HUBUNGAN ANTARA KEJADIAN PENYAKIT
KLOROSIS DAN KERUPUK DENGAN KEBERADAAN DUA
SPESIES KUTUKEBUL PADA TANAMAN TOMAT
ACEU WULANDARI AMALIA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Hubungan Antara Kejadian Penyakit Klorosis dan Kerupuk dengan Keberadaan Dua Spesies Kutukebul pada Tanaman Tomat adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Maret 2013
Aceu Wulandari Amalia
RINGKASAN
ACEU WULANDARI AMALIA. Hubungan Antara Kejadian Penyakit Klorosis dan Kerupuk dengan Keberadaan Dua Spesies Kutukebul pada Tanaman Tomat. Dibimbing oleh GEDE SUASTIKA, ALI NURMANSYAH, dan DEWI SARTIAMI.
Crinivirus dan Begomovirus telah dilaporkan banyak menimbulkan kerusakan pada berbagai komoditas penting dunia, khususnya pada pertanaman tomat di berbagai negara penghasil tomat. Crinivirus (Tomato infectious chlorosis virus/TICV dan Tomato chlorosis virus/ToCV) menyebabkan penyakit klorosis dan Begomovirus (Tomato yellow leaf curl virus/TYLCV) menyebabkan penyakit kerupuk pada tanaman tomat. TICV dan ToCV ditransmisikan oleh serangga vektor secara semipersisten, sementara itu TYLCV ditransmisikan secara persisten. Penyakit ini sudah ditemukan di Indonesia, terutama di Jawa Barat. Informasi tentang kedua penyakit tersebut yang menginfeksi tanaman tomat masih sangat terbatas. Penyebaran kedua penyakit pada tanaman tomat berdasarkan kelimpahan serangga vektor di berbagai ketinggian tempat dianalisis pada penelitian ini.
Penelitian terdiri atas tiga kegiatan utama, yaitu (1) survei kejadian penyakit yang dilakukan di areal pertanaman tomat di beberapa ketinggian tempat di Jawa Barat mulai dari 100 sampai dengan 1500 meter di atas permukaan laut (mdpl), dengan mengamati variabel-variabel berikut: (a) kejadian penyakit, (b) kondisi iklim (suhu dan kelembaban nisbi), (c) kelimpahan populasi Trialeurodes vaporariorum dan Bemisia tabaci, dan (d) kondisi tanaman, seperti umur tanaman, varietas tomat, karakteristik budidaya dan penggunaan pestisida; (2) deteksi virus dan identifikasi kutukebul, dan (3) analisis hubungan kejadian penyakit dengan faktor-faktor yang memengaruhinya.
Gen protein selubung minor Crinivirus sebesar 360 bp dan 417 bp serta gen protein selubung Begomovirus sebesar 780 bp dari sampel tanaman bergejala pada seluruh ketinggian tempat yang dikaji telah berhasil diamplifikasi dengan PCR. Beberapa stadia pertumbuhan T. vaporariorum dan B. tabaci yang ditemukan saat pengamatan telah berhasil diidentifikasi. Penyakit klorosis dan kerupuk ditemukan menginfeksi tanaman tomat di ketinggian tempat di atas 1200 mdpl sampai dengan 1500 mdpl. Juga, B. tabaci ditemukan menyerang tanaman tomat pada ketinggian tempat tersebut yang sebelumnya hanya ditemukan di bawah ketinggian 1200 mdpl. Hasil analisis regresi memperlihatkan adanya korelasi positif antara kejadian penyakit klorosis dan kerupuk dengan keberadaan kedua spesies kutukebul dan peningkatan ketinggian tempat. Faktor lain seperti kelembaban nisbi, varietas tomat, dan umur tanaman tidak memberikan kontribusi yang signifikan terhadap perubahan kejadian kedua penyakit tersebut.
Kata kunci: Begomovirus, Bemisia tabaci, Crinivirus, ketinggian tempat,
SUMMARY
ACEU WULANDARI AMALIA. Relationship between Disease Incidence of Chlorosis and Leaf Curl and Two Species of Whitefly in Tomato Crop. Under direction of GEDE SUASTIKA, ALI NURMANSYAH, and DEWI SARTIAMI.
Crinivirus and Begomoviruses have been reported to cause serious diseases in a number of economic crops in the world, especially in tomato crop. Crinivirus
(Tomato infectious chlorosis virus/TICV and Tomato chlorosis virus/ToCV) causes chlorosis while Begomovirus (Tomato yellow leaf curl virus/TYLCV) causes curl diseases. TICV and ToCV are transmitted by vector insects in semipersistent, but TYLCV in persistent manner. The diseases have been found in Indonesia, especially in West Java. Information about the infecstation of both diseases in tomato crops are still limited. The spread of both diseases on tomato crops based on vector insect abundance in various altitude in Indonesia was evaluated in this study.
The study consists of three main activities: (1) a field survey on disease incidence conducted in various altitudes from 100 to 1500 meters above sea level (m asl), which observed variables as follows: (a) diseases incidence, (b) climate condition (temperature and relative humidity (RH)), (c) abundaces of
Trialeurodes vaporariorum and Bemisia tabaci, and (d) cropping condition, such as age of plant, varieties, and planting system; (2) the detection and identification of viruses and whiteflies; and (3) the analysis of the relationship between disease incidence and its determinant factors.
Minor coat protein gene of Crinivirus (360 bp and 417 bp) and coat protein gene of Begomovirus (780 bp) from symptomed samples in all studied altitudes were successfully amplified by PCR. Some stadia of T. vaporariorum and B. tabaci found during observations had also been identified. The chlorosis and curl diseases infected tomato crops in altitude above 1200 m asl until 1500 m asl. Also, B. tabaci was found at those altitudeswhich previously only found at altitudes below 1200 m asl. The result of regression analysis showed there were positive correlations between the incidence of chlorosis and curl diseases with the presence of the two whitfly spesieses and the increase in altitude. Other factors such as relative humidity, tomato varieties, and plant age did not show significant contributions to the change in the incidence of both diseases.
© Hak Cipta Milik IPB, Tahun 2011
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada Mayor Fitopatologi Departemen Proteksi Tanaman
HUBUNGAN ANTARA KEJADIAN PENYAKIT
KLOROSIS DAN KERUPUK DENGAN KEBERADAAN DUA
SPESIES KUTUKEBUL PADA TANAMAN TOMAT
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
PRAKATA
Alhamdulillahhirabil’alamin. Puji dan syukur penulis panjatkan kepada Allah SWT karena hanya atas ridhonya dan karuniaNYA tesis dengan judul Hubungan Antara Kejadian Penyakit Klorosis dan Kerupuk dengan Keberadaan Dua Spesies Kutukebul pada Tanaman Tomat ini dapat diselesaikan dengan baik.
Terima kasih penulis ucapkan kepada Dr Ir Gede Suastika, Dr Ir Ali Nurmansyah, dan Dra Dewi Sartiami atas kesempatan dan kepercayaan yang diberikan kepada penulis serta segala bimbingan yang diberikan dengan setulus hati kepada penulis demi lahirnya sebuah karya tesis ini.
Terimakasih penulis sampaikan kepada: Dr Ir Sri Hendrastuti Hidayat, Saudari Tuti S. Legiastuti dan Ibu Aisyah yang telah banyak memberikan saran, nasihatnya dan pengarahan kepada penulis selama bekerja di Laboratorium Virologi dan Laboratorium Biosistematika Serangga, Departemen Proteksi Tanaman, Fakultas Pertanian, IPB. Di samping itu terima kasih kepada Dr Trijoko Santoso, Dr Ifa Manzila dan Saudari Fitrianingrum Kuriawati yang telah memberikan pengarahan bagi penulis selama bekerja di Lab Biologi Molekuler, BB-BIOGEN (Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian) Bogor, serta Saudari Dwi Astuti atas bantuannya saat penulis bekerja di Laboratorium Biologi Molekuler, LIPI (Lembaga Ilmu Pengetahuan Indonesia).
Terimakasih penulis ucapkan untuk semua anggota Laboratorium Virologi dan Biosistematika Serangga, kepada bapak/ibu staff kepegawaian dan laboratorium HPT yang selalu mendukung penulis agar tetap semangat, teman-teman Fitopatologi angkatan 2008, 2009, 2010 dan 2011, HPT’41, adik-adik HPT, JICA serta semua sahabat untuk kebersamaan dan dukungan moril selama ini kepada penulis. Semoga Allah senantiasa melimpahi kesuksesan dan kebahagiaan bagi kita semua. Amin.
Rasa terimakasih yang tiada henti berikut penghargaan yang tak terhingga penulis persembahkan kepada ayahanda Ir Ahmad Husen, ibunda Tutin Suryatin BSCf, kakak dan adik-adikku Susan Irmayani, Feri Ferdiansyah Solihin dan Fajar Ramadhani Hikmatullah, keluarga besar H. Nawi dan Hj. Siti Sadiah (Bandung) serta H. Furqon dan Hj. Djadja Tedjaningsih (Garut) atas limpahan cinta, kasih sayang serta doa sepenuh hati kepada penulis demi terselesaikan tesis ini. Semoga Allah SWT senantiasa memberikan ridhoNya bagi kita semua dunia dan akhirat dalam ikatan kekeluargaan yang indah selamanya. Amin.
Pada akhirnya penulis berharap semoga karya ini mampu berkontribusi bagi kemajuan dan kearifan ilmu pengetahuan di masa yang akan datang. Amin.
Bogor, Maret 2013
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 2
Manfaat Penelitian 3
Ruang Lingkup Penelitian 3
Kerangka Alur Penelitian 4
2 TINJAUAN PUSTAKA 5
Interaksi Organisme Pengganggu Tanaman dan Tanaman Tomat 5
Bioekologi Crinivirus (ToCV dan TICV) 5 Bioekologi Begomovirus (TYLCV) 7 Bioekologi Trialeurodes vaporariorum dan Bemisia
tabaci 8
Faktor Penentu Kejadian Penyakit Virus 9 Deteksi Asam Nukleat dengan Polymerase Chain Reaction (PCR) 11
3 METODE 13
Tempat dan Waktu Penelitian 13
Metode Penelitian 13
Survei Kejadian Penyakit dan Pengambilan Sampel 13 Deteksi Keberadaan Virus 13 Ekstraksi RNA Total TICV dan ToCV 14 Prosedur RT-PCR dan PCR TICV dan ToCV 14 Ekstraksi DNA Total TYLCV 15 Prosedur PCR DNA TYLCV 15 Identifikasi dan Penghitungan Populasi Kutukebul 16 Pembuatan Preparat Mikroskop dan Identifikasi
Kutukebul 16
Analisis Hubungan Kejadian Penyakit dan Kelimpahan Vektor 17
4 HASIL DAN PEMBAHASAN 18
Insiden Begomovirus dan Crinivirus pada Tanaman Tomat 18 Gejala Penyakit 18 Verifikasi Infeksi Virus 19 Kejadian Penyakit oleh Begomovirus dan Crinivirus 21 Kutukebul pada Pertanaman Tomat 22 Spesies Kutukebul 22 Kelimpahan Populasi Kutukebul 24
Hubungan Kejadian Penyakit oleh Crinivirus dan Begomovirus
dengan Kelimpahan Populasi Kutukebul
25
Pembahasan Umum 28
DAFTAR PUSTAKA 33
LAMPIRAN 38
DAFTAR TABEL
1 Kejadian penyakit Crinivirus dan Begomovirus di lapangan 21 2 Populasi kutukebul di pertanaman tomat di Jawa Barat 24 3 Koefisien model dan hasil uji kejadian penyakit 26
DAFTAR GAMBAR
1 Kerangka alur penelitian 4
2 Gejala Crinivirus di lahan pertanaman tomat 6
3 Organisasi genom TICV dan ToCV 6
4 Gejala penyakit Begomoviruspada tanaman tomat 7 5 Imago B. tabaci dan T. vaporariorum 8 6 Gejala Begomovirus dan Crinivirus saat pengamatan survei 18
7 Hasil amplifikasi DNA Crinivirus 20
8 Hasil amplifikasi DNA Begomovirus 21 9 Morfologi kutukebul T. vaporariorum dan B. tabaci 22
10 Identifikasi pupa kutukebul 23
DAFTAR LAMPIRAN
1 Hasil analisis regresi berganda kejadian penyakit Crinivirus terhadap ketinggian tempat dan kelimpahan T. vaporariorum 38 2 Plot kenormalan kejadian penyakit Crinivirus terhadap ketinggian
tempat dan kelimpahan T. vaporariorum 39 3 Plot residual versus fits kejadian penyakit Crinivirus terhadap ketinggian
tempat dan T. vaporariorum 39
4 Plot histogram kejadian penyakit Crinivirus terhadap ketinggian tempat dan kelimpahan T. vaporariorum imago B. tabaci dan T. vaporariorum 40 5 Plot residu versus order kejadian penyakit Crinivirus terhadap
ketinggian tempat dan kelimpahan T. vaporariorum 40 6 Hasil analisis regresi berganda kejadian penyakit Begomovirus terhadap
ketinggian tempat, kelimpahan B. tabaci, dan umur tanaman 41 7 Plot kenormalan kejadian penyakit Begomovirus terhadap ketinggian
tempat, kelimpahan B. tabaci, dan umur tanaman 42 8 Plot residual versus fits kejadian penyakit Begomovirus terhadap
ketinggian tempat, kelimpahan B. tabaci, dan umur tanaman 42 9 Plot residual histogram kejadian penyakit Begomovirus terhadap
ketinggian tempat, kelimpahan B. tabaci, dan umur tanaman 43 10 Plot residual histogram kejadian penyakit Begomovirus terhadap
ketinggian tempat, kelimpahan B. tabaci, dan umur tanaman 43 11 Hasil analisis regresi berganda kejadian penyakit virus ganda terhadap
ketinggian tempat, kelimpahan T. vaporariorum,kelimpahan B. tabaci,
12 Plot kenormalan kejadian penyakit virus ganda terhadap ketinggian tempat, kelimpahan T. vaporariorum, kelimpahan B. tabaci, dan umur
tanaman 45 13 Plot residual versus fits kejadian penyakit virus ganda terhadap
ketinggian tempat, kelimpahan T. vaporariorum,kelimpahan B. tabaci,
dan umur tanaman 45
14 Plot residual histogram kejadian penyakit virus ganda terhadap ketinggian tempat, kelimpahan T. vaporariorum,kelimpahan B. tabaci,
dan umur tanaman 46
15 Plot residual versus order kejadian penyakit virus ganda terhadap ketinggian tempat, kelimpahan T. vaporariorum,kelimpahan B. tabaci,
1
PENDAHULUAN
Latar Belakang
Crinivirus dan Begomovirus adalah dua kelompok virus yang
penyebarannya di lapangan sangat cepat dan telah dilaporkan banyak menimbulkan kerusakan pada berbagai komoditas penting dunia (Wisler et al. 1998). Beberapa tahun terakhir, ini kedua virus tersebut telah menjadi kendala penting bagi tanaman hortikultura dan menginfeksi tanaman khususnya pada pertanaman tomat di berbagai negara penghasil tomat di dunia (Dalmon et al. 2008; Louro et al. 2000). Crinivirus merupakan kelompok virus dalam famili
Closteroviridae yang mempunyai genom berupa dua jenis RNA (bipartite) (Wisler et al. 1998), sedangkan Begomovirus adalah salah satu genus dalam famili
Geminiviridae yang bergenom DNA (bipartite) (Tsai et al. 2006).
Tomato chlorosis virus (ToCV) dan Tomato infectious chlorosis virus
(TICV) adalah dua virus anggota Crinivirus yang dilaporkan menginfeksi tanaman tomat. Kedua spesies virus ini dapat menginfeksi tanaman tomat secara tunggal maupun ganda, sehingga dalam gejalanya seringkali tidak dapat dibedakan. Ciri khas dari penyakit klorosis yang ditimbulkannya adalah antara tulang daun menguning (interveinal yellowing), daun menjadi rapuh (leaf brittleness), adanya nekrotik yang menjadi merah keunguan (bronzing), kebugaran (vigor) tanaman berkurang, buah berukuran lebih kecil dari ukuran normal dan tanaman mudah gugur (early senescence) (Duffus et al. 1996; Dalmon
et al. 2008). Begomovirus yang diidentifikasi menyerang tanaman tomat adalah
Tomato yellow leaf curl virus (TYLCV). Pada tanaman tomat, virus ini menimbulkan penyakit kerupuk karena daun-daun yang tumbuh setelah terinfeksi mengalami pertumbuhan terhambat sehingga berukuran lebih kecil dan mengalami malformasi melengkung ke atas menyerupai kerupuk. Gejala lain yang mengikuti penyakit kerupuk ini adalah penebalan daun, lamina daun berkerut,daun menguning, tepi daun melengkung ke atas, daun keriting dan tanaman kerdil (Sugiarman dan Hidayat 2000).
Di Indonesia, insiden penyakit yang disebabkan oleh Crinivirus maupun
Begomovirus sudah sangat luas dan sampai saat ini telah dilaporkan ditemukan di sentra-sentra produksi tomat di daerah Jawa Barat, Jawa Tengah, Yogyakarta, Bali, dan Lombok (Sudiono et al. 2004; Hartono dan Wijonarko 2007; Suastika 2010). Epidemi kedua virus ini ditenggarai berhubungan dengan aktivitas serangga vektornya. Crinivirus ataupun Begomovirus ditularkan oleh kutukebul masing-masing secara semi persisten dan persisten (Duffus et al. 1994; Wisler et al. 1996; Wintermantel 2004). Kutukebul yang efektif menjadi vektor kedua virus ini adalah Bemisia tabaci dan Trialeurodes vaporariorum. Kedua spesies kutukebul ini mempunyai daya adaptasi yang berbeda terhadap lingkungan terutama terhadap temperatur udara (Xie et al. 2006). Kutukebul B. tabaci,
2
Populasi vektor yang tinggi pada pertanaman tomat di lapangan dapat menimbulkan terjadinya epidemik penyakit klorosis ataupun kerupuk. Faktor-faktor lain yang mendukung terjadinya epidemik penyakit klorosis ataupun kerupuk pada pertanaman tomat antara lain jenis tanaman tomat yang rentan, virus yang virulen, lingkungan yang mendukung untuk perkembangan penyakit seperti: suhu, kelembaban udara, dan teknik budidaya tanaman tomat secara monokultur dan terus-menerus (Wisler et al. 1998).
Informasi mengenai distribusi kejadian penyakit klorosis dan kerupuk pada tanaman tomat di Indonesia yang masing-masing disebabkan oleh Crinivirus dan
Begomovirus masih sangat terbatas. Penelitian untuk mempelajari kelimpahan kutukebul yang berasosiasi dengan penyakit klorosis di berbagai ketinggian tempat di Jawa Barat perlu dilakukan, mengingat Jawa Barat merupakan salah satu sentra produksi tomat di Indonesia dengan ketinggian tempat yang beragam (100 - > 1200 mdpl).
Perumusan Masalah
Penyakit klorosis oleh Crinivirus dan penyakit kerupuk oleh Begomovirus
dilaporkan sebagai virus yang berperan menyebabkan penurunan produksi yang sangat besar di pertanaman tomat baik di daerah sub tropis maupun tropis termasuk Indonesia. Kedua penyakit tersebut juga memiliki daerah penyebaran yang sangat luas akibat peran kutukebul sebagai serangga vektornya. Kelompok
Crinivirus yang menyerang tanaman tomat teridentifikasi sebagai ToCV dan TICV, sedangkan kelompok Begomovirus adalah TYLCV. Penyebaran ketiga
virus tersebut sangat cepat karena ditularkan dengan serangga vektor
T. vaporariorum dan B. tabaci. Kedua penyakit tersebut telah tersebar luas di Indonesia dengan kejadian penyakit sampai dengan 100%.
Salah satu upaya dalam pencegahan perkembangan penyakit tersebut adalah dengan melakukan pengendalian terhadap serangga vektornya. Pengendalian tersebut dengan cara menekan populasi kutukebulnya. Hal tersebut diperlukan pengetahuan mengenai habitat dan kondisi yang kondusif bagi perkembangan hidup kutukebul. Selain itu lingkungan yang mendukung bagi perkembangan penyakit dan tanaman inang juga sangat diperlukan. Dalam penelitian ini dilakukan pengamatan faktor-faktor berpengaruh terutama faktor ketinggian tempat serta analisis hubungan kejadian penyakit dengan kelimpahan populasi kutukebul.
Tujuan Penelitian
3 Manfaat Penelitian
Manfaat penelitian ini adalah memberikan informasi tentang daerah sebaran penyakit klorosis (Crinivirus) dan penyakit kerupuk (Begomovirus) serta populasi kutukebul Trialeurodes vaporariorum dan Bemisia tabaci pada pertanaman tomat di berbagai ketinggian tempat di Jawa Barat.
Ruang Lingkup Penelitian
Perkembangan penyakit yang disebabkan oleh Crinivirus (ToCV dan TICV) dan Begomovirus (TYLCV) pada pertanaman tomat meningkat secara cepat di Indonesia. Hal tersebut memerlukan upaya dalam menentukan faktor-faktor yang secara signifikan berpengaruh terhadap penyebarannya serta untuk mengetahui habitat vektor sebagai awal dari pencegahan terjadinya epidemi penyakit.
4
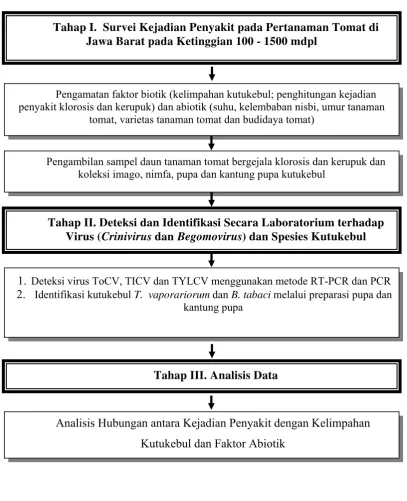
Tahap I. Survei Kejadian Penyakit pada Pertanaman Tomat di Jawa Barat pada Ketinggian 100 - 1500 mdpl
Pengamatan faktor biotik (kelimpahan kutukebul; penghitungan kejadian penyakit klorosis dan kerupuk) dan abiotik (suhu, kelembaban nisbi, umur tanaman
tomat, varietas tanaman tomat dan budidaya tomat)
Pengambilan sampel daun tanaman tomat bergejala klorosis dan kerupuk dan koleksi imago, nimfa, pupa dan kantung pupa kutukebul
Tahap II. Deteksi dan Identifikasi Secara Laboratorium terhadap Virus (Crinivirus dan Begomovirus) dan Spesies Kutukebul
Tahap III. Analisis Data
Analisis Hubungan antara Kejadian Penyakit dengan Kelimpahan Kutukebul dan Faktor Abiotik
1. Deteksi virus ToCV, TICV dan TYLCV menggunakan metode RT-PCR dan PCR 2. Identifikasi kutukebul T. vaporariorum dan B. tabaci melalui preparasi pupa dan
kantung pupa
Keterangan :
= Kegiatan induk utama penelitian = Aktivitas penelitian
= Alur kegiatan
5
2
TINJAUAN PUSTAKA
Interaksi Organisme Pengganggu Tanaman dan Tanaman Tomat Kendala utama pada budidaya tanaman hortikultura termasuk tanaman tomat adalah organisme pengganggu tanaman (OPT) yang terdiri dari hama, penyakit dan gulma. Pada dasarnya penyakit maupun hama hanya dapat terjadi jika adanya tiga faktor yang saling mendukung yaitu patogen, inang dan lingkungan yang berawal dari konsep Ilmu Penyakit Tumbuhan. Adanya perubahan dari salah satu komponen akan berpengaruh terhadap intensitas penyakit yang muncul. Inang dalam keadaan rentan, patogen bersifat virulen (daya infeksi tinggi) dan jumlah yang cukup, serta lingkungan yang mendukung. Lingkungan berupa komponen lingkungan fisik (suhu, kelembaban, cahaya) maupun biotik (musuh alami, organisme kompetitor).
Penyakit penting tanaman tomat adalah penyakit rebah kecambah (Rhizoctonia solani Kuhn), penyakit bercak daun Septoria (Septoria lycopersici
Speg), penyakit bercak daun (Alternaria solani Ell.), penyakit busuk daun (Phytophthora infestans (Mont) de Bary.), penyakit layu cendawan (Verticillium
albo-atrum Reinke & Berth.), penyakit layu fusarium (Fusarium oxyporum
Schlecht.), penyakit layu bakteri (Pseudomonas solanacearum Smith.), penyakit mosaik yang disebabkan oleh salah satu atau gabungan berbagai jenis virus seperti Tomato infectious chlorosis virus (TICV) dan Tomato chlorosis virus
(ToCV), virus tomat mosaik (tomato mosaic virus: ToMV), virus mosaik tembakau (tobbaco mosaic virus: TMV), virus mosaik ketimun (cucumber mosaic virus: CMV), virus kentang Y (potato virus Y: PVY) dan virus X kentang (potato virus X: PVX), penyakit kuning dan daun menggulung (Tomato yellow leaf curl virus: TYLCV), dan penyakit bengkak akar (Meloidogyne incognita). Hama penting yang menyerang tanaman tomat adalah ulat buah (Helicoverpa armigera
Hubn.), ulat tanah (Agrotis ipsilon Hufn.), kutukebul (Bemisia tabaci Genn. dan
Trialeurodes vaporariorum), ulat grayak (Spodoptera litura F.), dan penggorok daun (Liriomyza sp.) (Setiawati 1990).
Beberapa tahun terakhir ini penyakit yang menjadi kendala penting bagi pertanaman tomat di berbagai negara penghasil tomat di dunia adalah penyakit yang disebabkan oleh virus yaitu Crinivirus dan Begomovirus (Dalmon et al. 2008; Louro et al. 2000) dan juga menjadi ancaman penting karena ditransmisikan oleh serangga vektor. Adapun spesies virus yang termasuk ke dalam anggota kedua genus tersebut yang menginfeksi tomat adalah Tomato infectious chlorosis virus (TICV) dan Tomato chlorosis virus (ToCV) merupakan Crinivirus dan
Tomato yellow leaf curl virus (TYLCV) merupakan Begomovirus. Crinivirus
ditransmisikan kutukebul Trialeurodes vaporariorum dan Begomovirus
ditransmisikan kutukebul Bemisia tabaci.
Bioekologi Crinivirus (Tomato chlorosis virus/ToCV dan Tomato infectious chlorosis virus/TICV)
6
perkembangan selanjutnya daun-daun menjadi rapuh (leaf brittleness), mengalami nekrotik pada beberapa bagian dan warna bagian yang nekrotik menjadi merah keunguan (bronzing), kebugaran (vigor) tanaman menjadi sangat berkurang, dan apabila menghasilkan buah maka ukurannya jauh lebih kecil dari normal dan proses pematangannya terganggu, serta mudah gugur (early senescence) sehingga sangat menurunkan bahkan meniadakan nilai ekonomi tanaman yang terinfeksi (Duffus et al. 1996; Dalmon et al. 2008). Gejala terbatas pada jaringan floem dengan konsentrasi virus rendah dimana hal tersebut salah satu kendala untuk diagnosis keberadaan virus.
Gambar 2 Gejala klorosis di lahan pertanaman tomat (koleksi pribadi)
Partikel TICV dan ToCV berbentuk seperti benang, memanjang (filamentous) dan lentur (flexuous). Partikel TICV memiliki panjang 850 – 900 nm (Duffus et al. 1996, Liu et al. 2000), sedikit lebih panjang dibandingkan ToCV yaitu 800-850 nm. Keduanya masing-masing memiliki dua jenis genom (bipartite) berupa RNA utas tunggal yaitu RNA 1 dan RNA 2 (Gambar 3a). RNA 1 dan RNA 2 TICV berukuran 7.8 dan 7.4 kb, dan ToCV berukuran 7.8 dan 8.2 kb (Wintermantel et al. 2005). RNA 1 pada TICV dan ToCV berfungsi dalam mengkode dua jenis protein yang terlibat dalam replikasi virus (Martelli et al.
2000). a)
b)
Gambar 3 Organisasi genom TICV RNA 1 dan RNA 2 (a) dan ToCV sekuen RNA 2 (b) (Lozano et al. 2005)
Selubung protein minor (CPm) pada TICV dan ToCV yang membentuk bagian ujung atau ekor virion memiliki peranan dalam penularan dengan kutukebul. CPm dari kedua virus tersebut memiliki kespesifikan dengan reseptor
7 vektornya sangat ditentukan oleh reseptor yang ada pada stilet serangga dengan CP dari virus yang bersangkutan (Wintermantel 2006). Duffus et al. (1994) dan Wisler et al. (1996) melaporkan bahwa penularan virus TICV oleh kutukebul
T.vaporariorum secara semi-persisten selama 4 hari, dengan periode retensi sampai 2 hari (Wisler et al. 1998a).
Bioekologi Begomovirus (Tomato yellowing leaf curl virus/TYLCV)
Gejala yang ditimbulkan oleh TYLCV adalah khlorosis yang muncul dari tunas daun kemudian menyebar ke daun-daun bagian bawah disertai gejala keriting atau berbentuk seperti mangkok (cupping), keras, daun berkerut (puckering), bunga rontok dan terdapat enasi pada permukaan bawah daun juga khlorotik (Butter dan Rataul 1977; Jones et al. 1991). Menurut Sugiarman dan Hidayat (2000), gejala dari infeksi TYLCV di Indonesia pada umumnya menimbulkan penebalan daun, lamina daun berkerut, daun menguning, tepi daun melengkung ke atas, daun keriting dan tanaman kerdil.
Gambar 4 Gejala penyakit pada tanaman tomat yang disebabkan oleh TYLCV (koleksi pribadi)
Partikel TYLCV berbentuk isometrik ganda, yang dalam keadaan tunggal berdiameter 18-20 nm dan dalam keadaan berpasangan berdiameter 20-30 nm (Thsai et al. 2006). Virus ini memiliki genom bipartite dan memiliki asam nukleat single stranded DNA (ss-DNA). Genom berukuran 2.6-2.8 kb yang terselubung dalam virion ekosahedral kembar (gemini).
Vektor yang menyebarkan penyakit kuning ini adalah B. tabaci yang mendapatkan virus dengan menghisap tanaman tomat yang terinfeksi selama 15 sampai 30 menit. Vektor menularkan secara persisten. Vektor dapat mentransmisi virus ke tanaman sehat memerlukan waktu 15 menit setelah 24 jam inkubasi (setelah periode makan akuisisi). Adapun hasil penelitian dari Uzcategui dan Lastra (1978) menunjukkan bahwa periode makan akuisisi minimum B. tabaci
8
Bioekologi Kutukebul T. vaporariorum dan B. tabaci
Kutukebul T. vaporariorum dan B. tabaci termasuk ke dalam famili Aleyrodidae, superfamili Aleyrodoidea, ordo Hemiptera (Caerver et al. 1991). Kutukebul baik nimfa maupun imago memiliki lapisan lilin yang dapat digunakan sebagai dasar identifikasi karena penampilan dan pola lapisan lilin dapat membedakan antara satu spesies dengan spesies lainnya (Botha et al. 2000). Semua stadia kutukebul hidup dan makan di bawah permukaan daun hasil ekskresinya adalah embun madu yang dapat menjadi media untuk cendawan
Capnodium sp. atau embun jelaga (Hoddle 2004). Siklus hidup serangga ini dimulai dari telur, nimfa, pupa, dan imago. Cara reproduksi dengan cara seksual atau partenogenesis. Telur diletakkan di permukaan daun dan menetas menjadi nimfa instar pertama, kemudian akan bergerak untuk mencari tempat menghisap makanan yang sesuai dan menetap disana selama fase nimfa. Kutukebul menghentikan aktivitas makan pada stadia akhir (puparium). Kulit pupa akan terbentuk dan menuju proses pembentukan menjadi imago (Kalshoven 1981). Stadia nimfa dan imago merupakan stadia yang menyebabkan kerusakan tanaman (Morales 2001).
Gambar 5 Imago B. tabaci (A) dan T. vaporariorum (B) (koleksi pribadi) Kutukebul ini memiliki bentuk alat mulut menusuk-menghisap yang berisi empat struktur tubular yang dinamakan stilet. Melalui stilet inilah penularan virus ke tanaman inang terjadi. Proses penempelan partikel virus dimulai adanya interaksi antara protein selubung virus dengan alat mulut kutukebul. Selanjutnya partikel virus akan menempel pada reseptornya yang terletak di stilet tersebut. Virus akan berada dalam tubuh vektor saat diakusisi kemudian menuju sel epitel saluran pencernaan dan berasosiasi dengan kelenjar saliva serangga vektor. Virus bersirkulasi dalam tubuh serangga sampai akhirnya virus mencapai stilet dan masuk ke dalam tanaman sehat saat vektor makan cairan nutrisi di floem. Virus memerlukan waktu akuisisi dan inokulasi satu jam hingga satu hari dan periode laten satu hari hingga beberapa minggu dalam tubuh serangga (Gray dan Banerjee 1999).
Imago betina B. tabaci hidup sekitar 6 hari sampai 60 hari. Lama hidup imago jantan lebih pendek sekitar 9-17 hari. Imago bertahan hidup pada suhu 14-35°C, sedangkan pada tanaman tomat imago mampu bertahan sampai suhu 30°C selama sekitar 15 hari dan menyerang tanaman pada musim kemarau (Smith
9 2009). Cohen dan Berlinger (1986) menyatakan bahwa serendah apapun populasi
B. tabaci cukup efektif menyebabkan kerusakan langsung maupun tidak langsung. Imago T. vaporariorum meletakkan telur dan hidup di daun muda dekat titik tumbuh tanaman. Imago betina mampu bertelur sampai 250 butir dengan siklus hidup 30 sampai 45 hari (Smith 2009). Durasi perkembangan berbagai stadia
T. vaporariorum pada tanaman tomat adalah sebagai berikut: stadium telur adalah sekitar 8 hari, lama perkembangan nimfa instar 1 sekitar 6 hari, nimfa instar 2 sekitar 2 hari, nimfa instar 3 sekitar 3 hari, lama stadium pupa adalah 9 hari dan lama hidup imago rata-rata 7 sampai 8 hari (Roermund dan Lenteren 1992). Setiap fase perkembangan kutukebul T. vaporariorum dapat ditemukan pada lingkungan bersuhu dingin (Xie et al. 2006), dan Smith (2009) mengatakan fase imago dapat bertahan hidup pada suhu 22-25°C. Fase imago tidak mampu berkembang pada suhu 30°C dan akan mati pada suhu di atas 35°C (Smith 2009).
Faktor Penentu Kejadian Penyakit Virus
Perkembangan organisme pengganggu tanaman dipengaruhi oleh lingkungan sekitar. Topografi dan faktor iklim sangat menentukan keberlangsungan siklus hidup baik dari hama serangga maupun penyakitnya. Faktor iklim baik secara langsung maupun tidak langsung di antaranya temperatur, kelembaban relatif udara dan curah hujan, memiliki pengaruh yang berbeda untuk tiap spesies serangga dan dampak secara langsung dapat terlihat pada siklus hidup, keperidian, lama hidup, serta kemampuan diapause serangga (Ganaha et al. 2007; Lastuvka 2009). Keragaman iklim dapat mempengaruhi pertumbuhan populasi dan penyebaran serangga sehingga dalam kurun waktu singkat dapat menimbulkan ledakan populasi serangga hama tertentu (Wiyono 2007; Dale 1994). Serangga-serangga hama yang kecil seperti kutu-kutuan umumnya menjadi masalah pada musim kemarau atau rumah kaca karena tidak ada terpaan air hujan. Seiring dengan timbulnya ledakan populasi serangga hama itupun dapat meningkatkan kejadian penyakit sebagai akibat dari serangga hama yang berperan sebagai vektor, terutama penyakit virus. Pengaruh faktor iklim terhadap patogen pun dapat berpengaruh terhadap siklus hidup patogen, virulensi (daya infeksi), penularan, dan reproduksi patogen. Pengaruh perubahan iklim akan sangat spesifik untuk masing masing penyakit. Perubahan iklim berpengaruh terhadap penyakit melalui pengaruhnya pada tingkat genom, seluler, proses fisiologi tanaman dan patogen (Garret et al. 2006).
10
keadaan suhu rendah maupun suhu tinggi (Leather dan Awmack 1998). Keterangan tersebut telah dibuktikan pada beberapa studi tentang adanya keterikatan antara suhu dengan perkembangan hidup serangga yang selama ini telah dikembangkan diantaranya adalah studi oleh Xie et al. (2006) dan Smith (2009) tentang perkembangan hidup kutukebul. Menurut Xie et al. (2006), perkembangan optimum dan kemampuan bertahan kutukebul berbeda-beda. Lebih lanjut Xie et al. (2006) melaporkan bahwa B. tabaci dan T. vaporariorum
bisa beradaptasi pada suhu 2°C sampai dengan diatas 24°C. Selanjutnya, Xie et al.
(2006) menyatakan bahwa pada dasarnya semua stadium T. vaporariorum
memiliki kemampuan beradaptasi di suhu dingin. T.vaporariorum mampu bertahan di suhu -8°C selama 10 jam, kelangsungan hidupnya 78% pada suhu 5°C selama 2 hari dan berkembang baik pada suhu 15°-18°C, sedangkan menurut Smith (2009), suhu optimum untuk perkembangan T. vaporariorum adalah 20°C - 25°C. Imago masih dapat berkembang pada suhu di bawah 8°C dan akan berhenti meletakkan telur pada suhu di bawah 7°C (Smith 2009). Adapun suhu optimum untuk perkembangan B. tabaci adalah diatas 24°C (Xie et al. 2006). Menurut Smith (2009), B. tabaci cenderung lebih menyukai suhu yang lebih hangat dibandingkan dengan T. vaporariorum. Perkembangan B. tabaci terjadi di kisaran 14-35°C, tetapi optimum pada suhu 25-30°C. Pada lahan tomat yang bersuhu 30°C, waktu perkembangan dari mulai telur sampai dewasa ialah pendek yaitu selama 18 hari. Imago B. tabaci hidup selama 10-15 hari pada suhu 28-30°C. Pada suhu 25°C, B. tabaci betina memproduksi sekitar 195 telur dalam hidupnya. Pada suhu 16-19°C, perkembangan B. tabaci sangatlah lambat, dengan siklus hidup selama 140-70 hari berturut-turut, dan tingkat kematiannya sangat tinggi. Nimfa mati pada suhu 9°C dan kelembaban udara yang rendah. Selama musim dingin, imago B. tabaci dapat bertahan beberapa minggu tanpa tanaman inang atau di dalam sebuah rumah kaca yang kosong.
Salah satu hubungan penyakit dengan keberadaan serangga vektornya yang terpenting adalah peningkatan kejadian penyakit Crinivirus dan Begomovirus atau virus gemini yang disebabkan oleh ledakan populasi kutukebul T. vaporariorum
11 Deteksi Asam Nukleat dengan Polymerase chain reaction (PCR)
Reverse transcriptase-polymerase chain reaction (RT-PCR) tidak dapat dilakukan dengan menggunakan RNA sebagai cetakan maka terlebih dahulu dilakukan proses transkripsi balik (reverse transcriptation) terhadap molekul mRNA sehingga diperoleh molekul cDNA (complemantery DNA). Molekul cDNA tersebut kemudian digunakan sebagai cetakan dalam proses PCR. RT-PCR memerlukan enzim reverse transkiptase. Enzim ini adalah enzim DNA polimerase yang menggunakan molekul RNA sebagai cetakan untuk menyintesis molekul DNA (cDNA) yang komplementer dengan molekul RNA tersebut. Beberapa enzim reverse transkiptase yang dapat digunakan adalah mesophilic viral reverse transcriptase (RTase) yang dikodekan oleh virus avian myoblastosis (AMV) maupun oleh moloney murine leukimia (M-MuL V), dan Tht DNA polimerase. Menurut Yuwono (2005) Reaksi trankripsi balik dapat dilakukan dengan menggunakan beberapa macam primer yaitu Oligo dT (sepanjang 12 – 18 nukleotida yang akan melekat pada ekor poli-(A) pada ujung 3’ mRNA mamalia primer semacam ini pada umumnya menghasilkan cDNA yang lengkap; Heksanukleotida acak yang akan melekat pada cetakan mRNA yang komplementer pada bagian manapun. Primer semacam ini akan menghasilkan cDNA yang tidak lengkap (parsial); dan yang ketiga adalah urutan nukleotida spesifik yang dapat digunakan secara selektif untuk menyalin mRNA tertentu.
12
13
3
METODE
Tempat dan Waktu
Penelitian terdiri atas dua kegiatan utama, yaitu (1) survei kejadian penyakit di lapangan dan (2) deteksi virus dan identifikasi kutukebul. Kegiatan pertama dilakukan di areal pertanaman tomat pada lima strata ketinggian tempat yaitu: 100 – 300 m di atas permukaan laut (mdpl) (Anggada, Bogor), 300 – 600 mdpl (Rancamaya, Sukabumi), 600 – 900 mdpl (Cibedug dan Ciampea, Bogor; Leles dan Cilimus, Garut), 900 – 1200 mdpl (Bayongbong, Samarang, dan Cisurupan, Garut; Ciherang, Cianjur), dan 1200 – 1500 mdpl (Pacet, Cianjur; Lembang dan Pangalengan, Bandung). Kegiatan kedua dilakukan di Laboratorium Virologi Tumbuhan dan Laboratorium Biosistematika Serangga, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor; Laboratorium Biologi Molekuler, Lembaga Ilmu Pengetahuan Indonesia (LIPI); dan Laboratorium Biologi Molekuler, Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumber Daya Genetik Pertanian (BB-BIOGEN). Penelitian ini dilaksanakan pada Maret 2011 sampai dengan Maret 2012.
Metode
Survei Kejadian Penyakit dan Pengambilan Sampel Tanaman Tomat
Kegiatan ini bertujuan untuk mengamati tingkat kejadian penyakit klorosis dan kerupuk serta kelimpahan kutukebul di pertanaman tomat. Pada setiap lahan tanaman tomat yang terserang kedua penyakit tersebut dihitung tingkat kejadian penyakitnya dengan mengamati 100 tanaman sampel secara acak. Juga, pada setiap tanaman sampel yang diamati dilakukan pengumpulan pupa dan kantung pupa kutukebul yang ada pada tanaman tersebut. Pada kegiatan ini juga dilakukan pencatatan terhadap kondisi suhu udara, kelembaban nisbi, dan karakteristik budidaya seperti varietas tomat, umur tanaman, dan penggunaan pestisida. Untuk mengindentifikasi jenis virusnya, sampel daun dari tanaman yang memperlihatkan gejala kedua jenis penyakit tersebut diambil secara acak dari lima tanaman terserang. Selanjutnya, daun-daun sampel ini dibawa ke laboratorium untuk dianalisis lebih lanjut.
Deteksi Keberadaan Virus
Daun sampel diambil dari masing-masing tanaman yang memiliki gejala penyakit klorosis, kerupuk maupun infeksi ganda. Deteksi virus dilakukan melalui tahapan ekstraksi RNA/DNA total dan amplifikasi DNA dengan RT-PCR/PCR. Ekstraksi RNA total Crinivirus menggunakan Rneasy Plant Mini Kits (Philekorea Technology) mengikuti prosedur produsen sedangkan ekstraksi DNA
14 (5’-CTTCAAACATCCTCCATCTGCC-3’) yang berukuran 417 bp. Gen protein
selubung (coat proteion) Begomovirus diamplifikasi menggunakan primer spesifik untuk gen AV1 Begomovirus (AVRDC, Taiwan) yaitu CPPROTEIN-V1 (5’TAATTCTAGATGTCGAAGCGACCCGCCGA-3’) dan CPPROTEIN-C1 (5’-GGCCGAATTCTTAATTTTGAACAGAATCA-3’) berukuran 780 bp.
Ektraksi RNA Total TICV dan ToCV. Ekstraksi RNA total dengan jaringan daun tanaman tomat bergejala Crinivirus menggunakan Rneasy Plant Mini Kits
(Philekorea Technology) mengikuti prosedur produsen. Sebanyak 0.1 g sampel daun digerus menggunakan mortar dengan bantuan nitrogen cair. Hasil gerusan dimasukkan ke dalam tabung mikro 2 ml lalu ditambahkan campuran 450 µl
buffer ekstraksi (buffer XPRB) dan 4.5 µl merkaptoetanol (perbandingan 1:100) kemudian divorteks. Selanjutnya filter colomn berwarna putih disiapkan dan ditumpuk diatas tabung mikro 2 ml yang baru. Sampel dipipet dan dimasukkan ke dalam filter colomn berwarna putih, lalu disentrifugasi pada kecepatan 14000 rpm selama 2 menit. Setelah itu, supernatan dipipet dan dipindahkan ke dalam tabung mikro 2 ml yang baru kemudian volumenya diukur. Kemudian etanol absolut ditambahkan sebanyak 0,5 vol supernatan kemudian homogenkan dengan menaik turunkan pipet. Selanjutnya sampel dimasukan ke dalam XPPLR mini colomn
berwarna merah yang telah disiapkan dan ditumpuk diatas tabung mikro 2 ml yang baru lalu disentrifugasi pada kecepatan 14000 rpm selama 1 menit. Cairan yang tersisa lalu dibuang. Pada tahap pencucian, sebanyak 500 µl Wash buffer 1 dimasukan ke dalam tabung tadi, lalu disentrifugasi pada kecepatan 14000 rpm selama 1 menit. Cairan yang tersisa lalu dibuang. Selanjutnya sebanyak 750 µl
Wash buffer 2 dimasukan ke dalam tabung tadi, lalu disentrifugasi pada kecepatan 12000 rpm selama 1 menit. Cairan yang tersisa lalu dibuang. Kemudian tabung disentrifugasi kembali pada kecepatan 12000 rpm selama 3 menit. XPPLR mini colomn berwarna merah (tanpa tabung koleksi 2 ml tadi) dipindahkan ke tabung koleksi 2 ml yang baru. Kemudian RNase-free water 50 µl dipipet dan dimasukan ke pusat membrane colomn lalu didiamkan selama 1 menit. Selanjutnya disentrifugasi pada kecepatan 12000 rpm selama 2 menit untuk mengelusi RNA. Siapan RNA total disimpan di lemari pendingin -80°C dan digunakan sebagai
template dalam reaksi RT-PCR.
Prosedur RT-PCR dan PCR TICV dan ToCV. Reaksi RT dilakukan dengan volume total 25μl yang mengandung 3 μl RNA total; 0,75 pmol random primer, 500 mM dNTP, 5 mM MgCl2 , 4 μl buffer RT (250 mM Tris-HCl, pH 8,3; 375
mM KCl; 15 mM MgCl2; 50 mM DTT), 20unit RNasin ribonuclease inhibitor
15
buffer PCR 10x (500 mM KCl; 100 mM Tris-HCl, pH 9; 1% Triton X-100), dan 0,5 μl Taq DNA polymerase (Promega, Madison, WI). Reaksi PCR dengan denaturasi inisiasi pada 94°C selama 1 menit, dilanjutkan 30 siklus yang terdiri dari denaturasi pada 94°C selama 1 menit, penempelan primer pada 62°C selama 1 menit, dan pemanjangan pada 72°C selama 2 menit dan diikuti pemanjangan akhir pada 72°C selama 10 menit. Produk PCR kemudian dielektroforesis dengan mesin elektroforesis pada gel agarose 1% dalam buffer 0,5x TBE dan divisualisasi dengan mesin UV transilluminator.
Ektraksi DNA Total. Ekstraksi DNA total dengan jaringan daun tanaman tomat bergejala Begomovirus menggunakan metode Cetyl triethylammonium bromide
(CTAB) (Doyle and Doyle 1990). Proses ektraksi diawali dengan memanaskan terlebih dahulu 500 µl buffer ekstraksi/ CTAB buffer untuk masing-masing sampel ditambah dengan 1% merkaptoetanol dari volume total buffer di dalam
waterbath dengan suhu 65°C. Sementara itu sampel daun digerus dengan bantuan nitrogen cair. Selanjutnya ekstrak daun hasil penggerusan dimasukkan ke dalam tabung koleksi 2 ml dan ditambahkan 500 µl buffer ekstraksi yang telah dipanaskan tadi, kemudian campur merata. Selanjutnya campuran hasil gerusan tadi diinkubasi kembali dalam waterbath dengan suhu 65°C selama 60 menit dan setiap 10 menit tabung tersebut dibolak-balik untuk membantu proses lisis.
Setelah 60 menit, campuran diambil dan didiamkan sebentar (± 2 menit) pada suhu ruang, kemudian ditambahkan 500 µl campuran Chloroform : Isoamil (CI) dengan perbandingan 24:1 lalu divorteks selama 5 menit. Kemudian disentrifugasi dengan kecepatan 12000 rpm selama 15 menit. Supernatna yang terbentuk diambil dan dihitung volumenya, kemudian dimasukkan ke dalam tabung koleksi 2 ml yang baru. Selanjutnya tabung berisi supernatan ditambahkan 1/10 sodium asetat (CH3COONA) dan dicampur dengan merata. Setelah itu tambahkan lagi ke
dalamnya dengan 2/3 x volume isopropanol atau 2.5 x volume etanol absolut untuk presipitasi DNA. Selanjutnya diinkubasi semalaman dalam lemari pendingin dengan suhu -20°C. Keesokan harinya, tabung tersebut disentrifugasi kembali dengan kecepatan 12000 rpm selama 10 menit untuk mengendapkan DNA. Selanjutnya cairan supernatan (yang berada di lapisan paling atas) dibuang dan endapan DNA (yang berada di lapisan paling bawah tabung) dicuci dengan 500 μl etanol 70%, kemudian disentrifugasi kembali dengan kecepatan 12000 rpm selama 5 menit. Setelah itu cairan supernatan dibuang dan endapan DNA dikeringkan di atas kertas tisu bersih. Setelah kering, endapan DNA dilarutkan kembali dengan 100 μl buffer TE (pH 8). Selanjutnya DNA total disimpan di lemari pendingin -20°C dan dapat digunakan untuk proses amplifikasi PCR.
Prosedur PCR DNA TYLCV. Tabung koleksi 200 μl disiapkan, kemudian
buffer PCR 10x + Mg2+ (sebanyak 2.5 μl), sukrosa 10x (sebanyak 2.5 μl), deoksiribonukleotida triphosphat (dNTP) 10 mM (sebanyak 0.5 μl), forward primer (sebanyak 2 μl), reverse primer (sebanyak 2 μl), Taq DNA polymerase
(sebanyak 0.5 μl), ddH2O (sebanyak 14 μl), template DNA hasil ektraksi
(sebanyak 1 μl). Reaksi PCR dilakukan dalam volume 25 μl menggunakan mesin
16
dielektroforesis dengan mesin elektroforesis pada gel agarose 1% dalam buffer
0,5x TBE dan divisualisasi dengan mesin UV transilluminator.
Identifikasi dan Penghitungan Populasi Kutukebul
Pupa dan kantung pupa kutukebul yang diambil dari 100 tanaman sampel pada pengamatan kejadian penyakit selanjutnya diidentifikasi jenis spesiesnya menggunakan bantuan mikroskop cahaya binokuler dan kunci identifikasi Martin (1987). Pupa dan kantung pupa tersebut diambil dari daun tomat dengan menggunakan jarum dan dimasukkan ke dalam tabung eppendorf berisi alkohol 80% dan disimpan sebagai stok preparasi. Setelah jenis spesies kutukebul teridentifikasi, yaitu B. tabaci dan T. vaporariorum, kelimpahan populasi kedua spesies kutukebul tersebut dihitung dengan merata-ratakan jumlah pupa dan kantung pupa pada spesies yang sama dari seluruh daun yang diambil.
Populasi kedua kutukebul diketahui melalui penghitungan jumlah puparium dan kantung pupa masing-masing kutukebul. Jumlah populasi B. tabaci dan T. vaporariorum berupa pupa dan kantung pupa kemudian dicatat lalu dihitung rata-rata populasi kutukebul per kebun.
Pembuatan Preparat Mikroskop dan Identifikasi Kutukebul
Pupa dan kantung pupa kutukebul dari lapangan dilepaskan dari daun tomat dengan menggunakan jarum dan dimasukkan ke dalam tabung reaksi berbeda yang berisi larutan alkohol 80%. Pembuatan preparat pupa dan kantung pupa memiliki perbedaan yaitu pada pupa dilakukan proses pemanasan agar isi pupa bersih dan hasil akhir hanya berbentuk kantung pupa. Tabung reaksi berisi kantung pupa dimasukkan ke dalam gelas piala berisi kapas dan air, kemudian dipanaskan pada suhu 100°C selama 10 menit. Hasil rebusan tersebut dituang ke dalam cawan syracuse dengan memakai penjepit. Tahap selanjutnya pupa dimasukkan ke dalam tabung reaksi berisi larutan KOH 10% dan direbus seperti sebelumnya. Setelah pupa lunak dan berwarna transparan, tabung reaksi diangkat dan dituang ke dalam cawan Syracuse. Supaya cairan isi pupa keluar dan pupa tidak sobek, maka secara perlahan pupa ditekan dengan jarum halus. Selanjutnya, pada tahap pewarnaan, kantung pupa dan pupa yang telah bersih isinya (kantung pupa) kemudian dipindahkan ke cawan yang baru dan dibilas dengan aquades. Kantung pupa yang sudah dicuci bersih dipindahkan ke dalam cawan syracase
17 Analisis Hubungan Kejadian Penyakit dengan Kelimpahan Kutukebul
Hubungan antara kejadian penyakit klorosis (CRNV), kerupuk (BGMV), atau keduanya (ganda/ MLTV) dengan kelimpahan kutukebul (B. tabaci, BT, atau
T. vaporariorum, TV) dan faktor biotik dan abiotik lainnya seperti ketinggian tempat (ELV), kelembaban nisbi (RH), jenis varietas (VAR), dan umur tanaman (UTN) dianalisis menggunakan model regresi linier berganda yang didefinisikan sebagai berikut:
CRNV = b01 + b11 lnTV + b21 ELV + b31 RH + b41 VAR + b51 UTN + e1
BGMV= b02 + b12 lnBT + b22 ELV + b32 RH + b42 VAR + b52 UTN + e2
MLTV = b03 + b13 lnBT + b23 lnTV + b33 ELV + b43 RH + b53 VAR +
b63 UTN + e3
dengan e1, e2, dan e3 merupakan komponen galat. Pendugaan koefisien
regresi (b0i, b1i, b2i, ..., b6i; i = 1, 2, dan 3) menggunakan metode kuadrat terkecil
dan pengujiannya menggunakan uji t-student serta pengujian model menggunakan sidik ragam (Anova). Pendugaan dan pengujian tersebut dilakukan dengan program Minitab versi 14. Penentuan model terbaik selain menggunakan statistik kesesuaian model, yaitu R2 dan nilai F, juga didasarkan pada landasan teori tentang hubungan antara kejadian penyakit (klorosis dan kerupuk) dan vektornya. Model CRNV harus mencakup peubah TV (LnTV) sebagai salah satu peubah bebasnya, sedangkan model BGMV harus mencakup peubah BT (LnBT) sebagai salah satu peubah bebasnya.
18
4
HASIL DAN PEMBAHASAN
Insiden Crinivirus dan Begomovirus pada Tanaman Tomat Gejala Penyakit
Survei dan pengambilan sampel yang dilakukan di daerah sentra produksi tomat di Jawa Barat ditemukan dua penyakit virus dengan gejala yang mudah dibedakan yaitu (1) penyakit klorosis (chlorosis) dan (2) kerupuk (leaf curl). Penyakit klorosis dicirikan dengan perubahan warna lamina daun yaitu daerah antara tulang daun menjadi kuning karena mengalami klorosis. Gejala klorosis ini dimulai dari daun-daun tua di bagian bawah kemudian berkembang ke daun-daun di bagian atasnya dan pada akhirnya semua daun pada tanaman tersebut mengalami klorosis. Tanaman tidak tampak mengalami penghambatan pertumbuhan karena tanaman yang sakit umumnya setinggi tanaman sehat. Namun demikian, tanaman yang sakit klorosis menghasilkan buah dengan ukuran dan jumlah yang lebih kecil dari normal (Gambar 6A). Gejala-gejala yang terlihat secara visual di lapangan sesuai dengan deskripsi gejala berdasarkan penelitian-penelitian sebelumnya (Wisler et al. 1998; Dovas et al. 2002; Wintermantel 2004; Dalmon et al. 2005; Tsai et al. 2004; Vaira et al. 2002).
Gambar 6 Tanaman tomat yang terinfeksi Crinivirus memperlihatkan gejala klorosis (A) dan yang terinfeksi Begomovirus memperlihatkan gejala kerupuk (B); masing-masing telah diverifikasi dengan polymerase chain reaction (PCR) dan reverse transcription-PCR.
Menurut Wintermantel (2004), gejala penyakit klorosis oleh Tomato chlorosis virus (ToCV) ditunjukkan dengan klorosis di bagian antara tulang daun (interveinal yellowing) yang diawali pada daun bagian bawah kemudian berkembang ke bagian pucuk, mudah rapuh, ukuran buah mengecil, jumlah buah berkurang saat panen dan umur tanaman menurun. Selanjutnya, gejala penyakit klorosis oleh Tomato infectious chlorosis virus (TICV) adalah terjadinya klorosis dan nekrotik pada seluruh daun tanaman sakit, buah yang dihasilkan berukuran jauh lebih kecil dari normal dan proses pematangannya terganggu serta mudah gugur (Wisler et al. 1998a, Dalmon et al. 2005, Tsai et al. 2004, Vaira et al.
2008). Menurut Duffus et al. (1996) dan Dalmon et al. (2008) terjadinya infeksi ganda ToCV dan TICV pada tanaman tomat menyebabkan daun-daun tomat klorosis, yaitu menguning di antara tulang daun (interveinal yellowing) yang berasosiasi dengan berkurangnya kemampuan fotosintesisnya. Perkembangan
19 selanjutnya daun-daun menjadi rapuh (leaf brittleness), mengalami nekrotik pada beberapa bagian dan warna bagian yang nekrotik menjadi merah keunguan (bronzing), kebugaran (vigor) tanaman menjadi sangat berkurang, dan apabila menghasilkan buah maka ukurannya jauh lebih kecil dari normal dan proses pematangannya terganggu, serta mudah gugur (early senescence) sehingga sangat menurunkan bahkan meniadakan nilai ekonomi tanaman yang terinfeksi.
Penyakit kerupuk dicirikan dengan pertumbuhan terhambat pada tanaman sehingga nampak kerdil, demikian juga daun-daun yang tumbuh di bagian pucuk mengalami perubahan bentuk (malformasi) berat yaitu bagian pinggirnya melengkung ke atas sehingga tampak seperti kerupuk. Tanaman yang memperlihatkan gejala seperti ini umumnya tidak berbuah (Gambar 6B). Gejala kerupuk ini sudah banyak dideskripsikan oleh peneliti terdahulu (Butter dan Rataul 1977; Jones et al 1991; Sugiarman dan Hidayat 2000). Menurut Butter dan Rataul (1977) serta Jones et al. (1991), tanaman tomat yang terinfeksi Tomato yellow leaf curl virus (TYLCV) adalah klorosis yang muncul dari tunas daun kemudian menyebar ke daun-daun bagian bawah disertai gejala keriting atau berbentuk seperti mangkok (cupping), keras, daun berkerut (puckering), bunga rontok dan terdapat enasi pada permukaan bawah daun. Sugiarman dan Hidayat (2000) lebih lanjut menjelaskan bahwa gejala kerupuk di Indonesia pada umumnya menimbulkan gejala berupa penebalan daun, lamina daun berkerut, daun menguning, tepi daun melengkung ke atas, daun keriting dan tanaman kerdil. Adanya asosiasi dari ketiga penyakit tersebut dikonfirmasi lebih lanjut dengan deteksi keberadaan virus secara molekuler menggunakan metode RT-PCR untuk
Crinivirus dan PCR untuk Begomovirus. Verifikasi Infeksi Virus
Verifikasi infeksi virus dilakukan dengan menggunakan metode RT-PCR dan PCR yang sebelumnya dibutuhkan DNA total virus dalam menggunakan metode tersebut. DNA total virus didapatkan melalui teknik isolasi DNA total yaitu untuk DNA Crinivirus melalui cara ekstraksi RNA total Crinivirus
20
elektroforesis. Hasil yang baik diindikasikan dengan pita DNA yang tebal dan tegas serta bersih.
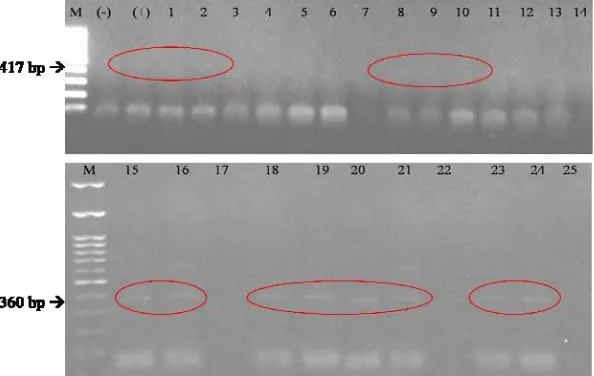
Verifikasi infeksi Crinivirus pada tanaman tomat sampel telah berhasil dilakukan dengan RT-PCR menggunakan primer spesifik terhadap ToCV yaitu ToCV-CF dan ToCV-CR yang dapat mengamplifikasi bagian minor coat protein
(CPm) sebesar 360 bp (Hirota et al. 2009) dan pasangan primer spesifik terhadap TICV yaitu TICV-CF dan TICV-CR yang dapat mengamplifikasi bagian gen CPm dengan amplikon yang berukuran 417 bp (Hirota et al. 2010). Sampel-sampel tanaman tomat dengan gejala khas penyakit klorosis pada berbagai strata ketinggian tempat memperlihatkan pita DNA dengan ukuran sesuai dengan prediksi primer yang digunakan untuk ToCV ataupun TICV (Gambar 7). Infeksi masing-masing virus tersebut pada tanaman tomat tidak menginduksi gejala berbeda. Melalui RT-PCR maka penghitungan insiden infeksi Crinivirus di lapangan yang dilakukan melalui pencatatan tanaman bergejala dapat diverifikasi.
Gambar 7 Hasil amplifikasi sampel DNA Crinivirus dari lapangan menggunakan metode RT-PCR. M, Marker DNA 100 bp; (Kn), Kontrol negatif/tanaman sehat; (Kp), Kontrol positif; 1-5, 6-10, 11-15, 16-20, dan 21-25 sampel tanaman tomat bergejala klorosis yang diambil berturut-turut dari ketinggian tempat 100-300 mdpl, 300-600 mdpl, 600-900 mdpl, 900-1200 mdpl, 1200-1500 mdpl.
Penyakit kerupuk pada tanaman tomat diketahui berasosiasi dengan infeksi
Begomovirus yaitu TYLCV. Penelitian ini telah dan berhasil dilakukan verifikasi asosiasi antara penyakit kerupuk dan infeksi TYLCV melalui PCR menggunakan primer spesifik untuk gen AV1 Begomovirus yaitu CPPROTEIN-V1 dan CPPROTEIN-C1 (Rojas et al. 1993) (Gambar 8). Sampel-sampel tanaman yang memperlihatkan gejala khas penyakit kerupuk yang dikoleksi dari berbagai strata ketinggian tempat memperlihatkan amplikon berukuran 780 bp, sesuai dengan prediksi desain primer yang digunakan (Rojas et al. 1993). Hasil ini memberikan legitimasi pada hasil pengamatan lapangan mengenai insiden infeksi Begomovirus
21
Gambar 8 Hasil amplifikasi sampel DNA Begomovirus dari lapangan menggunakan metode PCR. M, Marker DNA 100 bp; (Kn), Kontrol negatif/tanaman sehat; (Kp), Kontrol positif; 1-5, 6-10, 11-15, 16-20, dan 21-25 sampel tanaman tomat bergejala klorosis yang diambil berturut-turut dari ketinggian tempat 100-300 mdpl, 300-600 mdpl, 600-900 mdpl, 900-1200 mdpl, 1200-1500 mdpl.
Kejadian Penyakit oleh Crinivirus dan Begomovirus
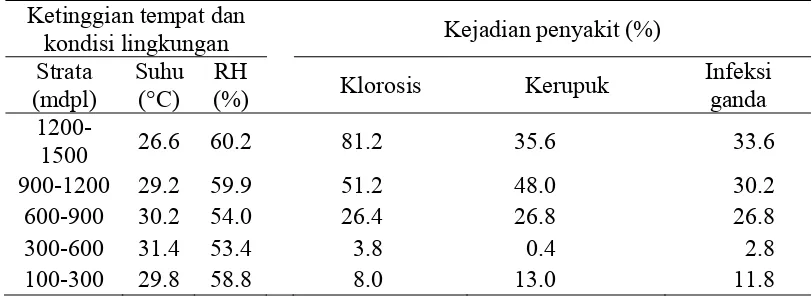
Tingkat kejadian penyakit atau insiden infeksi masing-masing virus pada suatu luasan lahan memiliki persentase yang beragam. Hal ini disebabkan oleh faktor-faktor yang mempengaruhi perkembangan penyakit seperti segitiga penyakit. Hubungan segitiga penyakit yang meliputi ketahanan tanaman inang, patogen yang virulen dan kondisi lingkungan yang mendukung sulit dipisahkan keterkaitannya. Ketinggian tempat saling terkait dengan kondisi suhu sekitar sehingga hal tersebut akan berpengaruh terhadap kejadian penyakit. Hasil survei kejadian penyakit (KP) di lapangan menunjukkan bahwa secara umum, semakin tinggi tempat, maka semakin besar rataan kejadian penyakit klorosis. Hal serupa terjadi pada rataan kejadian penyakit oleh kerupukdan infeksi ganda.
Tabel 1 Kejadian penyakit (insiden infeksi) klorosis dan kerupuk pada tanaman tomat di daerah Jawa Barat pada setiap strata ketinggian tempat
Ketinggian tempat dan
kondisi lingkungan Kejadian penyakit (%) Strata
(mdpl)
Suhu (°C)
RH
(%) Klorosis Kerupuk
Infeksi ganda
22
penyakit kerupuk, titik puncak kenaikan kejadian penyakit berada di strata ketinggian 900-1200 mdpl sebesar 48.0% yang kemudian terjadi penurunan yaitu 35.6% saat ketinggian tempat lebih dari 1200 mdpl.
Kondisi lingkungan dan waktu yang tepat sangat mendukung penyebaran kedua penyakit tersebut. Penyebaran kedua penyakit tersebut melalui serangga vektor yaitu kutukebul. Mobilitas kutukebul sebagai vektor dapat menyebarkan virus dari satu tanaman ke tanaman lain secara cepat (Duffus et al. 1994, Wintermantel 2004) sehingga kejadian penyakit dalam suatu areal dapat mencapai 90% (Duffus et al. 1996). Selain itu, persentase kejadian penyakit dapat pula dipengaruhi oleh jenis kutukebulnya.
Kutukebul pada Pertanaman Tomat Spesies Kutukebul
Selama survei, selain dilakukan pengamatan terhadap gejala penyakit oleh
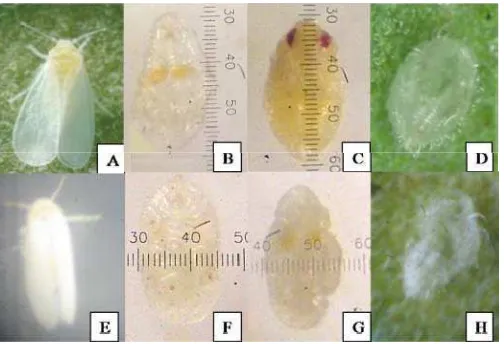
Crinivirus maupun Begomovirus, juga telah dilakukan pengamatan terhadap kutukebul yang merupakan serangga vektor virus tersebut. Spesies kutukebul yang diamati pada lahan pertanaman tomat sampel adalah Trialeurodes vaporariorum dan Bemisia tabaci dari ordo Hemiptera, famili Aleyrodidae. Hasil survei dan pengamatan di lapangan, telah berhasil ditemukan beberapa stadia perkembangan kutukebul tersebut yaitu stadia nimfa, pupa dan imago. Selain itu ditemukan pula kantung pupa dari kedua spesies kutukebul tersebut. Secara umum, imago kutukebul sering digambarkan seperti ngengat, memiliki bentuk putih kecil sekitar 1.0 mm, seluruh tubuh termasuk sayap memiliki penampilan tepung putih terang, biasanya ditemukan pada permukaan bawah daun tanaman tomat, dan terbang bila merasa terganggu. Nimfa berbentuk bulat panjang dan berwarna coklat muda sampai transparan. Pupa berbentuk bulat panjang, struktur abdomen jelas, bagian toraks cembung dan melebar. Kantung pupa berwarna transparan dan terdapat lubang berupa sobekan pada ujung kantung yang merupakan tempat keluarnya imago dari kantung pupa.
23 Hasil pengamatan secara morfologi yang dilakukan di laboratorium ditemukan bahwa bentuk morfologi antara imago, nimfa, pupa, dan kantung pupa
T. vaporariorum dan B. tabaci dapat dibedakan (Gambar 9). Imago kedua spesies kutukebul tersebut memiliki tubuh berwarna kuning dengan sayap berwarna putih. Ukuran tubuh T. vaporariorum lebih besar dibanding dengan B. tabaci.
Perbedaan lain terletak pada posisi sayap imago saat sedang istirahat yaitu posisi sayap T. vaporariorum lebih mendatar dibanding dengan posisi sayap B. tabaci
yang berbentuk seperti atap rumah. Perbedaan bentuk nimfa kedua spesies
kutukebul tersebut dapat dilihat dari bentuk tubuh nimfanya yaitu pada
T. vaporariorum berbentuk bulat lonjong, tepi tubuh nimfa tanpa lekukan, terdapat dua bulatan berwarna kuning pada bagian tengah tubuhnya, dan tubuh nimfa berwarna kuning transparan. Nimfa B. tabaci berbentuk lebih bulat dengan tepian tubuh berlekuk-lekuk, tidak terdapat dua bulatan berwarna kuning pada bagian tengah tubuhnya, dan tubuh nimfa berwarna kuning transparan. Ketebalan nimfa dari kedua spesies ini yaitu nimfa T. vaporariorum lebih tebal dibandingkan
B. tabaci. Bentuk pupa T. vaporariorum bulat lonjong, strukturnya cembung dan mempunyai ketebalan, tepi pupa halus tanpa lekukan, memiliki seta yang banyak dan ukuran yang pendek, pupa berwarna kuning kecoklatan, dan di ujung pupa terdapat dua bulatan bakal mata berwarna merah. Pupa B. tabaci berbentuk bulat dan agak melebar dibandingkan pupa T. vaporariorum, strukturnya tidak cembung dan tipis, tepi pupa memiliki lebih banyak lekukan besar seringkali menyerupai bentuk bunga, memiliki sedikit seta dan ukuran yang panjang, pupa berwarna kuning. Kantung pupa T. vaporariorum dan B. tabaci berbentuk seperti pupa masing-masing tetapi sudah tidak berisi imago lagi di dalamnya, dapat digambarkan seperti kantung yang kosong, kantung berwarna putih transparan, terdapat sedikit sobekan di ujung kantung sebagai tempat keluarnya imago.
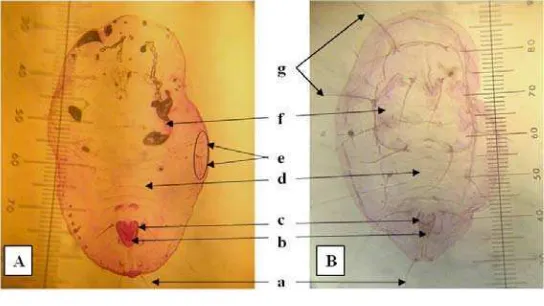
Gambar 10 Hasil identifikasi pupa kutukebul. (A) pupa T. vaporariorum. (B) pupa B. tabaci. (a) seta kauda; (b) lingula; (c) vasiform orifice; (d) ruas abdomen VII; (e) submargin papilla; (f) basal tungkai tengah dan belakang; (g) rambut dorsal.
Identifikasi dikonfirmasi juga melalui preparat kantung pupa (Gambar 10). Menurut Mound & Halsey (1978), kutukebul dapat diidentifikasi dengan mudah, diketahui spesies atau genusnya melalui struktur pupa ataupun kantung pupa. Hasil identifikasi menunjukkan bahwa pupa T. vaporariorum memiliki bagian-bagian berupa bentuk lingula yang membulat, vasiform orifice berbentuk segitiga,
24
belakang mempunyai seta yang halus dan kecil. Hasil identifikasi pupa B. tabaci
ialah bentuk lingula yang memanjang seperti lidah, submargin tanpa barisan papila, vasiform orifice berbentuk segitiga, pinggiran trakea seperti sisir, dan rambut dorsal berjumlah tujuh pasang. Hasil pengamatan dan identifikasi preparat kedua kutukebul tersebut sesuai dengan deskripsi Martin (1987).
Kelimpahan kutukebul sangat mempengaruhi perkembangan penyakit klorosis dan kerupuk. Rusli et al. (1999) menyatakan bahwa tingkat kejadian penyakit yang disebabkan oleh Crinivirus dan Begomovirus berkaitan dengan populasi kutukebul. Meningkatnya jumlah kutukebul yang viruliverous akan menyebabkan peningkatan persentase kejadian penyakit.
Kelimpahan Populasi Kutukebul
Jumlah populasi kedua spesies kutukebul pada pengamatan di beberapa ketinggian tempat di Jawa Barat menunjukkan hasil yang berbeda. Hasil pengamatan kelimpahan populasi kutukebul secara umum menunjukkan bahwa kelimpahan populasi T. vaporariorum dan B. tabaci semakin besar seiring dengan ketinggian tempat yang semakin tinggi. Kedua spesies kutukebul tersebut juga seringkali ditemukan berada dalam satu lahan pertanaman yang sama (Tabel 2). Hasil pengamatan juga menunjukkan keberadaan populasi T. vaporariorum pada dataran rendah cenderung lebih sedikit dibandingkan dengan B. tabaci. Hal tersebut menjelaskan bahwa perbedaan ketinggian tempat berpengaruh terhadap tingkat populasi kutukebul baik T. vaporariorum maupun B. tabaci. Adanya pengaruh dari ketinggian tempat terhadap populasi kutukebul tidak lepas dari peran serta kondisi lingkungan di sekitar pertanaman tomat dan budaya masing-masing masyarakat setempat dalam tata cara dalam pembudidayaan tomat. Tabel 2 Populasi kutukebul pada tanaman tomat di daerah Jawa Barat pada
setiap strata ketinggian tempat dan lokasi pengamatan Strata
(mdpl) Lokasi pengamatan
Suhu (°C)
RH (%)
Populasi (ekor/100 tanaman)
T. vaporariorum B. tabaci
1200-1500 Pacet, Lembang,
Pangalengan 26.6 60.2 257.6 187.2
600-900 Cibedug, Ciampea,
Leles, Cilimus 30.2 54.0 8.8 30.0 300-600 Rancamaya 31.4 53.4 0.0 70.0
100-300 Anggada 29.8 58.8 2.8 64.2
Populasi T. vaporariorum dan B. tabaci yang menyerang setiap 100 tanaman tomat di masing-masing kebun hampir sama jumlahnya pada ketinggian 900-1200 mdpl yaitu berturut-turut ditemukan sebanyak 33 ekor dan 36 ekor. Nampaknya kedua serangga tersebut cocok hidup dan beradaptasi pada ketinggian
tempat tersebut. Namun pada ketinggian lebih dari 1200 mdpl, populasi
25
sebanyak 257.6 ekor dan 187.2 ekor. Lingkungan dingin merupakan habitat
T. vaporariorum sehingga pada ketinggian tersebut populasi kutukebul lebih banyak, sedangkan lingkungan dingin bukan habitat B. tabaci, namun pada saat ini kutukebul tersebut mampu beradaptasi pada ketinggian tersebut. Pertanaman tomat yang memiliki jumlah rataan populasi B. tabaci dan T. vaporariorum paling besar terdapat pada ketinggian lebih dari 1200 mdpl dengan suhu rata-rata 26.6°C dan kelembaban nisbi 60.2%.
Hubungan Kejadian Penyakit oleh Crinivirus dan Begomovirus dengan Kelimpahan Populasi Kutukebul
Enam peubah bebas yang telah dianalisis, yaitu elevasi, kelimpahan
T. vaporariorum, kelimpahan B. tabaci, kelembaban nisbi, umur tanaman, dan varietas tanaman, ternyata hanya empat peubah yang terlihat berkorelasi dengan peningkatan kejadian penyakit klorosis dan kerupuk dari ketinggian tempat rendah ke ketinggian tempat tinggi. Keempat peubah bebas tersebut adalah elevasi, kelimpahan T. vaporariorum, kelimpahan B. tabaci, dan umur tanaman. Jumlah dan jenis peubah bebas yang berkorelasi dengan peubah terikat (kejadian penyakit) berbeda antara satu model kejadian penyakit ke model kejadian penyakit lainnya. Model kejadian penyakit klorosis, peubah bebas selain kelimpahan kutukebul (TV atau BT) yang berkontribusi nyata terhadap kejadian penyakit adalah elevasi (ELV), sementara pada model kejadian penyakit kerupuk adalah ELV dan umur tanaman (UTN). Hasil analisis regresi terhadap hubungan antara kejadian penyakit klorosis (Crinivirus), kerupuk (Begomovirus), dan gabungan keduanya secara rinci disajikan pada Tabel 3. Dua peubah bebas yang tidak terlihat kontribusinya adalah kelembaban nisbi (RH) dan umur tanaman (UTN). Hal ini mengindikasikan bahwa kedua faktor tersebut tidak mempengaruhi perubahan kejadian penyakit klorosis dan kerupuk.
Model Crinivirus, koefisien pada peubah ELV dan LnTV memiliki tanda positif dan signifikan pada taraf 5% (Nilai-P < 0.05). Hal ini berarti bahwa peningkatan ketinggian tempat dan kelimpahan T. vaporariorum akan meningkatkan kejadian penyakit klorosis. Nilai koefisien pada ELV sebesar 0.0004 atau 0.04%, yang berarti peningkatan ketinggian tempat sebesar 100 mdpl akan meningkatkan kejadian penyakit klorosis sebesar 4%. Sementara nilai koefisien pada LnTV adalah 0.0847 atau 8.47%, yang berarti bahwa setiap peningkatan LnTV sebesar 1 satuan atau dengan kata lain peningkatan kelimpahan T. vaporariorum sebesar e1 = 2.7 ekor per tanaman akan meningkatkan kejadian penyakit klorosis sebesar 8.5%. Hasil ini menunjukkan bahwa insiden penyakit klorosis semakin meningkat dengan semakin meningkatnya ketinggian tempat dan peningkatan insiden penyakit tersebut bertambah besar bila keberadaan kutukebul T. vaporariorum di pertanaman semakin melimpah.
26
Tabel 3 Koefisien model dan hasil uji terhadapnya dari tiga model kejadian penyakit klorosis, kerupuk, dan gabungan keduanya Peubah
bebas
Model Crinivirus Model Begomovirus Model Virus Ganda
Koef thit NilaiP Koef thit NilaiP Koef thit NilaiP
Intersep -0.1146 -1.36 0.187 0.1738 1.08 0.293 -0.3696 -2.42 0.025 ELV 0.0004 2.10 0.047 0.0005 3.25 0.004 0.0001 0.73 0.472
LnTV 0.0847 2.42 0.024 - - - -0.0445 -1.14 0.268
LnBT - - - 0.0151 0.43 0.675 0.1032 2.81 0.011
UTN - - - -0.0048 -2.15 0.043 0.0031 1.86 0.077
R2 = 73.2%; Nilai-P = 0.000
R2 = 42.2%; Nilai-P = 0.008
R2 = 49.6%; Nilai-P = 0.006