KONTROL HORMONAL PADA IMPLANTASI
S K R I P S I
oleh
EDI WIRYANA AHMAD
B 17.1432
FAKULTAS KEDOKTERAN HEWAN
INSTITUT PERTANIAN BOGOR
1 9 S 5
Edi VI. Ahmad. Kontrol Hormonal Pada Implantasi ( Di-bawah bimbingan
R.
Kurnia Achjadi ).Penulisan ini bertujuan untuk mengetahui dan mempela-jari peristiwa hormonal yang terjadi dalam proses implan-tasi, selain i tu, juga mempelajari peristiwa fertilisasi, perkembangan embrio sebelum mencapai bentukan fetus dan
jalannya implantasi.
Terjaninya implantasi diinduksi oleh kerja dari hor-mon progesteron, estrogen dan kemungkinan besar juga ィゥウセ@
min dan prostaglandin. Sekresi progesteron dan estrogen adalah hasil rangsangan LH (Luteinizing Hormon ) dan LTH
( Luteotropik Hormon ) atau prolaktin, jadi LH dan LTH エ・セ@ libat tidak langsung dalam peristiwa implantasi. Sebelum
terjadinya implantasi estrogen menginduksi uterus untuk ー・セ@ siapan penerimaan blostosis pada implantasi, dalam proses induksi tersebut dibantu pula oleh progesteron.
Progesteron merangsang sistim kelenjar endometrium オエセ@
rus, dan menghambat kontraksi miometrium, sehingga ォ・エ・ョ。ョセ@
an miometrium menjamin pemukiman blastosis dalam uterus. Dalam proses induksi uterus untuk implantasi estrogen me -rangsang sistem RNA uterus untuk mensintesa protein, meta-bolisme lipid, karbohidrat dan perturnbuhan endometrial. Estrogen merangsang kontraksi untuk pergese,ran atau pemba-gian tempat bagi embrio di dalam uterus. selain itu estrQ gen juga mengsensitifkan terhadap rangsangan progesteron.
in-pada kerja hormon tersebut SBcara sendiri-sendiri.
Beberapa peneliti telah membuktikan pentingnya kontrol hormon tersebut di atas dalam persiapan inplantasi maupun dalam implantasi. Ketidak hadiran hormon-hormon i tu terbu.!.£ ti menunda terjadinya impl-antasi. Mengenai keterlibatan histamin, prostaglandin dan CO2 embrio atau ion hikarbonat dalam implantasi, masih merupakan bahan perdebatan dianta-ra bebedianta-rapa peneli ti.
SKRIPSI
OLEH
EDI WIRYANA AHMAD
B.17.1432
Skripsi yang dajukan sebagai salah satu syarat uutuk memperoleh gelar Dokter Hewan
pada Fakultas Kedokteran Hewan Insti tu t Pertanian Bogor
Fakul tas Kedokteran Hewan Insti tut Pertanian Bogor
SKRIPSI
OLER
EDI WIRYANA AHMAD
B.17.1432
Telah dᄆウ・エオェオセ@ dan Diper1ksa.
Oleh :
( Drh. R. Kurnia Achja·di., MS ) Dasen Ilmu Rep=duksi dan KeMdanan.
Faku.l tas Kedakteran Rewan Institut Pertanian Bagar.
Penulis dilahirkan di Tanggerang ( Jawa Barat ) pada tanggal 18 Agustus 1960, dari ayah Ahmad Sakiman dan ibu Esti, sebagai anak pertama dari tujuh bersauctara.
Penulis menyelesaikan pendidikan Sekolah Dasar di SD negri Mamajang Ujung Pandang tahun 1972, SMP negri I Ujung Pandang tahun 1975. Pada tahun 1979 penulis rnenama tkan pendidikan pada SMA negri II Jakar ta.
Alhamdulillah, penulis ucapkan, atas segala petunjuk dan rahmatNya dalam peny.elesaian skripsi ini, karena dengan keberkahannya jualah penulis dapa t mengatasi segala rintan.E; an selama penulisan ini berlangsung.
Penulisan skripsi ini disusun berdasarkan hasil studi litei'atur, sebagai salah satu syarat untuk memperoleh gelar Dokter Hewan Pada Fakultas Kedokteran Hewan Institut Perta-nian Bogor.
Pada kesempa tan ini, penulis sampaikan penghargaan dan ucapan terima kasih yang sebesar-besarnya kepada semua do-sen yang pernah mendidik penulis, terutama kepada :
Drh R. Kurnia Achjadi, MS atas segala bimbingan, saran dan keritiknya, yang penuh perhatian serta kesungguhan dalam mengarahkan penulis selama penulisan skripsi ini.
Penulis ucapkan terima kasih dan penghargaan yang mea dalam untuk kedua orangtua dan adik-adik, yang senantiasa berdo' a dan berkorban untuk keberhasilan ini.
Ucapan yang sama juga penulis sampaikan kepada staf Perpustakaan FKH, Perpustakaan Pusat Institut Pertanian Bo gor, BPT Ciawi dan LBN atas bantuannya selama penulis men-cari li tel'atur sebagai bahan penulisan.
Akhir kata, betapa penulis sadari, bahwa tulisan ini masih jauh dari harapan sempurna, namun demikian penulis berharap semoga akan berman faa t bagi penulis sendiri ser·ta bagi mereka yang haus akan pengetahuan. Amien.
Ha1aman
DAFTAR TABEL .. .. ..
.
.. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. .. ..... ..
i i iDAFTARGAMBAR •••.•••..•••.•••••.•..•..•••••••..••.• iv
PEND.IlHULUAN •••.••••...•.•.•...•••.••••••••.•• 1
TINJA UAN PU STAKA ••••••••••••••••••••••••••••••.••••
3
1. Ferti1isasi •••••••..••.•..••..•.••••••••••• 3
2 • C1ea vage •••••••••.•••..•.••.•.•••.•••..•••• 13
3 • Imp1an tasi ••••••...•••••....••••••••••••••• 16
4.
Horman-harmon Yang Pen ting Un t.ukImplantasi •••••••••••..•...•••••••••..•. •.• •
27
PE!1BAHASAN
KESIMPULAN
...
..
..
.. .. .. .. .. .. .. ..... ..
39
43
DAFTAR TABEL
Nomor Halaman
Teks
1. Rata-rata Umur Sperma Di dalam Saluran
Kela-min, dan Umur Ovum Sesudah Ovulasi
...6
2. Draja t Perkembangan Embrio .••••..•...•.••.••••• 16
3.
Perbandingan '.'iaktu. Pengenalan IndukTerha-dap Kebuntingan dan Perlekatan Embrio De-ngan JariDe-ngan Uterus Pada Dornba, Sapi, Babi
dan Kuda .••••••... " . . . . . .. . . . . . . . . .. . .• • 27
4.
Efek Variasi Dosis Estradiol 17-betha Dalam Penginduksian Implantasi Pada Tikus Yang Di-ovariektomi Pada Awal Kebuntingan danDAFTAR GAMBAR
Nomor Halamallll
Teks
1. Hubungan Antara Waktu Insiminasi dan
Fer·ti-lisasi Pada Sapi, Domba dan Bab.i •...•••• 7
2. Reaksi Akrosom Sperma dan Penembusan
Lapi-san Luar Ovum •.•••••••..••...•.•...•••.•...
9
3.
Diagram Yang Menggambarkan Proses-prosesYang Ter jadi Selama Pembuahan Pada Tikus ••••••• 12
4.
dセ。ァイ。ュ@ Elastosis Mulai MenyentuhEndomet-rlum ••••...••.••••.•••....••.•.••...•••••••.• 23
5.
Pengaruh Progesteron dan Estrog·en Dalam me-rangsang Decidualisasi dan Implantasi PadaPengetahuan mengenai proses terjadinya kelahiran ada-lah penting, karena dengan mengetahui kejadian dari proses tersebut dapat diketahui penanganan yang terbaik hila in-duk mengalami kesulitan dalam proses kelahiran, sehingga fetus dapat dilahirkan secara normal dan indukpun selamat. Tetapi tidak kalah pentingnya juga mengetahui tahap dari kejadian sebelum terjadi kelahiran, yaitu peristiwa ferti-lisasi sampai implantasi, karena proses dalam tahap i tulah maka terjadi embrio sampai terjadi fetus.
Terjadinya fertilisasi sampai implantasi dan kontrol hormon dalam proses implantasi, merupakan salah satu
dian-tara keajaihan-keajaiban セャ。ュ@ ilmu biologi yang hendak 、セ@
ll1.nglr.apkan.
KejaUian implantasi penting untuk diketahui, karena pengetahuan mengenai kapan akan terjadi implantasi b.erguna untuk kepentingan Embrio Transfer. セオウ。ャョケ。@ dalam proses panen embrio untuk kepentingan Emhrio Transfer harus
dike-tahui saat terjadinya implantasi, sebab hila telah terjaUi implantasi embrio sudah tak dapat dipanen. Harus diketa-hui pula bahwa uterus hewan penerima harus sesuai dengan kondisi embrio, agar ter jadi implan tasi, sehingga emb-rio dapat tumbuh dan berkembang dengan baik.
Menurut Sukra (1982), implantasi adalah hersarangnya blastosis di dalam uterus, sampai terjadi hubungan antara
selaput ekstra embrionik dan selaput lendir uterus. iューャ。セ@
a. Implantasi superfisial, dimana hlastosis ada diruang lumen uterus, misalnya. pada ungula ta, kelinci, ォ。イョゥセ@
vora dan primata rendah.
b. Implantasi eksentrik, blastosis terletak di dalam ォイゥセ@
ta atau lipatan selaput lendir, misalnya pada rodensia. c. Implantasi interstitial atau profundal, blastosis
me-nembus lapisa epitel uterus dan berkembang di dalam endometrium, misalnya pada manusia dan marmut.
Hormon adalah senyawa organik yang diproduksi oleh sel-sel tertentu yang normal dan seha t di dalam tubuh, da-lam jumlah sediki t dan langsung dialirkan kedada-lam pembulu darah menuju tempat lain untuk mempengaruhi aktivitas ォッッセ@
dinasi dan iILtegrasi bagian-bagian diseluruh tubuh indivi-du. Hormon-hormon yang terpen ting dalam implan tasi, yai tu LE, LTH, progesteron dan .estrogen. Sedangkan hi.stamin, prostaglandtn dan produksi CO2 embrio atau ion bikarbonat, diduga juga terliba t dalam perangsangan proses implantasi. Tanpa kontrol hormon-hormon tersebut implantasi tidak akan terjadi. Tetapi mengenai histamin, prostaglandiIL dan pro-duksi CO2 embrio a tau ion bikarbona t, beberapa peneli ti セ@
sih meragukan keterlibatannya dalam implantasi.
Penelitian lebih lanjut masih perlu dilakukan untuk merrgetahui lebih jelas mengenai implantasi atau peranan ヲ。セ@
tor hormonal dalam kejadian tersebu t, karena sampai saa t ini belum banyak informasi yang menjelaskan peranan ィッイュッセ@
1. Fertilisasi
F.ertilisasi adalah penyatuan dari dua sel gamet jan-tan dan betina untUK membentuk sa tu sel zigote. Dalam fe;;: tilisasi terdapa
t
proses ganda, yai tu dalam aspek embrio-logik fertilisasi meliputi pengaktifan ovum oleh sperma,tanpa rangsangan fertilisasi ovum tidak akan melalui clea-va.ge dan tidakp.erkembangan embriologik. Dalam aspek ァN・ョセ@
tik, fertilisasi mel:!:.puti pemasukan faktor-faktor heredi-tas pejantan ke dalam ovum ( Me. Larendalam Hafez, 1968 ).
MenUl:'ut Austin (1975), penyatuan ovum dan sperma dalam proses fertilisasi memerlukan pendeka tan membran kedua sel ganet tersebut sehingga membentuk satu sel tunggal.
Umur sperma dan ovum relatif singkat, maka waktu bagi
ke dua sel tersebu t ama t penting. Ke tepa tan meneapai tem-pat fertilisasi menyebabkan semua proses perjalanan, pen-dewasaan dan pertemuan harus diatur sedemikian rupa. Pro-ses ini dia·tur ol.eh alam seeara otoma tik ( Partodihar
So,
19.80 ).
Pengangkutan sperma di dalam lumen uterus disebahkan oleh kontraksi kuat dinding uwrus yang dirangsang ol.eh ーセ@
lepasan oksitosin pada waktu kopulasi atau insemiaasi bUa-tan ( Van Demas dan Hays yang diku tip Nalbandov, 1958 ). Haf.ez (1968) menambahkan pula, walaupun sperma diletakkan ke dalam seluruh kelamin betina dalam jumlah berjuta-juta
waktu yang lebih singka tkira-kira 15 Ire ni t sesudah
perka-winan.
Andeson dalam Cole dan Cupps (1968) serta Partodihar-dJo (1980) menyatakan bahwa pada saat sperrna akan memasuki tuba fallopii maka terjadi rintangan bagi aperrna yai tu be-rupa cincin mukosa yang menandai ba tas an tara ue trus dan
tuba dan dikenal dengan narna uterotubal junction (
UTJ ).
UTJ
ini juga berperan menyeleksi sperma ( Partodihardjo, 1980). Seleksi berikutnya sepanjang tuba fallopii セ・イェ。ᆳdi di isthmus terutama batas antara isthmus dan ampula, ウセ@
lain seleksi kwalitatif juga terjadi reaksi biokimiawi ( termasuk reaksi kapasitasi ) untuk menamb?-h kemampuan セ@
ri セウー・イイョ。@ untuk menembus sel-sel corona radiata yang ュ・ョセ@
lilingi sel ovum.
Pada kelinci dan tikus, kemungkinan besar pada sapi ser'ta mungkin pula jenis hewan lainnya, sperrna harus
men-jalani perubahan-perubahan fisi<>log.tk yang disebut kapasi-tasi ( Me. Laran dalam Hafez, 1968 dan Anderson dalam Cole dan Cupps, 1:968 ). Selanjutnya Austin (1951) ,dan Change
(1951) yang dikutip Austin (1975), menemukanbukti adanya proseskapastasi pada tikus dan kelinci dengF'ln menginj.eksi kan suspensi sperrna ke dalamkapsula ovarium pada tikus atau ke dalam oviduk pada kelinci dan pe·ngamatan menujuk-kan bahwa sper-ma membu tuhmenujuk-kan waktu yang agF'lk lama untuk セ@
1970 dalam Toelihere, 1981 ).
Salysburry dan Van Denmark (1961) serta Partodihardjo
(1980) menerangkan bahwa saluran reproduksi betina yang
ィ。イセウ@ dilalui ovum adalan セオ「。@ fallopii, kornUa uterus dan
tujuan terakhir yai·tu kornU3. u teru3 a tau korpus uterus.
Tuba fal10pii seeara ィセエッM。ョ。エッュゥォ@ dapat dibagi menjadi
fimbriae, infundibulum, ampula dan isthmus. Mereka
menya-takan pula bahwa kegiatan silia sepanjang tuba fallopii
dan kontraksi dari otot tuba fallopii sebagai faktor
pen-ting dalam pengangkutan ovum. Menurut Me. Laren dalam
Ha-fez (1968) dan Partodihardjo (1980), keadaan ini juga
di-atur oleh kesinambungan kerja hormon yang bersal dari
ova-rium, menjelang ovulasi estrogen merupakan hormon yang
do-minan.
Estrogen dibantu olsh oksitosin menyebabkan
terjadi-nya gerak peristaltik yang aktif. Pada saat terjadi ovu-.
lasi dan terlepasnya ovum, gerakan tersebu
t
kua t -:ianllle-nyebabkan fimbriae secara aktif menurunkan ovum. Setelah
ovum digerkkan menuju infundibulum dan kemudian masuk ke
ampula. Per jalanan ovum dalam ampula termasuk cepa t seba b
pengaruh estrogen masih kua t. Pada ba tas an tara
ampula-i::;;thmu8 ovum geraknya di perlahan sekali. Pada ba tas ini
terdapa t konsen trasi ,j-.inE,gi dari phosphat yang diduga
radiata yang menghubungkan ovum hingga penembusan sperma untuk ュ・ョセ。ー。ゥ@ ovum lebih mudah ( Partodihardjo, 1980 ).
Ovum tiba 、ゥセューオャ。@ dalam waktu yang cukup lama sesu-dah tibanya sperma untuk menjamin terlaksananya kapasita-si. Umur OVum umumnya kurang dari 24 jam demikian pula umur sperma, hal tersebut dapat dilihat pada tabel 1.
Tabel 1. Rata -ra ta Umur Sp.erma Di Dalam Sil.luran Kelamin dan U.mur Ovum Sesudah Ovulasi.
Umur sperma Umur ovum
Jenis hewan ( jam) (jam)
Menei t 10 - 12 6 - 15
Kelinci 30
-3"6
6 - 8Domba 30 - 48 16 - 24
Sapi 30 - 48 8 - 12
Kuda 72 - 120
6 -
8Babi 24 - 48 8 - 10
Manusia 28 - 48 6 - 24
Sumber : Toelihere, 1981 oleh karena i tu waktu terbaik un tuk inseminasi harus bena£. benar diperhatikan agar fertilisasi berhasil. セqウ。ャョケ。@ ーセ@
da sapi, yang secara normal berovulasi kira-kira 14 jam ウセ@ aUdah akhir estrus, angka konsepsi dari inseminasi yang
di
lakukan pada waktu ov.ulasi sanga t rendah, dan waktu terba-ik untuk inseminasi adalah dari6
sampai 24 jam ovulasi100
セ@
\\'iLk .. L Nセョゥャ。セエ@
-l
I
ualJl Jumua :''''\>I I
Al
,j.1
/
"-80
/
!":.
(
,
.\=
/:\
セ@
c 60
/
セ@
I : \
セ@
/
-" 0 [ I \
'"
40/
I I \I
セ@
\/
\ \\
/
\ I,
,
,
20
\
\
0
48 24 0 24 48
_ Jam, sebelum セ@
t akhlr estrus
-+-- Jam. sesudah セ@
akhlr estrus
Waktu inseminasi ,-'"
-GambaI' 1. Hubungan Antara l'/·aktu I.nseminasidan
F.lirti11 tas :pada セG。ーゥL@ ,Domba dan Babi.
Diku tip dari : Toel1here, 1981.
KeterallJgan Gambar : - Sap1 - Bab1
- Domba
Pada hampir semua mamalia, fert11isasi dimulai
sesu-dah badan kutub pBrtama disingkirkan, sehingga sperma ュセM
nembus dan masuk ke dalam ovum sewaktu pemhelahan reduksi
kedua sedanr. berlangsung. Akan tetapi, pada kuda, sperma
dimu-lai (Mc. Laren dalam Hafez, 1968 ). Tempat fertilisasi pada hampir semua ternak adalah bagian bawah ampula tuba fallopii ( Salysbury Van Denmark, 1961). Sewaktu masuk ke dalam ampula, selubung ovum, zona pelusida masih dikeli
lingi olsh sel-sel granulosa yang masih disebut sel-sel c!!. mulus. Pada ternak-ternak mamalia kecuali babi, sel-sel
cumulus menghilang dari Ova dalam beherapa jam sesudah ov!!. lasi.
Secara normal hanya sa tu sperma yang membuahi ovum. Oleh karena i tu ovum mengadakan reaksi zona dan hambatan vitelin untuk mencegah sperma yang lain masuk ke dalam ovum. Reaksi zona untuk merubah sperma lain menembus zona pelu-sida, sedang reaksi hambatan vitelin mencegah sperma meneE hus membran vi telin.
Untuk masuk ke dalam ovum, sperma pertama harus ュ・ョセ@
bus Ca) massa cumulus oo.phorus, bila masih ada, (b) zo-na pelusida dan (c) membran vitelin. Sperma menerobos ュ。セ@
sa cumulus oophorus dengan pergerakannya sendiri sambil
lD9-larutkan selubung asam Hyaluronik pada massa tersebut de-ngan enzim Hyaluronidase yang dihasilkan sperma ( Balins-ky, 1970; Hafez, 1974 ).
Menurut Toelihere (1981), sebelum sel sperma dapat ュセ@
GambaI' 2.
i8
Ie)
Reakst Akrosorn $.perzna nan Penembusan
r.ap;
,>-,),
1uM' 0vum.Dikutip dari Cole dan Cupps, 1968
Keterangan GambaI' :
A • Sperma yang sudah men,ga1ami kapasi tasi ( kiri j,
ketika sperma mengalami kapasitasi ( kanan ).
B.
Reaksi akrosom sperm a ( kiri ), ketika spermamenembus corona radta ta ( kanan ).
C. Sperma yang mengalami reaksi akrosom ( kirt ),
ke tika sperma menembus zona pe1u-sida ( kan.an ).
Dilaporkan bahwa enzim pe1ebur zona ( zonalysin ),
me-mungkinkan sel sperma melebur dan membuat suatu celah mel;a lui zona kedalam ruang peri vi telin ovum ( S tambrugh dan Buckley, 1968 dalam Hafez, 1968 ).
Balinsky (1970) dan Hafez (1974), menyatakan bahwa ovum mengeluarkan sua tu Za t ( fertilisin ) yang bereaksi dengan sperma dan terjadilah aglutinasi. Proses aglutina-si tidak menghentikan pergerakan sperrr.a, karena sperma te-rus berenang melalui zona pelusida dan lIlenimbulkan aluran kecil. Pada saa t ini akrosom yan.g merenggang sewaktu kapg si tasi akhirnya menghilartg dan menimbulkan perfora torium. Mungkin aktivitas suatu enzim tertentu berhubung dengan perforatorium yang memungkinkan perterobosanzona pelusida.
Fase terakhir penembusan ovum melipu ti pertautan ke-pala sperma ォセ・イュオォ。。ョ@ vitelin. Priode ini sangat
pen-ting karena pada saat itulah terjadi aktivasi ovum. Te-rangsang oleh pendeka tan sperma, ovum terbangki t dari kea-daan diamnya dan ter jadilah perkembangan. Kepala sperma juga ekor sperma memasuki ovum. Suatu proyeksi pacta per-mUkaan vitelin menandakan jalan masuk sperma. Membran pla§. rna sperma dan ovum pecah, dan kemudian bersatu memben.tuk selubung bersama. Sebagai akibatnya, sperma memasuki vi te lin, meninggalkan selubungnya bertaut pada membran vi telin ( Toalihere, 1981 ). Pada alternatif lain, membran plasma sperma pecah ( Piko dan Tayler, 1964 dalam Toelihere, 1981 ) pada kedua kejadian tersebu t kepa la sperma secara terbulill
Nenuru t Shumaway dan Adarnstone (1942), pada ovum yang baru dibuahi maka akan terjadi ; pemagkatan aktivitas
me-tabolisme, peUingkatan viskositas sitop1asrna dan peningka-tan permeabi1itas membran.
Segera setelah sperma masuk vitelin kepala sperma mem-bengkak dan kehi1angan 「・ョセオォL@ membran sekelilingnya tak nampak lagi. Selama proses ini, material im menghilang dari akrosom, kemudian segera terbentuk nUkleus-nukleus di-mana pertama-tama sebagai ti tik-ti tik keeil. Nukleus-nuk-leus yang terbeIlJtuk bersa tu dan memhentuk membran disekeli-lingnya, maka terbentuklah pronukleus jaatan. Pembentukan pronukleus betina mengikuti pola yang sama dengan pembentu-kan pronukleus betina ( Salysbury dan Van Denmark, 1961 ). Nenurut Hafez (1974), pembentukan pronukleus betina segera
setelah badan kutub kedua dikeluarkan dari ovum.
Ukuran nUkleus bervariasi pada setiap indi vidu -dan spesies. Pronukleus juga ukurannya bervariasi, pronukleus
jantan pada tikus, menei t dan kueing besar, pada beberapa spesies dUa pronuldeus samahesarnya.
mem-Gambar
3.
Dia.gram Yang Menggamharkan Proses-Proses Yang Ter jadi. S-elama Pembuahan ?ada TikI! s.i ,---,,'
p「セzNpG@
セ@
\
2nCl
a
Keterangan Gambar :
a. Sperma berkontak dengan zona pelusida (Z.P) badan kutub p-ertama (Pb.l) telah disingkirkan ; inti sel telur sedang mengalami pembelahan miotik yang kedua (2nd M).
·b. Sperma telah menem1:Jus zona pelusida, dan kini ber.
taut pada vi telin (vi t). Hal ini merangsang イb。セ@
sizona, yang ditandai oleh pembayangan yang rne-nyelusur sekeliling zona pelusida.
c. Kepala sperrna masuk ke dalarn vitelin, dan terle-tak di bawah perrnukaan yang telah terungki t di atasnya.
d. Sperma hampir kini seluru}1nya herada di dalam
vi-telin. Kepala sperma membengkak. Vitelin
berku-rang v.olumenya, dan hadan ku tub kedua sUdah di-singkirkan.
e. Pronukleus jan tan dan betina mUlai berkernbang.
Mitochonria (Mit) berkumpul disekitar pronukleus.
f. Pronukleus b.erkernbang sempurna dan mengandung
ha-nyak nukleus. Pronukleus jantan lebih besar dari
pada betina.
g. Fertilisasi telah sempurna. Pronukleus telah meu£
hi lang dan diganti oleh kelornpok-kelompok krorno-sorn yang telah bersatu di dalarn prophase pada pem-hagian cleavage yang pertama.
( Sumber : Austin dan Bishop,
1957
dalarn [image:23.613.104.463.94.278.2]bran inti hilang. Kelompok kromosom mengganti inti yang hilang, dan dua kelompok kromosom menjadi satu. lni di-ikuti oleh penyusun kromosam dalam tahap metaphase,
gelen-dong pembelahan pertama segera nampak. Perkiraan.interval dari pembentukan pronukleus sampai pembentukan gelendong pembelehan pertama bervariasi, tetapi umumnya diperlukan 12 sampai 20 jam untuk perubahan ini.
2. Cleavage
Sesudah singami selesai, untuk beberapa hari lamanya zi.got.e atau embriohidup bebas di dalam tuba fallopii atau uterus induk. Di dalam uterus, makanan enbrio diperoleh dari sekresi kelenjar-kelenjar uterus. sesudah implantasi embrio memper.oleh makanannya dari saluran darah induk.
Pada awal priode hidup aebasnya ovum merupakan satu sel tunggal, dengan volume yang rela tif besar dibandingkaa. dBngan sel-sel tubuh lainnya. Sehingga perbandingan si to-plasma dengan nukleus sangat besar. Bahan-bahan makanan persediaan disimpan di dalam si toplasma dalam bentuk ku-mng telur ( deutoplasma ).
Sel tunggal ini lalu mengalami cleavage y,ai tu membagi. diri beberapa kali tanpa tamhahan volume si toplasma dan pertumbuhan. Proses im berlangsung terus sampai
implan-tasi dimana ukuran sel telah sama dengan ukuran sel tubuh ( mセN@ Laren dalam Hafez, 1968 ).
Fertilisasi satu sel telur ditransformasikan oleh
hagi-an-hagian mi tosis yang tera tur ke dalam kompleks multi celluar.
Tidak ter jadi pertumbuhan.
Bentuk umum dari embrio tidak men.galami perubahan, ke -cuali untuk pembentukan rongga dibagian dalam blastosul. Selain dari pada itu transpormasi bahan-bahan sitoplas-ma ke dalam bahan-bahan inti dan perubahan-perubahan kwalitatif di dalam komposisi kimia dari telur di bata-si.
Bagian-bagian dari si.toplasma telur tidak diganti lemh lanjut .dan bertahan dalam posisi yang sama seperti pada permulaan cleavage.
P.erbandingan inti terbadap si toplasma rendah pada perm),! laan cleava-ge kemudian pada akhir cleavage merucapai ba'" tas tertiruggi seperti yang terdapa t dalam sel soma tik.
Cleavage pada telur yang dibuahi diawali oleh pembela-han inti yal1g diiku ti oleh pembelapembela-han si toplasma, sehilligga dihasilkan dua sel anak. Dua b,lastomere yang p.ertama mem-bagi diri lagi sehingga menghasilkan Lf blastomere,
kemudi-an 8, 16 dkemudi-an seterusnya. Semua pembelahan sel tersebut bersifat mitosis sehingga setiap sel embrio mengandung
kh-romosom diploid {2n) { Balinsky, 1970; Anderson dalam Cole dan Cupps, 1977 ).
terhen-tuk bila jumlah sel blastomere dalam zona pelusida telah mencapai 16-32 bUah. Beberapa spesies seperti kelinci di-mana pembelaharmya kontinyu sehingga morula dapat mengan-dung 100 sel. Sedangkan ukuran dari sel hlastomere dari morula berkurang oleh mitosis yang kontinyu sampai sel blastomere mencapai bentuk sel normal seperti sel tubuh dewasa. Perubahan dalam perbandiagan si toplasma inti di-anggap penting dalam pengaturan ( regulasi ) kegiatan atau sifat genetlk di dalam embrio. Tidak hanya ukuran embrio, mamalia gagal untuk bertambah selama proses pemhelahaa, t.§. tapi bukti pada menci t menunjukkan terjadi juga pengurang-an jumlah sel selama beherapa hari pertama dari proses pel: kembangannya.
Menurut Partodihardjo (1980) bila terbentuk morula, cairan mUlai terlihat berkumpul diantara beb.erapa sel da-lam tubuh morula. Ruangan ini disebut blastosul, sedang embrio kini 1iisebu t blastosis. Jika blastosis telah ter-hentuk ·maka tuhuh embrio seolah-olah terbagi dua, karena
ada bagian sel yang tumbuh membentuk sel-sel tipis dibagi-an permukadibagi-an ydibagi-ang menyelubungi hampir seluruh tubuh
;11as-tosul. Bagian yang menyeluhungi ini disebut trofoblas, s.§. dang bagian yang diseluhungi disebut massa sel bagiab dalam ( inner cell mass ). TrofDblas mempunyai fungsi menyerap cairan yang mengandung nu trisi bagi embrio.
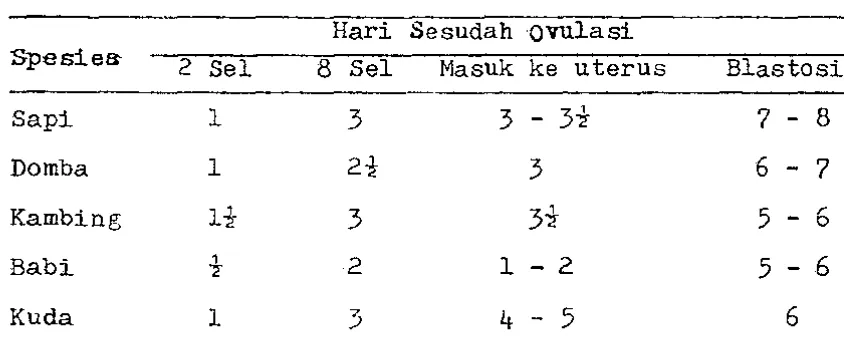
Tabel 2. Derajat Perkembangan Embrio.
Hari Sesudah ovulasi
Spesiea- 2 Sel 8 Sel Masuk ke uterus Elastosis
Sapi 1 3 3 - 3"2-
7 -
8Domba 1 2* 3 6 - 7
Kambing
H
3 3"2-5 -
6Babi
t
2 1 - 25
- 6
Kuda 1
3
4 -
5
6Sumber Toelihere, 1981. Dalam pertumbuhan selanjutnya trof'oblas akan tumbuh menjadi plasenta, sedangkan massa sel hagian dalam ( inner cell mass ) tUmbuh menjadi makhluk yang baru lahir·. Zona pelusida pada beberapa mamalia akan pecah setelah hlasto-sis menyentuh endometrium untuk proses implantasi. Tetapi pada beb;erapa mamalia lain misalnya pada marmut,zona pelu sida terkelupas oleh a.danya juluran-juluran protein yang b-erasal dari trcfoblas menjelang implantasi.
3.
Implantasi.Implantasi dapa t ter jadi bila embrio dan uterus telah sama-sama siap untuk menjalani proses terseuut. Untuk itu embrio harus mengalami beberapa perubahan sampai mencapai tahap hentuk terten tu yang siap diimplantasikan, demikian pula uterus mengalami beberapa perubahan sampai mencapai
[image:27.620.76.498.113.288.2]ber-langsung dengan baik, sehingga perkembangan embri-o selan-ju tnya ter jamin.
a. Pengertian Implal!ltasi
Implan tasi didefinisikan sebagai sua tu proses perleka-tan embrio mamalia dengan dindin-g uterus, dengan penembus-an epitelium dpenembus-an dikukuhkpenembus-an oleh suatu hubungpenembus-an dari
sis-tim sirku1asi induk. Implantasi perlu sua tu koordinasi interaksi antara embrio dan kondisi uterus yang seharusnya \ Weitlauf, 1978 ).
Menurut Hafez (1968) dan Partodihardjo (1980) istilah implantasi 1ebih sesuai un-:tuk-_,hewan- yang embrionya terta-nam dalam dinding uterus. sebagai contah pada hewan ー・ョァセ@
ra t ( raden tia ) blastosis tertanam dalam kripta endometri um dan se1uru·h tro.foblas berhubungan sanga t eEa t dengan
dinding ke1enjar endometrium. Sedang pada ternak mamalia sebaliknya embrio tetap di da1am lumen uterus dan pertau-tan yang dibentuk dengan dinding uterus sebelum pembentu-kan plasenta adalah sangat labil. Pergerapembentu-kan blastosis di dalam uterus semakin terbatas dengan perkembangannya.
Peristiwa terjadinya per1ekatan embrio, penembusan epitelium, pembentukan plasenta dan keadaan uterus berbeda paria setiap spesies mama1ia. Pada beberapa spesies, irnpla!!.
di antara ke 10 dan ke 22 se telah koi tus pada domba dan an tara hari ke 10 sampai hari ke 40 pada sapi ( Nalbandov, 1958; Toelihere, 1981 ).
b. Faktor Embrio.
Pembentukan blastosis diikuti oleh gatrulasi yang me-rupakan pendahulu dari pembentukan organ tubuh. Gastrula-si terdiri dari gerakan-gerakan sel atau sekelompok sel sedemikian rupa untuk merubah ・ュセイゥッ@ dari struktur dua ャセ@
pis menjadi tiga lapis dan membawa daerah-daerah bakat ー・セ@
bentuk organ kepasisi yang defini tif di dalam embrio. Pada mamalia, gastrulasi hanya meliputi sel-sel pacta discus embrional. Dari sini berdefrensiasilah tiga macam jaringan endoderm, mesoderm, ektoderm. Dari jaringan-jar;!. ngan ini terbentuk semua jaringan feotalis dan selubung-selubung embrional yang menghubungkan embrio dan fetus ke-induk. Sel-sel berimigrasi atau melepaskan diri dari mas-sa sel dibagian dalam discus embrional un tuk membentuk se-lapis endoderm yang menyebar ke_sekeliling bagian dalam bla§.
tosis, membentuk omphalopleura bilaminar. Pada waktu yang sarna notochord dan mesoderm terbentuk oleh invaginasi sel-sel didaerah garis primi tif pacta discus embriona1 ( Hafez, 1968 ).
Se1ama gastrulasi discus enbrional menebal. Baris primitif yang terbentuk menentukan sumbu crania-caudal em-brio. Sewaktu mesoderm menyebar keluar dari garis
lapis, dipisahkan oleh coelum. Notochord berkembang dari ujung anterior garis primitif.
Disebelah dorsal notochord ektoderm menebal membentuk dataran neural. Sesudah heberapa hari lipa tan-lipa tan ne-ural bertumbuh dan bersa tu membentuk sua tu pipa nene-ural, yaitu bakat otak sumsum tulang belakang. Sementara itu somit, kondensasi berganda mesoderm dorsal telah kelihat-an pada masing-masing sisi notochord. Sewaktu embriome-manjang, pasangan somi t-somit tambahan terus berkemhang, sehingga jumlah somi t dapat dipakai seb.agai indeks umur embrio muda.
Pada tingka tan somi t-7, somi t anterior berdefrensiasi menjadi
3
bagian, diperun.:tukan bagi pembenctukan urat-urat, aaging, tulang, ォ・イ。ョセォ。@ dan tenunan pengika t. Se gera se-slidah ini baka t-baka t telinga dan ma ta jelas terliha t pada kepala embrio dan jantuu,g mulai berkembang sedang saluran pencernaanterbentuk dari kantong-kantong di dalam endoderm dan mesoderm ( Hafez, 1"968 ).Pada jenis hewan polytocous, blastosis didistribusi-kan menurut panjang kornua uteri sebagai akibat pergerak-an dindill;g uterus. Mekpergerak-anisme ypergerak-ang pasti dalam pembagipergerak-an ruangan secara sempurna b.agi embrio selama implantasi be-lum diketahui. Tidak terdapa t bukti l:Iahwa suatu blastosis yang sudah berimplantasi akan menghan.ba t implantasi blast,Q. sis yang lain dideka tnya.
steraidogenik yang di tunjukan oleh pertumbuhan embrio kel:i.!!l ci menduga bahwa embrio mungkin mempunyai kemampuan untuk mensintesa ウエイ・イセゥ、@ dan hormon-hormon protein.
Akhir-akhir ini telah di temukan hahwa preimp1antasi pada baIod. menjadi kegia tan yang penting di dalam proses bi osintesa estrogen ( Perry dkk dalam Cook dan Hun.ter, 1982). Estrogen diproduksi oleh jaringan trofoblas pada ーセイゥ@ ke 12, produksi ini dimaksudkan terlibat dalam pemberian tan-da untuk peme1iharaan luteal. Bukti steroidogenesis oleh blastosis sapi dan domba tidak sejelas seperti pada babi.
Tetapi Aya10n dkk yang dikut1p Cook dan Hunter (1982) te-lah mengamati adanya produksi streroid oleh blastosis sapi, .clan mendukung pendapa t adanya kegia tan steroido.genik pada blastosis.Godsby dkk (1976) dalam Cook dan Hunter (1982) menyatakan ·bahwa steraido.genesis ti·mbu.L sebelum dan sesud9.h perlekatan. Produksi-produksi utama pada spesies i tu ada-lah steroid-steroid netral, produksi estrogen oleh hlasto-sis ren-dah a tau sarna sekali tidak ada.
Menurut Nartenz dkk (1976) yang dikutiP Cook dan Hun-ter (1982), sHun-teroid-sHun-teroid yang dihasilkan dapa t berfung-si sebagai pengisyarat pada induk akan adanya embri:o 、ゥオエセ@
c. Faktor uterus.
Pada waktu embrio mengalami cleavage dan pembentukan blastosis uterus juga men.galami perubahan-perubahan melJJye-diakan diri untuk implantasi.
Pada proses preimplantasi di dalam uterus terjadi ーセ@
nambahan vaskularisasi pada endometrium dan peningka tan peFtumbuhan serta kegiatan dari kelenjar uterus ( Eckstein
dan Kelly dalam Cole dan Cupps, 1968 ).
Weitlauf (1978), menyatakan bahwa untuk suatu implan-tasi pada mamalia biasanya uterus membentuk suatu reaksi decidua sebagai respon. Di dalam kejadian ini stroma end£ metrium, sel fibroblastik ditransformasikan ke dalam
ben-tuk sel decidua khusus. Sel ini ditandai dengan penolJJjol-an epithelloid, kehadirpenolJJjol-an imti poliploid, akumulasi glikQ-gen dan lipid di dalam sitoplasma, pembentukan banyak li-sosom dan terjadi kontak antara sel dengan suatu hubungan yang kompleks. stroma endometrium ini akan ュ・ョェ。、ゥッ・、・ュセ@
tus sebab terjadi vasodilatasi dan penambahan permiabili-tas pembulu kapiler, peningkatan mitosis dan kegiatan me-tabolisme.
Cairan uterus mempunyai ー・イ。ョセョ@ sangat penting dalam menunjang kehidupan embrio menjelang implantasi. Peneliti an mengenai hal ini telah dilakukan pada kelinci. Dalam
gliein, alanin, taurin dan glutamin eukup tinggi kadarnya dan asam-asam animo ini merupakan substra t yang baik ba.gi tumbuhnya .embrio sebelum terbentuknya plasenta ( Partodi-hardjo, . 1980 ).
Psyehoyos (1973), menerangkan bahwa adanya ウゥョォイッョゥセ@
si dari hubungan telur dan uterus menjaminembriosampai mencapai posisinya yang eukup untuk diimplantasikan dan ェセ@
ringan induk dilindungi dari penyerbuan yang tidak terkon-tro1 jaringan trofoblas dari b1astosis. Sedang endometri-um biasanya te1ah siap menerima pada saat emhrio telah meg ,capai tahap blastosis atau b1astulasis masih ber1angsung.
d. Ja1annya Implantasi.
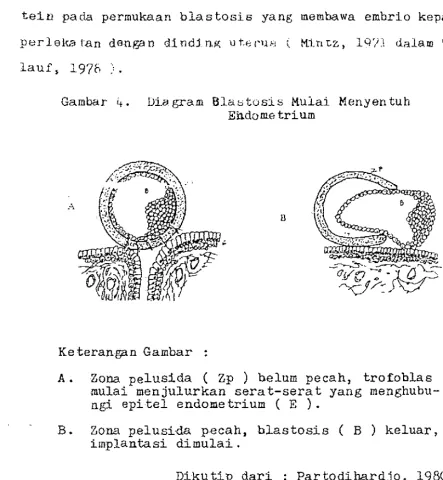
Menurut Partodihardjo (1980), implantasi berlangsung secara bertahap. Tahap-tahap ini adalah tahap persentuhan embrio d.engan endometrium, ter1.epasnya zona pelusida, per-geseran a tau pembagian tempa t dan yang terakhir ada1ah per. tautan antara trofoblas dengan epitel endometrium.
Tahap pelepasan zona pelusida adalah penting karena ·zona pe1usida merupalr..an sua tu penghalang un tuk imp1antasi.
g1ikopro-tein pada permukaan blastosis yang membawa embrio kepada
perleka tan dengan dinding ut.el'lW ( M.i.n t.z , ly?1 dalam Wei
t-lauf,
19'70
:"
Gambar 4. Di.agram Blaotosis Mulai Menyentuh
Endo me tri um
A
B
Keteran·gan Gambar :
A • Zona pelusida ( Zp ) belum pecah, trof.oblas
mulai menjulurkan serat-serat yang menghubu-ngi epi tel endometrium ( E ).
B. Zona pelusi-da pecah, blastosis ( B ) keluar,
implantasi dimulai.
,
Dikutip dari : Partodihardjo,
1980.
Sebalum implantasi .cairan blastosul men.gandung banyak
ion kalium dan bikarbona t, bahan ini berasal dari cairan
uterus. Setelah terjadi implantasi jumlah kalium dan
bi-karbonatberkurang sehinggp. sarna dengan kadar yane;
terda-pa t di dalam serum induk. Te tapi kadar pro tein dan
gluko-sa, fosfor dan khlor yang mula-mula rendah menjadi tinggi
[image:34.612.54.497.63.542.2]Pe-lepasan bikarbonat dari blastosis mempermudah trofoblas me-lekat pada selaput lendir uterus dengan demikian memperlan-car implantasi (Sukra, 1981 ).
Menurut Hafez (1974) dan Toe1ihere (1981), proses mor fologik imp1antasi ternak mamalia sebagai berikut :
Babi
Karena jumlah embrio banyak maka penempatan embrio oleh kontraksi uterus. Periode permu1aan pertautan ィ・イャ。セァ@
sung dari kir.a-kira hari ke-12 sampai hari ke-24 sesudah fertilisasi. Menjelang seki tar hari ke-7 zona pelusida di-sekeliling blastosis telah lepas sehingga sel-sel trofob-las berkontak langsung dengan epitel uterus. Trofobtrofob-las kini mulai memperbanyak diri secara cepat yang menyebabkan pelipatan dinding trofoblas, mungkin karena akumulasi cair-andi dalam rongga blastosis t:Ldak dapat mengikuti kecepa,!: an pengembangan dinding. Endoderm mulai muncul dan blastQ sis herubah dalam beberapa hari dari suatu gelembung bulat kecil menjadi suatu pipa herbentuk benang yang sangat me-manjang, kadang-kacang mencapai panjang beberapa kaki. Discus embrional menempati satti bagian yang besar pendek dipertengahan pipa. Pada saa t tersebu t dinding uterus her-lipat-lipat dalam dan lapisan luar ( khorion ) dari blast£ sis yang memanjang dihadapkan pada epi tel uterus, mengiku-ti jalannya lipatan-lipatan. Selama priode ini makanan
Do mba
Permulaan perkembangan blastosis domba sarna den§an babi. Sediki t pertautan telah terjadi seeepa tnya pada ha-ri ke-10 tetapi blastosis masih dapat digoyahkan keluar da ri uterus 16 sampai 17 hari sesudah perkawinan. Perpanja-ngan yang agak lambat dan tidak dimulai sebelum hari ke-l1 a tau 12 tetapi blastosis rnasih dapa t meneapai panjang 20
em menjelang minggu ketiga.
Proses imp1antasi domba berbeda dengan babi karena uterus domba mengandung karunkula. Menjelang hari ke-18 khorion telah berkembang sehingga memenuhi lumen uterus, membawa trofoblas berkontak dengan epitel uterus karunkula. Sel-sel trofoblas rnasuk ke dalam eelah vili pada karunkula dan terjadilah pertautan yang erat karena terjadi pelebur-an sel vili kemudipelebur-an seluruh epitel uterus berkontak de-ngan sel-sel tersebut.
Sapi
Jalannya implantasi pada sapi hampir sama dengan pada domba tetapi dimulai lebih lambat. Zona pelusida dilepas-kan pada seki tar hari ke-8 pada tingka tan blastosis muda dan beberapa hari kemudian blastosis mulai mernanjang. Gastrulasi selesai pada hari ke-13. Nenjelang hari ke-33 khorion telah membentuk sua tu pertau tan yang rapuh dengan 2 sampai 4 buah kotiledon sekeliling fetus dan dalam waktu
oleh ko tiled{)n. Kuda
Tahap hidup bebas blastosis kuda dalam lumen uterus dapat mencapai hampir dua bulan. Selama minggu ketiga b1a§
tosis memperoleh suatu komplek sel-sel kolumnar, discus trQ fob1astik dapat terlihat pada dinding trofoblas dari khori-on. Beberapa sel memilikiproses yang mungkin bersifat fa-gosi tik. Discus trofob1astik mungkin membantu pertau tan tetapi lebih mungkin untuk dihubungkan dengan ingesti susu uterus. Menjelang minggu ke-IO vili ( penonJo1an-penonjol-an ) dari khorion menju1ur masuk ke dalam lipatpenonJo1an-penonjol-an_mukosa pada dinding uterus dan menjelang minggu ke-4 pertautannya
te1ah sempurna.
Setelah implantasi dilan:jutkan dengan plasentasi, yai-tu pr{)ses pembellyai-tukan p1asenta an tara tenunan yai-tubuh dari embrio dan induk hewan untuk keperluan penya1uran makanan dari induk kepada anak dan za t buangan dari anak kepada in-duk.
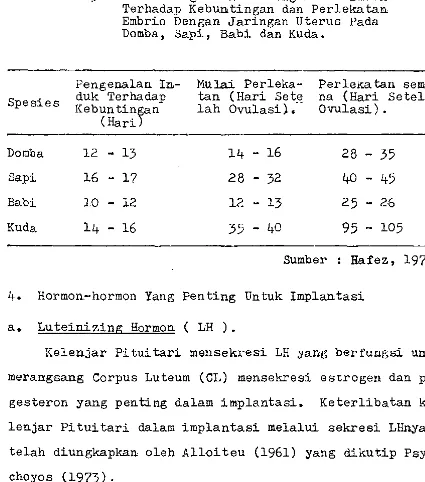
Tabel
3.
Perbandingan Waktu Pengenalan Induk Terhadap Kebuntingan dan Perlekataa Embrio Dengan Jaringan Uterus Pada Domba, Sapi, Babi dan Kuda.Spesies
Pengenalan In-duk Terhadap Kebuntingan
Mulai Perleka-tan (Hari Sete lah
Ovulasi):-Perlekatan sempu£ na (Hari Setelah Ovulasi) •
(Hari)
Domba 12 - 13 14 - 16 28 - 35
Sapi 16 - 17 28 - 32 40 - 45
Babi 10 - :1.2 l2 - 13 25 - 26
Kuda 14 - 16 35 - 40 95 - 105
Sumber : Hafez, 1974.
4. Hormon-hormon Yang Penting Untuk Implantasi a. Luteinizing Hormon ( LH ).
Kelenjar Pituitari mensekresi LH yang berfungsi untuk merangsang Corpus Luteum (CL) mensekresi es-crogen dan
pro-gesteron yang penting dalam implantasi. Keterlibatan ke-lenjar Pi tui tari dalam implantasi melalui sekresi LHnya
telah diungkapkan oleh Alloiteu (1961) yang dikutip Psy-choyos (1973).
Raj dkk (1968), dengan menggunakan anti FSH ( FOlikel
<V
[image:38.616.73.498.83.565.2]yang apabila diberikan pada hari ke-4 !nasa kebuntingan se-lama 10 jam berhasil dalam menghambat implantasi pada hari ke-8 masa kebuntingan. Hal tersebut terjadi karena dcsis itu anti serum menghambat banjirnya estrogen yang diperlu-kan un tuk implan tasi • Diusuldiperlu-kan juga bahwa pengeluaran IH
mendahului pengeluaran estrogen pada hari ke-4 :Jnasa kebun-tingan.
Perlakuan LH pada hari ke-12 sesudah kawin ternyata dapat menginduksi implantasi blastosis, sedangkan dengan perlakuan FSH dan LTH tidak memberi efek induksi un tuk im-plantasi. Hal inl mendukung hipothesa bahwa LH merangsang sekresi fisiologis jumlah estrogen dari lu teining ovarium ctengan atau tanpa kemampuan untuk mensekresi progesteron ( Macdonald dkk, 1967 ).
Penyelidikan baru-baru ini mengusulkan pel epa san estrQ gen dari dari ovarium pacta tengan hari pada hari ke-4 masa
ォ・「オョエゥョセョL@ didahului oleh banjir LH yang disekresikan
oleh Pi tui tari. Per"Cobaan dengan anti serum LH dalam 10 jam pada hari ke-4 masa kebuntingan menghasilkan penghambaj: an total terhadap implantasi ( Raj dkk, 1968). lni karena LH belum merangsang pembentukanestrogen oleh Dvarium se-hingga implantasi masih dapat berlangsung.
kare-na HOG mempunyai struktur protein serupa dengan LH, sehing ga HeG dapat merangsang sekresi estrogen pada ovarium, oleh karena itu implantasi masih dapat ber1angsung. b. Luteotropic Hormon ( LTH )
Keterlibatan LTH atau prolaktin da1am imp1antasi mer!!, pakan keterlibatan tidak 1angsung sama ha1nya dengan LH. Prolaktin merangsang OL untuk mensekresi progesteron yang diper1ukan un tuk menginduksi terjadinya imp1antasi.
Evans (1941) da1am Partodihardjo (1980), te1ah membuk tikan bahwa prolaktin dapat memper'tahankan hidupOL tikus. Kemudian Denamur (1973) yang dikutip Partodihardjo (1980), me1akulr..an percobaan untuk menguji apakah LTH dapat hekerja sendiri, tanpa 「。ョセオ。ョ@ LH dan FSH. Den,gan menggunakan doE!. ba yang te1ah dihipofisektomi, kemudian diberi suntikan hormon dengan kombi,nasi : LT"d, LTH '" LH dan LTH + FSH. Dari percobaan tersebu t disimpu1kan bahwa LTH dapa t
heker-ja secara maksima1 hanya bila di tambah LH. Sedang LTH mu£:
ni a tau yang ditamhah FSH efek bio1ogiknya sangat kecil { Partodihardjo, 1980 ).
Se1aIllju tnya 'I'oe1ihere (1981), menya takan ,hahwa LTR mampu meme1ihara aktivitas fungsiona1 'OL atau aktivitas Iu teo trOpic dan merangsang deciduoma pada uterus tikus dan: mencit. 01eh karena pembentukan deciduoma uterus
bahi ( Dunea dkk, 1961 da1am Toelihere, 1981 ).
Murphy (1979) juga telah membuktikan bahwa LTH diper-lukan pada perangsangan sekresi progesteron untuk kepenti-ngan implantasi pada ferret { sebangsa musang ) yang dihi-pofisektomi. Dari hasil penelitian yang dilakukannya
di-simpulkan juga bahwa prolaktin secara kwalitatif mampu me-melihara CL pada awal pertenganhan kebuntingan, dan seeara kwantitatif dapat memelihara sekresi progesteron dari CL
ferret yang dihipofisektomi. e. Progesteron
Menuru.t Partodihardjo (1980), progesteron mempunyai tiga pengaruh nyata pada uterus, yaitu :
Menghambat kontraksi miometrium, sehingga ketenangan
Ini
ometrium menjamin pemukiman hlastosis dalam uterus. Selanjutnya progesteron juga meniadakan pengaruh oksi t.Q. sin pada miometrum, agar tidak menginduksi kelahiran. Merangsang tumbuhnya kelenjar-kelenjar susu uterus paila mi,ome tri um.Pada spesies-spesies tertentu merangsang perkembangan sel-sel permukaan endometrium untuk menerima blastosis yang disebut deciduoma.
Fungsi proges teron suli t . dipisahkan dari hormon-hormon lain seperti estrogen. Hal ini disebahkan ォ。イセョ。@ kenyataan bahwa prog.esteron seeara normal beker ja sarna dengan
estro-ァ・セウ・イエ。@ steroid-steroid lainnya dan menghasilkan hanya
Pengaruh progesteron dalam implantasi yaitu merangsang pe£ tumbuhan sistim kelenjar pada endometrium uterus yang telah disensitifkan terlebih dahulu oleh estrogen. Endometrium sekretoris ini ditandai oleh penebalan mukosa, pertambahan
jumlah liku-liku kelenjar, oedema pada stroma, dan adanya butiran-butiran glikogen di dalam sel-sel kelenjar yang pentingEntuk makan ovum (Achjadi, 1979; Toelihere, 1981 ).
Menurut Psycoyos (1973), pada hewan yang
diovariekto-mi lebih awa1 dan den.gan per1akuan progesteron menunjukkan keterlambatan dalam implantasi sampai estrogen disuntikan. Sekresi progesteron pen ting un tuk pemeliharaal!l luteal, dimana sekresi ini memerlukan ran.gsangan hipotha1amus. yang diteruskrul pada Pituitari ( ke1enjar hipofise ) agar ュ・ョウ・セ@
r.esi LTH yang akan merangsang sekresi progesteron. Oleh karena i tu hipofisektomi selama pada hari pertama masa ke-buntingan menyehabkan tertundanya implantasi. Akan tetapi kejadian ini dapa t dicegah bila diberikan progesteron dan
estrogen \ Psychoyos, 1973 )., d. Estrogen
yang penting untuk imp1antasi pada hari ke-6.
Uterus pada tikus dan men·ci t yang diovariektomi disen-si tifkan dengan per1akuan estrogen.. Pertama disuntikan
de-ngan estrogen kemudian dede-ngan ーイッァ・ウエ・イッョセ@ dan terakhir de-ngan. progesteron dikomhinasi dengan ・ウエイッァ・ョセ@ dari perlaku-an ini terbentuk suatu deciduoma ( w・セエャ。オヲL@ 197£ ).
fin. dan Martin (1970) yang dikutip Weitlauf (1978), juga me1akukan pengama tan dengan pretrea tmen; estrogen, dan disimpu1kan laahwa estrogen penting di dalam perangsangan uterus untuk imp1antasi. Macdonald dkk (1967), menyatakan
juga bahwa implantasi b1astosis pada tikus diawali oleh lr.:erja estrogen.
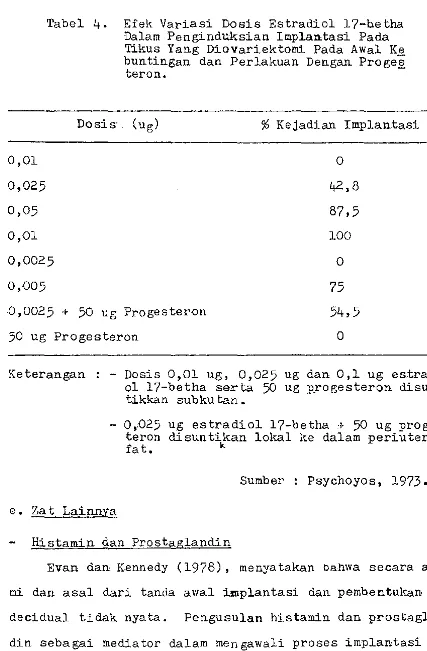
Cani v·ec dan LaffaTague dalam Psychoyos (1973), menemu-kan hahwa implantasi tertunda pada hewan yang diovariekto-mi pada hari ke-4 masa kebun tingan bila hanya diherikan 10 mg pro.gesteron setiap hari.Menurut mereka implantasi ha-nya dapat terjadi hila estradiol ditamhahkan pada perlaku-an dengperlaku-an progesteron itu. Variasi dosis estradiol yperlaku-ang
0,01 0,025 D,05
O,Dl
Tabel
4.
Efek Variasi Dosis Estradiol 17-hetha Dalarn Pengind1.±sian Implantasi Pada Tikus Yang Diovariektorni Pada Awal Ke buntingan dan Perlakuan Dengan pイッァ・セ@teron.
Dosis. (ug)
%
Kejadian Implantasi0
42,8
87,5 100
D,0025 0
0,D05 75
.0,002 5 + 50 ug Progesteron 54,5
50 ug Progesteron 0
Keterangan : - Dosis 0,01 ug, 0,025 ug dan 0,1 ug estradi-ol 17-betha serta 50 ug p.rogesteron disun-tikkan subku tan.
- 0,:025
taron fat.
e.Zat Lainnya
ug estradiol 17-betha :1- 50 ug
proges-disuntikan lokal ke dalam periuterin
l<.
Sumber Psychoyos, 1973.
Histamin dan Prostaglandin
[image:44.600.57.484.86.737.2]ma-sih merupakan bahan pertentangan dian tara beberapa
pene·-l i ti.
Shelennyak (1952) yang dikutip Wei tlauf (1978), yang
pertama beranggapan bahwa histamin mungkin meningkatkan ーセ@
ngaturan dalam proses nidasi. l1enurut we.itlauf (1978), be
herapa tahun kemudian timbul suatu hipothesa yang
menyata-kan bahwa histamin dilepasmenyata-kan dari sel mast uterus oleh ーセ@
ngaruh banjirnya estrogen, merangsang progestational
endo-metrium dan menimbulkan reaksi decidual. Hopothesa ini
berdasar dari kejadian
Anatagonis histamin diinstilasi ke dalam uterus mencegah
respon -decidual dan menguI'altngi jumlah implantasj. bun ... · _
ling palsu pacta ti.kus.
Histamin distintikkE:.rl intra luminal dan histamin atau ーセ@
lepas histamin. diberikan sistemik menginduksi r.eal(si de
cidual pada tikus bunting palsu.
Kandungp.n histamin dari uterus tikus berkurang pada saa t
implantasi dan herkurang pada. ovariektomi dengan
penyun-tikkan estrogen.
De Feo (1967) dalam Weitlauf (1978), menentang
hipo-thesa shelesnyak dengan alasan-alasan sehagai herikut :
Perangsang intra luminal dari respon decidual mungkin
tidak spesifik.
Anti histamin tidak efektif dalam menghamba.t reaksi
de-cidual.
tidak menirnbulkan respon decidual yang besar.
Pengosongan histarnin dari sel mast diuterus dengan 48;130 tidak mengharnbat reaksi decidual.
Histarnin dihas:i.lkan melalui kegiatan enzirn hestidine decarboxylase (HDC). Peningkatan kegiatan HDC dalam hari ke-6 blastosis, rnungkin dihubungkan pada suatu kepentingaa untuk peningkatan jurnlah histamin: untuk imJ'antasi. Usulan ini didukung oleh pengaraatan bahwa bi.la
6.
,mg DL--alphii-methyl histidin dihydrochloride ( DL-alpha-MH ), suatu pengham-bat spesifik dari HDC disuntikkan intra luminal pada hari ke-5 masa kebuntingan, laju implantasi menurun, yai tu hanya 36,0±
12,1%,
sedang laju implantasi normal 96,4±
3.6%
( Taylor dkk, 1973 dalam Dey dkk, 1979 ). Jadidapat disim pulkan bahwa implantasi dapa t dirintangi den.gan mengganggu k.egiatan enzim HDC dari ernbrio.
Ferrando dan Nalbandov (1968), menyatakan bahwa dengan menggunakan tehnik pendingin lokal dari en.dometrium dan de-ngan rnenghilangkan sel mast uterus tikus, ternyata perlaku-an ini mencegah terjadinya implperlaku-antasi dperlaku-an reaksidecidual pada daerah tersebut. Tetapi kemudian instilasi histamin intra luminal memulihkan respon decidual pacta daerah yang kekurangan r.istarnin i tu, dan terbukti me·nyebabkan
implan-tasi.
Peneli tian Dey dkk (1978), dengan menggunakan decidium cromoglycate ( suatu penghambat pelepas histamin )
Dengan perlakuan disodium cromoglycate terbukti terjqdi pe.-ngurangan kejadian implantasi pada beb.erapa blastosis
he-wan ferret.
Kemudian Dey dkk (1979), menemukan pula bahwa terjadi hambatan implantasi pada kelinci dengan perlakuan pengham-bat pelepas histamin.
Blastosic kelinci menganrlung PGE, PGF dan PGS, yang selanjutnya mungkin dikeluarkan dari blastosis untuk ningkatkan permeabilitas vaskular uterus atau mungkin me-mainkan peranan di dalam streroidogenesis blastosis ( Dyk-man dan SpilDyk-man, 1975 dalam Fimdaly, 1982 ). Selanjutnya
streroid yang dihasilkan blastosis mungkin mengawali implaQ tasi dengan dukungan prostaglandin lokal, sebab estrogen diketahui pula dapat merangsang produksi PGF ( Saksene dan Harper, 1972 dalam Findaly, 1982 ).
Sharma (1979) yang dikutip Hoos dan Hoffman (1982), menemukan adanya konsentrasi yang lebih tinggi dari PGE2 dalam daerah implantasi kelinci dari pad& dalam daerah nOQ implantasi dari uterus pada hari ke-6 dan ke-7 masa
kebun-tingan.
Produksi CO2 EmQxiQ
Lutwak dkk (1954) dalam Wei tlauf (1979), menemukan ada nya peruhahan dramastis konsentrasi bikarbonat yang diamati pada blastosis kelinci. Hereka mengusulkan kemungkinan セ・{@
libatnya coセ@ yang.diproduksi blastosis dalam proses
implan-t:.
(1978), menduga bahwa CO2 dirindahkan darl embrio kelinci
sebagai ion bikarbonat dan dirubah jadi asam karbon dan セ@
ram basa karbona t dalam epi thel uterus. A sam karbon dengan
bantuan enzim karbonik anhidrase kemudian dlrubah menjadi
'C02 dan air,C02 dipindahkan oleh sirkulasi sistem induk. Akibatnya terjadi peningkatan pH lokal, karena reaksi ini
meningkatkan sifat basa, kemudian terjadi perubahan pada
permukaan s81ubung embrlo yang menyebabkan embrio mudah me
lekat sendiri pada dinding uterus.
Tidak ada bukti langsung untuk membuktikan hipothesa
i tu, tetapi beb.erapa pengamatan telah diajukan untuk mend),!.
kung hipothesa tersebut, yaitu :
Kutub abembrionik dari b1astosis menjadi basa dalam
la-rutarunonbuffer ( pH mendekati 9 ). Trofoblas dan
sela-セ@
put noncelluler pacta blastosis menjadi 1engket, dan
si-fat lengket ini dapatdicegah dengan fikasasi
pengasam-an. Dari kenyataan ini diduga bahwa adisi yang
diiduk-si oleh basa mungkin merupakan mekanisme normal un tuk
awa1 per1ekatan ( Boving, 1963 da1am weitlauf, 1978 ).
Kegiatan enzim karbonik anhidrase meningkat dalam epitel
uterus kelinci pada kira-kira saat implantasi dan
mung-kin berguna memberikan fasilitas perpindahan asam
kar-bon sebagai CO2 •
We.itlauf {l978) mengutip pendapat Hetherington (1968),
menyatakan bahwa gas CO2 da1am lumen uterus menginduksi
Usulan C02 embrio terlibat dalam implantasi belum di-terima secara luas, dan timbul pertanyaan apakah CO2 meru-pakan isyara t dari embrio terhadap uterus pada saa t implag
Diantara beberapa peneliti masih terdapat adanya per-bedaan pendapat mengenai hormon-hormon yang terlibat dalam implantasi, dan belum diketemukan inforrnasi yang menerang-kan secara terperinci tentang memenerang-kanisme kerja hormon yang berperanan da1am implantasi. Seperti keterlibatan LTH, hi£
tamin dan prostaglandin, . beberapa peneli ti rnasih meragukan keterliba tannnya dalam implantasi.
Rangsangan LH dan LTH terhadap CL menghasilkan sekre-si estrogen dan progesteron. Progesteron bekerja sarna de-ngan estrogen menginduksi persiapan uterus untuk proses ゥセ@
plantasi.
Omalley dan Strott (1973), mencoba menerangkan bagai mana progesteron mentransformasi endometrium pada tahap ーセ@
nerimaan b1astosis baru menduga bahwa progesteron mungkin menekan sintesa agen pengbambat implantasi, dan progeste-ron mungkin menginduksi sin.tesa sua tu moleku1 protein yang menyediakan atau yang memheri fasi1itas untuk imp1antasi.
Di. bawah pengaruhestrogen endometrium mengalami pro-liferasi dan menjadi tebal. Tetapi kemudian progesteron menghambat ーイッャゥヲ・イセゥ@ tersehut, dan endometrium menjadi ak-tif bersekresi. Selaput kelenjar endometrium menjadi tak beraturan, kandungan glikogen meningkat dan stromanya
men-jadi oedematus, seterusnya endometrium siap menerima, meng-ikat dan memelihara blastosis ( Omalley dan Strott, 1973).
bera t ini disebabkan adanya pertumbuhan jaringan, teru tama oleh adanya proses hipertropi dan hiperplasia dari endomet-rium dan miometendomet-rium. Pada implantasi estrogen sebagai
pen-sensi tif uterus dari rangsangan progesteron dan berfungsi merangsang kontraksi uterus yang diperlukan untuk pergeser-an atau penggpergeser-antipergeser-an tempat embrio ( Partodihardjo, 1978 ).
Estrogen merangsang kontraksi uterus dengan penambahan am-plitudo dan derajat kontraksi ( Achjadi, 1979 ).
Henuru t Prasad dkk (1968), estrogen menginduksi peru-bahan dalam blastosis dan uterus untuk persiapan implanta-si dengan meningka tkan implanta-sintesa RNA, DNA dan protein di da-lam blastosi, serta pengaktifan sintesa RNA dan protein di dalam uterus.
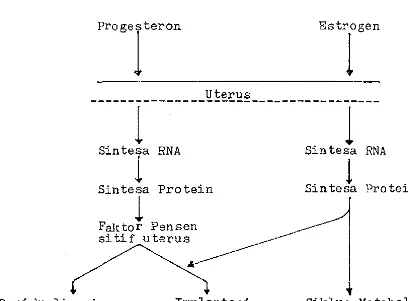
Sistim kerja progesteron dan estrogen dalam proses persiapan implantasi di terangkan pula oleh Omalley dan Strott (1973), progesteron dan estrogen merangsang uterus aktif dalam mensintesa RNA, kemudian mensintesa protein dan
faktor pensensi tif uterus ( diduga atau enzim ) yang aktif dalam merangsang decidualisasi dan ゥセーャ。ョエ。ウゥN@
Gambar
5.
Pengaruh Progesteron dan Estrogen Dalam Merangsang Decidualisasi daniューャ。セエ。ウゥ@ Pada Tikus.
MMMMMMMMMMMMMMMセセセセセᄃMMMMMMMMMMMMMMMMM
1
Sintesa RNA
1
Sintesa Protein
1
Faktor Pensen si ti f uterus
1
Sin tesa
RNA
sinte}a Protein
Decidualisa si Implantasi Siklus Metabolis-me
Sumber Omalley dan Strott, 1973.
Mekanisme kerja histamin dan prostaglandin dalam pro-ses implantasi belun diketahui, namun diduga bahwa kedua media tor i tu meningka tkan permeabili tas vaskular endometri urn untuk kepentingan implantasi ( Evans dan Kennedy, 1978; Hoos dan Hoffman, 1983 ). Brandon dan Roval (1979) yang diku tip oleh Johnson dan Dey (1980), menambahkan bahwa hi.2
bonat yang dipindahkan dari blastosis, kemudian ion
bikar-bonat dengan bantuan enzim karbonik anhidrase dirubah
men-jadi asam karbon dan garam basa karbonat. Asam karbon te£
urai idenjadi CO2 dan air, CO2 dipindahkan oleh si stim
sir-kulasi iruduk. Perubahan ini menyebabkan penil.gka tan suas§l:.
na basa yang mempengaruhi permukaan selubung embrio
bersi-fat lengket. Hal ini memudahkan embrio untuk melekat pada
Ovum tiba diarnpula dalam waktu yang cukup lama, sesu-dak tibanya sperrna untuk menjarnin terlaksananya kapasitasi. Urnur ovum umumlllfa kurang dari 24 jam, demikian pula umur
sperrna. Oleh karena i tu waktu terbaik untuk illseminasi harus benar-benar diperhatikan agar fertilisasi berhasil dan dapat terjadi kebuntingan.
Sebelum implantasi, embrio memperoleh makanan dari sekresi kelenjar-kelenjar uterus, sedang sesudah implanta-si embrio memperoleh makanan dari saluran darah induk.
Embrio diduga mensintesa steroid dan hormon-hormon protein, estrogen yang diproduksi oleh blastosis terlibat dalam pemberian tanda untuk pemeliharaan luteal.
Proses preimpIan.tasi pada uterus ter jadi p!.'!ningka tan vaskularisasi dan peningkata'n serta kegiatan pertumbuhan kel enjar uterus.
Implan tasi perlu sua tu koordinasi in teraksi antara
eJJl-brio dan kondi si uterus yang seharusnya, dan un tuk se tiap ternak marnalia kejadiannya hervariasi. Pada umumnya implag tasi herjalan secara hertahap mulai dari tahap persentuhan embrio dengan endometrium, teriepasn,ya zona pelusida, geseran atau pemhagian tempa t dan yang terakhir adalah per-tautan antara trofoblas dengan epi tel en,dometrium.
LTH sarna dengan LH yang juga berperan tidak lang sung dalam proses ゥューャ。ョエ。ウゥセ@ yaitu dengan merangsang corpus ャセ@
teum untuk mensekresikan progesteron yang akan menginduksi terjadinya implantasi.
Dalam implantasi progesteron merangsang pert;umbuhan sistem kelenjar pada endometrium uterus yang disensitifkan terlebih dahulu oleh estrogen. Progesteron menghambat kog traksi miDmetrium, ウ・ィゥョァァセ@ ketenangan meometrium ini men jamin pemukiman blastosi.s dalam uterus. Progesteron mung-kin juga dapat menginduksi sintesa suatu molekul protein yang menyediakan.atau memberi fasilitas untuk implantasi.
Estrogen berfungsi オセエオォ@ mensensitifkan uterus dari rangsangan progesteron dan berfungsi merangsang kontraksi uterus yang diperlukan untuk pergeseran atau pembagian tern pat dari embrio pada tahap implantasi.
Kerja sarna an tara estrogen dan progesteron panting un-tuk implantasi, membuat kondisi pemeliharaan unt;uk hlasto-sis agar dapat hidup.marangsang uterus aktif dalam
mensin-te sa RNA, kemudian mensin mensin-te sa pro mensin-tein dan rak tor per sen sitif uterus ( diduga suatu enzim ) yang aktif dalam merang ·sang decidualisasi dan implantasi.
Hista-min dan prostaglandin diduga sehagai media tor yang dapa t meningka tkan permeabili tas vaskular endometrium.
Austin, C. 1975. Hembrane Fuslon Events ln Fertillzation
J.
Reprod. Fert., 44 l 155 - 166.Achyadl. K, 1979. Diktat Pengantar Kuliah Hormon-hormon Reproduksi. FKH - IPB. Bogor. (Tidak
Dipublikasi-lean) •
Bindon, B.M. and D.R. Lamond. 1969. to my on Implantation in The Rat. 18 : 43 - 50.
Effect of Hypophysec J. Reprod.
Fert.,-Balinsky, B.I. 1970. An Intoduction to Embryology. J. Raprod. Fert.
W.B.
Saunders Co. pィゥャ。、・ャーセゥ。N@Cole, H.H. and P.T. Cupps. 1968. r・ーイッ、オNセエゥッョ@ in Domestic Animals. Second Edi tion. Academia Press. New York and
London.-Cook, Band R.H. Hunter. 1978. System1c and local Hormonal Requirements For Implantation. J. Reprod Fert., 54 : 471 - 482.
Dey, S.K., D.C. Production in Rabbit.
Johson and J .. G. Santos. 1979. Is Histamin by The Blastocyst Required For Implantation Biology of Reproduction., 21 : 1169 - 1173. Evans, C.A. and T.G. Kennedy. 1978. The Importance of ーイッセ@
taglandin Synthesis for The Initiation of Blastocyst Implantation in The Hamster. J. Reprod. Fert.,
54 : 255 - 261.
Ferrando, G. and A.V. Nalbandov. 1968. Relative Importance of Histamin and Eatrogen on Implantation in Rat •.
En--docrinology., 83 : 933 - 937.
Findlay, J.K. 1980. Blastosyst - Endometrial Interactions in Early Pregnancy in-The Sheep. J. Reprod. Fert.
sオーーャセN@ 30 セ@ L7.1 - 182.
Hafez, E.S.E. 1<)68. Reproduction in Farm Animals. ZndEd • Lea and Febiger. Phi delphia •
1974. Reproduction in Farm Animals. 4 thEd • Lea and Febiger. Phi delphia.
Hoos, P.C. and L.H. Hofflllan. 1983. Effecct of Histamine Receptor Antagonists and Indomethacin on I:nplanta tion ill The Rabbit. J. Biology of Reproduction.)
29 : 833 - 840.
Implantation: Dexamethasone ced Implantation in The Rat •.
22 : 1136 - 1141.
Inhibits Estradiol Indu-Biology of Reproduction.
Ma.cdonald, G.J.,.T.A. David and O.R. Roy.
1967.
Initia-tion of Blastocyst ImplantaInitia-tion by Luteiniz.:!:ng Hor,... mone. Endo crinolo gy., 80 :172 - 177.
mオイーィケセ@ B.D.
1.979.
The Role. of Prolactinand Luteal 11aintenance i.n The Ferret. roduction.,
21 : 517 - 521.
in Implan ta tion Biology of
Rep-Raj, H.G.M., M.R. Sairam and N.R. Moudgal.
JL968.
Invol-vement of Luteinizing Hormone in The Implantation Pro-cess of The Ra t . J . Raprod. Fer-t •. ,12 :
335 -
341..
Nalbandov, A.V.1958.
Reproductive Physi.oJ!.ogy. W.H.Fre-man and -Co., San Fransisco.
O'malley, B.W and C.A. Strott.
1973.
The Mechanism of Ac-tion ofProgesteron. Dalam: Hand Book of Physiology •. Endocrinology, vol. 2.part
1. American Phys:Lologycal Sosiety. Waverly Press, Inc.,. Ba:ltimore and Maryland. Prasad. M.R.N •• C.M.S. Dass and S. Mohla.1968.
Action ofOestrogen.on The Blastocyst and uterus in. Delayed Im-plantation-An Autoradio:graphilc Study. J. Reprod. Fert ••
16 : 97 - 104.
Psychoyos, A.
1973.
Endocrine Control of Egg Implantation. Dalam : Hand Book of Physiology. Endocrinology vol. II, Part 1. American pィケウゥッャッァケセ。ャ@ Sosiety.. Waverly Press Inc., Baltimore, Maryland.Partodihardjo, S.
1980.
Ilau Reprodksi Hewan. Mutliara. Jakarta.Shumaway, wセjN@ and F.D. Adamstone •.
1942.
Introduction to Vertebrate Emhriology. Jhon Willey and Son, Inc •• New York.Salysbury, G. W. and N. I. Van Denmark.
1961.
l?:lysiolo gy of Reproduction and Artifi:::ial Insemination. W.H. Freeman and Co. San Fransisco and London, pp.639.
Sukra, Y.1981.
Diktat Pengantar Kuliah Embriologi I.FKH - IPB, Bogor ( Tidak dゥーオ「Qゥォ。ウゥセ。ョ@ ).
tion and Fertili ty. Harper and Row PUb,lisher. Cam-bridge, New York, London, pィゥャ。、・ャーゥィゥ。セ@ Mexico cゥエケセ@
KONTROL HORMONAL PADA IMPLANTASI
S K R I P S I
oleh
EDI WIRYANA AHMAD
B 17.1432
FAKULTAS KEDOKTERAN HEWAN
INSTITUT PERTANIAN BOGOR
1 9 S 5
Edi VI. Ahmad. Kontrol Hormonal Pada Implantasi ( Di-bawah bimbingan
R.
Kurnia Achjadi ).Penulisan ini bertujuan untuk mengetahui dan mempela-jari peristiwa hormonal yang terjadi dalam proses implan-tasi, selain i tu, juga mempelajari peristiwa fertilisasi, perkembangan embrio sebelum mencapai bentukan fetus dan
jalannya implantasi.
Terjaninya implantasi diinduksi oleh kerja dari hor-