1
YANG DIINOKULASI CENDAWAN MIKORIZA
ARBUSKULA (CMA) DAN RHIZOBIUM
DI ULTISOL
A. RAHMAN SY
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2 Dengan ini saya menyatakan bahwa disertasi yang berjudul :
Respons Pertumbuhan dan Adaptasi terhadap Cekaman Kekeringan Tiga Jenis Tanaman Legum Pakan yang Diinokulasi Cendawan Mikoriza Arbuskula (CMA) dan Rhizobium di Ultisol
adalah karya saya sendiri dengan arahan komisi pembimbing, belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
3 Kekeringan Tiga Jenis Tanaman Legum Pakan yang Diinokulasi Cendawan Mikoriza Arbuskula (CMA) dan Rhizobium di Ultisol. Di bawah bimbingan SOEDARMADI HARDJOSUWIGNYO sebagai Ketua Komisi, dan SUDIRMAN YAHYA, KOMARUDDIN IDRIS, dan SRI WILARSO BUDI R sebagai Anggota Komisi.
Di daerah tropika pada umumnya, termasuk Indonesia, terdapat berbagai jenis tanaman legum pakan, tiga di antaranya yang sudah tersebar luas dan populer di kalangan petani peternak yaitu Sentro (Centrosema pubescens Benth.), Kalopo (Calopogonium mucunoides Desv.), dan Puero [Pueraria phaseoloides Roxb. (Benth.)]. Ketiga jenis tanaman legum pakan ini biasanya dibudidayakan pada tanah -tanah marginal seperti Ultisol, di mana pada tanah ini tanaman akan menghadapi masalah defisiensi unsur hara esensial tertentu, selain juga pada musim kemarau dapat mengalami kondisi kekurangan air (cekaman kekeringan). Akibatnya pertumbuhan tanaman menjadi tidak optimal. Hal ini sangat penting untuk diperhatikan, mengingat dalam pembuatan padang penggembalaan baru terutama pada tahap ”pasture establishment” (pembentukan awal padang rumput), pertumbuhan awal tanaman yang baik akan memberi jaminan terhadap pertumbuhan selanjutnya sepanjang siklus hidupnya. Penggunaan mikro -organisme bermanfaat (CMA dan Rhizobium) merupakan salah satu alternatif pemecahan masalahnya.
4 peningkatan pertumbuhan (biomassa total) tanaman Sentro 18 kali, Kalopo 8 kali, dan Puero 15 kali, sedangkan pada kadar air tanah 50 % KL peningkatan pertumbuhan tanaman secara umum lebih rendah, Sentro meningkat 8 kali dan Puero 13 kali dibandingkan tanpa inokulasi. Inokulasi Rhizobium meningkatkan pertumbuhan tanaman Sentro pada kadar air tanah 50 % KL (0.78 kali), sedangkan untuk tanaman Kalopo pada kadar air tanah 80 % KL terjadi penurunan pertumbuhan tanaman (- 0.27 kali) dibandingkan tanpa inokulasi. Inokulasi ganda Rhizobium dan CMA justru menurunkan pertumbuhan tanaman Sentro (- 0.44 kali) pada kadar air tanah 80 % KL dan (- 0. 33 kali) pada kadar air tanah 50 % KL dibandingkan inokulasi tunggal CMA. Pada kadar air tanah 50 % KL (cekaman kekeringan), tanaman yang bersimbiosis dengan CMA mengembangkan mekanisme adaptasi berupa pengurangan luas daun (Sentro, Puero) serta mempertahankan bobot kering akar (Kalopo, Puero), sedangkan tanaman Sentro yang bersimbiosis dengan Rhizobium secara tunggal tidak mengurangi luas daunnya, tetapi meningkatkan bobot kering akarnya, dan tanaman Sentro yang bersimbiosis secara ganda dengan Rhizobium dan CMA mengurangi luas daun serta bobot kering akarnya. Sebaliknya, tanaman yang tidak bersimbiosis dengan CMA (Puero), selain mengurangi luas daun dan bobot kering akarnya, juga meningkatkan kadar prolina daun.
Hasil penelitian di lapangan menunjukkan bahwa meskipun inokulasi tunggal CMA atau Rhizobium dan inokulasi ganda CMA dan Rhizobium belum mampu meningkatkan biomassa total tanaman, tatapi inokulasi tunggal CMA mampu meningkatkan rataan bobot kering akar ketiga jenis tanaman legum pakan (Sentro, Kalopo, dan Puero) yaitu sebesar 0.92 kali dibandingkan tanaman tanpa inokulasi. Inokulasi tunggal Rhizobium mampu meningkatkan rataan bobot kering akar ketiga jenis tanaman legum pakan (Sentro, Kalopo, dan Puero) yaitu sebesar 1.09 kali dibandingkan tanaman tanpa inokulasi. Inokulasi ganda CMA dan Rhizobium mampu meningkatkan rataan bobot kering akar ketiga jenis tanaman legum pakan (Sentro, Kalopo, dan Puero) yaitu sebesar 0.66 kali dibandingkan tanaman tanpa inokulasi.
Dari kedua penelitian ini yaitu penelitian di rumah kaca dan penelitian di lapangan terbukti bahwa inokulasi CMA mampu meningkatkan pertumbuhan ketiga jenis tanaman legum pakan (Sentro, Kalopo, dan Puero), baik pada kadar air 80 % KL maupun 50 % KL (kondisi cekaman kekeringan). Selain itu, keberadaan CMA (simbiosis CMA dan tanaman) juga dapat mempengaruhi mekanisme adaptasi tanaman terhadap kondisi kekeringan .
5 A. RAHMAN SY 2006. Growth Response and Adaptation to Drought Stress of Three Forage Legume Crops Inoculated with Arbuscular Mycorrhizal Fungi (AM-fungi) and Rhizobium at Ultisol. Supervised by SOEDARMADI HARDJOSUWIGNYO as chairman, SUDIRMAN YAHYA, KOMARUDDIN IDRIS, and SRI WILARSO BUDI R. as members of the advisory committee.
In general, there are several forage legume crops in the tropics, including Indonesia, three of them i.e. Centro (Centrosema pubescens Benth.), Calopo (Calopogonium mucun oides Desv.), and Puero [Pueraria phaseoloides Roxb. (Benth.)] are familiar and widely used by the farmers. The three tropical forage legumes usually grown on marginal soils such as Ultisol, where the plants face some essential nutrient deficiencies, and also soil water deficit in the dry season. The result is that the plants can not reach their optimal growth. It is very urgent issue that in the development of newly grassland program especially for pasture establishment, the early well growth of plants guarantees the long terms growth in the their life cycles. Using the useful microorganisms (AM-fungi and Rhizobium) is one of the alternative solution.
The research was conducted in two experiments, the green house and field experiments. The objectives of the experiments were 1) to evaluate the effectiveness of AM-fungi and Rhizobium inoculation on growth of three forage legume crops (Centro, Calopo, and Puero) at 80 % and 50 % soil water content, 2) to determine the adaptation mechanisms of three forage legume crops inoculated with AM-fungi and or Rhizobium on drought stress (at 50 % soil water content), 3) to prove the effectiveness of AM-fungi and Rhizobium inoculation on growth of three forage legume crops in the non-steril soil. The two parallel experiments were conducted from September 2005 to April 2006. In the green house experiment, three factors treatments were assigned in completely randomized design with three replications. The first factor : three forage legume species (Centro, Calopo, and Puero ), the second factor : two levels water content (80 % and 50 % of field capacity), and the third factor : four inoculation treatments (noninoculated, inoculated with Rhizobium, inoculated with AM-fungi, inoculated with Rhizobium and AM-fungi). In the field experiment, the treatments were assigned in split plot design, three forage legume species as the main plots and inoculation treatments as the subplots. Growth variables for green house experiment were : plant length, total biomass, shoot dry weight, root dry weight, leaf area, proline content, amount of spores per 50 g soil, root colonization, amount of nodules, nitrogen content, and growth variables for field experiment were : total biomass, shoot dry weight, root dry weight, amount of spores per 50 g soil, root colonization, amount of nodules, nitrogen content, and phosphor content.
6 content (drought stress), the mycorrhizal plants developed adaptation mechanism i.e. decreased their leaf area (Sentro, Puero) and maintenanced their root dry weight (Kalopo, Puero). Sentro inoculated with single Rhizobium did not decreas e its leaf area, and increased its root dry weight, but for Sentro inoculated with both Rhizobium and AM-fungi decreased their leaf area and root dry weight. In the other hands the nonmycorrhizal plants (Puero), decreased their leaf area and root weight, and increased their proline content in the leaf.
Field experiment showed that eventhough single AM-fungi or Rhizobium inoculation and both AM-fungi and Rhizobium inoculation did not affect total plant biomass, but the mean growth of three forage legume species (Centro, Calopo, and Puero) were increased in terms of root dry matter i.e. 0.92 folds for plants inoculated with AM-fungi, 1.09 folds for plants inoculated with Rhizobium, and 0.66 folds for plants inoculated with both Rhizobium and AM-fungi.
The two experiments (green house and field experiments) proved that AM-fungi inoculation can increase the growth of three forage legume crops (Centro, Calopo, and Puero), both at 80 % and 50 % soil water cointent. Other result, that the simbiosis between AM-fungi and plant can affect the adaptation mechanism to drought stress.
7 © Hak cipta milik Institut Pertanian Bogor, 2006
Hak cipta dilindungi
8
YANG DIINOKULASI CENDAWAN MIKORIZA
ARBUSKULA (CMA) DAN RHIZOBIUM
DI ULTISOL
A. RAHMAN SY
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Agronomi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
9
Nama : A. Rahman Sy
NIM : 995190
Disetujui
Komisi Pembimbing
Prof.Dr.Ir.H. Soedarmadi H., M.Sc. Prof.Dr.Ir.H. Sudirman Yahya, M.Sc. Ketua Anggota
Dr. Ir. Komaru ddin Idris, M.S. Dr. Ir. Sri Wilarso Budi R., M.S. Anggota Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Dr. Ir. Satriyas Ilyas, M.S. Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
10 Puji dan syukur penulis panjatkan ke hadirat Allah SWT, Tuhan semesta alam, atas segala karunia, rahmat dan hidayah -Nya sehingga penulis dapat menyelesaikan penelitian disertasi ini. Tema yang dipilih dalam penelitian yang dilaksanakan di rumah kaca dan kebun percobaan kampus Universitas Jambi (UNJA) Mendalo Darat ini adalah Pemanfaatan Cendawan Mikoriza Arbuskula (CMA) dan Rhizobium pada Tanaman Legum Pakan, dengan judul “Respons Pertumbuhan dan Adaptasi terhadap Cekaman Kekeringan Tiga Jenis Tanaman Legum Pakan yang Diinokulasi Cendawan Mikoriza Arbuskula (CMA) dan Rhizobium di Ultisol”. Adapun waktu pelaksanaan penelitian ini yaitu sejak bulan Oktober 2004 sampai bulan April 2006.
Selama pelaksanaan penelitian, penulis mendapat banyak bantuan dan bimbingan dari berbagai pihak. Untuk itu pada kesempatan ini penulis ingin menyampaikan ucapan terima kasih kepada :
1. Prof. Dr. Ir. H. Soedarmadi Hardjosuwignyo, M.Sc. selaku Ketua Komisi Pembimbing, dan Prof. Dr. Ir. H. Sudirman Yahya, M.Sc., Dr. Ir. Komaruddin Idris, MS., Dr. Ir. Sri Wilarso Budi R., MS. selaku Anggota Komisi Pembimbing atas segala bantuan, bimbingan dan arahan yang telah diberikan selama ini.
2. Pimpinan dan Staf Sekolah Pascasarjana, Pimpinan dan Staf Program Studi Agronomi Sekolah Pascasarjana Institut Pertanian Bogor yang telah memberi kesempatan kepada penulis mengikuti program doktor serta segala kemudahan administrasi yang diberikan.
3. Pimpinan dan Staf Proyek DUE II Universitas Jambi, Walikota Jambi, Badan Amil Zakat, Infak dan Sadak ah (Bazis) Kota Jambi atas bantuan sebagian dana penelitian yang diberikan kepada penulis.
11 Laboratorium Pengolahan Hasil Ternak, Laboratorium Nutrisi Ruminans ia, Laboratorium Kimia Tanah, Laboratorium Pusat Studi Pemuliaan Tanaman IPB, Laboratorium Pusat Penelitan Tanah Bogor, Kepala Kebun Percobaan Fakultas Pertanian atas kesediaan analisis tanaman, tanah dan izin menggunakan laboratorium serta lahan percobaan . 6. Rekan-rekan sejawat di Laboratorium Hijauan Makanan Ternak,
khususnya Bapak Ir.Ubaidillah MP atas bantuan dalam pengolahan data, Bapak Ir. Ahmad Yani MP atas izin menggunakan kamera digital.
7. Bapak Dr. Ir. Dadang Sukandar MSc. (staf pengajar Departemen Gizi Masyarakat dan Keluarga) IPB yang telah membantu dalam menafsirkan data secara statistik, serta rekan-rekan seperjuangan : Dr. Ir. Hamzah MS, Ir. Asmadi Saad MS, Ir. Suandi MS, Haryadi SE MSc, Arman Delis SE, MS, Ir. Yatno MS, Ir. Hutwan Syarifudin MS, Ir. Suryono MS, Ir. Jalius MS, Ir. Rainiyati MS, Dr. Ir.Elis Kartika MS, Rahmi Dianita SPt. MSc, Sofia San di, SPt., MSi, Nurul Febriani, SPt, Meisji Liana, SPt., MS, Ir. Lizawati MS, dan Ir. Henny MS.
8. Rekan-rekan di Laboratorium Bioteknologi Tanaman, khususnya Bapak Ir. Dede Martino MP, Sdr. Ir. Efrizal, Sdr. Era Wahyudi SP, Mbak Tuti Kurniati SP, dan Ayuk Bariyah.
9. Mahasiswa : Dedi, Nining, Dorma, Wahyuni, Anisa, Alha, Ayu, dan Lina yang telah membantu di laboratorium, di rumah kaca, dan di lapangan. 10.Kakak, adik, keponakan, dan semua keluarga yang telah membantu
khususnya di lapangan : Wak Nizar, Arbain, dan Rahmaini SP.
11.Yang mulia ayahanda Syarifuddin, yang tercinta isteriku Sahinah serta anak-anak tersayang Riza, Ifadah, dan Lisa yang telah memberi inspirasi, bersabar dan berdoa, demi kesuksesan penulis.
Harapan penulis, semoga karya ilmiah ini dapat bermanfaat bagi perkembangan ilmu pengetahuan khususnya di bidang hijauan makanan ternak.
12 Penulis dila hirkan di Teluk Rendah, Kabupaten Tebo Jambi pada tanggal 13 Pebruari 1959, adalah anak kedua dari pasangan Syarifuddin dan Zubaidah (alm). Pada tanggal 14 April 1985 penulis menikah dengan Sahinah, dan saat ini telah dikaruniai tiga orang anak yaitu Muhammad Riza Alfajri (20 tahun), Rafiqotul Ifadah (17 tahun), dan Raihani Elkhalishah (12 tahun).
Penulis menyelesaikan pendidikan Sekolah Dasar Negeri No. 10/IV Tahtul Yaman Jambi tahun 1971, Sekolah Menengah Pertama Negeri I Jambi tahun 1974, dan Sekolah Menengah Tingkat Atas Negeri I Jambi tahun 1977.
Pendidikan sarjana ditempuh di Jurusan Nutrisi dan Makanan Ternak, Fakultas Peternakan Universitas Jambi, lulus pada tahun 1984. Pada tahun 1990 penulis meneruskan pendidikan program master pada divisi Crop P roduction and Management, Agronomy Departement, University of the Philippines Los Banos (UPLB) di Filipina, dengan beasiswa dari SEAMEO -SEARCA, dan tamat tahun 1993. Selanjutnya pada tahun 1999, penulis mendapat kesempatan melanjutkan pendidikan program doktor pada Program Studi Agronomi, Sekolah Pascasarjana Institut Pertranian Bogor (IPB) dengan beasiswa dari proyek DUE II Universitas Jambi.
13 Halaman
DAFTAR TABEL ... vii
DAFTAR GAMBAR ... viii
DAFTAR LAMPIRAN ... ix
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 4
Manfaat Penelitian ... 5
Hipotesis ... 5
Strategi Penelitian ... 6
TINJAUAN PUSTAKA ... 8
Sifat Tanah Ultisol ... 8
Deskripsi Tanaman Legum Pakan ... 9
Mikoriza ... 12
Bakteri Bintil Akar ... 17
Simbiosis Tripartit ... 25
Respons Tanaman terhadap Cekaman Kekeringan ... 27
PENGUJIAN KEEFEKTIFAN CMA DAN RHIZOBIUM TERHADAP PERTUMBUHAN TANAMAN LEGUM PAKAN PADA KADAR AIR TANAH YANG BERBEDA ... 31
Abstrak ... 31
Abstract ... 32
Pendahuluan ... 32
Bahan dan Metode ... 34
Hasil dan Pembahasan ... 38
14
Abstrak ... 62
Abstract ... 62
Pendahuluan ... 63
Bahan dan Metode ... 64
Hasil dan Pembahasan ... 68
Kesimpulan ... 80
PEMBAHASAN UMUM ... 81
Peranan CMA dan Rhizobium dalam Peningkatan Produktivitas Tanaman Legum Pakan ... 81
Aplikasi Penggunaan Teknologi Inokulasi CMA dan Rhizobium dalam Pembuatan Padang Penggembalaan ... 85
KESIMPULAN UMUM DAN SARAN ... 85
Kesimpulan Umum ... 85
Saran ... 86
DAFTAR PUSTAKA ………. 87
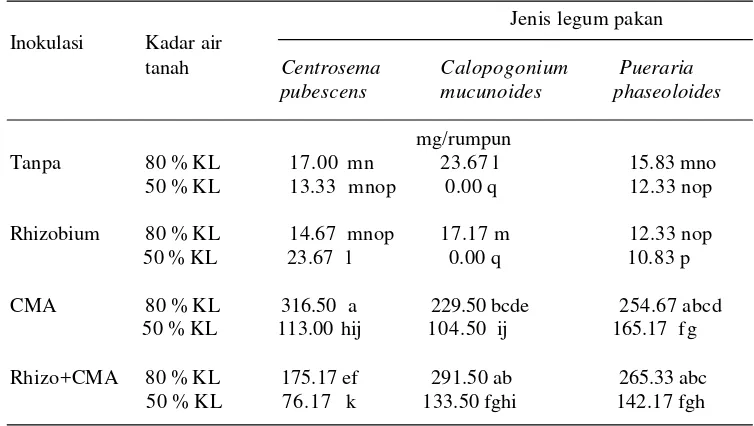
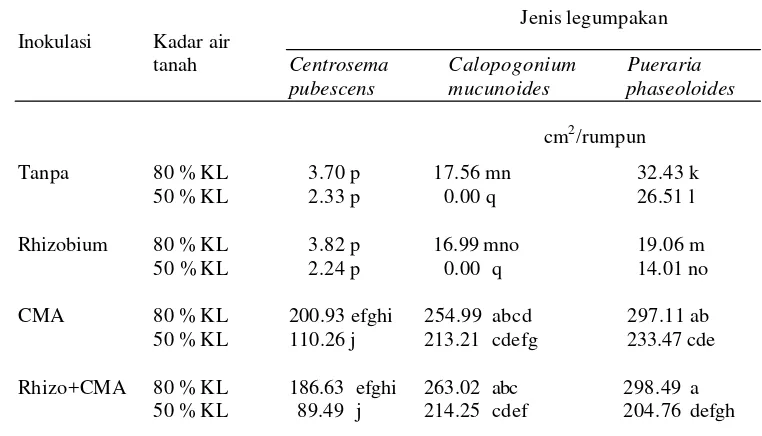
15 Halaman 1. Pengaruh inokulasi CMA dan Rhizobium terhadap biomassa total
tiga jenis tanaman legum pakan pada kadar air tanah yang berbeda ……… 41 2. Pengaruh inokulasi CMA dan Rhizobium terhadap bobot kering tajuk
tiga jenis tanaman legum pakan pada kadar air tanah yang berbeda ……… 42 3. Pengaruh inokulasi CMA dan Rhizobium terhadap luas daun
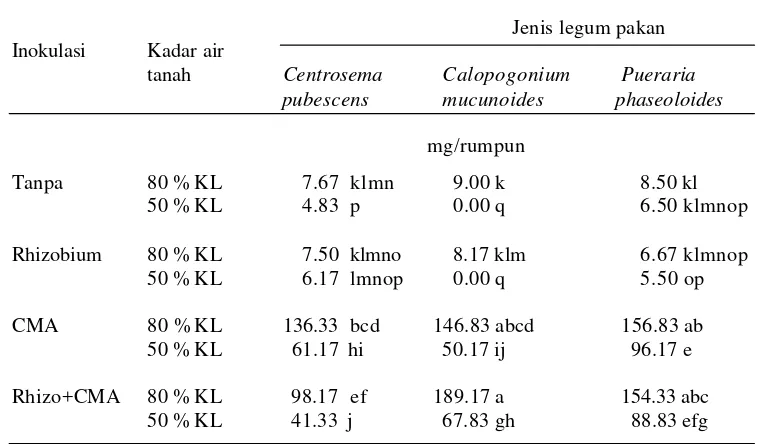
tiga jenis tanaman legum pakan pada kadar air tanah yang berbeda ……… 49 4. Pengaruh inokulasi CMA dan Rhizobium terhadap bobot kering akar
tiga jenis tanaman legum pakan pada kadar air tanah yang berbeda ……… 50 5. Pengaruh inokulasi CMA dan Rhizobium terhadap biomassa total
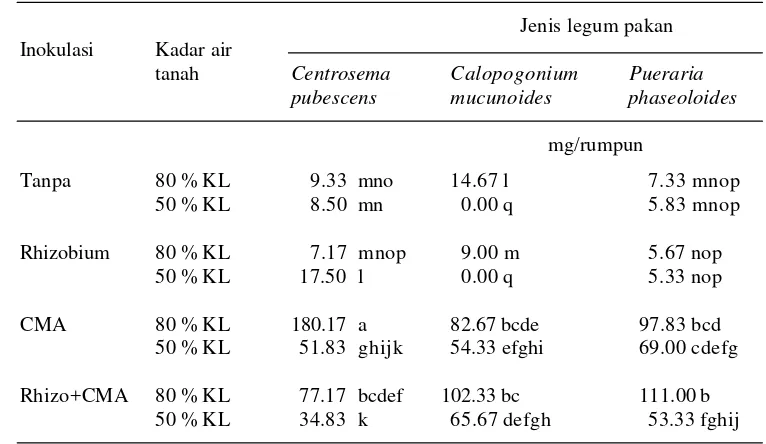
tiga jenis tanaman legum pakan ……… 68 6. Pengaruh inokulasi CMA dan Rhizobium terhadap bobot kering tajuk
tiga jenis tanaman legum pakan ……….. 69 7. Pengaruh inokulasi CMA dan Rhizobium terhadap bobot kering akar
tiga jenis tanaman legum pakan ……… 70 8. Pengaruh inokulasi CMA dan Rhizobium terhadap infeksi akar
tiga jenis tan aman legum ……….. 74 9. Pengaruh inokulasi CMA dan Rhizobium terhadap jumlah spora
per 50 g tanah tiga jenis tanaman legum pakan ……… 75 10. Pengaruh inokulasi CMA dan Rhizobium terhadap kadar nitrogen
daun tiga jenis tanaman legum pakan ….……… 78 11. Pengaruh inokulasi CMA dan Rhizobium terhadap kadar fosfor daun
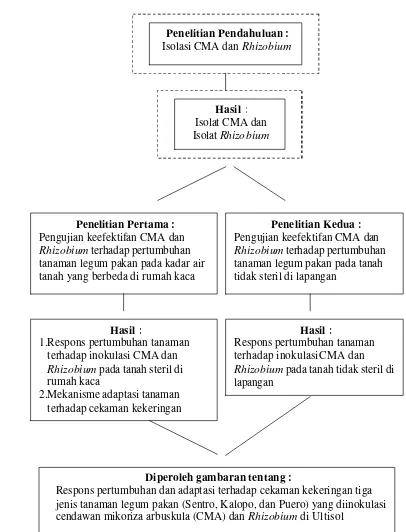
16 Halaman 1. Bagan alir penelitian respons pertumbuhan dan adaptasi terhadap
cekaman kekeringan tiga jenis tanaman legum pakan yang diinokulasi
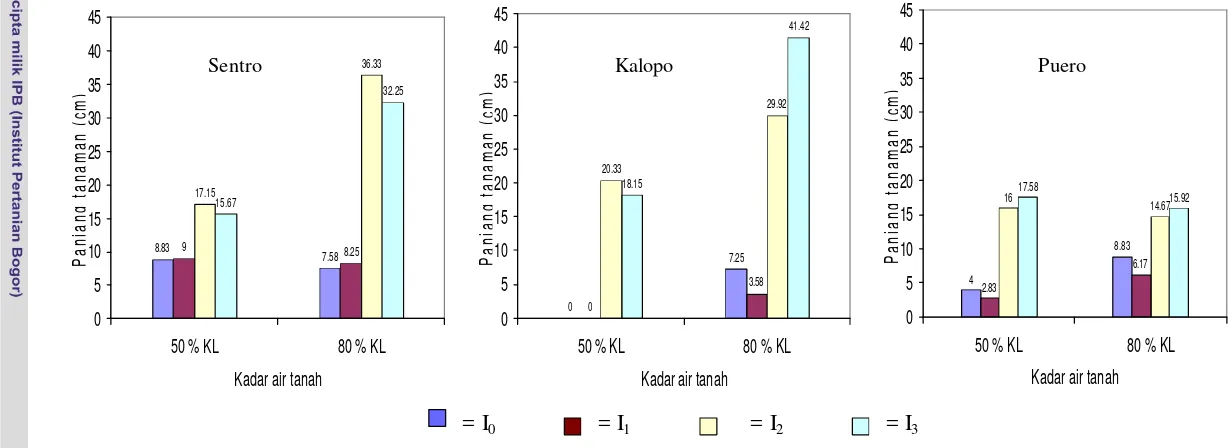
cendawan mikoriza arbuskula (CMA) dan Rhizobium di Ultisol ……. 7 2. Panjang tanaman ketiga jenis tanaman legum pakan sebagai respons
terhadap perlakuan tanpa inokulasi (I0), inokulasi Rhizobium (I1), inokulasi
CMA (I2), dan inokulasi Rhizobium dan CMA (I3) pada kadar air
tanah yang berbeda ………... 40 3. Penampilan morfologi Sentro empat bulan setelah tanam sebagai respons terhadap perlakuan : tanpa inokulasi (L1A1I0), inokulasi Rhizobium (L1A1I1),
inokulasi CMA (L1A1I2), dan inokulasi Rhizobium dan CMA (L1A1I3) pada
kadar air tanah 80 % KL (A1) dan kadar air tanah 50 % KL (A2) ……... 43
4. Penampilan morfo logi Kalopo empat bulan setelah tanam sebagai respons terhadap perlakuan : tanpa inokulasi (L2A1I0), inokulasi Rhizobium (L2A1I1),
inokulasi CMA (L2A1I2), dan inokulasi Rhizobium dan CMA (L2A1I3) pada
kadar air tanah 80 % KL (A1) dan kadar air tanah 50 % KL (A2) ……... 44
5. Penampilan morfologi Puero empat bulan setelah tanam sebagai respons terhadap perlakuan : tanpa inokulasi (L3A1I0), inokulasi Rhizobium (L3A1I1),
inokulasi CMA (L3A1I2), dan inokulasi Rhizobium dan CMA (L3A1I3) pada
kadar air tanah 80 % KL (A1) dan kadar air tanah 50 % KL (A2) …… 45
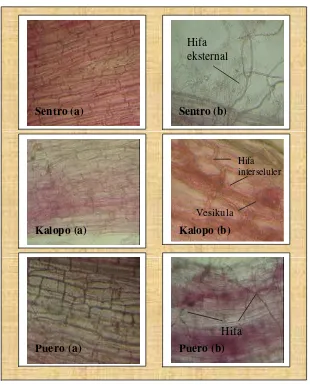
6. Akar tanaman legum pakan (korteks akar) yang tidak terinfeksi (a)
dan yang terinfeksi (b) dengan pembesaran 400 x ..……….. 46 7. Kad ar prolina daun ketiga jenis tanaman legum pakan sebagai respons
terhadap perlakuan tanpa inokulasi (I0), inokulasi Rhizobium (I1), inokulasi
CMA (I2), dan inokulasi Rhizobium dan CMA (I3) pada kadar air
tanah yang berbeda ………... 52 8. Bintil akar yang terbentuk pada tanaman Sentro dan Puero,
tetapi tidak terbentuk pada tanaman Kalopo ………. 56 9. Penampilan morfologi Sentro empat bulan setelah tanam sebagai respons
terhadap perlakuan tanpa inokulasi (L1I0), inokulasi Rhizobium (L1I1),
inokulasi CMA (L1I2), dan inokulasi Rhizobium dan CMA (L1I3) ……… 71
10. Penampilan morfologi Kalopo empat bulan setelah tanam sebagai respons terhadap perlakuan tanpa inokulasi (L2I0), inokulasi Rhizob ium (L2I1),
17 12.Infeksi akar oleh CMA pada ketiga jenis tanaman legum pakan,
(a) yang mendapat perlakuan inokulasi, (b) tanpa inokulasi CMA …… 74 13.Tipe spora CMA asli (Glomus sp-1a, sp -2a, sp -3a, sp -4a, dan
Gigaspora sp) ………. 76 14.Tipe spora CMA introduksi (Glomus sp-1, sp -2, sp-3, sp -4) …………. 76 15.Bintil akar yang terbentuk pada tanaman Sentro dan Puero, tetapi
18
Halaman
1. Analisis sifat-sifat tanah percobaan ……… 99 2. Data curah hujan, suhu dan radiasi selama pelaksanaan
pengamatan di lapangan ... 100 3. Ekstraksi spora teknik tuang aring basah menurut Pacioni (1992) dan
dilanjutkan teknik sentrifugasi menurut Brudrett et al. (1996) ……... 102 4. Pengamatan kolonisasi CMA pada akar tanaman dengan cara
pewarnaan akar (metode Kormanik dan McGraw, 1982) dan penghitungan akar yang terinfeksi dengan metode panjang slide
menurut Giovannetti dan Mosse (1980) ... 103 5. Prosedur pengukuran kadar prolina (metode Bates et al. 1973) ……... 104 6. Prosedur penentuan kadar nitrogen daun (metode Kjeldahl) ...……. 105 7. Prosedur penentuan kapasitas lapang tanah percobaan, metode
grafimetri (Somasegaran dan Hoben 1994) ... 106 8. Analisis ragam panjang tanaman tiga jenis tanaman legum pakan
yang diinokulasi CMA dan Rhizobium pad a kadar air tanah
yang berbeda ...… 108 9. Analisis ragam biomassa total tiga jenis tanaman legum pakan
yang diinokulasi CMA dan Rhizobium pada kadar air tanah
yang berbeda ...… 108 10. Analisis ragam bobot kering tajuk tiga jenis tanaman legum pakan
yang diinokulasi CMA dan Rhizobium pada kadar air tanah
yang berbeda ...…. 109 11. Analisis ragam luas daun tiga jenis tanaman legum pakan
yang diinokulasi CMA dan Rhizobium pada kadar air tanah
yang berbeda ... 109 12. Analisis ragam bobot kering akar tiga jenis tanaman legum
pakan yang diinokulasi CMA dan Rhizobium pada kadar
air tanah yang berbeda ...…. 110 13. Analisis ragam biomassa total tiga jenis tanaman legum pakan
yang diinokulasi CMA dan Rhizobium ...……….. 110 14. Analisis ragam bobot kering tajuk tiga jenis tanaman legum pakan
19 16. Analisis ragam infeksi akar tiga jenis tanaman legum pakan
yang diinokulasi CMA dan Rhizobium ..……… 112 17. Analisis ragam jumlah spora per 50 g tanah tiga jenis tanaman
legum pakan yang diinokulasi CMA dan Rhizobium …….………… 112 18. Analisis ragam kadar nitrogen tiga jenis tanaman legum pakan
yang diinokulasi CMA dan Rhizobium ...……….………. 113 19. Analisis ragam kadar fosfor tiga jenis tanaman legum pakan
yang diinokulasi CMA dan Rhizob ium ...………. 113 20. Karakteristik tipe spora introduksi (inokulum) yang diberikan
pada tanaman legum pakan, berasal dari rizosfer tanaman pisang raja nangka, Kabupaten Merangin,
Provinsi Jambi (Rainiyati 2004) ... 114 21. Karakteristik tipe spora asli yang diisolasi dari tanah lapisan atas
Kebun Percobaan Fakultas Pertanian Universitas Jambi
Latar Belakang
Sejalan dengan peningkatan kesejahteraan dan kenaikan pendapatan masyarakat, kesadaran akan pentingnya nilai gizi dari makanan yang dikonsumsi juga semakin meningkat. Hal ini membawa konsekuensi kepada peningkatan konsumsi protein hewani, sehingga perlu diimbangi dengan peningkatan produktivitas ternak secara umum, termasuk di antaranya ternak ruminansia seperti sapi, kerbau, kambing dan domba. Peningkatan produktivitas ternak ruminansia ini harus ditunjang oleh adanya jaminan penyediaan pakan dalam arti baik produksi, kualitas, maupun kontinuitas ketersediaannya. Kendala umum yang ditemui dalam penyediaan pakan ternak ruminansia di daerah tropika yaitu rendahnya produktivitas tanaman pakan.
Tanaman pakan secara garis besar terdiri atas jenis rumput (famili Gramineae) dan legum (famili Leguminosae). Tanaman legum pakan tropika yang sudah tersebar luas dan populer di kalangan patani peternak antara lain adalah Sentro (Centrosema pubescens Benth.), Kalopo (Calopogonium mucunoides Desv.) dan Puero [Pueraria phaseoloides Roxb.(Benth.)].
Sentro adalah tanaman legum pakan penting, disukai ternak (palatable), merupakan spesies pertama yang digunakan secara luas untuk perbaikan padang penggembalaan karena sangat produktif dengan kualitas hijauan yang tinggi. Tanaman ini mampu tumbuh baik sebagai tanaman campuran dengan spesies rumput (Bogdan 1977), tetapi tidak dapat tumbuh baik pada tanah dengan kandungan Al dan Mn tinggi atau bila pH di bawah 5.0. Sentro dapat bertahan pada curah hujan tahunan 750 mm (Kretschmer dan Pitman 2001), serta tahan terhadap kekeringan (Bogdan 1977), tetapi lebih produktif bila menerima curah hujan tahunan 1.000 mm atau lebih (Kretschmer dan Pitman 2001).
Puero termasuk tanaman legum pakan yang lebih palatable dibandingkan Kalopo. Tanaman ini dapat tumbuh baik di daerah tropika basah dengan curah hujan tahunan 1.200 – 1.500 mm, dan tahan terhadap periode kering yang singkat sekitar 2 – 3 bulan serta toleran terhadap tanah masam (pH 4.0 – 5.5) dan tanah yang kekurangan P (Bogdan 1977; Whiteman 1980). Puero dapat membentuk tanaman campuran dengan beberapa spesies rumput pakan seperti Brachiaria mutica, Panicum maximum, Pennisetum purpureum, Melinis minutiflora,
Hyparrhenia rufa, dan Digitaria decumbens (Bogdan 1977).
Meskipun potensi pertumbuhan tanaman legum pakan daerah tropika tidak jauh berbeda dibandingkan tanaman legum pakan daerah iklim sedang (temperate), tetapi apabila tumbuh pada kondisi lingkungan yang tidak menunjang maka potensi tersebut menjadi terhambat. Keadaan demikian selain disebabkan oleh faktor genetik tanaman itu sendiri, juga berkaitan dengan faktor lingkungan seperti rendahnya tingkat kesuburan tanah. Menurut Beinroth (2001) kendala produksi tanaman pakan yang berhubungan dengan sumberdaya lahan di daerah tropika antara lain adalah defisiensi unsur hara, kemasaman dan toksisitas, serta kekurangan kandungan air tanah.
Di daerah tropika umumnya termasuk Indonesia, tanah yang digunakan untuk budidaya tanaman legum pakan seperti Sentro, Kalopo, dan Puero biasanya adalah tanah -tanah marginal. Menurut Hardjowigeno (1995) tanah marginal banyak dijumpai di luar pulau Jawa terutama di Sumatera, Kalimantan, Sulawesi, dan Irian Jaya yaitu didominasi oleh Ultisol (disebut juga tanah Podzolik Merah Kuning). Ultisol termasuk ke dalam kategori tanah masam yaitu dengan ciri: reaksi masam, unsur hara rendah, kadar bahan organik rendah, kejenuhan basa rendah, tetapi kadar besi, aluminium dan mangan tinggi (Sudjadi 1984; Hardjowigeno 1995).
(Mengel dan Kirkby 2001), sehingga tanaman mudah mengalami kondisi kekurangan air (cekaman kekeringan). Hal ini terutama terjadi di musim kemarau , baik yang mempunyai pola kekeringan secara periodik pada daerah dengan tipe iklim tropika basah ataupun yang mempunyai pola kekeringan selama beberapa bulan secara terus-menerus pada daerah dengan tipe iklim tropika basah kering dan tipe iklim tropika kering.
Untuk mengatasi faktor yang menghambat pertumbuhan tanaman legum pakan pada Ultisol, selama ini dilakukan upaya pengapuran dan pemupukan dengan menggunakan pupuk buatan seperti pupuk nitrogen (Urea), fosfor (TSP atau SP-36) dan kalium (KCl). Meskipun pupuk buatan ini dapat meningkatkan pertumbuhan tanaman pakan secara signifikan, namun sistem produksi pupuk buatan tersebut membutuhkan masukan dengan biaya tinggi dan tidak dapat diperbarui (non -renewable) serta penggunaannya dalam jangka panjang mempunyai dampak merugikan bagi kelestarian lingkungan. Oleh karena itu perlu dicarikan pupuk alternatif dengan sistem produksi ramah lingkungan yang menunjang sistem pertanian berkelanjutan serta dapat memberikan harapan untuk memenuhi kebutuhan di masa datang, yaitu pupuk hayati berupa cendawan mikoriza arbuskula (CMA) dan Rhizobium. Karena peran CMA selain dapat meningkatkan pertumbuhan tanaman, juga mampu meningkatkan ketahanan tanaman terhadap cekaman kekeringan, maka penggunaan CMA ini juga memungkinkan untuk mengatasi kendala kekurangan air yang dihadapi tanaman legum pakan pada kondisi kekeringan (musim kemarau).
kekeringan (Nelson dan Safir 1982; Setiadi 1989; Brundrett et al. 1996; Al-Karaki dan Al-Raddad 1997; Al-Al-Karaki et al. 1998; Aboul-Nasr 1998; Al-Karaki dan Clark 1999).
Kenyataan di lapangan menunjukkan bahwa tanaman legum yang membentuk bintil akar, biasanya juga dikolonisasi oleh cendawan mikoriza arbuskula (Barea et al., 1992; Fitter dan Garbaye 1994), sehingga terjadi suatu hubungan atau simbiosis tripartit antara tiga jenis organisme yaitu tanaman legum, bakteri Rhizobium, dan cendawan mikoriza arbuskula (CMA). Beberapa penelitian (Aries et al.1991; Soliman et al. 1996; Sylvia 1998) menunjukkan bahwa inokulasi Rhizobium dan cendawan mikoriza arbuskula mampu meningkatkan pertumbuhan, kandungan N dan P tanaman pangan kedelai dan tanaman legum pakan tropika.
Namun penelitian-penelitian tentang peranan CMA dan Rhizobium tersebut pada umumnya dilakukan di rumah kaca di mana kondisi penelitian lebih
terkontrol dengan media tanam biasanya steril. Untuk aplikasi di lapangan, di mana kondisi penelitian kurang atau bahkan tidak dapat dikontrol dengan media tanam (tanah) tidak steril, kemungkinan besar hasil yang diperoleh akan berbeda dengan apa yang telah terbukti.
Berdasarkan alasan di atas, dilakukan penelitian tentang respons pertumbuhan dan adaptasi terhadap cekaman kekeringan tiga jenis tanaman legum pakan yaitu Sentro, Kalopo, dan Puero yang diinokulasi CMA dan Rhizobium di Ultisol, yang dilakukan di rumah kaca dan di lapangan.
Tujuan Penelitian
Penelitian ini bertujuan untuk :
1. Menguji keefektifan inokulasi CMAdan Rhizobium terhadap pertumbuhan tiga jenis tanaman legum pakan (Sentro, Kalopo dan Puero) pada kadar air tanah 80 % kapasitas lapang (KL) dan 50 % KL di rumah kaca.
3. Membuktikan keefektifan inokulasi CMA dan Rhizobium terhadap pertumbuhan tiga jenis tanaman legum pakan (Sentro, Kalopo dan Puero) pada tanah tidak steril di lapangan.
Manfaat Penelitian
Manfaat yang dapat diperoleh dari penelitian ini adalah :
1. Dengan ditemukannya inokulum CMA dan Rhizobium yang efektif pada tanaman legum pakan, maka dapat digunakan untuk program peningkatan produktivitas tanaman legum pakan, khususnya Sentro, Kalopo, dan Puero.
2. Dengan diketahuinya mekanisme adaptasi tiga jenis tanaman legum pakan (Sentro, Kalopo dan Puero) terhadap cekaman kekeringan, diharapkan dapat menjadi dasar pertimbangan dalam memanipulasi teknik budidaya khususnya untuk mengatasi kondisi musim kemarau.
3. Hasil penelitian ini dapat memberikan informasi sebagai dasar pertimbangan penggunaan pupuk hayati (teknologi inokulasi CMA dan Rhizobium) dalam rangka pengelolaan padang rumput atau padang penggembalaan.
Hipotesis
Hipotesis yang diajukan dalam penelitian ini adalah :
1. Inokulasi CMA dan Rhizobium baik secara tunggal maupun ganda dapat meningkatkan pertumbuhan tiga jenis tanaman legum pakan (Sentro, Kalopo dan Puero), baik pada kadar air tanah 80 % KL maupun 50 % KL (cekaman kekeringan).
2. Mekanisme adaptasi setiap jenis tanaman legum pakan terhadap cekaman kekeringan dipengaruhi oleh interaksi tanaman dengan CMA dan atau Rhizobium.
Strategi Penelitian
Disertasi ini disusun berdasarkan hasil penelitian yang terdiri atas dua penelitian utama yaitu di rumah kaca dan di lapangan. Sebelumnya, dilakukan Penelitian Pendahuluan yaitu “Isolasi CMA dan Rhizobium”, yang bertujuan untuk memperoleh isolat CMA dan Rhizobium yang akan digunakan sebagai inokulum untuk penelitian di rumah kaca dan di lapangan.
Penelitian pertama yaitu penelitian di rumah kaca dengan judul “Pengujian Keefektifan CMA dan Rhizobium terhadap Pertumbuhan Tanaman Legum Pakan pada Kadar Air Tanah yang Berbeda”, bertujuan untuk : 1) membuktikan keefektifan CMA dan Rhizobium terhadap pertumbuhan tiga jenis tanaman legum pakan (Sentro, Kalopo, dan Puero) pada kadar air tanah 80 % KL dan 50 % KL (cekaman kekeringan), 2) mengetahui mekanisme adaptasi tiga jenis tanaman legum pakan terhadap cekaman kekeringan, baik yang mendapat perlakuan tanpa inokulasi, inokulasi tunggal CMA atau Rhizobium, maupun inokulasi ganda CMA dan Rhizobium.
Penelitian kedua dilaksanakan di lapangan dengan judul “Pengujian Keefektifan CMA dan Rhizobium terhadap Pertumbuhan Tanaman Legum Pakan pada Tanah Tidak Steril di Lapangan”, bertujuan untuk membuktikan keefektifan CMA dan Rhizobium terhadap pertumbuhan tiga jenis tanaman legum pakan (Sentro, Kalopo, dan Puero) pada tanah tidak steril di lapangan.
Gambar 1. Bagan alir penelitian respons pertumbuhan dan adaptasi terhadap cekaman kekeringan tiga jenis tanaman legum pakan yang diinokulasi cendawan mikoriza arbuskula (CMA) dan Rhizobium di Ultisol. Penelitian Pertama :
Pengujian keefektifan CMAdan Rhizobium terhadap pertumbuhan tanaman legum pakan pada kadar air tanah yang berbeda di rumah kaca
Penelitian Kedua : Pengujian keefektifan CMAdan Rhizobium terhadap pertumbuhan tanaman legum pakan pada tanah tidak steril di lapangan
Diperoleh gambaran tentang :
Respons pertumbuhan dan adaptasi terhadap cekaman kekeringan tiga jenis tanaman legum pakan (Sentro, Kalopo, dan Puero) yang diinokulasi cendawan mikoriza arbuskula (CMA) dan Rhizobium di Ultisol
Hasil :
1.Respons pertumbuhan tanaman terhadap inokulasi CMAdan Rhizobium pada tanah steril di rumah kaca
2.Mekanisme adaptasi tanaman terhadap cekaman kekeringan
Hasil :
Respons pertumbuhan tanaman terhadap inokulasiCMA dan Rhizobium pada tanah tidak steril di lapangan
Penelitian Pendahuluan : Isolasi CMA dan Rhizobium
TINJAUAN PUSTAKA
Sifat Tanah Ultisol
Lahan yang digunakan untuk budidaya tanaman pakan atau pembuatan padang penggembalaan biasanya adalah lahan yang tidak subur dan termasuk kategori lahan marginal. Ultisol termasuk salah satu lahan marginal dalam arti mempunyai tingkat kesuburan yang rendah.
Di seluruh wilayah tropika, luas Ultisol (disebut juga tanah podzolik merah kuning) menempati urutan kedua setelah Oxisol, sedangkan di Asia, Ultisol menempati urutan pertama dan Oxisol urutan kedua (Giller 2001). Di Indonesia, luas Ultisol yaitu 48.3 juta ha, yang tersebar di Sumatera 20.6 juta ha, di Kalimantan 16.1 juta ha, di Sulawesi 2.0 juta ha dan di Irian Jaya 9.6 juta ha (LPT 1969). Ultisol bersama Oxisol termasuk ke dalam kelompok tanah tropika yang telah mengalami pelapukan dan pencucian berat. Pencucian menghilangkan banyak unsur hara dari tanah. Tanah yang sangat melapuk di daerah tropika banyak mengandung oksida dan hidroksida besi dan aluminium serta mempunyai mineral liat yang dominan berupa kaolinit (tipe 1 : 1) yaitu mempunyai permukaan bermuatan negatif lebih sedikit dibandingkan mineral liat illit atau vermikulit (tipe 2 : 1). Akibatnya kapasitas tukar kation (KTK) tanah menjadi rendah dan sebagian besar permukaan jerapan didominasi oleh ion aluminium (persentase kejenuhan aluminium tinggi). Tanah demikian mempunyai tingkat kesuburan rendah yang ditandai dengan rendahnya ketersed iaan unsur hara seperti N, P, Ca, Mg, S, Zn, B, dan Cu. Selain itu sifat tanah bereaksi masam (Giller 2001). Menurut Beinroth (2001) Ultisol mempunyai sifat bereaksi masam (pH rendah), kejenuhan basa rendah, kejenuhan Al tinggi, dan kandungan unsur hara Ca, Mg, K, dan P rendah.
Pada umumnya Ultisol mempunyai pH sangat masam, pada lapisan atas mempunyai pH 3.7 sampai 5.8, tekstur sangat beragam dengan kadar liat 10-80 %, KTK berkisar 7-20 me/100 g . Tanah Ultisol di Lampung Utara mempunyai kadar Al dd 0.01-2.66 me/100 g, kejenuhan Al sangat beragam 0-90 %, kadar Fe2O3
dengan kadar rendah dan kurang tersedia bagi tanaman. Pada tanah masam di daerah tropika sebagian besar P tanah terdapat dalam bentuk occluded -P yang terbentuk oleh adanya selaput besi oksida dan aluminium oksida karena tingginya kadar besi dan aluminium dalam tanah (Tisdale et al. 1990).
Untuk kepentingan budidaya tanaman, upaya yang perlu dilakukan pada Ultisol adalah dengan cara pengapuran dan pemupukan serta pengelolaan yang baik (Hardjowigeno 1995). Biasanya untuk pemupukan tanaman pakan, dilakukan dengan menggunakan pupuk buatan seperti pupuk N, P dan K. Sebagai alternatif lain, dapat pula dilakukan dengan menggunakan pupuk hayati (mikroorganisme bermanfaat seperti Rhizobium dan CMA). Namun, penggunaannya di tingkat petani belum memasyarakat. Kelebihan pupuk hayati ini adalah bersifat ramah lingkungan, dan dapat diterapkan oleh petani dengan teknologi yang sederhana serta membutuhkan biaya relatif murah.
Deskripsi Tanaman Legum Pakan
Sentro (Centrosema pubescens Benth)
Nama lain dari Centrosema pubescens adalah
C
entro (Australia), Butterfly pea (Inggeris), Dilang-butiki, Lesu-kesu (Filipina), lai, Thua-sentro (Thailand), Day trung chao long (Vietnam). Sentro termasuk famili Leguminosae dan sub famili Papilionaceae, berasal dari Amerika Selatan dan Tengah, telah tersebar luas di daerah tropika dan subtropika, dan merupakan salah satu jenis legum yang paling banyak penyebarannya di daerah yang mempunyai tipe iklim tropika basah (Skerman 1977; ‘t Mannetje dan Jones 1992).linear dengan panjang 4–17 cm dan lebar 6–7 mm, pipih, pinggir menonjol, lurus atau agak membengkok, semakin meruncing ke ujung; kalau sudah masak berwarna coklat tua, mengandung biji 12–20 buah. Biji Sentro berbentuk oblong pendek berwarna coklat kemerahan dengan garis -garis hitam. Ukuran bijinya yaitu panjang 4–5 mm, lebar 3 mm, dan tebal 2 mm. Jumlah biji Sentro per kg yaitu 40,000–45,000 (Bogdan 1977; Whit eman et al. 1974; ‘t Mannetje dan Jones 1992).
Di daerah tropika basah, Sentro dan Stylo dapat tumbuh baik pada tanah yang subur maupun yang tidak subur. Pada tanah yang mengalami defisiensi mineral, apabila dilakukan pemupukan dan inokulasi biji dengan Bradyrhizobium efektif, Sentro lebih produktif dibandingkan dengan Stylo (‘t Mannetje dan Jones 1992).
Sentro sangat selektif dalam kebutuhan Rhizobium (Bogdan 1977). Menurut Giller (2001) berdasarkan laporan Chen et al. (1997) bahwa rhizobia yang diisolas i dari Sentro dimasukkan ke dalam suatu genus Rhizobium baru yaitu Rhizobium hainanense. Sentro menghendaki curah hujan tahunan 1500 mm atau lebih, meskipun juga bisa bertahan pada curah hujan lebih rendah yaitu pada kondisi pastura yang menerima curah hujan tahunan 800 mm. Sentro juga toleran terhadap kondisi tergenang jika dengan penggembalaan ringan, selain itu centro masih dapat bertahan pada kondisi musim kering 3-4 bulan, tetapi tidak tahan pada kondisi kekeringan yang lebih lama dari itu. Sentro dap at tumbuh baik dengan spesies lain pada pastura campuran rumput-legum. Sentro termasuk legum penambat nitrogen yang efisien dengan kemampuan menambat N berkisar 120 sampai 270 kg N per ha per tahun dan mempunyai kadar N berkisar 2.4 sampai 2.7 %, dan sangat disukai oleh ternak (‘t Mannetje dan Jones 1992).
Kalopo (Calopogonium mucunoidesDesv.)
kemudian, Kalopo dibawa ke Malaysia dan di sana dimanfaatkan sebagai tanaman penutup tanah pada perkebunan karet. Pada tahun 1930 Kalopo dibawa ke Australia, tetapi tidak digunakan secara meluas di sana (‘t Mannetje dan Jones 1992).
Kalopo adalah legum herba perenial yang tumbuh kuat, menjalar dan membelit dengan panjang mencapai beberapa meter, membentuk massa jalinan dedaunan setebal 30–50 cm. Batang Kalopo sukulen dan ditutupi bulu panjang berwarna coklat, serta terbentuk akar pada buku-buku yang kontak dengan tanah. Tipe daun Kalopo trifoliat dengan helai daun berbulu pada kedua sisi permukaannya, helai daun terminal berbentuk ovat melebar sampai ovat jajaran genjang, sedangkan helai daun lateral berbentuk ovat melebar miring. Panjang helai daunnya 4–10 cm dan lebar 2-5 cm. Kalopo mempunyai bunga kecil sebanyak 4–12 buah terdapat dalam tandan aksilar yang pendek, corolla berwarna biru atau ungu. Polongnya linear, lurus atau melengkung dengan panjang 2– 4 cm dan lebar 3.5–5.0 mm ditutupi oleh bulu -bulu kasar berwarna coklat kemerahan, terdapat 3 – 8 biji dalam polong. Biji Kalopo berbentuk agak persegi dengan sudut bundar berwarna coklat tua atau coklat kekuningan dengan panjang 2.5–4.0 mm dan lebar 2.5–3.0 mm. Jumlah biji Kalopo per kg yaitu 70,000–75,000 butir (Bogdan 1977; Whiteman et al. 1974).
Puero (Pueraria phaseoloides(Roxb.) Benth.)
Istilah lain dari Pueraria phaseoloides adalah Puero, Tropical kudzu (Australia dan Inggeris), Krandang (Indonesia), Kudzu tropika (Malaysia), Singkamasaso, Bahay, Vaay (Filipina), Pied, Suak pied (Laos), Thua sian paa (Thailand), Dau ma, Dau dai, Dau rung (Vietnam). Puero berasal dari daerah dataran rendah Asia Tenggara dan Timur. Sekarang telah tersebar luas ke daerah tropika lainnya dan menjadi tanaman alami pada berbagai negara termasuk di antaranya daerah tropika basah Australia dan Amerika (‘t Mannetje dan Jones 1992).
lembab, pada bukunya dapat terbentuk akar. Pada batang lateral Puero tumbuh cabang pada buku, pucuk muda ditutupi oleh bulu-bulu tebal berwarna coklat. Daun Puero lebar berbentuk trifoliat, terdapat pada petiol yang panjangnya 2–13 cm. Helai daunnya tipis triangular atau ovat dengan panjang 2–20 cm dan lebar 2–15 cm; bagian pangkal berbentuk triangular atau agak jajaran genjang dan bercuping sangat dangkal; helai daun lateral miring, biasanya panjang dan lebarnya hampir sama yaitu 6–7 cm, permukaan atas berbulu tipis dan permukaan bawah berbulu lebih padat serta berwarna hijau keabuan. Bunganya kecil berwarna biru kehijauan sampai ungu tua, terdapat berpasangan terpencar pada tandan aksilari yang panjangnya 15–30 cm. Korollanya panjang sampai 2 cm, berwarna biru kehijauan, ungu atau putih; bendera berwarna keputihan dengan bintik biru kehijauan. Polongnya linear, lurus atau agak melengkung, kompak, berbentuk silinder dengan panjang 4–11 cm dan lebar 3–5 mm, mengandung 10-20 biji, ditutupi oleh bulu-bulu yang jarang dan kaku. Bijinya berbentuk oblong sampai agak persegi dengan sudut bulat, berukuran 3 mm x 2 mm, berwarna coklat sampai hitam kecoklatan. Jumlah biji Puero per kg adalah 81,000–88,000 (Bogdan 1977; Whiteman et al. 1974; ‘t Mannetje dan Jones 1992).
Mikoriza Cendawan Mikoriza Arbuskula (CMA)
atau pergerakan karbon dari tanaman inang ke cendawan dan pergerakan unsur hara yang difasilitasi oleh cendawan ke tanaman inang (Sylvia 1998).
Berdasarkan struktur tumbuh dan cara infeksinya pada sistem perakaran tanaman inang (host), mikoriza dikelompokkan ke dalam dua golongan besar yaitu Ektomikoriza dan Endomikoriza. Di dalam kelompok endomikoriza terdapat enam subtipe yaitu mikoriza arbuskula, ectendo, arbutoid, monotropoid, ericoid, dan orchid. Akhir -akhir ini tipe arbuskula banyak menjadi perhatian para ahli biologi dan lingkungan. Cendawaan mikoriza arbuskula (CMA) lebih populer dipakai untuk tanaman pertanian, hortikultura, perkebunan, dan pastura. Sedangkan ektomikoriza untuk tanaman kehutanan (Setiadi 2000; Sylvia 1998). Berbeda dengan ektomikoriza, kolonisasi CMA tidak menyebabkan terjadinya perubahan morfologi akar sehingga kuantifikasi kolonis asi CMA ini harus melalui pengamatan dengan mikroskop. Dalam asosiasi ini, baik dinding sel cendawan maupun membran sel tanaman inang tidak diterobos. Ketika cendawan tumbuh, membran sel tanaman inang melipat ke dalam membentuk rongga dan membungkus cendawan dalam suatu kompartemen (ruang apoplastik) di mana bahan yang mengandung molekul kompleks ditimbun di sana. Ruang apoplastik ini mencegah kontak langsung antara sitoplasma cendawan dan tanaman dan menyebabkan transfer nutrien antara simbion menjadi efis ien (Sylvia 1998).
diferensiasi hifa vegetatif, berupa spora reproduktif yang dapat dibentuk baik di dalam akar maupun yang paling sering di dalam tanah (Brundrett et al. 1996; Sylvia 1998)
Cendawan mikoriza arbuskula (CMA) diklasifikasikan ke dalam ordo Glomales (Morton 1988 yang dikutip Sylv ia 1998). Taksonomi lengkapnya yaitu ordo Glomales, terdiri dari dua subordo yaitu subordo Gigasporinae dan subordo Glomineae. Gigasporinae terdiri atas famili Gigasporaceae, mencakup genus Gigaspora dan genus Scutellospora. Glomineae terdiri atas famili Glomaceae mencakup genus Glomus dan genus Sclerocystis, dan famili Acaulosporaceae mencakup genus Acaulospora dan genus Entrophospora (Brundrett et al. 1996; Sylvia 1998). Di INVAM (2002) telah ditemukan dua famili CMA tambahan, yaitu Paraglomaceae dengan genus Paraglomus terdiri atas dua spesies P. occultum Morton dan Redecker dan P. brasilianum serta famili Archaesporaceae dengan genus Archaespora terdiri atas tiga spesies A. trappel, A. leptoticha, dan A. gerdeman (http://invam.caf.wvu.edu/myc-info/taxonomy/authors/authors.htm).
Simbiosis CMA-Tanaman Inang
Terjadinya simbiosis antara cendawan dengan tanaman inang melalui suatu proses kolonisasi atau infeksi akar oleh cendawan. Proses kolonisasi cendawan mikoriza arbuskula terdiri dari beberapa tahapan yaitu : prakolonisasi, kolonisasi primer (penetrasi akar), pembentukan arbuskula dan vesikel, penyebaran cendawan di dalam akar, pertumbuhan cendawan di dalam rhizosfer, dan fase reproduktif cendawan (Sukarno 2003).
ekstraradikal terdiri dari miselia, spora, dan hifa pelengkap. Struktur ekstrardikal tumbuh menjauhi akar, mengeksplorasi rhizosfer, serta melakukan penyerapan air dan nutrisi (Sukarno 2003).
CMA mempunyai berbagai peran penting baik dalam peningkatan produktivitas tanaman melalui peningkatan penyerapan unsur hara, toleransi terhadap kondisi stres air dan toksisitas logam berat, serta resistensi terhadap patogen tanah (Killham 1999; Marschner 1995) maupun dalam kelestarian sumberdaya tanah melalui peningkatan siklus hara, konservasi, serta perbaikan sifat fisik dan mikrobiologi tanah (Brundrett et al. 1996; Mason et al. 1992). Setiadi (2000) merinci berbagai peran CMA sebagai berikut : 1) perbaikan nutrisi tanaman dan peningkatan pertumbuhan, 2) sebagai pelindung hayati ( bio-protection), 3) meningkatkan resistensi tanaman terhadap kekeringan, 4) terlibat dalam siklus bio -geo-kimia, 5) sinergis dengan mikroorganisme lain, dan 6) mempertahankan keanekaragaman tumbuhan. Menurut Marschner (1995) CMA dapat meningkatkan penyerapan unsur hara tanaman terutama P, tetapi termasuk juga unsur hara Zn, Cu, K, dan NH4+. Crowley dan Rengel (2000) menyatakan
bahwa dari hasil pengujian, dapat disimpulkan CMA dapat menyediakan sejumlah unsur hara untuk tanaman yaitu masing-masing 80 % P, 25 % N, 10 % K, 25 % Zn, dan 60 % Cu.
Berdasarkan kepada banyaknya peranan mikoriza dalam peningkatan produktivitas tanaman, maka CMA dapat merupakan salah satu alternatif teknologi berupa pupuk hayati (biofertilizer) untuk pengembangan tanaman pertanian pada umumnya. Pemanfaatan pupuk hayati tersebut adalah berupa inokulasi cendawan mikoriza arbuskula dari isolat yang telah terbukti efektif dalam meningkatkan produktivitas tanaman.
tanaman bergantung kepada depedensi (ketergantungan terhadap tanaman inang) mikoriza, sifat tanah terutama ketersediaan hara seperti fosfor, dan kemampuan cendawan dalam memberi keuntungan kepada tanaman inang. Penelitian pot di rumah kaca yang dilakukan oleh Karti et al. (2000) menyimpulkan bahwa inokulasi campuran tiga jenis cendawan mikoriza arbuskula (Gigaspora margarita, Glomus manihotis, G. etunicatum) mampu meningkatkan pertumbuhan, produksi, dan serapan fosfor enam jenis rumput pakan (Brachiaria humidicola, B. decumbens, Cynodon plectostachyus, Chloris gayana, Digitaria
decumbens, Setaria splendida) yang ditanam pada tanah masam yang rendah unsur hara. Lukiwati (1996) membuktikan bahwa inokulasi cendawan mikoriza arbuskula yang dikombinasikan dengan pemupukan batuan fosfat mampu meningkatkan produksi dan nilai nutrisi hijauan legum centro dan puero pada tanah steril (penelitian pot), tetapi tidak pada tanah yang tidak steril (penelitian lapang).
Menurut Fakuara (1988), bahwa infeksi CMA dipengaruhi oleh berbagai faktor meliputi : pemupukan, nutrisi tanaman, pestisida, intensitas cahaya, musim dan kelembaban tanah, pH, kepadatan inokulum, dan kerentanan tanaman. Efektifitas CMA sangat tergantung kepada kesesuaian antara faktor jenis CMA, tanaman dan tanah, serta interaksi ketiga faktor tersebut. Kodisi tanah sangat mempengaruhi perkembangan CMA, di mana perkembangan asosiasi antara CMA dengan tanaman inang sangat tertekan pada kondisi ketersediaan N dan P yang tinggi. Demikian juga dengan pH tanah, yang dapat mengubah ketersediaan baik hara maupun toksin (Killham 1999). Khusus mengenai kandungan air tanah, di mana pada kondisi cekaman kekurangan air tanah, CMA justru dapat meningkatkan resistensi tanaman terhadap kekeringan (Marschner 1995; Setiadi 2000), karena CMA membantu meningkatkan penyerapan air (de la Cruz 1986).
CMA tetapi tidak mempengaruhi keanekaragaman spesies. Hasil penelitian pada beberapa tipe pemanfaatan lahan di Provinsi Jambi, ditemukan jenis CMA yang paling beragam (11 spesies) yaitu pada padang alang -alang, dengan jumlah spora terbanyak kedua setelah lahan singkong yang didominasi oleh spesies Glomus sp.4.
Bakteri Bintil Akar
Biologi Rhizobium
Rhizobium adalah merupakan salah satu genus bakteri penambat nitrogen secara simbiotik dalam bintil akar tanaman legum (bakteri bintil akar). Terjadinya penambatan nitrogen dalam bintil akar tanaman legum untuk pertama kalinya dideskripsikan oleh Helriegel dan Wilfarth pada tahun 1886, dan kemudian pada tahun 1888 Beijerinck berhasil mengisolasi organisme yang bertanggung jawab dalam penambatan nitrogen pada tanaman legum yang waktu itu disebut sebagai Bacillus radicicola, selanjutnya diganti dengan nama generik Rhizobium (Paul dan Clark, 1989), yang secara resmi diadopsi pada tahun 1926; dan sekarang di dalam Manual of Bacteriology Bergey ditempatkan di bawah genus Rhizobium (Mishra et al. 1999).
Bradyrhizobium (Graham 1998) beserta tanaman inangnya (di dalam tanda kurung) yaitu terdiri dari R. leguminosarum bv. phaseoli (Phaseolus), bv. Viciae (Vicia, Pisum), bv. Trifoli (Trifolium), R. tropici (Phaseolus), R. etli (Phaseolus, Mimosa), R. gallicum (Phaseolus), R. giardinii (Phaseolus), R. galegae (Galega), R. huautlense (Sesbania), R. mongolense (Medicago, Leucaena), dan R. hainanense (Centrosema, Desmodium, Stylosanthes, Tephrosia). Untuk genus Bradyrhizobium mencakup beberapa spesies seperti B. japonicum (Glycine, Soybean), B. elkanii (Glycine), dan B. liaoningense (Glycine).
Rhizobium dan Bradyrhizobium merupakan bakteri pembentuk bintil akar, mempunyai ukuran sel sedang dengan lebar 0.5–0.9 µm dan panjang 1.2–3.0 µm,
berbentuk batang, tidak membentuk endospora, gram negatif, bersifat mobile (bergerak) dengan satu flagel polar tunggal atau 2–6 flagel peritrikus
(Somasegaran dan Hoben, 1994). Rhizobium dan Bradyrhizobium berbeda dalam beberapa hal, di antaranya : kecepatan pertumbuhan dalam media agar MEK (Rhizobium tumbuh cepat yaitu membentuk koloni dalam 2 hari, Bradyrhizobium tumbuh lambat yaitu membentuk kolo ni setelah 4–5 hari), legum yang diinfeksi (Rhizobium menginfeksi kisaran sempit legum umumnya legum temperate, Bradyrhizobium menginfeksi kisaran luas legum tropika), gen nodulasi (gen nodulasi Rhizobium terdapat pada plasmid, sedangkan untuk Bradyrhizobium terdapat pada kromosom), reaksi metabolis pada media agar MEK yang ditambahkan BTB (brontimol biru) 0.0025 % (Rhizobium mengubah warna medium menjadi kuning atau bereaksi masam, sedangkan Bradyrhizobium tetap berwarna biru atau bereaksi basa) (Mishra et al. 1999; Somasegaran dan Hoben 1994).
yang dapat menginduksi bintil pada legum dari kelompok inokulasi-silang yang berbeda (Fred et al. 1932) yang dikutip Giller (2001). Menurut Giller (2001), meskipun konsep kelompok inokulasi-silang ini secara praktis dapat digunakan dalam menentukan galur rhizobia yang perlu diinokulasikan pada tanaman legum tertentu, dan mempunyai nilai ilmiah dalam mengangkat isu bahwa terdapat kelompok tertentu tanaman legum yang cenderung memakai bersama simbion bakteri yang lazim, namun seperti telah lama diketahui (Wilson 1944) yang dikutip Giller (2001) bahwa konsep tersebut tidak sempurna karena terdapat banyak anomali seperti misalnya adanya kisaran legum yang membentuk bintil dengan galur rhizobia apasaja dan dapat menjadi tumpang tindih, meskipun berbeda jelas dengan galur rhizobia lainnya yang berasal dari tanaman inang yang sama. Selain itu pengetahuan tentang kisaran tanaman inang dapat menjadi bias yang disebabkan oleh pilihan terhadap tanaman inang yang diuji. Kelemahan lainnya dari konsep inokulasi silang ini yaitu bahwa konsep tersebut hanya mengacu kepada kemampuan membentuk bintil pada tanaman inang, tanpa menghubungkannya dengan kemampuan penambatan nitrogen (Giller 2001).
Dalam perkembangan sistem klasifikasi selanjutnya, telah dibuat berbagai metode pendekatan klasifikasi yang lebih baik karena menggunakan banyak informasi dan pertimbangan terbaru, sehingga saat ini terdapat metode taksonomi numerik, metode analisis sekuens 16S rRNA atau SSU rRNA, dan hibridisasi DNA-DNA (Mishra et al. 1999; Giller 2001). Berdasarkan rekomendasi dari subkomiti internasional mengenai taksonomi Rhizobium dan Agrobacterium bahwa untuk menggambarkan genus atau spesies baru suatu bakteri pembentuk bintil akar dan bintil batang dianjurkan menggunakan pendekatan terpadu yang disebut sebagai taksonomi polifasik yaitu merupakan integrasi data fenotipe, genotipe, dan pilogenetik (Giller 2001).
Simbiosis Rhizobium-Legum
mikrosimbion saling menerima keuntungan, di mana legum sebagai tanaman inang mendapatkan nutrisi berupa amonia yang dihasilkan oleh Rhizobium, sedangkan Rhizobium mendapatkan nutrisi beru pa senyawa kaya gula yang dihasilkan oleh legum (Mishra et al. 1999; Tjahjoleksono 2003).
Terwujudnya sistem penambatan nitrogen secara biologis ini sangat bergantung pada struktur kompleks yang merupakan asosiasi antara bagian tanaman dengan bakteri. Struktur kompleks yang dikenal dengan nama nodule atau bintil ini terbentuk melalui suatu seri atau tahapan morfogenetik yang akhirnya memerangkap bakteri di dalam kompartemen intraseluler pada sistem perakaran tanaman (dan pada batang dalam interaksi dengan Azorhizobium) yang berisi bakteri. Jadi, bintil akar adalah merupakan bagian dari tanaman yang berisi bakteri. Bakteri yang berada di dalam bintil akar ini dikenal dengan sebutan bakteroid. Di dalam bintil akar inilah berlangsungnya sistem penambatan nitrogen yang dibarengi dengan sistem pertukaran nutrisi antar kedua simbion yang berfungsi dan memberikan keuntungan mutualistik bagi tanaman dan bakteri yang berasosiasi (Tjahjoleksono 2003).
Schroder (2001) mengemukakan pendapat yang lebih baru, yaitu molekul penginduksi yang terdapat dalam eksudat akar diidentifikasi sebagai flavonoid, isoplavonoid, dan senyawa yang berkerabat, untuk selanjutnya bakteri memberi respons terhadap sinyal dari tanaman dengan cara mengeluarkan senyawa berupa suatu molekul lipo-oligosakarida yang bersifat spesifik untuk setiap galur rhizobia. Giller (2001) menambahkan bahwa terjadi suatu stimulasi aktivitas biokimia pada Rhizobium dengan adanya molekul flavonoid dan isoflavonoid yang terdapat dalam eksudat akar, menyebabkan aktivitas gen nod (“nodulation”) yaitu berupa gen yang produknya diperlukan untuk kemampuan membentuk bintil akar pada legum yang berkerabat sebagai tanaman inang. Lambers et al. (1998) dan Mishra et al. (1999) menyatakan bahwa terdapat tiga tipe gen nod yaitu gen nod pengatur (regulatory), gen nod umum (common), dan gen nod spesifik ( host-specific). Flavonoid yang dilepaskan oleh tanaman legum bergabung dengan gen nod pengatur, mengadakan transkripsi menghasilkan oligo-polisakarida, kemudian gen nod umum menyandi enzim lain yang terlibat dalam sintesis lipo-oligosakarida. Flavonoid juga mengaktifkan transkripsi gen nod spesifik menyandi enzim yang terlibat dalam dekorasi lipo-oligosakarida. Lipo-oligosakarida yang mengalami dekorasi ini disebut sebagai “nod factor” (faktor nod), yang merupakan suatu molekul sinyal yang disekresikan ke dalam rhizosfer tanaman. Menurut Giller (2001) struktur dasar faktor nod adalah suatu tulang punggung N-asetil-D-glukosamin yang berhubungan pada β-1,4 dengan 3 – 6 unit gula yang mengalami asilasi (mengandung gugus asil) pada residu gula terminal yang tidak mereduksi. Peranan faktor nod yaitu menggerakkan respon tanaman yang mengakibatkan infeksi Rhizobium dan pembentukan bintil akar (nodulasi). Faktor nod juga menggerakkan sejumlah respon di dalam epidermis tanaman : depolarisasi membran sel di daerah akar rambut yang sedang muncul, deformasi akar rambut dan menginduksi ekspresi gen -gen spesifik bintil akar.
deformasi, dan dinding sel bagian lainnya di sebelah dalam didegradasi sehingg a bakteri dapat masuk, sambil membentuk benang -benang infeksi. Benang-benang infeksi ini tumbuh di dalam akar rambut dan menyediakan suatu saluran yang memungkinkan bakteri dapat mencapai korteks akar. Kemudian ujung benang infeksi membuka, dan bakteri dilepaskan ke dalam korteks akar (Lambers et al. 1998). Mekanisme infeksi demikian disebut melalui penetrasi akar rambut disertai pembentukan benang-benang infeksi seperti yang dijumpai pada tanaman clover dan bean (Graham 1998). Tetapi ada juga mekanisme infeksi lain seperti yang ditemui pada tanaman Arachis dan Stylosanthes yaitu masuk melalui celah luka (crack entry) pada sisi akar lateral (Graham 1998; Schroder 2001).
Menurut Mengel dan Kirkby (2001), Lambers et al. (1998), dan Giller (2001), bersamaan dengan proses invasi bakteri, sel-sel tanaman yang terinfeksi mengadakan pembelahan. Selama proses pembelahan sel, organel-organel seperti mitokondria, ribosom, dan retikulum endoplasma melipatgandakan diri. Bakteri yang berada di dalam sel yang terinfeksi, selama waktu tertentu mengalami pembelahan dan berdiferensiasi membentuk bakteroid. Bakteroid umumnya dikurung dalam suatu membran yang disebut membran peribakteroid dan membentuk simbiosom. Hasil dari pembelahan sel-sel tanaman membentuk suatu meristem baru, yang selanjutnya tumbuh menjadi nodul atau bintil akar. Terdapat dua pola perkembangan bintil akar, yaitu beberapa tanaman tertentu mempunyai meristem yang bersifat tetap bertahan sampai dewasa, sehingga disebut sebagai bintil indeterminate seperti yang ditemui pada alfalfa dan leucaena, tetapi pada tanaman tertentu lainnya meristem ini tidak bertahan sampai dewasa sehingga tidak terlihat lagi adanya meristem di dalam bintil akarnya, disebut sebagai determinate seperti yang dijumpai pada tanaman Arachis dan Glycine.
Proses penambatan N2 terjadi di dalam bintil akar dengan adanya enzim
nitrogenase yang disentesis oleh bakteri. Simbiosis yang efektif, disertai dengan produksi leghaemoglobin yang berfungsi menyediakan O2 yang cukup untuk
atmosfer menjadi NH3 oleh enzim nitrogenase, serta lebih lanjut mengubahnya
menjadi glutamat.
Menurut Heldt (1999) substrat utama yang disediakan oleh sel tanaman inang untuk bakteroid adalah malat yang berasal dari sukrosa yang dikirim melalui tabung pembuluh. Sukrosa dimetabolisme oleh enzim sukrosa sintase, didegradasi melalui glikolisis menjadi fosfoenolpiruvat, kemudian dikarboksilasi menjadi oksaloasetat, dan direduksi menjadi malat. Sedangkan hasil penambatan N2 oleh bakteroid dalam bentuk NH3 (atau NH4+), dikirim ke sel tanaman inang
melalui suatu saluran khusus pada membran simbiosom, di dalam sel tanaman pada bintil akar tipe indeterminate (pengekspor amida), akan diubah menjadi asparagin melalui pembentukan glutamin, glutamat dan aspartat, untuk selanjutnya asparagin ini dikirim ke pucuk (bagian at as tanaman) (Taiz dan Zeiger 1991; Salisbury dan Ross 1992; Graham 1998; Heldt 1999). Tetapi di dalam sel tanaman pada bintil akar tipe determinate (pengekspor ureida), glutamat dan aspartat yang dihasilkan digunakan untuk mensintesis purin seperti xantin, kemudian dikirim ke sel-sel tetangga yang tidak terinfeksi untuk diubah menjadi ureida alantoin dan asam alantoat, untuk selanjutnya dikirim ke pucuk (Taiz dan Zeiger 1991; Graham 1998).
terdapat jenis legum yang sama atau legum yang bersimbiosis dengan Rhizobium pada masa yang baru lampau, 2) tidak terjadi nodulasi yang baik jika tanaman legum yang sama ditanam sebelumnya, 3) apabila legum ditanam sesudah tanaman non-legum dalam suatu rotasi, 4) pada reklamasi tanah, 5) apabila kondisi lingkungan tidak menunjang bagi kehidupan Rhizobium.
Date (1977) yang dikutip Schroder (2001) membuat postulasi pengelompokan legum pakan tropika berdasarkan kepada kebutuhan Rhizobium yaitu : 1) kelompok PE (Promiscuousand Effective), adalah kelompok tanaman legum yang membentuk bintil akar secara efektif dengan suatu kisaran luas galur Rhizobium dari genus dan spesies yang berbeda di dalam kelompok ini. Genus yang termasuk ke dalam kelompok ini adalah : Arachis, Calopogonium, Cajanus, Canavalia, Clitoria, Crotalaria, Cyamopsis, Desmanthus, Dolichos, Galactica,
Gliricidia, Glycine, Indigofera, Lablab, Macroptilium, Macrotyloma,
Psophocarpus, Rhynchosia, Stizolobium, Stylosanthes (beberapa subkelompok), Teramnus, Tephrosia, Vigna, dan Zornia, 2) kelompok PI (Promiscuous but Ineffective), yaitu kelompok tanaman legum yang sering membentuk bintil akar dengan suatu kisaran luas galur Rhizobium, yang sebagian besar tidak efektif dalam penambatan nitrogen. Genus dari kelompok ini adalah Adesmia, Aeschynomene, Centrosema (dua subkelompok), Desmodium (dua subkelompok), Psoralea, Sesbania (dua subkelompok), dan Stylosanthes, 3) kelompok S (Specific), kelompok genus dan spesies tanaman legum yang membentuk bintil akar secara efektif dengan suatu kisaran sempit atau terbatas dari galur Rhizobium yang umumnya hanya berasal dari bintil akar dari spesies yang homolog. Sebagian besar anggota kelompok ini tidak membentuk bintil akar dengan jenis dari genus atau spesies lain , dan banyak tanaman membentuk bintil akar secara tidak teratur apabila jenis Rhizobium yang sama digunakan pada kesempatan yang berbeda. Kelompok ini mencakup genus : Coronilla, Leucaena, Lotononis, Lotus (dua subkelompok), Lupinus (dua subkelompok), Medicago-Trigonella, Mimosa, Stylosanthes (dua subkelompok), dan Trifolium (Afrika, empat subkelompok).
kelompok PE (Schroder 2001; Giller 2001). Penelitian Nurhayati dan Gunawan (1989) membuktikan bahwa legum Sentro (Centrosema pubescens) dapat membentuk simbiosis yang efektif dengan rhizobium, baik galur lokal atau rekomendasi (BPT 115) maupun galur komersial (CB 1923), dan Sentro dikelompokkan menjadi legum yang lebih spesifik dalam kebutuhan rhizobium.
Secara teoritis untuk kelompok PE tidak perlu diberikan inokulasi. Namun menurut Date (1977) yang dikultip Giller (2001) bahwa meskipun suatu legum termasuk ke dalam kelompok PE, inokulasi dapat memberikan respon, jika galur yang kompatibel yang terdapat di dalam tanah kurang efektif dan atau kurang kompetitif dibandingkan dengan galur yang digunakan dalam inokulum.
Keberhasilan suatu inokulasi Rhizobium dipengaruhi oleh banyak faktor, selain faktor Rhizobium dan tanaman, faktor lingkungan dan pengelolaan juga akan berpengaruh. Salah satu faktor lingkungan penting adalah kandungan air tanah. Menurut Mishra et al. (1999) kondisi cekaman akibat kekurangan air akan menurunkan aktivitas nitrogenase yang berperan penting adalam proses penambatan N2. Ditambahkan oleh Mishra et al. (1999) bahwa kandungan air
tanah optimum untuk pembentukan bintil akar yaitu berkisar 65–70 % atau sekitar 80 % (Ciptadi 1992).
Menurut Schroder (2001) kapasitas penambatan nitrogen oleh tanaman legum pakan tropika berkisar antara 80 sampai 300 kg N per ha per tahun. Peoples dan Craswell (1992) melaporkan bahwa di Malaysia Centrosema pubescens menambat nitrogen sebanyak 150 kg N per ha per tahun, sedangkan Pueraria phaseoloides pada komposisi 0-40 % dalam campuran dengan tanaman lainnya selama tiga bulan mampu menambat nitrogen sebanyak 9 kg N per ha. Sedangkan perkiraan jumlah nitrogen yang dit ambat oleh padang rumput campuran legum dan rumput di dunia adalah berkisar antara 13 sampai 682 kg N per ha per tahun (Ledgard dan Steele, 1992).
Simbiosis Tripartit
Fitter dan Garbaye 1994), sehingga terjadi hubungan antara tiga jenis organisma yaitu tanaman legum, bakteri Rhizobium, dan cendawan mikoriza arbuskula. Hubungan demikian disebut sebagai simbiosis tripartit.
Aries et al. (1991) melaporkan bahwa simbiosis tripartit memberikan pengaruh positif terhadap pertumbuhan, pembentukan bintil akar, penambatan nitrogen, serta penyerapan fosfat. Menurut Linderman (1991) kebanyakan tanaman legum yang bersimbiosis dengan Rhizobium dan CMA (simbiosis tripartit) umumnya tumbuh lebih baik jika dibandingkan dengan yang hanya bersimbiosis dengan salah satunya saja. Sylvia (1998) menyatakan bahwa simbiosis tripartit merupakan suatu hubungan yang sinergis dan dapat meningk atkan kandungan N dan P tanaman. Dengan adanya CMA pada tanaman legum, dapat meningkatkan nutrisi tanaman terutama fosfor yang sangat dibutuhkan oleh tanaman legum yang melakukan penambatan N, dan berarti secara tidak langsung CMA dapat meningkatkan pemben tukan bintil akar dan penambatan N (Schroder 2001). Menurut Mansur (2000) terdapat pengaruh yang menguntungkan dari CMA pada simbiosis antara tanaman legum dengan Rhizobium dan CMA pada media tumbuh yang mempunyai tingkat kesuburan yang rendah yaitu berupa peningkatan nodulasi (pembentukan nodul) dan penambatan N.
kandungan hormon, kemungkinan besar melalui peningkatan status hara P, karena status hara P juga mempengaruhi kandungan hormon.
Peningkatan pertumbuhan tanaman, selain berkaitan dengan peran CMA dalam peningkatan hara tanaman, juga disebabkan oleh kemampuan CMA dalam meningkatkan toleransi tanaman terhadap kondisi kekurangan air (kekeringan) (Marschner 1995; Setiadi 2000). Hal ini berkaitan dengan adanya perkembangan hifa ekstraradikal CMA pada akar tanaman sehingga mampu menjelajahi lebih besar volume tanah (Davies et al. 1992) dan peningkatan penyerapan air tanah (Faber et al. ; Ruiz-Lozano dan Azcon 1995).
Respons Tanaman terhadap Cekaman Kekeringan
Tanaman selama hidupnya kadang-kadang atau bahkan selalu menghadapi suatu kondisi lingkungan yang menyimpang dari kondisi optimal atau disebut juga dengan kondisi suboptimal. Dalam biologi, menurut Levit (1980) kondisi suboptimal berupa perubahan kondisi lingkungan yang mungkin akan menurunkan atau merugikan pertumbuhan atau perkembangan tanaman disebut sebagai cekaman. Salah satu cekaman yang umum dijumpai adalah cekaman air (water stress). Cekaman air ini dapat dibedakan menjadi dua yaitu cekaman kekurangan air (cekaman kekeringan) dan cekaman kelebihan air. Cekaman kekeringan (drought stress) dapat disebabkan oleh kekurangan suplai air di daerah perakaran atau laju kehilangan air (transpirasi) lebih besar dari absorbsi air oleh akar meskipun kadar air tanahnya cukup.
(tanaman tahunan) yaitu sebagai penghindar kekeringan (drought avoider) dan sebagai penahan kekeringan (drought tolerator). Tanaman penghindar kekeringan mempunyai jaringan yang sangat peka terhadap dehidrasi dan cend erung menghindari terjadinya defisit air pada jaringannya jika terjadi kondisi air tanah yang terbatas, yaitu melalui perakaran yang dalam untuk memaksimumkan penyerapan air serta pergerakan daun atau kepekaan stomata untuk meminimumkan kehilangan air melalui daun. Tanaman penahan kekeringan mempunyai daun yang tahan terhadap dehidrasi dan cenderung memiliki respon yang kurang baik untuk menghindari dehidrasi. Tanaman penahan ini bergantung kepada pengaturan osmotik untuk dapat bertahan terhadap kekeringan.
Taiz dan Zeiger (1991) menjelaskan, pengaturan osmotik atau akumulasi solut oleh sel adalah suatu proses di mana potensial air dapat diturunkan tanpa diikuti oleh penurunan turgor. Perubahan potensial air dalam jaringan dihasilkan oleh perubahan dalam tekanan osmotik, biasanya dilakukan melalui peningkatan konsentrasi berbagai solut umum seperti gula, asam organik, dan ion (terutama K+). Akumulasi ion selama pengaturan osmotik terjadi terutama di dalam vakuola. Karena kompartementasi ion ini, solut lain harus diakumulasikan ke dalam sitoplasma untuk mempertahankan keseimbangan potensial air di dalam sel. Solut lain tersebut adalah berupa senyawa organik yang tidak mengganggu fungsi enzim yang disebut sebagai solut kompatibel atau osmolit kompatibel yang paling umum yaitu prolina, gula alkohol, sorbitol, amin kuarterner, dan glisin betain.
lebih toleran terhadap kekeringan dan kadar garam tinggi dibandingkan dengan legum yang menghasilkan bintil akar determinate.
Adapun respons tanaman yang bersimbiosis dengan cendawan mikoriza arbuskula menunjukkan gejala terjadinya peningkatan resistensi tanaman terhadap cekaman kekeringan. Ini terbukti dari banyak penelitian seperti yang dilaporkan oleh Auge (2001) bahwa keberadaan cendawan mikoriza arbuskula pada simbiosis CMA-tanaman meningkatkan resistensi tanaman terhadap cekaman kekeringan yaitu dengan cara mengembangkan strategi penghindaran kekeringan (mempertahankan potensial air internal tetap tinggi) dan peningkatan toleransi terhadap kekeringan (bertahan pada potensial air internal yang rendah).
PENGUJIAN KEEFEKTIFAN CMA DAN RHIZOBIUM
TERHADAP PERTUMBUHAN TANAMAN LEGUM
PAKAN PADA KADAR AIR TANAH
YANG BERBEDA
Evaluation on the Effectiveness of Arbuscular Mycorrhizal Fungi
and Rhizobium on Growth of Forage Legumes
under Different Soil Water Regimes
Abstrak
Tanaman legum pakan yang tumbuh pada tanah yang tidak subur seperti Ultisol mempunyai pertumbuhan yang tidak optimal.