KARAKTER MORFOLOGI DAN MOLEKULER PARASITOID
TELUR,
Telenomus
spp. (HYMENOPTERA: SCELIONIDAE)
DARI BEBERAPA DAERAH DI JAWA
OLEK
:NETT1 YULIARTI
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
NETTI YULIARTI. Karakter Morfologi dan Molekuler Parasitoid Telur, Telenomus spp. (Hymenoptera: Scelionidae) dari Beberapa Daerah di Jawa. D i b i i i g oleh PURNAMA HIDAYAT dan DAMAYANTI BUCHORT.
Studi karakter morfologi dan karakter molekuler Telenomus spp., dilakukan dengan eksplorasi parasitoid telur di beberapa lokasi di Jawa Barat, Jawa Tengah dan Jawa T i u r , dengan mengumpulkan telur Lepidoptera dan Hemiptera di pertanaman padi, tebu dan kedelai. Untuk pengamatan karakter morfologi, setiap spesies Telenomus diamati 25 pasang individu. Studi karakter molekuler dilakukan dengan teknik RAPD-PCR di Laboratorium Virologi Tumbuhan HPT IPB. Data diolah dengan program NTSYS versi 2.1. Penelitian diiulai bulan Februari -Desember 2001.
Dari penelitian ini diperoleh 9 spesies Telenomus yang tersebar dari wilayah Barat sampai ke Bagian Timur Pulau Jawa dengan persentase parasitisasi bervariasi dari 2,50-83,40%. Dari 9 spesies Telenomus, 4 spesies berhasil diidentifikasi satnpai tingkat spesies yaitu T. renlus, T. rouwzi, T. dignus dan T. dignoides. Terdapat perbedaan karakter morfologi diantara keenlpat spesies Telenomus tersebut. Perbedaan terlihat pada antena, abdomen betina, genitalia jantan, tungkai, sayap dan rumbai sayap. Hasil amplifikasi DNA dengan teknik RAPD-PCR pada keernpat spesies Telenomus, dengan satu pruner menunjukkan adanya pola pita spesifx setiap spesies. Masing-masing spesies mempunyai 3 pola pita DNA kecuali T. dignoides hanya mempunyai 2 pola pita yang jelas. Pola pita spesifik untuk T. remus berukuran 600, 850 dan 1150 pasang basa @b) sedangkan T. rou~ani mempunyai pita yang diperkiiakan berukuran 650, 850, dan 1100 pb. Adapun T. dignus dan T. dignoides ukuran pola pitanya berturut-turut adalah 550, 800, 1050 pb dan 700, 1450 pb.
Hasil amplifikasi DNA dengan teknik RAPD-PCR pada 5 populasi T. row~uni tidak memperlihatkan adanya keragaman dalam spesies. Foto profil DNA menunjukkan 3 pola pita yang diperkirakan berukuran sama yaitu 300, 430 dan 480 pb. Fenogram yang dihasilkan dari analisis pengelompokan berdasarkan I6 karakter morfologi dari 4 spesies Telenomus, membentuk 2 kelompok. Pada kelompok I
SURAT
PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudui:
Karakter Morfologi dan Molekuler Parasitoid Telur Telenomus spp.
(Hymenoptera: Scelionidae) Dari Beberapa Daerah di Jawa
Adalah benar merupakan hasil karya saya sendiri dan belum pernah
dipublikasikan. Semua sumber data dan informasi yang digunakan telah
dinyatakan secara jelas dan dapat diperiksa kebenarnnya.
Bogor, 28 Maret 2002
KARAKTER MORFOLOGI DAN MOLEKULER PARASITOID
TELUR,
Telenomus
spp. (HYMENOPTERA: SCELIONDAE)
DARI BEBERAPA DAERAH DI JAWA
NETT1
YULIARTI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Entomologi/Fitopatologi
PROGRAM PASCASARJANA
INSTITUT PERTANlAN BOGOR
Judul Tesis : Karakter Morfologi dan Molekuler Parasitoid Telur, Telenomus spp. (Hymenoptera : Scelionidae) dari Beberapa Daerah di Jawa
Nama : Netti Yuliarti
NRP : 99190
Program Studi : Entomologi/Fitopatologi
Menyetujui:
1. Komisi Penlbilnbing
Dr. Ir. Damavanti Buchori, M.Sc
Ketua A~iggota
Mengetahui:
2. Ketua Program Studi,
Penulis dilahirkan di Lubuk Lintali Padang, pada tanggal 30 Juli 1962,
sebagai anak ke-empat dari tujuh bersaudara, dari pasangan Syafei Noer
(almarhum) dan Raihanah Abbas.
Penulis menamatkan Sekolah Lanjutan Tingkat Atas pada SMA PGRI
Padang pada tahun 1981. Pada tahun yang sama, penulis melanjutkan
pendidikan pada Fakultas Matematika dan llmu Pengetahuan Alaln Universitas
Andalas. Penulis meniilih jumsan Biologi dengan spesialisasi Entoniologi dan
berhasil meraih gelar sarjaiia pada tahun 1986.
Sejak tahun 1987 sanipai 1990 penulis me11.iadi guru di SMA PGRI
Padaiig. Selanjutnya pada tahun 1990 penulis diterima sebagai staf pengajar di
Politeknik Pertanian Universitas Aidalas.
Penulis mendapatkan kesenipatan untuk melanjutkan pendidikan S2
tahun 1999 pada Program Pascasarjana Institut Pertanian Bogor dengan
Program Studi Entomologi/Fitopatologi. Pada jenjang ini penulis mendapat
bantuzn biaya dari Beasiswa Program Pascasarjana (BPPS), Direhorat Jenderal
PRAKATA
Syukur Alhamdulillali penulis ucapkan ke hadirat Allah Yang Maha
Kuasa karena atas rahmat dan karunia-Nya penulis dapat menyelesaikan
pelielitian dan penulisan tesis ini
Dalam kese~npatan ini penulis mengucapkan terima kasih kepada komisi
pembimbing: Dr. Ir. Purnarna Hidayat, M.Sc. sebagai ketua dan Dr. Ir.
Damayanti Buchori, M.Sc. sebagai anggota, atas bimbingan dan pengarahan
yang diberikan mulai penpusunan usulan penelitian, pelaksanaan penelitian
sampai penulisan tesis ini. Ucapan terima kasih juga disampaikan keljada
Ditjen. Dikti , liektor IPU, Direktur Prograin Pascasarjana IPB dan seluruh Staf
Peligajar Program St~idi Entomologi/Fitopatologi, yang telah meinberikari
kese~npatan kepada penulis u~ituk menyelesaikan studi ini.
Teriring do'a buat Ayahanda Syafei Noer (almarhum) yang tidak
sempat menyaksikan karya ini dan Ibunda Raihanah Abbas. Penulis
mengucapkan terima kasih yang setulus-tulusnya buat suami tercinta, serta
seluruh keluarga yang selalu mendo'akan penulis dalam penyelesaian studi.
Terima kasih juga penulis sampaikan kepada Ibu Aisyah, Noor Aidawati dan
Yuni, Bapak Chandra Irsan, Terri Pakki, Hasmiandi Hamid, serta teman-teman
di Laboratorium Bioekologi Parasitoid dan Predator yang teiah banyak
membantu Semoga semua bantuan yang telah diberikan mendapat balasan dari
yang Maha Kuasa.
Bogor, Pebruari 2002
DAPTAR IS1
Halaman
DAFTAR TABEL
...
viiiDAFTAR GAMBAR
...
IxDAFTAR LAMPIRAN
...
70PENDAHLTLUAN
...
1Latar Belakang
...
1. .
Tujuan Peneht~an
...
3...
TINJAUAN PUSTAKA 4
...
Taksonorni 4
...
Karakter Morfologi Untuk Identifikasi 5
Keragaman Antara Spesies dan Dalam Spesies
...
6DNA Sebagai Penanda Genetik
...
8Random An~plified Polymorphic DNA (RAl'D)
...
9...
Inang Telenomus spp 11
...
BAHAN DAN METODE 16
...
Waktu dan Tempat 16
...
Koleksi Parasitoid 16
...
Identifikasi 17
...
Karakter Morfologi 17
...
Analis'i Pengelompokan 18
...
Ekstraksi DNA
:. ...
18...
Amplifikasi DNA Dengan Teknik PCR-RAPD 19
...
...
Visualisaii DNA Dengan Elektroforesis 20
...
Penyiapan Gel agarosa 20
...
Penyiapan Sampel DNA dan Elektroforesis 21
...
Pengamatan dan Pengolahan Data 21
...
HASIL 22
...
Karakter Morfologi Telenontus spp
...
...
Telenomus remus Nixon
Telenomus rowani Gahan
...
...
Telenomus dignus Gahan
...
Telenomus dignoides Nixon
Telenomus sp
.
1...
...
.
Telenonius sp 2
...
.
Telenomus sp 3
...
Telenon7us sp . 4
Telenomus sp
.
5...
...
Kunci Identifikasi
.
....
Analis~s Pengelolnpokan
Karakter Molekuler Teler7ornus spp
...
...
Kombinasi Karakter Morfologi dan Genetika...
Karakter Molekuler T. ro~rlat7i
...
PEMBAHASAN
...
Inang dan Sebaran Telenomus spp
...
Perbedaan Karakter Morfologi Telenomus sp
...
Karakter Molekuler Telenomus spp
...
Analisis Pengelompokan
...
Karakter Molekuler T
.
rowani...
KESIMl'ULAN
...
E.WTAR PUSTAKA
DAFTAR TABEL
Haiaman
I
.
Telenomus spp.
yang pemah dilaporkan ada di Indonesia...
13 2.
Telenomus spp.
yang pemah dilaporkan di beberapa negara lain...
153
.
Daur dan waktu pemanasan yang digunakan untuk PCR.RAPD...
20....
4
.
Telenomus spp.
yang didapat dari berbagai inang dan lokasi di Jawa 225
.
Matriks data 4 spesies Telenomus berdasarkan karakter morfologi...
366
.
Karakter rnorfologi yang diamati serta kodenya...
377 . Matriks jarak 4 spesies Telenomus berdasarkan karakter morfologi
...
38. Perkiraan ukuran pita DNA spesifik spesies-spesies Telerzonms
...
429
.
Matriks data karakter nlolekuler (RAPD-PCR) Telenonnis spp...
4210
.
Matriks jarak 4 spesies Telenomus berdasarkan karakter molekuler....
4211
.
K o m b i i i karakter morfologi dan molekuler 4 spesies Telenomus....
4412
.
Matriks jarak 4 spesies Telenomus yang disusun berdasarkan karakter...
DAFTAR GAMBAR
Halaman
1 . Peta persebaran Telenomus di Pulau Jawa
...
232
.
Imago Telenomus remus...
243
.
Karakter morfologi T . remus...
254 . Imago Telenomus rowani
...
265
.
Karakter morfologi T.
rowani...
26 6.
Telenomus dignus, imago betina dan jantan...
287
.
Karakter morfologi Telenomus dignus...
298
.
Imago Telenonzus dignoides...
29...
9
.
Karakter nlorfologi T . dignoidzs 30. ...
10
.
Imago betina Teler7omu.s sp 1 31...
.
11
.
Telei7onzus sp 2, imago janlan dan imago betina 3212 . Imago betina Telenomus sp
.
3...
13
.
Imago betina Telenomus sp.
4...
14
.
Imago betina Telenomus sp.
5...
15
.
Fenogram 4 spesies Telenomus berdasarkan karakter morfologi...
16
.
Fenogram 9 spesies Telenomus berdasarkan karakter morfologi...
...
.
.
17 Pola pita DNA total Telenomus spp dengan primer P2
...
18
.
Fenogram 4 spesies Telenomus berdasarkan karakter molekuler19
.
Fenogram 4 spesies Telenomus berdasarkan karakter morfologi dan...
molekuler
...
. 20
.
Fenogram populasi Telenomus spp.
dari berbagai lokasi di Jawa...
.
DAFTAR LAMPIRAN
Halaman
Telur-telur inang Telenomus spp. a: kelompok telur Hesperidae pada padi, b: telur Nezara viridula, c: telur penggerek tebu, d: telur penggerek batang padi, e: te!ur S. litura dan f telur Spodoptera sp..
.
72Pengukuran T. dignoides asal desa Pandes Kabupaten Bantul DIY..
.
73Pengukuran T. rowani asal desa Lemah Karya Kabupaten Karawang 74
Pengukuran T. dignus asal desa Cibatu Girang Subang..
. ... . . ..
....
76 Pengukuran T. remus asal desa Cipeuyeum Kabupaten Cianjur... . . . ..
77 Pengukuran T. rowani populasi desa Cibatu Girang Subang...
... . . ...
79 Pengukuran T. rowani populasi Bantul.. ...
. . .. .
. ... . .
. . .. . .
. . . 79 Pengukuran T. ron~ani populasi desa Pondok Lakah Purwokerto...
. . . 80Pengukuran T. ron~ani populasi Sindang Barang..
. . . .
..
. . . .. . .
. .. .
..
81Pengukuran T. rowani populasi Bantar Kemang-Bogor.
.. . ..
..
.. . . . ....
82PENDAHULUAN
Latar Belakang
Telenornus spp. secara eksklusif merupakan parasitoid pada telur
serangga lain (Polaszek & I(imani i990). Inang dari Telenomus spp. kebanyakan adalah Lepidoptera dan Hemiptera, tetapi ada pula yang diketahui
muncul dari telur Diptera dan Neuroptera (Johnson 1984).
Banyak spesies dari Telenomus sangat potensial secara ekonomi karena
perannya dalam menekan populasi hama secara alami. Selain itu beberapa
spesies Telenomus dilaporkan bersifat fores; atau membonceng pada tubuh
inang sehingga menlungkillkan keberhasilannya yang tinggi dalam memparasiti
telur inangnya yang baru diletakkan seperti pada T. transversus (Bin & Johnson 1982). Di Indonesia misalnya terdapat T. rowani dan T. dignus yang berperan dalam menekan serangan penggerek batang padi (Kalshoven 1981 dan Sasmita
& Baehaki 1997). Di Afiiika terdapat T. busseolae yang menyerang penggerek tanaman serealea (Polaszek et a1 1993). Di California Selatan, Honda &
Trjapitzin (1995) justru menemukan T. hugi yang menyerang telur hama
alpokat, Sabulodes aegrotata (~uenee') (Lepidoptera : Geometridae) 2 m u s h
setelah diadakan pelepasan augmentatif Trichogramma platneri untuk
mengendalikan hama tersebut bahkan nampaknya T. hugi lebii efektii dalam
menekan hama alpokat di lapangan.
Contoh spesies yang telah sukses digunakan dalam program
Barbados d m India (Mani & Khrisnanloorthy 1986) dan T. alsophilae untuk
mengendalikan Oxydia trichiata di Columbia (Bustillo & Drooze 1977 diacu
dalam Polaszek & Kimani 1990).
Sejauh ini mash sedikit dilaporkan tentang kesuksesan Telenomus spp.
dalam pengendalian biologi klasik. Salah satu penyebabnya adalah karena
kesulitan dalam identifikasi pada tingkat spesies. Untuk lebih meningkatkan
perannya dalam program pengendalian hama terpadu, perlu dilakukan
penelitian-penelitian dasar mengenai Telenornus spp. yang dapat mendorong
pemanfaatannya sebagai agens hayati secara optimal.
Di Indonesia, penelitian Telenomus masih terbatas pada aspek biologi
atau bionomi (Agus 1991; Buchori et al. 1997) dan potensi serta parasitisasinya
sebagai agens biologi (Laba et al. 1977; Priyatna dan Baehaki 1997 dan
Kartohardjono et 01. 1999). Penelitian tentang aspek taksonomi dan ekologi sediit diiaporkan. Identifikasi yang benar dari parasitoid mempakan langkah
pertama menuju keberhasilan pengendalian hayati. Studi karakter morfologi
dapat memberikan manfaat dalam mengenali spesies. Tetapi karakter morfologi
mempunyai plastisitas yang tinggi dan sangat dipengaruhi oleh faktor
-
lingkungan (Masutti 1994). Oleh sebab itu diperlukan teknik yang l e b i sensitif
untuk mengatasi kesulitan taksonomi pada tingkat spesies.
Saat ini tersedia teknik molekuler yang merupakan salah satu cara baru
untuk mempelajari ekologi dan genetika populasi serangga. Analisis DNA
dispersal, keragaman antar spesies dan dalarn spesies, interaksi serangga-tanaman
dan spesies yang mirip secara morfologi (Hoy 1994). Teknik PCR-RAPD
(Polymerase
Chaii
Reaction- Random Amplified Polymorphic DNA) merupakansalah satu teknik analisis DNA yang dapat digunakan untuk menemukan penanda
genetik karakter spesies beberapa ordo serangga tennasuk Hymenoptera (Edwards
& Hoy 1993). Sejauh
ini
teknik tersebut belum pemah diterapkan untukmengidentifikasi spesies Telenomus yang ada di Indonesia.
Berdasarkan uraian di atas, dilakukan penelitian mengenai karakter
morfologi dan molekuler Telenomus spp. yang terdapat di berbagai habitat di
Jawa. Dengan menggunakan teknik PCR-RAPD diharapkan dapat memberikan
informasi yang bermanfaat dalam mempelajari karakter molekuler parasitoid telur,
Telenomus spp.
Tujuan Penelitian
Tujuan dari penelitian ini adalah :
1. Eksplorasi parasitoid telur Telenomus spp. pada telur hama dari ordo
Lepidoptera dan Hemiptera di berbagai daerah pertanian di pulau Jawa.
2. Menguraikan karakter morfologi dari setiap spesies Telenomus yang -
ditemukan.
3. Menemukan karakter spesifik spesies Telenomus dengan teknik PCR-RAPD.
4. Membuat fenogram dari spesies Telenomus yang ditemukan berdasarkan
TINJAUAN PUSTAKA
Taksonorni
Parasitoid telur Telenomus spp. diklasifikasikan dalam Klas : Insekta;
Ordo Hymenoptera; Sub ordo Apocrita; Superfamili : Platygasteroidea; Famili :
Scelionidae. Selanjutnya famili Scelionidae dibagi atas 3 sub famili yaitu
Telenominae, Teleasinae dan Scelioninae (Masner 1993; Csiro 1996;). Sedangkan
Kalshoven (1981) dan Borror et al. (1992) memasukkan genus Telenomus ke
dalam superfamili Proctotrupoidea dan fanlili Scelionidae.
Saat
ini
genus Telenomus mempunyai lebih dari 500 spesies yangkeseluruhannya adalah parasitoid telur. Di Akiia, 11 spesies diantaranya
diiaporkan menyerang hama dari famili Pyralidae dan 7 spesies telah dikenal
sebagai parasitoid pada telur penggerek batang serealea (Polaszek et al. 1993).
Selanjutnya Bin & Johnson (1982), diacu dalam Honda & Trjaptzin (19951 melaporkan bahwa hampir 600 spesies Telenomus yang sudah dikenal, namun
baru 10-25% yang sudah diakukan studi biologi dan sistematikanya.
Masalah taksonorni dari parasitoid telur
ini
merupakan persoaian besaryang hampir tidak terpecahkan bahkan sampai -- saat ini (Nixon 1937, Ein &
Johnson 1982 dan Polaszek & Kimani - 1990). Walaupun kekurangan informasi
dasar mengenai biosistematik, Telenomus spp. merniliki s e j d a h karakter yang
menjadikan mereka sebagai agens pengendali biologi yang efektif seperti kapasitas
5
penyebaran yang juga tinggi, kisaran ekologi yang lebar serta lebii spesifik dan
sinkron terhadap hang (Honda & Trjaptzin 1995).
Karakter Morfologi Untuk Identifikasi
Johnson (1984) rnenyatakan bahwa sebagian besar spesies Telenomus dicirikan oleh beberapa karakter sebagai berikut: antena betina 11 ruas (10 mas pada beberapa spesies T. californicus kompleks), antena jantan 12 ruas ( 11 mas pada T. pachycoris). Antena betina dengan 5 ruas gada ( 6 mas pada beberapa spesies kelornpok T. podisi, 4 ruas pada beberapa spesies kelompok T. luricis), kadang-kadang gada tidak jelas batasannya, frons licin ai bagian tengah (terukir
pada beberapa spesies dari kelompok T. Jorlol7dunus d m kelornpok T. crassiclava), rnata berambut, kepala hampir bentuk segi empat, scutellum licin, sayap depan transparan, vena marginal lebih pendek daripada stigma, vena postmarginal lebii panjang daripada stigma, sayap belakang sempit sampai lebar, tergit metasoma
pertama dengan sepasang atau lebii rambut sub lateral; tergit kedua sama atau
lebii panjang daripada lebamya; tubuh berwama coklat sampai hitam (kuning
sebagian atau keseluruhan pada T. xanthosoma, jantan kelompok T. tabanivorus, T. melanogaster dan beberapa spesies dari T. arzamae, T. floridanus dan kelompok T. crassiclava); mesosoma bin metalik atau hijau pada beberapa spesies.
identifikasi sehingga sering menyebabkan kesalahan dalam identifikasi spesies.
Hal ini pemah terjadi pada Telenomus yang menyerang Pyralidae di AEika, dari
12 spesies yang telah diiaporkan, hanya 1 yang didasarkan pada identifikasi
yang benar (Polaszek & i(imani 1990).
Pada famili Scelionidae beberapa karakter dapat digunakan untuk
identifikasi genus sampai spesies. Alba (1989) menggunakan beberapa karakter
untuk identifikasi spesies Telenomus yaitu antara lain sayap, torak, abdomen,
tergit, tibia dan warna. Sedangkan Polaszek et al. (1993) menggunakan karakter
genitalia jantan, antma jantan, tarsus, tibia, femur dan warna tubuh. Barrion &
Litsinger (1989) menambahkan beberapa karakter seperti rumbai sayap, vertex
dan preoccipital carina.
Keragaman antar spesies dan dalam spesies
Nixon (1937) yang telah banyak mengidentifikasi Telenomus
mengemukakan bahwa kesulitan dalam mendeskripsi dan mengklasifikasikan
sejumlah besar anggota genus ini mash belum bisa diatasi karena sulitnya
menangkap perbedaan diantara mereka. Satu spesies sering diciriian oleh
karakter yang bercampur baur dengan karakter yang juga d ioleh semua spesies sehingga sulit diterima oleh mata yang tidak biasa dalam menghadapi
perubahan spesifik yang terjadi pada genus ini. Dijelaskan bahwa genitalia
jantan merupakan karakter yang tidak terhingga nilainya dalam identifikasi.
membentuk kelompok spesies yang kompak sangat sulit dibedakan dengan
metode biasa.
Bin & Johnson (1982) menemukan 4 spesies baru Telenomus dari telur
Pyralidae di daerah tropik. Selain itu ada satu spesies yang dideskripsi ulang
yakni T. alecto. Identifikasi ini diiakukan selain dengan menggunakan karakter
genitalia jantan, juga menerapkan pengukuran morfometrik dan karakter nunus
sensila pada ruas-ruas antena Empat spesies baru tersebut adalah:
I:
applanatus Bin & John., T. versicolor Bin & John., T. transversus Bin & John.
dan T. globosus Bin & John
Pengetahuan mengenai perbedaan spesies dalam genus Teler7onzus sering
didasarkan pada adanya perbedaan yang sangat sedikit pada morfologinya
(Johnson et al. 1987). Studi morfologi yang dilakukan pada spesies kompleks
T.
busseolae Gah. dari Afiika telah berhasil memisahkan spesies kompleks inimenjadi 4 spesies yang berbeda. Dua dari empat Tvlenornus ini merupakan
spesies simpatrik yang berhubungan melalui distribusi, hang dan periode
oviposisi. Variasi yang ditemukan pada spesies-spesies yang menyerang
Pyralidae dan Noctuidae ini terdapat pada kepala, mesosoma dan genitalia
jantan (Polaszek et al. 1993).
Keragaman dalam spesies pada Hymenoptera parasit sangat
berhubungan dengan perbedaan inang seperti yang diingatkan oleh Salt (1941)
juga melaporkan adanya variasi pada Telenomus sp. yang dipelihara pada telur
Spodoptera litura (F.) dan S. exigua (Hbn) ( Lepidoptera: Noctuidae). Variasi terjadi pada lebar kepala, lama hidup dan keperidian.
Fenomena ini juga ierjadi pada T. upplanatus Bin & John. yang dipelihara pada dua macam penggerek tebu yaitu Eldana dan Maliarpha.
Perbedaan terdapat pada struktur genitalia jantan dan sensila pada antenomer
jantan. Selanjutnya Johnson et al. (1987) juga melaporkan adanya variasi pada
T. alsophilae Vier. yang dipelihara pada 5 spesies hang yaitu Caenurgina crassiuscula Haworth (Lepidoptera: Noctuidae); Campaea perlata Guene'e,
Tetracis cachexiata G ~ ~ e n e e, Plugodis seriiraria Herich-SchaeRer, Pergu~nrr
honestaria Walker (Lepidoptera: Geometridae). Variasi tersebut mencakup
lebar kepala serta panjang dan lebar antenomer.
DNA Sebagai Penanda Genetik
Perkiiaan keragaman antara dan dalam spesies perlu untuk optimisasi
dalam strategi pemeliharaan parasitoid guna meningkatkan efisiensinya dalam
program pengendalian biologi (Masutti 1994). Teknik molekuler yang
menggunakan analisis DNA dapat mendeteksi keragaman antara spesies d m
dalam spesies serangga parasitoid. Penggunaan analisis DNA juga dapat
membedakan dua populasi parasitoid yang mirip secara morfologi (spesies
kriptik), spesies yang berhubungan dekat, biotipe dan klon pada spesies yang
DNA yang telah banyak digunakan dalam studi sistematika adalah DNA
inti, DNA mitokondria, DNA ribosom dan DNA satelit. DNA mitokondria
biasanya digunakan untuk mempelajari struktur populasi, alian gen, hibridisasi,
biogeografi dan Nogenetik. Sekuen DNA satelit biasanya menggambarlcan
spesifik spesies, sehingga dapat digunakan untuk diagnosis spesies (Hoy 1994).
Random Amplified Polymorphic DNA (RAPD)
Polymerase Chain Reaction merupakan metode in vitro untuk
n~engamplifikasi sejumlah kecil DNA atau RNA. Beberapa macanl ballan yang
diperlukan antara lain DNA polynlerase, dNTPs, n~olekul DNA fenz17late dan
dua oligonukleotida sebagai primer. Reaksi ampliiiasi sangat bergantung pada
keberadaan enzim polymerase sebagai katalisator, terutama yang tahan panas.
Enzim yang paling terkenal dan banyak digunakan adalah tag DNA polyrneruse
(taq polymerase) yang diisolasi dari bakteri tahan panas Themus aquaticus.
Deoksinukleotida tripospat (dNTPS) terdiri atas 4 nukleotida yaitu
deoksiadenosin tripospat (dATP), deoksiguanidiin tripospat (dGTP),
deoksisitid'in tripospat (dCTP) dan deoksitimidii tripospat (dTTP) (Kessing et
al. 1989).
Proses PCR terdiri atas beberapa tahap reaksi dengan perlakuan panas
yang berbeda-beda secara berulang-ulang dalam beberapa siklus tertentu yaitu
denaturasi, anealisasi dan sintesis DNA. Dengan pengulangan reaksi secara
simultan tersebut, maka junllah DNA sasaran yang diamplifiiasi menjadi
PCR-RAPD adalah teknik PCR yang menggunakan satu primer acak
guna menghasilkan ampliiasi DNA. Hasil pola pita DNA dengan teknik PCR-
RAPD menyediakan informasi tentang variasi genetik dalam keseluruhan
genom serangga (Dowdy & McGaughey 1996).
Tigkat variasi genetik (DNA) dapat dideteksi dengan menggunakan
teknik RAPD. Dalam analisis DNA pada Hymenoptera, teknik ini lebh baik
daripada teknik allozyme yang mendeteksi tingkat protein. RAPD mendeteksi
keragaman berdasarkan pada amplifikasi di daerah-daerah yang bervariasi pada
suatu genom dengan menggunakan satu primer acak dan tidak memerlukan
pengetahuan tentang sekuen DNA (Edward & Hoy 1993).
Keuntungan menggunakan teknik PCR-RAPD antara lain adalah
prosesnya yang berlangsung cepat karena mulai dari awal prosedur sampai
visualisasi DNA yang diamplitikasi pada suatu gel dapat ditentukan kia-kira 24
jam (Kambhampati et al. 1992); relatif murah dibandiigkan dengan sekuensing;
dapat digunakan untuk berbagai studi antara lain analisis filogenetik, hubungan
kekerabatan, mengidentifikasi spesies yang mirip secara morfologi,
membedakan strain serta mengidentsasi asal geografi beberapa serangga yang
dintroduksi (Cenis et al. 1993; Heimpel et ul. 1997; Dowdy & Mc.Maughey 1996). Faktor-faktor yang perlu diperhatikan dalam PCR-RAPD yaitu
konsentrasi DNA contoh, panjang primer, komposisi basa primer, konsentrasi
secara hati-hati agar bisa diperoleh pita-pita DNA yang utuh dan baik (&rani et
al. 1998).
Hoy (1994) menambahkan bahwa analisis variabilitas genetik melalui
amplifikasi DNA dengan primer acak menghasilkan data yang tidak kodorninan,
tetapi karena kemudahan dan kecepatan menghasilkan data, teknik ini telah
banyak digunakan untuk mengidentifkasi spesies-spesies yang sulit dibedakan
secara morfologi.
Inang Telenon~us spp.
Kebanyakan inang Telenomus spp. merupakan hama penting pertanian,
kehutanan dan peternakan. %saran hang dari genus
ini
cukup lebar terutarnadari ordo Lepidoptera dan Hemiptera. Pernah diiaporkan bahwa parasitoid
genus ini juga ditemukan pada Diptera. Spesies tertentu dari genus ini
inangnya ada yang terbatas pada genus tertentu tetapi ada pula yang dapat
hidup pada beberapa famili. Misalnya
T.
clisiocampae Riley merupakanTelenomus yang hanya berinang pada genus Malacosoma ( Lepidoptera:
Lasiocampidae) sedangkan T. alsophilae berhasil dipelihara pada 17 genus
dari famili Geometridae dan Noctuidae (Johnson 1984).
Polaszek dan Kimani (1990) menambahkan bahwa walaupun kisaran
inang dari Telenomus spp. cukup lebar, nmnun spesilisitas inang bervariasi antar
spesies, beberapa spesies dapat menyerang telur dari beberapa genus dalam satu
ordo dan yang lain kelihata sangat terbatas pada satu spesies saja. Ada
laricis adalah semua spesies yang memarasit telur Miridae dan kelompok
spesiesfloridanus khusus memarasit telur Lygaeidae.
Telenomus spp. merupakan parasitoid yang telah terbukti sebagai agens
pengendali biologis yang penting. Beberapa proyek telah diiakukan untuk
pengendalian biologi populasi Pyralidae dengan menggunakan parasitoid telur
genus Telenomus dengan hasil yang menjanjikan. Contohnya adalah di Ivory
Coast untuk mengendalikan Eldana saccharina Wk., Maliarpha separatella
Rag. dan Scirpophaga melanoclysta Mey. pada tebu, padi dan jagung. Di India
digunakan untuk melawan serangan Chilo sacchariphagus (Boyer) pada tebu. di
Bolivia untuk mengendalikan serangan D[ulr.uea rufescens Box. dan D.
saccharalis (F.) juga pada tebu (Bin & Johnson 1982).
Telenomus remus Nix. telah digunakan untuk program pengendalian
hayati hama Spodoptera spp. di Barbados dan telah dicapai hasil yang baik
Parasitoid yang sama juga telah diintroduksi ke India dan diiepas pada
pertanaman tembakau, kubis dan kapas (Mani & Khrisnamoorthy 1986).
Beberapa spesies Telenomus yang berinang pada Lepidoptera famili
Pyralidae antara lain adalah T. rowani Gah., T. dignus Gah., T. dignoides Nix.
yaitu menyerang Scirpophaga spp. (Kalshoven 1981). Sasmita dan Baehaki
(1997) melaporkan dua spesies Telenomus pada penggerek padi putih di
Kalshoven (1981) menyatakan bahwa pada Chilo spp. dietahui satu
betina Telenomus spp. dapat menyerang 30-50 telur sedangkan pada kelompok
telur penggerek padi putih yang biasanya berjumlah sekitar 200 telur, 50-75%
diantaranya terparasit dan parasitoid tersebut diduga T. rowani atau T.
dignoides.
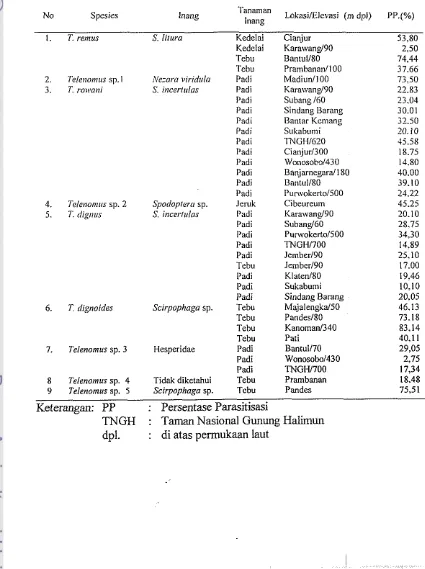
[image:134.608.90.540.308.798.2]Pada Tabel 1 berikut dapat dilihat beberapa spesies Telenomus serta inangnya yang pernah dilaporkan ada di Indonesia.
Tabel 1. Telenomus spp. yang pemah dilaporkan ada di Indonesia
No Spesies Inang Pustaka Lokasi
1 Telenonius Scirpophaga ii?iiotata (Wlk.) Nixon ( 1937) Jawa
diznzrs . S. iiicerfztlas (Wlk.) &Chilo Kalshoven ( 1 98 1) SPP.
2 T. rowani S. innotata dan
S. incerlulas
3 T. dignoides S. aurijlua, Scirpophaga spp.
4 T. luculIus Nocruidae
5 T. bene$ciens Diatraea slrialalis Sn,
6 T. transversus Tvporiza irrnotata
Nixon (1937), Jawa Klashoven (1981)
Nixon (1 937), Cirebon Kalshoven (1981)
Nixon (1937) Jawa
Nixon (1937), Cirebon Klashoven (1 981)
Bin &Johnson Sulawesi (1982)
7 T. spodopterae Spodoplera litura (F.) ~ a l s h b v e n (198 1) 8 T. javae Hidari irava (Mr.)
(Lepidoptera: Hesperidae) 9 T. tirathabae Tirathaba sp. (Lepidoptera :
Wlk. Pyralidae)
I0 T. c e s Nezara viridula L. (Hemiptera: Pentatomidae)
11 T. remus S. litura
12 Telenomus sp. A Piezoderus hybneri (Gmelin) (Hemiptera: Pentatomidae) 13 Telenomus sp. B Nezara viridula
Kalshoven (1981) Jawa
Nixon (1937), Jawa Kalshoven (198 1)
Nixon (1937) Lembang
Shephard & Barrion
(1998) Indonesia Shepard & Barrion Indonesia (1998)
Spesies Telenomus yang diketahui menyerang ordo Lepidoptera famili
Noctuidae adalah T remus Nix., T. Spodopterae Dodd., T. nawai Ash. (Polaszek 2001) dan T. alsophilae Ash. (Fedde 1977). Honda & Trjapitzin (1995) melaporkan spesies baru T. hugi Hond. & Trjap. yang menyerang ulat daun alpokat Sabulodes aegrotata (Guenee') (Lepidoptera: Geometridae) di
Kalifornia Selatan. Sedangkan T. attaci Nix. merupakan spesies yang
menyerang telur kupu- kupu gajah, Attacus atlas L. (Lepidoptera: Saturniidae)
di Kuala Lumpur dan Bangkok (Nixon 1937).
Pada ordo Diptera diietahui ada 3 famili yang menjadi inang Telenornus yaitu Tabanidae, Asilidae dan Stratiomyidae. Spesies Telenomus tersebut
adalah T. tabanivorus Ash. dan T. en~ersoifii Gir. (Johnson 1984). Sedangkan T.
cyrus Nix. dan T. triptus Nix. adalah dua spesies Telenomus yang dilaporkan
menyerang telur kepik (Hemiptera: Pentatomidae) di Jawa dan Malaysia (Nixon
Pada tabel berikut ini dapat dilihat Telenomus spp. yang pernah
dilaporkan di beberapa negara lain
Tabel 2. Parasitoid telur, Telenomus spp. yang pernah diaporkan di beberapa negara lain
No Spesies Inang I Tanaman Daerah
sebaran Pustaka Inang
1 T. nawai Spodoptera spp. Jepang Arakaki, Noda &
Yamagishi (2000)
2 1: isis Polasz. Sesamia spp. IJagung Afiika Polaszek et al. (1993)
3 T. bussealae Gah
4 T. nagarajae
Polasz.
5 T. sesamiae Wu &
Chen.
6 T. triptus Nix.
7 T. padisi Ash.
8 T. alsophilae Ash.
9 T. heliothidis Ash.
10 T. sphingis Ash.
11 T fariai
12 T. manolus Nix.
Bussealafisca
(Lepidoptera: Noctuidae)l Jagung
Sesamia spp. lPadi &
tebu
Sesamia sp. IJagung
Scotinophara caarctata
Padisus maculiventris
Kedelai
Alsophilae IPinus &
Eucalyptus Heliothis virescensl Tomat Ifeliathis virescens Triatoma phyllosoma Hemiptera:Reduviidae
Afiika, Polaszek et al. (1993) Mesir, Iran,
Irak , Israel
Papua New Polaszek el al. (1993) Guinea,
Bangladesh
China Polaszek et al. (1993)
Malaysia Nixon (1937)
Amerika Barrion & Litsinger (1989), Okuda &
Yeargan (1988)
Amerika Bustillo & Drooz (l977), Fedde (1977)
Amerika Strand & Vinson (1983)
Amerika Farrar et al. (1994)
Venezuela Rabinovich, Jorda &
Bemstein (2000)
[image:136.602.107.512.160.557.2]BAHAN DAN METODE
Waktu dan Tempat
Penelitian ini diiakukan mulai bulan Februari 2001 sampai Desember
2001. Parasitoid dikoleksi dari beberapa peitanaman padi, tebu dan kedelai di
beberapa lokasi di Pulau Jawa. Lokasi pengambilan sampel parasitoid di Jawa
Barat antara lain adalah Bogor, Subang, Karawang, Sukaburni, Cianjur,
Majalengka, Taman Nasional Gunung Halirnun dan di Jawa Tengah adalah
Pwwokerto, Wonosobo, Banjarnegara, Pati, di Daerah Istimewa Yogyakarta
adalah Klaten, Bantul, Prambanan sedangkan untuk wilayah Jawa Timur yaitu
Madiun dan Jernber.
Kajian karakter morfologi dilakukan di Laboratorium Taksonorni
Serangga Jurusan Hama dan Penyakit Tumbuhan Institut Pertanian Bogor.
Sedangkan penelitian keragaman karakter molekuler dilakukan di Laboratorium
Virologi Jwusan Hama dan Penyakit Tumbuhan IPB.
Koleksi Parasitoid
Parasitoid dikoleksi dari areal pertanaman padi, tebu dan kedelai dengan
cara mengumpulkan kelompok telur Lepidoptera dan Heniptera yang diiketahui
rnerupakan inang Telenomus. Telur-telur tersebut d i i s u k k a n ke dalam t a b h g
kaca yang bagian daIamnya sudah diolesi madu dan diberi label menurut
tanaman inang dan lokasinya. Telur-telur yang sudah berada dalam tabung
parasitoid muncul. J d a h parasitoid yang muncul dan j d a h dari telur hang
d i t u n g untuk mengetahui persentase parasitisasinya. Rumus yang digunakan
untuk menghitung persentase parasitisasi adalah :
Persentase parasitisasi (%) = (C telw terparasit / C telur seluruhnya) x 100%
Identifikasi
Untuk mengetahui spesies Telenomus, dilakukan identifikasi dengan
menggunakan beberapa karakter morfologi yang dapat menusahkan spesics,
misal sayap, antena, metasorna, tibia, tarsus dan genitalia jantan. Kunci
identifikasi yang digunakan adalah Nixon (1937), Nishida & Torii (1970), Alba
(1989) dan Barrion & Litsinger (1989).
Karakter Morfologi
Untuk mempelajari karakter morfologi Telenornus spp. diperlukan
preparat awetan basah dalam alkohol 70%, dan slide mikroskop. Untuk
pengamatan karakter rnorfologi diperlukan 25 pasang imago Telenomus dari
setiap spesies. Sernua individu diukur panjang total dan ukuan bazian-bagian
tubuhnya. Diseksi dilakukan untuk rnengamati karakter genitalia jantan.
Analisis Pengelompokan
Semua karakter spesifik dari setiap spesies d i i p u k a n dalam suatu
tabel. Dalam ha1 ini karakter atau ciri dapat dibedakan berdasarkan tipenya
yaitu ciri kualitatif dan ciri kcantitatit Ciri kuantitatif adalah ciri yang dapat
diitung atau diukur pada interval tertentu, misal panjang atau lebar metasoma.
Sedangkan ciri kualitatif misalnya ada atau tidak adanya penebalan pada
genitalia jantan. Ciri-ciri ini dmyatakan dengan kode tertentu misal jika ada
penebalan pada genitalia jantan diberi kode 1 sedangkan jika tidak ada
penebalan diberi kode 0.
Setelah pemilihan ciri dan dan hal-ha1 yang berkaitan dengan ciri
ditentukan, tahapan selanjutnya adalah penyusunan sifat ciri dari sejumlah n ciri
dan sejumlah p unit takson atau populasi dalam suatu matriks n x p. Matriks
data ini selanjutnya diolah dengan program NTSYS. versi 2.1 sehingga
diperoleh pengelompokan spesies-spesies Telenomus.
Ekstraksi DNA
Ekstraksi DNA Telenomus spp. dilakukan dengan metode ekstraksi
DNA total yaitu yang dideskripsikan oleh Goodwin et al. (1994). Imago
Telenomus spp. diambil 1-3 ekor dari setiap spesies atau popdasi, kemudian
d i i u k k a n ke dalam tabung mikro yang berisi 10 p1 buffer CTAB 2% d m
ditambahkan 0,2 gram pecahan bubuk kaca clan digerus dengan mikropistil
dalam kondisi dingin Ke dalam tabung yang sama dimasukkan lagi 115 pl
pada suhu 65 OC selama 5 menit. Kemudian ditambahkan 125 p1 cNorofom
isoamilalkohol (24 : 1 vlv) dan dibiarkan pada suhu ruang selama 20 menit. Lalu disentrihgasi pada kecepatan 800 rpm selama 5 menit. Hasil supernatan
yang akan digunakan sebanyak 90 p1 dipindahkan ke tabung yang lain. Ke
dalam tabung tersebut lalu ditambahkan 10 p1 NaOAc 3 M (pH 5,2) dan 250 p1
etanol absolut (-20 "C). Kemudian campuran ini disimpan dalarn fi.eezer selama
30 menit. Selanjutnya disentrihgasi pada kecepatan 11500 rpln selama 15
menit, supernatan yang terbentuk dibuang. Hasil pellet dicuci dengan 200 p1
alkohol 70% (-20 OC) dan disentrifugasi dengan kecepatan 11500 rpm selama 2
menit.. Kemudian etanol dibuang, pellet dikeringanginkan dan diresuspensi
dengan 30 p1 akuades steril.
Amplifikasi DNA dengan Teknik PCR-RAPD
Hasil ekstraksi DNA diamplifikasi dengan teknik RAPD-PCR dengan
menggunakan beads dan primer acak dari Amersham Pharmacia Biotech
(Ready to go RAPD analysis beads). Beads ini biia dilarutkan dalam volume 25
pl campuran maka akan mengandung thermostable polymerases (amplitaq TM
DNA polimerase dan Stoffel fragment), 0,4 m M tiap dNTP, BSA (2,5 pg) dan buffer (3 mM MgC12,30 mM KC1 dan 10 mM Tris, (pH 8,3).
Primer yang digunakan adalah primer P2 (5'-~[GTTTCGCTCCI-3') dari
Amersham Pharmacia Biotech. Volume total campuran berbagai komponen
5 p1 primer P2 dengan konsentrasi 25 pmol dan 5 p1 DNA hasil ekstraksi.
Pencampuran ini diiakukan dalam kondisi dingin. Kemudian disentrifUgasi
dalam mikrosentrifus selama lebih kurang 1 menit agar campuran bahan
mengumpul di bagian bawah tabung.
Tabung-tabung ditempatkan pada mesin PCR (thermal cycler) pada suhu
95 OC selama 5 menit untuk pemanasan awal. Selanjutnya mesin PCR diatur
melalui program untuk melakukan 45 daur suhu pemanasan beserta waktunya
yang telah ditentukan seperti tertera pada Tabel 3. Jika siklusnya telah selesai maka hasil PCR disin-,pm dalam freezer sebelum dipergunakan lebii lanjut.
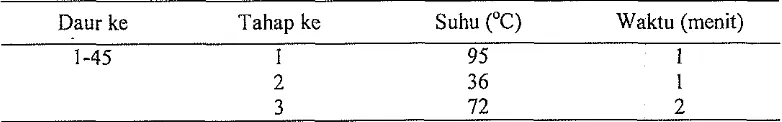
Tabel 3. Daur dan waktu pemanasan yang digunakan untuk PCR-RAPD.
Daur ke Tahap ke Suhu ("C) Waktu (menit)
3 72 2
Akhir daur dipertahankan pada suhu 4°C sampai tabung diambii
Visualisasi DNA dengan Elektroforesis
DNA hasil amplitikasi PCR-RAPD dieiektroforesis dengan gel agarosa
1,5% pada buffer TBE dengan alat elektroforesis horizontal. Untuk mengukur
berat molekul DNA digunakan marker 100 pasang basa @b).
Penyiapan Gel Agarosa
Agarosa 1,5 g dan 100 ml buffer TBE konsentrasi l x dimasukkan ke
[image:141.602.116.507.369.430.2]Selanjutnya agarosa dibiarkan diigin dan keras, kemudian sisiran dilepaskan
sehingga akan terbentuk sumuran (~jells). Buffer TBE i x ditambahkan ke
dalam baki sampai gel tergenang.
Penyiapan Sampel DNA dan Elektroforesis
Sampel-sampel DNA sebanyak 5 p1 dicampur dengan 2 p1 buffer
pemberat. Masing-masing sampel tersebut diisikan ke dalam sumuran pada gel
dengan menggunakan pipet mikro. Gel dielektroforesis pada tegangan 100 volt
selama lebii kurang 2,5 jam. Gel selanjutnya diirendam dalam Etidium Bromide
(EtBr) sekitar 3 menit kemudian dicuci dengan air sebanyak 2 kali. Hasil
elektroforesis tersebut lalu dilihat dengan menggunakan transluminator ultra
violet dan dipotret dengan kamera Polaroid.
Pengamatan dan Pengolahan Data
Pita polimorfik masing-masing DNA sampel diamati untuk menentukan
adanya perbedaan spesies. Pola pita yang jelas ada diberi kode 1 dan jika tidak
Inang dan Penyebaran Telenomus
Dari hasil eksplorasi Telenomus, diperoleh 9 spesies Telenomus dari
[image:143.602.90.515.210.779.2]berbagai lokasi di P. Jawa seperti terlihat pada Tabel 4 berikut:
Tabel 4. Telenomus spp. yang didapat dari berbagai inang dan lokasi di Jawa
Spesies lnang Tanaman
lnang LokasiIElevasi (m dpl)
1. T. remus S. litura Kedelai Cianjur Kedelai Karawand90 Tebu
Tebu 2. Teleno!nus sp. l r\'ezara i,irid!~la Padi 3. T ro~cani S incerlalas Padi I'adi I'adi I'adi Padi Padi I'adi Padi Padi Padi Padi 4. l'elei~o~~!rrs sp. 2 Spodopoprera sp. Jeruk 5. 7: digntts S bicerlulas I'adi
Padi Padi Padi Padi Tebu Padi Padi Padi 6. T dignoides Scirpophaga sp. Tebu
Tebu Tebu
~ a n t u l l 8 < Pramhananl100 Madiud100 Karawang!90 Subang I60 Sindang Barang Uaniar Kcmanz Cianjurl300 Wonosobd430 Banjarnegarafl80 Bantu1180 Purwokerto/SOO Cibeureum Karawang!90 Suband60 Jemberl90 Jemberl90 KlatenISO Sukabu~ni Sindang Barang MajalengkalSO Pandes180 Kanoman/340
Tebu Pati 40.1 1
7. Telenomus sp. 3 Hesperidae Padi Bantu1170 29,05
Padi Wonosobo1430 2.75
Padi TNGW700 17,34
8 Telenomus so. 4 Tidak diketahui Tebu Prambanan 18.48 9 Telenomus sp. 5 Scirpophago ~~ - sp. ~ Tcbu Pandes 75.51
Keterangan: PP : Persentase Parasitisasi
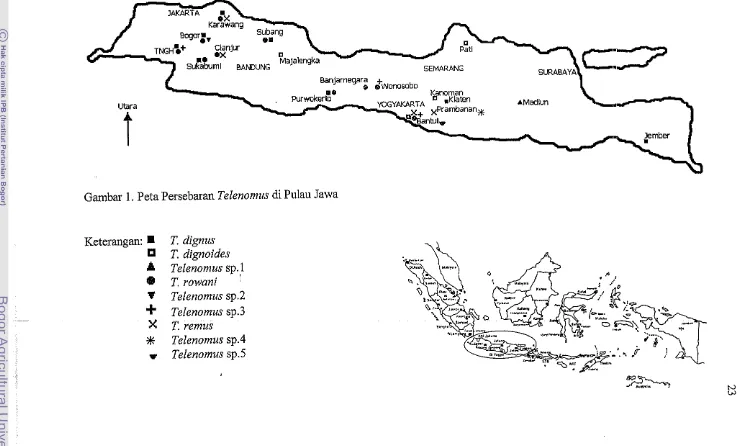
Gambar 1. Peta Persebaran Telenomus di Pulau Jawa Keterangan:
A
9 V-I-
X*
T. dignus T. dignoides Telenomus sp.1 T rowani I Telenon~us sp.2 Telenomus sp.3T.
remusKarakter Morfologi Telenomus spp.
Dari hasil identifikasi yang telah dilakukan, maka didapat 4 spesies
Telenomus yaitu T. remus dari inang S. litura, T. rowani dan T. dignus dengan inangnya penggerek Satang padi (Scirpophaga spp.) dan T. dignoides dari hang penggerek pucuk tebu. Berikut ini diuraikan deskripsi karakter morfologi
masing-masing spesies.
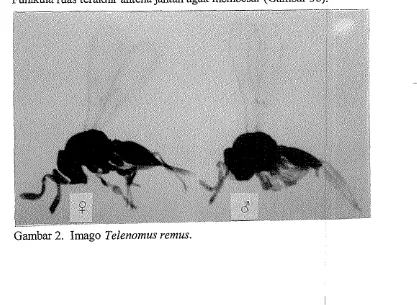
1. Telenomus remus Nixon
Merupakan spesies yang kecil dengan ukuran tubuh berkisar 0,44 - 0,60
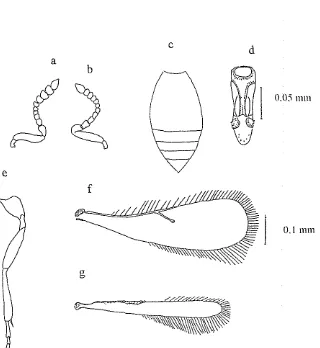
mm. Torak menonjol atau lebih tinggi daripada abdomen (Gambar 2). Antena
betina dengan funikula 1-4 (Fl-4 ) sangat kecil, jauh lebih kecil daripada pedisel, funikula 6-8 hampir sama ukurannya. Genitalia jantan pendek dan agak
lebar dengan median plate yang cekung dekat digiti (Gambar 3c). Tungkai dan antena jantan benvarna leb'i pucat dibandimg tungkai dan antena betina.
Funikula ruas terakhir antena jantan agak membesar (Gambar 3b).
[image:145.602.111.528.471.776.2]Gambar 3. Karakter morfologi T. remus; a : antena betina b: antenna jantan; c: abdomen betina d: genitalia jantan; e: tungkai belakang jantan; f:
sayap depan jantan g: sayap belakang jantan (skala sama kecuali genitalia jantan).
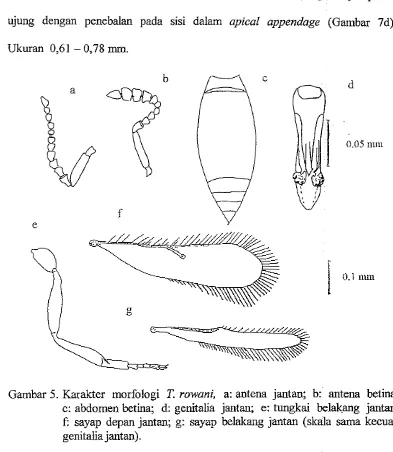
2. Telenomus rowani Gahan
Separuh bagian bawah abdomw meruncing dengan rata ke ujung.
Betina berwama hitam sedangkan jantan benvama coklat kenlerahan dengan
mata dan ujung abdomen benvama lebiih gelap (Gambar 4). Tubuh betina
biasanya berukuran lebii besar daripada yang jantan dengan kisaran bertumt-
turut adalah 0,75-0,93 mm dan 0,56-0,75 mm. Gada pada antena betina
atau sediiit lebih panjang dari lebarnya (Gambar 5b). Funikula 1-3 antena
jantan lebih besar daripada ruas-ruas berikutnya (Gambar Sa). Metatarsus dari
semua tungkai jantan melebar (Gambar 5 e). Genitalia jantan dengan penis
[image:147.608.115.504.161.540.2]sheatlf tidak menyempit pada bagian ujungnya (Gambar 5 d). Sayap depan jantan lebih sempit dengan rumbai yang ~anjang (Gambar 5 f).
Gambar 4. Imago Telenomus rowani
3. Telenomus dignus Gahan
- Abdomen betina kira-kia 2 kali lebarnya dan menyempit tajam ke arah
ujung (Gambar 7c). Pedisel betina sama atau sediiit lebih panjang dari
gabungan F1 dan F2 (Gambar 7b). Tungkai kuning cerah, kecuali koksa depan
agak gelap. Antena betina berwarna l e b i gelap dari pada antena jantan kecuali
skapus separuhnya benvama pucat. Antena jantan benvarna kuning kecuali 3-4
kali metatarsus. Genitalia jantan mempunyai penis sheath yang menyempit ke ujung dengan penebalan pada sisi dalam apical appendage (Gambar 7d). Ukuran 0,61 - 0,78 rnm.
Gambar 5. Karakter morfologi T. rowani, a: antena jantan; b: antena betina; c: abdomen betina; d: genitalia jantan; e: tungkai belakang jantan; f: sayap depan jantan; g: sayap belakang jantan (skala sama kecuali genitalia jantan).
4. Telenomus dignoides Nixon
Abdomen betina lebii kurang 3 kali lebamya yang secara berangsur- angsur menyempit ke arah ujung (Gambar 9c). Kepala lebii lebar daripada
torak, mata besar. Torak sedikit lebii panjang dari lebarnya. Sayap lebar dengan
kurang 3 kali segmen berikutnya. Antena betina berwama kehitaman (gelap)
kecuali skapus lebih berwarna pucat. Tiga atau empat ruas ujung antena jantan
berwana lebih gelap dibandiig ruas sebelumnya (Gambar 8). Tungkai jantan
hanya koksa depan yang berwarna gelap. Penis sheath n~enyenlpit ke arah
ujung, tetapi tidak ada penebalan pada apical appendages (Gambar 9d). Jantan
[image:149.602.116.477.280.523.2]berukuran 0,82- 0,97 mm sedangkan betina berukuran 0,95 -1,12
mm.
Garnbar 7. Karakter morfologi Telenomus dignus ; a: antena jantan; b: antenna betina; c:abdomen betina; d: genitalia jantan; e: tungkai belakang jantan; fi sayap depan jantan; g: sayap belakang jantan (skala sama kecuali genitalia jantan).
Gambar 9. Karakter morfologi
T.
dignoides, a: antena jantan; b: antena betina; c: abdomen betina; d: genitalia jantan; e: tungkai belakang jantan;f:
sayap depan jantan; g: sayap belakang jantan (skala sama kecuali genitalia jantan).
Telenomus sp. 1
Secara umurn tubuh bemama hitam. Abdomen pendek, perbandiigan
parjang dan lebar abdomen adalah 1,l dengan ujung membulat. Panjang torak
sedikit lebii panjang daripada lebamya. Perbandingan panjang dan lebar sayap
depan adalah 1,6 dengan rumbai pendek. Kepala lebjh lebar dari torak. Gada
pedisel, F4 paling kecil diantara ruas-ruas lain, bentuknya agak bulat. Tungkai
berwarna kuning. Tungkai belakang betina jauh lebii panjang daripada tungkai
yang lain. Ukuran tubuh 65 -70,4 rnm. Inang: Nezara viridula pada tanaman
[image:152.602.118.485.184.476.2]padi.
Gambar 10. Imago betina Telenomus sp. 1.
Telenomus sp. 2
Ukuran tubuh 0,47 - 0,65 mm, dengan torak yang menonjol atau lebih
tinggi daripada abdomen (Gambar 10). Abdomen betina berwarna hitam
mengkiiat. Secara umum morfologinya menyerupai
T.
remus dengan genitaliajantannya pendek dan agak lebar dengan bagian yang cekung dekat digiti. Selain
itu individu jantannya juga mempunyai sayap belakang dengan rumbai yang
panjang melebii lebar sayap. Funikula paling ujung dari antena jantannya
melebar. Funikula ke-4 antena betina paling kecil dibandimg ruas lain. Warna
tungkainya lebii gelap dibandiigkan tungkai jantan. Perbedaannya dengan
T.
remus adalah ukuran yang lebii besar dan sayap depan yang lebih lebar. inang:
Spodoptera sp. pada tanaman jeruk.
Garnbar 11. Telenomus sp.2, imago jantan (kiri) dan imago betina (kanan).
Telenomus sp. 3
Tubuh betina memanjang, kira-kia 113 abdomen bagian bawah
menyempit ke ujung dengan ovipositor yang panjang (Gambar 10). Ukuran
betina herkisar 1,07 - 1,17 mm. Perbandiigan panjang d i lebar abdomen
betina antara 2,8 - 3,4. Warna tub& kehitaman, tungkai kecokiatan. Gada
dari antena betina 4 mas. Panjang skapus kira-kia 3 kali panjang pedisel.
Funikula 1 lebii besar dibandiig F2, F3 dan F4 sedangkan F2 dan F3 sama
besar. Sayap sangat tipis sehingga sangat mudah robek. Perbandingan panjang
Ganlbar 12. Imago betina Telenornus sp.3
Telenomlrs sp. 4
Ukuran tubuh 0,68 - 0,72 mnl. Tubuh dan antena berwarna hitam
sedangkan tungkai coklat. Torak sedikit lebih panjang daripada lebarnya.
Abdomen pendek dengan perbandingan panjang dan lebar antara 1.14 - 1,35.
Tungkai dan antena banyak ditumbuhi rambut-rambut halus. Perbandingan
panjang dan lebar sayap depan 2,75 - 3. Rumbai sayap belakang lebii pendek
daripada lebar sayap. Inang: tidak diketahui, namun mempunyai ciri-ciri: telur
Ganlbar 13. Imago betina Telenontus sp. 4.
Telenomus sp. 5
Antena mempunyai pedisel yang lebii pendek dari funikula 1. Funikula
1, 2 dan 3 antena jantan hampir sama panjangnya. Funikula 4-9 bulat seperti manik. Abdomen betina lebar, perbandingan panjang dan lebarnya 2,2 - 2,6.
"
Tibia belakang betina lebih kurang dua kali metatarsusnya. Funikula 1 sedikit
Gambar 14. Imago betina Telenomus sp. 5
Kunci Identifikasi
Dari beberapa karakter morfologi spesies parasitoid Telenomus yang
ditemukan dapat disusun kunci identifikasi sebagai berikut:
1. a. Ukuran betina lebii besar dari 0,6 mrn..
...
..2b. Betina berukuran sama atau lebii kecil dari 0,6
mm.
Rumbai sayapbelakang jantan lebih panjang dari lebar sayap (Gambar 3g). Genitalia
[image:156.599.116.477.95.354.2]jantan pendek dan agak lebar, median plate lebar di bagian pangkal dan
...
cekung dekat digiti (Gambar 3d) ..T. remus
2. a. Penis sheath genitalia jantan membulat di bagian ujung (Gambar 5d).
Separuh bagian bawah abdomen meruncing dengan rata ke ujung
panjang dari lebarnya (Gambar 5b), antena jantan dengan funikula 1-3
lebih lebar daripada ruas-ruas berikutnya (gambar 5a). Betina berwarna
hitam sedangkan jantan coklat kemerahan (Gambar 4). Betina 0,75 -
0,93 mm dan jantan 0,56 - 0,75 mrn.
...
T. rowanib. Penis sheath pada genitalia jantan menyempit di bagian ujung (Gambar
7d dan 9d)
...
33. a. Panjang abdomen betina lebih kurang dua kali lebarnya, menyempit tajam ke arah ujung (Gambar 7c). Pedisel lebih panjang dari gabungan
funikula 1 dan 2 (Gambar 7b). Ada penebalan pada sisi dalan~ apical appendage genitalia jantan (Ganlbar 7 d)
...
..T. dignusb. Panjang abdomen betina kua-kira 3 kali lebarnya (Gambar 9c).
Genitalia jantan tidak ada penebalan pada sisi dalam apical appendage
(Gambar 9d)
...
..T. dignoides.Analisis Pengelompokan
Tabel 5. Matriks data 4 spesies Telenomus berdasarkan 16 karakter morfologi
- - -
T. rowani 1 l O O O l l l l O O O I O O O
T. remus 0 0 0 0 0 0 0 1 0 0 0 0 1 1 1 0
T. dignus 0 0 1 1 0 0 0 0 0 0 0 1 0 0 0 0
Keterangan: OTU (Operation Taxon Unit)=spesies
Tabel 6. Karakter morfologi yang diarnati serta kodenya.
No Karakter lnorfologi Kode
Ujung abdomen betina membulat
Separuh bagian bawah abdomen meruncing dengan rata ke ujung Abdomen betina menyempit tajam ke ujung
Metasoma betina panjang dan silinder Metasoma betina pendek
Panjang abdomen betina lebih kurang 3 kali lebar yang terlebar Panjang abdomen betina dua kali lebar
Panjang abdomen betina tidak seperti di atas
Pedisel betina lebih panjang dari gabungan F1 dan F2 Pedisel betina tidak lebih panjang dari gabungan F1 dan F2 Panjang tubuh betina berkisar dari 0,95 mm sanipai l,17 mm Panjang tubuh betina kurang dari ukuran tersebut di atas
Funikula 1-111 jantan sama dengan dua kali lebar segmen berikutnya
Funikula 1-111 jantan tidak seperti di atas
Sayap depan jantan seinpil dengan rumbai yang patijang Sayap depan jantan tidak seperti di alas
Rutnbai sayap belakang jantan lebih panjang dari lebar sayap Runibai dari sayap belakang jantan tidak melebihi lebar saqap Metatarsus dari semua tungkai jantan melebar
Metatarsus dari tungkai jantan tidak melebar
Tarsus belakang jantan panjang dan silinder sama dengan 3 segmen berikutnya
Tarsus belakang jantan tidak seperti di atas
Hanya koksa tungkai depan jantan yang berwarna gelap Semua koksa jantan berwarna gelap
Pada genitalia jantan ada penebalan seperti pita gelap di sisi dalarn
apical uppenduge
Tidak ada penebalan seperti pita gelap pada apical appendage Pe~iis .sheath tidak menyempit ke ujung
Penis sheath menyempit ke ujung Genitzlia jantan pendek dan lebar Genitaliajantan tidak seperti di atas
Median plale pada genitalia jantan cekung di bagian ujung dekat
digiti
Medianplate tidak cekung di bagian ujung dekat digiti
Panjang tubuh jantan berkisar dari 0,82-0,97 mm
[image:158.605.85.513.103.807.2]Tabel 7. Matriks jarak 4 spesies Telenornus berdasarkan 16 karakter morfologi
OTU T. dignoides T. rowani T. dignus T. rernus
T. dignoides 0.0000
T. rowani 1,5666 0,0000
T. dignus 1,2766 1,6338 0,0000
T. renius 1,6609 1,3185 1,5475 0,0000
Karakter Molekuler Telerzomus spp.
Hasil anlplifikasi DNA total dengan