid disimpan dalam bentuk grid.gpf. Proses optimasi merupakan proses docking untuk menentukan tapak interaksi yang paling disukai oleh setiap ligan. Tapak tersebut kemungkinan adalah tapak aktif dari protein.
Docking
Autogrid merupakan bagian dari proses docking untuk menentukan area interaksi antara protein dan ligan. Grid yang digunakan merupakan hasil dari proses optimasi sebelumnya. Setelah ukuran grid disesuaikan dengan grid optimum, disimpan dalam bentuk grid.gpf. Setelah itu, Autogrid 4.2 diolah menggunakan terminal ubuntu pada direktori fail tersebut. Setelah autogrid selesai, akan muncul beberapa fail yang akan digunakan saat docking. Setelah itu, parameter docking disesuaikan dengan menggunakan pilihan algoritma genetik dan disimpan dalam bentuk docking.dpf. Docking dilakukan pada terminal Ubuntu menggunakan Autodock 4.2 sehingga menghasilkan fail docking.dlg. Fail docking.dlg dianalisis menggunakan Autodock Tools 1.5.6 menggunakan menu analyze untuk mengetahui nilai ∆G. Fail tersebut kemudian disimpan dalam bentuk kompleks untuk divisualisasikan menggunakan Pymol 1.7.
Visualisasi Fail Docking
Fail keluaran docking dengan ekstensi .pdbqt divisualisasikan menggunakan Pymol 1.7 untuk melihat tapak pengikatan, jumlah interaksi hidrogen, dan jarak antaratom yang berikatan.
Uji Toksisitas Ligan
Uji toksisitas ligan yang merupakan senyawa bahan alam, dilakukan menggunakan perangkat lunak TEST 4.1. yang dapat membaca fail dengan format .sdf atau .mol. Setelah fail ligan atau senyawa bahan alam dimasukkan, kemudian dihitung menggunakan metode oral rat untuk mengetahui nilai LD50.
HASIL DAN PEMBAHASAN
5 yang baik dan tanpa efek samping.
Jumlah senyawa ligan bahan alam yang berpotensi sebagai obat kanker sangat banyak. Pencarian kandidat obat menggunakan metode konvensional di laboratorium akan sangat lama dan tidak efisien. Oleh karena itu digunakan metode komputasi atau in silico untuk mengoptimalkan pencarian tersebut. Metode yang digunakan tergantung pada informasi struktural yang tersedia untuk protein target dan ligan dari hasil kristalografi sinar-X, tapak aktif protein dan ligan, jenis pengikatan ligan, serta interaksi yang mungkin terjadi antara protein dan ligan. Hal ini dapat dilakukan dengan cara docking menggunakan media elektronik/komputer (Madsen et al. 2010). Simulasi docking ini melibatkan beberapa protein antiapoptosis sel kanker pankreas serta ligan-ligan yang berasal dari senyawa bahan alam. Protein dan ligan tersebut diinteraksikan untuk menentukan senyawa bahan alam terbaik yang dapat menginhibisi protein antiapoptosis berdasarkan beberapa parameter seperti energi bebas Gibbs (∆G), tetapan inhibisi, dan interaksi hidrogen. Interaksi kemudian dibandingkan dengan hasil docking antara protein antiapoptosis dan standar gemsitabin (Gambar 2). Namun sebelum docking dilakukan, perlu ditentukan tapak aktif yang akan diinhibisi atau daerah interaksi docking.
Gambar 2 Gemsitabin
Tapak aktif Protein
Pertumbuhan sel kanker disebabkan protein-protein yang meregulasi kelangsungan hidup sel mengalami malafungsi sehingga sel tersebut terus tumbuh secara abnormal. Protein tersebut harus diinaktivasi dengan cara inhibisi melalui pada tapak aktifnya. Dalam hal ini, inhibisi protein akan dilakukan secara simulasi docking menggunakan perangkat lunak Autodock Tools 4.2. Digunakan ligan-ligan dari senyawa bahan alam dengan gemsitabin sebagai standar. Sebelum dilakukan docking, terlebih dahulu ditentukan daerah pada protein yang akan diinhibisi atau grid.
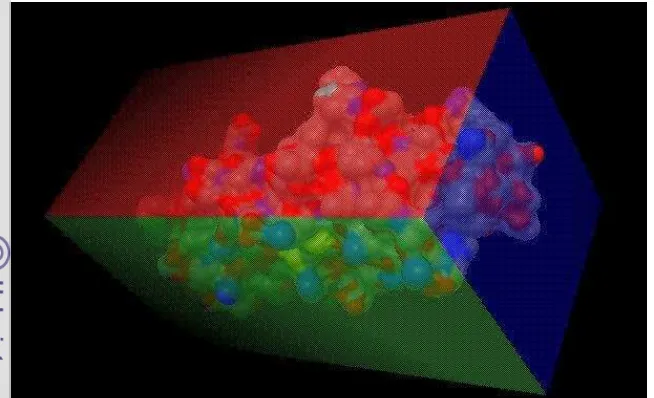
6
Gambar 3 Grid protein Hsp27 x, y, z berturut-turut merah, hijau, dan biru. Protein Hsp27 diinteraksikan dengan ligan brusein D menggunakan parameter LGA (Morris et al. 2009b). Protein dan ligan diinteraksikan melalui docking menggunakan perintah “Autogrid4 -p grid.gpf -l grid.glg” dan “Autodock4 -p docking.dpf -l docking.glg” melalui Terminal Ubuntu. Setelah docking selesai, fail docking.glg dianalisis menggunakan Autodock Tools untuk mengetahui letak tapak aktif protein. Total konformasi terbaik yang ditampilkan oleh Autodock Tools adalah 10 interaksi ligan dengan 7 di antaranya menempel pada bagian tengah protein Hsp27 yang berlubang (Gambar 4).
Gambar 4 Interaksi Hsp27 dengan ligan brusein D
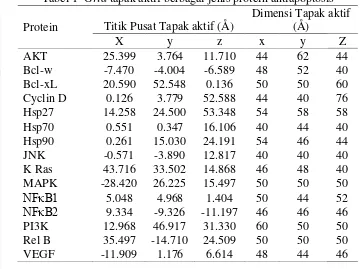
7 aktif seluruh protein (Tabel 1) yang kemudian digunakan untuk docking secara fleksibel dan kaku.
Tabel 1 Grid tapak aktif berbagai jenis protein antiapoptosis Protein Titik Pusat Tapak aktif (Å)
Dimensi Tapak aktif
Docking atau penambatan protein-ligan dilakukan untuk mencari interaksi terbaik antara protein-ligan dengan parameter tertentu seperti energi bebas Gibbs (∆G), tetapan inhibisi, dan interaksi hidrogen. Docking dilakukan 2 macam perlakuan terhadap ligan, yaitu kaku dan fleksibel. Perlakuan ini bertujuan untuk membandingkan interaksi yang lebih baik antara ligan kaku dan fleksibel. Docking secara kaku berbasis pada teori enzim kunci dan gembok yang pertama kali dikemukakan pada tahun 1894 oleh Emil Fischer. Teori ini dikemukakan oleh Koshland (1994) menganalogikan enzim sebagai gembok dan substrat sebagai kuncinya. Substrat akan menginduksi enzim tepat pada tapak aktif untuk mencapai reaksi. Docking secara fleksibel berbasis pada teori enzim Induced Fit, teori ini mengasumsikan bahwa substrat berperan dalam menentukan bentuk akhir dari enzim dan sebagian dari enzim tersebut bersifat fleksibel. Hal ini menjelaskan mengenai senyawa tertentu dapat mengikat enzim, tetapi tidak bereaksi karena enzim tersebut terdistorsi. Senyawa tersebut mungkin teralu kecil untuk menginduksi enzim, sehingga enzim hanya dapat bereaksi dengan substrat yang tepat.
Interaksi Protein-Ligan secara Fleksibel
8
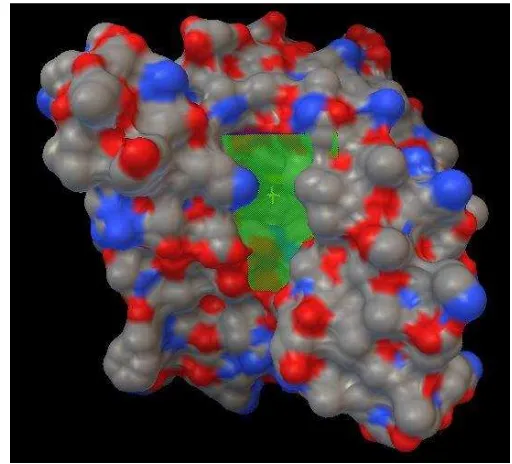
terbuka (McMurry 2012). Pada dasarnya prosedur docking protein-ligan secara fleksibel sama dengan saat menentukan tapak aktif protein, hanya saja menggunakan grid yang lebih spesifik dan terarah pada tapak aktif protein. Pada protein Hsp90 yang di-docking oleh ligan Epigalokatekin-3-galat, menggunakan grid dengan titik pusat x, y, z 0.261; 15.030; 24.191 serta dimensi x, y, z 54, 46, 44 (Gambar 5). Fail protein diolah menggunakan Autodock Tools, kemudian protein dibersihkan dari molekul-molekul air dan dilakukan penambahan hidrogen. Penambahan atom hidrogen dilakukan karena terdapat kemungkinan hilangnya atom hidrogen pada saat kristalisasi yang dapat memengaruhi interaksi (Zusapa 2013).
Gambar 5 Grid tapak aktif protein Hsp90
Setelah grid terbentuk, dilakukan autogrid menggunakan Autogrid 4.2 dan docking menggunakan Autodock 4.2 melalui Terminal Ubuntu. Kemudian akan diperoleh 10 nilai ∆G paling rendah atau negatif. Kriteria ∆G ˂ 0 pada suhu dan tekanan konstan, memiliki arti bahwa reaksi kimia tersebut bersifat spontan dengan penurunan energi berbas Gibbs. Jika pada hasil reaksi G menurun, maka reaksi memiliki kecenderungan spontan untuk mengkonversi reaktan menjadi produk. (Atkins 2006).
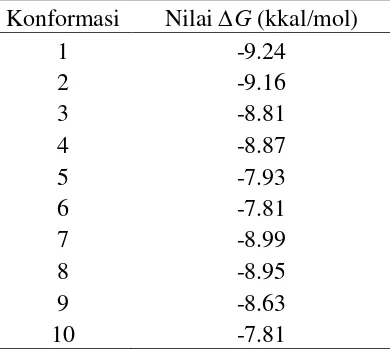
9 Tabel 2 Nilai ∆G terbaik dari interaksi Hsp90 dan Epigalokatekin-3-galat
Konformasi Nilai ∆G (kkal/mol)
1 -9.24
2 -9.16
3 -8.81
4 -8.87
5 -7.93
6 -7.81
7 -8.99
8 -8.95
9 -8.63
10 -7.81
Konformasi docking dengan nilai ∆G terbaik adalah konformasi yang memiliki ∆G sebesar -9.24 kkal/mol. Berdasarkan nilai ∆G tersebut menggunakan persamaan Gibbs maka akan diperoleh nilai tetapan inhibisi (Ki).
∆G = nRTlnKi
(Morris et al. 1998a) Nilai dari tetapan inhibisi konformasi tersebut adalah 0.17. Konformasi yang dihasilkan kemudian divisualisasi menggunakan Pymol untuk mengetahui jumlah interaksi hidrogennya. Jumlah interaksi hidrogen pada konformasi ini sejumlah 9 interaksi (Gambar 6).
Gambar 6 Interaksi hidrogen Hsp90 dengan Epigalokatekin-3-galat
10
jumlah interaksi hidrogen dirangkum dan dibandingkan untuk mencari interaksi yang lebih baik dari standar (Lampiran 3).
Nilai ΔG dan tetapan inhibisi standar gemsitabin terbaik terdapat pada docking protein Hsp90 berturut-turut, yaitu sebesar -5.97 kkal/mol dan Kisebesar
41.81 (Lampiran 3), sedangkan interaksi hidrogen terbanyak terdapat pada docking
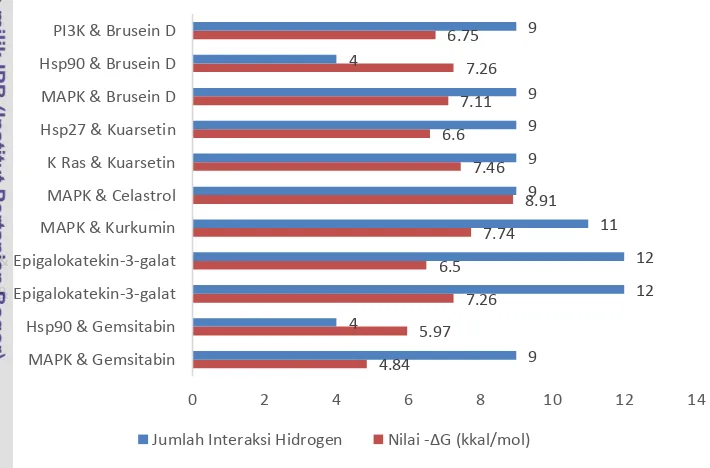
MAPK dengan jumlah interaksi hidrogen sebanyak 9 interaksi. Ligan yang berpotensi untuk inhibisi protein sel kanker pankreas, harus memiliki nilai ΔG dan tetapan inhibisi yang lebih kecil serta memiliki interaksi hidrogen lebih besar dari standar. Dari seluruh hasil docking hanya ada 8 interaksi protein-ligan yang memenuhi sebagai interaksi yang lebih baik dari standar (Gambar 7). Terdapat lima jenis ligan berbeda, yaitu brusein D, kuarsetin, celastrol, kurkumin, dan epigalokatekin-3-galat.
Gambar 7 Nilai ΔG interaksi protein-ligan dan jumlah interaksi hidrogen secara fleksibel
Interaksi Protein-Ligan secara Kaku
Docking secara kaku pada umumnya sama dengan docking secara fleksibel, perbedaannya hanya terdapat pada ikatan σ ligan. Pada docking secara kaku, ikatan σ dibuat menjadi kaku sehingga tidak dapat berotasi. Seperti pada ligan gemsitabin, memiliki lima ikatan σ yang dapat berotasi (Gambar 8) kemudian ikatan σ tersebut diatur sehingga tidak dapat berotasi.
4.84
11
Gambar 8 Ikatan σ gemsitabin yang dapat berotasi
Setelah protein dan ligan disiapkan, kemudian dilakukan autogrid dan docking dengan metode yang sama dengan cara fleksibel (Lampiran 4). Nilai ΔG dan tetapan inhibisi standar gemsitabin terbaik terdapat pada docking protein K Ras, yaitu sebesar -5.06 kkal/mol dan Kisebesar 194.42 (Lampiran 4), dengan jumlah
interaksi hidrogen terbanyak adalah 11 (Gambar 9). Dari seluruh interaksi docking hanya ada interaksi yang memiliki nilai ΔG dan tetapan inhibisi yang lebih kecil dari standar, tetapi tidak ada yang memiliki interaksi hidrogen lebih besar atau sama dengan 11 interaksi. Ada beberapa interaksi yang memiliki nilai ΔG dan tetapan inhibisi yang lebih baik dari standar, dengan jumlah interaksi hidrogen yang cukup banyak antara lain. Namun interaksi antara protein K Ras dengan gemsitabin merupakan interaksi yang memiliki interaksi hidrogen paling banyak.
Gambar 9 Nilai ΔG interaksi protein-ligan dan jumlah interaksi hidrogen secara kaku
Jika dibandingkan antara docking secara kaku dan fleksibel, maka docking secara fleksibel memberikan lebih banyak kemungkinan interaksi, karena pada docking secara kaku, ikatan antara karbon-karbon sp3 dibuat kaku sehingga tidak
dapat berputar. Ligan-ligan yang masuk ke dalam lubang protein membutuhkan penyesuaian dengan mengubah konformasi ligan agar ligan bisa menginhibisi dengan baik. Senyawa ligan bahan alam memiliki struktur yang cukup besar, rerata terdiri lebih dari 20 karbon sehingga cukup sulit bagi ligan tersebut untuk masuk ke dalam protein jika tidak melakukan penyesuaian. Kesulitan tersebut membuat interaksi protein-ligan menjadi kurang spontan dibandingkan dengan docking secara fleksibel, terlihat dari nilai ΔG yang menurun pada hampir seluruh interaksi protein-ligan secara kaku. Jumlah interaksi hidrogen juga mengalami penurunan
5.06
12
akibat sulitnya ligan berinteraksi dengan protein. Hal tersebut juga terjadi pada interaksi MAPK-brusein D yang mengalami penurunan nilai ΔG dan interaksi hidrogen (Gambar 10).
Gambar 10 Perbandingan nilai ΔG dan jumlah interaksi hidrogen pada docking ligan brusein D terhadap protein MAPK
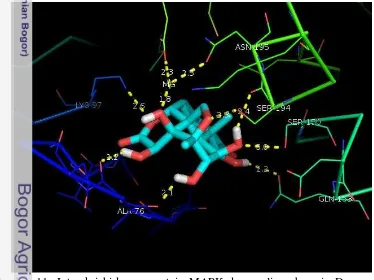
Interaksi antara protein MAPK dengan brusein D yang di-docking secara fleksibel (Gambar 11) dan kaku (Gambar 12) divisualisasikan menggunakan perangkat lunak Pymol.
Gambar 11 Interaksi hidrogen protein MAPK dengan ligan brusein D yang di-docking secara fleksibel
7.11
9
6.76 7
-ΔG (Kkal/mol) ∑Interaksi Hidrogen
fleksibel
13
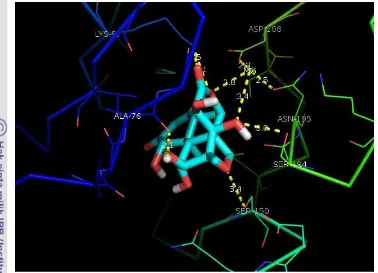
Gambar 12 Interaksi hidrogen protein MAPK dengan ligan brusein D yang di-docking secara kaku
Hasil interaksi di atas diinterpretasikan lebih jelas ke dalam Tabel 3 untuk menentukan residu protein MAPK yang berinteraksi hidrogen dengan brusein D dan jarak interaksinya. Pada docking MAPK-brusein D secara fleksibel berinteraksi terhadap 9 residu protein dengan jarak interaksi yang lebih pendek, sedangkan docking MAPK-brusein D secara kaku berinteraksi dengan 7 residu dengan jarak interaksi yang lebih jauh. Hal tersebut menujukan bahwa interaksi secara fleksibel lebih stabil dibandingkan dengan interaksi secara kaku.
Tabel 3 Residu dan jarak interaksi hidrogen MAPK-brusein D secara fleksibel dan kaku
Residu Fleksibel (Å) Kaku (Å)
ALA-76 2.1 2.1
ASN-195 2.5 2.5
ASP-208 2.3 2.3
GLN-153 2.2 -
GLY-77 2.2 -
LYS-97 2.6 2.5
LYS-97 - 3.1
SER-150 3.0 3.4
SER-194 2.3 3.0
14
Hasil-hasil di atas menunjukan brusein D merupakan ligan yang paling potensial dibandingkan ligan-ligan lainnya yang diujikan dalam penelitian ini. Dari segi strukturnya, brusein D juga memiliki banyak gugus fungsi yang dapat berinteraksi hidrogen dengan protein sehingga interkasi brusein D dengan protein akan semakin stabil. Brusein D berpotensi besar untuk menggantikan gemsitabin dalam pengobatan kanker terutama kanker pankreas. Berbeda dari gemsitabin yang didistribusikan secara transfusi intravena kepada penderita, menurut aturan Lipinski of five (1997) brusein D dapat didistribusikan secara oral pada penderita. Aturan yang telah terpenuhi antara lain, brusein D memiliki hidrogen donor tidak lebih dari 5 atom dan brusein D memiliki hidrogen donor tepat 5 atom, bobot molekul kurang dari 500 g/mol, yaitu 410.15 g/mol, nilai koefisien partisi oktanol-air kurang dari 5, yaitu -0.9220, jumlah akseptor hidrogen tidak lebih dari 10, yaitu terdapat 9 akseptor hidrogen.
Brucea javanica (L.) Merr. atau biasa dikenal dengan nama buah makasar (Gambar 13) merupakan tumbuhan obat yang biasa digunakan sebagai antimalaria, antipiretik, dan efek homeostatis (Wagih et al. 2008). Kandungan utama dari buah ini adalah brusein D yang telah diketahui memiliki aktifitas antiproliferasi pada sel kanker. Pada penelitian sebelumnya diketahui bahwa ekstrak etanol dari Brucea javanica (L.) Merr.atau lebih dikenal dengan nama buah makasar dapat menghambat proliferasi sel adenokarsinoma pankreas manusia dan menyebabkan apoptosis secara in vitro (Lau et al. 2008b). Brusein D menghambat protein p38 atau dikenal juga sebagai MAPK dan mengaktifkan jalur sinyal apoptosis pada sel kanker pankreas (Lau et al. 2009a). Interaksi docking dengan protein MAPK secara fleksibel dan kaku memiliki nilai ΔG yang tidak berbeda jauh dan lebih baik dari standar gemsitabin.
Gambar 13 Tanaman Brucea javanica (L.) Merr.
Toksisitas Ligan berdasarkan LD50
15 pelepasan protein proapoptosis mitokondria yang dikenal sebagai caspases. Caspases berada di bawah kendali protein Bcl-2 atau ekspresi dari protein proapoptosis pada sel kanker, kemudian interaksi dari protein Bcl-2 dengan ligannya mengaktifkan sinyal apoptosis (Narang dan Desai 2009). Ligan-ligan pada docking juga diuji toksisitasnya menggunakan perangkat lunak TEST atau Toxicity Estimation Software Tool versi 4.1 dengan metode Oral Rat LD50.
Oral Rat LD50 adalah dosis zat kimia yang dapat membunuh separuh populasi
hewan (tikus). Tes ini adalah cara yang paling umum untuk menentukan toksisitas suatu zat kimia. Menurut Hodge dan Sterner yang diacu pada Lancaster (2002) toksisitas dibagi menjadi beberapa kelas berdasarkan dosis LD50 (Lampiran 5).
Metode ini juga mengacu pada metode konsensus Quantitative Structure Activity Relationship atau QSAR yang saat ini digunakan secara luas dalam berbagai disiplin ilmu. Penggunaan metode ini ditujukan untuk mengurangi uji toksisitas secara in vivo sekaligus menggurangi penggunaan bahan kimia (Hewitt et al. 2007). Fail ligan dimasukan dengan format .sdf atau .mol, InChl, SMILES, nomor CAS, dan ID molekul dari zat yang akan diujikan. Hasil dari 12 senyawa bahan alam dan 1 standar yang diujikan. Berikut Tabel 4 adalah nilai toksisitas LD50 setelah
pengujian menggunakan TEST.
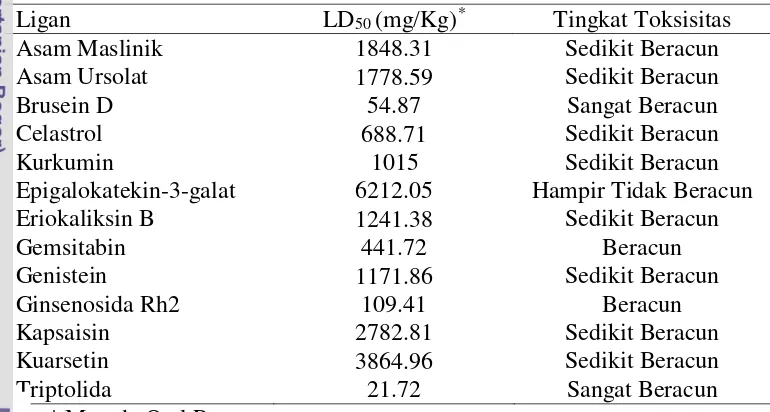
Tabel 4 Tingkat toksisitas ligan berdasarkan nilai LD50
Ligan LD50 (mg/Kg)* Tingkat Toksisitas
Asam Maslinik 1848.31 Sedikit Beracun
Asam Ursolat 1778.59 Sedikit Beracun
Brusein D 54.87 Sangat Beracun
Celastrol 688.71 Sedikit Beracun
Kurkumin 1015 Sedikit Beracun
Epigalokatekin-3-galat 6212.05 Hampir Tidak Beracun
Eriokaliksin B 1241.38 Sedikit Beracun
Gemsitabin 441.72 Beracun
Genistein 1171.86 Sedikit Beracun
Ginsenosida Rh2 109.41 Beracun
Kapsaisin 2782.81 Sedikit Beracun
Kuarsetin 3864.96 Sedikit Beracun
Triptolida 21.72 Sangat Beracun
* Metode Oral Rat
Brusein D merupakan ligan yang memiliki toksisitas yang sangat beracun berdasarkan nilai LD50 sebesar 54.87 mg/Kg, sehingga memiliki potensi yang
16
SIMPULAN DAN SARAN
Simpulan
Senyawa bahan alam brusein D sangat berpotensi menginhibisi protein antiapoptosis mitogen-activated protein kinases (MAPK) yang berfungsi sebagai pengatur proliferasi, diferensiasi, transformasi, dan kelangsungan hidup sel kanker pankreas. Interaksi antara MAPK dengan brusein D merupakan interaksi yang lebih baik berdasarkan nilai energi bebas Gibbs (ΔG), tetapan inhibisi, interaksi hidrogen,
serta toksisitas dibandingkan dengan interaksi dari standar gemsitabin secara in silico.
Saran
Perlu dilakukan pengujian lebih lanjut terhadap senyawa brusein D secara in vivo dan in vitro terhadap sel kanker pankreas.
DAFTAR PUSTAKA
Atkins P, Paula DJ. 2006. Physical Chemistry 8th Edition. New York: W. H.
Freeman and Company.
Attwood TK, Cammack R, Campbell PN, Parish JH, Smith AD, Stirling JL, Vella F. 2006 Oxford Dictionary of Biochemistry and Molecular Biology. Oxford:
Oxford University Press.
Berman HM, Westbrook J, Feng Z, Gilliland G, Bhat TN, Weissig H, Shindyalov IN, Bourne PE. 2000. The protein data bank. Nucleic Acid Research.
28(1):235-242. doi:10.1093/nar/28.1.235.
Bolton EE, Wang Y, Thiessen PA, Bryant SH. 2008. Pubchem: integrated platform of small molecules and biological activities. An Rep Comp Chem. 4:217-241. doi:10.1016/S1574-1400(08)00012-1.
Cerqueira NMFSA, Fernandes PA, Ramos MJ. 2007. Understanding ribonucleotide reductase inactivation by gemcitabine. Chem Eur J. 13:8507-8515. doi:10.1002/chem.200700260.
Chiang AC, Massagua J. 2008. Molecular basis of metastasis. The New England J Med. 359:2814-2823. doi:10.1056/NEJMra0805239.
Hewitt M, Cronin M, Madden JC, Rowe PH, Johnsen C, Obi A, Enoch SJ. 2007. Consensus QSAR models: do the benefits outweigh the complexity?. J. Chem. Inf. Model. 47:1460-1468. doi:10.1021/ci700016d.
Koshland DE. 1994. The key-lock theory and the induced fit theory. Angew Chem Int Ed Engl. 33:2375-2378. doi: 10.1002/anie.199423751.
Kuno T, Tsukamoto T, Hara A, Tanaka T. 2012. Cancer chemoprevention trough the introduction of apoptosis by natural compound. Journal of Biophysical Chemistry. 2:156-173. doi:10.4236/jbpc.2012.32018.
17 Lau ST, Lin ZX, Liao Y, Zhao M, Cheng HK, Leung PS. 2009a. Brucein D induces apoptosis in pancreatic adenocarcinoma cell line PANC-1 through the activation of p38-mitogen activated protein kinase. Cancer Letters. 281:42– 52. doi:10.1016/j.canlet.2009.02.017.
Lau ST, Lin ZX, Zhao M, Leung PS. 2008b. Brucea javanica fruit induces cytotoxicity and apoptosis in pancreatic adenocarcinoma cell lines. Phytother. Res. 22:477-486. doi:10.1002/ptr.2344.
Lin L, Leung PS. 2014. Use herbal medicines and natural product: an alternative approach to overcoming the apoptotic resistance of pancreatic cancer. Int J Biochem & Cell Bio. 53:224-236. doi:10.1016/j.biocel.2014.05.021.
Lipinski CA, Lombardo F, Dominy BW, Feeney PJ. 1997. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Advanced Drug Delivery Reviews. 23:3-25. Doi: 10.1016/S0169-409X(96)00423-1.
Madsen U, Povl KL, Tommy L. 2010. Textbook of Drug Design and Discovery Edisi ke-4. Washington DC (US): Taylor & Francis.
Martin TM. 2008. ToxPredictor: a Toxicity Estimation Software Tool. Washington: EPA Science Forum.
McMurry J. 2012. Organic Chemistry 8thEdition. Brooks/Cole: Cengage Learning.
Morris GM, Goodsell DS, Halliday RS, Huey R, Hart WE, Belew RK, Olson AJ. 1998a. Automated docking using a lamarckian genetic algorithm and an empirical binding free energy function. J Comput Chem. 19(14):1639-1662. doi:10.1002/(SICI)1096987X(19981115)19:14<1639::AIDJCC10>3.0.CO;2 -B
Morris GM, Huey R, Lindstrom W, Sanner MF, Belew RK, Goodsell DS, Olson AJ. 2009b. AutoDock4 and AutoDockTools4: automated docking with selective receptor flexibility. J Comput Chem. 30(16):2785-2791. doi:10.1002/jcc.21256.
Moysan E, Bastiat G, Benoit JP. 2013. Gemcitabine versus modified gemcitabine: A review of several promising chemical modifications. J Am Chem Soc. 10:430-444. doi:10.1021/mp300370t.
Nakamura T, Masuda K, Harada S, Akioka K, Sako H. 2013. Pancreatic cancer: Slow progression in the early stages. Int J Surgery Case Rep. 4:693-696. doi:10.1016/j.ijscr.2013.04.040.
Narang AS dan Desai DS. 2009. Pharmaceutical Perspectives of Cancer Therapeutics. New York: Springer.
Oettle H. 2014. Progress in the knowledge and treatment of advanced pancreatic cancer: from benchside to bedside. Cancer Treatment Rev. 40:1039-1047. doi:10.1016/j.ctrv.2014.07.003.
Oliveira SHP, Ferraz FAN, Honorato RV, Xavier-Neto J, Sobreira TJP, Oliveira PSL. 2014. KVFinder: steered identification of protein cavities as a PyMOL plugin. BMC Bioinformatics. 15:197-204. doi:10.1186/1471-2105-15-197. Ördög R dan Grolmusz V. 2008. Evaluating genetic algorithms in protein-ligand
docking. Lecture Notes in Comp Sci. 4983:402-413. doi:10.1007/978-3-540-79450-9_37.
18
natural products: mechanisms of action and molecular targets. Anti-Cancer Agents in Med Chem. 12:1-26. doi:10.2174/187152012803833035.
Wagih ME, Alam G, Wiryowidagdo, Attia K. 2008. Improved production of the indole alkaloid canthin-6-one from cell suspension culture of Brucea javanica(L.) Merr. Indian Journal of Science and Technology. 1(7):1-6. doi:10.17485/ijst/2008/v1i7/29591
Wolfgang CL, Herman JM, Laheri DA, Klein AP, Erdek MA, Fishman EK, Hruban RH. 2013. Recent progress in pancreatic cancer. CA Cancer J Clin. 63:318-348. doi:10.3322/caac.21190.
Yan C, Siegel D, Newsome J, Chiloux A, Moody CJ, Ross D. 2012. Antitumor indolequinones induced apoptosis in human pancreatic cancer cells via inhibition of thioredoxin reductase and activation of redox signaling.
Molecular Pharmacology. 81(3):401-410. doi:10.1124/mol.111.076091.
Zhao G, Deng S, Zhu S, Wang B, Li X, Liu Y, Qin Q, Gong Q, Niu Y, Xiang C et al. 2014. Chronic pancreatitis and pancreatic cancer demonstrate active
epithelial-mesenchymal transition profile, regulated by miR-217-SIRT1 pathway. Cancer Lett. 355:184-191. doi:10.1016/j.canlet.2014.08.007. Zusapa GA. 2013. Studi komponen aktif temu lawak terhadap patogenesis kanker
19
Menentukan protein yang akan diinhibisi dan ligan penginhibisi
Pengumpulan data protein dan ligan
Autodock 4.2
Molecular grid dan molecular docking
Kanker Pankreas
Uji Toksisitas Ligan
Energi bebas Gibbs (∆G)
Tetapan Inhibisi (Ki)
Visualisasi
20
21 Lampiran 3 Hasil docking protein-ligan secara fleksibel
Protein Ligan ΔG
Epigalokatekin-3-galat -8.58 0.51 9
Eriokaliksin B -7.77 2.00 3
Epigalokatekin-3-galat -6.63 13.72 5
Eriokaliksin B -6.62 13.95 2
Epigalokatekin-3-galat -9.28 0.16 7
22
Epigalokatekin-3-galat -4.68 369.37 4
Eriokaliksin B -5.23 145.90 0
Epigalokatekin-3-galat -6.76 11.01 9
Eriokaliksin B -8.28 0.85 2
Epigalokatekin-3-galat -6.50 17.08 12
23
Epigalokatekin-3-galat -9.24 0.17 9
Eriokaliksin B -6.29 24.36 0
Epigalokatekin-3-galat -7.09 6.31 8
Eriokaliksin B -7.44 3.49 2
Epigalokatekin-3-galat -7.26 4.73 12
24
Epigalokatekin-3-galat -8.26 0.87 9
Eriokaliksin B -7.34 4.14 2
Epigalokatekin-3-galat -4.22 803.24 8
Eriokaliksin B -5.78 57.63 3
Epigalokatekin-3-galat -4.90 254.74 9
25
Epigalokatekin-3-galat -8.43 0.66 10
Eriokaliksin B -8.26 0.87 6
Epigalokatekin-3-galat -6.22 27.41 7
Eriokaliksin B -7.17 5.51 3
Epigalokatekin-3-galat -5.61 76.80 7
26
Lampiran 4 Hasil docking protein-ligan secara kaku
Protein Ligan ΔG
Epigalokatekin-3-galat -4.50 500.58 9
Eriokaliksin B -6.76 11.01 3
Epigalokatekin-3-galat -2.60 1.24x104 1
Eriokaliksin B -6.37 21.28 1
Epigalokatekin-3-galat -3.38 3318.49 4
27
Epigalokatekin-3-galat -2.98 6521.10 3
Eriokaliksin B -5.57 82.16 2
Epigalokatekin-3-galat -4.58 437.32 7
Eriokaliksin B -8.01 1.33 3
Epigalokatekin-3-galat -2.07 3.03x104 5
Eriokaliksin B -5.81 54.78 4
Gemsitabin -2.71 1.03x104 5
Genistein -4.91 250.47 4
Ginsenosida Rh2 62.04 3.19x1051 2
Kapsaisin -3.13 5061.78 1
Kuarsetin -4.61 415.72 8
28
Epigalokatekin-3-galat 0.72 3.37x106 4
Eriokaliksin B -6.39 20.57 2
Gemsitabin -4.36 634.10 4
Genistein -7.23 4.98 3
Ginsenosida Rh2 64.00 8.72x1052 1
Kapsaisin -4.62 408.76 0
Epigalokatekin-3-galat -0.71 3.01x105 4
Eriokaliksin B -7.46 3.38 2
Gemsitabin -3.99 1184.50 3
Genistein -5.58 80.79 2
Ginsenosida Rh2 101.12 1.47x1080 2
Kapsaisin -3.86 1475.32 1
Epigalokatekin-3-galat -4.98 222.55 8
Eriokaliksin B -8.42 0.67 4
Gemsitabin -5.06 194.42 11
Genistein -6.59 14.67 6
Ginsenosida Rh2 28.04 3.68x1026 5
Kapsaisin -3.78 1688.73 1
Kuarsetin -6.20 28.35 6
29
Epigalokatekin-3-galat -3.06 5696.98 10
Eriokaliksin B -7.22 5.06 1
Epigalokatekin-3-galat -3.28 3929.03 4
Eriokaliksin B -5.49 94.05 3
Epigalokatekin-3-galat -3.24 4203.62 3
30
Epigalokatekin-3-galat -5.14 169.85 6
Eriokaliksin B -6.75 11.20 4
Epigalokatekin-3-galat -5.83 52.97 6
Eriokaliksin B -6.65 13.26 4
Epigalokatekin-3-galat -3.91 1355.85 7
31 Lampiran 5 Tingkat toksisitas berdasarkan nilai LD50
Tingkat Toksisitas Oral Rat LD50 (mg/kg)
Amat Sangat Beracun < 1
Sangat Beracun 1 - 50
Beracun 50 - 500
Sedikit Beracun 500 - 5000
Hampir Tidak Beracun 5000 - 15000
32
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 31 Agustus 1993. Penulis merupakan anak pertama dari 2 bersaudara, dari pasangan Djainul Abidin dan Yuliya. Tahun 2010, penulis lulus dari SMA Negeri 2 Jakarta dan diterima melalui jalur Seleksi Nasional Masuk Perguruan Tinggi Negeri (SNMPTN) secara tertulis di Departemen Kimia, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.