(Skripsi)
Oleh Adek Purnawati
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS LAMPUNG
THE PRODUCTION OF CGT-ase FROM LOCAL ISOLATE’S AMYLOLITIC BACTERIA STRAIN LTi-21-3
By
Adek Purnawati
Microorganisms are potential resources for the production of various enzymes as well as its application on industries. Enzyme is a functional protein that enable to bind and catalyze a substrate biochemically into new compounds or products. In this research, a local amylolytic isolates named as LTi-21-3, which has been isolated by a previous researcher was used as ase producing bacteria. CGT-ase enzymes are enzymes that enable to convert starch into cyclodextrins. Optimization of CGT-ase enzyme production was carried out by varying the carbon’ssource (soluble starch, corn starch, sweet potato starch, sago starch, and
the addition of maltose), nitrogen’ssources (yeast extract, urea and NH4Cl), metal ion’ssources (CuSO4, ZnSO4 , FeSO4, and CaCl2) and the pH (on pH 7, 8, 9 and 11) of the modified culture medium Horikoshi's II. The results showed that the optimum conditions for production of the enzyme was on a culture medium containing cassava starch, NH4Cl, MgSO4 at pH 8 with optimum incubation time for 36 hours. In these conditions were obtained the value of the unit and specific activity are 546,86 U/mL and 595,72 U/mg, respectively. The specific activity on this condition was approximately 253% higher than that of the modified Horikoshi's II medium without treatment.
TERHADAP PRODUKSI ENZIM CGT-ase
DARI BAKTERI AMILOLITIK ISOLAT LOKAL LTi-21-3
Oleh
Adek Purnawati
Mikroorganisme merupakan sumber yang potensial sebagai bahan baku untuk produksi enzim. Saat ini, enzim dalam bentuk hasil isolasi maupun langsung dari mikroorganisme dapat dimanfaatkan dalam berbagai bidang industri. Enzim merupakan protein fungsional yang dapat berikatan dengan suatu substrat spesifik untuk mengkatalisasi reaksi biokimia menjadi suatu senyawa baru yang disebut produk. Pada penelitian ini, digunakan bakteri amilolitik isolat lokal LTi-21-3 yang telah berhasil diisolasi oleh peneliti sebelumnya, yang merupakan bakteri penghasil enzim CGT-ase. Enzim CGT-ase yaitu enzim ekstraseluler yang dapat mengubah pati menjadi siklodekstrin. Optimasi produksi enzim CGT-ase ini dilakukan dengan cara memvariasikan: sumber karbon (pati soluble, pati jagung,
pati ubi jalar, pati sagu, dan penambahan maltosa), sumber nitrogen (ekstrak ragi, urea dan NH4Cl), sumber ion logam (CuSO4, ZnSO4, FeSO4 dan CaCl2) dan variasi pH (7, 8, 9 dan 11) pada medium kultur Horikoshi’s II termodifikasi.
Hasil penelitian yang diperoleh menunjukkan bahwa kondisi optimum untuk produksi enzim CGT-ase dari isolat LTi-21-3 yaitu pada medium kultur yang mengandung pati singkong, NH4Cl, MgSO4 pada pH 8 dengan waktu inkubasi optimum ada pada 36 jam. Pada kondisi tersebut diperoleh nilai aktivitas unit dan spesifik secara berurutan yaitu 546,86 U/mL dan 595,72 U/mg. Persentase aktivitas spesifik pada kondisi optimum ini sekitar 253% lebih tinggi jika
dibandingkan dengan medium Horikoshi’s II termodifikasi tanpa perlakuan.
1. Tim Penguji
Ketua :Mulyono, Ph.D. …………....
Sekretaris :Dra. Aspita Laila, M.S. …………....
Penguji
Bukan Pembimbing :Prof. Dr. Tati Suhartati, M.S. …………....
2. Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam
Prof. Suharso, Ph.D.
NIP 196905301995121001
Jadilah kamu manusia yang pada kelahiranmu semua orang
tertawa bahagia, tetapi hanya kamu sendiri yang menangis; dan
pada kematianmu semua orang menangis sedih, tetapi hanya kamu
sendiri yang tersenyum.
(Mahatma Gandhi)
Kebanyakan dari kita tidak mensyukuri apa yang sudah kita
miliki, tetapi kita selalu menyesali apa yang belum kita capai.
(Schopenhauer)
Ilmu itu lebih baik daripada harta. Ilmu akan menjaga engkau
dan engkau menjaga harta. Ilmu itu penghukum (hakim)
sedangkan harta terhukum. Kalau harta itu akan berkurang
apabila dibelanjakan, tetapi ilmu akan bertambah apabila
dibelanjakan.
(Sayidina Ali bin Abi Thalib)
Boleh jadi kamu membenci sesuatu padahal ia amat baik bagimu
dan boleh jadi kamu menyukai sesuatu padahal ia amat buruk
bagimu. Allah mengetahui sedang kamu tidak mengetahui.
Dengan penuh rasa syukur dan bangga kupersembahkan karyaku ini sebagai bukti rasa cintaku kepada:
Allah SWT yang telah memberiku kemudahan dalam penyelesaian karya sederhana ini,
Kedua orang tuaku Bapak Mangsuri dan Ibu Sunarsih yang sangat ku cinta dan ku sayangi,
Kakak-kakakku tersayang
Siti Khotimah, Muhammad Khozin, Ahmad Jaelani dan Nita Komalasari,
Segenap Keluarga besarku yang selalu mendoakan keberhasilanku,
Sahabat dan teman-temanku yang selalu berbagi kebahagiaan,
Seseorang yang kelak akan mendampingiku,
I. PENDAHULUAN
A. Latar Belakang
Perkembangan ilmu dan teknologi serta pendayagunaan enzim semakin
meningkat dilihat dari segi diversifikasi maupun skala penggunaannya. Saat
ini enzim dalam bentuk hasil isolasi maupun langsung dari mikroorganisme
dapat digunakan dalam bidang produksi pangan. Selain itu, dapat juga
digunakan pada produksi antibiotika, vitamin, asam-asam organik, etanol,
tekstil, kertas, industri kulit, analisis biokimia, kedokteran dan untuk
penanganan limbah industri (Judoamidjojo et al., 1989).
Enzim merupakan protein khusus yang dapat bergabung dengan suatu substrat
spesifik untuk mengkatalisasi reaksi biokimia dari substrat tersebut (Maier
et al.,2000). Dalam reaksi tersebut enzim mengubah senyawa yang disebut
substrat menjadi bentuk suatu senyawa baru yang disebut produk. Enzim
memiliki substrat spesifik dan reaksi kimia yang spesifik untuk dikatalisnya
(Palmer, 1985). Aktivitas enzim dipengaruhi oleh suhu, pH dari lingkungan
tempat enzim bekerja, konsentrasi substrat, aktivator dan inhibitor enzim.
Pada penelitian ini, digunakan bakteri amilolitik yang telah berhasil diisolasi
oleh peneliti sebelumnya yaitu bakteri amilolitik isolat lokal LTi-21-3 yang
merupakan mikroorganisme yang mampu memecah pati menjadi senyawa yang
lebih sederhana. Kebanyakan mikroorganisme amilolitik tumbuh subur pada
bahan pangan yang banyak mengandung pati atau karbohidrat, misalnya pada
berbagai jenis tepung. Siklodekstrin pada umumnya diproduksi dari pati
menggunakan enzim CGT-ase (Leeet al.,1992).
Siklodekstrin merupakan oligosakarida non pereduksi produk pati dengan
struktur kimia berbentuk cincin, dan terbentuk melalui proses siklisasi oleh
aktivitas CGT-ase (Cyclodextrin Glycocyltransferase) (Szejtli, 1988; Schmid et al., 1989; Tonkova, 1998). Berdasarkan jumlah glukosa yang menyusunnya
siklodekstrin dibedakan atas α-siklodekstrin (6 unit glukosa), β-siklodekstrin
(7 unit glukosa), dan γ-siklodekstrin (8 unit glukosa) (Szejtli, 1988; Tonkova,
1998).
Siklodekstrin dapat digunakan dalam berbagai industri, seperti pada industri
kimia, farmasi, pangan dan kosmetika. Hal ini karena siklodekstrin
mempunyai sifat enkapsulasi, termasuk peningkatan kelarutan dan
perlindungan komponen kimia yang labil dari pengaruh oksidasi (Laga, 2001).
Produk siklodekstrin yang dihasilkan dipengaruhi oleh jumlah amilosa dalam
pati (Whistleret al., 1984).
Untuk memproduksi enzim yang memiliki aktivitas yang tinggi, maka perlu
diperhatikan faktor-faktor yang penting seperti suhu, substrat, dan pH yang
digunakan pada medium kultur (Suriawiria, 1990). Dalam penelitian ini, telah
LTi-21-3 dengan cara memvariasikan sumber karbon, nitrogen, ion logam dan
pH pada medium kultur.
Dengan ini diharapkan dapat diketahui kondisi yang optimum bagi bakteri
amilolitik isolat lokal LTi-21-3 untuk menghasilkan enzim CGT-ase yang
dapat mengkatalisis pembentukan siklodekstrin.
B. Tujuan Penelitian
Tujuan dari penelitian ini adalah :
1. Mempelajari pengaruh pemberian variasi sumber karbon, nitrogen, ion
logam dan pH pada medium kultur terhadap bakteri amilolitik isolat lokal
LTi-21-3 dalam memproduksi enzim CGT-ase.
2. Mengetahui kondisi kultur yang optimum dari bakteri amilolitik isolat lokal
LTi-21-3 untuk memproduksi enzim CGT-ase yang memiliki aktivitas unit
dan spesifik yang tinggi.
C. Manfaat Penelitian
Manfaat yang dapat diperoleh dari penelitian ini antara lain yaitu mengetahui
kondisi yang optimum dari bakteri amilolitik isolat lokal LTi-21-3 dalam
memproduksi enzim CGT-ase yang dapat mengkatalisis pembentukan
siklodekstrin. Siklodekstrin yang dihasilkan dapat dilakukan penelitian
II. TINJAUAN PUSTAKA
A. Mikroorganisme
Populasi mikroorganisme yang ada di alam sekitar kita ini sangatlah besar dan
cukup kompleks. Beratus spesies mikroba menguasai setiap bagian tubuh kita.
Mereka terdapat dalam jumlah yang cukup besar. Sebagai contoh, sekali kita
bersin dapat menebarkan beribu- ribu mikroorganisme. Satu gram tinja dapat
mengandung jutaan bakteri. Alam di sekitar kita, baik itu tanah, air, maupun
udara juga dihuni oleh kumpulan mikroorganisme. Penelitian yang layak
mengenai mikroorganisme dalam berbagai habitat ini memerlukan teknik untuk
memisahkan populasi campuran yang rumit ini, atau yang biasanya dikenal
dengan istilah biakan campuran, menjadi spesies yang berbeda- beda yang
dikenal dengan istilah biakan murni. Biakan murni ini terdiri dari satu populasi
sel yang semuanya berasal dari satu sel induk (Pelczar, 1986).
Mikroorganisme dapat ditumbuhkan dan dikembangkan pada suatu substrat
yang disebut medium. Medium yang digunakan untuk menumbuhkan dan
mengembangbiakkan mikroorganisme tersebut harus sesuai susunannya
dengan kebutuhan jenis-jenis mikroorganisme yang bersangkutan. Beberapa
mikroorganisme dapat hidup baik pada medium yang sangat sederhana yang
gula. Sedangkan mikroorganisme lainnya memerlukan suatu medium yang
sangat kompleks yaitu berupa medium ditambahkan darah atau bahan-bahan
kompleks lainnya (Volk dan Wheeler,1993).
Mikroorganisme adalah sumber yang potensial sebagai bahan baku untuk
produksi enzim. Hal ini disebabkan karena beberapa faktor yaitu: ekonomis,
karena dapat dihasilkan dalam waktu yang cukup pendek dan media yang
cukup murah, kondisi reaksi seperti pH dan temperatur mudah diatur, dan
peningkatan produksi enzimnya dapat dikondisikan dengan cara penambahan
induser tertentu (Wang, 1979).
B. Bakteri
Bakteri, berasal dari kata Latinbacterium, adalah kelompok raksasa dari
organisme hidup. Ukuran bakteri sangatlah kecil (mikroskopik) dan
kebanyakan uniselular, dengan struktur sel yang relatif sederhana tanpanucleus
(inti sel),cytoskeleton, dan organel lain seperti mitokondria dan kloroplas.
Bakteri digolongkan menjadi dua, yaitu bakteri yang menguntungkan dan
bakteri yang merugikan. Bakteri tersebar di tanah, air, dan sebagai simbiosis
dari organisme lain. Bakteri rata-rata mengandung 40-70% protein, 13-34%
asam nukleat, 10-30% lipid (angka-angka ini menunjukkan persen berat
kering) (Suhartono, 1989).
Ada tiga bentuk dasar bakteri, yaitu bentuk bulat atau kokus, bentuk batang
silindris, bentuk lengkung atau vibri. Bentuk bulat atau kokus dapat dibedakan
stafilokokus. Bakteri berbentuk batang dapat dibedakan ke dalam bentuk
batang panjang dan batang pendek dengan ujung datar atau lengkung. Bakteri
berbentuk lengkung dapat dibagi menjadi bentuk koma (vibrio), jika
lengkungnya kurang dari setengah lingkaran. Bentuk bakteri dipengaruhi oleh
umur dan syarat pertumbuhan tertentu (Hidayatet al., 2006).
1. Nutrien untuk Pertumbuhan Bakteri
Bakteri memerlukan nutrien dengan komposisi tertentu untuk tumbuh dan
membelah diri. Kebutuhan nutrien meliputi unsur makroesensial dan unsur
mikroesensial. Unsur makro digunakan oleh mikroba dalam metabolisme
sel, sedangkan unsur mikro digunakan untuk mengaktifkan enzim
(Suhartono, 1989).
Organisme hidup memerlukan nutrisi untuk pertumbuhannya. Substansi
kimia organik dan inorganik diperoleh dari lingkungan dalam berbagai
macam bentuk. Nutrien diambil dari lingkungan kemudian
ditransformasikan melalui membran plasma menuju sel. Di sel beberapa
nutrisi diolah menghasilkan energi yang digunakan dalam proses seluler
(Lim, 1998).
Jasad renik heterotrof membutuhkan nutrien untuk kehidupan dan
pertumbuhannya, yakni sebagai: (1) sumber karbon, (2) sumber nitrogen,
(3) sumber energi, (4) dan faktor pertumbuhan, yakni mineral dan vitamin.
Nutrien tersebut dibutuhkan untuk membentuk energi dan menyusun
komponen-komponen sel. Setiap jasad renik bervariasi dalam
Ada tujuh komponen utama yang dibutuhkan semua makhluk hidup, yaitu
karbon, oksigen, nitrogen, hidrogen, fosfor, sulfur dan kalium. Untuk
kebutuhan akan sumber karbon dipenuhi oleh adanya gula, pati, serta
karbohidrat lainnya. Ada dua macam mikronutrien yakni mikronutrien
organik dan mikronutrien anorganik. Zat-zat yang bertindak sebagai
mikronutrien organik pada beberapa asam amino (triptofan) dan pada
beberapa komponen-komponen DNA dan RNA (purin dan pirimidin).
Beberapa unsur logam yang termasuk dalam mikronutrien anorganik adalah
Co, Mo, Cu, Zn. Unsur logam ini sangat diperlukan untuk kehidupan sel
meskipun jumlahnya sangat sedikit (Irianto, 2006).
Menurut Waluyo (2005), peran utama nutrien adalah sebagai sumber energi,
bahan pembangun sel, dan sebagai akseptor elektron dalam reaksi
bioenergetik (reaksi yang menghasilkan energi). Oleh karenanya bahan
makanan yang diperlukan terdiri dari air, sumber energi, sumber karbon,
sumber akseptor elektron, sumber mineral, faktor pertumbuhan, dan
nitrogen. “Selain itu, secara umum nutrien dalam media pembenihan harus
mengandung seluruh elemen yang penting untuk sintesis biologik organisme
baru (Jawetz, 2001).”
a. Air
Air merupakan komponen utama sel mikroba dan medium. Fungsi air
adalah sebagai sumber oksigen untuk bahan organik sel pada respirasi.
Selain itu air berfungsi sebagai pelarut dan alat pengangkut dalam
b. Sumber Energi
Ada beberapa sumber energi untuk mikroba yaitu senyawa organik atau
anorganik yang dapat dioksidasi dan cahaya terutama cahaya matahari.
c. Sumber Karbon
Sumber karbon untuk mikroba dapat berbentuk senyawa organik maupun
anorganik. Senyawa organik meliputi karbohidrat, lemak, protein, asam
amino, asam organik, garam asam organik, polialkohol, dan sebagainya.
Senyawa anorganik misalnya karbonat dan gas CO2yang merupakan
sumber karbon utama terutama untuk tumbuhan tingkat tinggi.
d. Sumber Akseptor Elektron
Proses oksidasi biologi merupakan proses pengambilan dan pemindahan
elektron dari substrat. Karena elektron dalam sel tidak berada dalam
bentuk bebas, maka harus ada suatu zat yang dapat menangkap elektron
tersebut. Penangkap elektron ini disebut akseptor elektron. Akseptor
elektron ialah agensia pengoksidasi. Pada mikrobia yang dapat berfungsi
sebagai akseptor elektron ialah O2, senyawa organik, NO3-, NO2-, N2O,
SO42-, CO2, dan Fe3+.
e. Sumber Mineral
Mineral merupakan bagian dari sel. Unsur penyusun utama sel ialah C,
O, N, H, dan P. Unsur mineral lainnya yang diperlukan sel ialah K, Ca,
Mg, Na, S, Cl. Unsur mineral yang digunakan dalam jumlah sangat
sedikit ialah Fe, Mn, Co, Cu, Bo, Zn, Mo, Al, Ni, Va, Sc, Si, Tu, dan
jumlah besar disebut unsur makro, dalam jumlah sedang unsur oligo, dan
dalam jumlah sangat sedikit unsur mikro. Unsur mikro sering terdapat
sebagai ikutan (impurities) pada garam unsur makro, dan dapat masuk ke
dalam medium lewat kontaminasi gelas tempatnya atau lewat partikel
debu. Selain berfungsi sebagai penyusun sel, unsur mineral juga
berfungsi untuk mengatur tekanan osmosis, kadar ion H+(kemasaman,
pH), dan potensial oksidasi reduksi (redox potential) medium.
f. Faktor Tumbuh
Faktor tumbuh ialah senyawa organik yang sangat diperlukan untuk
pertumbuhan (sebagai prekursor, atau penyusun bahan sel) dan senyawa
ini tidak dapat disintesis dari sumber karbon yang sederhana. Faktor
tumbuh sering juga disebut zat tumbuh dan hanya diperlukan dalam
jumlah sangat sedikit. Berdasarkan struktur dan fungsinya dalam
metabolisme, faktor tumbuh digolongkan menjadi asam amino, sebagai
penyusun protein; basa purin dan pirimidin, sebagai penyusun asam
nukleat; dan vitamin sebagai gugus prostetis atau bagian aktif dari enzim.
g. Sumber Nitrogen
Mikroba dapat menggunakan nitrogen dalam bentuk amonium, nitrat,
asam amino, protein, dan sebagainya. Jenis senyawa nitrogen yang
digunakan tergantung pada jenis jasadnya. Beberapa mikroba dapat
menggunakan nitrogen dalam bentuk gas N2(zat lemas) udara. Mikroba
Untuk mendapatkan bakteri yang potensial dilakukan dengan penapisan
mikroorganisme dari lingkungan. Umumnya isolat bakteri yang diperoleh
sesuai dengan lingkungan tempat hidupnya. Hal ini dikarenakan kondisi
lingkungan tersebut biasanya digunakan bakteri sebagai substrat utamanya.
Misalnya, bakteri amilolitik dapat diisolasi dari sampel yang berasal dari
limbah pengolahan singkong, limbah sayur (Chakrabarty and Sen, 1984),
rumen (Freer, 1993), serta hasil fermentasi ikan dan bahan makanan dari beras
(Olympiaet al., 1995).
2. Fase Pertumbuhan Bakteri
Suatu mikroorganisme mempunyai siklus pertumbuhan tertentu tergantung
produk yang akan dihasilkan. Fase pertumbuhan bakteri dapat dibagi
menjadi 4 fase, yaitu fase lag, fase logaritma (eksponensial), fase stasioner
dan fase kematian. Fase lag merupakan fase penyesuaian bakteri dengan
lingkungan yang baru. Lama fase lag pada bakteri sangat bervariasi,
tergantung pada komposisi media, pH, suhu, aerasi, jumlah sel pada
inokulum awal dan sifat fisiologis mikroorganisme pada media sebelumnya.
Ketika sel telah menyesuaikan diri dengan lingkungan yang baru maka sel
mulai membelah hingga mencapai populasi yang maksimum. Fase ini
disebut fase logaritma atau fase eksponensial.
Fase eksponensial ditandai dengan terjadinya periode pertumbuhan yang
cepat. Variasi derajat pertumbuhan bakteri pada fase eksponensial ini
sangat dipengaruhi oleh sifat genetik yang diturunkannya. Selain itu,
inkubasi, kondisi pH dan aerasi. Ketika derajat pertumbuhan bakteri telah
menghasilkan populasi yang maksimum, maka akan terjadi keseimbangan
antara jumlah sel yang mati dan jumlah sel yang hidup.
Fase stasioner terjadi pada saat laju pertumbuhan bakteri sama dengan laju
kematiannya, sehingga jumlah bakteri keseluruhan akan tetap.
Keseimbangan jumlah keseluruhan bakteri ini terjadi karena adanya
pengurangan derajat pembelahan sel. Hal ini disebabkan oleh kadar nutrisi
yang berkurang dan terjadi akumulasi produk toksik sehingga mengganggu
pembelahan sel. Fase stasioner ini dilanjutkan dengan fase kematian yang
ditandai dengan peningkatan laju kematian yang melampaui laju
pertumbuhan, sehingga secara keseluruhan terjadi penurunan populasi
bakteri (Volk dan Wheeler, 1993).
C. Amilum
Amilum atau dalam bahasa sehari-hari disebut pati terdapat pada umbi, daun,
batang dan biji-bijian (Poedjiadi, 1994). Pati (C6H10O5)nadalah homopolimer
glukosa dengan ikatan α-glikosidik yang merupakan rantai gula yang panjang. Sifat berbagai macam pati tidak sama, tergantung dari panjang rantai C dan
cabang rantai molekul. Pati terdiri dari dua fraksi yang dapat dipisahkan
dengan air panas. Fraksi terlarut disebut amilosa dan fraksi tidak larut disebut
amilopektin (Winarno, 1986). Berbagai macam pati ditemukan di alam karena
dapat disintesis oleh berbagai macam tumbuhan. Setiap macam pati memiliki
untuk membedakan berbagai pati alamiah. Pati terbagi menjadi dua golongan
yaitu pertama adalah amilosa (15-20%) yang merupakan rantai panjang tidak
bercabang yang terdiri dari molekul-molekul α-D-glukopiranosa yang
bersambungan dengan ikatan α-1,4. Amilosa terdiri atas 250-300 unit D-glukosa yang terikat dengan ikatan α-1,4-glikosidik, jadi molekulnya
merupakan rantai terbuka. Kedua adalah amilopektin (80-85%) yang
merupakan rantai bercabang sebanyak 20-30 molekul α-D-glukopiranosa yang
bersambungan dengan ikatan α-1,4 dan α-1,6. Adanya ikatan α-1,6-glikosidik
ini menyebabkan terjadinya cabang, sehingga molekul amilopektin berbentuk
rantai terbuka dan bercabang (Poedjiadi, 1994). Struktur amilosa dan
amilopektin dapat dilihat pada Gambar 1.
(B) Amilopektin
Gambar 1.Struktur kimia dari (A) Amilosa dan (B) Amilopektin
Amilum dapat dihidrolisis sempurna dengan menggunakan asam sehingga
menghasilkan glukosa. Hidrolisis juga dapat dilakukan dengan bantuan enzim
amilase. Pada reaksi hidrolisis parsial, amilum terpecah menjadi
molekul-molekul yang lebih kecil yang dikenal dengan nama dekstrin. Jadi dekstrin
adalah hasil antara pada proses hidrolisis amilum sebelum terbentuk maltosa
(Poedjiadi, 1994). Untuk mengetahui adanya pati maka dilakukan pengujian
dengan menggunakan larutan iodium (I2dalam KI). Bila terdapat amilosa,
polimer-polimer glukosa yang lebih besar dari 20 maka akan menghasilkan
warna biru. Bila polimer-polimer glukosa kurang dari 20 maka akan
menghasilkan warna merah. Dekstrin dengan polimer enam, tujuh, dan
delapan akan memberikan warna coklat. Polimer yang lebih kecil dari lima
tidak memberikan warna dengan iodium (Winarno, 1986).
Dalam kehidupan manusia amilum berperan sebagai sumber makanan
penghasil energi utama dari golongan karbohidrat, di samping itu amilum juga
dapat berperan sebagai bahan aditif pada proses pengolahan makanan,
misalnya sebagai penstabil dalam proses pembuatan puding. Amilum juga
berperan dalam pembuatan sirup dan pemanis buatan seperti sakarin. Dalam
bidang non makanan, amilum digunakan untuk bahan baku dalam proses
pembuatan kertas, pakaian dari katun, industri cat, maupun untuk produksi
hidrogen. Tabel 1. di bawah ini menunjukkan peran amilum di berbagai bidang
Tabel 1.Penggunaan amilum di bidang industri (Liu, 2005)
Jenis Industri Penggunaan Amilum/Amilum Termodifikasi
Makanan
Bahan Perekat Kertas dan Papan Tekstil
Pengental, pelapis makanan, film makanan
Pembuat lem
Kertas penjilid, pembungkus, pengepak Dalam prosessizing,finishingdan printing
Kapsul obat, bahan pelarut obat Modifikasi pengental
Surfaktan, bahan pensuspensi, bahan pemutih, aktivator pemutih
Pembungkus biji, pembungkus pestisida benang pintal
Pembungkus makananfiller
Bedak dantalk
Flokulan
Scaffold,plasma eksterder, produk
absorben untuk sanitasi
D. Enzim
Enzim merupakan katalisator protein yang mempercepat reaksi kimia dalam
makhluk hidup atau dalam sistem biologik. Sebagai protein, enzim memiliki
sifat-sifat umum protein, seperti enzim terdenaturasi pada suhu tinggi atau
kondisi ekstrim lainnya. Beberapa oksidator, keadaan polaritas larutan,
tekanan osmotik yang abnormal juga dapat menghambat kerja enzim
Menurut Page (1997), sebagai katalis enzim adalah satu-satunya dibanding
dengan katalis-katalis anorganik atau organik sederhana. Sifat-sifat katalitik
dari enzim termasuk hal-hal berikut:
1. Enzim meningkatkan laju reaksi pada kondisi biasa (fisiologik) dari
tekanan, suhu, dan pH. Hal ini merupakan keadaan yang jarang dengan
katalis-katalis lain.
2. Enzim berfungsi dengan selektivitas dan spesifisitas bertingkat luar biasa
tinggi terhadap reaktan yang dikerjakan dan jenis reaksi yang
dikatalisasikan.
3. Enzim memberikan peningkatan laju reaksi yang luar biasa dibanding
dengan katalis biasa.
Peranan enzim sebagai biokatalisator dalam industri semakin meningkat seiring
dengan pesatnya perkembangan industri, khususnya industri makanan,
minuman, industri tekstil, industri kulit dan industri kertas. Hal ini disebabkan
karena enzim bersifat sangat spesifik dibandingkan dengan katalis anorganik.
Selain itu, enzim bekerja sangat efisien, bekerja pada pH yang relatif netral dan
suhu yang relatif rendah, aman, mudah dikontrol, dapat menggantikan bahan
kimia yang berbahaya, dan dapat didegradasi secara biologis (Page, 1997).
Klasifikasi enzim secara internasional berdasarkan atas reaksi yang dikatalisis
antara lain:
1. Oksidoreduktase, mengkatalisis berbagai macam reaksi oksidasi reduksi
serta sering menggunakan koenzim seperti NAD, NADP, FAD, atau lipoleat
2. Transferase, mengkatalisis berbagai jenis transfer kelompok seperti
aminotransferase, karnitin asil transferase, dan transkarboksilase.
3. Hidrolase, mengkatalisis pembelahan ikatan antara karbon dan beberapa
atom lain dengan adanya penambahan air.
4. Liase, mengkatalisis pemecahan ikatan karbon-karbon, karbon sulfur dan
karbon nitrogen tertentu.
5. Isomerase, mengkatalisis isomer optik dan geometrik dan oksidasi reduksi
intramolekuler tertentu.
6. Ligase, mengkatalisis pembentukan ikatan antara karbon dan oksigen
(Lehninger, 1997).
Semua enzim adalah protein, dan aktivitas katalitiknya bergantung kepada
integritas strukturnya sebagai protein. Enzim seperti protein lain, mempunyai
berat molekul berkisar dari kira-kira 12.000 sampai 1 juta. Oleh karena itu,
enzim berukuran amat besar dibandingkan dengan substrat atau gugus
fungsional targetnya. Penataan tertentu pada rantai samping asam amino suatu
enzim di sisi aktifnya menentukan tipe molekul yang dapat terikat dan bereaksi
di situ. Biasanya ada sekitar lima rantai samping seperti itu dalam enzim
apapun. Selain itu, banyak enzim yang molekul-molekul nonprotein kecil yang
terhubung dengan sisi aktif atau didekatnya. Molekul-molekul enzim ini
disebut kofaktor atau koenzim (Ngili, 2009).
Beberapa enzim memerlukan kofaktor atau koenzim untuk aktivitas
katalitiknya, dan enzim lain mungkin membutuhkan koenzim maupun satu atau
ion) bersifat stabil sewaktu pemanasan, sedangkan bagian apoenzim (protein)
terdenaturasi oleh pemanasan (Poedjiadi dan Supriyanti, 1994).
1. Faktor-Faktor yang Mempengaruhi Aktivitas Enzim a. Suhu
Pengaruh suhu sangat menentukan aktivitas enzim pada waktu
mengkatalisa suatu reaksi. Seluruh enzim memerlukan jumlah panas
tertentu untuk dapat aktif. Sejalan dengan meningkatnya suhu, makin
meningkat pula aktivitas enzim. Secara umum, setiap peningkatan 10oC
di atas suhu minimum, aktivitas enzim akan meningkat sebanyak dua kali
lipat. Aktivitas enzim meningkat pada kecepatan ini hingga mencapai
kondisi optimum. Peningkatan suhu yang melebihi suhu optimumnya
menyebabkan lemahnya ikatan di dalam enzim secara struktural (Pratiwi,
2008). Pada suhu maksimum enzim akan terdenaturasi karena struktur
protein terbuka dan gugus non polar yang berada di dalam molekul
menjadi terbuka keluar, kelarutan protein di dalam air yang polar menjadi
turun, sehingga aktivitas enzim juga akan turun (Lehninger, 1997).
b. Konsentrasi Substrat
Aktivitas enzim dipengaruhi oleh konsentrasi substrat. Pada konsentrasi
substrat rendah, enzim tidak mencapai konversi maksimum akibat
sulitnya enzim menemukan substrat yang akan direaksikan. Seiring
dengan meningkatnya konsentrasi substrat, kecepatan reaksi juga akan
meningkat akibat makin cepatnya substrat terikat pada enzim.
Peningkatan konsentrasi substrat pada titik jenuh tidak lagi dapat
proximaty, atau kemudahan berdekatan yang disebut sebagai efek
keakraban, yang berarti bahwa laju reaksi antara dua molekul
ditingkatkan bila dalam larutan encer keduanya dijaga dalam jarak dekat
dalam sisi aktif enzim, sehingga menaikkan konsentrasi efektif reaktan
(Ngili, 2009).
c. pH
pH lingkungan juga berpengaruh terhadap kecepatan aktivitas enzim
dalam mengkatalisis suatu reaksi. Hal ini disebabkan konsentrasi ion
hidrogen mempengaruhi struktur tiga dimensi enzim dan aktivitasnya.
Setiap enzim memiliki pH optimum yang dapat mengikat substrat dengan
struktur tiga dimensi yang paling kondusif. Bila konsentrasi ion hidrogen
berubah dari konsentrasi optimal, aktivitas enzim secara progesif hilang
sampai pada akhirnya enzim menjadi tidak fungsional (Lehninger, 1997).
d. Inhibitor
Selain suhu, pH dan konsentrasi substrat, aktivitas enzim juga
dipengaruhi oleh ada tidaknya inhibitor. Jika terdapat pengurangan laju
reaksi oleh suatu senyawa, senyawa tersebut dinamakan inhibitor.
Inhibitor dapat bersaing dengan substrat dalam berikatan dengan enzim,
sehingga menghalangi substrat terikat pada tapak aktif enzim (Poedjiadi
dan Supriyanti, 1994). Peningkatan laju reaksi yang disebabkan oleh
2. Mekanisme Reaksi Enzim
Enzim bekerja dengan dua cara, yaitu menurut Teori Kunci-Gembok (Lock and Key Theory) dan Teori Kecocokan Induksi (Induced Fit Theory).
Mekanisme kerja enzim dapat dilihat pada Gambar 2.
Gambar 2.Mekanisme kerja reaksi enzim
Teori Kunci-Gembok (Lock and Key Theory) dikemukakan oleh Emil
Fisher yang menyatakan bahwa kerja enzim seperti kunci dan anak kunci,
melalui hidrolisis senyawa gula dengan enzim invertase. Terjadinya reaksi
antara substrat dengan enzim adalah karena adanya kesesuaian bentuk
ruang antara substrat dengan sisi aktif (active site) dari enzim. Dengan
begitu sisi aktif enzim cenderung kaku. Substrat berperan sebagai kunci
(key) dan sisi aktif (lock) berperan sebagai gembok. Substrat masuk ke
dalam sisi aktif sehingga terjadi kompleks enzim-substrat. Hubungan
antara enzim dan substrat membentuk ikatan yang lemah. Pada saat ikatan
kompleks enzim-substrat terputus, produk hasil reaksi akan dilepas dan
enzim akan kembali pada konfigurasi semula.
Teori Kecocokan Induksi (Induced Fit Theory) dikemukakan oleh Daniel
Koshland yang menyatakan bahwa ini sisi aktif tidak bersifat kaku tetapi Enzim
Substrat Sisi Aktif Enzim
(Active Site)
Enzim
Sisi Aktif Enzim (Active Site)
Substrat
Teori Kunci Gembok Sisi Aktif cenderung kaku
lebih fleksibel. Sisi aktif secara terus menerus berubah bentuknya sesuai
dengan interaksi antara enzim dan substrat. Ketika substrat memasuki sisi
aktif enzim, bentuk sisi aktif akan termodifikasi menyesuaikan bentuk
substrat sehingga terbentuk kompleks enzim substrat. Sisi aktif akan terus
berubah bentuknya sampai substrat terikat secara sepenuhnya, yang mana
bentuk akhir dan muatan enzim ditentukan. Ketika substrat terikat pada
enzim, sisi aktif enzim mengalami beberapa perubahan sehingga ikatan
yang terbentuk antara enzim dan substrat menjadi menjadi lebih kuat.
Interaksi antara enzim dan substrat disebutInduced fit(Shahib, 2005).
E. Enzim CGT-ase
Enzim CGT-ase yaitu enzim ekstraseluler yang dapat mengubah pati menjadi
siklodekstrin (Tonkova, 1998). CGT-ase diklasifikasikan menjadi tiga tipe
berdasarkan jenis-jenis siklodekstrin yang dihasilkan dari ikatan
α-1,4-glikosida seperti pati, melalui reaksi transglikosilasi intramolekuler, yaitu tipeBacillus maceransyang menghasilkan α-siklodekstrin, tipeBacillus
megateriumyang menghasilkan β-siklodekstrin, dan tipeBacillussp.
menghasilkan γ-siklodekstrin (Moriet al.,1994).
CGT-ase merupakan keluarga dari enzim α-amilase (Leemhuiset al., 2003).
CGT-ase memiliki berat molekul yang bervariasi dari 60-110 kDa dan terdiri
dari 700 asam amino. Sebagian memerlukan kalsium sebagai agen pelindung
terhadap denaturasi panas (Bovettoet al., 1992) dan CGT-ase alkalofilik
optimum pada pH 9-10 (Tao, 1991). Suhu maksimal untuk bakteri yang
CGT-ase diinhibisi kuat oleh Zn2+, Cu2+dan Fe2+ (Tonkova, 1998).
Reaksi katalisis oleh enzim CGT-ase dapat terjadi secara intramolekul
(siklisasi) dan antarmolekul (kopling, disproporsionasi), serta reaksi hidrolisis
(Penninga, 1996). Reaksi siklisasi yaitu transfer residu gula akhir ke residu
gula yang lain pada rantai oligosakarida yang sama untuk membentuk suatu
senyawa siklik. Ini merupakan reaksi intramolekul pada pati yang
dihubungkan dengan ikatan α-1,4-glukan yang dikonversi ke dalam
siklodekstrin. Reaksi ini bersifatreversibledan cincin dapat dibuka oleh
CGT-ase untuk reaksi lebih lanjut (Hedges, 1992). Siklisasi adalah reaksi utama dari
CGT-ase untuk menghasilkan siklodekstrin. Produksiα-siklodekstrin,
β-siklodekstrindan γ-siklodekstrin oleh enzim CGT-ase tergantung pada
beberapa faktor seperti pemilihan substrat pati untuk degradasi enzim,
karakteristik enzim, komposisi medium dan kondisi yang digunakan pada
proses reaksi katalisis pati oleh enzim CGT-ase (Salvaet al.,1997). Reaksi
siklisasi enzim CGT-ase dapat dilihat pada Gambar 3.
Gambar 3.Reaksi siklisasi enzim CGT-ase (Van der Veenet al., 2000)
F. Siklodekstrin
Siklodekstrin adalah merupakan oligosakarida yang tersusun atas 6, 7 atau 8
melingkar seperti kue donat. Berdasarkan jumlah unit glukosa penyusun,
siklodekstrin dipilah menjadi tiga, yaitu alfa (6 unit), beta (7 unit) dan gamma
(8 unit). Cincin luar struktur siklodekstrin bersifat polar (hidrofilik) sedangkan
bagian dalam rongga bersifat lebih nonpolar (hidrofobik) (Szetjli, 1982).
Produk siklik dapat terbentuk secara kompleks inklusi dengan senyawaan
anorganik maupun organik. Sifat ini menyebabkan siklodektrin banyak
digunakan dalam berbagai industri seperti pangan, kosmetika, farmasi,
agrokimia serta untuk penanganan polusi (Bender, 1977; Kaneto and
Fumithasi, 1996; Szetjli, 1982).
Adapun struktur α-siklodekstrin, β-siklodekstrindan γ-siklodekstrin dapat
dilihat pada Gambar 4 serta sifat-sifat siklodekstrin dapat dilihat pada Tabel 2.
Gambar 4.Strukturα-, β-dan γ-siklodekstrin (Van der Veenet al.,2000)
Tabel 2.Sifat-sifat siklodekstrin (Van der Veenet al.,2000)
Tipe Siklodekstrin
Karakteristik Alfa Beta Gamma
Berat molekul (g/mol) Monomer glukosa
Diameter rongga internal (Å)
Kelarutan dalam air (g/100mL:25°C) Tegangan permukaan (mN/m) Rentang lebur (°C)
Air kristalisasi
Molekul air dalam rongga
972
G. Pengujian Aktivitas Enzim CGT-ase
Pada uji aktivitas, larutan pati berperan sebagai substrat untuk enzim CGT-ase.
Enzim ini akan mengurai pati menjadi siklodekstrin. Proses penguraian pati
menjadi siklodekstrin, berlangsung saat inkubasi pada suhu 55°C selama
10 menit. Penghentian reaksi antara pati dengan enzim dilakukan dengan
penambahan Na2CO3. Selanjutnya, ditambahkan larutan fenolftalein untuk
mengetahui terbentuk kompleks siklodekstrin dengan fenolftalein dengan
menunjukkan perubahan warna menjadi merah muda keunguan. Besarnya
pengurangan pati dapat dideteksi dari menurunnya serapan larutan pada daerah
visible(campuran reaksi berwarna merah muda keunguan) dengan
spektrofotometerUV-Vis. Nilai serapan yang terukur pada spektrofotometer UV-Visdapat digunakan untuk menghitung besarnya aktivitas unit enzim
CGT-ase. Sedangkan satu unit enzim β-siklodekstrin didefinisikan sebagai
H. Penentuan Kadar Protein
Penentuan kadar protein dengan metode Lowry didasarkan pada pengukuran
serapan cahaya oleh ikatan kompleks yang berwarna ungu. Ini terjadi karena
protein bereaksi dengan tembaga dalam lingkungan alkali yang mudah larut,
kompleks Cu2+dengan ikatan peptida akan tereduksi menjadi Cu+. Lalu, Cu+
akan mereduksi folin-ciocalteu yang mengikat protein sekitar pH 10. Sehingga
kompleks fosfomolibdat-fosfotungstat menghasilkanheteropolymolybdenum
dari warna kuning menjadi biru. Ini disebabkan karena oksidasi gugus
aromatik terkatalis Cu, sehingga menghasilkan kompleks berwarna biru dalam
derajat yang berbeda tergantung pada komposisi triftofan dan tirosinnya.
Karena itu, protein yang berbeda akan memberikan tingkat warna yang berbeda
(Alexander and Griffith, 1993).
Metode ini merupakan metode relatif sederhana dengan biaya yang relatif
murah juga. Tetapi juga mempunyai kelemahan yaitu sensitif terhadap
perubahan pH dan konsentrasi protein yang rendah. Hal ini dapat diatasi
dengan menggunakan volume sampel yang sangat kecil sehingga tidak
mempengaruhi reaksi (Lowryet al.,1951).
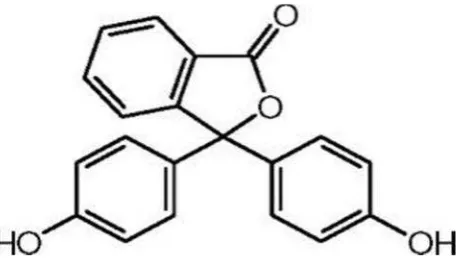
I. Fenolftalein
Sifat fisik dan kimia dari fenolftalein:
(a) Gambaran: Putih atau kekuningan-putih, kristal triklinik
(c) Kelarutan: Praktis tidak larut dalam air; larut dalam dietil eter, etanol dan
larutan cair alkali hidroksida, sangat sedikit larut dalam kloroform
(d) Konstanta disosiasi: pKa 9,7 pada suhu 25oC
(Budavari, 1996).
Berikut struktur kimia dari
fenolftalein(3,3-Bis(4-hidroksifenil)-1-(3H)-isobenzofuranon) yang terdapat pada Gambar 5.
III. METODOLOGI PENELITIAN
A. Waktu dan Tempat Penelitian
Penelitian ini telah dilakukan pada bulan April sampai dengan Juni 2012 di
Laboratorium Biokimia Jurusan Kimia Fakultas Matematika dan Ilmu
Pengetahuan Alam Universitas Lampung.
B. Alat dan Bahan
Alat-alat yang digunakan pada penelitian ini antara lain adalah alat-alat gelas,
Bunsen, kasa, kapas, spatula, rak tabung reaksi, jarum ose, autoklaf
(model S-90N),laminar air flow(CURMA model 9005-FL), neraca analitik,
inkubator,shaker incubator(Stuart SSL2),waterbath, sentrifuga, tabung
sentrifuga, mikropipet, pH indikator, dan spektrofotometerUV-Vis
(Cary Win UV 32).
Sedangkan bahan-bahan yang digunakan pada penelitian ini adalah pati
singkong, pati ubi jalar, pati jagung, pati sagu, maltosa, urea, NH4Cl, CuSO4,
FeSO4, ZnSO4, CaCl2.2H2O, pepton, ekstrak ragi, K2HPO4, MgSO4.7H2O,
fenolftalein (PP), metil jingga, agar, natrium karbonat (Na2CO3), NaCl,
pereaksi C, pereaksi D, alkohol, dan akuades. Sampel yang digunakan adalah
bakteri isolat lokal LTi-21-3 yang telah diperoleh dari peneliti sebelumnya.
C. Prosedur Penelitian
1. Peremajaan Bakteri dan Produksi Enzim CGT-ase dari Isolat LTi-21-3
Kegiatan rutin yang dilakukan untuk peremajaan bakteri yang digunakan
pada penelitian ini adalah :
a. Peremajaan Bakteri Isolat LTi-21-3
Bakteri isolat LTi-21-3diremajakan dalam medium Horikoshi’s II
termodifikasi yang memiliki komposisi 1 g pati singkong, 0,5 g pepton,
0,5 g ekstrak ragi, 0,1 g K2HPO4, 0,02 g MgSO4.7H2O, 0,03 g
fenolftalein, 0,01 g metil jingga dan 1,5 g agar. Kemudian dimasukkan
ke dalam Erlenmeyer dan dilarutkan dalam 80 mL akuades disebut
larutan I. Larutan I dipanaskan sampai larutan jernih sambil diaduk.
Selanjutnya membuat larutan natrium karbonat (Na2CO3) 1% dengan
cara menimbang 0,25 g Na2CO3dilarutkan dalam 25 ml akuades dan
dimasukkan ke dalam Erlenmeyer disebut larutan II. Sejumlah akuades
disiapkan dan dimasukkan ke dalam Erlenmeyer disebut larutan III.
Larutan I, larutan II dan larutan III disterilisasi dengan menggunakan
autoklaf pada suhu 121oC dengan tekanan 1 atm selama 30 menit.
Medium Horikoshi’s IItermodifikasi disiapkan dengan mencampurkan
larutan I sebanyak 80 mL, larutan II sebanyak 17 mL dan larutan III
sebanyak 3 mL yang dilakukan di dalamlaminar air flow. Setelah
dimasukkan ke dalam tabung reaksi sebanyak ± 5 mL, dimiringkan dan
didiamkan pada suhu kamar. Setelah media memadat dilakukan
inokulasi bakteri isolat LTi-21-3 tersebut pada media agar miring.
Kemudian diinkubasi selama 3 hari dalam inkubator.
b. Pembuatan MediumNutrien Broth(NB)
Medium NB dibuat dengan komposisi 0,3% (b/v) ekstrak ragi, 0,5% (b/v)
pepton dan 0,5% (b/v) NaCl, kemudian disterilisasi. Medium NB
digunakan sebagai penyiapan inokulum (starter) dengan cara
menginokulasikan satu ose galur bakteri isolat LTi-21-3 dari agar miring
yang telah berusia 3 hari ke dalam masing-masing Erlenmeyer yang
berisi 5 mL medium NB. Setelah diinokulasi, kemudian diinkubasi
dalamshaker incubatordengan kecepatan 105 rpm pada suhu 37ºC
selama 1 malam (overnight: 16-20 jam).
c. Pembuatan Medium Kultur dan Ekstraksi Enzim CGT-ase
Medium kultur menggunakan mediumHorikoshi’s IItermodifikasi
(Parket al.,1989) yang mengandung pati singkong, pepton, ekstrak ragi,
K2HPO4, MgSO4.7H2O, Na2CO3tanpa fenolftalein dan metil jingga
dengan pH medium 10. Produksi enzim CGT-ase dilakukan dengan
menambahkan 1% inokulum daristarterisolat LTi-21-3 diinokulasikan
ke dalam media kultur volume 50 mL dalam Erlenmeyer 250 mL. Kultur
kemudian diinkubasi padashaker incubatordengan kecepatan 105 rpm,
berkala setiap 12 jam dengan cara sentrifugasi, untuk mendapatkan
ekstrak kasar enzim CGT-ase.
2. Penentuan Pertumbuhan Sel
Penentuan pertumbuhan sel ini digunakan untuk mengetahui pertumbuhan
dari sel bakteri. Sebanyak 0,3 mL kultur dimasukkan ke dalam tabung reaksi
kemudian ditambahkan 2,7 mL akuades lalu diukur serapannya
menggunakan spektrofotometerUV-Vispada panjang gelombang 600 nm.
3. Uji Aktivitas Enzim CGT-ase
a. Pembuatan Larutan Buffer Asetat 0,1 M (pH 5,5)
Sebanyak 100 mL Na-asetat 0,1M dimasukkan ke dalam Erlenmeyer
kemudian ditambahkan 0,2 mL asam asetat 0,1M.
b. Pereaksi CGT-ase(Kanekoet al.,1987; Alves-Pradoet al.,2008)
Pereaksi CGT-ase digunakan untuk menguji aktivitas ekstrak kasar
enzim CGT-ase. Pereaksi CGT-ase yaitu larutan patisoluble1% (b/v).
Larutan patisolubledisiapkan dengan melarutkan 1 g patisolubleke
dalam 100 mL 0,1M buffer asetat pH 5,5 kemudian dipanaskan hingga
larut.
c. Penentuan Aktivitas Enzim CGT-ase
Ekstrak kasar enzim CGT-ase yang diperoleh kemudian diuji aktivitas
enzim CGT-ase nya. Sebanyak 100 µL larutan enzim ditambahkan
800 µL patisoluble1% yang disiapkan dalam buffer asetat 0,1M
pH 5,5 dan diinkubasi pada suhu 55ºC selama 10 menit. Reaksi
larutan fenolftalein 1 mM. Absorbansi diukur menggunakan
spektrofotometerUV-Vispada λmaks550 nm. Kontrol dibuat dengan cara
menginaktifkan larutan enzim pada suhu 100ºC selama 30 menit, dan
selanjutnya diperlakukan sama dengan sampel.
4. Uji Kadar Protein Enzim CGT-ase a. Pereaksi Lowry(Lowryet al., 1951)
Pereaksi Lowry terdiri atas 4 macam, yang meliputi Pereaksi A, B, C,
dan D. Masing-masing pereaksi tersebut disiapkan sebagai berikut :
Pereaksi A : 2 g Na2CO3dilarutkan dalam 100 mL NaOH 0,1N; Pereaksi
B : 5 mL larutan CuSO4.5H2O 1% (b/v) ditambahkan 5 mL larutan
Na(K)-tartarat 1% (b/v); Pereaksi C : 2 mL pereaksi B ditambah 100 mL
pereaksi A; dan Pereaksi D : reagenFolin-Ciocelteaudiencerkan dengan
akuades 1:1. Larutan standar protein digunakan BSA (Bovine Serum Albumine) dengan konsentrasi 500-5000 ppm.
b. Penentuan Kadar Protein Enzim CGT-ase
Ekstrak kasar enzim CGT-ase yang diperoleh kemudian diuji kadar
proteinnya. Sebanyak 0,1 mL larutan enzim ditambahkan 0,9 mL
akuades lalu ditambahkan dengan 5 mL pereaksi C. Campuran diaduk
secara merata dan dibiarkan selama 10 menit pada suhu kamar.
Kemudian ditambahkan dengan cepat 0,5 mL pereaksi D dan diaduk
dengan sempurna. Setelah itu didiamkan selama 30 menit pada suhu
kamar. Sebagai kontrol, larutan enzim diganti dengan akuades.
serapan dilakukan menggunakan spektrofotometerUV-Vispada λmaks
600 nm.
5. Pengaruh Beberapa Faktor Pada Medium KulturHorikoshi’s II Termodifikasi Terhadap Pertumbuhan Bakteri dan Produksi Enzim CGT-ase dari Isolat LTi-21-3
a. Pengaruh Sumber Karbon
Pada medium kulturHorikoshi’s II termodifikasidengan sumber karbon
pati singkong dianggap sebagai medium kultur tanpa perlakuan variasi.
Selanjutnya dilakukan variasi sumber karbon dengan cara mengganti pati
singkong 1% (b/v) dengan patisoluble, pati jagung, pati ubi jalar, pati
sagu masing-masing 1% (b/v) dan penambahan maltosa 0,5% (b/v).
Kemudian dilakukan perlakuan yang sama dengan prosedur 2, 3 bagian c
dan 4 bagian b lalu hasilnya dibandingkan antara tanpa dan dengan
perlakuan variasi.
b. Pengaruh Sumber Nitrogen
Sumber karbon terbaik kemudian dilakukan variasi terhadap sumber
nitrogen. Medium kultur Horikoshi’s IItermodifikasi yang mengandung
pepton sebagai sumber nitrogen dianggap sebagai tanpa perlakuan
variasi. Selanjutnya dilakukan variasi sumber nitrogen dengan cara
mengganti pepton 0,5% (b/v) dengan ekstrak ragi, urea dan NH4Cl
masing-masing 0,5% (b/v). Kemudian dilakukan perlakuan yang sama
dengan prosedur 2, 3 bagian c dan 4 bagian b lalu hasilnya dibandingkan
c. Pengaruh Sumber Ion Logam
Sumber karbon dan nitrogen terbaik kemudian dilakukan variasi terhadap
sumber ion logam. Medium kultur Horikoshi’s IItermodifikasi yang
mengandung MgSO4sebagai sumber ion logam dianggap sebagai tanpa
perlakuan variasi. Selanjutnya dilakukan variasi sumber ion logam
dengan cara mengganti MgSO40,02% (b/v) dengan CuSO4,FeSO4,
ZnSO4dan CaCl2masing-masing 0,02% (b/v). Kemudian dilakukan
perlakuan yang sama dengan prosedur 2, 3 bagian c dan 4 bagian b lalu
hasilnya dibandingkan antara tanpa dan dengan perlakuan variasi.
d. Pengaruh pH
Sumber karbon, nitrogen dan ion logam terbaik kemudian dilakukan
variasi pH-nya. Medium kultur Horikoshi’s IItermodifikasi dengan pH
10 dianggap sebagai tanpa perlakuan variasi. Selanjutnya dilakukan
variasi pH-nya yaitu pada pH 7, 8, 9 dan 11 dengan cara perbedaan
penambahan larutan Na2CO3. Kemudian dilakukan perlakuan yang sama
dengan prosedur 2, 3 bagian c dan 4 bagian b lalu hasilnya dibandingkan
6. Diagram Alir Prosedur Penelitian
-divariasikan sumber karbon pada medium Horikoshi’s II
termodifikasi
-divariasikan sumber nitrogen pada medium Horikoshi’s II
termodifikasi
-dilakukan seperti prosedur 4
--divariasikan sumber ion logam pada medium Horikoshi’s II
termodifikasi
-dilakukan seperti prosedur 4
--divariasikan pH pada medium Horikoshi’s IItermodifikasi
-dilakukan seperti prosedur 4
1. Peremajaan bakteri isolat LTi-21-3
2. Penanaman bakteri ke dalam media NB
3. Inokulasi dari media NB ke medium
Horikoshi’s IItermodifikasi
1.
4. Produksi Enzim CGT-ase - Penentuan pertumbuhan sel - Penentuan aktivitas enzim - Penentuan kadar protein
Medium Horikoshi’s IItermodifikasi dengan
sumber karbon terbaik
Medium Horikoshi’s IItermodifikasi dengan
sumber karbon dan nitrogen terbaik
Prosedur 1, 2, 3
Prosedur 1, 2, 3
Medium Horikoshi’s IItermodifikasi dengan
sumber karbon, nitrogen dan ion logam terbaik
Prosedur 1, 2, 3
Medium Horikoshi’s IItermodifikasi dengan
IV. HASIL DAN PEMBAHASAN
A. Pengaruh Beberapa Faktor Pada Medium Kultur Terhadap Pertumbuhan Sel dan Produksi Enzim CGT-ase Isolat LTi-21-3
Pertumbuhan mikroorganisme tertentu sangat dipengaruhi oleh sumber karbon,
nitrogen, ion logam dan pH, dari sumber-sumber tersebut ada yang berfungsi
mengkatalisis jumlah sel dan jumlah produksi enzim dan ada juga yang
berfungsi menghambat pertumbuhan dan produksi enzim.
Pada penelitian ini, variasi komposisi medium sumber karbon, nitrogen, ion
logam dan pH terbaik dipilih berdasarkan aktivitas spesifik tertinggi dari isolat
LTi-21-3. Aktivitas spesifik merupakan hasil bagi antara aktivitas unit
terhadap kadar protein. Dengan adanya aktivitas spesifik yang tinggi ini akan
memudahkan dalam proses pemurnian enzim CGT-ase.
Hasil dari keseluruhan penelitian ini menunjukkan bahwa waktu optimum
untuk menghasilkan enzim CGT-ase ekstraseluler untuk isolat LTi-21-3 ada
pada 36 jam. Pada waktu optimum ini, enzim CGT-ase ekstraseluler banyak
diproduksi oleh isolat bakteri. Hal ini ditunjukkan dengan aktivitas yang tinggi
1. Pengaruh Sumber Karbon
Menurut Posciet al.(1998), patisolubleadalah sumber karbon terbaik
untukB. firmusdanB. maceransdalam memproduksi enzim CGT-ase.
PadaB. circulansDF 9R sumber karbon terbaiknya yaitu 1,5% pati
singkong (Rossoet al., 2002) danBacillussp. TS1-1 alkalofilik dan
Bacillus stearothermophilusHR1 secara berurutan dengan 1,48% dan 1,5%
pati sagu (Mahatet al., 2004; Rahmanet al., 2004) dalam memproduksi
enzim CGT-ase. Menurut Ibrahimet al. (2005),BacillusG1 memiliki
aktivitas CGT-ase tertinggi ketika digunakan pati singkong sebagai sumber
karbonnya.
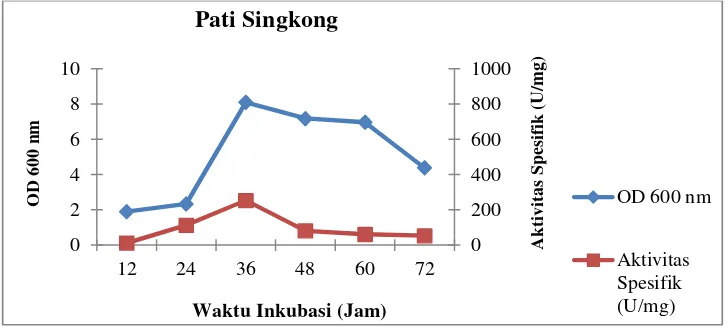
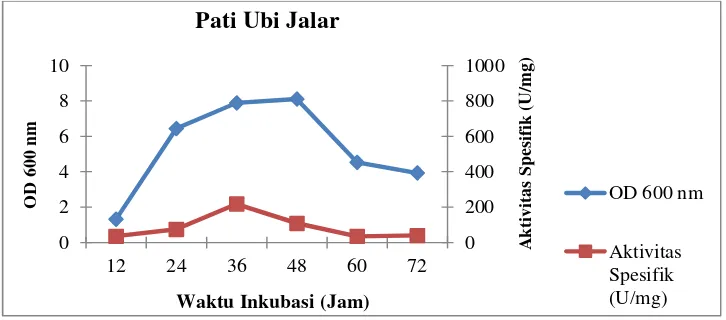
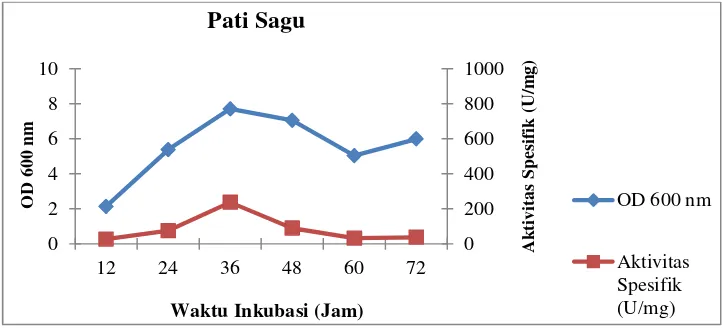
Pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3 pada variasi
komposisi medium sumber karbon dapat dilihat pada Gambar 6–11
berikut:
Gambar 6.Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s II dengan sumber karbonpati
singkong
Waktu Inkubasi (Jam) Pati Singkong
OD 600 nm
Gambar 7.Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s II dengan sumber karbonpati
soluble
Gambar 8.Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s II dengan sumber karbon pati jagung
Gambar 9.Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s II dengan sumber karbonpati ubi
jalar
Waktu Inkubasi (Jam)
PatiSoluble
Waktu Inkubasi (Jam) Pati Jagung
Waktu Inkubasi (Jam) Pati Ubi Jalar
OD 600 nm
Gambar 10. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s II dengan sumber karbonpati
sagu
Gambar 11. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
karbon penambahan maltosa
Pada Tabel 3. berikut dapat dilihat pertumbuhan sel, aktivitas unit, kadar
protein dan aktivitas spesifik dari isolat LTi-21-3 pada variasi komposisi
medium sumber karbon dengan waktu inkubasi optimum 36 jam. Data
lengkap untuk pertumbuhan sel, aktivitas unit, kadar protein dan aktivitas
spesifik dari isolat LTi-21-3 pada variasi komposisi medium sumber
karbon dapat dilihat pada Lampiran 4.
0
Waktu Inkubasi (Jam) Pati Sagu
Waktu Inkubasi (Jam) Penambahan Maltosa
OD 600 nm
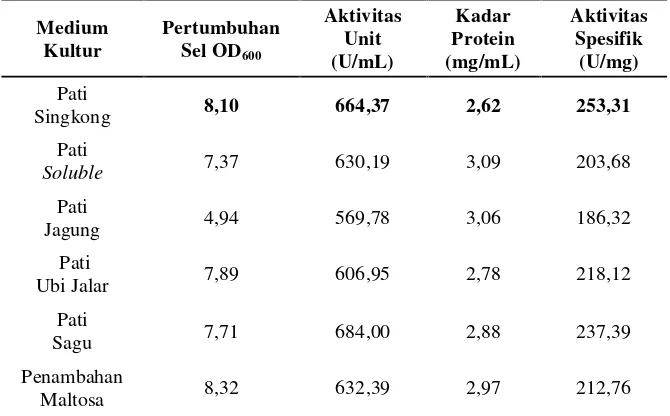
Tabel 3. Pertumbuhan sel, aktivitas unit, kadar protein dan aktivitas
spesifik isolat LTi-21-3 pada variasi komposisi medium sumber karbon dengan waktu inkubasi optimum 36 jam.
Medium
Singkong 8,10 664,37 2,62 253,31
Pati
Soluble 7,37 630,19 3,09 203,68
Pati
Jagung 4,94 569,78 3,06 186,32
Pati
Ubi Jalar 7,89 606,95 2,78 218,12
Pati
Sagu 7,71 684,00 2,88 237,39
Penambahan
Maltosa 8,32 632,39 2,97 212,76
Tabel 4. Perbandingan pertumbuhan sel, aktivitas unit dan aktivitas
spesifik antara medium tanpa dan dengan perlakuan variasi komposisi medium sumber karbon
Jenis
Pati Singkong 8,10 664,37 253,31 100 100 100
Dengan Perlakuan
Variasi
PatiSoluble 7,37 630,19 203,68 91 95 80
Pati Jagung 4,94 569,78 186,32 61 86 74 Pati
Ubi Jalar 7,89 606,95 218,12 97 91 86 Pati Sagu 7,71 684,00 237,39 95 103 94 Penambahan
Maltosa 8,32 632,39 212,76 103 95 84 Keterangan: OD = pertumbuhan sel, AU = aktivitas unit dan
AS = aktivitas spesifik
Dari kedua data yang disajikan di atas, diketahui bahwa isolat LTi-21-3
menghasilkan aktivitas unit tertinggi pada sumber karbon pati sagu yaitu
termodifikasi hanya meningkatkan pertumbuhan sel tanpa diikuti dengan
meningkatnya aktivitas unit dan aktivitas spesifiknya. Terdapatnya gula
sederhana dalam substrat hidrolisat seperti glukosa, maltosa dan
maltotriosa, menyebabkan aktivitas CGT-ase tidak dapat mengkonversi
substrat menjadi siklodekstrin secara optimal. Hal tersebut disebabkan
gula sederhana menghambat reaksi siklisasi akibat terjadinya reaksi
couplingyaitu pemecahan cincin siklodekstrin sehingga terbentuk
maltooligosakarida yang menghasilkan maltooligosakarida lain dan gula
pereduksi (Charoenlapet al.,2004; Burhanet al., 2005). Namun, pada
medium kultur Horikoshi’s II termodifikasiyang mengandung sumber
karbon pati singkong menghasilkan aktivitas unit dan aktivitas spesifik
secara berurutan yaitu 664,37 U/mL dan 253,31 U/mg. Nilai aktivitas
spesifik ini merupakan nilai tertinggi dari sumber karbon yang lainnya.
Untuk itu, pada variasi komposisi medium sumber karbon dengan pati
singkong dipilih sebagai sumber karbon terbaik.
2. Pengaruh Sumber Nitrogen
Ibrahimet al. (2005) mengamati adanya pengaruh sumber nitrogen
organik dan anorganik pada produksi CGT-ase dariBacillusG1. Mereka
menemukan bahwa produksi CGT-ase tertinggi ketika sumber nitrogen
organik ada di dalam medium. Menurut Illiaset al. (2003) sumber
nitrogen terbaik untuk produksi CGT-ase adalah pepton. Tetapi Mahat
et al. (2004) mengamati bahwa pati sagu dan ekstrak ragi mempunyai efek
yang signifikan untuk produksi CGT-ase. Gawande and Patkar (1999),
sumber nitrogen untukKlebsiella pneumoniaeAS-22. Urea dan NH4NO3 diketahui menghambat pertumbuhan bakteri dan selanjutnya tidak ada
enzim yang dihasilkan.
Pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3 pada variasi sumber
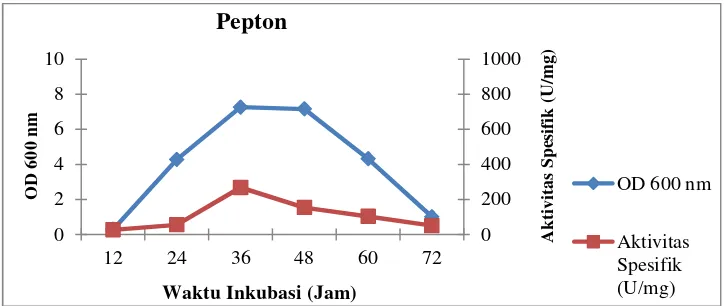
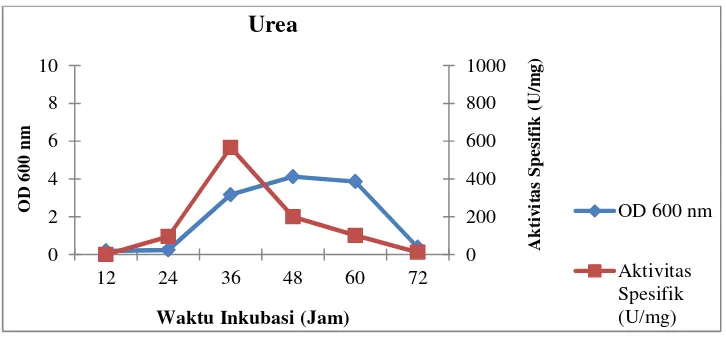
nitrogen dapat dilihat pada Gambar 12–15 berikut:
Gambar 12. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
nitrogen pepton
Gambar 13. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
nitrogen ekstrak ragi
0
Waktu Inkubasi (Jam) Pepton
Waktu Inkubasi (Jam) Ekstrak Ragi
OD 600 nm
Gambar 14. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
nitrogen urea
Gambar 15. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
nitrogen NH4Cl
Berikut data pertumbuhan sel, aktivitas unit, kadar protein dan aktivitas
spesifik dari isolat LTi-21-3 pada variasi komposisi medium sumber
nitrogen dengan waktu inkubasi optimum 36 jam dapat dilihat pada Tabel
5. Data lengkap untuk pertumbuhan sel, aktivitas unit, kadar protein dan
aktivitas spesifik dari isolat LTi-21-3 pada variasi komposisi medium
sumber nitrogen dapat dilihat pada Lampiran 5.
0
Waktu Inkubasi (Jam) Urea
Waktu Inkubasi (Jam) NH4Cl
OD 600 nm
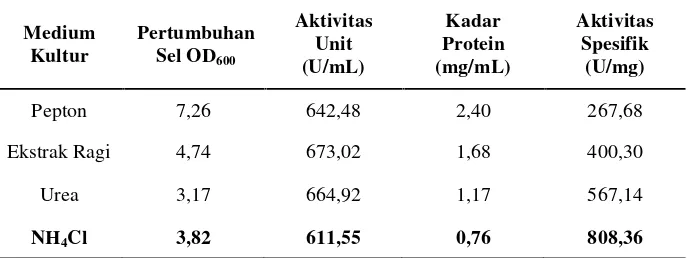
Tabel 5. Pertumbuhan sel, aktivitas unit, kadar protein dan aktivitas
spesifik isolat LTi-21-3 pada variasi komposisi medium sumber nitrogen dengan waktu inkubasi optimum 36 jam
Medium
Pepton 7,26 642,48 2,40 267,68
Ekstrak Ragi 4,74 673,02 1,68 400,30
Urea 3,17 664,92 1,17 567,14
NH4Cl 3,82 611,55 0,76 808,36
Tabel 6. Perbandingan pertumbuhan sel, aktivitas unit dan aktivitas
spesifik antara medium tanpa dan dengan perlakuan variasi komposisi medium sumber nitrogen
Jenis
Variasi Pepton 7,26 642,48 267,68 100 100 100
Dengan Perlakuan
Variasi
Ekstrak
Ragi 4,74 673,02 400,30 65 105 150 Urea 3,17 664,92 567,14 44 103 212
NH4Cl 3,82 611,55 808,36 53 95 302 Keterangan: OD = pertumbuhan sel, AU = aktivitas unit dan
AS = aktivitas spesifik
Dari kedua data di atas, diketahui bahwa aktivitas unit pada waktu
inkubasi optimum 36 jam dengan sumber nitrogen pepton dan ekstrak ragi
secara berurutan yaitu 642,48 U/mL dan 673,02 U/mL. Nilai aktivitas unit
kedua sumber nitrogen tersebut lebih tinggi jika dibandingkan dengan
sumber nitrogen NH4Cl (611,55 U/mL). Walaupun pepton dan ekstrak
ragi merupakan sumber nitrogen organik sedangkan urea dan NH4Cl
adalah sumber nitrogen anorganik. Namun, sumber nitrogen NH4Cl
memiliki aktivitas spesifik yang lebih tinggi jika dibandingkan dengan
menggunakan NH4Cl mengalami penurunan sekitar 47% dari medium
tanpa perlakuan. Dengan demikian, sumber nitrogen NH4Cl dipilih
sebagai variasi komposisi medium terbaik.
3. Pengaruh Sumber Ion Logam
Menurut Ibrahimet al.(2005), Gawandeet al.(1998), Gawande and
Patkar (1999) dan Jin-Bonget al.(1990) menemukan bahwa magnesium
diperlukan untuk produksi CGT-ase. Cu2+dan Zn2+merupakan efek
inhibitor yang signifikan dalam kestabilan CGT-ase untukBrevibacterium
sp. No. 9605 (Moriet al., 1994),B. agaradhaerens (Martins and
Hatti-Kaul, 2002) danBacillussp.AL-6 (Fujitaet al., 1990). Sebagian besar
CGT-ase diinhibisi kuat oleh Zn2+, Cu2+dan Fe2+ (Tonkova, 1998).
Pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3 pada variasi
komposisi medium sumber ion logam dapat dilihat pada Gambar 16–20
berikut:
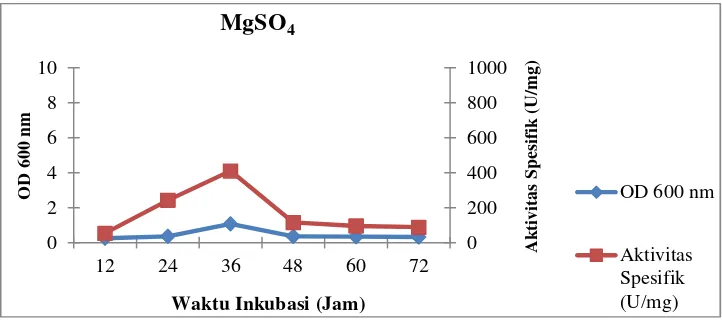
Gambar 16. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
ion logam MgSO4
0
Waktu Inkubasi (Jam) MgSO4
OD 600 nm
Gambar 17. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
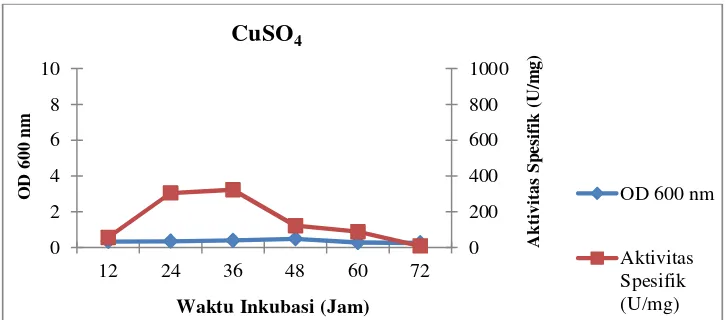
ion logam CuSO4
Gambar 18. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
ion logam ZnSO4
Gambar 19. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
ion logam FeSO4
0
Waktu Inkubasi (Jam) CuSO4
Waktu Inkubasi (Jam) ZnSO4
Waktu Inkubasi (Jam) FeSO4
OD 600 nm
Gambar 20. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi dengan sumber
ion logam CaCl2
Medium Horikoshi’s IItermodifikasi yang mengandung sumber ion logam
MgSO4merupakan medium tanpa perlakuan. Selain logam yang
berkombinasi dengan anion sulfat (SO42-), digunakan juga logam yang
berkombinasi dengan anion klorida yaitu CaCl2.
Berikut data pertumbuhan sel, aktivitas unit, kadar protein dan aktivitas
spesifik dari isolat LTi-21-3 pada variasi komposisi medium sumber ion
logam dengan waktu inkubasi optimum 36 jam dapat dilihat pada Tabel 7.
Data lengkap untuk pertumbuhan sel, aktivitas unit, kadar protein dan
aktivitas spesifik dari isolat LTi-21-3 pada variasi komposisi medium
sumber ion logam dapat dilihat pada Lampiran 6.
0
Waktu Inkubasi (Jam) CaCl2
OD 600 nm
Tabel 7. Pertumbuhan sel, aktivitas unit, kadar protein dan aktivitas
spesifik isolat LTi-21-3 pada variasi komposisi medium sumber ion logam dengan waktu inkubasi optimum 36 jam
Medium
MgSO4 1,08 529,19 1,29 410,23
CuSO4 0,40 504,63 1,56 322,71
ZnSO4 0,42 480,37 1,42 338,40
FeSO4 0,38 522,55 1,53 340,54
CaCl2 1,18 416,27 1,53 272,86
Tabel 8. Perbandingan pertumbuhan sel, aktivitas unit dan aktivitas
spesifik antara medium tanpa dan dengan perlakuan variasi komposisi medium sumber ion logam
Jenis
MgSO4 1,08 529,19 410,23 100 100 100
Dengan Perlakuan
Variasi
CuSO4 0,40 504,63 322,71 37 95 79
ZnSO4 0,42 480,37 338,40 39 91 82
FeSO4 0,38 522,55 340,54 35 99 83
CaCl2 1,18 416,27 272,86 109 84 80 Keterangan: OD = pertumbuhan sel, AU = aktivitas unit dan
AS = aktivitas spesifik
Dari data yang disajikan di atas, isolat LTi-21-3 memiliki aktivitas unit
dan aktivitas spesifik yang tinggi pada sumber ion logam MgSO4secara
berurutan yaitu 529,19 U/mL dan 410,23 U/mg. Untuk sumber ion logam
CaCl2menghasilkan aktivitas unit dan aktivitas spesifik yang terendah dan
pertumbuhan selnya tertinggi dibandingkan dengan sumber ion logam
dengan sumber ion logam MgSO4mengalami penurunan begitu juga
dengan pertumbuhan selnya dibandingkan dengan penelitian sebelumnya
pada variasi komposisi medium sumber nitrogen. Sumber ion logam
MgSO4dipilih sebagai sumber ion logam terbaik karena memiliki aktivitas
spesifik tertinggi sesuai dengan beberapa literatur yang telah disebutkan di
atas.
4. Pengaruh Variasi pH
Pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3 pada variasi
komposisi pH medium dapat dilihat pada Gambar 21- 25 berikut:
Gambar 21. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi pH 7
Gambar 22. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi pH 8 0
Waktu Inkubasi (Jam) pH 7
Waktu Inkubasi (Jam) pH 8
OD 600 nm
Gambar 23. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi pH 9
Gambar 24. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi pH 10
Gambar 25. Profil pertumbuhan sel dan aktivitas spesifik isolat LTi-21-3
pada mediumHorikoshi’s IItermodifikasi pH 11 0
Waktu Inkubasi (Jam) pH 9
Waktu Inkubasi (Jam) pH 10
Waktu Inkubasi (Jam) pH 11
OD 600 nm
Pada Tabel 9 berikut dapat dilihat pertumbuhan sel, aktivitas unit, kadar
protein dan aktivitas spesifik dari isolat LTi-21-3 pada variasi komposisi
pH medium dengan waktu inkubasi optimum 36 jam. Data lengkap untuk
pertumbuhan sel, aktivitas unit, kadar protein dan aktivitas spesifik dari
isolat LTi-21-3 pada variasi komposisi pH medium dapat dilihat pada
Lampiran 7.
Tabel 9. Pertumbuhan sel, aktivitas unit, kadar protein dan aktivitas
spesifik isolat LTi-21-3 pada variasi komposisi pH medium dengan waktu inkubasi optimum 36 jam
Medium
pH 7 3,87 520,44 0,98 529,58
pH 8 3,75 546,86 0,92 595,72
pH 9 1,75 467,39 1,29 361,15
pH 10 0,93 494,01 1,45 340,24
pH 11 0,76 520,55 0,93 556,93
Tabel 10.Perbandingan pertumbuhan sel, aktivitas unit dan aktivitas
spesifik antara medium tanpa dan dengan perlakuan variasi komposisi pH medium
Jenis
pH 10 0,93 494,01 340,24 100 100 100
Dengan Perlakuan
Variasi
pH 7 3,87 520,44 529,58 416 105 156
pH 8 3,75 546,86 595,72 403 111 175
pH 9 1,75 467,39 361,15 188 95 106
pH 11 0,76 520,55 556,93 82 105 164 Keterangan: OD = pertumbuhan sel, AU = aktivitas unit dan
Dari data yang disajikan di atas, pH 8 merupakan pH optimum untuk isolat
LTi-21-3 dengan dihasilkan aktivitas unit dan aktivitas spesifik secara
berurutan yaitu 546,86 U/mL dan 595,72 U/mg. Pertumbuhan sel dan
aktivitas spesifik pada pH 8 lebih tinggi dibandingkan pada pH 10 dengan
komposisi medium yang sama. Pada pH 8, pertumbuhan sel pada waktu
inkubasi optimum 36 jam mencapai 3,75 sedangkan pada pH 10 hanya
0,93.
B. Profil Kondisi Optimum Isolat LTi-21-3
Berdasarkan hasil penelitian yang telah dilakukan, kondisi optimum untuk
isolat LTi-21-3 yaitu pada medium kulturHorikoshi’s IIyang mengandung 1%
(b/v) pati singkong, 0,5% (b/v) NH4Cl, 0,02% (b/v) MgSO4dengan pH
medium 8. Berikut merupakan profil pertumbuhan sel dan aktivitas spesifik
isolat LTi-21-3 pada kondisi optimum yang dapat dilihat pada Gambar 26.
Gambar 26. Profil pertumbuhan sel dan produksi enzim CGT-ase isolat
LTi-21-3 padamedium Horikoshi’s II termodifikasi dengan
kondisi optimum
Waktu Inkubasi (Jam) Kondisi Optimum
OD 600 nm
Tabel 11.berikut merupakan perbandingan antara medium Horikoshi’s II
termodifikasi dengan medium variasi terbaik.
Tabel 11. Perbandingan pertumbuhan sel, aktivitas unit, dan aktivitas spesifik
isolat LTi-21-3antara medium Horikoshi’s IItermodifikasi dengan
medium variasi terbaik pada waktu inkubasi optimum 36 jam
Jenis Perlakuan Pertumbuhan Sel OD600
Aktivitas Unit (U/mL)
Aktivitas Spesifik (U/mg)
Nilai/Persentase (%)
OD AU AS
Medium Horikoshi’s II
Termodifikasi 8,10 664,37 253,31 100 100 100
Medium Horikoshi’s II
termodifikasi dengan Variasi Terbaik
3,75 546,86 595,72 46 82 253
Keterangan: OD = pertumbuhan sel, AU = aktivitas unit dan AS = aktivitas spesifik
Dari data di atas, isolat LTi-21-3 dari variasi terbaik menghasilkan aktivitas
spesifik 253% lebih tinggi jika dibandingkan dengan medium Horikoshi’s II
tanpa perlakuan variasi. Dengan aktivitas spesifik yang tinggi ini akan
V. SIMPULAN DAN SARAN
A. Simpulan
Berdasarkan hasil yang diperoleh dari penelitian yang telah dilakukan maka
dapat ditarik simpulan sebagai berikut:
1. Kondisi optimum untuk isolat LTi-21-3 yaitu pada mediumHorikoshi’s II
yang mengandung pati singkong 1% (b/v), 0,5% (b/v) NH4Cl, 0,02% (b/v)
MgSO4dengan pH 8. Nilai aktivitas unit dan spesifik yang dihasilkan
secara berurutan yaitu 546,86 U/mL dan 595,72 U/mg.
2. Pada medium Horikoshi’s II termodifikasi dengan perlakuan variasi
terbaik menghasilkan kenaikan persentase aktivitas spesifiknya sekitar
253% lebih tinggijika dibandingkan pada medium Horikoshi’s II
termodifikasi tanpa perlakuan.
B. Saran
1. Perlu dilakukan identifikasi lebih lanjut terhadap bakteri isolat LTi-21-3
untuk mengetahui jenis dan morfologinya serta perlu diketahui juga
komponen kimia dalam pati singkong yang dapat menghasilkan enzim
2. Kedepan, aplikasi enzim yang diharapkan dari isolat LTi-21-3 adalah untuk
menghasilkan siklodekstrin dari pati singkong yang akan dimanfaatkan
untuk proses pembentukan obat-obatan. Oleh karena itu, perlu dilakukan
tahap pemurnian enzim CGT-ase terhadap isolat LTi-21-3 dengan kondisi