SERTA DERAJAT KOMPETISI BEBERAPA AKSESI
GULMA Echinochloa crus-galli (L.) Beauv.
TERHADAP TANAMAN PADI SAWAH
DWI GUNTORO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PENDAHULUAN
Latar Belakang
Tanaman padi (Oryza sativa) merupakan komoditas yang strategis di Indonesia karena beras merupakan sumber makanan pokok bagi hampir seluruh
rakyat Indonesia. Kebutuhan beras di Indonesia semakin meningkat sejalan
dengan pertambahan jumlah penduduk. Jumlah penduduk Indonesia tahun 2011
mencapai 241 juta jiwa dan kebutuhan beras mencapai 34 juta ton per tahun (BPS,
2011). Produksi padi pada tahun 2011 berdasarkan Angka Ramalan II (ARAM II)
diperkirakan mencapai 68.06 juta ton gabah kering giling (GKG) atau 38.2 juta
ton beras. Kebutuhan beras pada tahun 2025 diperkirakan mencapai 48.5 juta ton
atau setara dengan 70 juta ton GKG.
Untuk memenuhi kebutuhan beras tersebut diperlukan usaha peningkatan
produksi dan produktivitas padi di Indonesia. Peningkatan produksi beras tahun
2011 dibandingkan tahun 2010 disebabkan oleh peningkatan luas areal (0.11%)
dan peningkatan produktivitas (1.24%) (BPS 2011). Pada tahun-tahun
mendatang, upaya peningkatan produksi beras akan menghadapi banyak kendala
diantaranya perubahan fungsi lahan sawah menjadi lahan non pertanian yang
mencapai 187 720 hektar/tahun (Ditjen Pengelolaan Lahan dan Air 2005),
degradasi kesuburan lahan yang saat ini mencapai 4 juta hektar dari degradasi
berat (50%) hingga degradasi rendah (0.8%) (Badan Litbang Pertanian 2011),
serta adanya serangan organisme pengganggu tanaman.
Salah satu organisme pengganggu tanaman yang dapat menurunkan
produksi dan produktivitas tanaman padi adalah gulma. Kehilangan hasil akibat
gulma di seluruh dunia diperkirakan mencapai 10-15% (Smith 1968; Smith 1983;
Zoschke 1990; Baltazar dan De Datta 1992), bahkan kehilangan hasil dapat
mencapai 86% jika tanpa dilakukan pengendalian gulma (Kropff 1993).
Salah satu gulma penting dan dominan pada lahan padi sawah di Indonesia
adalah gulma jajagoan (Echinochloa crus-galli) (Ali dan Sankaran 1984; Ali 1985). Gulma E. crus-galli menjadi masalah utama pada budidaya tanaman padi sawah dan merupakan penyebab kehilangan hasil produksi yang utama pada
produksi padi akibat gulma E. crus-galli dapat mencapai 46-59% (Sultana 2000;
Chin 2001), 57-95% (Ahn dan Chung 2000), bahkan hingga 97% (Islam dan
Karim 2003).
Penurunan produksi tanaman padi oleh gulma E. crus-galli dapat terjadi karena kompetisi, alelopati, dan menjadi inang hama penyakit tanaman padi
(alelomediasi). Kompetisi gulma E. crus-galli menyebabkan penurunan hasil produksi akibat penurunan jumlah anakan, jumlah malai, dan jumlah gabah per
malai (Tindall et al. 2005). Gulma E. crus-galli berpotensi mengeluarkan senyawa alelopati yang dapat menghambat pertumbuhan tanaman (Putnam 1986).
Eksudat akar E. crus-galli yang berupa senyawa p-Hidroxymandelic menunjukkan efek alelopati dan menekan pertumbuhan tajuk dan pemanjangan akar tanaman
padi (Yamamoto et al. 1999; Xuan et al. 2006). Gulma E. crus-galli dapat menjadi inang perantara bagi Leptocorisa oratorius, Acrocylindricum oryzae,
Corticium sasakii, dan Rhynchosporium oryzae (Tjitrosemito 1994). Selain menurunkan kuantitas hasil, keberadaan gulma E. crus-galli juga menyebabkan penurunan kualitas dalam produksi benih akibat tercampurnya benih padi dengan
biji-biji E. crus-galli.
Gulma E. crus-galli sulit dikendalikan karena kemiripan morfologi dengan tanaman padi. Gulma ini menjadi lebih bermasalah pada budidaya tanaman padi
ketika cara tanam padi berubah dari cara tanam pindah (transplanting) menjadi cara tebar benih langung (direct planting). Keberadaan gulma E. crus-galli pada pertanaman padi sawah dapat menurunkan pendapatan petani padi akibat
peningkatan biaya pengendalian gulma.
Usaha peningkatan produksi yang dilakukan oleh pemerintah harus
diimbangi dengan upaya penyelamatan kehilangan hasil akibat organisme
pengganggu tanaman. E. crus-galli memiliki daya adaptasi yang luas pada
kondisi lingkungan yang bervariasi (Galinato et al. 1999). E. crus-galli yang berasal dari habitat yang berbeda diduga memiliki daya kompetisi yang berbeda
pula. Studi tentang potensi aksesi gulma E. crus-galli dalam menurunkan produksi padi masih jarang dilakukan di Indonesia. Studi keragaman morfologi
tentang karakter dan perilaku gulma tersebut dapat menjadi dasar bagi
pengembangan teknik pengendalian di lapangan sehingga dapat mendukung usaha
peningkatan produksi padi nasional.
Tujuan Penelitian
Secara umum penelitian ini bertujuan untuk mengkaji keragaman
morfologi dan genetik aksesi gulma E. crus-galli serta menganalisis tingkat kompetisinya terhadap tanaman padi sawah. Adapun tujuan khusus penelitian ini
adalah :
1. Menganalisis keragaman morfologi dan genetik aksesi E. crus-galli asal Jawa Barat
2. Mengidentifikasi potensi alelopati aksesi E. crus-galli asal Jawa Barat 3. Mempelajari pengaruh aksesi dan tingkat populasi E. crus-galli terhadap
pertumbuhan dan produksi padi sawah
4. Menduga derajat kompetisi gulma E. crus-galli melalui pendekatan
replacement series
5. Mempelajari fisiologi kompetisi antara padi dengan gulma E. crus-galli.
Hipotesis
1. Aksesi gulma E. crus-galli asal Jawa Barat memiliki keragaman karakter morfologi dan genetik. Keragaman morfologi yang terjadi di lapangan
disebabkan oleh keragaman genetik dan oleh kemampuan plastisitas fenotipik.
2. Tiap aksesi gulma E. crus-galli asal Jawa Barat memiliki potensi alelopati yang berbeda dan memiliki perbedaan kemampuan dalam menurunkan
pertumbuhan dan produksi tanaman padi sawah.
3. Gulma E. crus-galli memiliki daya kompetisi yang lebih kuat dibandingkan
dengan tanaman padi sawah dalam memperebutkan sumberdaya.
4. Perbedaan populasi dan asal aksesi gulma E. crus-galli menyebabkan perbedaan respon pertumbuhan dan produksi tanaman padi sawah.
Untuk menguji hipotesis yang dirumuskan maka dilakukan rangkaian
penelitian dengan tahapan seperti disajikan pada Gambar 1 berikut.
Gambar 1. Bagan alir tahapan penelitian Studi Keragaman Morfologi dan Genetik Aksesi Gulma
E. crus-galli Asal Jawa Barat
Pendugaan Derajat Kompetisi Gulma E. crus-galli
dengan Metode Replacement Series
Uji Potensi Allelopati Aksesi Gulma E. crus-galli
Percobaan Pendahuluan tentang Pertumbuhan dan Produksi Padi pada Berbagai Tingkat Populasi Gulma E. crus-galli
yang Berasal dari Tiga Sentra Produksi Padi
Studi Pengaruh Aksesi dan Tingkat Populasi E. crus-galli terhadap Pertumbuhan dan Produksi Padi
A. Percobaan Rumah Kaca B. Percobaan Lapangan
Studi Fisiologi Kompetisi Gulma E. crus-galli pada Beberapa Varietas Padi Sawah
Diperoleh informasi keragaman morfologi, genetik, dan derajat kompetisi aksesi gulma E. crus-galli
TINJAUAN PUSTAKA
Botanidan Morfologi Echinochloa crus-galli Botani
Gulma E. crus-galli merupakan gulma semusim kelas Monocotyledon, subkelas Commelinidae, ordo Cyperales, suku Poaceae/Graminae, dan marga
Echinochloa. Spesies E. crus-galli meliputi 3 subspesies, yaitu E. crus-galli var.
crus-galli, E. crus-galli var. particola dan E. crus-galli var. formosensis (Kim 1994). E. crus-galli adalah tumbuhan hexaploid (2n=6x=54). Gulma E. crus-galli
memiliki distribusi geografis yang luas dan keragaman dari morfologi dan
ekologi yang diduga terkait dengan sifat allohexaploid dari spesies ini (Yabuno
1983).
E. crus-galli diperkirakan berasal dari India, tersebar pada daerah tropis dan sub tropis di seluruh negara Asia Tenggara dan Asia selatan serta Australia
(Holm et al. 1977). Beberapa nama umum untuk rumput ini antara lain barnyard grass, japanese millet, cockspur, dan watergrass. Di Indonesia E. crus-galli
dikenal dengan nama gagajahan, jajagoan, jawan, jawan pari, suket ngawan
(Jawa) (Moenandir 1993; Galinato et al. 1999).
Morfologi
Rumput E. crus-galli sangat mirip dengan tanaman padi pada saat masih muda. E. crus-galli memiliki penampilan tegak, memiliki tinggi sekitar 20-150 cm (Soerjani et al. 1987), bahkan bisa mencapai 200 cm (Galinato et al. 1999).
E. crus-galli memiliki daun yang tegak atau rebah pada dasarnya.
Daunnya memiliki ukuran panjang sampai 35 cm dan lebar 0.5-1.5 cm. Warna
daun rumput ini hijau sampai hijau keabuan. Setiap daun memiliki pelepah yang
tidak berambut dan memiliki panjang 9-13 cm. Pelepah daun umumnya berwarna
kemerahan di bagian bawahnya. Helaian daun berukuran 5-65 cm x 6-22 mm,
bersatu dengan pelepah, berbentuk linear dengan bagian dasar yang lebar dan
melingkar dan bagian ujung yang meruncing. Permukaan daun rata, agak kasar
dan menebal di bagian tepi (Duke 1996). Helaian daun memiliki beberapa rambut
Daerah pangkal daun dapat digunakan untuk membedakan daun E. crus-galli dan
daun padi. Pangkal daun E. crus-galli tidak memiliki ligula dan aurikel, sedangkan pangkal daun padi memiliki ligula yang bermembran dan aurikel yang berbulu (Itoh 1991).
Gambar 2. Echinochloa crus-galli (L.) Beauv (Soerjani et al. 1987)
Batang berbentuk silindris dengan pith yang menyerupai spons putih di
bagian dalamnya. Di lahan sawah, anakan pertama dari E. crus-galli muncul 10 hari setelah perkecambahan, dan biasanya sekitar 15 anakan yang terbentuk
(Galinato et al. 1999). E. crus-galli memiliki jenis akar serabut dan tebal. Akar
E. crus-galli dihasilkan pada setiap ruasnya (Soerjani et al. 1987).
Bunga berupa malai yang berada di ujung dengan 5-40 bunga majemuk
bulir yang mempunyai tipe raceme, dengan cabang-cabang pendek yang menaik. Bunga majemuk terdiri atas banyak spikelet yang berbelok pada satu sisi, berbentuk tegak pada awalnya tetapi selanjutnya sering membengkok ke bawah.
Panjang malai bisa mencapai 5-21 cm. Malai kaku dengan permukaan yang agak
kasar. Bulir terbawah merupakan bulir yang paling panjang, sekitar 1.75-8 cm,
sedangkan bulir yang paling atas sangat pendek. Setiap bulir terdapat susunan
spikelet yang berselang-seling di setiap sisinya (Soerjani et al. 1987).
Spikelet tersusun soliter pada bulir paling atas dan susunannya bisa mencapai 2-4 spikelet dan pada bulir bagian bawah susunan spikelet bisa mencapai 4-10 spikelet (Soerjani et al. 1987). Spikelet tebal dan padat, sedikit berbentuk elips dengan panjang 3.2-3.5 mm. Spikelet biasanya sedikit berambut dan terkadang terdapat rambut yang tebal dan kaku yang panjangnya dapat
mencapai 13 mm. Spikelet berwarna kehijauan dan sedikit berwarna ungu (Ampong-Nyarko dan De Datta 1991).
Stamen pada E. crus-galli berjumlah 3 dengan anther berwarna kuning. Jumlah putik ada 2 dengan stigma berbulu, berwarna ungu, menonjol keluar di
bawah ujung spikelet. Buah E. crus-galli disebut caryopsis, berbentuk lonjong dengan panjang 1.5-2 mm, berbentuk ovoid sampai obovoid (Galinato et al. 1999). Lemma dari floret yang pertama memiliki permukaan yang datar atau sedikit cembung atau tumpul. Glume bagian bawah memiliki panjang sekitar
1.5-2.5 mm, berbentuk ovate, memendek dan memiliki ujung yang memendek secara bertahap. Glume bagian atas memiliki panjang yang sama dengan spikelet, berbentuk ovate-oblong, runcing, memiliki rambut yang tebal dan kaku sepanjang 0.5-3 mm serta berambut pendek (Galinato et al. 1999).
Biji yang tua berwarna kecoklat-coklatan sampai kehitaman. Produksi biji
(Ampong-Nyarko dan De Datta 1991). E. crus-galli mampu menghasilkan lebih
dari 1 000 kg benih/ha (Galinato et al. 1999).
Perbanyakan dan Penyebaran
Gulma E. crus-galli berperan sebagai gulma pada 36 jenis tanaman
budidaya di 61 negara. Jenis rumput ini memperbanyak diri secara generatif
melalui biji yang seringkali tercampur dengan benih padi (Galinato et al. 1999). Gulma ini bereproduksi dengan cara penyerbukan sendiri atau penyerbukan
silang. E. crus-galli melakukan penyerbukan silang dengan menggunakan bantuan angin. Biji E. crus-galli dapat menyebar melalui saluran irigasi, hewan, burung, pengangkutan biji padi dan mesin pertanian atau peralatan pertanian lainnya (Itoh
1991).
Perkembangan yang sangat cepat dan agresif dari E. crus-galli terkait dengan pertumbuhannya yang sangat cepat, produksi benih yang tinggi, dormansi
benih dan daya adaptasi yang tinggi di bawah kondisi lahan pertanian yang
berbeda (Bahrendt dan Hanf 1979). E. crus-galli adalah spesies yang sangat bervariasi, memiliki banyak bentuk dan variasi dengan waktu berbunga dan
menghasilkan biji yang sangat berbeda antara satu dengan yang lain.
Ekologi Gulma E. crus-galli Lingkungan Tumbuh
E. crus-galli merupakan gulma yang beradaptasi pada daerah berair, dan
tumbuh baik pada kelembaban tanah 80 % dari water capasity (Ampong-Nyarko dan De Datta 1991). Pertumbuhan E. crus-galli sangat baik pada jenis tanah berpasir dan berlempung terutama apabila kandungan nitrogennya tinggi (Kropff
dan Van Laar 1993).
Gulma ini mampu terus tumbuh walaupun hanya sebagian dari benih yang
terendam. Perkecambahan 30% lebih baik di tanah padat daripada di tanah yang
kurang padat. E. crus-galli juga mampu tumbuh dengan baik pada tanah yang lebih kering, tetapi memiliki pertumbuhan yang lebih kecil dan menghasilkan
jumlah malai, anakan dan jumlah biji yang lebih sedikit dibandingkan pada tanah
Menurut Soerjani et al. (1987) benih E. crus-galli tidak dapat berkecambah
pada kedalaman air lebih dari 12 cm, sedangkan menurut Kropff dan Van Laar (1993)
kedalaman air maksimum bagi perkecambahan benih E. crus-galli adalah 15 cm. Benih yang terendam pada kedalaman lebih dari 15 cm tidak dapat berkecambah.
Benih E. crus-galli dapat hidup terus dalam waktu yang lama. Benih yang
terdapat di dalam tanah dapat hidup terus sampai 1 tahun. Benih yang disimpan di
tempat penyimpanan dalam kondisi kering dapat hidup terus sampai 7 tahun.
Kelembaban optimum untuk perkecambahan benih E. crus-galli tergantung dari karakteristik tanah, tetapi umumnya pada 70-90% kapasitas lapang. Benih E. crus-galli
yang berada dekat dengan permukaan tanah akan berkecambah baik pada hari yang
panas (Galinato et al. 1999).
E. crus-galli dapat tumbuh pada daerah dataran rendah sampai sedang. Gulma ini tumbuh baik pada tempat dengan penyinaran penuh sepanjang tepi perairan
(Soerjani et al. 1987). E. crus-galli membutuhkan waktu 42-64 hari untuk melengkapi siklus hidupnya. Benih akan langsung tumbuh setelah ditanam tetapi sebagian lagi
mengalami dormansi yang bisa mencapai selama 4-48 bulan. Fotoperiodisme
mempengaruhi jumlah benih yang dorman dan intensitas dari dormansi tersebut
(Zimdahl et al. 2004).
Pembungaan dipengaruhi oleh panjang hari. Pada hari pendek pembungaan
lebih cepat terjadi. Jumlah malai dan anakan lebih besar pada hari pendek, tetapi
ukurannya kecil. Pada hari panjang (16 jam), gulma ini menghasilkan malai dengan
ukuran yang lebih besar dan jumlah benih yang lebih banyak. E. crus-galli yang
tumbuh pada daerah dengan penyinaran penuh memiliki bobot kering empat kali lebih
besar serta jumlah malai dan anakan dua kali lebih banyak daripada E. crus-galli yang tumbuh pada daerah dengan naungan 50% (Galinato et al. 1999).
Suhu lingkungan optimum untuk perkecambahan biji adalah 32-37°C. Tingkat
perkecambahan akan menurun drastis pada suhu lingkungan di bawah 10°C atau di atas
40°C dan berhenti berkecambah pada suhu 5o
Distribusi geografis dari E. crus-galli yaitu dari 50 °LU sampai 40 °LS (Holm
et al. 1977). Di Kanada telah dilaporkan bahwa E. crus-galli ditemukan pada tempat C. Pemanjangan kecambah sangat
tergantung pada persediaan oksigen. Benih tetap memiliki viabilitas yang tinggi
di atas 50 °LU, seperti di Edmonton (53° 33’ LU), Saskatoon (52° 07’ LU) dan Prince
Albert (53° 12’ LU) (Maun dan Barret 1986).
Plastisitas Fenotipik
Spesies tumbuhan dapat menyebar pada kondisi lingkungan secara luas, namun
kemampuan penyebaran setiap genotip terbatas. Kemampuan spesies menyebar secara
luas ditandai oleh kemampuan plastisitas fenotipik dan tingkat variasi genetik yang
tinggi (Santamaria et al. 2003). Plastisitas fenotipik adalah kemampuan suatu organisme untuk mengubah morfologi atau fisiologinya setelah terpapar atau berada
pada kondisi lingkungan yang berbeda atau ekspresi fenotipik yang tergantung pada
lingkungan (Schlichting 1986; Thompson 1991; Sultan 2000; deWitt & Scheiner
2004). Plastisitas fenotipik memainkan peranan yang penting dalam distribusi ekologi
suatu organisme (Sultan 2003). Plastisitas fenotipik membiarkan organisme untuk
menerima kondisi lingkungan yang berbeda dengan mengubah fenotipiknya untuk
menyesuaikan diri dengan lingkungannya sehingga menjadi adaptif (Plante & Hendry
2011). Sumber perbedaan lingkungan tumbuh seperti iklim, cahaya, dan ketinggian
tempat dapat menyebabkan adanya plastisitas fenotipik (Santamaria 2003; Sultan
2003). Kemampuan plastisitas fenotipik gulma E. crus-galli telah dilaporkan oleh banyak peneliti. Menurut Yamasue (2003) gulma E. crus-galli memiliki plastisitas fenotipik yang luas pada karakter tinggi tanaman yang tergantung dari tanaman
tetangganya dan membentuk daun dan malai yang lebih tinggi daripada kanopi
tanaman padi.
E. crus-galli dapat memiliki kemampuan “mimikri” yaitu menyerupai tanaman padi pada tahapan pertumbuhan tertentu dalam siklus hidupnya. Sifat ini muncul
melalui seleksi alami akibat tindakan penyiangan pada sistem pertanian yang intensif
(Barret 1983; Baki et al. 2003). Kemampuan mimikri membantu gulma E. crus-galli
untuk menghindar dari penyiangan manual (Yamasue 2003).
Fisiologi E. crus-galli
tanaman/tumbuhan C3 (Ampong-Nyarko dan De Datta 1991). Gulma dengan
siklus C4 mempunyai kapasitas tinggi dalam berproduksi dan berkompetisi serta
mempunyai kebutuhan air yang lebih rendah. Pada kondisi ketersediaan air yang
rendah, gulma ini secara umum mampu bersaing terhadap tanaman padi (Baki dan
Azmi 2003).
Alelopati Gulma E. crus-galli
Tumbuh-tumbuhan juga dapat bersaing antar sesamanya secara interaksi
biokimiawi, yaitu salah satu tumbuhan mengeluarkan senyawa beracun ke
lingkungan sekitarnya dan dapat mengakibatkan gangguan pertumbuhan pada
tumbuhan yang ada di dekatnya. Interaksi biokimiawi antara gulma dan
pertanamanan antara lain menyebabkan gangguan perkecambahan biji, kecambah
jadi abnormal, pertumbuhan memanjang akar terhambat, perubahan susunan
sel-sel akar dan lain sebagainya. Beberapa species gulma menyaingi pertanaman
dengan mengeluarkan senyawa beracun dari akarnya (root exudates atau lechates) atau dari pembusukan bagian vegetatifnya. Persaingan yang timbul akibat
dikeluarkannya zat yang meracuni tumbuhan lain disebut alelopati dan zat
kimianya disebut alelopat.
Menurut Sastroutomo (1990) senyawa alelopati dapat mempengaruhi
aktivitas tumbuhan antara lain menghambat penyerapan hara oleh akar tanaman,
pembelahan sel-sel akar, pertumbuhan tanaman, aktivitas fotosintesis,
mempengaruhi respirasi, sitesis protein, menurunkan daya permeabilitas membran
sel dan menghambat aktivitas enzim. Senyawa-senyawa kimia yang mempunyai
potensi alelopati dapat ditemukan di semua jaringan tumbuhan termasuk daun,
batang, akar, rizoma, umbi, bunga, buah, dan biji. Senyawa-senyawa alelopati itu
dapat dilepaskan dari jaringan-jaringan tumbuhan dalam berbagai cara termasuk
melalui penguapan, eksudat akar, pencucian dan pembusukan organ tumbuhan.
Beberapa gulma yang berpotensi alelopati baik yang masih hidup atau yang sudah
mati sama-sama dapat melepaskan senyawa alelopati melalui organ yang berada
di atas tanah maupun yang di bawah tanah. Menurut Putnam dan Weston (1986)
Selama perkecambahan dan awal pertumbuhan, E. crus-galli menekan
pertumbuhan beberapa tanaman pertanian termasuk padi dan E. crus-galli itu sendiri. Eksudat akar dari E. crus-galli muda menunjukkan efek alelopati dan menekan pemanjangan akar padi. Senyawa p-Hidroxymandelic acid merupakan
allelochemical yang dikeluarkan dari akar E. crus-galli muda yang dapat menekan
kuat pertumbuhan tajuk dan pemanjangan akar padi (Yamamoto et al. 1999). Eksudat akar E. crus-galli menekan perkecambahan dan pertumbuhan tanaman padi, lettuce dan monochoria. Komponen yang berpotensi terlibat dalam aktivitas
phytotoxic E. crus-galli telah teridentifikasi antara lain phenolic, long-chain fatty acids, loctones, diethyl phthalate, acenaphthene, phthalic acids, benzoic acid dan
decane. Penghambat pertumbuhan terbesar ditunjukkan oleh lactones, diikuti oleh
phenolic dan phthalic acid. Phytotoxins yang dikeluarkan akar E. crus-galli
memperlihatkan hambatan terhadap pertumbuhan tanaman indikator berdaun
lebar, tetapi kurang efektif pada tanaman padi dan E. crus-galli itu sendiri (Xuan
et al. 2006).
Produksi senyawa alelopati sangat dipengaruhi oleh beberapa faktor
lingkungan, diantaranya : 1) kualitas, intensitas dan lamanya masa penyinaran
cahaya dapat mempengaruhi produksi. Senyawa alelopati lebih banyak dihasilkan
pada kondisi cahaya ultraviolet dan periode penyinaran yang panjang; 2) jumlah
senyawa alelopati akan lebih banyak dihasilkan pada keadaan dengan kondisi
yang kekurangan hara; 3) senyawa alelopati lebih banyak dihasilkan dalam
keadaan yang mengalami gangguan kekeringan; 4) senyawa alelopati lebih
banyak dihasilkan dalam keadaan dengan suhu yang lebih rendah jika
dibandingkan dengan suhu normal bagi pertumbuhannya; 5) penggunaan hormon
seperti 2,4-D atau hidrasid maleat dapat meningkatkan senyawa alelopati (Rice
1974).
Kompetisi Padi dengan Gulma E. crus-galli
Gulma menurunkan pertumbuhan dan produksi tanaman melalui kompetisi
dalam memperebutkan cahaya, hara, air, dan ruang tumbuh (Kropff dan van Laar
1993). Tingkat kehilangan hasil akibat kompetisi tergantung pada tingkat
pertumbuhan gulma akan terhambat. Sebaliknya, apabila tanaman kurang vigor
dan tidak dapat menguasai sarana tumbuh, maka gulma akan tumbuh dengan
subur. Kompetisi tanaman dengan gulma tergantung pada beberapa faktor antara
lain lingkungan, pertumbuhan tanaman, kepadatan tanaman, fase pertumbuhan
tanaman, jenis gulma dan kepadatan gulma, fase pertumbuhan gulma, dan teknik
budidaya yang dilakukan (De Datta 1981).
Gulma E. crus-galli merupakan kompetitor yang sangat kuat terhadap tanaman padi sehingga menurunkan pertumbuhan dan produksi tanaman padi
(Chin 2003). Pertumbuhan tinggi tanaman padi menurun akibat kompetisi dengan
gulma E. crus-galli (Perera et al. 1992; Sultana 2000). Trung et al. (1995) melaporkan bahwa pada pertanaman padi pindah tanam, kepadatan gulma E. crus-galli 5-10 gulma per m2 menyebabkan kehilangan hasil 7-13% dan kepadatan 15-35 gulma per m2 menyebabkan kehilangan hasil sebesar 23-27%. Islam et al. (2003) juga melaporkan bahwa ketika tanaman padi berkompetisi dengan 8 gulma
E. crus-galli per pot, tinggi tanaman padi menurun hingga 42.9%, indeks luas daun (ILD) menurun sekitar 92%, jumlah anakan per tanaman menurun sekitar
72.7%, jumlah malai per rumpun menurun hingga 88.5%, dan jumlah gabah per
malai menurun hingga 63.8%.
Peubah-peubah Kompetisi
Terdapat dua jenis kompetisi yang biasa terjadi di alam yaitu kompetisi
intra spesifik dan interspesifik. Kompetisi intraspesifik adalah interaksi negatif
yang terjadi pada tumbuhan dengan jenis yang sama. Kompetisi interspesifik
adalah interaksi negatif yang terjadi pada tumbuhan yang berbeda jenis atau
disebut kompetisi antar jenis tumbuhan (Sastroutomo 1990). Menurut Eussen dan
Zulfadli (1981) kompetisi total merupakan gabungan dari kompetisi dan pengaruh
alelopati. Beberapa peubah kompetisi antara lain total hasil relatif (THR),
koefisien pendesakan, penguasaan sarana tumbuh (PST), dan agresivitas.
Total Hasil Relatif (THR) merupakan salah satu model untuk mempelajari kompetisi antara tumbuhan dengan tumbuhan lainnya (De Wit 1960).
THR adalah peubah yang dapat digunakan untuk mengetahui adanya kompetisi
percobaan replacement series atau seri penggantian yaitu pengaturan populasi
relatif agar campurannya tetap satu (satu). Pengaturan populasi dimulai dengan
menyusun suatu seri kerapatan monokultur tumbuhan pertama dengan populasi
relatif satu, kemudian berangsur-angsur diganti sampai menjadi monokultur
tumbuhan kedua dengan populasi relatif satu. Nilai THR dirumuskan :
THR = tII cII tI cI Y Y Y Y + Keterangan : YcI Y
= bobot kering tumbuhan 1 pada pertanaman campuran
tI
Y
= bobot kering tumbuhan 1 pada pertanaman tunggal
cII
Y
= bobot kering tumbuhan 2 pada pertanaman campuran
tII = bobot kering tumbuhan 2 pada pertanaman tunggal
Kelebihan pendekatan ini adalah bahwa terjadi kompetisi atau tidak dapat
diketahui dari nilai THR yang diperoleh. Nilai THR < 1 menunjukkan pengaruh
yang saling merugikan atau interaksi negatif diantara kedua tumbuhan atau
menunjukkan adanya kejadian kompetisi. Nilai THR = 1 menunjukkan tidak ada
kompetisi atau menunjukkan adanya pemakaian sarana tumbuh secara
bersama-sama. Nilai THR = 1 dapat pula menunjukkan kejadian kompetisi jika salah satu
tumbuhan lebih dominan menguasai sarana tumbuh yang ada. Nilai THR >1
menunjukkan kejadian penambahan sarana tumbuh yang tidak terukur,
penggunaan sarana tumbuh yang berbeda dari masing-masing tumbuhan, kejadian
simbiosis mutualisme atau interaksi positif diantara kedua tumbuhan, dan
ketiadaan kompetisi.
Relative Crowding Coefficient (Koefisien Pendesakan). Koefisien Pendesakan (KP) dapat menunjukkan kemampuan kompetisi suatu tumbuhan
terhadap tumbuhan lain (De Wit, 1960) yang dirumuskan sebagai berikut :
ij
c
=Keterangan :
Cij
Y
= koefisien pendesakan tumbuhan i terhadap tumbuhan j
i
Y
(campuran) = hasil tumbuhan i pada pertanaman campuran
i
Z
(monokultur) = hasil tumbuhan i pada pertanaman monokultur
i
Z
= rasio densitas tumbuhan i pada pertanaman campuran
j = rasio densitas tumbuhan j pada pertanaman campuran
Nilai KP yang lebih besar menunjukkan kemampuan kompetisi suatu
tanaman terhadap tanaman lain. Persamaan di atas berlaku pula sebaliknya, yaitu
koefisien pendesakan tumbuhan II terhadap tumbuhan I. Perbandingan koefisien
pendesakan antara kedua tumbuhan dapat menunjukkan tumbuhan yang lebih
kompetitif. Nilai koefisien pendesakan lebih tinggi menunjukkan derajat
kompetisi lebih besar.
Penguasaan Sarana Tumbuh (PST). Kompetisi antara gulma dan tanaman terjadi karena faktor tumbuh yang terbatas. Faktor yang dikompetisikan
berupa air, hara, CO2, cahaya, dan ruang tumbuh, namun sulit bagi kita untuk
menjelaskan faktor mana yang berperan dalam peristiwa kompetisi tersebut. Studi
kompetisi dari salah satu faktor penunjang pertumbuhan (unsur kompetisi) secara
terpisah sangat sulit dilakukan karena banyaknya faktor lingkungan yang terlibat
dalam pertumbuhan. De Wit (1960) memperkenalkan konsep penguasaan sarana
tumbuh yang mencakup semua faktor yang mempengaruhi kompetisi dan
merumuskan pertumbuhan tanaman dalam persamaan berikut :
Keterangan :
Y = hasil nyata
d = densitas tumbuhan Ymax
b = penguasaan sarana/faktor tumbuh (space occupation) atau kemampuan memanfaatkan sarana tumbuh
Persamaan garis tersebut masih bersifat hiperbolik. Resiprokal dari
persamaan tersebut akan memberikan persamaan linear sebagai berikut :
=
Angka Penguasaan Sarana Tumbuh ditetapkan dengan rumus :
PST = (Y/Ymax) x 100%
PST menyatakan besarnya penguasaan sarana tumbuh dan dinyatakan
dalam (%). Nilai PST yang lebih besar menunjukkan dominasi suatu tumbuhan
terhadap tumbuhan lain.
Agresivitas merupakan salah satu pendekatan untuk mengukur peningkatan hasil relatif dari suatu spesies dalam sistem tumpangsari yang
dikembangkan oleh McGilchrist dan disimbolkan dengan A (Whiley 1979).
Konsep ini didasarkan atas percobaan rangkaian substitusi dan untuk suatu
kombinasi substitusi. Nilai agresivitas dirumuskan sebagai berikut :
Nilai agresivitas sama dengan nol berarti semua spesies dalam pertanaman
campuran mempunyai daya kompetisi yang sama besarnya. Pada sisi lain, nilai
agresivitas mempunyai angka yang sama, tetapi spesies yang dominan memiliki
nilai positif sedangkan spesies yang didominasi memiliki nilai negatif.
Marka Molekuler Simple Sequence Repeat (SSR)
Penanda morfologi telah digunakan untuk mengatasi masalah duplikasi
plasma nutfah di lapang, penentuan jarak genetik dan hubungan kekerabatan antar
plasma nutfah/klon/kultivar (Vuylsteke et al. 1988; Ortiz et al. 1993; Swennen
et al. 1995; Soejono et al. 2001). Namun, penanda morfologi sering menunjukkan hasil yang bias karena pengaruh lingkungan terhadap penampakan fenotipik.
Untuk memperkuat informasi data penanda morfologi, diperlukan dukungan
aksesi berdasarkan karakteristik molekulernya (DNA) (Jarret dan Gawel 1995).
Penanda molekuler didasarkan pada polimorfisme yang dideteksi pada tingkat
makromolekul dalam sel. Pengertian ini akhir-akhir ini banyak digunakan untuk
mendiskripsikan hanya pada DNA. Penanda DNA ini dapat tidak terbatas dalam
jumlah dan dapat memberikan kegunaan yang besar untuk beragam tujuan yang
relevan terhadap perbaikan tanaman.
Penanda DNA yang memiliki tingkat akurasi cukup tinggi salah satunya
adalah simple sequen repeat (SSR) atau short tandem repeat (STR) atau mikrosatelit. SSR memiliki keunggulan mudah dan murah (pada tahapan setelah
ditemukan primer spesifiknya), keberadaannya melimpah dan tersebar di seluruh
genom tanaman, dan dengan sampel dalam jumlah sedikit, mencukupi untuk
amplifikasi dengan PCR (Ribaut et al 2002). Salah satu teknik yang memanfaatkan mikrosatelit adalah Sequence-tagged microsatellite sites (STMSs) atau Sequence-tagged sites (STS). Keuntungan STMSs adalah menggunakan sepasang primer yang sudah didisain khusus untuk tiap spesies (Sulyo 1997),
sehingga menyebabkan penanda ini bersifat ko-dominan (Hiu LB 1998;
Sulyo 1997). Penanda STMS memungkinkan mendapat derajat polimorfisme dan
variasi yang tinggi karena sekuen DNA mikrosatelit mengandung urutan basa
berulang-ulang secara bergandengan dengan panjang berbeda-beda pada genom.
Bentuk berulang yang umum adalah dinukleotida sederhana. Frekuensinya yang
tinggi dalam genom lebih mudah dideteksi dibandingkan mikrosatelit dengan
ulangan tri- dan tetranukleotida (Hiu Liu 1998). Mikrosatelit tri- dan
tetranukleotida lebih sedikit dalam genom dan tingkat keragamannya lebih rendah
dari dinukleotida (Scotti et al. 2002). Variasi terjadi dalam ukuran panjang mikrosatelit pada lokus-lokus individu yang spesifik, sehingga penanda ini
polialelik dan ko-dominan secara alami, yang menjadikan penanda ini mempunyai
manfaat lebih banyak (Puspendra et al. 2002) dan memiliki tingkat reprodusibilitas tinggi dibandingkan penanda RAPD dan RFLP (McGregor et al. 2000; Powell et al. 1996).
Studi yang membandingkan reprodusibilitas dari beberapa penanda DNA
pada tanaman kedelai menyimpulkan bahwa SSR mempunyai nilai informasi
heterosigositas yang diharapkan) dan spesifik lokus, sehingga menjadi penanda
pilihan untuk beragam tujuan termasuk pemuliaan tanaman (Powell et al. 1996). Pada studi kesamaan dan keanekaragaman genetik anggur, dari data molekuler
yang membandingkan tingkat efektivitas SSR dan AFLP, menunjukkan bahwa
Echinochloa crus-galli
YANG BERASAL DARI TIGA LOKASI
ABSTRAK
E. crus-galli merupakan gulma utama pada pertanaman padi sawah yang dapat menurunkan hasil panen. Tujuan penelitian adalah mempelajari pengaruh aksesi dan populasi gulma E. crus-galli terhadap pertumbuhan dan produksi tanaman padi. Penelitian dilaksanakan di rumah kaca, Kebun Percobaan IPB, Cikabayan, Bogor mulai bulan Mei 2006 hingga September 2006. Penelitian dilakukan dengan menggunakan rancangan petak terbagi dengan tiga ulangan. Aksesi gulma
E. crus-galli sebagai petak utama terdiri atas aksesi Karawang, Cikampek, dan Sukabumi. Populasi gulma E. crus-galli sebagai anak petak terdiri atas lima taraf yaitu 0, 1, 2, 3, dan 4 gulma E. crus-galli/pot. Hasil penelitian menunjukkan bahwa aksesi gulma E. crus-galli berpengaruh terhadap tinggi tanaman, jumlah anakan, dan kepadatan malai. Daya kompetisi gulma E. crus-galli aksesi Cikampek lebih tinggi dibandingkan dengan aksesi Sukabumi dan Karawang. Populasi E. crus-galli berpengaruh terhadap terhadap pertumbuhan dan produksi tanaman padi. Populasi 4 gulma E. crus-galli/pot menurunkan berat kering gabah total sebesar 48.0% dan berat kering gabah isi sebesar 46.2%. Interaksi antara aksesi dan populasi gulma E. crus-galli tidak berpengaruh terhadap pertumbuhan dan produksi tanaman padi.
GROWTH AND PRODUCTION OF RICE PLANT ON SOME
POPULATION DENSITY OF
Echinochloa crus-galli
WEED
FROM THREE LOCATIONS
ABSTRACT
Echinochloa crus-galli is a major weed in paddy field that reduces rice yield. The objective of the research was to study the effect of E. crus-galli accession and population on rice growth and production. The research was conducted in a green house using split plot design with three replications. The main plot consisted of three E. crus-galli accession i.e. Karawang, Cikampek, and Sukabumi accession.
E. crus-galli population as sub plot consisted of 0, 1, 2, 3, and 4 E. crus-galli per pot. The results showed that accession of E. crus-galli affected plant height, number of tiller, and panicle density. The competitiveness against rice of E. crus-galli accession Cikampek was higher than that of Sukabumi dan Karawang accession. Population E. crus-galli affected rice growth and production. Population of 4 E. crus-galli/pot decreased spikelets weigth about 48.0% and filled spikelets weigth about 46.2%. Interaction of accession and population of E. crus-galli did not affected rice growth and production.
Pendahuluan
Kebutuhan beras semakin meningkat sejalan dengan peningkatan jumlah
penduduk. Pada tahun 2030 nanti, kebutuhan beras Indonesia diperkirakan
mencapai 41.7 juta ton (BPS 2008). Upaya peningkatan produksi beras pada
masa yang akan datang dihadapkan pada berbagai kendala seperti alih fungsi
lahan pertanian ke non pertanian, degradasi kesuburan lahan, dan serangan
organisme pengganggu tanaman (OPT).
Salah satu OPT yang dapat menurunkan produksi tanaman padi adalah
gulma. Penurunan produksi yang diakibatkan oleh gulma pada beberapa situasi
secara ekonomis lebih penting daripada penurunan produksi yang disebabkan oleh
insekta, cendawan, atau organisme pengganggu lainnya (Savary et al. 1997; 2000). Selain penurunan produksi, adanya gulma di pertanaman padi sawah juga
menyebabkan biaya pengendalian yang besar sehingga menurunkan pendapatan
petani (Tungate et.al. 2007).
Salah satu spesies gulma dominan pada lahan sawah adalah Echinochloa crus-galli (Ali dan Sankaran, 1984). Kehadiran gulma E. crus-galli di pertanaman padi sawah dapat menurunkan produksi tanaman padi hingga 50-59%
(Sultana 2000; Chin 2001), 57-95% (Ahn dan Chung 2000), dan bahkan dapat
menurunkan produksi gabah hingga 97% (Islam dan Karim 2003). Penurunan
produksi tersebut disebabkan oleh adanya kompetisi antara gulma dan tanaman
padi terhadap sumberdaya yang tersedia (Zimdahl 2004). Gulma ini juga dapat
menjadi tumbuhan inang bagi Leptocorisa oratorius, Acrocylindricum oryzae,
Corticium sasakii, dan Rhynchosporium oryzae (Tjitrosemito 1994).
Gulma E. crus-galli memiliki daya adaptasi yang luas pada kondisi lingkungan yang beragam (Galinato et al. 1999). Karena kemampuan adaptasi yang luas, maka gulma E. crus-galli dari tiap aksesi diduga memiliki daya
kompetisi yang berbeda pula. Perubahan praktek agronomis pada berbagai lokasi
dari waktu ke waktu seperti penggunaan herbisida baru, inovasi cara pengolahan
tanah, penggunaan kultivar baru dapat mempengaruhi distribusi gulma dan
kemampuan kompetisi gulma terhadap tanaman budidaya (Froud-Williams et al. 1984; Clement et al. 1996). Perbedaan karakter daya kompetisi dari aksesi gulma
Indonesia. Penelitian bertujuan untuk mempelajari pengaruh aksesi gulma E. crus-galli pada beberapa tingkat populasi terhadap pertumbuhan dan produksi
tanaman padi sawah.
Bahan dan Metode
Penelitian dilakukan dalam pot di rumah kaca Kebun Percobaan Cikabayan,
Dramaga, Bogor, pada bulan Mei 2006 sampai dengan September 2006. Bahan
yang digunakan antara lain benih padi varietas IR-64, pupuk urea, SP-36, dan
KCl, dan biji E. crus-galli. Peralatan yang digunakan antara lain pot berukuran 30 cm - 40 cm (diameter - tinggi), tray, neraca, oven, dan leaf area meter.
Percobaan menggunakan rancangan petak terbagi (split plot design) dalam rancangan acak kelompok dengan tiga ulangan. Percobaan terdiri atas dua faktor,
yaitu aksesi E. crus-galli sebagai petak utama yang terdiri atas tiga aksesi (aksesi Karawang, Cikampek, dan Sukabumi) dan populasi E. crus-galli sebagai anak petak yang terdiri atas lima taraf, yaitu 0, 1, 2, 3, dan 4 bibit gulma E. crus-galli
per pot. Satuan percobaan terdiri atas 3 pot sehingga total terdapat 135 pot
percobaan.
Media tanam yang digunakan adalah tanah latosol dramaga yang berasal
dari lahan sawah kebun percobaan IPB Sawah Baru. Sebelum digunakan sebagai
media, tanah dikeringanginkan terlebih dahulu, kemudian dihaluskan dan diayak.
Tiap pot diisi media tanah sebanyak 10 kg/pot. Media tanah dalam pot
selanjutnya dilumpurkan dan digenangi dengan air hinga ketinggian 5 cm dari
permukaan media. Benih padi varietas IR-64 dan biji E. crus-galli disemai sebelum penanaman di pot dengan menggunakan bak semai. Bibit padi
dipindahtanam ke dalam pot pada saat berumur 21 hari setelah semai (HSS).
Bibit padi ditanam tepat di tengah-tengah pot. Bibit gulma E. crus-galli yang
berumur 14 HSS ditanam pada jarak 7 cm dari tanaman padi dengan jumlah bibit
sesuai dengan perlakuan.
Pemeliharaan tanaman yang dilakukan meliputi pemupukan, penyiraman,
pengendalian hama penyakit. Pupuk SP-36 dan KCl dengan dosis masing-masing
sebesar 0.5 g/pot diberikan seluruhnya pada saat tanam, sedangkan pupuk urea
pada 4 minggu setelah tanam (MST), dan 1/3 dosis pada 8 MST. Penyiraman
dilakukan 2 hari sekali sampai ketinggian genangan sekitar 5 cm. Penyiangan
gulma selain E. crus-galli dilakukan secara manual. Pengendalian penyakit tungro dilakukan dengan cara membuang bagian tanaman yang terserang. Panen
padi dan gulma E. crus-galli dilakukan bersamaan pada 13 MST.
Peubah yang diamati antara lain tinggi tanaman, jumlah anakan, bobot
biomassa tajuk dan akar, panjang dan lebar daun bendera, luas daun bendera,
kadar nitrogen daun bendera, jumlah anakan produktif, panjang malai, kepadatan
malai, dan produksi gabah. Data hasil pengamatan dianalisis menggunakan
analisis ragam (uji F) dengan uji lanjut DMRT (Duncan’s Multiple Range Test) pada taraf nyata 5%.
Hasil dan Pembahasan
Pertumbuhan Tanaman Padi
Aksesi gulma E. crus-galli berpengaruh terhadap tinggi tanaman padi pada saat 7 MST (Tabel 1). Tinggi tanaman padi yang ditanam dengan gulma E. crus-galli aksesi Sukabumi lebih pendek dibandingkan dengan yang ditanam dengan gulma aksesi lainnya.
Tabel 1. Pengaruh aksesi gulma terhadap tinggi tanaman padi
Aksesi Tinggi Tanaman Padi (cm)
2 MST 3 MST 4 MST 5 MST 6 MST 7 MST 8 MST
Karawang 39.8 54.6 67.0 80.9 89.4 92.4a 95.1
Cikampek 38.7 53.0 66.4 80.2 87.6 91.4a 93.7
Sukabumi 40.2 52.5 67.7 80.4 86.6 89.4b 92.0
Keterangan: angka yang diikuti huruf yang sama pada kolom yang sama, tidak berbeda nyata pada uji DMRT taraf 5%.
Populasi dan interaksi antara aksesi dengan populasi E. crus-galli tidak berpengaruh terhadap tinggi tanaman padi. Hasil berbeda dilaporkan oleh Islam et al. (2003) bahwa keberadaan gulma E. crus-galli mulai populasi 2 per pot menurunkan tinggi tanaman padi. Perera et al. (1992), Sultana (2000) dan Purba (2007) juga melaporkan adanya penurunan tinggi tanaman padi akibat kompetisi
Aksesi E. crus-galli berpengaruh terhadap jumlah anakan tanaman padi
pada 2 MST; populasi E. crus-galli berpengaruh pada 2 MST dan 5-8 MST; sedangkan interaksi antara keduanya tidak berpengaruh terhadap jumlah anakan
tanaman padi (Tabel 2).
Tabel 2. Pengaruh aksesi dan populasi E. crus-galli terhadap jumlah anakan tanaman padi
Perlakuan Jumlah Anakan
2 MST 3 MST 4 MST 5 MST 6 MST 7 MST 8 MST Aksesi
Karawang 2.2ab 3.9 6.5 9.4 12.0 13.9 14.2
Cikampek 2.1b 3.3 6.0 8.0 9.8 11.0 11.7
Sukabumi 2.6a 4.8 7.8 11.1 13.5 14.8 14.5
Populasi E. crus-galli/pot
0 2.2b 4.0 6.2 8.9b 12.3b 15.0ab 17.3a
1 2.6a 4.5 7.8 11.4a 14.7a 17.1a 16.9a
2 2.3ab 3.9 7.1 9.6ab 11.7bc 12.6bc 12.2b
3 2.3ab 4.0 6.6 9.2b 10.8bc 11.7c 11.5b
4 2.1b 3.6 6.2 8.3b 9.4c 9.8c 9.6b
Keterangan: angka yang diikuti huruf yang sama pada kolom dan faktor perlakuan yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Aksesi Cikampek pada 2 MST menyebabkan jumlah anakan padi lebih
rendah dibandingkan dengan aksesi Sukabumi dan Karawang, namun pada
pengamatan 3-8 MST tidak menunjukkan perbedaan. Populasi 2 gulma E. crus-galli/pot pada pengamatan 8 MST nyata menurunkan jumlah anakan padi dibandingkan terhadap kontrol. Populasi 4 gulma/pot menyebabkan jumlah
anakan padi menurun hingga 53.8% dibandingkan terhadap kontrol. Hasil
tersebut sesuai dengan penelitian Sutrisno dan Turanto (1981) bahwa tanaman
padi yang ditanam dengan E. crus-galli pada saat tanam menunjukkan jumlah anakan total yang lebih rendah dibandingkan dengan padi yang ditanam tanpa
E. crus-galli.
Jumlah daun tanaman padi dipengaruhi oleh populasi E. crus-galli, namun tidak dipengaruhi oleh aksesi E. crus-galli maupun interaksi antara aksesidengan populasi E. crus-galli (Tabel 3). Populasi E. crus-galli mulai 2 gulma per pot nyata menurunkan jumlah daun tanaman padi dibandingkan terhadap kontrol.
crus-galli. Namun demikian, populasi 4 gulma E. crus-galli menghasilkan jumlah
daun yang sebanding dengan populasi 3 gulma E. crus-galli mulai pengamatan 9 MST sampai dengan 13 MST.
Tabel 3. Jumlah daun tanaman padi pada perlakuan populasi E. crus-galli
Populasi E. crus-galli/pot
Jumlah Daun Tanaman Padi (helai)
9 MST 10 MST 11 MST 12 MST 13 MST
0 78.3a 76.0a 60.4a 56.2a 53.9a
1 79.1a 71.7ab 50.0ab 45.3ab 43.7ab
2 66.1a 59.3b 42.7b 38.2b 36.6b
3 49.9b 42.9c 29.2c 25.4c 24.6c
4 43.1b 35.6c 23.0c 19.7c 18.9c
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Jumlah daun tanaman padi terkait dengan jumlah anakan per rumpun.
Semakin tinggi populasi E. crus-galli jumlah anakan tanaman padi semakin menurun dan akhirnya menurunkan jumlah daun per rumpun. Penurunan jumlah
anakan dan jumlah daun tanaman padi diduga disebabkan oleh adanya kompetisi
antar spesies antara tanaman padi dengan gulma E. crus-galli dalam memperebutkan hara. Kekurangan hara P dapat menyebabkan penurunan jumlah
anakan tanaman padi. Selain itu, penurunan jumlah anakan dan jumlah daun
diduga disebabkan oleh adanya zat alelopati yang dikeluarkan oleh gulma E. crus-galli. Yamamoto et al. (1999) dan Xuan et al. (2006) menyatakan bahwa eksudat akar E. crus-galli yaitu senyawa hidroxymandelic acid dan lactones,
menyebabkan penurunan perkecambahan dan pertumbuhan tanaman padi.
Bobot kering tajuk tanaman padi pada 2 bulan setelah tanam (BST) dan
3 BST dipengaruhi oleh populasi E. crus-galli. Aksesi E. crus-galli maupun interaksi antara aksesi dengan populasi E. crus-galli tidak berpengaruh terhadap
bobot kering tajuk tanaman padi (Tabel 4). Gulma E. crus-galli pada pengamatan 3 BST menurunkan bobot kering tajuk tanaman padi mulai populasi 2 gulma/pot
dengan penurunan sebesar 30.9% dibandingkan terhadap kontrol. Eussen dan
Zulfadli (1981) menyatakan bahwa produksi bahan kering tanaman menurun
akibat ditanam bersama dengan gulma pada seluruh siklus pertumbuhannya.
tanaman padi dengan gulma E. crus-galli dalam memperebutkan unsur hara serta
adanya alelopati gulma E. crus-galli.
Bobot kering akar tanaman padi pada 3 BST dipengaruhi oleh populasi
E. crus-galli, namun tidak dipengaruhi oleh aksesi E. crus-galli maupun interaksi antara aksesi E. crus-galli dengan populasi. Pada pengamatan 3 BST terlihat
bahwa semakin tinggi populasi E. crus-galli maka bobot kering akar tanaman padi semakin rendah (Tabel 4).
Tabel 4. Bobot kering tajuk dan akar padi pada perlakuan populasi E. crus-galli
Populasi E. crus-galli/pot
Bobot Kering Tajuk Bobot Kering Akar 1 BST 2 BST 3 BST 1 BST 2 BST 3 BST
--- (g/pot) ---
0 1.4 16.6ab 23.0a 1.2 11.4 5.0a
1 1.5 19.2a 19.0ab 1.9 10.7 5.0a
2 1.4 13.4b 15.9bc 0.7 7.8 3.9ab
3 1.4 15.2ab 12.6cd 1.3 7.6 3.2bc
4 1.3 12.1b 10.2d 1.0 5.6 2.2c
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Penurunan bobot kering akar tanaman padi dibandingkan dengan kontrol
terlihat mulai populasi 3 gulma E. crus-galli per pot. Populasi 4 gulma E. crus-galli per pot menghasilkan bobot kering akar yang paling rendah yaitu sebesar 2.2 g/pot. Penurunan bobot kering akar akibat kompetisi dengan gulma E. crus-galli
juga dilaporkan oleh Ranasinghe dan Crabtree (1999) bahwa efek kompetisi dari
E. crus-galli pada tanaman padi yaitu menurunkan bobot kering tanaman padi dan penurunan meningkat dengan peningkatan kepadatan E. crus-galli. Zimdahl (2004) menyatakan bahwa kompetisi antara dua tanaman terjadi karena
memperebutkan sumberdaya dalam ruang tumbuh yang sama. Penurunan bobot
kering akar ini diduga disebabkan oleh adanya hambatan dalam perkembangan
akar tanaman padi akibat kompetisi dalam mendapatkan ruang tumbuh.
Panjang, lebar dan luas daun bendera tanaman padi dipengaruhi oleh
dibandingkan dengan kontrol. Semakin tinggi populasi E. crus-galli panjang daun
dan luas daun bendera semakin rendah.
Daun merupakan bagian tanaman yang dipengaruhi oleh unsur nitrogen
dan salah satu fungsi dari unsur nitrogen adalah meningkatkan ukuran daun.
Menurut Takeda (1961) nitrogen yang diserap tanaman dapat meningkatkan luas
daun. Panjang dan lebar daun bendera adalah dimensi dari luas daun bendera.
Tanaman padi yang ditanam dengan E. crus-galli mengalami kompetisi dalam mendapatkan unsur nitrogen, sehingga panjang, lebar dan luas daun bendera lebih
rendah dibandingkan dengan yang ditanam tanpa gulma E. crus-galli.
Tabel 5. Panjang, lebar dan luas daun bendera padi dan kadar N daun bendera padi pada perlakuan populasi E. crus-galli
Populasi E. crus-galli/pot
Daun Bendera Panjang Daun
(cm)
Lebar Daun (cm)
Luas Daun (cm2
Kadar N
) (%)
0 30.6a 1.3a 381.2a 1.5a
1 26.7b 1.2b 245.8b 1.1b
2 27.6ab 1.1b 204.7bc 1.1b
3 28.6ab 1.1b 127.5bc 1.0b
4 25.5b 1.1b 115.5c 0.9b
Keterangan: angka-angka yang diikuti huruf yang sama pada kolom yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Kadar nitrogen pada daun bendera tanaman padi dipengaruhi oleh populasi
gulma E. crus-galli, tetapi tidak dipengaruhi oleh aksesi gulma E. crus-galli
maupun interaksi antara aksesi dengan populasi gulma E. crus-galli. Populasi
gulma E. crus-galli sebanyak 1 per pot menyebabkan penurunan kadar nitrogen pada daun bendera tanaman padi sebesar 26.7% dibandingkan dengan kontrol.
Hasil ini menunjukkan bahwa gulma E. crus-galli berkompetisi dengan tanaman padi dalam mendapatkan hara nitrogen. Arai dalam Sutrisno dan Turanto (1981)
Komponen Produksi Padi
Populasi gulma E. crus-galli berpengaruh terhadap jumlah anakan produktif tanaman padi, namun aksesi E. crus-galli dan interaksi antara aksesi dengan populasi E. crus-galli tidak berpengaruh terhadap jumlah anakan produktif tanaman padi (Tabel 6). Jumlah anakan produktif tanaman padi mulai menurun
pada perlakuan populasi 2 gulma E. crus-galli per pot. Semakin tinggi populasi gulma E. crus-galli, maka jumlah anakan produktif semakin menurun. Tanaman padi yang ditanam dengan populasi 4 gulma E. crus-galli per pot memiliki jumlah anakan produktif yang terendah yaitu 7.6 anakan.
Tabel 6. Jumlah anakan produktif, panjang malai dan kepadatan malai padi pada perlakuan aksesi dan populasi E.crus-galli
Perlakuan Jumlah Anakan
Produktif
Panjang Malai (cm)
Kepadatan Malai (butir/cm) Aksesi
Karawang 12.6 21.9 4.2a
Cikampek 11.1 21.6 2.8b
Sukabumi 12.6 21.7 4.2a
Populasi E. crus-galli per Pot
0 17.0a 21.8 4.8a
1 15.3a 21.8 4.7a
2 12.0b 22.0 3.8ab
3 8.6c 21.4 3.0bc
4 7.6c 21.7 2.4c
Keterangan: angka yang diikuti huruf yang sama pada kolom dan faktor perlakuan yang sama tidak berbeda nyata pada uji DMRT taraf 5%.
Perlakuan aksesi E. crus-galli dan populasinya berpengaruh terhadap kepadatan malai, namun tidak berpengaruh terhadap panjang malai padi. Interaksi
antara aksesi E. crus-galli dengan populasi tidak berpengaruh terhadap panjang malai padi dan kepadatan malai. Panjang malai rata-rata dari perlakuan aksesi
E. crus-galli dan populasi E. crus-galli berturut-turut adalah 21.73 cm dan 21.74 cm (Tabel 6). Hasil penelitian Tobing dan Chozin (1980) juga menunjukkan
bahwa rata-rata panjang malai pada perlakuan penyiangan gulma dan kontrol
tidak berpengaruh secara nyata.
Kepadatan malai menurun dengan meningkatnya populasi E. crus-galli.
kepadatan malai terendah yaitu 2.4 butir/cm. Penurunan kepadatan malai pada
populasi 4 E. crus-galli per pot sebesar 50.1% dibandingkan terhadap kontrol (Tabel 6). Gulma E. crus-galli aksesi Cikampek memiliki daya kompetisi yang lebih besar dalam menurunkan kepadatan malai yang ditunjukkan dengan
kepadatan malai yang lebih rendah dibandingkan dengan aksesi Karawang dan
Sukabumi (Tabel 6). Daya kompetisi yang lebih besar ini diduga sebagai akibat
praktek budidaya tanaman padi sawah di lokasi Cikampek yang dinamis
dibandingkan dengan aksesi Sukabumi dan aksesi Karawang. Menurut
Froud-Williams et al. (1984) dan Clement et al. (1996) praktik agronomi tanaman yang tidak statis dalam waktu dan ruang seperti penggunaan herbisida kelas baru,
kultivar, inovasi pengolahan tanah, penggunaan irigasi, dapat mempengaruhi
distribusi geografis gulma dan daya kompetisi gulma dalam menurunkan
produksi.
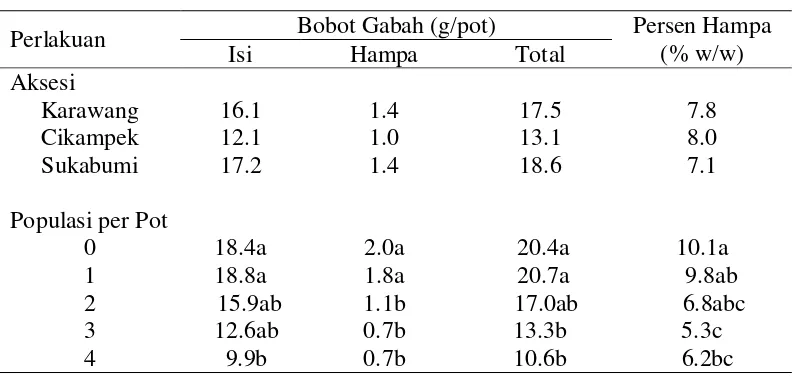
Produksi Gabah
Populasi gulma E. crus-galli berpengaruh terhadap bobot gabah total, bobot gabah isi, bobot gabah hampa, dan persentase gabah hampa. Aksesi gulma
tidak berpengaruh terhadap bobot gabah total, bobot gabah isi, bobot gabah
[image:30.595.113.509.527.715.2]hampa, dan persentase gabah hampa (Tabel 7).
Tabel 7. Pengaruh aksesidan populasi E. crus-galli terhadap bobot gabah dan persentase kehampaan
Perlakuan Bobot Gabah (g/pot) Persen Hampa
(% w/w)
Isi Hampa Total
Aksesi
Karawang 16.1 1.4 17.5 7.8
Cikampek 12.1 1.0 13.1 8.0
Sukabumi 17.2 1.4 18.6 7.1
Populasi per Pot
0 18.4a 2.0a 20.4a 10.1a
1 18.8a 1.8a 20.7a 9.8ab
2 15.9ab 1.1b 17.0ab 6.8abc
3 12.6ab 0.7b 13.3b 5.3c
4 9.9b 0.7b 10.6b 6.2bc
Populasi gulma E. crus-galli sebanyak 4 gulma per pot menurunkan bobot
gabah isi sebesar 46.2% dan menurunkan bobot gabah total sebesar 48.0%
dibandingkan terhadap kontrol. Namun demikian, persen kehampaan pada
populasi gulma E. crus-galli sebanyak 4 gulma per pot menurun dibandingkan dengan tanpa gulma. Bobot gabah isi yang rendah diduga disebabkan oleh adanya
kompetisi dalam mendapatkan unsur nitrogen. Menurut De Data (1981) salah satu
fungsi nitrogen pada tanaman padi adalah meningkatkan jumlah gabah isi.
Kesimpulan
Perbedaan aksesi gulma E. crus-galli menyebabkan perbedaan pertumbuhan dan produksi tanaman padi khususnya pada peubah tinggi tanaman
pada 7 MST, jumlah anakan pada 2 MST, dan kepadatan malai pada saat panen.
Gulma E. crus-galli aksesi Cikampek menunjukkan daya kompetisi yang lebih kuat dibandingkan aksesi Karawang dan Sukabumi berdasarkan penurunan jumlah
anakan pada 2 MST dan penurunan kepadatan malai pada saat panen.
Kepadatan polulasi gulma E. crus-galli per pot menentukan tingkat pertumbuhan dan produksi tanaman padi sawah. Semakin tinggi populasi gulma
E. crus-galli pengaruh kompetisi terhadap tanaman padi semakin besar. Populasi gulma E. crus-galli sebanyak 4 per pot menurunkan bobot gabah sebesar 48.0% dan menurunkan bobot gabah isi sebesar 46.2%.
Penelitian lanjutan tentang keragaman morfologi dan genetik aksesi gulma
E. crus-galli dari berbagai lokasi di Jawa Barat penting dilakukan untuk
mengetahui apakah keragaman aksesi gulma disebabkan oleh perbedaan geografis
KERAGAMAN MORFOLOGI DAN GENETIK
AKSESI GULMA
Echinochloa crus-galli
(L.) Beauv.
ASAL JAWA BARAT
ABSTRAK
Gulma E. crus-galli merupakan gulma dominan pada tanaman padi sawah yang menunjukkan variasi morfologi dan genetik. Penelitian bertujuan untuk menganalisis keragaman morfologi dan genetik aksesi gulma E. crus-galli dari berbagai lokasi di Jawa Barat. Sebanyak 16 aksesi E. crus-galli dikoleksi dari tujuh kabupaten di Jawa Barat. Hasil penelitian menunjukkan bahwa aksesi gulma E. crus-galli asal Jawa Barat menunjukkan adanya keragaman morfologi dan genetik. Aksesi gulma E. crus-galli menunjukkan keragaman morfologi baik pada habitat asal maupun pada kondisi rumah kaca. Berdasarkan marka molekuler SSR, aksesi gulma E. crus-galli dapat dikelompokkan menjadi empat sub grup, dimana hampir seluruh aksesi asal Jawa Barat mengelompok ke dalam sub grup A dan C. Pusat keragaman aksesi gulma E. crus-galli diduga berasal dari Subang, Karawang, dan Pangalengan. Implikasi penelitian adalah bahwa tindakan agronomi terpadu harus dilakukan agar penyebaran aksesi gulma dapat dikendalikan.
MORPHOLOGY AND GENETICAL DIVERSITY OF
Echinochloa crus-galli
(l.) Beauv. WEED ACCESSION
FROM WEST JAVA
ABSTRACT
E. crus-galli is a major weed in the paddy field in Indonesia. This weed shows morphology and genetical variations. The objective of the research was to analyze the morphology and genetic diversity of E. crus-galli accession from some locations in West Java, Indonesia. Genetic diversity of E. crus-galli accession analyze by using SSR markers. The E. crus-galli was collected from 7 district in West Java, Indonesia. The results showed that genetic variations were observed among districts and within district. E. crus-galli accession showed morphology diversity both in origin habitat and in greenhouse. Based on SSR marker, accessions could be grouped into four sub groups, where sub group A and C consisted the most accessions from West Java. It is likely that center of diversity of E. crus-galli is located in Subang, Karawang and Pangalengan. This experiment implies that integrated agronomic treatments should be conducted in order to control distribution of E. crus-galli in West Java.
Pendahuluan
Echinochloa crus-galli (L.) Beauv. merupakan gulma semusim yang
memiliki kemampuan adaptasi ekologi yang luas dan daya kompetisi yang kuat
sehingga menjadi spesies gulma penting dan dominan pada tanaman padi sawah
(Altop et al. 2011). Gulma E. crus-galli (L.) Beauv. dapat menyebabkan
kehilangan hasil produksi tanaman padi sawah (Ali dan Sankaran 1984; Ali 1985;
Gealy et al. 2003; Haefele et al. 2004) dengan penurunan hasil produksi padi mencapai 46-59% (Sultana 2000; Chin 2001; Guntoro et al. 2009), 57-95% (Ahn dan Chung 2000), dan bahkan mencapai 97% (Islam dan Karim 2003).
Gulma E. crus-galli (L.) Beauv. memiliki distribusi yang luas, mampu beradaptasi pada berbagai ekologi, toleran terhadap kondisi iklim kering dan
kondisi anaerob, memiliki kemampuan mimikri, perkecambahan dan pertumbuhan
yang cepat, produksi biji yang banyak, sehingga spesies ini menjadi gulma di
lebih dari 60 negara (Barret 1983; Altop et al. 2011). Aoki dan Yamaguchi (2008) juga melaporkan bahwa gulma E. crus-galli memperlihatkan keragaman yang sangat tinggi dalam morfologi dan kemampuan beradaptasi pada kondisi
lingkungan yang beragam. Tasrif et al. (2004) melaporkan adanya keragaman karakter tinggi gulma, panjang malai, luas daun, jumlah biji per malai, panjang
bulu, dan karakter fenotipik lainnya dari E. crus-galli yang disebabkan oleh perbedaan geografi lokasi aksesi. Menurut Altop et al. (2011), perubahan morfologi seperti perubahan susunan kanopi akan mempengaruhi kemampuan
kompetisi spesies. Oleh karena itu, informasi tingkat keragaman morfologi dalam
spesies sangat penting untuk menentukan strategi pengendalian yang efektif.
Identifikasi menggunakan karakter morfologi merupakan salah satu upaya
untuk mendapatkan informasi mengenai keragaman genetik. Penanda morfologi
telah digunakan untuk mengatasi masalah duplikasi plasma nutfah di lapang,
penentuan jarak genetik dan hubungan kekerabatan antar plasma
nutfah/klon/kultivar (Vuylsteke et al. 1988; Ortiz et al. 1993; Swennen et al.
1995; Soejono et al. 2001). Untuk memperkuat informasi data
penanda morfologi, diperlukan dukungan penanda molekuler (Jarret dan Gawel
1995). Penggunaan teknik molekuler untuk mengetahui keragaman genetik aksesi
beradaptasi dengan lingkungannya, juga untuk merencanakan strategi
pengendalian yang tepat (Nissen et al. 1995).
Biologi molekuler saat ini memainkan peranan penting dalam studi
ekologi. Dengan berkembangnya teknik molekuler, marka molekuler yang lebih
beragam telah tersedia untuk menginvestigasi genetik ekologi gulma (Moodie
et al. 1997; Ash et al. 2003; Altop et al. 2011). Metode berdasarkan polymerase chain reaction (PCR), seperti random amplified polymorphic DNA (RAPD) (Williams et al. 1990) dan inter simple sequence repeats (ISSR) (Zietkewicz et al. 1994) telah banyak diaplikasikan untuk survey keanekaragaman populasi genetik. Simple sequence repeat (SSR) atau short tandem repeat (STR) atau mikrosatelit diyakini memiliki tingkat akurasi yang lebih tinggi dibandingkan dengan penanda
molekuler yang lain. SSR digunakan sebagai penanda karena relatif lebih mudah
dan murah, keberadaannya tersebar di seluruh genom tanaman, dan sampel yang
diperlukan lebih sedikit (Ribaut et al. 2002).
Studi variasi genetik gulma E. crus-galli di Indonesia dengan menggunakan marka molekuler RAPD telah dilakukan oleh Tasrief et al. (2004). Namun demikian, informasi tentang keragaman morfologi dan genetik antar
aksesi gulma E. crus-galli dari berbagai lokasi masih sangat terbatas. Penelitian bertujuan untuk menganalisis keragaman morfologi dan genetik aksesi gulma
E. crus-galli dari berbagai lokasi di Jawa Barat.
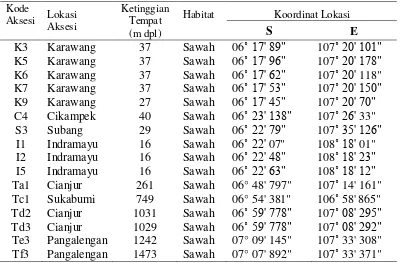
Metode Penelitian Evaluasi Keragaman Morfologi Aksesi E. crus-galli
Evaluasi dilakukan di rumah kaca Kebun Percobaan IPB Darmaga, Bogor
yang berada pada ketinggian 250 m dpl. Sebanyak 16 aksesi E. crus-galli diperoleh dengan cara mengumpulkan biji-biji gulma yang telah matang dari
berbagai habitat sawah di Jawa Barat yaitu Indramayu, Subang, Cikampek,
Karawang, Sukabumi, Cianjur, dan Pangalengan-Bandung. Koleksi biji gulma
dilakukan pada bulan Maret – April 2007. Lokasi pengambilan contoh aksesi
ditetapkan berdasarkan ketinggian tempat (Tabel 8). Selanjutnya, biji disemai pada
bak semai dan pada saat 14 hari setelah semai bibit gulma dipindahtanam ke dalam
Darmaga sebanyak 9 kg/pot. Setiap aksesi ditanam pada tiga pot. Pemeliharaan
gulma di dalam pot meliputi pemupukan dengan dosis pupuk 0.5 g N/pot, 0.5 g
[image:36.595.113.515.187.450.2]P2O5/pot, dan 0.5 g K2O/pot, dan penyiraman dilakukan tiap pagi hari.
Tabel 8. Deskripsi asal-usul 16 aksesi gulma E. crus-galli di Jawa Barat
Kode
Aksesi Lokasi
Aksesi
Ketinggian Tempat (m dpl)
Habitat Koordinat Lokasi
S E
K3 Karawang 37 Sawah 06˚ 17' 89" 107˚ 20' 101" K5 Karawang 37 Sawah 06˚ 17' 96" 107˚ 20' 178" K6 Karawang 37 Sawah 06˚ 17' 62" 107˚ 20' 118" K7 Karawang 37 Sawah 06˚ 17' 53" 107˚ 20' 150" K9 Karawang 27 Sawah 06˚ 17' 45" 107˚ 20' 70" C4 Cikampek 40 Sawah 06˚ 23' 138" 107˚ 26' 33"
S3 Subang 29 Sawah 06˚ 22' 79" 107˚ 35' 126"
I1 Indramayu 16 Sawah 06˚ 22' 07" 108˚ 18' 01" I2 Indramayu 16 Sawah 06˚ 22' 48" 108˚ 18' 23" I5 Indramayu 16 Sawah 06˚ 22' 63" 108˚ 18' 12" Ta1 Cianjur 261 Sawah 06° 48' 797" 107˚ 14' 161" Tc1 Sukabumi 749 Sawah 06° 54' 381" 106˚ 58' 865" Td2 Cianjur 1031 Sawah 06˚ 59' 778" 107˚ 08' 295" Td3 Cianjur 1029 Sawah 06˚ 59' 778" 107˚ 08' 292" Te3 Pangalengan 1242 Sawah 07° 09' 145" 107˚ 33' 308" Tf3 Pangalengan 1473 Sawah 07° 07' 892" 107˚ 33' 371"
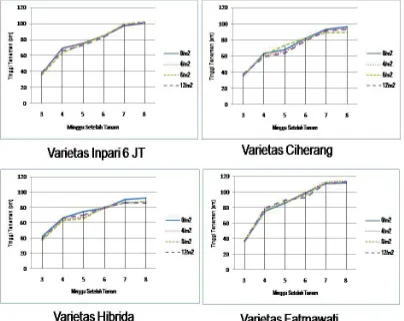
Karakter yang diamati pada habitat asal antara lain tinggi gulma, jumlah
anakan, jumlah daun, panjang dan lebar daun, serta panjang malai. Karakter yang
diamati pada kondisi rumah kaca antara lain karakter vegetatif meliput i tinggi
gulma, jumlah anakan, jumlah daun, panjang daun, lebar daun, sudut daun, dan
diameter batang. Karakter panjang daun, lebar daun, dan sudut daun diamati dari
daun ketiga dari atas yaitu daun yang pertama kali membuka sempurna. Karakter
vegetatif diamati pada 10 minggu setelah tanam.
Karakter generatif yang diamati antara lain umur berbunga, umur panen,
panjang malai, jumlah biji per malai, bobot 100 butir biji, panjang biji, diamater
biji, panjang daun bendera, lebar daun bendera, luas daun bendera. Karakter
generatif diamati pada saat panen, kecuali karakter umur berbunga yang diamati
Evaluasi Keragaman Genetik Aksesi E. crus-galli
Evaluasi keragaman genetik 16 aksesi gulma E. crus-galli asal Jawa Barat dilakukan dengan menggunakan marka molekuler Simple SequenceRepeat (SSR). Primer SSR yang digunakan untuk evaluasi keragaman genetik adalah primer yang
dikembangkan oleh Danquah et al. (2002), yaitu EC01, EC02, EC03, EC04, dan
EC05 (Tabel 9).
Tabel 9. Primer Simple Sequence Repeat (SSR) dan sekuen nukleotida yang digunakan dalam analisis genetik aksesi E. crus-galli
Lokus GenBank
Accession no. Sekuen 5’ to 3’ Repeat Motif
EC1 AY050530 F:ATTACTGGTCAGACGGAAAC (CA)6
R:GCAGTTATCTCCGTGGGCAC
EC2 AY050531 F:GGCTCCAAACAAGGCAATTC (CA)
5 R:TTCAGGGAATTTAGTACAAG
EC3 AY050532 F:GAAAGGAAATGGGTTGGCTG (CA)8(TA)3(CA)
12 R:CTTCGCACCATGATCTTCTC
EC4 AY050533 F:AGTAGAAGGCTGCAAGAAGG (GA)4AGAG(GA)
3 R:TCTCAGCCCACTTTGTATAG
EC5 AY050534 F:CAGAGCCTTCAATCATGGTG (CA)
6 R:TGCTTCAAGTTCTAGGAGAC
Bahan-bahan yang digunakan meliputi : buffer ekstraksi (10% CTAB;
0.5 M EDTA (pH 8.0); 1 M Tris-HCl (pH 8.0), 5 M NaCl; 1% β-mercaptoethanol),
buffer purifikasi/buffer CIA (Chloroform : Isoamil Alcohol = 24:1 v/v),
2-propanol, TE (1 M Tris-HCl (pH 8.0); 0.5 M EDTA (pH 8.0)), agarose,
ethidium bromida 1%, Tris-HCL (pH 9.0), MgCl2, dNTPs (1:1:1:1), 1 unit enzim
taq DNA polymerase, dan air bebas ion. Peralatan yang digunakan antara lain
tabung mikro steril (1.5 ml), vorteks, waterbath, sentrifuse, microtube steril,
vacum, bak elektroforesis, lampu UV, mesin PCR, dan Kamera Polaroid.
Isolasi DNA dilaksanakan di laboratorium Departemen Konservasi
Sumberdaya Hutan, Fakultas Kehutanan, IPB. Isolasi DNA dilakukan
berdasarkan metode Doyle & Doyle (1987) yang telah dimodifikasi oleh Sobir
(2000), metode Gawel & Jarret (1991) yang telah dimodifikasi oleh Crouch
et al. (1998a), metode Orozco-Castillo (1995) yang telah dimodifikasi oleh Toruan-Mathius dan Hutabarat (1997) dan metode Nguyen (2002) yang
crus-galli dipotong dan diambil sekitar 0.2-0.5 g, kemudian ditumbuk dalam
tabung mikro steril (1.5 ml) berisi 0.6-0.8 ml buffer ekstraksi (10% CTAB; 0.5 M
EDTA (pH 8.0); 1 M Tris-HCl (pH 8.0), 5 M NaCl; 1% β-mercaptoethanol), lalu
ditutup rapat, kemudian divorteks agar homogen. Campuran selanjutnya
diinkubasi di dalam waterbath pada suhu 65o
Pemurnian DNA dilakukan dengan penambahan 0.6-0.7 ml buffer
purifikasi/buffer CIA (Chloroform : Isoamil Alcohol = 24:1 v/v), dan pemisahan
fraksi di dalam campuran dilakukan dengan sentrifugasi 13000 rpm selama
10 menit. Fase cair (supernatan) yang diperoleh dipindahkan ke microtube steril yang baru, lalu ditambahkan 500-600 µl 2-propanol dingin, diinkubasi 4
C selama 15 menit, lalu dikocok.
o
Pengujian kuantitas dan kualitas DNA dilakukan dengan menggunakan
metode Doyle & Doyle (1987) dengan beberapa modifikasi (Sobir 2000). DNA
hasil ekstraksi sebanyak 5 µ l ditambah dengan loading dye sebanyak 1 µ l dimasukkan pada sumur agarose gel 1.2% pada bak elektroforesis selama
45 menit pada tegangan 110 volt. Hasil elektroforesis diwarnai dengan ethidium
bromida 1% dan dibilas dengan aquades, selanjutnya pita DNA hasil dilihat pada
lampu UV.
C
selama 30 menit, lalu disentrifuse 13 000 rpm selama 15 menit. Fase cair dibuang
dan fase padat/pelet dikeringkan menggunakan vacum selama 5 menit.
Selanjutnya pelet dilarutkan dalam 100 µ l TE (1 M Tris-HCl (pH 8.0); 0.5 M
EDTA (pH 8.0); Aquades).
Reaksi PCR meliputi 50 ng DNA template, primer forward dan reverse masing-masing 1.2 µM, 10 mM Tris-HCL (pH 9.0), 2.5 mM MgCl2, 0.2 mM
atau 200 µM dNTPs (1:1:1:1) gabungan dari dATP, dCTP, dGTP dan dTTP,
1 unit enzim taq DNA polymerase, dan ditambahkan air bebas ion hingga volume
total 25 µ l dimasukkan ke dalam tabung PCR dan diamplifikasi pada mesin ABI
Prism 270.
PCR reaksi diatur denaturasi selama 4 menit pada 94oC, diikuti amplifikasi
sebanyak 30 siklus yaitu 1 menit denaturasi pada 94oC, annealing selama 1 menit sesuai suhu masing-masing primer dan elongasi selama 45 detik pada 72oC dan
diakhiri 4 oC. Hasil amplifikasi dilihat dengan menggunakan gel agarose 1.4%
DNA hasil amplifikasi dicampur dengan loading dye dengan perbandingan DNA : loading dye adalah 10: 2, selanjutnya dielektroforesis (di-running) pada 50 V
selama 3 jam. Gel hasil elektroforesis direndam dalam 1% ethidium bromide
selama ± 30 menit, kemudian divisualisasikan menggunakan transimulator UV
(312 nm) dan didokumentasikan dengan Kamera Polaroid. Profil pita DNA hasil
amplifikasi pada laju elektroforesis yang sama untuk setiap tanaman diskor nilai
nol (0) jika tidak ada pita dan satu (1) jika ada pita pada posisi yang sama individu
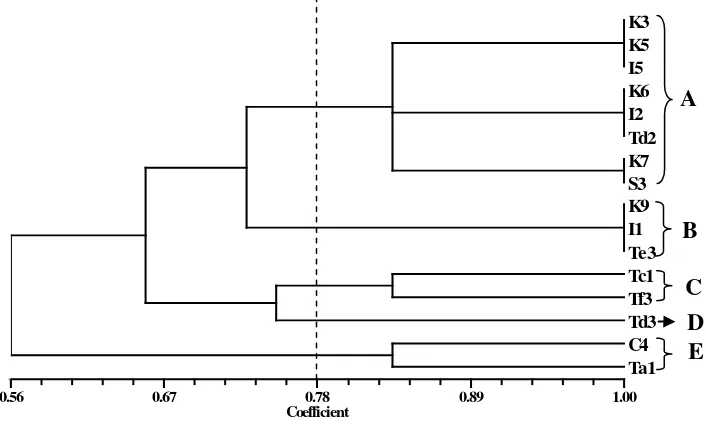
yang dibandingkan. Kesamaan genetik antar dua individu dihitung menurut
koefisien Dice (Rohlf 1998). Analisis pengelompokan dilakukan berdasarkan
kesamaan genetik dan disajikan dalam bentuk dendrogram. Data SSR dianalisis
dengan