Indian Society of Pulses Research and Development
Indian Institute of Pulses Research
Kanpur, India
Number 3 & 4
of
Journal
Food Legumes
September - December 2013
I SPRD
1987
ISSN
0970-6380
Online ISSN
0976-2434
www.isprd.in
The Indian Society of Pulses Research and Development (ISPRD) was founded in April 1987 with the following objectives:
To advance the cause of pulses research
To promote research and development, teaching and extension activities in pulses
To facilitate close association among pulse workers in India and abroad
To publish “Journal of Food Legumes” which is the official publication of the Society, published four times a year.
Membership : Any person in India and abroad interested in pulses research and development shall be eligible for membership of the Society by becoming ordinary, life or corporate member by paying respective membership fee.
Membership Fee Indian (Rs.) Foreign (US $)
Ordinary (Annual) 350 25
Life Member 3500 200
Admission Fee 20 10
Library/ Institution 3000 100
Corporate Member 5000
-INDIAN SOCIETY OF PULSES RESEARCH AND DEVELOPMENT (Regn. No.877)
The contribution to the Journal, except in case of invited articles, is open to the members of the Society only. Any non-member submitting a manuscript will be required to become annual member. Members will be entitled to receive the Journal and other communications issued by the Society.
Renewal of subscription should be done in January each year. If the subscription is not received by February 15, the membership would stand cancelled. The membership can be revived by paying readmission fee of Rs. 10/-. Membership fee drawn in favour of Treasurer, Indian Society of Pulses Research and Development, through M.O./D.D. may be sent to the Treasurer, Indian Society of Pulses Research and Development, Indian Institute of Pulses Research, Kanpur 208 024, India. In case of outstation cheques, an extra amount of
Rs. 40/- may be paid as clearance charges.
EXECUTIVE COUNCIL : 2013-2015
Zone I : Dr Brij Nandan
(SKUAST) Sambha (J&K) Zone II : Vacant
Zone III : Vacant Zone IV : Dr OP Khedar
Durgapura, Jaipur, Rajasthan
Councillors
Dr Jagdish Singh, IIPR, Kanpur
Dr A Amarendra Reddy, ICRISAT, Hyderabad Dr CS Praharaj, IIPR, Kanpur
Chief Patron
Dr S Ayyappan
Patron
Dr SK Datta
Co-patron
Dr N Nadarajan
Zone V : Dr DK Patil Badnapur Zone VI : Vacant Zone VII : Vacant
Zone VIII : Dr Devraj Mishra IIPR, Kanpur, U.P.
Dr IP Singh, IIPR, Kanpur Dr Mohd Akram, IIPR, Kanpur
President
Dr NP Singh
Secretary
Dr GP Dixit
Joint Secretary
Dr KK Singh
Treasurer
Dr KK Singh (Acting)
Vice President
Dr Guriqbal Singh
Journal of Food Legumes
(Formerly Indian Journal of Pulses Research)
Vol. 26 (3 & 4)
September - December 2013
CONTENTS
RESEARCH PAPERS
1. Heavy metal toxicity to food legumes: effects, antioxidative defense and tolerance mechanisms 1 Navneet Kaur and Harsh Nayyar
2. Assessment of genetic diversity at molecular level in mungbean (Vigna radiata (L.) Wilczek) 19 S. K. Gupta, R. Bansal, U. J. Vaidya and T. Gopalakrishna
3. Effectiveness and efficiency of gamma rays and Ethyl Methane sulphonate (EMS) in mungbean 25 Kuldeep Singh and M.N. Singh
4. Combining ability analysis in medium duration CGMS based hybrid pigeonpea (Cajanus cajan (L.) Millsp.,) 29 M. P. Meshram, A.N. Patil and Abhilasha Kharkar
5. Genetic variability, character association and path analysis in clusterbean (Cyamopsis tetragonoloba (L.) Taub) 34 A. Manivannan and C. R. Anandakumar
6. Genetic analysis for quantitative traits in pigeonpea (Cajanus Cajan L. Millsp.) 38 C. K. Chethana, P. S. Dharmaraj, R. Lokesha, G. Girisha, S, Muniswamy, Yamanura,
Niranjana Kumar and D. H. Vinayaka
7. Genetic variabilty and association studies in cowpea (Vigna unguiculata L. walp.) 42 Hasan Khan, K. P. Vishwanatha and H.C. Sowmya
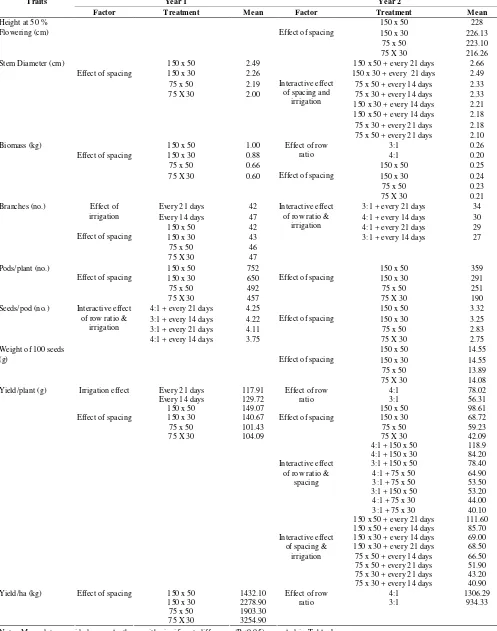
8. Yield and yield attributes of hybrid pigeonpea (ICPH 2671) grown for seed purpose as influenced by 46 plant density and irrigation
M.G. Mula, KB Saxena, A. Rathore and R.V. Kumar
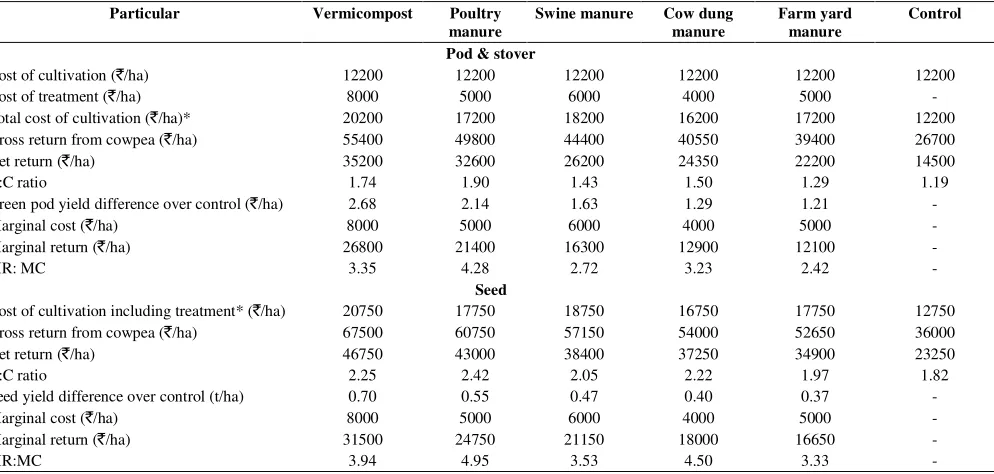
9. Influence of organic nutrient sources on growth, seed yield and economics of cowpea under 51 mid hills of Arunachal Pradesh
V.K. Choudhary, P. Suresh Kumar and R. Bhagawati
10. Pathogenic variation and compatibility groups in Sclerotium rolfsii isolates causing collar rot on 55 chickpea (Cicer arietinum L.)
O.M. Gupta, Sachin Padole and Madhuri Mishra
11. Efficacy of bioinoculants in combination with insecticides against insect pests of 59 blackgram Vigna Mungo (L.) Hepper
P.S. Singh and V. Chourasiya
13. Ovipositional preference of bruchid (Callosobruchus Maculatus Fabricius) on pod character and pod maturity 70 S. Nandini and G. Asokan
14. Beneficial traits of endophytic bacteria from field pea nodules and plant growth promotion of field pea 73 S. Narula, R.C. Anand and S.S. Dudeja
15. Effect of temperature-tolerant rhizobial isolates as PGPR on nodulation, growth and yield of 80 Pigeonpea [Cajanus cajan (L) Milsp.]
Simranjit Kaur and Veena Khanna
16. Phenotypic characterization of rhizobacteria associated with mungbean rhizosphere 84 Navprabhjot Kaur and Poonam Sharma
17. Root morphology and architecture (CRIDA indigenous root chamber-pin board method) of two 90 morphologically contrasting genotypes of mungbean under varied water conditions
V. Maruthi, K. Srinivas, K.S. Reddy, B.M.K. Reddy, B.M.K. Raju, M. Purushotham Reddy, D.G.M. Saroja and K. Surender Rao
18. Selection parameters for pigeonpea (Cajanus cajan L. Millsp.) genotypes at early growth stages 97 against soil moisture stress
Anuj Kumar Singh, J.P. Srivastava, R.M. Singh, M.N. Singh and Manoj Kumar
19. Optimization of extrusion process variables for development of pulse-carrot pomace 103 incorporated rice based snacks
Md. Shafiq Alam, Baljit Singh, Harjot Khaira, Jasmeen Kaur and Sunil Kumar Singh
20. Area expansion under improved varieties of lentil through participatory seed production programme 115 in Ballia District of Uttar Pradesh
S. K. Singh, Riyajuddeen, Vinay Shankar Ojha and Sanjay Yadav
21. Performance of chickpea in varied conditions of Uttar Pradesh 120
Lakhan Singh and A.K. Singh
22. Role of pulses in the food and nutritional security in India 124
Shalendra, K. C. Gummagolmath, Purushottam Sharmaand S. M. Patil
SHORT COMMUNICATIONS
23. Genetic variability and character association analysis in french bean (phaseolus vulgaris L.) 130 Anand Singh and Dhirendra Kumar Singh
24. Assessment of heritable components in chickpea (Cicer arietinum L.) 134 Sudhanshu Jain, S. C. Srivastava, Y. M. Indapurkar and H.S. Yadava
25. Genetic variability and character association for yield and its components in black gram 137 (Vigna mungo (L.) Hepper)
A. Narasimhan, B. R. Patil and B. M. Khadi
26. Studies on genetic variability, heritability and genetic advance in chickpea (Cicer arietinum L) 139 Shweta, A.K. Yadav and R.K. Yadav
27. Effect of zinc, molybdenum and Rhizobium on yield and nutrient uptake in summer urdbean (Vigna mungo L.) 141 Khalil Khan and Ved Prakash
28. Effect of seed dressers against root rot of cowpea 145
D. B. Patel, S. M. Chaudhari, R.G. Parmar and Y. Ravindrababu
29. Development of tempeh a value added product from soyabeans and other underutilised 147 cereals/millets using Rhizophus Oryzae PGJ-1
Journal of Food Legumes 26(3 & 4): 1-18, 2013
Heavy metal toxicity to food legumes: effects, antioxidative defense and tolerance
mechanisms
NAVNEET KAUR and HARSH NAYYAR*
Department of Botany, Panjab University, Chandigarh, India; *E-mail:[email protected]
ABSTRACT
Heavy metal stress has emerged as one of the most detrimental for the major food crops due to persistent soil pollution. These metals are known to replace essential metals in pigments or enzymes disrupting their function and have thus proved to be toxic. Their toxicity results in chlorosis, weak plant growth, yield depression accompanied by reduced nutrient uptake, disorders in plant metabolism and, in leguminous plants, a red uced ability to fix molecula r nitrog en. Most of the leguminous crops are affected by metal stress present in the soil mainly due to contaminated agrochemicals and sewage sludge. They are known to cause deleterious effect on the cell division of the pla nts and cause ma ny chromosomal abnormalities which depend upon the concentration and the intensity of the exposure of the same. Uptake and accumulation of metals at higher concentrations cause ultra-structural and an atomical cha nges in the plan t cells. Various pla nt physiological a ctivities like seed germination, nutrition distribution, enzymatic activity, nitrogen fixation, photosynthesis and pollen function are adversely affected. Uptake into the seeds adversely affects the nutritional quality of seeds. However, the plants possess various defense and tolerance mechanisms to cope up with stresses. In this review, we describe various effects, defense and tolerance mechanisms due to heavy metals, especially for legumes.
Key words:Heavy metals, Legumes, Tolerance mechanism, Toxicity
1. Introduction
Plants experience various biotic and abiotic stresses which adversely affect the growth,deteriorate the yield and physiological parameters of the crops. Among these stresses, heavy metal stress has emerged as one of the most detrimental for the major food crops. Toxic metals have also contributed towards most of the soil and water pollution in the environment. The term heavy metal refers to any metallic chemical element that has relative high density and molecular weight and is toxic even at low concentrations. These include the transition metals, metalloids, lanthanides and actinides in the periodic table, mostly having a specific gravity greater than 5.0 (Nieboer and Richardson 1980). According to another view, heavy metals can include elements lighter than carbon and can exclude some of the heaviest metals. While speaking of heavy metals, we generally mean lead, mercury, iron, copper, manganese, cadmium, arsenic, nickel, aluminium, silver and beryllium. Some heavy metals are essential to the living organisms but at higher concentrations they produce toxicity.
In general, heavy metals produce toxicity by forming complexes or “ligands” with organic compounds. These modified biological molecules lose their ability to function properly, and result in malfunction or death of the affected cells. The most common groups involved in ligand formation are oxygen, sulphur, and nitrogen.Carboxylic acids and amino acids such as citric acid, malic acid and histidine are potential ligands for the heavy metals(Rauser 1999). When the metals bind to these groups they may inactivate the important enzyme systems, or affect protein structure of various biological molecules and thus may block the pathways of major physiological processes. These have affected the biosphere in many places worldwide (Cunningham et al. 1997). Heavy metals are one of the main abiotic stressors for the plants because of their increased use in the industries and agrotechnics and also because of their high bioaccumulation ability and toxicity (Maksymiec 2007). Heavy metals are known to replace essential metals in pigments or enzymes disrupting their function (Ghosh and Singh 2005) and have thus proved to be toxic. Roots are the first target to the heavy metals, which get damaged and lead to inhibition of the plant growth and ultimately crop failures. A long-term exposure of whole plants to enhanced metal concentrations may also affect the chlorophyll synthesis and thus have an important role in both
D
r. Harsh Nayyar is a Professor of Stress Physiology at Department of Botany, Panjab University, Chandigarh, India. He received his M.Sc. and Ph.D from Punjab Agricultural University, Ludhiana India. He worked as a Scientist (A.R.S.) under I.C.A.R., New Delhi for some time and2 Journal of Food Legumes 26(3 & 4), 2013
the chloroplast development in young leaves and the inhibition of photosynthesis (Boddi et al. 1995).
The sources of heavy metals pollution can be both natural and anthropogenic. Natural sources include the mother rocks and minerals of the metals while the anthropogenic sources are agriculture, black and colored metallurgy and transport (Budnikov 1998).There are many records that the places adjacent to the industrial places have been affected mainly by the toxic heavy metals (Rao 1979). Human activity has contributed metal concentrations in soil range from less than 1 mg/kg to as high as 100,000 mg/kg (Blaylock and Huang 2000), which have lead to the disruption of natural, aquatic and terrestrial ecosystems. Above certain concentrations and over a narrow range these heavy metals can turn into toxic products (Babich and Stotzky 1978). It is reported that the continuous exposure to the heavy metals have severely affected the health of human beings, plants, animals and the microbial populations that have disrupted the ecological habitats (Sterritt and Lester 1980, Brynhildsen and Rosswall 1997).
2. Toxic effects of various heavy metals on plants Heavy metals cause acute toxic effects on plants grown on the contaminated soils. Their toxicity results in chlorosis, weak plant growth, yield depression accompanied by reduced nutrient uptake, disorders in plant metabolism and, in leguminous plants, a reduced ability to fix molecular nitrogen (Chaudri et al. 2000, Dan et al. 2008). Agricultural yield is decreased due to their prevalent use and cause dangerous health effects as they enter into the food chains (Schickler and Caspi 1999). Heavy metals are transmitted through the food chains and strongly affect the human health (Brun et al. 2001, Gincchio et al. 2002). There are metals like Pb, Hg, Cd, Ar and Cr, which have no biological function but are toxic to life even at very low concentration (Salt et al. 1995). Many metals are nutrients at permissible levels but act as toxins when the concentrations found in the soil exceed to those required as the nutrients. Arsenic is non-essential element and generally toxic to plants. Symptoms include poor seed germination and marked reductions in root growth. Plants may show reduced growth, nutrient deficiencies and chlorosis resulting from reduced chlorophyll biosynthesis (Mascher et al. 2002).Cadmium is also a non-essential element, the accumulation of which may cause several physiological, biochemical and structural changes in plants (Feng et al. 2010). It also alters mineral nutrients uptake, cause inhibition in stomatal opening by interacting with the water balance of plant (Hossain et al. 2010), disturbs the Calvin cycle enzymes, photosynthesis, carbohydrate metabolism (Shi et al. 2010) and ultimately reduces the crop productivity (di Toppi and Gabreilli 1999).There are reports that excess of chromium causes inhibition of plant growth, chlorosis in young leaves, nutrient imbalance, wilting of tops, and root injury (Scoccianti
et al. 2006). It inhibits the chlorophyll biosynthesis in terrestrial plants (Vajpayee et al. 2000). High levels of Cr affected total dry matter production and yield of plants (Shanker et al. 2005). Like other heavy metals, chromium toxicity produces chlorosis and necrosis in plants (Cervantes et al. 2001).
Copper is considered as an important micronutrient for plants (Thomas et al.1998) and plays important role in CO2 assimilation and ATP synthesis. It is known to be an essential component of various proteins like plastocyanin of photosynthetic system and cytochrome oxidase of respiratory electron transport chain (Demirevska-kepova et al. 2006) as well as other various proteins. In acute Cu toxicity, leaves may become wilted before eventually becoming necrotic (Yau et al. 1991). Plants can accumulate some amount of cobalt from the soils but the uptake and distribution of cobalt in plants is species-dependent and is controlled by different mechanisms (Kukier et al. 2004, Li et al. 2004 and Bakkaus et al. 2005). There are some reports that Co adversely affects the shoot growth and biomass in some plants (Li et al. 2009). Lead is a major heavy metal that has gained importance as a potent environmental pollutant (Sharma and Dubey 2005). Lead affects a number of plants by decreasing the productivity of plants growing in lead- polluted soils (Johnson and Eaton 1980). Its phytotoxicity leads to inhibition of number of enzyme activities, disturbances in the mineral nutrition, water imbalance, and changes in the hormonal status and alteration in the membrane permeability. At high concentrations, lead may even lead to cell death (Seregin and Ivanov 2001). Mercury is also a very important toxic metal. Exposure to the mercurials in crop plants are by direct administration as antifungal agents, through the seed treatment or foliar spray, or accidentally (Patra and Sharma 2000). It affects both the light and dark reactions of photosynthesis by the substitution of magnesium of chlorophyll thereby resulting in the breakdown of photosynthesis. It binds to water channel proteins, thus inducing leaf stomata to close and physical obstruction of water flow in plants (Zhang and Tyerman 1999). High levels of mercury interferes with the mitochondrial activity and induces oxidative stress by triggering the generation of ROS thus leading to the disruption of biomembrane lipids and cellular metabolism in plants ( Israr et al. 2006, Cargnelutti et al. 2006).
Nickel also causes phytotoxicity by altering physiological reactions and diverse toxicity symptoms such as chlorosis and necrosis in different plant species (Zornoza et al. 1999, Pandey and Sharma 2002, Rahman et al. 2005), including rice (Samantaray et al. 1997). Yadav (2009) have reported that plants grown in high Ni2+ containing soil showed
Kaur & Nayyar : Heavy metal toxicity to food legumes: effects, antioxidative defense and tolerance mechanisms 3
the application of zinc (Fontes and Cox 1998). Prolonged exposure to zinc causes chlorosis in the younger leaves, which can extend to older leaves (Ebbs and Kochian 1997). 3. Effects on Legumes
Legumes belonging to the family Fabaceae are responsible for substantial part of the global flux of nitrogen from atmospheric nitrogen to the fixed forms such as ammonia, nitrate and organic nitrogen. Most of these are agriculturally important food crops and are a rich source of proteins to the animals and human beings(Gupta 1987). Most of the leguminous crops are affected by metal stress present in the soil mainly due to contaminated agrochemicals and sewage sludge. Due to this, various plant physiological activities like seed germination, nutrition distribution, enzymes activity, alternation of the membrane permeability, nitrogen fixation, photosynthesis, transport of the assimilates and respiration are adversely affected.
3.1. a. Cytology:
Heavy metals adversely affect the cytology of the plant cells. The genotoxicity of heavy metals influences the DNA synthesis, duplication of DNA and chromosomes causing many chromosomal aberrations (Cheng 2003). They are known to cause deleterious effect on the cell division of the plants, which depend upon the concentration and the intensity of the exposure of the same.
Fig: 1 Heavy metal induced chromosomal abnormalities in plant cells
The effects of Cd on the cell division of the root tips in beans were studied by Mo and Li 1992. The treatment of beans by Cd, Pb , and Hg at concentrations 0.01,1.0 and 10 ppm, respectively shortened the cell division but extended the cell cycle. The chromosomes of beans exposed to Cd, Pd, Hg got injured and revealed polyploidy, C-karyokinesis and various chromosomal abnormalities like formation of chromosomal bridges, chromosomal rings and chromosome fragmentation, chromosome micro-nuclei and nuclear decomposition (Mo and
Li 1992, Liu et al.1992, Duan and Wang 1995). ( Fig. 1). Gomez-Arroyo et al. (2001) reported that the salts of nickel, cobalt and cadmium increased the frequency of the sister chromatid exchanges (SCE) in Vicia faba. It has been observed in the some plants that mercury poisoning leads to the disturbance in the spindle activity resulting in the formation of polyploid and aneuploid cells and c-tumours. Kumar (2007) studied the mutagenic potential of lead (25, 50, 100, 200, and 300 ppm) in Lathyrus sativus and showed that chromosomal abnormalities increased with the lead nitrate concentration. These abnormalities included condensed bivalents, laggards, bridges, cytomixis and stickiness of the chromosomes. Siddiqui et al.(2009) studied the effect of cadmium on the root tips of Pisum sativum L. Seeds of P. sativum were treated with a series of concentrations ranging from 0.125, 0.250, 0.500 and 1 mM Cd for 6 h. It was reported that the overall percentage of aberrations generally increased with increasing concentrations of Cd. The most common chromosomal abnormalities were laggards, bridges, stickiness, precocious separation and fragments. Zhang et al. (2009) investigated the effects of different concentrations of Cd (1-50µM) on cell division and nucleoli in root tip cells of Vicia faba. Results revealed that Cd induced c-mitosis, chromosome bridges, chromosome stickiness and lagging chromosomes. Small amounts of nucleolus materials were extruded from the nucleus into the cytoplasm at 1µM Cd when the root tips were exposed for 24 hours. Zhang et al. (2009) studied the effects of different concentrations of Al (10 µM, 50 µM, 100 µM) on nucleoli in root tip cells in hydroponically grown Vicia faba L. It was revealed that aluminum significantly inhibited root growth of V. faba treated with 50 µM and 100 µM concentrations. In the nucleolus of root tip cells, some particulates containing agyrophilic proteins were extruded from the nucleus into the cytoplasm, and some were scattered in the nucleus after Al stress in the plants. In the root tips of green gram (Vigna radiata. L), arsenic and manganese induced chromosomal stickiness at higher concentrations. However it was reported that arsenic had more toxic effect than manganese on the root tip cells of greengram during mitosis (Mumthaz 2010).
Studies were carried out by Muneer et al. (2011) on root tips and leaves of Vigna radiata, where 15 days old plantlets grown in nutrient Hoagland media were exposed to various levels of cadmium chloride (0.05, 0.10 and 0.50 mM) for 48 and 72 hours. It was shown that Cd exhibited inhibitory effect on cytological studies namely, mitotic index and chromosome number. Chromosomal studies showed various chromosomal abnormalities such as laggard chromosomes, anaphasic bridges, and uni-distribution of chromosomes.
4 Journal of Food Legumes 26(3 & 4), 2013
tail length (CTL), comet tail DNA (CT, DNA), and tail moment (TM) with increased concentration of three heavy metals, like Cd, Pb and Hg. The most prominent abnormalities induced by heavy metals were micronucleus, precocious separation, and laggard formation. Ritambhara et al.(2010) carried out some cytogenetic studies to evaluate the genotoxic effect of lead (Pb) and zinc (Zn) on the gametic cells of grass pea (Lathyrus sativus). A severe chromosome stickiness in meiosis impairing normal chromosome segregation was reported, which persisted upto microspore stage, ultimately leading to chromosome degeneration. Recently, Oladele et al. (2013) reported thatincreased metal pollution can lead to some irreversible cytogenetic effects in plants and higher organisms. Their group investigated the effects of lead and zinc nitrates at different concentrations: 25, 50 and 100 mg/L on the chromosomes of bambara groundnut (Vigna subterranean). The results show the most frequent chromosomal anomalies induced by these heavy metals as stickiness and bridges. 3.1.b. Anatomy and Ultrastructure
Han et al. (2004) have reported that uptake and accumulation of metals at higher concentrations cause ultra-structural and anatomical changes in the plant cells. These include structural modifications in the chloroplast, thylakoid membranes, deposition of electron dense globules in vacuoles, increase or decrease in the size of cells, reduction of the intercellular spaces and also changes in the turgor pressure of the plant cells. In the thylakoids, modifications include swelling and curling of the thylakoidal membranes. Also, it has been reported that heavy metal stress causes degradation of polypeptide compositions of the thylakoid membrane, reduction, disappearance or swelling of the grana cascade of chloroplast mitochondria (Cheng 2003). Heavy metal induced cellular and ultrastructural modifications have been illustrated in Fig.2.
There was accumulation of callus on the sieve plates of phloem of Phaseolus vulgaris seedlings exposed to excess of Co, Ni and Zn (Peterson and Rauser 1979). In case of beans, toxicity of chromium has also found to decrease the diameter of the treachery vessels, which resulted in reduced longitudinal water movement (Vazques et al.1987). Reduced turgor pressure and plasmolysis in the epidermal and cortical cells of bush bean plants, which were exposed to chromium was also observed (Vazques et al.1987). Formation of wall ingrowths in hypodermal cells of the cadmium treated Phaseolus vulgaris roots was reported (Vazques et al.1989). Cadmium treatments to Arachis hypogea plants induced xerophyte anatomic features of leaves i.e. thick lamina, upper epidermis, palisade mesophyll, high palisade to spongy thickness ratio, as well as abundant and small stomata (Shi and Cai 2008). In 5µg/ml cadmium treated roots of bush bean plants (Phaseolus vulgaris L. cv. Contender) grown on perlite, plastid ultrastructure was hardly affected, while in the upper parts of the plant the chloroplasts showed severe alterations. Younger trifoliate leaves showed greater disruption of chlorophyll synthesis and plastid ultrastructure (Barcelo et al. 1987). In the experiments of Baszynski et al. (1980) and Stoyanova and Chakalova (1990), it was established that cadmium, applied in toxic concentrations, disturbs the chloroplast envelope and the integrity of the membrane system and leads to increased plastoglobule number, changing the lipid composition and the ratios of the main structural components of thylakoid membranes. Heavy metals effects on the structure and functions of the photosynthetic membranes of the higher plants showed that the sub-microstructure of the chloroplast was changed (Yang 1991). Ultra structural alternations in the cortical root cells of pigeon pea in response to zinc and nickel both at cellular and the organelle level were observed (Sresty et al. 1999). Unusual deposition of electron dense globules in the vacuoles of the root cortical cells and appearance of unusual two nucleoli occurred. The ultra structural analysis of the leaves of pea plants grown with 50µM CdCl2 showed the internal cell disturbances characterized by an increase of mesophyll cell size, a reduction of the intercellular spaces and severe disturbances in the chloroplast structure (Sandalio et al. 2001). In the pea leaves treated with Cd, there was disorganization of the chloroplast structure, with an increase in the number of plastoglobuli and formation of vesicles in the vacuoles (Mc Carthy et al. 2001). The effect of Cu2+ at concentrations
(50 and 75uM) on the ultra-structure of the chloroplasts of the bean seedlings revealed that excess of copper induced changes in the ultra-structure of chloroplasts visible in form of deterioration in the grana structure and the accumulation and swelling of starch grains in the stroma (Bouazizi et al. 2010).
Kaur & Nayyar : Heavy metal toxicity to food legumes: effects, antioxidative defense and tolerance mechanisms 5
3.2 Germination of legume seeds
Germination of the seed is one of the most highly sensitive processes and toxicity with the essential and non essential heavy metals invariably affects the germination rate and subsequent seedling growth. It represents a limiting stage of plant life cycle under heavy metal stress situation (Rahoui et al. 2008, Smiri et al. 2009). High levels (500 ppm) of hexavalent Cr in soil reduced germination up to 48% in the bush bean Phaseolus vulgaris (Parr and Taylor 1982). The growth of embryonic axis of germinating pea seeds (Pisum sativum cv. Bonneville) was significantly inhibited by as low as 0.25 mM cadmium and the elongation of the radicle was affected more severely than that of the plumule (Chugh and Sawhney 1996). In another experiment, Al-Yemini and Nasser (2001) reported a significant decrease in the percentage of seed germination, seedling growth and an increase in radicle length of Vigna ambacensis L.after treatment with cadmium, mercury and lead at concentrations (0.05-50mM). Seed germination of the alfalfa plant (cultivar Malone) was seriously affected by a concentration of 20 ppm of Cd(II), Cr(VI), and by 40 ppm of Cu(II) and Ni(II) (Peralta et al. 2001). However, the root and shoot growth of the alfalfa plant was stimulated by a concentration of 5 ppm of Cr(VI), Cu(II), Ni(II), and Zn(II). In a study, Al-Rumaih et al. (2001) revealed that cadmium chloride adversely influenced the germination process of Vigna unguiculata seeds. The germination percentage and the germination rate index (GRl) expressed as percentage germination per day showed a significant (p~0.05) decrease at 40 ppm, and the decline in these measures became highly significant (p~0.01) at 80 and 160 ppm cadmium chloride concentrations, as compared with the untreated control ones. Cr (VI) at 40 ppm reduced seed germination by 23% of seeds of lucerne (Medicago sativa cv. Malone) and growth in the contaminated medium (Peralta et al.2001).The effect of several doses of As (V), Cd (II), Pb (II), Hg (II), Cu (II), Zn (II) on the seed germination of four common pulses (Vigna mungo (L.)Hepper, Vigna radiata (L.)Wilzek, Pisum sativum L. and Lens culinaris L.) were observed by Mandal and Bhattacharyya 2007. Certain levels of some heavy metals were favourable for seed germination while others were the inhibitory ones. Exposure of 20 ppm Hg (II) showed higher toxicity than other heavymetals and reduced the germination potential to 50% as compared to control. It was also shown that the order of toxicity of the metal elements on the four pulses decreased as follows: Hg> As> Cd> Pb> Cu> Zn. In alfalfa, it was reported that the seed germination is seriously affected by 20 ppm of Cd+2, Cr+6, and by 40 ppm of Cu+2, Ni+2
while the root and shoot growth are stimulated by 5 ppm of Cr+6, Cu+2, Ni+2, and Zn+2(Aydinalp and Marinova 2009). In
another experiment Al-Qurainy (2009) studied the toxicity of Al and Ni individually on P. vulgaris which affected root and shoot length and their effects on length was appeared after 3 days of germination but % germination was not inhibited in both metal treatments. A reduction in seed germination and
seedling growth in chickpea treated with 50, 100, 200 and 400 ppm of nickel and cobalt was reported by Khan and Khan et al. 2010. The germination of Vigna unguiculata seeds after treatment in solution containing varying concentration of cadmium chloride (CdCl2.H2O) was observed (Egharevba and Omoregie 2010).The concentrations of cadmium (Cd) in the solution used for the treatment were 0.00 ppm, 0.80 ppm, 8.00 ppm, 40.00 ppm, 100.00 ppm and 180 ppm. Results showed that the percentage germination and rate of increment in shoot height decreased as cadmium level in the treatment solution increased. However, no growth was observed at 100 and
120 ppm.
There can be a number of reasons behind the decreased rate of seed germination in plants. The decrease in seed germination of heavy metal treated plants can be attributed to the accelerated breakdown of stored food materials in seed (Shafiq et al. 2008). Several authors reported that the inhibition of root elongation caused by heavy metals may be due to metal interference with cell division, including inducement of chromosomal aberrations and abnormal mitosis (Radha et al. 2010, Liu et al. 2003), which can affect the seedling growth. These observations indicated variable effects of metals, though inhibitory in each of the legume species tested. 3.3. Nitrogen fixation and symbiosis
It is reported that excessive metal concentrations in the polluted soils cause damage to Rhizobia, legumes and their symbiosis (Ahmad et al. 2012). The establishment of symbiosis i.e. root nodulation is an orderly process, which is influenced by various edaphic factors and the presence of pollutants in the soil. However, very little is known about how legume– Rhizobium symbiosis is affected by varying metal concentration (Huang et al.1974, Mcilveen and Cole 1974, Decarvalho et al. 1982, Paivoke 1983, Yakoleva 1984). Various effects of heavy metals on nitrogen fixation are shown in Fig.3.
3.3.1 Heavy metals inhibit the activity of the symbiotic nitrogen fixers
Heavy metals affect the nitrogen fixation process by interfering with the performance of the symbiotic bacteria.
6 Journal of Food Legumes 26(3 & 4), 2013
Various metals (e.g. Cu, Ni. Zn, Cd, As) are known to inhibit the growth, morphology and activities of various symbiotic N2 fixers (Stan et al. 2011) like R. leguminosarum, Mesorhizobium ciceri, Rhizobium sp. and Bradyrhizobium sp. and Sinorhizobium (Arora et al. 2010, Bianucci et al. 2011). The population of R. leguminosarum bv. trifolii was radically altered by long-term exposure to heavy metals and it lost the ability to form functional symbiosis with white and red clover (Hirsch et al. 1993). Chaudri et al. (2000) reported a decrease in two agriculturally important species of Rhizobia, R. leguminosarum bv. viciae and R. leguminosarum bv. trifolii, in soils, which were irrigated with sewage sludge containing Zn or Cu or mixture of Zn and Cu. Similarly, there are numerousreports where elevated amounts of heavy metals have been found to limit the rhizobial growth and their host legumes (Heckman et al. 1987, Broos et al.2005) and therebyreducing the total crop yields (Moftah 2000). 3.3.2 Inhibition and delay in nodulation in the legume roots
Nodulation in the soybean roots was greatly inhibited by the addition of Cd (10-20mg/kg) to the soil (Chen et al. 2003) but the nitrogen fixation of the root nodule was stimulated with low concentrations of Cd, which decreased sharply with the further additions. The impact of heavy metals such as cadmium (23mg/kg) and lead (390mg/kg) on nitrogen uptake in chickpea was studied by Wani et al. 2008. It was reported that cadmium and lead reduced the number of nodules by considerable percentages. The total nitrogen content of the shoots, nodule weight, nodule number and N2 (C2H2 )-fixation were reduced significantly in dry beans treated with 10µM Cd /L (Vigue et al.1981). The responses of Lablab purpureus-rhizobium symbiosis to the effect of different levels of heavy metals Cd, Zn Co and Cu at concentrations (control, 50, 100, 150 and 200 mg/kg soil) was reported (Younis 2007). It was reported that there was enhancement in the nodule number and their mass in the soil treated with 100 mg/kg soil of Co and Cu, respectively while there was inhibition at other levels. There was severe inhibition in the nitrogenase activity. Delay in the nodulation process in some legume crops has also been observed. For example, with increasing concentration of arsenic (As) in the nutrient solution, there was greater time required for Bradyrhizobium japonicum strain CB1809 to inoculate soybean. (Riechman 2007). 3.3.3 Decrease in the rate of symbiosis
A considerable decrease in the total yield, nitrogen content in the plant tissue and the protein content of the seeds was noticed chickpea-rhizobium and green gram-bradyrhizobium symbiotic systems when treated with cadmium, lead, copper, zinc, chromium and nickel added in combinations and separately (Athar and Ahmad 2002). Adverse effects of sludge application on N2 fixation in faba bean (Vicia faba) have been reported (Chaudri et al.1993). In white clover, cadmium, lead and zinc caused reduction in growth and symbiosis when they were grown in soils highly contaminated
with these metals (Rother et al.1983). Chickpea-rhizobium symbiotic system was more sensitive to the metals toxicity than green gram-bradyrhizobium system (Antipchuk et al. 2000).The adverse effects of mercury, cadmium, nickel (1mg/ml) on nodulation and nitrogen fixation in Cicer arietinum-rhizobium symbiotic system were reported as decrease in dry weight of both total nodules and effective nodules indicating disturbances in the nodule function (Pal 1996).
3.3.4.Nitrogenase activity and protein content
Soybean (Glycine max L.) nodules and roots in plants were subjected to two different concentrations (50 and 200 µM) of CdCl2 (Karina et al. 2003). Nitrogenase activity decreased in nodules treated with 200 µM Cd2+. In 50 µM
Cd2+-treated plants, NH
4+ content increased by 55% in roots.
Glutamate (Glu) and protein contents remained unaltered in nodules treated with 50 µM Cd2+, while at the higher Cd2+
concentration, both were decreased.Cadmium significantly depressed biological nitrogen fixation in 0-10mM treated one month old pea (Pisum sativum) and also decreased nitrate reductase and glutamate synthase activities 6 days after treatment (Chugh et al. 1999). In Lablab purpureus, nodulation and nitrogenase activity were severely affected with 50-200 mg/kg metal concentrations of Cd, Zn, Co, and Cu (Younis 2007). (Fig.3)
3.4 Photosynthetic efficiency of some legumes
Heavy metals are known to interfere with many vital processes of the plants including photosynthesis (Clijsters and Assche 1985) and cause inhibition in this process. This inhibition has been attributed with an indirect action on plant water balance, stomatal conductance and CO2 availability. The direct effects include effect on chloroplast organization, chlorophyll biosynthesis, electron transport and enzymes of photosynthetic carbon metabolism (Fig. 4).
3.4.1 Interference in the photosynthetic function and machinery
Heavy metals lead to oxidative stress in the plants. As a result, there is decreased photosynthetic activity and growth of tissues, which is followed by reduction of plant productivity (Ouzounidou 1995 Maksymiec 1997). The insufficiently utilized assimilatory force by Calvin cycle slowed down due to heavy
Kaur & Nayyar : Heavy metal toxicity to food legumes: effects, antioxidative defense and tolerance mechanisms 7
metal stress may, in consequence, enhance proton gradient formed in chloroplasts and increase non-photochemical dissipation of light energy and/or decrease photochemical efficiency (Maksymiec and Baszyn´ski 1996, Maksymiec 1997). Many heavy metals are known to interfere with the photosynthetic machinery (Fig.4). For example cadmium interferes with the chloroplast function and electron transport system by damaging PSII of photosynthesis. Copper shows negative effect on the components of both the light reactions e.g., PSII, thylakoid membrane structure andchlorophyll content (Ralph and Burchett 1998, Szalontai et al.1999, Pätsikkä et al. 2002 and CO2-fixation reactions ( Angelov et al.1993). However, in studies that examined both light and CO2-fixation components, the relative sensitivity of each to Cu varies among studies ( Moustakas et al. 1994). Krupa and Baszynski (1995) investigated the heavy metals treated legumes and reported that they severely affect the rate of photosynthesis by inhibiting the light and dark photosynthetic reactions, inhibiting the enzymes of the carbon reduction pathways and disturbing the photosynthetic apparatus.
3.4.2.Interference in chlorophyll synthesis
Studies indicate that heavy metals have deleterious effects on the total chlorophyll content in plants (Fig.4). The effect of Cr on chloroplast pigment content in mungbean showed that irrespective of concentration, chlorophyll a, chlorophyll b and total chlorophyll decreased in 6-day-old mungbean seedlings (Bera et al.1999). Effects of some heavy metals on content of chlorophyll in bean Phaseolus vulgaris seedlings was investigated by Zengin and Munzuroglu 2005 grown in Hoagland solution spiked with various concentrations of Pb, Cu, Cd and Hg. It was reported that the total chlorophyll content declined progressively with increasing concentrations of heavy metals. The total chlorophyll content, chlorophyll b content and carotenoids content was severely affected with in blackgram varieties treated with lead and copper (Bibi and Hussain 2005). Also, it was concluded that application of lead and copper to both the black gram cultivars caused significant reduction in the photosynthetic gas exchange, inactivation of enzymes such as Rubisco, Rubisco activase and carbonic anhydrase. Total chlorophylls and carotenoids were calculated from the seedlings of Cyamopis tetraganoloba treated with heavy metals Cd, Pd, Ni, Zn and Cu. It was concluded that Cd and Pb in comparison to Zn, Cu, and Ni reduced the total chlorophyll content at 1000 ppm. Shi and Cai (2008) reported the effects of cadmium treatments on Arachis hypogea plants and concluded that these treatments caused a decrease in the net photosynthetic rate and reduced the content of the photosynthetic pigments as well. Phaseolus vulgaris L. plants grown in soil supplemented with different Pb and Cd concentrations (2,4, 6, 8 g kg-1 for lead and 1.5, 2.0, 2.5, 3.0 g
kg-1 for cadmium) showed decrease in the content of
photosynthetic pigments, total soluble sugars, starch content as well as soluble protein. However, total free amino acid content and lipid peroxidation were increased with increasing concentration of heavy metals (Bhardwaj et al. 2009). Kamel (2008) treated Vicia faba plants with different concentrations of lead nitrate ranging from 0-48 mM in hydroponic solution. It was observed that low doses of Pb (0.49 mM) increased the chlorophyll content while the chl-a content decreased at high concentrations of Pb (48 mM). It was also observed that the
14C-fixation decreased at all the applied Pb concentrations.
3.4.3. Reduction in the activities of photosynthetic enzymes and inhibition of photosynthetic rate
Heavy metals interfere with chlorophyll synthesis either through direct inhibition of an enzymatic step or by inducing deficiency of an essential nutrient (Van Assche and Clíjsters 1990). Sheoran et al.(1990) studied the effect of Cd2+ and Ni2+ on the rate of photosynthesis and activities of key enzymes of the photosynthetic carbon reduction cycle in leaves from pigeonpea (Cajanus cajan L., cv. UPAS-120) grown in nitrogen free sand culture. It was concluded that the application of Cd2+ and Ni2+ (0.5 and 1.0 mM) at an early vegetative stage (30
days after sowing) resulted in about 50% and 32% reduction in net photosynthesis, respectively. The activities of the photosynthetic enzymes were decreased to different levels (2–61%) depending upon the enzyme and the concentration of the metal ion. It was found that Cd toxicity caused notable reduction in photosynthetic rate in different plant species (Baszynski et al.1980). In case of pigeon pea, Cd concentrations of 56 and 112 mg/L inhibited net photosynthesis to about 50% at early development stages (30-day-old plants) and did not exert any significant effect on that process at later stages (70-day-old plants). Sheoran et al.(1990) also reported that at the early stage of pigeonpea, CO2 exchange rate was quite susceptible to Cd stress. Application of copper and lead at concentrations 25 or 50 mg L-1 in two Mungbean cultivars Vigna radiata (L.) Wilczek]
(Mung-1 & Mung-6) caused significant reduction in the CO2 exchange and photosynthetic pigments. Also, there was significant inhibition of photosynthetic and transpiration rates and stomatal conductance compared to the same doses of copper at higher concentration of lead (50 mg L1) compared to
8 Journal of Food Legumes 26(3 & 4), 2013
3.5 Reproductive biology: Pollen function
It has been found, that inhibitory effects of sulfur dioxide on pollen germination and tube elongation have occurred at concentrations lower than those at which foliar effects have been recorded (Varshney and Varshney 1980). This suggests that heavy metals, even if present at low concentrations in plant tissue, may affect pollen germination and pollen tube growth. Pollen tubes are excellent standard systems with which the effects of drugs and pollutants can be investigated (Kristen et al.1993). Studies report that heavy metals at toxic levels inhibit pollen germination, pollen tube growth (Tuna et al. 2002) by causing ultra-structural changes. Xiong and Peng(2001)tested 5 herb species (Vicia augustifolia, V.tetrasperma, Pisum sativum, Plantago depressa, and Medicago hispida) for their responses against Cd exposure for pollen germination and tube growth to Cd exposure in vitro. Results revealed that pollen germination of all the species was inhibited at Cd concentrations of 2.51µg/ml and higher, and tube growth was inhibited at concentrations of 1.58µg/ml and higher. The pollen response to Cd stress exhibited interspecies differences. Vicia angustifolia and V. tetrasperma were sensitive to Cd and were inhibited in either pollen germination or tube growth by Cd at 0.01 µg/ml. At 1 µg/ml, pollen tube growth of V. angustifolia, V. tetrasperma, and P. sativum was inhibited. Results suggested that cadmium at such a low concentration as 0.01 lg/mL is able to exert adverse effects on pollen germination in some sensitive species, while it fails to do so for less-sensitive species.
3.6 Phyto-hormones
Quantitative determination of endogenous chemical compounds including hormones in plants growing under controlled environmental conditions in presence and absence of heavy metals is used to study the effect of heavy metals (Varga et al.1999). Atici et al. (2005) investigated the changes in abscissic acid, gibberellic acid, zeatin and zeatin riboside hormones of chickpea seeds germinating under Pb or Zn heavy metals exposure. Pb increased abcisic acid and zeatin contents while decreased gibbrellic acid content in the germinating seeds. High concentrations of Zn (1.0 and 10 mM) decreased contents of zeatin, zeatin riboside and giberellic acid while 0.1 mM Zn increased the content of the same hormones. ABA content was enhanced by Zn in all concentrations used.Cakmak et al. (1989) estimated the concentrations of phytohormones particularly IAA and concluded that its concentration in zinc-deficient bean (Phaseolus vulgaris L.) plants are clearly lower compared to those of Zn-sufficient plants(Zn2+), changes in concentrations of ABA are less
distinct. Re-supply of Zn to deficient plants restores the IAA level to that of the Zn-rich plants within 96 h, whereas the ABAconcentrations are only slightly increased after Zn resupply.In contrast, the effect of Zn nutritional status oncytokinin levels is less clear. Information on effect of phytohormones is sparse and is required to be worked out.
3.7 Adverse effects on nutritional value of legumes Limited data suggests that some heavy metals induce substantial reduction in the nutritional quality of the seeds in terms of accumulation of starch, proteins, amino acids, and minerals. It has been reported that As interferes in the uptake and accumulation of minerals in seeds and shoots (Paivoke and Simola 2002) and may alter nutritional composition (Tu and Ma 2005). Paivoke and Simola (2002) reported that As (12.5 to 73.3 mg of sodium arsenate/kg dry weight of soil) caused interference in mineral nutrient balance of Zn, Mg, and Mn in peas. Similarly, in pea, growing in 2.5mM cadmium a decrease in starch content of its seeds was observed, however, the protein content remained unaffected (Dewan and Dhingra 2004). In case of Phaseolus vulgaris L. plants supplemented with different Pb and Cd concentrations (2, 4, 6, 8 g Kg-1 for lead and 1.5, 2.0, 2.5, 3.0 g Kg-1 for cadmium),
total soluble sugars, starch content as well as soluble proteins content decreased as concentration of metals was increased in comparison of control plants. However, the total free amino acid content was increased with increasing concentration of heavy metals (Bhardwaj et al. 2009). In case of chickpea grown in As (5mg/kg of dry soil), there was a significant inhibition in the accumulation of seed reserves such as starch, proteins, sugars, and minerals as compared to the controls, which indicated that As application markedly reduced the quality of the chickpea seeds (Malik et al. 2011).
4. Antioxidative Defense mechanisms
Heavy metals cause oxidative stress by damaging the cells and disrupting the cellular homeostasis by the enhanced generation of toxic reactive oxygen species (ROS). The ROS produced during stress are harmful for the plants and can pose a threat to cells by damaging membranes, nucleic acids and chloroplast pigments (Chen and Goldsbrough 1994, Dra,_zkiewicz et al. 2004). The plants possess anti-oxidative systems to protect themselves against the damage produced by ROS. This system is composed of antioxidant enzymes: ascorbate peroxidase (APOX), glutathione reductase (GR), superoxide dismutase (SOD), catalase (CAT) and non-enzymatic compounds (ascorbic acid, glutathione, carotenoids, -tocopherol) (Gill and Tuteja 2010).
4.1 Antioxidative Enzymatic systems
There is evidence that in pea plants exposed to Cd2+,
Kaur & Nayyar : Heavy metal toxicity to food legumes: effects, antioxidative defense and tolerance mechanisms 9
and guaiacol peroxidase activities when treated with CdCl2 (0-50 mM). No significant changes in the glutathione reductase activity were shown by the treated plants (Sandalio et al. 2001). Dixit et al. (2001) also reported the effects of 4 and 40uM cadmium on the antioxidants and antioxidant enzymes in the pea roots and leaves, separately. The results indicated that the levels of lipid peroxidation and H2O2 increased in both the roots and leaves. Activities of SOD, APX, GST and GR were more at 40 uM concentration while GPOX decreased in the roots. Aluminium phytotoxicity causes oxidative stress in developing green gram seedlings and a significant increase in lipid peroxidation, peroxide content accompanied by a decrease in catalase activity (Panda et al. 2003). However, superoxide dismutase, peroxidase and glutathione reductase activities increased with increasing aluminium concentrations. Both the contents of glutathione and ascorbate decreased with the elevated metal concentrations. In another experiment the effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max) were investigated (Cakmak and Horst 1991). Soybean seedlings treated with Al (AlCI3) concentrations ranging from 10 to 75 µM showed the enhancement of lipid peroxidation in the crude extracts of the root tips, the activities of SOD and peroxidase increased while catalase decreased. The effects of cadmium 5 uM and zinc 100 uM on the antioxidant enzyme activities in bean (Phaseolus vulgaris) were reported (Chaoni et al.1997). Lipid peroxidation was enhanced in all plant organs of the plant and the catalase activity was decreased in both roots and leaves but not in stems. Mercury toxicity in alfaalfa (Medicago sativa) by treating the plants with 0–40 µM HgCl2 for 7 d resulted in oxidative stress (Zhou 2007). It was observed that treatment with Hg2+ increased the activities of NADH oxidase and
lipoxygenase (LOX) and damaged the biomembrane lipids. There was enhancement in the total activities of APX, POD and CAT. Several antioxidative metabolites such as ascorbate and glutathione (GSH) differentially accumulated in leaves.
Table:1 Location and function of nonenzymatic antioxidants in plant cell
Antioxidant Location in the plant Function
α –tocopherol Chloroplast envelope, thylakoid membranes and plastoglobuli. Deactivating the photosynthesis-derived ROS and scavenging lipid peroxyl radicals in thylakoid membranes
Ascorbic Acid Usually higher in photosynthetic cells and meristems (and some fruits) and highest in mature leaves with fully developed chloroplast
Powerful water soluble antioxidant, prevent or minimize the damage caused by ROS.
Glutathione Cell compartments like cytosol, ER, vacuole, mitochondria,
chloroplasts, peroxisomes and in apoplast
Role in antioxidative defense, regulation of sulfate transport, signal transduction, detoxification of xenobiotics and the expression of stress-responsive genes
Carotenoids Leaves, fruits and floral parts Photoprotective role in addition to scavenging of ROS.
Proline Cytosol Osmoregulation, seed germination, membrane integrity, inhibition
of water loss and an antioxidant
Salicylic Acid Cytosol Seed germination, stomatal closure, an antioxidant
Flavonoids Leaves, floral parts, and pollens. Role as an antioxidant, flowers, fruits, and seed pigmentation,
protection against UV light, defence against phytopathogens role in plant fertility and germination of pollen.
4.2 Non enzymatic defense systems
The non enzymatic defense systems of the plants include various molecules such as glutathione, proline, -tocopherols, carotenoids and flavonoids. Various antioxidants like cysteine, proline, ascorbic acid and non-protein thiols also play an important role in detoxification of toxic metal ions (Singh and Sinha 2005). Their involvement in defense against metals in described below in legumes. 4.2. a. -tocopherol (Vitamin E)
It is the major vitamin E compound found in leaf chloroplasts, where it is primarly located in the chloroplast envelope, thylakoid membranes and plastoglobuli. (Table.1). It is responsible for deactivating the photosynthesis-derived reactive oxygen species and thus preventing the propagation of lipid peroxidation by scavenging lipid peroxyl radicals in thylakoid membranes. The alpa-tocopherol levels are very sensitive to environmental stresses and change differentially depending on the magnitude of the stress and species-sensitivity to stress. Recently, it has been found that oxidative stress activates the expression of genes responsible for the synthesis of tocopherols in higher plants (Wu et al. 2007). Srivastava et al. (2005) reported a general induction in -tocopherol content in Anabaena doliolum under NaCl and Cu2+ stress. The effects of lead, copper, cadmium and mercury on the content of chlorophyll, proline, retinol, -tocopherol and ascorbic acid were investigated in 17-day-old bean seedlings (Phaseolus vulgaris L.) A significant increase in the levels of -tocopherol was indicated, also increase in proline and ascorbic acid was reported (Zengin and Munzuroglu 2005).
4.2.b. Ascorbic acid (Vitamin C)
1 0 Journal of Food Legumes 26(3 & 4), 2013
tissues, usually being higher in photosynthetic cells and meristems in and some fruits. Its concentration is reported to be highest in mature leaves with fully developed chloroplast and with highest chlorophyll. Usually, ASC remains available in reduced form in leaves and chloroplast under normal conditions (Smirnoff 2000). Studies report that the ASC content of the roots and shoots of two cultivars of pigeonpea (Cajanus cajan) decreased with an increasing concentration (Madhava Rao and Sresty 2000) of Zn and Ni. The effect of ascorbic acid on soybean seedlings grown on medium containing a high concentration of copper were investigated (Golan-Goldhirsh et al. 1995) and reported that ascorbic acid prevented the entry of Cu into the plant roots and did not let copper toxicity signs. There are some reports where a decrease in the ascorbic acid in the roots and nodules of Glycine max under Cd stress has also been observed (Balestrasse et al. 2004). Cd also decreases the ascorbic acid content in the leaves of A. thaliana and P. sativum (Romero-Puertas et al. 2007).
4.2.c. Glutathione
Glutathione is one of the crucial metabolites in plants, which is considered as most important intracellular defense against ROS induced oxidative damage. It generally occurs abundantly in reduced form (GSH) in plant tissues and is mostly localized in all cell compartments like cytosol, endoplasmic reticulum, vacuole, mitochondria, chloroplasts, peroxisomes as well as in apoplast and plays a central role in several physiological processes, including regulation of sulfate transport, signal transduction, conjugation of metabolites, detoxification of xenobiotics and the expression of stress-responsive genes (Mullineaux et al. 2006). GSH plays a key role in the antioxidative defense system by regenerating another potential water soluble antioxidant like ASC, via the ASC-GSH cycle (Foyer and Halliwell 1976). With increase in stress, GSH concentrations usually decline and redox state becomes more oxidized, leading to degradation of the plant system (Tausz et al. 2004). During heavy metal stress, GSH concentration in the cell elevates. For example, increased concentration of GSH has been observed with the increasing Cd concentration in P. sativum (Metwally et al.2005) and V. mungo (Molina et al. 2008). On the other hand, Srivastava et al. (2005) reported an appreciable decline in GR activity and GSH pool under Cu stress in Anabaena dolicum and significantly higher increase under salt stress.
4.2.d. Carotenoids
Carotenoids are the protective pigments that are found in plants and microorganisms. These are lipid soluble antioxidants playing many functions in plant metabolism including oxidative stress tolerance. A decrease in carotenoid and chlorophyll contents in V. mungo plants with increasing Cd concentration was observed (Rai et al. 2004, Singh et al. 2008). In H. vulgare seedlings also, a reduction in carotenoids content was observed under Cd-stress (Demirevska-Kepova et al. 2006).
4.2.e. Proline
Proline accumulation, accepted as an indicator of environmental stress, is also considered to have important protective roles.Some authors have suggested that proline acts as an antioxidant in Cd-stressed cells and thereby improves metal tolerance (Sharma and Dietz 2006, Siripornadulsil et al. 2002). Others have reported that many plants have been reported to accumulate proline (Pro) when exposed to heavy metals (Alia and Saradhi 1991, Talanova et al. 2000). However, the precise mechanism and the functional significance of proline accumulation in plants under heavy metal stress have not been elucidated to date. Costa and Morel (1994) have suggested that proline accumulation in plants under Cd stress is induced by a Cd imposed decrease of the plant water potential. However, proline maintains the water balance under Cd stress thereby suggesting that proline-mediated alleviation of water deficit stress could substantially contribute to the Cd tolerance of the plant. But the direct conclusive evidence as to the role for the water potential in heavy metal-induced proline accumulation is still lacking. A recent study has shown that proline alleviates Cd toxicity by detoxifying ROS, and increasing the activity of SOD and CAT and glutathione content (Xu et al. 2009). Rai (2002) suggests that the possible role of proline against heavy metals is by forming chelates with the metals. There are indications that proline forms a proline-metal complex and protects the activity of glucose-6-phosphate dehydrogenase and nitrate reductase against inhibition by Cd and Zn (Sharma et al.1998). Apart from acting as an heavy metal alleviator, proline also acts as an important cytoplasmic osmoticum, a scavenger of free radicals, source of nitrogen and carbon for post stress growth, a stabilizer of membranes, machinery for protein synthesis and a sink for energy to regulate redox potential (Rai et al. 2004). Muneer et al. (2011) reported that proline content increase at all concentrations of cadmium exposure in Vigna radiata and maximum increase was found at 0.50 mM which was about 119 to 120%. In Cajanus cajanand Vigna mungo there was an accumulation of proline under heavy metals (Co, Cd. Zn and Pb) treatment (Alia and Saradhi 1991). Similar results were observed in case of Cajanus cajan when given aluminium treatment (Bhamburdekar and Chavan 2011). 4.2.f. Phenolics
Salicylic acid is an important signal molecule mediating many biotic and environmental stress-induced physiological responses in plants. The role of SA in regulating Hg-induced oxidative stress was investigated in the roots of alfalfa (Medicago sativa) by Zhou et al. 2006. It was seen that the plants pretreated with 0.2mM SA for 12 h and subsequently exposed to 10 µM Hg2+ for 24 h attenuated toxicity to the root
Kaur & Nayyar : Heavy metal toxicity to food legumes: effects, antioxidative defense and tolerance mechanisms 1 1
commonly found in leaves, floral parts, and pollens. They usually accumulate in the plant vacuole as glycosides, but they also occur as exudates on the surface of leaves and other aerial plant parts. Flavonoids are suggested to have many functions in flowers, fruits, and seed pigmentation, protection against UV light, defence against phytopathogens (pathogenic microorganisms, insects, animals), role in plant fertility and germination of pollen and, molecules in plant-microbe interaction. Apart from the above roles, flavonoids have as antioxidative activity (Brown et al.1998). Besides having the function of ROS scavenging , flavonoids are able to function as chelators for metals, depending on the molecular structure (Brown et al.1998) and hence can take part in plant defence. In Arabidopsis thaliana, the relation between flavonoids and heavy metal tolerance were investigated. Both Arabidopsis wild type and mutant lines with a defect in flavonoid biosynthesis were grown on media containing different heavy metals. Results revealed that root length and seedling weight were reduced in mutants more than in the wild type when grown on cadmium, while on zinc only root length was affected (Keilig and Muller 2009).
5. Tolerance mechanisms
Heavy metals in the plant environment operate as stress factors that cause physiological strain and in doing so they reduce the plant vigour and totally inhibit the plant growth in extremes However, plants have evolved several physiological mechanisms which enable them to tolerate metal toxicity (Baker 1987). The development of metal tolerance in plants is a major way to reduce the harmful effects of excessive exposure to heavy metal ions (Tyler et al.1989). There are various potential cellular and other mechanisms available for metal detoxification and tolerance in higher plants (Hall 2002), which have been reported to function in legumes.
5.a. Role of Arbuscular mycorrhizal fungi
Mycorrhizal association is a symbiotic non-pathogenic relationship between plant roots and fungal hyphae with a fungal connection between the soil and the root (Harley and Smith 1983). It has been reported that the host plant receives support from AM fungi, with the help of its symbiotic association, in the aspect of uptake of phosphorus and other nutrients, enhancement of growth hormones, increase of protein content, increase of lipid, sugars, amino acid levels, increase of tolerance to heavy metals, increase of salinity tolerance, and resistance to root-borne pathogens (Upadhyaya et al. 2010). Recently, the symbiotic association with mycorrhizal fungi has been proposed as one of the major mechanisms of plant HM-tolerance (Hall 2002, Joachim et al. 2009). However, alleviating heavy metal toxicity by AMF colonization can vary to a large extent, depending on which heavy metal is involved, its concentration in the soil, the fungal symbiosis partner and the conditions of plant growth (Turnau
1998).
There are many strategies adopted by AM which can alleviate heavy metal threats in mixed culture systems and, thus, from the food chains (Joschim et al. 2009). These include the immobilisation of metal compounds, precipitation of p o l y p h o s p h a t e granules in the soil, adsorption to chitin in the fungal cell walls and chelation of
heavy metals inside the fungus (Joachim et al. 2009). Generally, AM binds to heavy metals beyond the plant rhizosphere by releasing an insoluble glycoprotein commonly known as glomalin (Gonzalez- Chavez et al. 2004). The roles of AM are summarized in the Fig.5.
Numerous studies have indicated that AMF can decrease the metal uptake of the host plants, thus protecting them against HMs toxicity (Leyval et al. 1997). Many heavy metal contaminated sites are reported to have mycorrhizae (Weissenhorn and Leyval 1993).This indicates that these fungi have evolved a HM-tolerance and that they may play a role in the phytoremediation of the site (Khan et al. 2000). Mycorrhizae were found to ameliorate the toxicity of trace metals in polluted soils growing in soybean and lentil plants (Jamal et al. 2002). Increased heavy metal tolerance of plants by dual inoculation of an arbuscular mycorrhizal fungi and nitrogen-fixer Rhizobium bacterium was reported in cowpea (Al-Garni 2006). The effects of dual inoculation with arbuscular mycorrhizal (AM) fungus and Rhizobium (N-fixing bacteria, NFB) on the host plant cowpea Vigna sinensis in pot cultures were investigated at six concentrations of Zn (0.0-1000 mg/kg dry soil) and Cd (0.0-100 mg/kg dry soil). The study provides evidence for benefits of NFB to AM fungi in the protection of host plants against the detrimental effects of heavy metals and provides the mechanisms for metal tolerance against them.A greenhouse pot experiment was done to investigate the effects of the colonization of arbuscular mycorrhizal fungus (AMF) Glomus mosseae on the growth and metal uptake of three leguminous plants (Sesbania rostrata, Sesbania cannabina, Medicago sativa) grown in multi-metal contaminated soil (Lin et al. 2007). The results revealed that AMF colonization increased the growth of the legumes thereby indicating that AMF colonization increased the plant’s resistance to heavy metals. The effect was also enhanced on the formation of root nodules and N and P uptake increased, which may be due to the heavy metal tolerance mechanisms conferred by the AMF.
1 2 Journal of Food Legumes 26(3 & 4), 2013
5.b. Role of metallothioneins and phytochelatins
Metallothioneins (MT’s) belong to a family of cysteine-rich low molecular weight metal-binding proteins generally induced during the metal stress (Corbett and Goldsbrough 2002). Metallothioneins generally form complexes with heavy metal ions and are present in almost all forms of life and have a role in protecting cells from the deleterious effects of high concentration metal ions. The function of MT is to detoxify non-essential metals such as mercury, cadmium and essential metals such as zinc and copper.
Phytochelatins (PCs), a type of MT’s, are synthesized in plants in response to heavy metal stress and due to various metals. PCs consist of only the three amino acids: Glu, Cys and Gly, the Glu, and Cys residues linked through a g-carboxylamide bond(Cobbet 2000).Recent research indicates that PCs are present in a wide variety of plant species and in some microorganisms. They are structurally related to glutathione (GSH, g-Glu-Cys-Gly) and were presumed to be the products of a biosynthetic pathway. In addition, a number of structural variants, for example, Glu-Cys)n-b- Ala, (g-Glu-Cys)n-Ser, and (g-Glu-Cys)n-Glu, have been identified in some plant species (Rauser 1999, Zenk 1996).
Activation of the detoxicative-phytochelatin system was observed in the cytosol of root cells of three legume species, Vicia faba, Pisum sativum, and Phaseolus vulgaris when they were exposed to lead ions (Piechalak et al. 2002). This system was composed of phytochelatins (PCs) in roots of V. faba, homophytochelatins (hPCs) in P. vulgaris roots, and both PCs and hPCs in P. sativum roots.
5.c. Organic acids and amino acids
Some amino acids, particularly histidine and proline, also play very important roles in the chelation of metal ions both within plant cells and in the xylem sap (Rai 2002). Kerkeb and Krämer (2003)have reported that in Alyssum lesbiacum and Brassica juncea , an enhanced release of Ni into the xylem is associated with concurrent release of histidine from an increased root free His pool. Other amino acids such as citrate, malate and histidine are potential ligands for heavy metals and could play a role in tolerance and detoxification (Rauser 1999). Citrate, malate and oxalate have been involved in transport of metal ions through the xylem and vacuolar sequestering (Rauser 1999). It is reported that citric acid to be a major Cd2+ ligand at low Cd2+ concentrations (Wagner 1993)
and has been shown to form complexes with Ni2+ in
Ni-hyperaccumulation plants (Sagner et al. 1998). It is also suggested that malate is a cytosolic zinc chelator in zinc-tolerant plants (Mathys 1977). Kramer et al.(1996) reported that the significant and proportional change in amino acid or organic acid concentration elicited by a change in metal exposure was shown by histidine response in plants that accumulate nickel. The presence of different concentrations of organic acids among various ecotypes of metal-tolerant
plants in their natural habitat has deemed these substances as likely cellular chelators (Rauser 1999).
5.d. Polyamines
Polyamines (PAs) are nitrogenous compounds present in all living cells. They are not only involved in various cellular processes like growth promotion and cell division but also in the inhibition of ethylene production and senescence (Tiburcio et al.1997). They influence a variety of growth and development processes in plants which have been suggested to be a class of plant growth regulators and to act as second messengers (Evans and Malmberg 1989, Kakkar and Sawhney 2002). The polyamines are cations due to protonation at cytoplasmic pH, i.e. putrescine2+, spermidine3+, and spermine4+,
which accounts for their binding ability to nucleic acids (Flink and Pettijohn 1975).It has been reported that the levels of polyamines and the activities of their biosynthetic enzymes in plants increase under environmental stresses (Evans and Malmberg 1989). Polyamine contents are highly altered in response to the exposure to heavy metals. For example, the response of different polyamines to Cd treatment strongly varied in Phaseolus vulgaris in an organ-specific manner. Putrescine increased in root, hypocotyl, and epicotyl whereas spermidine increased in hypocotyl, decreased in leaves, and did not change in roots.In soybean phospholipids, using membrane vesicles (Weinstein et al. 1986). Tadolini et al. (1984) showed that polyamines inhibit lipid peroxidation when bound to the negative charges on the vesicle surface. In addition, polyamines namely, spermine, spermidine, putrescine, and cadaverine have been demonstrated to scavenge free radicals in vitro (Drolet et al.1986). Furthermore, polyami