78 Sosroseno and H e nninaj eng Med
J
Univ IndotrIts
Role
on
B
and
T
Cell
Interleukin
6
(IL-6)
:
The Biochemistry
and
Development
Wihaskoro
Sostroseno*'***,
EndangHerminajeng""
Abstrak
Aktivasi respon iuunitas neubutuhkan interoksi antara sel inunokonpeten, uolekul asesori dan sitokin. Dari sekian banyak sitokin, IL-6 adalah polipeptida dengan berat ùolekul 25 kD tlan,iletneganç peran penting pada proses inutùtas. Paper ini adalalt telaah pustaka singkat tentang peranan IL-6 pada sel B dart T. Bekerja satna dengan ienis sitokin ktinnya, IL-6 berfingsi pada tahap akhir diferensiasi sel B yang diindulcsi oleh antigen dan neningkatkan produksi antibodi. Sitokin ini tna,npu neningkatkan proliferasi sel thitttosit dan aktivasi sel T. Dibahas pula peranan lL-6 pada itnunopatogetresis AI DS.
Abstract
The inductiott of innuute response requires the interadiort atnong ittttttutroco,ilpetent cells, accessory tnolecules and cytokines.
Of
these cytokines, IL-6, a 26 kD potypeptide, plays a
najor
role on the innune re.\ponse. The roles of this cytokine on the B and T cell developntent are reviewed brieJly. In the presence of other cytokines, IL-6 act.s on antige,t dependent-tertninal dffirentiation of B cells and increases significantly the antibody productiorr. Both proliJèratiorr oJ'thynocytes and T cell activalion can be prontoted by IL-6. The roles of lL-6 on tlrc intnLuropalhogenesis of AIDS are discussedKeywords : IL-6, B cells, T cells
INTRODUCTION
The immune
system
is
an
extremely
impressive
andcomplex
network equipped
to
defend specifically
against a
wide
variety of
self
and nonself
antigens.It
is
now known that
cell to cell
contact mediated
byadhesion molecules and
costimulatory signals
provided
by
cytokines are
necessaryto
activate
theimmune
system.l-3
Of
thesecytokines, interleukin
6(IL-6),
a26Y,D polypeptide, is
apleiotropic
cytokine,
in
that
it
produces
by
a
variety of
cells
and actson a
wide
range
of
cells. The
role
of IL-6
on the immune
response and tissue damage such as acute phase
protein
has beenwell
recoqnized.a-1A
mechanism
by
which
IL-6
involves in
the
acute phaseprotein occurring
in
theliver
is beyond
the scopeof
this article.
Thus,this
article
islimited
only to
theimmunoregulatory role
of
IL-6
on B and Tcell
response aswell
as itsbiochemical
properties.Biochemistry of IL-6
Historically,
an attempt topurify
proteins
derivedfrom
the
fibroblast
culture resulted
in
apolypeptide
termedinterferon 82.8
Subsequently, there were
afew
otherproteins
such ashepatocyte
stimulating factor
(HSF),
B cell stimulatory factor-2 (BSF-2), 26
kD
protein,
hybridoma-plasmacytoma
growth
factor-2
(HPGF)
and
myeloid blood
cell differentiation-inducing
pro-tein
(MGI-2A),
isolatedfrorrr
different
sources andall
shared
amino acid
sequencehomology
atboth
cDNA
and
mature
form level;
thus,
they
aretermed
asIL-6
(seeref.
4,for
review).
The
geneencoding
IL-6
has beenidentified
andlocated
on
the short arm
chromosome
7pll-p2l
(human)
and chromosome
5
(murine).9-12 Human
cDNA of
IL-6
mRNA
from
aT
cell linel3 or
humanmonocytesl4
was cloned. ThemRNA
is translatedinto
a precursor
protein containing 212
amino acids
with
molecular weight
of 26kD.
Theremoval
of
a28amino
Vol 3, No 2, April-June 1994
13. Sawchenko PE, Swanson LW. The distribution and cells of origin of some afferent projections to the paraventricular and supraoptic nuclei of the rat (abstract). Soc Neurosci
lggl;
7:325.14. Ferguson
AV,
Day TA, Renaud Lp. Subfornical organ ef_ferents influence the excitability of neurohypophyseal and tuberoinfundibular paraventricular nucleus
in
the rat.N e ur oe ndo c ri no lo
gy
1984; 39 :423 -g.15. Swanson
LW. An
autoradiographic studyof
the efferent connections of the preoptic region in the rat. J Conp Neurol1976; 167:227-56.
16. Tanaka J, Kaba H, Saito H, Seto K. Lateral hypothalamic area stimulation excites neurons
in
the subfornical organ with efferent projections to the hypothalamic paraventricular nucleus in the rat. Brain res 1986;379'2OO-3.17. Lind RW. Bi-directional, chemically specified neural con-nections between the subfornical organ and midbrain raphe system. Brain Res 1986; 384:250-61.
18. Simpson
JB,
RouttenbergA.
Subfornical organ lesions recluce intravenous angiotensin-induced drinking. B rain
Res 1975;88:154-61.19. Eng R, Miselis RR. Polydipsia and abolotion of angiorensin_ induced drinking after transections
of
subfornical organ efferent projections in the rar. Brain Res. l98l;225:2OO-6. 20. Lind RW, Iohnson AK. Subfomical organ-median preopticconnections and drinking and pressor responses to ang.ioten_ sin-Il. J of Neuroscience 1982;2 (8); l0a3-5
l.
21. Mangiapane ML, Simpson JB. pharmacologic independence ofsubfornical organ receptors mediating drinking. Brain Res
1979; 178:507-t7.
22. PadmowijoyoD. An investigaîion
rf the
subfornical orgon as a receptorfor hista,,tine, acetylcholine or angiotensi,t_[litr ntediating drinking behaviour (AMaster thesis). Bangkok
: Mahidol University, I976.
23. Simpson JB, Routtenberg
A.
Subfomical organ: acetyl_
choline application elicits drinking . Brrtin Res 1914;69:157 -64.24.
Lind RW,
SwansonLW,
GantenD.
Angiotensin_Il_im_ munoreactive pathways in the central nervous system oftheFiber Connection ofsubfornical
Organ
77rat: evidence for a projection from the subfornical organ to the paraventricular nucleus of the hypothalarnus. Clin and Exper - Theory and Practice. 1984;A6 (10
&
ll):1915_20. 25. McKinley MJ, Denton DA and Weisinger RS. Sensors for antidiuresis and thirst-osmoreceptors of CFS sodium detec_ tors? Brain Res 1978; l4l:89-103.26. Paxinos G, Watson C. The rat brain, in steretotctxic coor_ dinates. San Diego, New York, London, Toronto : Academic
Press Harcourt Brace Jovanovich publishers, 19g6.
27.Heimet
L,
RobardsM!.
Neuroanatonical-tract TracingMethods. New York, London : plenum press,
l9gl.
28. FergusonAV,
Renaud LP. Hypothalamic paraventricularnucleus lesions decrease ptessor responses to subfornical organ stimulation. Brain Res 1984; 305:361_4.
29. Ferguson
AV,
Day TA, RenaudLp.
Subfomical organ ef-ferents influence the excitability of neurohypophyseal and tuberoinfundibular paraventricular nucleus neuronsin
theraT. Neu roendocrinology 1984 39:423-g.
30. Ferguson
AV,
Day TA, RenaudLp.
Subfornical organ stimulation excites paraventricular neurons projecting to dorsal medulla . Ant J physiol 1984; 247:Rl0gg-R1092.3I. Renaud
LP, FergusonAV,
Day TA, Bourque CW, Sgro S.Electrophysiology
of
the subfornical organ and its hypo_ thalamic connections an In-Vivo Study in the rat. Brain ResBull
1985; l5:83-6.32. Ferguson AV, Kasting NW. Electrical stimulation in subfor_ nical organ increase plasma vasopressin concentrations in the concious rat.
A,l
J
physiol 1986; 25 1:R425-R42g. 33. Sgro S, FergusonAV,
RenaudLp.
Subfornical organ_supraoptic nucleus connections: an electrophysiologic study in the rat. Brain Res 1984;303:7-13.
34. Yamashita
H,
OsakaT,
KannanH. Effect
of
electrical stimulation ofthe paraventricular nucleus on neurons in the subfomical organ of cats. Brain Res l9g4;323:176_gO. 35. Weindl A, SofroniewMV.
Neurohonnones and circum_VoI 3, No 2, April-June 1994
acid signal
sequenceyields
a secreted 184amino
acid protein.''
ThecDNA for
murine
IL-6 was
also clonedfrom
a
T
cell line
and
it
showed that murine IL-6
contains
2l
I amino
acids including
a24 residue signalpeptide.l5 Both
human and
murine
IL-6
only share
homo,logy
of
65%
at
cDNA
and
42%
at
protein
level.l5'lÔ
The amino acid
sequenceof
IL-6
is
dis-played
in Fig
1.IL-6
receptor
has also beenidentified
andcDNA
for IL-6R from
COST
cells
was cloned.lT The
IL-6R
contains
468 amino
acids
including a
19amino
acid signal peptidewith
a presumed trans membrandomain
of
28amino
acids, acytoplasmic domain
of
82amino
acids and an
extracellular
portion of
339amino
acids.The
domain
of
IL-6R
reveals a close
similarity with
that
of
immunoglobulin;
thus,IL-6R
isincluded in
theimmunoglobulin
superfamily.T
IL-6R
is
expressed ina
wide variety
ofcells
such as activated B cells,resting
T
cells, plasma
cell
line,
histiocytes
cell line
andglioblastoma cell
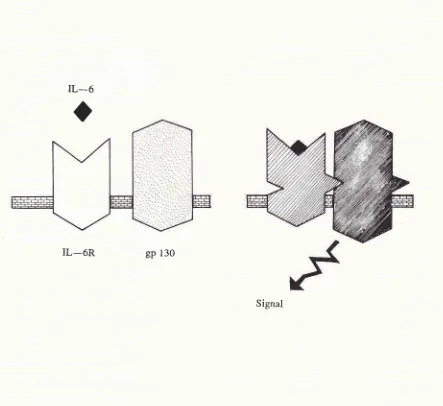
line.rEInterestingly,
thecytoplasmic
domain of
IL-6R
doesnot
act as asignal tranducer. In
this
respect, a surfaceprotein
receptor(gpl30)
closely
associated
with IL-6R is capable
of
transducing
thegrowth signal;
thus,following
interaction of
IL-6 with
its
receptor,
the
IL-6R
is
-intimate
with
gpl30
andsignal tranduction
occursle'201.."
Fig.2).
The
biological activities of
IL-6
IL-6
canbe produced
from different
sourcessuch
asfibroblast,
mononuclear cells, endothelial cells,
mac-rophages,
keratinocytes and
T-cell line,
as
well
ascertain
tumor cells (such
asT24 bladder carcinoma;.21This growing
list
of
Il-6-producing
cells suggests thatIL-6
is
secretedby both normal
andneoplastic
cells.l'
The effects
of IL-6
onB
cells
B cell differentiation into
plasmacells
can bedivided
into 3 major
steps,
viz., activation, proliferation
andIn this
respect,B cell
activation
mayy two
pathways,
i.
e., antigen_depend_independent activation.12
Briefly,
freeantigens can
be
recognized and
captureimmunoglobulin
attached
on
B
cells.
S the cellswith
their
receptors (TcRs) recogplus
MHC
neswhich
in
1
anddifferentia
f
B
cells is
mediatedby T
cell-
derived lymphokines
such as IL-4 andwill
not
befurther
discussed.It
is now known that
B
cell activation is
undercontrol of
thecytokine
network.
IL-4
andIL-5
play
aThe lwnunology of
IL-6
79major role
on B cellsactivation
andproliferation.22
On the other hand, therole of
IL-6 on
B cells
is somewhatdifferent.
In humans,antibody production
in
S. aureus CowànI-stimulated
Bcell
and a transformed Bcell line
culture
is induced
when
IL-6
is
added.l3
Additional
IL-6
at
concentration
up
to
l0
ng/ml
in
pokeweedmitogen (PWM) stimulated mononuclear
cells induceshigh
titer IgG, IgM
andIgA
antibody production.2s It
seems therefore thatIL-6
only
acts on activatedB
cellsundergoing terminal
differentiation
(seeFig.
3).It
should however be noted that the biological
functions
of
IL-6
on
B
cell
activation require the
presenceofother
cytokines. This
is based upon thefact
that
only in
the
presence
of
IL-1, TNF-
andTNF-B,
IL-6
supportsthe long term
growth of B
cells lines jn
vitro.''
Cocktail
of
IL-lB
and
TNF-cr.only
inducesIL-6
production
butfails to
stimulate antibody
produc-tion of
human bonemarrow
B
cells.28The
antibodiesof IL-lB
and
TNF-o,-stimulated
BM
cells
were secretedif
IL-6 was
added, suggestingthat
the formercytokines provoke the
secreted
IL-6
which
in
turn
stimulates
theantibody
production.28IL-6 as a
factor
for
B cell terminal
differentiation
is
also supported
by
rhefact
that
peyer's
patch (pp)
cell culture
produceshigh
rate IgAantibody
whenIL-6
is
added." In
this study, the effect
of IL-6
was much greater than thatof
IL-5.
Further
more,it also
revealed thatIL-6
did not induce
B cell proliferation,
but
it
did
increase
the number
of
IgA-secreting
B cells.
More-over,
in
theIgA sJ.slem
whereIL-5
is a potentdifferen-tiation
factor,2a'ro'JI
the
effect
of
cocktail
lL-5
andIL-6 on IgA
production
is farsuperior
than thatof
IL-5
or
IL-6 alone." In
humans,additional
IL-6
stimulatesappendix-derived
B cells to producehigh
rate of IgA.33These
results
suggest
therefore that
in
the
mucosalimmune
system,
IL-6 is
a
potent costimulator for
terminal
differentiation of IgA-committed B
cells
and actssynergically
with IL-5.
2. The
effects of
IL-6
onT cells
Thymocyte
differentiation
leading to maturation
of T
cells
haslong
been anenigma.
A
complex interaction
among
components
of
the thymic
microenviroment
directs
theselection
of T
cells
in
which
T
cells
recog-nizing the self
antigens
will
be
neglected.
Of
thesecomponents, cytokines produced
by
both thymocytes
and
other thymic cells provide
a
birectional
com_munication
in controlling
T cell selection
and matura_tion.34
Along with
other cytokines,
theability of IL-6
to
induce thymocyte proliferation has been
docu-mented.
In
the presenceof lectins,
Il-6
stimulates
the80
Sosroseno and HerninajengMNSFSTSAFG
PVAFSLGLLLERIDKQIRYI
CFQSGFNEET
CLVKIITGLLIQFLQKKAKN
LDAITTPDPTFLQSSLRALR
QMMed
J
Univ Indon50
VLPAAFPAPV
PPGEDSKDVA
APHRQPLTSS100
LDGISALRKE
TCNKSNMCES
SKEALAENNL
NLPKMAEKDG150
EFEVYLEYLQ NRFESSEEQA
RAVQMSTKVL [image:4.595.91.534.290.696.2]200
TNASLLTKLQ
AQNQWLQDMTTHLILRSFKEFigure 1. Auino acid sequence of IL-6 Nunrbers seen represent the positiorr ofthe residue. Redrawnfron ref.
I.
IL-6R
gp
130Signal
Figure 2. Signal transducrion induced by the binding of IL-6 and IL-6R.
gp ) 30 nolecule and the sigttal occurs. Redrawn
fron
ref. 7.IL_6
o
Vol 3, No 2, April-June 1994
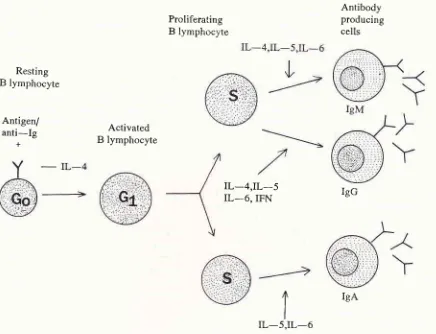
Resting
B
lymphocyte
Antigen/
anti-Ig
+
Proliferating
B
lymphocyte
The Imrnunology of IL-6
Antibody
producing
cells
8l
Activated
B
lymphocyte
IL_4,IL_5
IL-6,IFN
IgG
[image:5.595.99.535.95.429.2]IgA
Figure 3' The role of IL-6 on the antigen-rlependent B cell activatiotr. This ttrodel shows the interactiott between IL-6 and other cytokines in regulating B cell
dffirentiation.
Therefore, the ittunune network seen is not exclnsively ttte4iated by IL-6 ratherthe function of IL-6 on the target cells requires synergically other type
of
cytokines. Moclifted frontref.
26.1
L-5,IL-6
proliferation
but
it
does stimulate
L3T4*Lyt2-
andL3T4-Lyt2+-mature
thymocytes.36'37Ind"ed, it
recent-development
of
immature
thymo_diated by IL-7.38 Thus,
one
mayacts
on
the thymocyte
develop_action of
IL-7.
Therole
of
IL_6
onthe thymocytes
is further
evidenced
that
IL-1_stimu_ latedthymocyte
proliferation
requires the presenceof
IL-6;
thus,IL- I
inducesIL-6
production
and increases thesensitivity of
thymocytes
for
IL-6.3e
The
exactmechanism
of T cell activation
is
still
far from
clear.
However,
IL-2
has been
known
toimu
this cell
activation;40L-2
ell
anergy,leading
toof
".ot
Euià",r"es
haveemerged that
Il-6
may
be agood
candidatefor
T cell
activation in Il-2-independent fashion.
Ceuppens, etal., pointed out that
IL-6
actssynergically
with
pHA
to induce
stroJrgT cell proliferation without
the addi_tion of
IL-z."
Likewise,
thymocyte
proliferation
can beinduced
by
IL-6,
inIL-2
independent
fashion.43It
remains
however to be
determined
what the
physio-logical
relevance
of
these observations are,
iince
it
appers
that IL-2
and
IL-2R
may involve
in
IL_6_in_duced
T cell
proliferation.a For example,
IL-6 is
ableto
restore theproliferation of ConA
or Anti_TcR
anti_a second signal in
primary-antigen-receptor-dependent
T cell activation.
IL-6
activities
on
T
cells
activation
may therefore
bereferred
as an,,IL-2
inducer".37'4sThe exact mechanism
of T cell activation is still
82
Sosroseno and Henninajengthus' lack of
IL-2
may
induceT cell anergy,leading
tothe induction
of
T
cell
tolerance.al Euid.n"es
haveemerged that
Il-6
may
be a good candidatefor T
cell
activation
in
ll-2-independent fashion.
Ceuppens, etal., pointed out that
IL-6 acts
synergically
with PHA
to
g
T
cell
without
theaddi-tio
Likewis
proliferation
canbe
IL-6,
in
dent fashion.a3It
remains however
to be
determined
what
the
physio-logical
relevance
of
these observations are, since
it
appers
that
IL-2
and
IL-2R
may involve
in
IL-6-in-duced
T cell proliferation.a
For example,
IL-6 is
ableto
restore theproliferation of ConA
or
Anti-TcR
anti-body-srimulated
T.cells,
but
this
effect is Blocked by
antiIL-2
antibody.aaThis study
also showes thatpuri-fied
CD8-
T
cells
can beactivated
by IL-6 in
thepre-sence
of ConA
to
secreteIL-2,
suggesting thatIL-6
isa second signal in
primary-antigen-receptor-dependent
T cell activation.
IL-6
activities
on
T
cells activation
may therefore
bereferred
as an"IL-2
induce{'
.37,4sIt becomes
apparentthat
theabilities
of
IL-6
onT
cell proliferation
have somehow
beencomplicated
by
the fact that
this
cytokine
inhibits
delayed type
hypersensitivity
(DTH)
in
an animal
model.aôGiven
that
DTH
responseis mediated
by
Thl
cells
and sup-pressed b7,the
cytokines
(such
asIL-4 and
IL-10) of
Th2
cells*',
it would
appear thatIL-6
of Th2 cellsdown
regulates
the
function
of Thl
cells.
If
so,should
Th1cells
be suppressedby
IL-6, it
needsarguably
anothermechanism
other than mentioned above. One
pos-sibility
is thatIL-6
does notinduce CD4
cells inrotype
I
andtype
2cells,
it rather stimulates this
cell
subsetinto
ThOtype cells
which
secretevarious
cytokines.48Thus, no
effector T
cells
for DTH
responsewould
begenerated
if
IL-6
is
present. Whether
Il-6-mediated
DTH
unresponsiveness is orchestrated by thispathway
remains to
beinvestigated.
Perspective
The
relevancy of
IL-6
on theB
and Tcell development
is more appreciated
if
the
associatedpathological
fea-tures
are discussed.IL-6 has been
believed
to play
arole on
cardiac
myxoma, rheumatoid arthritis,
pso-riasis, lymphoma
and leukemia.THowever,
therole of
this
cytokine
on
thepatho-genesis
of
AIDS
(acquired immunodeficiency
syn-drome) is perhaps one
of
the mostintriguing
questions. Recent studies showed thatIL-6
isbelieved
to beinvolved
in
the
spreading
of
HIV
(human
immuno-deficiency virus) infection
as demonstratedby
ahigh
level of
serumIL-6 and
soluble
IL-6
receptor
and anincreased
number
of
Il-6-producing
monocyte cells
from
HIV- infected
donor..4e-52HIV
is
capable
toMed
J
Univ Indoninduce the excessive
production
of IL-6
in
human
B
cells, in the
presence
of IL-4.53
These
results
may
therefore
explain partly
the occurrenceofB
cell
hyper-activation in HlV-infected
patients.54Furthermore,
areport of Schnittman,
et
al.,
revealed that thymic
epithelial cells
produce
IL-6
which
induces the
HIV
replication in chronically
infected
HIV cells.ss
Theseresults
suggestedthat
in
AIDS
patients,
IL-6 in
the thymus maymaintain
and promote theHIV replication
of
intrathymic
precursor
T
cells. Hence, treatment
of
HlV-infected
patients
with
antiretroviral
agentsmay
only partly
suppress theHIV
replication,
but the Tcell
defect
still
occurs due to thefact
thatIL-6
prevents theintrathymic
development of T cells by
enhancingHIV
replication in
theinfected
T
cell
precursor.V/hilst the role
of
IL-6
on the previously
men-tioned
pathological
features
is
still
being
profoundly
investigated, the
useof this cytokine for clinical
pur-poses has been putforward
in
theanimal
model.Injec-tion of
recombinant
IL-6
reduces thetumor growth
in
mice, by enhancing the
activities
ofcytotoxic
T cells.56These
findings
reveal
thebroad
spectrum
of
IL-6 for
the
clinical
immunotherapy
and hence raise thepossi-bility
as
an
alternative
for IL-2 usage
in
tumor
regresion.
CONCLUSION
It becomes apparent
that
IL-6
is
oneof
the cytokines
which
serves as
an immunoregulator. This
cytokine
shows
to
act
on
the antigen-
dependentterminal
dif-ferentiation
of
B cells
and
to
enhance
the
antibody
production. Both thymocyte proliferation
and
T
cell
activation can
be
up-regulated
by IL-6.
Thus,
thepathological
features associatedwith
theseabilities of
Il-6
on
T
andB cell
Development
would
beobvious.
For
example,
increasedserum
IL-6
and
HIV
replica-tion
inthymus in
HIV-infected
patients reveal, to some degree, theinfluence
of IL-6
on the courseof AIDS.
REFERENCES
l.
Arai K, Lee F, Miyajima A, Miyatake S, Arai N, Yokota T.Cytokines: coordinators
of immune and
inflammatory responses. Ann Rev Biochem 1990;59:783-836.2. Springer
TA.
Adhesion receptorsof
the immune system. Nature 1990;346:425-34.3. Patarroyo M. l,eukocyte adhesion in host defence and tissue injury. Clin Immunol Immunopathol l99l;60:333-48. 4. Le
I,
VilcekI:
Interleukein 6:A
multifunctional cytokineVol 3, No 2, April-June 1994
5. Akira S, Hirano T, Taga T, Kishimoto T. Biology of multi-functional cytokines:
IL 6 and related molecules
(IL I
andTND. FASEB
I
199o;4:2860-7.6. Heinrich PC, Castell IV, Andus
T. Interleukin-6 and
the acute phase response. Biochem J 1990;265:621-36. 7. Hirano T, Kishimoto T. Biological and clinical aspects ofinterleukin 6. Immunol Today 1990;ll:443-9.
8. Weissenbach J, Chernajovsky Y, Zeevi M, Shulman L, et al.
Two interferon mRNAs in human fibroblast: in yitro
trans-lation and Escherichia coli cloning studies. Proc Natl Acad Sci USA l98O;77 :7 152-6.
9. Sehgal PB, Zilberstein
A, Ruggieri
M-R, may LT, et al.Human chromosome 7 carries the beta2 interferon gene. Proc natl Acad Sci USA 1986;83:5219-22.
10. Bowcock AM, Kidd JR, Lathorp M, Danashvar L. et al. the
human "beta-2 interferon/hepatocyte stimulating factor/in-terleukin-6" gene: DNA polymorphism studies and locatiza-tion to choromoseTp2l. Genomics 1988;3:8-16.
I
l.
Ferguson-Smith AC, Chen YF, Nerman MS, May LT, et al.Regional localization of the'R2-interferoniB-cell stimula-tory factor 2/hepatocyte stimulating factor"gene to human chromosome 7 p2 l-21. Genomics 1988;2 :203-8.
12. Mock BA, Nordan RP, Iustice MJ, et al. The murine IL-6 gene maps
to
the proximal regionof chromosome
5.
JImmunol 1989;142:1372- 6.
13. Hirano
T, Yasukawa
K,
HaradaH, Taga
T,
et al. Com-plementary DNA for novel human interleukin (BSF-2) rhat induces B lymphocytes to produce immunoglobulin. Nature1986;324:73-6.
14. Brakenhoff JPJ, DeGroot ER, Evers RF, Pannekoek H, Aar-den
LA.
Molecular cloning and expressionof hybridoma
growth factorin Escherichia
coli. J
Immunol 1987;139:4116-21.
15. Van Snick J, Cayphas S, Szikora J, Renauld J, et al. cDNA cloning
of murine interleukin-HPl:homology
with humaninterleukin 6, Eur J Immunol 1988;18:193-7.
16. Simpson RJ, Moritz RL, RUbira MR, Van Snick J. Murine hybridoma/plasmacytoma growth factor:complete amino acid sequence and relation to human interleukin-6. Eur J
Biochem 1988;176: 187- 97.
17. Yamasaki K, Taga T, Hirata Y, Yawata H, et al. Cloning and
expression
of
the human interleukin-6 (BSF-2/IFN R2)receptor. Science 1988 ;24 I :825-8.
18. Taga T, Kawanishi K, hardy RR, Hirano T, Kishimoto T. Receptor for B cell stimulatory faclor 2 @SF-2): quantita-tion, specificity, distribution and regulation of the
expres-sion. J Exp med 1987;166:967-81.
19. Taga T, Hibi M, Hirata Y, Yamasaki K, et al. Interleukin-6 triggers the association of its receptor with possible signal
transducer, gpl30. Cell 1989:58:573-81.
20. Saito M, Yoshida K, Hibi M, taga T, Kishimoro T. Molecular
cloning
of
a murine IL-6 receptor-associated signal
transducer, gpl30, and its regulated expressionin
vivo. JImmunol 19921,148: 4066-7
l.
21. Van Snick J, Nordan RP. Interleukin 6. In: Habenicht A, editor. Growth factors, differentiation factors and cytokines.
berlin : Sprin ger-Verlag, l99O 1 63 -7 6.
The Inmunology of
IL-6
8322. Kishimoto
T,
HiranoT. Molecular regulation
of B
iym-phocyte response. Ann Rev Ummunol 1988;6:485-512. 23, Kourilsky P, ClaverieI-M. MHC-antigen interaction: what
does T cell receptor see?. Adv Immunol 1989;45:LO7-93.
24. Haniman GR, StroberW. Theimmunobiology of interleukin
5. Year Immunol. 1989;5:160-77.
25. Muraguchi A, Hirano T, Tang B, matsuda T, et al. The essen
tial role of B cell stimulatory factor-2 (BSF-2/IL-6) for the
terminals differentiation
of
B cells.
J Exp Med
1988;167:332-44.
26. Abbas
AK. A
reassessment of the mechanisms of antigen specifis T- cell-dependent B cell activation. Immunol Today 1988;9:89-94.27. Abken H, Fluck J, Willecke K. Four cell-secreted cytokines act synergisticallly to maintain long term proliferation of
human B cell lines in vitro. J Immunol 1992;149:2685-794. 28. Roldan
E, Rodriguez
C, Navac
G, Pana
C,
BrievalA.
Cytokine network regulating terminals maturation of human bone marrow B cells capable of Spontaneous and high rateIg secretion in vitro. J Immunol 1992;149:236-71. 29. Beagley KW, Eldridge JH, Lee F, Kiyono H, et al.
Inter-leukins and IgA synthesis. HUman and murine interleukin 6
induce high rate IgA secretion in lgA-commited B cells. J
Exp Med 1989; 169:21 33-48.
30. Beagley KW, Eldridge JH, Kiyono H, Everson MP, er al.
Recombinant murine IL-5 induces high rate IgA synthesis in cycling
IgA- positive Peyer's patch
B cells. J Immunol
1988; l4l:2035-45.
31. Schoenbeck S, Mckenzie DT, Kagnoff MF. Interleukin 5 is
a differentiation
factorfor IgA
B
cells. EurJ Immunol
1989; l9:965-9.
32. Kunimoto
DY, Nordan
RP, Strober W. IL-6 is a potent cofactor of IL-I IgM
synthesis and of IL-5 in IgA synthesis.J Immunol 1989;143:223O-5.
33. Fujihashi K, McGhee JR, Lue C, Beagley KW, et al. Human appendix B cells naturally express receptors for and respond
to interleukin-6 with selective
IgAl and
IgA2 synthesis. JClin Invest 199 I ;88:248-52.
34. Ritter MA, Boyd RL. Development in the thymus:
it
takestwo to tango. Immunol Today 1993;L4:462-9.
35.Lotz M, Jirik
F, Kabouridis R, et al.B cells srimularing
factor2/interleukin 6 is a costimulant for human thymocytesand T lymphocytes. J Exp Med 1988;167:1253-8.
36. Uyttenhove C, Coulie PG, Van Snick JT. T cell growth and
differentiation induced by interleukin-HP1/IL-6, the murine hybridomaiplasmacytoma growth factor. J Exp Med 1988;
167:1417-27.
37. Quentmeier H, Klaucke J, Muhlradt PF, Drexler HG. Role of IL-6, IL-2, and IL-4 in the in vitro induction of cytotoxic T cells. J Immunol 1992;149:3316-2O.
38. Bertagnolli M, Herrmann S. IL-7 Supports the generation of cytotoxic T lymphocytes from thymocytes. J Immunol 1990;
145:17O6- 12.
39. Helle M, Boeije L, Aarden L. IL-6 is an intermediate in
IL-l-induced thymocyte proliferation.
J Immunol
1989;142:4335-8.
84
Sosroseno and Hentinajeng41. Schwartz RH. A cell culture model for T lymphocyte clonal
anergy. Science l99O:,248 1349-56.
42. Ceuppens JL, Baroja
ML,
I-orre K, Van Damme J, BilliauA, Human T cell activation with phytohemagglutinin. The
function
of IL-6
asan
accessorysignal.
J
Immunol1988;14 1:3868-74.
43. Iæ J, Fredrickson G, Reis LFL, Diamantstein T, et
al.Inter-leukin 2-dependent and interleukin 2-independent pathways
of regulation of thymocyte function by interleukin 6. Proc
natl Acad Sci USA 1988;85:8643-7.
44. Garman
RD,
JacobsKA,
Clark
SC, RauletDH.
B-cellstimulatory Ïactor 2 (R2 interferon) functions as a second
signal for interleukin 2 production by mature murine T cells.
Proc Natl Acad Sci USA 1987:7629-33.
45. Houssiau F, Van Snick J. 116 and the T-cell response. Res
Immunol 1992;143 :7 4O-3.
46. Mihara M, Ikuta M, Koishihara Y, Ohsugi Y. Interleukin 6
inhibits delayed-type hypersensitivity and the development
of adjuvant arthritis. Eur J Immunol L99l;212321-31.
47. Street
NE,
MosmanTR.
Functional diversityof
T
lym-phocytes due
to
secretionof
different cytokine pattems.FASEB J
l99l;5:17l-
8.48. Croft M, Swain SL. B cell response to helper cell subsets. II.
Both stage of T cell differentiation and the cytokines screted
determine the extent and nature of helper activity. J Immunol
199 l;r47 :3679-89 .
Med J Univ Indon
49. Birx DL, Redfield RR, Tencer K, Fowler A. et al. induction
of interleukin 6 during immunodeficiency virus infectioin.
Blood 1990;76:2303- 10.
50. Breen EC, Rezai AR, Nakajima K, Beall GN, et al. Infection
with HIV is associated with elevated IL-6 levels and
produc-tion.
I
Immunol 1990;144:480-8.51. Twigg HL,Iwamoto GK, Soloman DA. Role of cytokine in
alveolar macrophage accessory cell function in HIV-infected
individuals. J Immunol 1992;1 49 : I 462-9.
52. Honda
M,
Yamamoto S, ChengM,
YasukawaK,
et al.Human soluble
IL-6
receptor:its detection and enhancedrelease by HIV infection. J Immunol 1992;148:2175-80.
53. Boue F, Wallon C, Gourjard C, Bane-Sinoussi F, et al. HIV
induces IL-6 productiion by human B lymphocytes. J
lm-munol 1992; 148:37 6l -7.
54. Rosenberg ZF, Fauci AS. Immunopathogenic mechanisms
of
HIV
infection:cytokine inductionof
HIV.
ImmunolToday 1990;ll:176-80.
55. Schnittman S, Singer KH, Greenhouse JJ, Stanley SK, et al,
Thymic
microenviroment
inducesHIV
expression.Physiologic secretion of IL-6 by thymic epithelial cells
up-regulates virus expression
in
chronically infected cells. JImmunol 1992;147 :2553-8.
56. Mule JJ, Custer MC, Travis WD, Rosenberg SA. Cellular
mechanisms of the antitumor activity of recombinant IL06