EFEKTIVITAS CENDAWAN MIKORIZA ARBUSKULA (CMA)
TERHADAP PERTUMBUHAN DAN PRODUKTIVITAS
Cynodon dactylon (L.) Pers YANG DIBERI
LEVEL SALINITAS BERBEDA
SKRIPSI
EFEKTIVITAS CENDAWAN MIKORIZA ARBUSKULA (CMA)
TERHADAP PERTUMBUHAN DAN PRODUKTIVITAS
Cynodon dactylon (L.) Pers YANG DIBERI
LEVEL SALINITAS BERBEDA
SKRIPSI
TANTI PATRIYASARI
PROGRAM STUDI NUTRISI DAN MAKANAN TERNAK FAKULTAS PETERNAKAN
EFEKTIVITAS CENDAWAN MIKORIZA ARBUSKULA (CMA)
TERHADAP PERTUMBUHAN DAN PRODUKTIVITAS
Cynodon dactylon (L.) Pers
YANG DIBERI
LEVEL SALINITAS BERBEDA
TANTI PATRIYASARI D24102070
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
EFEKTIVITAS CENDAWAN MIKORIZA ARBUSKULA (CMA)
TERHADAP PERTUMBUHAN DAN PRODUKTIVITAS
Cynodon dactylon (L.) Pers
YANG DIBERI
LEVEL SALINITAS BERBEDA
Oleh
TANTI PATRIYASARI D24102070
Skripsi ini telah disetujui dan disidangkan di hadapan Komisi Ujian Lisan pada Tanggal 5 Juni 2006
Pembimbing Utama
Dr. Ir. Panca Dewi M.H.K.S, MSi. NIP. 131 672 157
Pembimbing Anggota
Dr. Ir. Luki Abdullah, MSc.Agr. NIP. 131 955 531
Dekan Fakultas Peternakan Institut Pertanian Bogor
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 20 September 1984. Penulis adalah putri pertama dari dua bersaudara dari pasangan Bapak Fridel Dawar dan Ibu Ratnamaida.
Pendidikan dasar diselesaikan pada tahun 1996 di SDN Mekarjaya XVIII Depok Timur, pendidikan lanjutan menengah pertama diselesaikan pada tahun 1999 di SMPN 3 Depok dan pendidikan lanjutan menengah atas diselesaikan pada tahun 2002 di SMAN 38 Jakarta.
Penulis diterima sebagai mahasiswa pada jurusan Ilmu Nutrisi dan Makanan Ternak, Fakultas Peternakan, Institut Pertanian Bogor melalui jalur Seleksi Penerimaan Mahasiswa Baru (SPMB) pada tahun 2002.
KATA PENGANTAR
Alhamdulillahirobbil’alamin. Puji syukur kehadirat Allah SWT yang telah memberikan rahmat dan karuniaNya, sehingga penulis dapat menyelesaikan skripsi dengan judul ”Efektivitas Cendawan Mikoriza Arbuskula Terhadap Pertumbuhan dan Produktivitas Cynodon dactylon (L.) Pers yang diberi Level Salinitas Berbeda”. Skripsi ini ditulis berdasarkan hasil penelitian yang penulis lakukan mulai bulan Maret sampai dengan Juli 2005. Penanaman, pemeliharaan dan pemanenan dilakukan di rumah kaca Laboratorium Lapang Agrostologi, analisa infeksi akar di Laboratorium Agrostologi Fakultas Peternakan, analisa jumlah spora di Laboratorium Bioteknologi Kehutanan dan Lingkungan, Pusat Penelitian Sumber Daya Hayati dan Bioteknologi, IPB untuk analisa jumlah spora.
Skripsi ini merupakan karya tulis yang berisi tentang upaya untuk memanfaatkan daerah pantai yang memiliki tingkat salinitas tinggi. Tingginya tingkat salinitas menyebabkan tanaman kurang dapat tumbuh dengan baik karena ketersediaan air yang rendah akibat terikat oleh senyawa-senyawa garam. Untuk mengatasi hal tersebut dapat dilakukan beberapa upaya, diantaranya adalah pemilihan jenis tanaman yang toleran terhadap kondisi salinitas dan penggunaan pupuk hayati cendawan mikoriza arbuskula (CMA). Pengetahuan tentang adanya asosiasi antara cendawan mikoriza arbuskula (CMA) dengan vegetasi pantai memberikan harapan kemungkinan pemanfaatannya pada kegiatan penghijauan didaerah pantai dimasa yang akan datang.
Akhir kata semoga skripsi ini bermanfaat bagi yang membacanya.
Bogor, Juni 2006
DAFTAR ISI
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... ix
Peran Mikoriza dan Sistem Serapan P... 5
Infeksi Akar dan Spora Cendawan Mikoriza Arbuskula (CMA)... 7
Salinitas ... 8
Cendawan Mikoriza Arbuskula dan Tingkat Salinitas...9
Cynodon dactylon (L.) Pers (Rumput Bermuda)... 9
Media Tumbuh Cynodon dactylon (L.) Pers... 10
Keadaan Umum Percobaan... 17
Jumlah Stolon... 18
Jumlah Individu... 21
Produksi Berat Kering Tajuk dan Akar... 22
Berat Kering Tajuk... ... 23
Berat Kering Akar... 26
KESIMPULAN DAN SARAN ... 32
Kesimpulan ... 32
Saran ... 32
UCAPAN TERIMAKASIH ... 33
DAFTAR PUSTAKA ... 34
DAFTAR TABEL
Nomor Halaman
1. Rekapitulasi Sidik Ragam Penelitian ... 18
2. Rataan Jumlah Stolon Panen 1, 2 dan 3 Cynodon dactylon (buah) ... 19
3. Rataan Jumlah Individu Cynodon dactylon (buah) ... 21
4. Rataan Berat Kering Tajuk Panen 1, 2 dan 3 Cynodon dactylon (gram) ... ... 23
5. Rataan Berat Kering Akar Cynodon dactylon (gram)... ... 26
6. Rataan Persentase Infeksi Akar . ... ... 28
DAFTAR GAMBAR
Nomor Halaman
1. Penampang Longitudinal Akar yang Terinfeksi CMA ... 5
2. Skema Dugaan Penghambatan Pembentukan Mikoriza... 7
3. Hifa, Arbuskula dan Vesikel didalam Jaringan Korteks ... 8
4. Cynodon dactylon (L.) Pers Varietas Tifgreen ... 10
5. Cynodon dactylon Saat Penelitian ... 17
6. Perakaran yang Terinfeksi CMA ... 28
7. Perakaran yang Tidak Terinfeksi CMA ... 28
DAFTAR LAMPIRAN
Nomor Halaman
1. Lampiran 1. ANOVA Jumlah Stolon Panen 1 ... 38
2. Lampiran 2. ANOVA Jumlah Stolon Panen 2 ... 38
3. Lampiran 3. ANOVA Jumlah Stolon Panen 3 ... 38
4. Lampiran 4. ANOVA Jumlah Iindividu ... 39
5. Lampiran 5. ANOVA Produksi Berat Kering Tajuk Panen 1 ... 39
6. Lampiran 6. ANOVA Produksi Berat Kering Tajuk Panen 2 ... 39
7. Lampiran 7. ANOVA Produksi Berat Kering Tajuk Panen 3 ... 40
8. Lampiran 8. ANOVA Produksi Berat Kering Akar ... 40
9. Lampiran 9. ANOVA Persentase Infeksi Akar ... 40
10. Lampiran 10. ANOVA Jumlah Spora... 41
11. Lampiran 11. Komposisi Pupuk Hyponex ... 41
RINGKASAN
TANTI PATRIYASARI. D24102070. 2006. Efektivitas Cendawan Mikoriza Arbuskula Terhadap Pertumbuhan dan Produktivitas Cynodon dactylon (L.) Pers yang diberi Level Salinitas Berbeda. Skripsi. Program Studi Nutrisi dan Makanan Ternak, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Dr. Ir. Panca Dewi MHKS, MSi. Pembimbing Anggota : Dr. Ir. Luki Abdullah, MSc.Agr.
Daerah pantai dan kawasan kering umumnya memiliki tingkat salinitas yang relatif tinggi sehingga tanaman kurang dapat tumbuh dengan baik. Upaya yang dapat dilakukan untuk memanfaatkan lahan berkadar garam tinggi adalah dengan mencari jenis tanaman yang toleran terhadap kondisi salinitas, perbaikan lahan pra tanam dan dengan penggunaan pupuk hayati cendawan mikoriza arbuskula (CMA). Cynodon dactylon (L.) Pers (rumput Bermuda) memiliki berbagai macam kegunaan, diantaranya sebagai pada padang penggembalaan permanen, rumput taman dan rumput pada lapangan golf. Mikoriza merupakan suatu bentuk simbiosis mutualiasme antara cendawan dengan perakaran tumbuhan tingkat tinggi. Cendawan mikoriza arbuskula (CMA) dapat memberikan berbagai keuntungan kepada tanaman inang, antara lain meningkatkan penyerapan unsur hara, meningkatkan ketahanan terhadap kekeringan, salinitas serta dapat memproduksi hormon dan zat pengatur tumbuh.
Penelitian ini bertujuan untuk menguji ketahanan Cynodon dactylon (L.) Pers
yang diberi level salinitas yang berbeda serta untuk mengetahui pengaruh pemberian CMA terhadap pertumbuhan dan produktivitas Cynodon dactylon (L.) Pers yang diberi level salinitas berbeda. Penelitian didesain menggunakan rancangan perlakuan pola faktorial dengan dua faktor yaitu cendawan mikoriza arbuskula (CMA) dan level salinitas yang disusun dalam rancangan acak kelompok. Faktor CMA terdiri atas dua taraf yaitu tanpa pemberian CMA (M0) dan dengan pemberian CMA (M1). Faktor level salinitas terdiri atas lima taraf yaitu tanpa salinitas (S0), salinitas 2.500 ppm, salinitas 5.000 ppm, salinitas 7.500 ppm dan salinitas 10.000 ppm. Penelitian dilakukan di rumah kaca Laboratorium Lapang Agrostologi, Fakultas Peternakan IPB untuk masa penanaman, pemeliharaan dan pemanenan (Maret-Juli 2005), serta di Laboratorium Agrostologi Fakultas Peternakan IPB dan di Laboratorium Bioteknologi kehutanan dan lingkungan, Pusat Penelitian Sumber Daya Hayati dan Bioteknologi, IPB untuk analisa persentase infeksi akar dan jumlah spora.
Level salinitas sangat nyata (p<0,01) mempengaruhi jumlah stolon panen ke-2, produksi berat kering tajuk panen ke-3 dan produksi berat kering akar, serta nyata (p<0,05) mempengaruhi jumlah stolon panen ke-3 dan produksi berat kering tajuk panen ke-2, sedangkan pemberian CMA sangat nyata (p<0,01) meningkatkan jumlah stolon panen ke-1dan ke-3, jumlah individu, produksi berat kering akar dan tajuk panen ke-3, persentase infeksi akar dan jumlah spora. Semua peubah tidak memberikan respon yang nyata terhadap interaksi antara level salinitas dengan pemberian CMA.
ABSTRACT
Effectivity of Arbuskula Mycorrhi zal Fungi (CMA ) on Cynodon dactylon (L.) Pers Growth and Production
by Salinity level Differences
T. Patriyasari., P .D.M.H. K arti and L. Abdullah
The present work was undertaken to investigate the tolerance of Cynodon dactylon (L.) Pers grass in salinity condition. This experiment was also conducted to study the effectivity of arbuskula mycorrhizal fungi (CMA) on Cynodon dactylon (L.) Pers growth by salinity level differences. This experiment used Factorial Complete Randomized Block Design consisted of 2 factors by 3 replications. The first factor was CMA (with CMA and without CMA). The second factor was level of salininity (0, 2.500, 5.000, 7.500 and 10.000 ppm). Statistical analysis was conducted using ANOVA; when the result indicated a significant treatment effect (p<0.01), the Duncan Multiple Range Test was used. The result showed that addition of CMA produced significant effects (p<0.01) on stolon number period I and III, plant individual number, production dry weight of root-shoot period III, root infection percentage and amount of spora formed. Level of salinity had significant effects (p<0.01) on stolon number period II, production dry weight of root-shoot period III. There was no interaction between CMA and the level of salinity. It is concluded that addition of arbuskula mycorrhizal fungi (CMA) can stimulate
Cynodon dactylon (L.) Pers growth on salinity level differences condition.
PENDAHULUAN
Latar Belakang
Indonesia merupakan negara kepulauan dengan garis pantai terpanjang di dunia sehingga menjadikan Indonesia memiliki area pantai yang luas. Areal yang luas ini sangat potensial untuk dimanfaatkan dalam kegiatan pertanian. Namun, pada daerah pantai dan kawasan kering umumnya memiliki tingkat salinitas yang tinggi sehingga tanaman tidak dapat tumbuh dengan baik karena ketersediaan air yang relatif rendah akibat terikat oleh senyawa-senyawa garam. Kelebihan kadar garam dalam tanah akan meracuni tanaman dan merupakan stres lingkungan bagi tanaman, karena dapat menghambat perkecambahan benih, perkembangan akar, tinggi tanaman, kualitas serta kuantitas produksi, daya serap tanaman terhadap unsur hara, menghambat pertukaran gas dan menurunkan potensial total air serta potensial osmotik yang akhirnya dapat merusak jaringan tanaman (Bintoro, 1984). Faktor inilah yang menjadi salah satu kendala untuk mengembangkan pertanian di kawasan pantai, karena sangat sedikit jenis tanaman yang mampu hidup di tempat dengan salinitas tinggi.
Menurut Setiadi (1989), beberapa upaya yang dapat dilakukan untuk memanfaatkan lahan berkadar garam tinggi adalah dengan: perbaikan lahan pra tanam, penggunaan pupuk hayati cendawan mikoriza arbuskula (CMA) dan pemilihan jenis tanaman yang toleran terhadap kondisi salinitas. Perbaikan lahan pra tanam dapat dilakukan melalui kegiatan irigasi, yang bertujuan untuk mengurangi garam yang berlebihan sehingga sesuai dengan kebutuhan tanaman. Namun, metode ini tidak efektif dan praktis karena membutuhkan biaya yang besar (Tan, 1991).
Pemanfaatan cendawan mikoriza arbuskula (CMA) merupakan alternatif lain dalam menanggulangi masalah rendahnya produktivitas tanaman pada tanah salin. Cendawan ini mendapat perhatian dari para ahli pertanian, karena merupakan salah satu alternatif teknologi dimasa mendatang yang tidak saja efektif, tetapi juga murah dan ramah lingkungan, untuk membantu meningkatkan produktivitas dan kualitas tanaman terutama yang ditanam pada lahan-lahan marjinal yang kurang subur.
sebagai pakan ternak serta padang penggembalaan permanen karena sangat tahan terhadap injakan dan penggembalaan berat, selain itu, rumput Bermuda sering digunakan untuk keindahan taman dan lahan untuk berolahraga, khususnya olahraga golf (Gohl, 1981). Untuk lapangan golf di Indonesia ada beberapa varietas yang sering digunakan, seperti varietas Tifway, Tifgreen dan Tifdwarf.
Beard (1973) menyebutkan bahwa Cynodon dapat digunakan sebagai spesies utama pembentuk lapangan rumput di daerah beriklim tropis karena memiliki banyak keunggulan, antara lain mempunyai tekstur daun yang halus, kepadatan pucuk yang tinggi, dapat beradaptasi dengan baik di daerah beriklim panas, berakar dalam, toleran terhadap kekeringan dan pemangkasan pendek (3.5-4 mm) yang merangsang pertumbuhan serta cukup responsif terhadap pemupukan. Rumput ini dapat tumbuh pada kondisi tanah bersalinitas tinggi karena memiliki stolon, yang berfungsi untuk mengakumulasikan NaCl sehingga tidak menyebabkan kerusakan jaringan yang berarti pada tanaman.
Perumusan Masalah
Salah satu faktor yang menjadi kendala untuk mengembangkan pertanian di kawasan pantai adalah tingginya tingkat salinitas, sehingga sangat sedikit jenis tanaman yang dapat tumbuh dengan baik. Sehubungan dengan kemampuan rumput
Cynodon untuk hidup pada kondisi salinitas maka dilakukanlah penelitian menggunakan rumput Cynodon dactylon (L.) Pers varietas Tifgreen pada kondisi salinitas yang berbeda dengan menambahkan cendawan mikoriza arbuskula (CMA) untuk memperbaiki pertumbuhan rumput ini. Pengetahuan tentang adanya asosiasi antara cendawan mikoriza arbuskula (CMA) dengan vegetasi pantai memberikan harapan kemungkinan pemanfaatannya pada kegiatan penghijauan didaerah pantai dimasa yang akan datang.
Tujuan
Penelitian ini bertujuan untuk menguji ketahanan Cynodon dactylon (L.) Pers
TINJAUAN PUSTAKA
Mikoriza
Menurut Imas et al., (1989) mikoriza merupakan suatu bentuk hubungan simbiosis mutualistik antara cendawan (mykes) dan perakaran (rhiza) tumbuhan tingkat tinggi. Simbiosis mutualistik yang berlangsung antara mikoriza dengan tanaman inang dimana tanaman inang dapat menyediakan fotosintat untuk mikoriza sebagai sumber energi, sedangkan mikoriza mensuplai mineral-mineral anorganik yang berasal dari tanah untuk tanaman inang.
Setiadi (1989) menjelaskan, beberapa manfaat yang dapat diperoleh tanaman inang dari adanya asosiasi mikoriza antara lain: (1) meningkatkan ketersediaan unsur hara dan air; (2) meningkatkan ketahanan terhadap kekeringan, salinitas dan toksisitas ion logam berat; (3) meningkatkan ketahanan terhadap serangan patogen akar serta (4) mikoriza dapat memproduksi hormon dan zat pengatur tumbuh seperti auksin, sitokinin dan giberilin. Menurut Abbot dan Robson (1984), ada tiga hal yang menyebabkan mikoriza arbuskula dapat meningkatkan penyerapan unsur hara dalam tanah, yaitu karena mikoriza dapat (a) mengurangi jarak bagi hara untuk memasuki akar tanaman; (b) meningkatkan rata-rata penyerapan hara dan konsentrasi hara pada permukaan penyerapan dan (c) merubah secara kimia sifat-sifat hara sehingga memudahkan penyerapannya kedalam akar tanaman.
Berdasarkan struktur dan cara infeksinya terhadap tanaman inang, mikoriza dapat dikelompokkan kedalam tiga golongan besar, yaitu: ektomikoriza, endomikoriza dan ektendomikoriza (Imas et al., 1989). Selanjutnya, Kabirun (1990) memaparkan bahwa endomikoriza dibagi menjadi tiga kelompok, yaitu: (1)
Ericaceous mykoryza, (2) Orchidaceous mikoryza dan (3) Mikoriza Vesikular Arbuskula (MVA).
Arbuskula diduga sebagai pemindah unsur hara diantara simbion-simbion. Arbuskula terbentuk antara 1-3 hari setelah infeksi akar dan mempunyai siklus hidup yang relatif pendek, berkisar 4-6 hari, sedangkan vesikel merupakan struktur yang banyak mengandung lemak dan berfungsi sebagai organ penyimpanan. Umumnya vesikel terbentuk setelah arbuskula terbentuk dan bertambah banyak pada waktu tanaman bertambah dewasa. Jika korteks mengelupas, beberapa vesikel keluar dari jaringan akar dan berada didalam tanah. Vesikel ini akan berkecambah dan bertindak sebagai propagul tidak efektif.
Menurut Sanders (1986), pembentukan interaksi cendawan mikoriza arbukula (CMA) dan tanaman inang adalah sebagai berikut:
1. Pembentukan kecambah dari propagul CMA yang diikuti pembentukan hifa dalam tanah.
2. Infeksi primer yang merupakan proses pembukaan cendawan ke dalam tanaman inang. Saat menembus sel, hifa cendawan mengalami pengerutan diameter, kemudian diameter hifa tersebut kembali pada ukuran yang semula lagi
3. Infeksi sekunder yang merupakan proses penyebaran/penjalaran cendawan ke bagian-bagian akar yang lain.
4. Transfer karbon hasil asimilasi dari tanaman inang ke cendawan.
5. Pertambahan biomassa cendawan yang terdapat dalam korteks akar dan yang terdapat disekitar akar.
6. Penyerapan hara mineral (terutama fosfor) oleh miselium dalam tanah dan ditransfer ke tanaman inang.
7. Meningkatnya pertumbuhan tajuk dan akar yang merupakan hasil perbaikan hara tanaman.
8. Meningkatnya pertumbuhan cendawan akibat meningkatnya persediaan ruang dan substrat tumbuh.
5 Gambar 1. Penampang Longitudinal Akar yang Terinfeksi CMA
(Sumber: Brundrett et al., 1994)
Peran Mikoriza dan Sistem Serapan P
Menurut Hardjowigeno (1992) phospor didalam tanah pada umumya dalam bentuk tidak larut sehingga hanya sebagian kecil yang dapat diserap oleh tanaman. Fosfor yang ada didalam tanah sama banyaknya antara yang organik dan anorganik. Tanah yang kering akan mengurangi pengambilan fosfor dan menyebabkan kekurangan fosfor. Phospor memiliki beberapa peranan bagi tanaman, diantaranya: (1) Berperan dalam metabolisme karbohidrat; (2) Sebagai aktivator; (3) Berperan dalam proses fisiologis; (4) Berperan dalam memberi pengaruh yang menguntungkan untuk pembelahan sel, kematangan tanaman, perkembangan akar halus, kualitas hasil tanaman, pembentukkan bunga, buah dan biji serta (5) Berperan dalam memberi ketahanan tanaman terhadap penyakit.
Tumbuhan yang diinokulasi mikoriza dapat tumbuh dengan baik daripada tumbuhan yang tidak diinokulasi. Hal ini disebabkan oleh kerja hifa eksternal yang mampu meningkatkan penyerapan unsur hara makro dan beberapa unsur hara mikro, selain itu hifa juga mampu mengakumulasi dan mengtranslokasikan logam (Indriani, 2004). Unsur hara yang telah terakumulasi pada hifa eksternal akan ditranslokasikan ke hifa internal oleh suatu sistem transport dan kemudian ke jaringan inang melalui arbuskula. Diameter hifa mikoriza yang jauh lebih kecil daripada diameter akar, panjang serta tersebar luas mengisi rongga dalam media menyebabkan semakin luasnya permukaan untuk menyerap unsur hara dan air, sehingga tambahan hara (NPK) dapat diserap tanaman lebih banyak dan kebutuhan air tercukupi (Bonfate dan Scannerini, 1992).
Setiadi (1989) menjelaskan dugaan bahwa tanaman yang terinfeksi mikoriza dapat menyerap P lebih banyak karena adanya peningkatan aktivitas enzim fosfatase pada rizosfir dan akar tanaman. Potensi mikoriza untuk memperbaiki nutrisi tanaman, sebagai ‘’bioprotection’’, meningkatkan resistensi terhadap kekeringan, terlibat dalam siklus biogeokimia (mengefektifkan daur ulang unsur hara untuk mempertahankan poduktivitas lahan, stabilitas dan keanekaragaman hayati) sinergis dengan mikroorganisme lainnya. Mikoriza juga dapat dijadikan salah satu teknologi alternatif untuk membantu pertumbuhan, meningkatkan produktivitas dan meningkatkan kualitas tanaman.
7 Tanaman Kahat P
P Tanah tersedia Rendah
Kandungan P Akar Rendah
Fosfolipid Membran Akar Menurun
Permeabilitas Membran Akar Meningkat
Perombakan Gula Reduksi dan Asam Amino Meningkat
Pembentukan Mikoriza Meningkat
Tanaman cukup P
P Tanah tersedia Tinggi
Kandungan P Akar Tinggi
Fosfolipid Membran Akar Meningkat
Permeabilitas Membran Akar Menurun
Perombakan Gula Reduksi dan Asam Amino Menurun
Pembentukan Mikoriza Menurun
Gambar 2. Skema Dugaan Penghambatan Pembentukan Mikoriza (Gunawan, 1993)
Infeksi Akar dan Spora Cendawan Mikoriza Arbuskula (CMA)
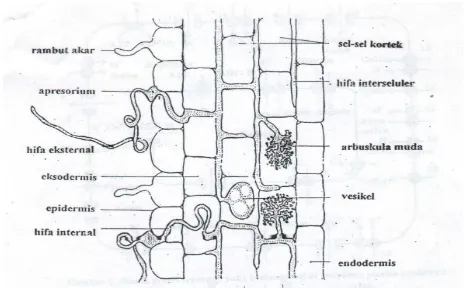
Menurut Fakuara (1988), infeksi pada endomikoriza terjadi pada sekitar epidermis, aksodermis dan korteks parenkim. Perakaran yang terinfeksi CMA ditandai dengan adanya struktur vesikel, hifa dan arbuskula di dalam korteks akar. Intensitas infeksi akar oleh CMA dipengaruhi oleh berbagai faktor, meliputi pemupukan dan nutrisi tanaman, pestisida, intensitas cahaya, musim, kelembaban tanah, pH, kepadatan inokulum dan tingkat ketahanan tanaman.
oleh Arifanti (1999) yang menyebutkan bahwa tidak terdapat korelasi antara jumlah spora dengan persentase akar terinfeksi.
Gambar 3. Hifa, Arbuskula dan Vesikel didalam Jaringan Korteks
Salinitas
Yahya (1987), menjelaskan bahwa salinitas merupakan akumulasi garam-garam terlarut didalam profil tanah, akibat dari proses alam atau karena kegiatan irigasi. Hal ini terjadi akibat penguapan air yang mengandung garam-garam terlarut . Garam-garam yang terlarut dalam larutan tanah, pada konsentrasi tertentu merupakan hara yang esensial bagi pertumbuhan tanaman. Namun pada konsentrasi yang tinggi, garam-garam terlarut akan menjadi masalah dalam pengelolaan tanaman.
Kelebihan kadar garam dalam tanah akan meracuni tanaman dan merupakan stres lingkungan bagi tanaman. Kadar garam yang tinggi akan menghambat perkecambahan benih, perkembangan akar, tinggi tanaman, kualitas serta kuantitas produksi, daya serap tanaman terhadap unsur hara, menghambat pertukaran gas dan menurunkan potensial total air serta potensial osmotik yang akhirnya dapat merusak jaringan tanaman (Bintoro, 1984).
9 kandungan garam melalui kelenjar pada tajuk atau secara pasif dengan mengakumulasikannya pada daun-daun yang tua yang diikuti dengan gugurnya daun (Fitter dan Hay, 1991).
Cendawan Mikoriza Arbuskula dan Tingkat Salinitas
Menurut Tsang dan Maun (1999), tingginya konsentrasi garam NaCl akan berpengaruh negatif terhadap pertumbuhan tanaman dan cendawan mikoriza arbuskula dapat meringankan pengaruh negatif tersebut. Hal ini disebabkan karena CMA mampu meningkatkan penyerapan unsur hara dan transpor phospor tanaman dari media serta mampu meningkatkan toleransi tanaman terhadap serangan patogen akar. Mekanisme mikoriza dalam menekan pengaruh negatif garam NaCl adalah: (1) adanya klorofil yang lebih banyak pada tanaman terinfeksi dapat meningkatkan asupan CO2 oleh tanaman dan (2) CMA meningkatkan nutrisi dan asupan air untuk
tanaman serta mampu memperbaiki daya tahan tanaman pada tanah bersalinitas sehingga secara konsisten dapat meningkatkan produksi berat tajuk.
Cynodon dactylon (L.) Pers (Rumput Bermuda)
Menurut Munandar dan Hardjosuwignyo (1992), Cynodon dactylon (L.) Pers
atau rumput Bermuda merupakan salah satu jenis rumput lansekap yang adaptasinya luas di daerah panas (tropis). Rumput ini terdapat secara alami di Afrika timur, menyebar keseluruh daerah tropika basah dan sub tropika.
Rumput Bermuda dalam taksonomi tumbuhan diklasifikasikan sebagai berikut; kingdom: Plantae, divisi: Embryophyta, Subdivisi: Phana, Kelas:
Monocotyledonae, Subkelas: Glumiflora, Ordo: Chlorideae (Poales), Famili:
Gramineae (Poaceae), Subfamili: Eragrostoideae, Genus: Cynodon L. Rich, Spesies:
Cynodon dactylon (L) Pers.
menambahkan, rumput Bermuda berkembang biak dengan stolon dan rhizoma serta membentuk kumpulan hijauan yang lebat. Rumput ini cepat membentuk pertanaman yang penuh dengan perakaran.
Gambar 4. Cynodon dactylon (L.) Pers Varietas Tifgreen
Cynodon dactylon (L.) Pers memiliki banyak varietas dengan kemampuan pertumbuhan yang berbeda. Varietas Tifway, Tifgreen dan Tifdwarf merupakan turunan pertama (F1) hasil persilangan Cynodon dactylon dengan Cynodon transvaalensis.
11
Media Tumbuh Cynodon dactylon (L.) Pers
Munandar dan Hardjosuwignyo (1992) menyebutkan bahwa pada prinsipnya lahan untuk media tumbuh rumput Bermuda harus menjamin pertumbuhan akar dan tajuk dengan baik. Akar tidak boleh tergenang air, cukup memperoleh udara serta media sehomogen mungkin dan tidak boleh ada gangguan ranting-ranting, batu, batang, kayu dan sejenisnya.
METODE
Lokasi dan Waktu
Penelitian dilaksanakan di rumah kaca Laboratorium Lapang Agrostologi,
Fakultas Peternakan, Institut Pertanian Bogor untuk masa penanaman, pemeliharaan
dan pemanenan (Maret-Juli 2005) serta di Laboratorium Agrostologi Fakultas
Peternakan dan di Laboratorium Bioteknologi kehutanan dan lingkungan, Pusat
Penelitian Sumber Daya Hayati dan Bioteknologi, Institut Pertanian Bogor untuk
menganalisis persentasi infeksi akar dan jumlah spora (November 2005-Februari
2006).
Materi
Bahan-bahan yang digunakan adalah rumput Cynodon dactylon varietas
Tifgreen dalam bentuk rumpun, pasir dengan ukuran 10 mess sebagai media tanam,
air untuk penyiraman, NaCl untuk perlakuan salinitas, isolat CMA Mycofer
(campuran Glomus manihotis, Glomus etunicatum, Gigaspora margarita dan
Acaulospora sp.) dan pupuk Hyponex. Analisis infeksi akar dan penghitungan
jumlah spora menggunakan larutan KOH 2,5 %, HCL 2 %, sukrosa 60 % dan larutan
staining.
Peralatan yang digunakan adalah, pot persegi sebanyak 30 buah, timbangan
analitik, oven 70 0C, sentrifuse mikro, cawan petri, gelas piala 500 ml dan 1 L,
pinset, saringan nematoda berukuran 710 μm,425μm dan 45 μm, gelas objek, cover glass dan label. Analisis infeksi akar menggunakan mikroskop compound dengan
perbesaran 10X10, sedangkan penghitungan jumlah spora menggunakan mikroskop
binokuler.
Rancangan
Penelitian dilakukan dengan menggunakan rancangan perlakuan faktorial
13 salinitas), salinitas dengan konsentrasi 2.500 ppm (5,49 μmhos/cm) , salinitas dengan konsentrasi 5.000 ppm (9,77 μmhos/cm), salinitas dengan konsentrasi 7.500 ppm (14,05 μmhos/cm) dan salinitas dengan konsentrasi 10.000 ppm (18,25 μmhos/cm). Terdapat 10 kombinasi perlakuan dan 3 ulangan sehingga terdapat 30 satuan
percobaan. Tiap satuan percobaan terdiri atas sebuah pot plasik yang berbentuk
persegi panjang.
Model rancangan yang digunakan adalah :
Yijk = μ + τk + αi + βj + (αβ)ij + εijk
Keterangan :
Yijk = nilai pengamatan karena adanya pengaruh mikoriza ke-i, level salinitas ke-j
dan kelompok ke-k μ = rataan umum
τk = pengaruh kelompok ke-k (k= 1, 2 dan 3) αi = pengaruh perlakuan mikoriza ke-i (i = 1 dan 2)
βj = pengaruh perlakuan level salinitas ke-j (j = 1, 2, 3, 4 dan 5) (αβ)ij = pengaruh interaksi mikoriza ke-i, level salinitas ke-j
εijk = pengaruh galat perlakuan mikoriza ke-i, level salinitas ke-j dan kelompok ke-k
Data yang diperoleh dianalisa dengan Sidik Ragam (ANOVA) dan jika berbeda
nyata dilanjutkan dengan Uji Jarak Berganda Duncan (Mattjik dan Sumertajaya,
2000).
Prosedur
1. Persiapan bahan dan alat
Bahan yang digunakan adalah pasir ± 300 kg sebagai media tanam, bibit
rumput Cynodon dactylon, isolat CMA Mycofer, air dan garam NaCl. Alat yang
digunakan adalah pot plastik 30 buah, timbangan, gelas piala, sendok, label.
2. Pengolahan media tanam
Pasir yang telah dibersihkan kemudian dijemur selama ± 1 minggu, lalu
dimasukkan kedalam 30 buah pot plastik, masing-masing pot ± 8,5 kg. Setelah itu,
margarita dan Acaulospora sp.) sebanyak 100 g/8.5 kg pasir ditebarkan kedalam 15
pot, sedangkan 15 pot lagi tidak diberi CMA. Masing-masing pot disiram 3 L air.
Bibit rumput Cynodon dactylon dalam bentuk rumpun yang telah dibersihkan
ditanam pada masing-masing pot hingga menutupi seluruh bagian pot. Terakhir,
siram kembali masing-masing pot dengan 500 ml air.
3. Pemeliharaan
Cynodon dactylon yang telah ditanam setiap hari disiram air 1 kali sehari
dengan volume penyiraman ± 400 ml/pot. Rumput dipupuk dua kali seminggu
sebanyak 100 ml/pot tiap pemupukan dengan menggunakan pupuk Hyponex. Pupuk
dilarutkan dengan konsentrasi 0,5 g Hyponex/1 L.
Perlakuan salinitas garam (NaCl) dilakukan dengan mengencerkan garam
NaCl murni sesuai level salinitas yang diperlukan. Level salinitas 2.500 ppm
diperoleh dengan mengencerkan NaCl murni 2,5 g didalam 1 L air. Level salinitas
5.000 ppm diperoleh dengan mengencerkan NaCl murni 5,0 g didalam 1 L air.
Untuk level salinitas 7.500 ppm diperoleh dengan mengencerkan NaCl murni 7,5 g
didalam 1 L air. Terakhir level salinitas 10.000 ppm diperoleh dengan mengencerkan
NaCl murni 10,0 g didalam 1 L air.
Perlakuan salinitas baru dilakukan dua minggu setelah tanam tujuannya
supaya tanaman dapat beradaptasi terlebih dahulu dengan lingkungannya. Untuk
tanaman yang mati sebelum perlakuan dilakukan penyulaman. Perlakuan salinitas
dilakukan secara bertahap supaya tanaman tidak langsung mengalami stres. Selama
6 minggu volume salinitas diberikan sebanyak 100 ml/pot, sedangkan sisanya ± 300
ml/pot disiram air biasa, karena tidak ada perubahan, volume salinitas ditingkatkan
menjadi 200 ml/pot, sisanya ± 200 ml/pot disiram air biasa selama 8 minggu.
Terakhir, volume penyiraman salinitas ditingkatkan hingga menjadi 400 ml/pot dan
tanpa disiram air biasa.
4. Pemanenan
Pemanenan dilakukan 3 kali, panen pertama dilakukan setelah tanaman
berumur 8 minggu, yang dipanen adalah bagian tajuk tanaman. Sebelum dilakukan
15 kedua dilakukan 8 minggu setelah panen pertama. Seperti halnya panen pertama,
sebelum tajuk dipanen terlebih dahulu dihitung jumlah stolon yang ada. Terakhir
adalah panen ketiga yang dilakukan 4 minggu setelah panen kedua. Pada panen
terakhir ini, selain tajuk dilakukan pula pemanenan akar. Sebelum tajuk serta akar
dipanen terlebih dahulu dilakukan perhitungan jumlah individu pada masing-masing
pot, setelah itu diambil sampel akar dan pasir untuk analisa persentase infeksi akar
dan jumlah spora.
Peubah yang diamati
1. Jumlah Stolon
Jumlah stolon dihitung sesaat sebelum panen pertama, kedua dan ketiga.
Stolon yang dihitung adalah yang panjangnya sudah melewati bagian pinggir pot.
2. Jumlah Individu
Jumlah individu tanaman yang terdiri dari batang dan daun yang berasal dari
tiap-tiap stolon disetiap pot dihitung sesaat sebelum panen ketiga.
3. Berat Kering Tajuk dan Akar
Pengukuran berat kering tajuk dan akar dilakukan dengan cara menimbang
tajuk dan akar yang telah dioven pada suhu 70ºC selama 48 jam dengan
menggunakan neraca analitik.
4. Persentase Infeksi Akar
Untuk menghitung jumlah akar yang terinfeksi oleh CMA dilakukan melalui
teknik pewarnaan akar (Phyllip dan Hayman, 1970 yang dimodifikasi oleh teknik
Koske dan Gemma, 1989). Pewarnaan akar dilakukan dengan cara akar dicuci
hingga bersih terlebih dahulu dan dimasukkan ke dalam tabung film, setelah itu KOH
2.5 % ditambahkan dan tabung kemudian ditutup. Setelah akar berwarna bening
(sekitar 7-10 hari) KOH 2.5 % dibuang, kemudian akar dicuci dibawah air mengalir
dan disaring menggunakan saringan teh, kemudian dimasukkan kembali ke dalam
tabung film dan ditambahkan dengan HCl 2 %. Setelah 24 jam, HCl dibuang,
terlalu pekat larutan destaining ditambahkan. Untuk menghitung infeksi akar, akar
dengan panjang sekitar 1 cm diambil sebanyak 10 buah, kemudian diletakkan di
objekglass dan kemudian ditutup dengan coverglass. Penghitungan jumlah akar yang
terinfeksi dilakukan dibawah mikroskop dengan perbesaran 10X10. Persentase akar
yang terinfeksi dihitung dengan rumus sebagai berikut :
% infeksi = jumlah akar yang terinfeksi X 100 %
jumlah contoh akar
4. Jumlah Spora
Penghitungan jumlah spora dilakukan dengan mengisolasi spora lebih
dahulu melalui metode penyaringan basah (Nicholson dan Gadermann, 1963).
Isolasi dilakukan dengan cara menimbang 50 gram pasir, lalu ditambah air, diaduk
dan didiamkan selama 5 menit sehingga terbentuk suspensi. Setelah itu disaring
bertingkat sebanyak 3 kali menggunakan saringan dengan ukuran 710 μm, 425 μm
dan 45 μm. Pasir yang terendap pada saringan 45 μm dimasukkan kedalam tabung
sentrifuse dan ditambahkan larutan sukrosa 60 %, setelah itu disentrifuse selama 5
menit dengan kecepatan 2.500 rpm. Supernatan yang diperoleh, ditampung dengan
saringan 45 μm (jangan sampai endapannya terbawa), kemudian dibilas dengan
aquades agar sukrosanya hilang, setelah itu ditampung dalam cawan petri dan
HASIL DAN PEMBAHASAN
Keadaan Umum Percobaan
Penanaman dilakukan di rumah kaca, dengan tujuan agar faktor-faktor yang mempengaruhi pertumbuhan tanaman, seperti intensitas cahaya matahari, suhu lingkungan, kelembaban dan angin yang diterima oleh tanaman relatif sama. Selama penelitian berlangsung, tanaman pernah mengalami gejala serangan hama sehingga dilakukan penyemprotan pestisida. Gambar 5 memperlihatkan Cynodon dactylon
varietas Tifgreen umur saat penelitian umur 8 minggu.
Gambar 5. Cynodon dactylon Saat Penelitian
Pengamatan visual di lapang menunjukkan tanaman baru mengalami 100 % kematian di minggu keduapuluh, yang mengalami hal tersebut adalah tanaman yang tidak diberi CMA pada level salinitas 10.000 ppm. Terjadinya kematian disebabkan tanaman sudah tidak mampu lagi mentolerir konsentrasi salinitas yang terlalu tinggi. Hal ini berbeda dengan penelitian Prihantoro (2003) yang menguji salinitas pada tanaman Lamtoro. Pada penelitian tersebut, lamtoro mengalami kematian diatas konsentrasi salinitas 5.000 ppm delapan minggu setelah penanaman.
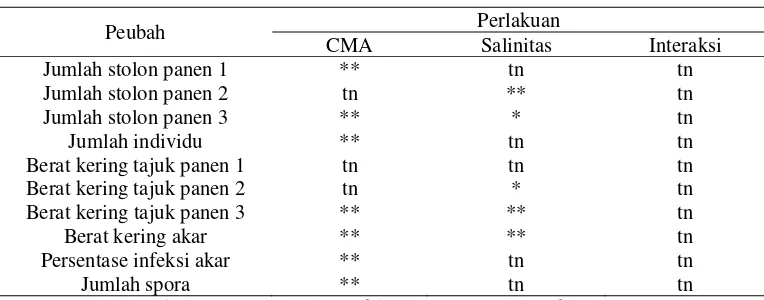
Tabel 1. Rekapitulasi Sidik Ragam Penelitian
Peubah Perlakuan
CMA Salinitas Interaksi
Jumlah stolon panen 1 Jumlah stolon panen 2 Jumlah stolon panen 3
Jumlah individu Berat kering tajuk panen 1 Berat kering tajuk panen 2 Berat kering tajuk panen 3
Berat kering akar
Rekapitulasi sidik ragam memperlihatkan pemberian cendawan mikoriza arbuskula (CMA) sangat nyata mempengaruhi jumlah stolon panen 1 dan 3, jumlah individu, berat kering tajuk panen 3, berat kering akar, persentase infeksi akar dan jumlah spora (p<0,01), sedangkan faktor tunggal penyiraman dengan berbagai level salinitas sangat nyata (p<0,01) berpengaruh terhadap jumlah stolon panen 2, berat kering tajuk panen 3 serta bahan kering akar dan nyata (p<0,05) mempengaruhi jumlah stolon panen 3 dan berat kering tajuk panen 2. Semua peubah tidak memberikan respon yang nyata terhadap interaksi antara level salinitas dengan pemberian CMA.
Jumlah Stolon
Pertumbuhan merupakan proses yang dilakukan oleh tanaman hidup pada lingkungan tertentu untuk menghasilkan kemajuan perkembangan dengan menggunakan faktor lingkungan seperti CO2, unsur-unsur hara, air dan radiasi dari
matahari (Sitompul dan Guritno, 1995). Salah satu cara yang dapat dilakukan untuk mengetahui pertumbuhan rumput Cynodon dactylon adalah dengan menghitung jumlah stolon yang dihasilkan, karena rumput ini berkembang biak menggunakan stolon.
19 salinitas tidak nyata mempengaruhi jumlah stolon Cynodon dactylon untuk semua panen. Rataan jumlah stolon Cynodon dactylon panen ke-1, 2 dan 3 ditunjukkan pada Tabel 2 dibawah ini.
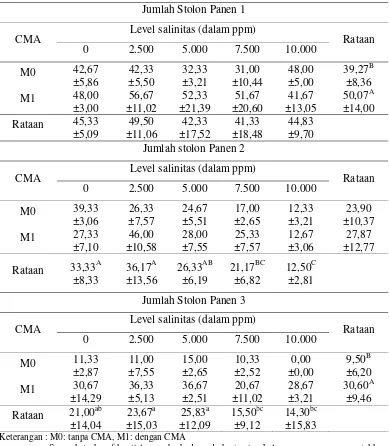
Tabel 2. Rataan Jumlah Stolon Panen 1, 2 dan 3 Cynodon dactylon (buah). Jumlah Stolon Panen 1
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000
Jumlah stolon Panen 2
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000
Jumlah Stolon Panen 3
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000 Keterangan : M0: tanpa CMA, M1: dengan CMA
Superskrip huruf kapital yang berbeda pada baris atau kolom yang sama menunjukkan sangat berbeda nyata (p<0,01).
Tidak menurunnya jumlah stolon Cynodon dactylon pada panen pertama memperlihatkan bahwa rumput ini memiliki tingkat toleransi yang tinggi terhadap kondisi salinitas. Menurut Delvian (2003), suatu tanaman dapat dikatakan memiliki toleransi terhadap kondisi salinitas apabila: (1) Mampu untuk hidup pada tanah salin; (2) Mampu berproduksi pada tanah salin; (3) Terdapat hasil relatif pada tanah salin yang dapat dibandingkan dengan hasil pada tanah normal; (4) Salinitas maksimum yang dapat dialami tanaman tanpa terjadi penurunan hasil dan (5) Persentase penurunan hasil setiap unit peningkatan salinitas tanah.
Pada panen ke-2, pemberian salinitas sangat nyata (p<0,01) menurunkan jumlah stolon rumput Cynodon mulai level 5.000, 7.500 dan 10.000 ppm dan pada panen ke-3 pemberian salinitas nyata (p<0,05) menurunkan jumlah stolon pada level 7.500 dan 10.000 ppm. Hal ini disebabkan pada panen ke-2 dan ke-3, volume penyiraman salinitas yang diberikan lebih tinggi dibandingkan pada panen pertama. Saat panen pertama, volume salinitas yang diberikan adalah 100 ml, sedangkan pada panen kedua volume salinitas yang diberikan ditingkatkan menjadi 200 ml dan saat panen terakhir volume penyiraman salinitas diberikan sebanyak 400 ml. Menurunnya jumlah stolon panen ke-2 dan ke-3 pada level salinitas 7.500 dan 10.000 ppm disebabkan tanaman yang tumbuh dalam kondisi tanah salin atau yang mendapat perlakuan NaCl akan mengakumulasikan prolin, suatu asam amino yang dapat larut. Akumulasi prolin tersebut merupakan usaha tanaman untuk menyesuaikan tekanan osmotik. Penyesuaian tekanan osmotik ini membutuhkan energi sehingga akan mengurangi pertumbuhan tanaman (Delvian, 2003).
Pemberian CMA sangat nyata meningkatkan (p<0,01) jumlah stolon Cynodon
21 hifa eksternal memiliki ukuran diameter sangat kecil (2-4 μm) dibanding dengan diameter rambut akar yang hanya sekitar 10 μm.
Jumlah Individu
Hasil sidik ragam pada Lampiran 4 menunjukkan level salinitas tidak berpengaruh nyata terhadap jumlah individu Cynodon. Pemberian CMA sangat nyata (p<0,01) meningkatkan jumlah individu Cynodon, sedangkan jumlah individu tidak memberikan respon yang nyata terhadap interaksi antara level salinitas dan pemberian cendawan mikoriza arbuskula (CMA). Rataan jumlah individu rumput
Cynodon dactylon ditampilkan pada Tabel 3.
Tabel 3. Rataan Jumlah Individu Cynodon dactylon (buah)
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000 Keterangan : M0: tanpa CMA, M1: dengan CMA
Superskrip huruf kapital yang berbeda pada baris yang sama menunjukkan sangat berbeda nyata (p<0,01).
Tidak menurunnya jumlah individu rumput Cynodon meskipun diberi siraman salinitas, karena diduga jumlah potensial air yang tersedia masih mencukupi kebutuhan tanaman untuk melakukan fotosintesis, selain itu Cynodon termasuk tumbuhan golongan C4 (McNaughton dan Wolf, 1990 dalam Kurniasari, 2005), yang
mampu bertahan pada potensial air serendah -1000 bar dan memiliki CER (Carbon dioxide Exchange Rate) dan WUE (Water Use Efficiency) yang tinggi. Mekanisme-mekanisme ini penting bagi tanaman dalam beradaptasi dengan kondisi stres garam (Fitter dan Hay, 1991).
Cynodon dengan semakin tingginya level salinitas yang diberikan disebabkan karena rumput ini memiliki mekanisme di dalam tubuhnya untuk mengatasi pengaruh negatif salinitas, salah satunya dengan cara adaptasi morfologi. Pada tanaman yang toleran terhadap kondisi salinitas NaCl akan ditimbun dalam vakuola sel daun, sedangkan di dalam sitoplasma dan organela konsentrasi garam tetap rendah sehingga tidak mengganggu aktivitas enzim dan metabolisme (Epstein, 1997). Tanaman yang toleran terhadap kondisi salin juga mampu mencapai keseimbangan termodinamik tanpa terjadi kerusakan jaringan yang berarti, karena tanaman dapat menyesuaikan tekanan osmotik selnya untuk mencegah terjadinya dehidrasi.
Pemberian CMA menunjukkan pengaruh yang sangat nyata (p<0,01) dalam meningkatkan jumlah individu rumput Cynodon dactylon. Rumput yang tidak diberi CMA memiliki rataan jumlah individu ± 393 buah, sedangkan rumput yang diberi
CMA memiliki rataan jumlah individu ± 435 buah. Hal ini disebabkan CMA mampu
membantu meningkatkan penyerapan unsur hara dengan mensuplai mineral-mineral anorganik yang ada di dalam tanah yang dibutuhkan oleh tanaman (Sanders, 1986).
Tumbuhan yang diinokulasi mikoriza dapat tumbuh lebih baik daripada tumbuhan yang tidak diinokulasi disebabkan meningkatnya penyerapan unsur hara makro terutama phospor dan beberapa unsur hara mikro pada tanaman yang terinfeksi karena adanya hifa eksternal dari cendawan yang berperan sebagai sistem perakaran yang menyebabkan tersedianya daerah serapan yang lebih luas dalam menyerap hara. Unsur hara yang telah terakumulasi pada hifa eksternal akan ditranslokasikan ke hifa internal oleh suatu sistem transport dan kemudian ke jaringan inang melalui arbuskula yang langsung dipindahkan ke tanaman inang, sehingga hifa ini sering disebut sebagai jalan bebas hambatan (Gunawan, 1993).
Produksi Berat Kering Tajuk dan Akar
23
Berat Kering Tajuk
Hasil sidik ragam produksi berat kering tajuk yang terdapat dalam Lampiran 5, 6 dan 7 menunjukkan pemberian cendawan mikoriza arbuskula (CMA) pada panen pertama dan kedua tidak memberikan pengaruh yang nyata, sedangkan pada panen terakhir, adanya CMA sangat nyata (p<0,01) meningkatkan produksi berat kering tajuk Cynodon dactylon. Tabel 4 menyajikan rataan produksi berat kering tajuk Cynodon dactylon panen ke-1,2 dan 3.
Tabel 4. Rataan Produksi Berat Kering Tajuk Panen 1, 2 dan 3
Cynodon dactylon (gram).
Produksi Berat Kering Tajuk Panen 1
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000
Produksi Berat Kering Tajuk Panen 2
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000 Produksi Berat Kering Tajuk Panen 3
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000 Keterangan : M0: tanpa CMA, M1: dengan CMA
Superskrip huruf kecil yang berbeda pada kolom yang sama menunjukkan berbeda nyata (p<0,05)
Pemberian level salinitas nyata (p<0,05) menurunkan produksi berat kering tajuk Cynodon pada panen kedua di level 7.500 dan 10.000 ppm, sedangkan pada panen terakhir pemberian salinitas sangat nyata (p<0,01) meningkatkan produksi berat kering tajuk di level 5.000 hingga 10.000 ppm. Interaksi antara pemberian CMA dan level salinitas tidak berpengaruh nyata terhadap produksi berat kering tajuk panen 1, 2 dan 3.
Tidak berpengaruhnya level salinitas pada panen pertama produksi berat kering tajuk disebabkan rumput ini mampu menghindari terjadinya keracunan atau ketidakseimbangan hara dengan tiga cara, yaitu: eksklusi, ekskresi dan sekresi. Eksklusi terjadi secara pasif dengan adanya dinding sel yang tidak permeabel terhadap garam atau ion-ion dari garam tersebut. Ekskresi dan sekresi merupakan pemompaan ion secara aktif masing-masing ke luar tanaman dan ke dalam vakuola (Levitt, 1980). Selain itu, pada panen pertama ini volume penyiraman salinitas hanya 100 ml, masih lebih rendah jika dibandingkan dengan volume penyiraman saat panen ke-2 yang diberikan sebesar 200 ml.
Pada panen ketiga terlihat bahwa level salinitas tidak menyebabkan produksi berat kering tajuk Cynodon dactylon mengalami penurunan. Berat kering tajuk terbesar terdapat pada rumput dengan kondisi salinitas 5.000 ppm, yaitu sebesar 17,25 g, tidak berbeda nyata dengan berat kering tajuk yang berada dalam kondisi salinitas 7.500 dan 10.000 ppm, namun berbeda sangat nyata (p<0,01) dengan berat kering tajuk yang berada dalam kondisi salinitas 0 dan 2.500 ppm. Berat kering tajuk terkecil justru terdapat pada rumput dengan kondisi salinitas 0 ppm, yaitu sebesar 9,78 gram. Hal ini diduga karena Cynodon merupakan jenis rumput yang tahan hidup dalam kondisi salinitas, sehingga akan tumbuh lebih baik jika terdapat garam dalam tubuhnya. Terbukti adanya konsentrasi salinitas mulai dari 5.000 ppm akan meningkatkan produksi berat kering tajuk rumput ini. Hal ini sesuai dengan hasil penelitian Bintoro (1984) yang menyebutkan bahwa Na+ dapat menggantikan sebagian peran K+ dalam meningkatkan pertumbuhan vegetatif dan reproduktif tanaman tomat dan terung.
25 pertumbuhan tanaman di lingkungan bergaram berhubungan langsung dengan ketahanan tanaman terhadap tekanan osmotik dan keracunan oleh ion-ion spesifik, misalnya Na dan Cl. Tanaman yang tumbuh pada derah-daerah berkadar garam tinggi akan banyak menyerap ion Na dan Cl. Ion-ion tersebut bergerak menuju perakaran tanaman melalui aliran massa. Sebelum mencapai ambang kritis, akumulasi ion masih dapat ditolerir tanaman sehingga tidak terjadi efek toksik.
Analisa ragam pada Lampiran 5 dan 6 yang menunjukkan belum mampunya CMA meningkatkan produksi berat kering tajuk pada panen pertama dan kedua disebabkan tertundanya perkecambahan spora dari cendawan ini. Pada panen ketiga, meskipun volume salinitas yang diberikan semakin tinggi dan penyerapan unsur hara serta air semakin berkurang namun adanya CMA sangat nyata (p<0,01) meningkatkan produksi berat kering tajuk. Hal ini disebabkan terjadinya pemulihan kemampuan berkecambah dari spora-spora CMA setelah masa adaptasi yang lebih lama. Selain itu, pada panen terakhir ini, hifa eksternal yang dimiliki mikoriza telah bekerja maksimal untuk membantu penyerapan unsur hara dan air.
Pada panen ketiga terlihat, rumput yang tidak diberi CMA memiliki rataan produksi berat kering tajuk sebesar 11,55 g, sedangkan rumput yang diberi CMA memiliki rataan produksi berat kering sebesar 17,27 g, sehingga dapat terlihat bahwa pemberian CMA mampu meningkatkan produksi berat kering rumput pada panen ketiga sebesar ± 50%. Kehadiran CMA mampu memperbaiki daya tahan tanaman
pada tanah bersalinitas, sehingga secara konsisten dapat meningkatkan produksi berat kering tajuk yang lebih tinggi. Hal ini disebabkan mikoriza mempunyai suatu mekanisme untuk menekan pengaruh negatif garam NaCl melalui: (1) adanya klorofil yang lebih banyak pada tanaman terinfeksi akan meningkatkan asupan CO2 oleh
tanaman dan (2) CMA meningkatkan nutrisi dan asupan air untuk tanaman (Tsang dan Maun, 1999).
terinfeksi penyerapan hara (N, P dan K) akan lebih banyak dan kebutuhan air tercukupi.
Berat Kering Akar
Menurut Harjadi (1996), peranan akar dalam pertumbuhan tanaman sangat berhubungan dengan tajuk, karena tajuk berfungsi dalam proses fotosintesis dan akar berfungsi dalam menyediakan unsur hara dan air yang digunakan dalam metabolisme tanaman. Perlakuan level salinitas dan pemberian cendawan mikoriza arbuskula (CMA) menunjukkan pengaruh yang sangat nyata (p<0,01) terhadap peningkatan akumulasi berat kering akar Cynodon dactylon (Lampiran 8). Interaksi antara level salinitas dengan pemberian CMA tidak berpengaruh nyata terhadap produksi berat kering akar Cynodon dactylon. Hal ini dapat dilihat pada Tabel 5.
Tabel 5. Rataan Berat Kering Akar Cynodon dactylon (gram)
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000 Keterangan : M0: tanpa CMA, M1: dengan CMA.
Superskrip huruf kapital yang berbeda pada baris atau kolom yang sama menunjukkan sangat berbeda nyata (p<0,01).
27 Kemampuan Cynodon untuk hidup dalam kondisi salinitas disebabkan karena rumput ini merupakan salah satu jenis tanaman yang mampu mengurangi kandungan garam yang terdapat didalam jaringan osmotiknya dengan cara: (1) mengakumulasi kandungan garam didalam akar (dalam hal ini didalam stolon); (2) mengembangkan jaringan penyimpan air untuk mengurangi tekanan osmotik yang tinggi; (3) mengekskresikan secara aktif kandungan garam melalui kelenjar pada tajuk atau secara pasif dengan mengakumulasikannya pada daun-daun yang tua yang diikuti dengan gugurnya daun (Fitter dan Hay, 1991).
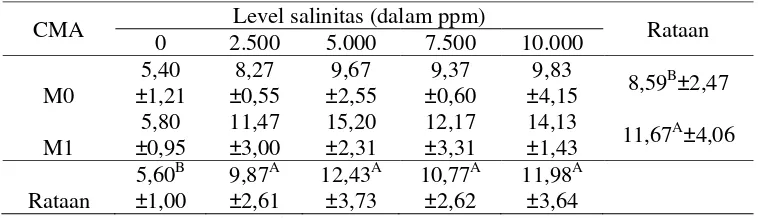
Pemberian CMA menunjukkan pengaruh yang sangat nyata (p<0.01) dalam meningkatkan produksi berat kering akar rumput Cynodon dactylon. Rumput yang tidak diberi CMA memiliki berat kering akar sebesar 8,59 g, sedangkan rumput yang diberi CMA memiliki berat kering akar sebesar 11,67 g. Sehingga dapat terlihat bahwa adanya CMA dapat meningkatkan produksi berat kering akar sebesar ± 36%.
Hal ini disebabkan CMA mampu membantu meningkatkan penyerapan unsur hara dengan mensuplai mineral-mineral anorganik yang ada di dalam tanah yang dibutuhkan oleh tanaman, khususnya mineral phospor sehingga dapat meningkatkan berat akar dan tajuk (Sanders, 1986).
Persentase Infeksi Akar
Akar yang terinfeksi CMA merupakan sumber inokulum yang efektif dan efektifitasnya lebih tinggi daripada spora (Gunawan, 1993). Intensitas infeksi akar oleh CMA dipengaruhi oleh berbagai faktor, meliputi pemupukan dan nutrisi tanaman, pestisida, intensitas cahaya, musim, kelembaban tanah, pH, kepadatan inokulum dan tingkat ketahanan tanaman. Rataan persentase infeksi akar ditampilkan pada Tabel 6.
Gambar 6. Perakaran yang Terinfeksi CMA
Perakaran yang terinfeksi CMA ditandai dengan adanya struktur vesikel, hifa dan arbuskula di dalam korteks akar (Fakuara, 1988). Gambar 6 dan 7 memperlihatkan perakaran yang terinfeksi dan tidak terinfeksi CMA.
29 Tabel 6. Rataan Persentase Infeksi Akar
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000 Keterangan : M0: tanpa CMA, M1: dengan CMA
Superskrip huruf kapital yang berbeda pada baris yang sama menunjukkan sangat berbeda nyata (p<0,01).
Dari hasil sidik ragam yang terdapat pada Lampiran 9 terlihat bahwa faktor tunggal level salinitas dan interaksi antara level salinitas dengan pemberian CMA tidak berpengaruh nyata terhadap persentase infeksi akar, sedangkan pemberian CMA sangat nyata (p<0,01) meningkatkan persentase infeksi akar. Rataan persentase infeksi akar menunjukkan tanaman yang diberi CMA memiliki persentase infeksi akar sebesar 91,42 %, sedangkan tanaman yang tidak diberi CMA memiliki persentase infeksi akar 69,08 %. Adanya akar yang terinfeksi pada tanaman yang tidak diberi CMA diduga berasal dari endofit yang terdapat dalam media (Fakuara, 1988).
bermikoriza akan mampu meningkatkan kapasitasnya dalam menyerap unsur hara dan air dan ketersediaan air bagi tanaman akan meningkat.
Jumlah Spora
Untuk berkembang biak dengan cara aseksual, mikoriza menghasilkan spora yang dibentuk oleh hifa eksternal dengan ukuran 20-1000 µm didalam tanah. Spora merupakan bentuk pertahanan diri mikoriza pada kondisi stres kering (Fakuara, 1988).
Menurut Smith dan Read (1997), tidak dapat dipastikan bahwa tanaman dengan persentase infeksi akar yang tinggi akan menghasilkan jumlah spora yang tinggi pada rhizosfer atau sebaliknya. Hal ini dinyatakan pula oleh Arifanti (1999) yang menyebutkan bahwa tidak terdapat korelasi antara jumlah spora dengan persentase akar terinfeksi. Gambar 8 merupakan isolat CMA Mycofer (campuran
Glomus manihotis, Glomus etunicatum, Gigaspora margarita dan Acaulospora sp) yang digunakan pada penelitian ini.
Gambar 8. Isolat CMA Mycofer yang digunakan
31 Tabel 7. Rataan Jumlah Spora (per 50 gram sampel)
CMA Level salinitas (dalam ppm) Rataan 0 2.500 5.000 7.500 10.000 Keterangan : M0: tanpa CMA, M1: dengan CMA
Superskrip huruf kapital yang berbeda pada baris yang sama menunjukkan sangat berbeda nyata (p<0,01).
Jumlah spora terbanyak dihasilkan pada tanaman dengan kondisi salinitas 2.500 ppm, sedangkan jumlah spora paling kecil dihasilkan pada tanaman dengan kondisi salinitas 10.000 ppm. Hal ini diduga terjadi akibat tingginya konsentrasi garam NaCl yang mengakibatkan terhambatnya isolat mikoriza dalam memproduksi spora. Selain itu, jumlah spora tidak hanya dipengaruhi oleh satu faktor saja, melainkan dipengaruhi oleh akumulasi dari beberapa faktor, diantaranya: cendawan itu sendiri, varietas tanaman inang dan kondisi lingkungan, seperti cahaya dan suhu, karena cahaya matahari berperan dalam pembentukan karbohidrat melalui asimilasi karbon yang selanjutnya CMA akan menggunakan karbon tersebut sebagai sumber energi bagi pertumbuhannya (Fakuara, 1988).
Analisa ragam menunjukkan interaksi antara salinitas dengan CMA tidak berpengaruh nyata pada semua peubah. Hal ini menunjukkan bahwa rumput
KESIMPULAN DAN SARAN
Kesimpulan
Cynodon dactylon (rumput Bermuda) merupakan jenis rumput yang memiliki tingkat toleransi tinggi hingga kondisi salinitas 10.000 ppm. Dalam keadaan terinfeksi cendawan mikoriza arbuskula (CMA), Cynodon mampu meningkatkan toleransinya terhadap media tanam bersalinitas tinggi. Pemberian cendawan mikoriza arbuskula (CMA) terbukti mampu memperbaiki pertumbuhan dan produktivitas rumput Cynodon dactylon (L.) Pers.
Saran
UCAPAN TERIMAKASIH
Alhamdulillahirobbil’alamin, segala puji dan syukur penulis panjatkan ke hadirat-Mu Ya Allah, karena dengan kekuatan, rahmat dan hidayah dari-Mu, tugas akhir ini dapat diselesaikan dengan baik.
Terima kasih untuk Mama, Papa dan adikku Yopi atas motivasi, dukungan, perhatian, kasih sayang serta doa’ tulus yang selalu terucap untuk penulis. Untuk keluarga besar Kamaruddin dan keluarga besar Dawar, terima kasih atas dukungan, semangat dan doa yang telah diberikan.
Terima kasih penulis ucapkan kepada Dr. Ir. Panca Dewi M.H.K.S, Msi. selaku pembimbing utama dan Dr. Ir. Luki Abdullah, MSc.Agr. selaku pembimbing anggota yang telah membimbing, mengarahkan dan membantu penulis dengan penuh kesabaran dan keikhlasan mulai dari penyusunan usulan proposal hingga tahap akhir penulisan skripsi ini. Kepada Ir. Anita Sardiana Tjajkradidjaya, MRur.Sc. selaku penguji seminar dan Ir. Sri Rahayu, MSi selaku penguji sidang yang telah banyak memberikan saran demi kesempurnaan skripsi ini. Terima kasih tidak lupa penulis ucapkan Kepada Dr. Ir. Rachjan G. Pratas, MSc dan Ir. Kukuh Budi Satoto, MS selaku pembimbing akademik yang telah memberikan bimbingan dan arahan selama penulis menjalani studi di Fakultas Peternakan.
Kepada seluruh staf Laboratorium Agrostologi, Fakultas Peternakan dan staf Laboratorium Bioteknologi kehutanan dan lingkungan Pusat Antar Universitas, IPB yang telah banyak membantu selama penelitian berlangsung penulis ucapkan terima kasih. Terima kasih penulis ucapkan kepada seluruh dosen di Fakultas Peternakan IPB yang telah banyak memberikan ilmu selama penulis menuntut ilmu di IPB
DAFTAR PUSTAKA
Abbot, L.K. and A.D. Robson. 1984. The effect of mycorrhizal on plant growth.
Dalam: C.L. Powell and D.J Bagyaraj (Editor). Vesikular arbuskula mychorrizal. CRC Press. Inc. Boca Raton, Florida.
Arifanti, V.B. 1999. Pengaruh penggunaan media tumbuh, pupuk NPK dan cendawan endomikoriza Glomus etunicatum terhadap pertumbuhan bibit
Tectona grandis L.f. Skripsi. Fakultas Kehutanan. Institut Pertanian Bogor, Bogor.
Beard, J.B. 1973. Turfgrass Science and Culture. Prentice-Hall. Inc, New Jersey.
Bintoro, M.H. 1984. Ketahanan tanaman sayuran terhadap lingkungan bergaram. Laporan Penelitian. Fakultas Pertanian. Institut Pertanian Bogor, Bogor.
Bintoro, M.H. 1990. Pengaruh NaCl terhadap pertumbuhan kultivar tomat. Laporan Penelitian. Fakultas Pertanian. Institut Pertanian Bogor, Bogor.
Bonfate, F.P and S. Scannerini. 1992. The celluler basis of plant fungus interchanges in mycorrhizal association. Dalam: M.F. Allen (Editor). Mycorrhizal function: an integrative plant-fungal process. Routledge Chapman&Hall. Inc, New York.
Brundett, M.C., L. Melville and L. Peterson. 1994. Practical Methods in Mycorrhizal Research. Mycologue Publications, Ohio.
Delvian. 2003. Keanekaragaman cendawan mikoriza arbuskula (CMA) di hutan pantai dan potensi pemanfaatannya (studi kasus di hutan cagar alam Leuweung Sancang, kabupaten Garut, Jawa Barat). Disertasi. Institut Pertanian Bogor, Bogor.
Epstein, E. 1997. Better crops for food. CIBA Foundation Simposium, London.
Fakuara, M.Y. 1988. Mikoriza, Teori dan Kegunaan dalam Praktek. Pusat Antar Universitas. Institut Pertanian Bogor, Bogor.
Fitter, A.H dan R.K.M. Hay. 1991. Fisiologi Lingkungan Tanaman. Universitas Gadjah Mada, Yogyakarta.
Gendermann, J.W and T.H Nicholson. 1963. Spores of mycorrhizal endogene species extracted from soil by wet sieving dan decanting. Transactions of the British mycological. Soc. 426 (2): 235-244.
Gohl, Bo. 1981. Tropical Feeds. Food and Agriculture Organization of the United Nations, Rome.
Hardjowigeno, S. 1992. Ilmu Tanah. PT. Mediyatama Sarana Perkasa, Jakarta.
Harjadi, S.S 1996. Pengantar Agronomi. PT Gramedia Pustaka Utama, Jakarta.
Imas, T., R.S. Hadioetomo, A.W. Gunawan dan Y. Setiadi. 1989. Mikrobiologi Tanah. Pusat Antar Universitas Bioteknologi. Institut Pertanian Bogor, Bogor.
Indriani, N.P. 2004. Kontribusi mikoriza dengan batuan fosfat dan waktu penjarangan pada tanaman jagung (Zea mays L.) penghasil jagung semi dan hijuan pakan ternak. Disertasi. Institut Pertanian Bogor, Bogor.
Jakobsen, I. 1991. Carbon metabolism in mycorrhizal. Dalam: J.R. Norris., D.J. Read and A.K. Varma (Editor). Methods in microbiology. Academic Press, London.
Jakobsen, I. 1992. Phosphorus transport by external hyphae of vesicular-arbuscular mycorrhizas. Dalam: D.J. Read., D.H. Lewis., A.H. Fitter and I.J. Alexander (Editor). Mycorrhizal in ecosystem. CAB International, London.
Jayadi, S. 1991. Pengenalan jenis tanaman pakan. Diktat Kuliah. Institut Pertanian Bogor, Bogor.
Kabirun, S. 1990. Peranan Mikoriza dalam Pertanian. Pusat Antar Universitas. Bioteknologi. Institut Pertanian Bogor, Bogor.
Karti, P.D.M.H. 2003. Respon morfofisiologi rumput toleran dan peka alumunium terhadap penambahan mikroorganisme dan pembenah tanah. Disertasi. Institut Pertanian Bogor, Bogor.
Kurniasari, D. 2005. Karakterisasi fisiologi rumput bermuda (Cynodon dactylon L.) lokal dalam rangka pengembangan turfgrass asli Indonesia. Skripsi. Fakultas Pertanian. Institut Pertanian Bogor, Bogor.
Koske, R.E dan J.H. Gemma. 1989. A modified procedure for staining roots to detect vesicular arbuscular mycorrhizal. Mycological. Res. 92 (4): 486-505.
Lakitan, B. 2004. Dasar-dasar Fisiologi Tumbuhan. Rajawali Pers, Jakarta.
Levitt, J. 1980. Responses of plant to environtmental stresses. Vol II. Academic Press, New York.
Mattjik, A.A dan M. Sumertajaya. 2000. Perancangan Percobaan dengan Aplikasi SAS dan Minitab Jilid I. Institut Pertanian Bogor, Bogor.
Munandar, A dan S. Hardjosuwignyo. 1992. Rumput Lansekap. Pusat Antar Universitas Ilmu Hayat. Institut Pertanian Bogor, Bogor.
36 assessment of infection. Transactions of the British mycological. Soc. 55: 158-160.
Prihantoro, I. 2003. Pengaruh pemberian kultur Campuran cendawan mikoriza arbuskula (Glomus sp., Gigaspora sp. dan Acaulospora sp.) terhadap pertumbuhan lamtoro (Leucaena leucocephala) pada media zeolit dengan tingkat salinitas yang berbeda. Skripsi. Fakultas Peternakan. Institut Pertanian Bogor, Bogor.
Sanders, I.E. 1986. The Role of mycorrhizal in plant nutrient uptake from soil.
Dalam: V. Jensen, A. Kjoller and L.H. Sorensen (Editor). Mycrobial communities in soil. El Sevier Applied Sci. Pub, New York.
Setiadi, Y. 1989. Pemanfaatan Mikroorganisme dalam Kehutanan. Pusat Antar Universitas. Institut Pertanian Bogor, Bogor.
Sitompul, S.M dan B. Guritno. 1995. Analisis Pertumbuhan Tanaman. Universitas Gajah Mada, Yogyakarta.
Smith, S.E dan D.J Read. 1997. Mycorrhizal Symbiosis. 2nd Edition. Academic Press, London.
Soepardi, G. 1983. Sifat dan Ciri Tanah. Jurusan Tanah, Fakultas Pertanian. Institut Pertanian Bogor, Bogor.
Tan, K.H. 1991. Principles of Soil Chemistry. Madison Venue New York. Inc, New York.
Tjijtrosoepomo, G. 2000. Taksonomi Tumbuhan (Spermatophyta). Universitas Gajah Mada, Yogyakarta.
Tsang, A and M.A. Maun. 1999. Mycorrhizal fungi increase salt tolerance of
strophostyles helvola in Coastal Foredunes. Journal of Plant Ecology. 144: 159-166.
38
Lampiran 1. ANOVA Jumlah Stolon Panen 1
Sumber
Lampiran 2. ANOVA Jumlah Stolon Panen 2
Sumber
Lampiran 3. ANOVA Jumlah Stolon Panen 3
Lampiran 4. ANOVA Jumlah Individu
Sumber keragaman
db JK KT Fhit F.05 F.01
Kelompok 2 180546,47 90273,24 4,48
Faktor A 1 442867,50 442867,50 21,98** 4,41 8,29
Lampiran 5. ANOVA Produksi Berat Kering Tajuk Panen 1
Sumber
Lampiran 6. ANOVA Produksi Berat Kering Tajuk Panen 2
40
Lampiran 7. ANOVA Produksi Berat Kering Tajuk Panen 3
Sumber
Lampiran 8. ANOVA Produksi Berat Kering Akar
Sumber
Lampiran 9. ANOVA Persentase Infeksi Akar
Lampiran 10. ANOVA Jumlah Spora
Lampiran 11. Komposisi Pupuk Hyponex
Total Nitrogen
Nitrate nitrogen Urea Nitrogen
Available Phosphoric Acid (P2O5)
Soluble Potash (K2O)
Total Available Primary Plant Food Trace Elements :
42
Lampiran 12. Bagan pengacakan
Keterangan:
M0 ; Tanpa mikoriza M1 ; Mikoriza
A ; 0 ppm B ; 2.500 ppm C ; 5.000 ppm D ; 7.500 ppm E ; 10.000 ppm M1B(2)
M1C(2)
M1D(1) M1A(1)
M1B(3)
M1A(3)
M1E(1) M1B(1) M1D(2)
M1C(1) M1D(3) M1C(3) M1A(2)
M0D(1)
M0A(3) M0B(1)
M0E(1) M0D(3) M0D(2) M0B(2)
M0C(3) M0C(1) M0A(1)
M0A(2) M0E(2)
M0E(3) M0B(3)
pintu M1E(2)
M0C (2) M1E
EFEKTIVITAS CENDAWAN MIKORIZA ARBUSKULA (CMA)
TERHADAP PERTUMBUHAN DAN PRODUKTIVITAS
Cynodon dactylon (L.) Pers YANG DIBERI
LEVEL SALINITAS BERBEDA
SKRIPSI
EFEKTIVITAS CENDAWAN MIKORIZA ARBUSKULA (CMA)
TERHADAP PERTUMBUHAN DAN PRODUKTIVITAS
Cynodon dactylon (L.) Pers YANG DIBERI
LEVEL SALINITAS BERBEDA
SKRIPSI
TANTI PATRIYASARI
PROGRAM STUDI NUTRISI DAN MAKANAN TERNAK FAKULTAS PETERNAKAN
EFEKTIVITAS CENDAWAN MIKORIZA ARBUSKULA (CMA)
TERHADAP PERTUMBUHAN DAN PRODUKTIVITAS
Cynodon dactylon (L.) Pers
YANG DIBERI
LEVEL SALINITAS BERBEDA
TANTI PATRIYASARI D24102070
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
EFEKTIVITAS CENDAWAN MIKORIZA ARBUSKULA (CMA)
TERHADAP PERTUMBUHAN DAN PRODUKTIVITAS
Cynodon dactylon (L.) Pers
YANG DIBERI
LEVEL SALINITAS BERBEDA
Oleh
TANTI PATRIYASARI D24102070
Skripsi ini telah disetujui dan disidangkan di hadapan Komisi Ujian Lisan pada Tanggal 5 Juni 2006
Pembimbing Utama
Dr. Ir. Panca Dewi M.H.K.S, MSi. NIP. 131 672 157
Pembimbing Anggota
Dr. Ir. Luki Abdullah, MSc.Agr. NIP. 131 955 531
Dekan Fakultas Peternakan Institut Pertanian Bogor
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 20 September 1984. Penulis adalah putri pertama dari dua bersaudara dari pasangan Bapak Fridel Dawar dan Ibu Ratnamaida.
Pendidikan dasar diselesaikan pada tahun 1996 di SDN Mekarjaya XVIII Depok Timur, pendidikan lanjutan menengah pertama diselesaikan pada tahun 1999 di SMPN 3 Depok dan pendidikan lanjutan menengah atas diselesaikan pada tahun 2002 di SMAN 38 Jakarta.
Penulis diterima sebagai mahasiswa pada jurusan Ilmu Nutrisi dan Makanan Ternak, Fakultas Peternakan, Institut Pertanian Bogor melalui jalur Seleksi Penerimaan Mahasiswa Baru (SPMB) pada tahun 2002.
KATA PENGANTAR
Alhamdulillahirobbil’alamin. Puji syukur kehadirat Allah SWT yang telah memberikan rahmat dan karuniaNya, sehingga penulis dapat menyelesaikan skripsi dengan judul ”Efektivitas Cendawan Mikoriza Arbuskula Terhadap Pertumbuhan dan Produktivitas Cynodon dactylon (L.) Pers yang diberi Level Salinitas Berbeda”. Skripsi ini ditulis berdasarkan hasil penelitian yang penulis lakukan mulai bulan Maret sampai dengan Juli 2005. Penanaman, pemeliharaan dan pemanenan dilakukan di rumah kaca Laboratorium Lapang Agrostologi, analisa infeksi akar di Laboratorium Agrostologi Fakultas Peternakan, analisa jumlah spora di Laboratorium Bioteknologi Kehutanan dan Lingkungan, Pusat Penelitian Sumber Daya Hayati dan Bioteknologi, IPB untuk analisa jumlah spora.
Skripsi ini merupakan karya tulis yang berisi tentang upaya untuk memanfaatkan daerah pantai yang memiliki tingkat salinitas tinggi. Tingginya tingkat salinitas menyebabkan tanaman kurang dapat tumbuh dengan baik karena ketersediaan air yang rendah akibat terikat oleh senyawa-senyawa garam. Untuk mengatasi hal tersebut dapat dilakukan beberapa upaya, diantaranya adalah pemilihan jenis tanaman yang toleran terhadap kondisi salinitas dan penggunaan pupuk hayati cendawan mikoriza arbuskula (CMA). Pengetahuan tentang adanya asosiasi antara cendawan mikoriza arbuskula (CMA) dengan vegetasi pantai memberikan harapan kemungkinan pemanfaatannya pada kegiatan penghijauan didaerah pantai dimasa yang akan datang.
Akhir kata semoga skripsi ini bermanfaat bagi yang membacanya.
Bogor, Juni 2006
DAFTAR ISI
DAFTAR TABEL ... viii
DAFTAR GAMBAR ... ix
Peran Mikoriza dan Sistem Serapan P... 5
Infeksi Akar dan Spora Cendawan Mikoriza Arbuskula (CMA)... 7
Salinitas ... 8
Cendawan Mikoriza Arbuskula dan Tingkat Salinitas...9
Cynodon dactylon (L.) Pers (Rumput Bermuda)... 9
Media Tumbuh Cynodon dactylon (L.) Pers... 10
Keadaan Umum Percobaan... 17
Jumlah Stolon... 18
Jumlah Individu... 21
Produksi Berat Kering Tajuk dan Akar... 22
Berat Kering Tajuk... ... 23
Berat Kering Akar... 26
KESIMPULAN DAN SARAN ... 32
Kesimpulan ... 32
Saran ... 32
UCAPAN TERIMAKASIH ... 33
DAFTAR PUSTAKA ... 34
DAFTAR TABEL
Nomor Halaman
1. Rekapitulasi Sidik Ragam Penelitian ... 18
2. Rataan Jumlah Stolon Panen 1, 2 dan 3 Cynodon dactylon (buah) ... 19
3. Rataan Jumlah Individu Cynodon dactylon (buah) ... 21
4. Rataan Berat Kering Tajuk Panen 1, 2 dan 3 Cynodon dactylon (gram) ... ... 23
5. Rataan Berat Kering Akar Cynodon dactylon (gram)... ... 26
6. Rataan Persentase Infeksi Akar . ... ... 28
DAFTAR GAMBAR
Nomor Halaman
1. Penampang Longitudinal Akar yang Terinfeksi CMA ... 5
2. Skema Dugaan Penghambatan Pembentukan Mikoriza... 7
3. Hifa, Arbuskula dan Vesikel didalam Jaringan Korteks ... 8
4. Cynodon dactylon (L.) Pers Varietas Tifgreen ... 10
5. Cynodon dactylon Saat Penelitian ... 17
6. Perakaran yang Terinfeksi CMA ... 28
7. Perakaran yang Tidak Terinfeksi CMA ... 28