KEANEKARAGAMAN
CENDAWAN MIKORIZA ARBUSKULA (CMA)
DI HUTAN PANTAI UJUNG GENTENG,
SUKABUMI-JAWA BARAT
RITA TRI PUSPITASARI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRACT
RITA TRI PUSPITASARI. Diversity of Arbuscular Mycorrhizal Fungi (AMF) at Coastal Forest Ujung Genteng, Sukabumi-West Java. Supervised by Nampiah Sukarno, Kartini Kramadibrata, dan Dede Setiadi.
Indonesia is one of the countries that has a megadiversity of microorganisms and has a large area of coastal forests. However, research on microbial diversity particularly AMF as one of the dominant microbes in a coastal forest ecosystems has not been done extensively. The main objective of this research was to study the diversity and distribution of AMF in coastal forest and to analyse effects of salinity on the fungal and plant community at Coastal Forest Ujung Genteng Cape, Sukabumi - West Java.
This research was carried out in two parts: field, green house and laboratorium experimental activities. Field experimental activities were conducted for vegetation analysis, collecting of plant root system, rhizosphere soils from tree and pole phase,. seedling of understorey plants for fungal colonization analysis, isolation and identification of AMF and soil characteristics, include salinity. Green house and laboratorium activities were carried out for isolation, identification and colonization analysis of AMF. Field research location was divided into three zones, first zone was 0 – 70 m distance from coastal area (P), second zone was 71 – 140 m distance from coastal area (T), and third zone was 141 m distance from coastal until forest up to area in the center (H).
KEANEKARAGAMAN
CENDAWAN MIKORIZA ARBUSKULA (CMA)
DI HUTAN PANTAI UJUNG GENTENG,
SUKABUMI-JAWA BARAT
RITA TRI PUSPITASARI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Depertemen Biologi Fakultas Matematika dan dan Ilmu Pengetahuan Alam
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Keanekaragaman Cendawan Mikoriza Arbuskula (CMA) di Hutan
Pantai Ujung Genteng, Sukabumi-Jawa Barat
Nama
: Rita Tri Puspitasari
NIM
: G425010141
Disetujui
Komisi Pembimbing
Dr. Ir. Nampiah Sukarno
Ketua
Dr. Kartini Kramadibrata
Dr. Ir. H. Dede Setiadi, MS.
Anggota
Anggota
Diketahui
Ketua Departemen Biologi
Dekan Sekolah Pascasarjana
Dr. Ir. Dedi Duryadi S., DEA
Prof. Dr. Ir. Syafrida Manuwoto, M.Sc.
SURAT PERNYATAAN
Saya yang bertanda tangan di bawah ini :
Nama
: Rita Tri Puspitasari
NIM
: G425010141
Program Studi : Biologi (BIO)
dengan ini menyatakan bahwa tesis dengan judul ”
Keanekaragaman Cendawan
Mikoriza Arbuskula (CMA) di Hutan Pantai Ujung Genteng, Sukabumi-Jawa
Barat”
adalah benar-benar merupakan hasil penelitian yang saya lakukan di bawah
bimbingan Komisi Pembimbing saya, dan belum pernah diteliti oleh peneliti lain.
Penelitian tersebut dilakukan di Semenanjung Ujung Genteng, Sukabumi-Jawa Barat,
dan di Laboratorium Mikologi IPB serta di Laboratorium Mikoriza Herbarium
Bogoriense sejak Februari 2003 sampai dengan Juli 2005.
Bogor,
Desember
2005
Rita
Tri
Puspitasari
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya
sehingga penulisan Tesis yang berjudul Keanekaragaman Cendawan Mikoriza
Arbuskula (CMA) di Hutan Pantai Ujung Genteng, Sukabumi-Jawa Barat berhasil
diselesaikan.
Terima kasih dan penghargaan yang setinggi-tingginya penulis ucapkan kepada:
•
Ibu Dr. Ir. Nampiah Sukarno, Ibu Dr. Kartini Kramadibrata dan Bapak Dr. Ir. H.
Dede Setiadi, MS. selaku pembimbing, yang telah banyak memberi arahan sejak
penulisan proposal, penelitian hingga penulisan tesis ini selesai. Serta Ibu Dr. Ir.
Panca Dewi Manu Hara Karti, M.Si. sebagai penguji yang telah memberi koreksi dan
masukan.
•
Rektor Universitas Muhammadiyah Jakarta dan Dekan Fakultas Pertanian
Universitas Muhammadiyah Jakarta yang telah memberi izin untuk studi lanjut.
•
Pimpinan Institut Pertanian Bogor, Dekan Sekolah Pascasarjana dan Ketua Program
studi Biologi, atas kesempatan yang telah diberikan untuk mengikuti program S2 ini.
•
Kepala Botani Pusat Penelitian Biologi-LIPI, dan Kepala Laboratorium Mikologi
IPB yang telah memberi fasilitas penelitian.
•
Staf pengajar dan pegawai di lingkungan Program Studi Biologi, FMIPA IPB.
•
Teman-temanku: Dr. Ir. Happy Widiastuti, M.Si. yang telah memberi masukan dan
dukungan, Rida Oktorida, S.Si, Ijoh, Hadi Prastyo, S.Si, Yenni Lucia, SP, M.Si.,
Bapak Ujang Hapid, Nury Nuryada Aradea, S.Si. yang telah membantu selama di
laboratorium,Vivi Sumarna, S.Si pemandu saat survey, dan Nurhasanah, S.Si, M.Si.
serta Ir. Elfarisna, M.Si. yang telah memberi semangat dan bantuan selama penulisan.
•
Ayahanda Prof. Dr. H. Noeng Muhadjir, Ibunda Hj. Hafni Zahara yang telah
memberi nasehat, semangat dan doa. Khususnya pada suamiku Ir. Tejo Padmono dan
anak-anakku tercinta, Poppy, Affan dan Sekar atas segala dukungan semangat,
kesabaran, pengertian dan doa sehingga tesis ini dapat diselesaikan.
Terima kasih, semoga tesis ini bermanfaat.
RIWAYAT HIDUP
Penulis dilahirkan di Jogjakarta pada tanggal 31 Mei 1966 dari Ayah Prof. Dr.
H. Noeng Muhadjir dan Ibu Hj. Hafni Zahara. Penulis merupakan anak ke tiga dari
empat bersaudara. Pada tanggal 12 Januari 1991 penulis menikah dengan Ir.Tejo
Padmono dan dikaruniai tiga orang anak yaitu Aphrodita Puspateja (Poppy) (14
tahun), Affan Ibrahim Puspateja (11½tahun), dan Aulia Sekar Puspateja (9 tahun).
Tahun 1979 penulis lulus SD Negeri II Laboratorium IKIP Jogjakarta, Sekolah
Menengah Pertama di SMP Negeri 8 Jogjakarta lulus tahun 1982, dan Sekolah
Menengah Atas di SMA Negeri 6 Jogjakarta lulus tahun 1985. Penulis masuk
Universitas Kristen Satya Wacana, Salatiga tahun 1985. Penulis memilih sub
Program Mikrobiologi Industri, Program Studi Biologi Lingkungan, Fakultas Biologi
dan lulus Sarjana Biologi tahun 1990. Pada tahun 2001 penulis melanjutkan studi di
Program Pascasarjana IPB, FMIPA Program Studi Biologi, sub Program
Mikrobiologi dengan beasiswa dari Universitas Muhammadiyah Jakarta selama 1
tahun, selanjutnya BPPS sampai selesai.
DAFTAR ISI
Halaman
DAFTAR TABEL ………...……….
ix
DAFTAR GAMBAR ………..….………... x
DAFTAR LAMPIRAN………..…….………... xii
PENDAHULUAN ………... 1
Latar Belakang ………... 1
Tujuan Penelitian ………...… 3
Pendekatan ………... 3
Hipotesis ………... 4
KEADAAN UMUM LOKASI PENELITIAN ………... 5
TINJAUAN PUSTAKA ... 6
Salinitas Tanah dan Beberapa Sifat Tanah lain ………... 6
Simbiosis Mikoriza Arbuskula (MA) …...………... 8
Kolonisasi
………..
10
Proses Infeksi CMA ..……….... 11
Pertumbuhan Hifa Eksternal dan Produksi Spora ………. 15
Cendawan Mikoriza Arbuskula ... 17
Ekologi dan Fisiologi CMA pada Kondisi Salin ……...………… 18
Peranan CMA dalam Penyerapan Unsur Hara ……….. 20
Keanekaragaman CMA dan Keanekaragaman Tumbuhan pada
berbagai Ekosistem ... 22
Peranan CMA dalam Komunitas Tumbuhan ... 24
BAHAN DAN METODE ………... 26
Waktu dan Tempat ………...
26
Bahan dan Alat ………...
26
Metode Penelitian ………... 26
Analisa Data ………...
32
Analisa Pendukung ………..
33
Penyimpanan Spesimen ………...
33
HASIL DAN PEMBAHASAN ………...
34
Analisa Vegetasi ………..
34
Keanekaragaman CMA ………...
41
Hubungan antara CMA dan Vegetasi ………..
75
PEMBAHASAN UMUM ... 95
SIMPULAN DAN SARAN……….
98
DAFTAR TABEL
Halaman
1 Nilai penting spesies tumbuhan fase pohon dan tiang pada zona 0-70 m
dari pantai (P) daerah Ujung Genteng ..……….. 36
2 Nilai penting spesies tumbuhan fase pohon dan tiang zona di 71-140 m
dari pantai (T) daerah Ujung Genteng …….………... 37
3 Nilai penting spesies tumbuhan fase pohon dan tiang zona di 141 m
dari pantai sampai tengah hutan (H) daerah Ujung Genteng ……….. 38
4 Identitas spora CMA yang berhasil diisolasi dari vegetasi di Ujung Genteng ... 41
5 Seri pengamatan dan jumlah spesies CMA hasil identifikasi ……… 76
6 Frekuensi mutlak dan frekuensi relatif keterdapatan setiap spesies CMA …… 77
7 Nilai penting tumbuhan, dan kerapatan, kekayaan serta keragaman CMA
di lapang pada rizosfer tumbuhan ……… 80
8 Spesies tumbuhan, kekayaan, kerapatan dan keragaman CMA pada biakan pot
dengan inang tumbuhan anakan ……… 81
9 Sifat tanah Semenanjung Ujung Genteng ………... 85
DAFTAR GAMBAR
Halaman
1 Struktur taksonomi secara umum pada CMA dan kaitannya dengan cendawan
lain berdasarkan urutan gen SSU rRNA (Walker dan Schü
β
ler 2004) ……... 17
2 Bagan alir penelitian ………. 28
3 Peta lokasi penelitian dan letak jalur pengamatan di hutan pantai Ujung
Genteng, Desa Batu, Kabupaten Sukabumi ………. 30
4 Spora
Acaulospora foveata
……….. 43
5 Spora
Acaulospora longula
………..……….... 44
6 Spora
Acaulospora scrobiculata
……….………... 45
7
Acaulospora tuberculata
……….………... 46
8 Spora
Acaulospora cf. undulata
………... 47
9 Spora
Gigaspora cf. gigantea
……….. 48
10 Spora
Gigaspora ramisporophora
……….. 48
11.
Glomus clavisporum
………..….. 49
12
Glomus cf. deserticola
………..……….. 50
13 Spora
Glomus etunicatum
……….. 52
14
Glomus geosporum
.………..…….. 53
15
Glomus cf. microaggregatum
….………..….. 54
16
Glomus rubiforme
……….. 56
17 Spora
Glomus tortuosum
….……….... 56
18
Glomus
sp1. ……….………..….... 57
KEANEKARAGAMAN
CENDAWAN MIKORIZA ARBUSKULA (CMA)
DI HUTAN PANTAI UJUNG GENTENG,
SUKABUMI-JAWA BARAT
RITA TRI PUSPITASARI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRACT
RITA TRI PUSPITASARI. Diversity of Arbuscular Mycorrhizal Fungi (AMF) at Coastal Forest Ujung Genteng, Sukabumi-West Java. Supervised by Nampiah Sukarno, Kartini Kramadibrata, dan Dede Setiadi.
Indonesia is one of the countries that has a megadiversity of microorganisms and has a large area of coastal forests. However, research on microbial diversity particularly AMF as one of the dominant microbes in a coastal forest ecosystems has not been done extensively. The main objective of this research was to study the diversity and distribution of AMF in coastal forest and to analyse effects of salinity on the fungal and plant community at Coastal Forest Ujung Genteng Cape, Sukabumi - West Java.
This research was carried out in two parts: field, green house and laboratorium experimental activities. Field experimental activities were conducted for vegetation analysis, collecting of plant root system, rhizosphere soils from tree and pole phase,. seedling of understorey plants for fungal colonization analysis, isolation and identification of AMF and soil characteristics, include salinity. Green house and laboratorium activities were carried out for isolation, identification and colonization analysis of AMF. Field research location was divided into three zones, first zone was 0 – 70 m distance from coastal area (P), second zone was 71 – 140 m distance from coastal area (T), and third zone was 141 m distance from coastal until forest up to area in the center (H).
KEANEKARAGAMAN
CENDAWAN MIKORIZA ARBUSKULA (CMA)
DI HUTAN PANTAI UJUNG GENTENG,
SUKABUMI-JAWA BARAT
RITA TRI PUSPITASARI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Depertemen Biologi Fakultas Matematika dan dan Ilmu Pengetahuan Alam
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Keanekaragaman Cendawan Mikoriza Arbuskula (CMA) di Hutan
Pantai Ujung Genteng, Sukabumi-Jawa Barat
Nama
: Rita Tri Puspitasari
NIM
: G425010141
Disetujui
Komisi Pembimbing
Dr. Ir. Nampiah Sukarno
Ketua
Dr. Kartini Kramadibrata
Dr. Ir. H. Dede Setiadi, MS.
Anggota
Anggota
Diketahui
Ketua Departemen Biologi
Dekan Sekolah Pascasarjana
Dr. Ir. Dedi Duryadi S., DEA
Prof. Dr. Ir. Syafrida Manuwoto, M.Sc.
SURAT PERNYATAAN
Saya yang bertanda tangan di bawah ini :
Nama
: Rita Tri Puspitasari
NIM
: G425010141
Program Studi : Biologi (BIO)
dengan ini menyatakan bahwa tesis dengan judul ”
Keanekaragaman Cendawan
Mikoriza Arbuskula (CMA) di Hutan Pantai Ujung Genteng, Sukabumi-Jawa
Barat”
adalah benar-benar merupakan hasil penelitian yang saya lakukan di bawah
bimbingan Komisi Pembimbing saya, dan belum pernah diteliti oleh peneliti lain.
Penelitian tersebut dilakukan di Semenanjung Ujung Genteng, Sukabumi-Jawa Barat,
dan di Laboratorium Mikologi IPB serta di Laboratorium Mikoriza Herbarium
Bogoriense sejak Februari 2003 sampai dengan Juli 2005.
Bogor,
Desember
2005
Rita
Tri
Puspitasari
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya
sehingga penulisan Tesis yang berjudul Keanekaragaman Cendawan Mikoriza
Arbuskula (CMA) di Hutan Pantai Ujung Genteng, Sukabumi-Jawa Barat berhasil
diselesaikan.
Terima kasih dan penghargaan yang setinggi-tingginya penulis ucapkan kepada:
•
Ibu Dr. Ir. Nampiah Sukarno, Ibu Dr. Kartini Kramadibrata dan Bapak Dr. Ir. H.
Dede Setiadi, MS. selaku pembimbing, yang telah banyak memberi arahan sejak
penulisan proposal, penelitian hingga penulisan tesis ini selesai. Serta Ibu Dr. Ir.
Panca Dewi Manu Hara Karti, M.Si. sebagai penguji yang telah memberi koreksi dan
masukan.
•
Rektor Universitas Muhammadiyah Jakarta dan Dekan Fakultas Pertanian
Universitas Muhammadiyah Jakarta yang telah memberi izin untuk studi lanjut.
•
Pimpinan Institut Pertanian Bogor, Dekan Sekolah Pascasarjana dan Ketua Program
studi Biologi, atas kesempatan yang telah diberikan untuk mengikuti program S2 ini.
•
Kepala Botani Pusat Penelitian Biologi-LIPI, dan Kepala Laboratorium Mikologi
IPB yang telah memberi fasilitas penelitian.
•
Staf pengajar dan pegawai di lingkungan Program Studi Biologi, FMIPA IPB.
•
Teman-temanku: Dr. Ir. Happy Widiastuti, M.Si. yang telah memberi masukan dan
dukungan, Rida Oktorida, S.Si, Ijoh, Hadi Prastyo, S.Si, Yenni Lucia, SP, M.Si.,
Bapak Ujang Hapid, Nury Nuryada Aradea, S.Si. yang telah membantu selama di
laboratorium,Vivi Sumarna, S.Si pemandu saat survey, dan Nurhasanah, S.Si, M.Si.
serta Ir. Elfarisna, M.Si. yang telah memberi semangat dan bantuan selama penulisan.
•
Ayahanda Prof. Dr. H. Noeng Muhadjir, Ibunda Hj. Hafni Zahara yang telah
memberi nasehat, semangat dan doa. Khususnya pada suamiku Ir. Tejo Padmono dan
anak-anakku tercinta, Poppy, Affan dan Sekar atas segala dukungan semangat,
kesabaran, pengertian dan doa sehingga tesis ini dapat diselesaikan.
Terima kasih, semoga tesis ini bermanfaat.
RIWAYAT HIDUP
Penulis dilahirkan di Jogjakarta pada tanggal 31 Mei 1966 dari Ayah Prof. Dr.
H. Noeng Muhadjir dan Ibu Hj. Hafni Zahara. Penulis merupakan anak ke tiga dari
empat bersaudara. Pada tanggal 12 Januari 1991 penulis menikah dengan Ir.Tejo
Padmono dan dikaruniai tiga orang anak yaitu Aphrodita Puspateja (Poppy) (14
tahun), Affan Ibrahim Puspateja (11½tahun), dan Aulia Sekar Puspateja (9 tahun).
Tahun 1979 penulis lulus SD Negeri II Laboratorium IKIP Jogjakarta, Sekolah
Menengah Pertama di SMP Negeri 8 Jogjakarta lulus tahun 1982, dan Sekolah
Menengah Atas di SMA Negeri 6 Jogjakarta lulus tahun 1985. Penulis masuk
Universitas Kristen Satya Wacana, Salatiga tahun 1985. Penulis memilih sub
Program Mikrobiologi Industri, Program Studi Biologi Lingkungan, Fakultas Biologi
dan lulus Sarjana Biologi tahun 1990. Pada tahun 2001 penulis melanjutkan studi di
Program Pascasarjana IPB, FMIPA Program Studi Biologi, sub Program
Mikrobiologi dengan beasiswa dari Universitas Muhammadiyah Jakarta selama 1
tahun, selanjutnya BPPS sampai selesai.
DAFTAR ISI
Halaman
DAFTAR TABEL ………...……….
ix
DAFTAR GAMBAR ………..….………... x
DAFTAR LAMPIRAN………..…….………... xii
PENDAHULUAN ………... 1
Latar Belakang ………... 1
Tujuan Penelitian ………...… 3
Pendekatan ………... 3
Hipotesis ………... 4
KEADAAN UMUM LOKASI PENELITIAN ………... 5
TINJAUAN PUSTAKA ... 6
Salinitas Tanah dan Beberapa Sifat Tanah lain ………... 6
Simbiosis Mikoriza Arbuskula (MA) …...………... 8
Kolonisasi
………..
10
Proses Infeksi CMA ..……….... 11
Pertumbuhan Hifa Eksternal dan Produksi Spora ………. 15
Cendawan Mikoriza Arbuskula ... 17
Ekologi dan Fisiologi CMA pada Kondisi Salin ……...………… 18
Peranan CMA dalam Penyerapan Unsur Hara ……….. 20
Keanekaragaman CMA dan Keanekaragaman Tumbuhan pada
berbagai Ekosistem ... 22
Peranan CMA dalam Komunitas Tumbuhan ... 24
BAHAN DAN METODE ………... 26
Waktu dan Tempat ………...
26
Bahan dan Alat ………...
26
Metode Penelitian ………... 26
Analisa Data ………...
32
Analisa Pendukung ………..
33
Penyimpanan Spesimen ………...
33
HASIL DAN PEMBAHASAN ………...
34
Analisa Vegetasi ………..
34
Keanekaragaman CMA ………...
41
Hubungan antara CMA dan Vegetasi ………..
75
PEMBAHASAN UMUM ... 95
SIMPULAN DAN SARAN……….
98
DAFTAR TABEL
Halaman
1 Nilai penting spesies tumbuhan fase pohon dan tiang pada zona 0-70 m
dari pantai (P) daerah Ujung Genteng ..……….. 36
2 Nilai penting spesies tumbuhan fase pohon dan tiang zona di 71-140 m
dari pantai (T) daerah Ujung Genteng …….………... 37
3 Nilai penting spesies tumbuhan fase pohon dan tiang zona di 141 m
dari pantai sampai tengah hutan (H) daerah Ujung Genteng ……….. 38
4 Identitas spora CMA yang berhasil diisolasi dari vegetasi di Ujung Genteng ... 41
5 Seri pengamatan dan jumlah spesies CMA hasil identifikasi ……… 76
6 Frekuensi mutlak dan frekuensi relatif keterdapatan setiap spesies CMA …… 77
7 Nilai penting tumbuhan, dan kerapatan, kekayaan serta keragaman CMA
di lapang pada rizosfer tumbuhan ……… 80
8 Spesies tumbuhan, kekayaan, kerapatan dan keragaman CMA pada biakan pot
dengan inang tumbuhan anakan ……… 81
9 Sifat tanah Semenanjung Ujung Genteng ………... 85
DAFTAR GAMBAR
Halaman
1 Struktur taksonomi secara umum pada CMA dan kaitannya dengan cendawan
lain berdasarkan urutan gen SSU rRNA (Walker dan Schü
β
ler 2004) ……... 17
2 Bagan alir penelitian ………. 28
3 Peta lokasi penelitian dan letak jalur pengamatan di hutan pantai Ujung
Genteng, Desa Batu, Kabupaten Sukabumi ………. 30
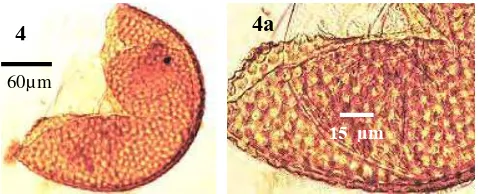
4 Spora
Acaulospora foveata
……….. 43
5 Spora
Acaulospora longula
………..……….... 44
6 Spora
Acaulospora scrobiculata
……….………... 45
7
Acaulospora tuberculata
……….………... 46
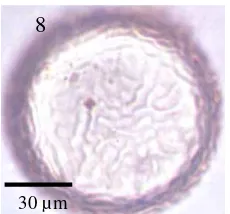
8 Spora
Acaulospora cf. undulata
………... 47
9 Spora
Gigaspora cf. gigantea
……….. 48
10 Spora
Gigaspora ramisporophora
……….. 48
11.
Glomus clavisporum
………..….. 49
12
Glomus cf. deserticola
………..……….. 50
13 Spora
Glomus etunicatum
……….. 52
14
Glomus geosporum
.………..…….. 53
15
Glomus cf. microaggregatum
….………..….. 54
16
Glomus rubiforme
……….. 56
17 Spora
Glomus tortuosum
….……….... 56
18
Glomus
sp1. ……….………..….... 57
20 Spora
Glomus
sp3 ………... 59
21
Glomus
sp4. ………..……….… 60
22 Kelompok
Glomus
sp5. ……….…… 61
23 Spora
Glomus
sp6. ……….…… 61
24 Spora
Paraglomus occultum
……… 62
25
Scutellospora cf. auriglobosa
………... 64
26 Spora
Scutellospora cf. heterogama
……….. 64
27 Frekuensi relatif keterdapatan spesies CMA di lapang, biakan pot dan anakan.. 78
28 Perbandingan kerapatan spora dan persentase kolonisasi akar dari lapang, dan
biakan pot ……… 83
29 Perbandingan kerapatan spora dan persentase kolonisasi akar dari lapang,
biak pot dan anakan dengan spesies tumbuhan yang sama ………....……….. 84
30 Sebaran CMA dan tumbuhan di Ujung Genteng ………... 86
DAFTAR LAMPIRAN
Halaman
1 Hasil Analisa Vegetasi Metoda Kuadran seluruh Semenanjung ... 107
2 Hasil Analisa Vegetasi Metoda Garis Menyinggung ……….... 108
3 Hasil Analisa Vegetasi Metoda Kuadran 0-70 m dari pantai ……… 110
4 Hasil Analisa Vegetasi Metoda Kuadran 71-140 m dari pantai ……… 111
5 Hasil Analisa Vegetasi Metoda Kuadran 141 m dari pantai sampai tengah
hutan ... 112
6 Persentase kolonisasi CMA di lapang, biakan pot dan anakan ………. 113
7 Sebaran CMA dan Tumbuhan di Semenanjung Ujung Genteng ……...……… 114
8 Gambar Biakan Pot ………... 115
9 Gambar Kolonisasi CMA ………... 116
10 Gambar Tumbuhan di Tepi Pantai ………. 117
11 Gambar Tumbuhan di Tengah Hutan ……… 118
12 Gambar Semenanjung Ujung Genteng, Sukabumi – Jawa Barat ……….. 119
13 Pembuatan Larutan Stock – Johnson Nutrient Solution ……… 120
PENDAHULUAN
Latar Belakang
Dalam ekosistem alami, cendawan mikoriza terutama cendawan mikoriza arbuskula (CMA) merupakan cendawan tanah yang paling dominan baik dalam jumlah maupun fungsinya. Hal ini disadari tidak saja oleh peneliti-peneliti dalam bidang mikrobiologi tetapi juga oleh peneliti-peneliti dalam bidang ekologi tumbuhan. Interaksi simbiosis mutualisme antara tumbuhan dan CMA ini merupakan bagian penting dalam ekosistem karena tumbuhan merupakan sumber utama penghasil karbon bagi mikroorganisme tanah termasuk CMA. Cendawan mikoriza arbuskula merupakan cendawan simbion obligat sehingga seluruh kebutuhan unsur karbonnya sangat bergantung pada tumbuhan. Sebaliknya, CMA mempunyai kemampuan untuk menyerap nutrisi dan air dari tanah dalam jumlah yang cukup besar dan mentransfernya kepada tumbuhan. Eratnya hubungan interaksi antara tumbuhan dengan CMA memungkinkan adanya peranan yang sangat besar dari tumbuhan dalam menentukan struktur keanekaragaman dan fungsi CMA dalam komunitas alami tersebut. Hal sebaliknya juga mungkin dapat terjadi yaitu keanekaragaman CMA merupakan faktor penentu dalam terpeliharanya keanekaragaman tumbuhan dalam komunitas alami (Johnson et al.
2005).
penyakit pada tumbuhan. Struktur keanekaragaman tumbuhan pada suatu ekosistem dapat terjaga dengan adanya simbiosis ini melalui mekanisme transfer nutrisi. Transfer nutrisi dilakukan oleh jaringan miselia atau hifa-hifa CMA dari satu akar tumbuhan dewasa ke akar tumbuhan anakannya. Jaringan miselia ini tidak saja berfungsi untuk menyerap air dan nutrisi dari dalam tanah, tetapi juga membantu mengalokasikan unsur karbon dari tumbuhan induk kepada anakannya. Mekanisme ini berperan sangat penting dalam membantu proses kelangsungan hidup suatu spesies tumbuhan terutama bagi anakan yang tumbuh dibawah kanopi yang cukup rapat. Sinar matahari yang sampai ke dasar hutan sangat rendah sehingga menghambat proses fotosintesa. Simbiosis mutualisme antara tumbuhan dan CMA tersebut selain dapat menjaga kesuburan tanah, dan nutrisi tumbuhan, juga dapat memelihara stabilitas dan biodiversitas komunitas tumbuhan (Smith & Read 1997).
Hutan pantai semenanjung Ujung Genteng merupakan salah satu hutan pantai yang masih tersisa di pulau Jawa. Ekosistem hutan pantai ini terletak di Desa Gunung Batu, Kecamatan Ciracap, Sukabumi, Jawa Barat. Posisi Semenanjung menghadap ke Samudra Hindia, dikelilingi batu karang yang cukup landai, dan jauh menjorok ke laut. Oleh karena areal hutan dikelilingi laut, maka ekosistem hutan tersebut dipengaruhi oleh salinitas.
terhadap produktivitas tumbuhan pada ekosistem tersebut (Al-Karaki 2000 dan Sumner 2000).
Tumbuhan bermikoriza umumnya memiliki toleransi lebih baik terhadap salinitas tanah sehingga pemanfaatan CMA dapat menjadi suatu pilihan dalam menanggulangi masalah rendahnya produktivitas tumbuhan di tanah salin. Namun penelitian terhadap peranan CMA pada tanah salin belum banyak dilakukan di Indonesia. Begitu pula penelitian tentang keanekaragaman CMA dengan faktor-faktor yang berkaitan di ekosistem hutan pantai, terutama keanekaragaman vegetasi tumbuhan yang tumbuh di atasnya dan sifat-sifat tanah khususnya salinitas belum dilakukan. Oleh karena itu penelitian mengenai keanekaragaman CMA di hutan pantai ini perlu dilakukan. Luaran dari penelitian ini diharapkan dapat memberikan data dasar mengenai keanekaragaman CMA di Indonesia, khususnya di Semenanjung Ujung Genteng yang dalam jangka panjang data ini dapat memberikan tambahan informasi untuk pemanfaatan CMA yang potensial sebagai sumber pupuk hayati yang dapat beradaptasi pada kondisi salin.
Tujuan Penelitian
Mempelajari keanekaragaman dan sebaran CMA pada tumbuhan hutan pantai dan hubungannya dengan sifat-sifat tanah, khususnya salinitas di Semenanjung Ujung Genteng, Sukabumi, Jawa Barat.
Pendekatan
2. Mengamati perbedaan populasi, keragaman dan kolonisasi CMA dari berbagai tumbuhan dari lapang setelah dibiakan dalam pot menggunakan inang Pueraria phaseoloides dan Shorghum sp.
3. Mengamati perbedaan populasi, keragaman dan kolonisasi CMA pada biakan pot menggunakan inang anakan / tumbuhan bawah dari lapang. 4. Hubungan keragaman tumbuhan dan keragaman CMA
Hipotesis
1. Terdapat hubungan antara keragaman tumbuhan dengan CMA.
KEADAAN UMUM LOKASI PENELITIAN
Letak Lokasi Penelitian
Secara geografis daerah penelitian terletak antara 07o15’ Lintang Selatan – 106o15’ Bujur Timur. Lokasi penelitian berada di Desa Gunung Batu, Kecamatan Ciracap, Sukabumi, Jawa Barat (Badan Meteorologi dan Geofisika Balai Wilayah II Stasiun Klimatologi Klas I Darmaga Bogor 2003). Semenanjung Ujung Genteng posisinya menghadap ke Samudra Hindia, dikelilingi batu karang yang cukup landai, dan jauh menjorok ke laut. Semenanjung ini merupakan tempat penunjang pangkalan Radar AURI Surade, dengan luas area hutan kurang lebih 34.7 Ha dari luas Desa Gunung Batu 85.82 Ha.
Kondisi Lahan
Pantai semenanjung berpasir putih dengan kandungan pasir berkisar dari 43.98% sampai 96.12 %, jenis tanah litosol, ditengah hutan tanah banyak bercampur dengan karang laut. Tanah mempunyai pH agak alkalis sampai alkalis. Salinitas berkisar antara 420 sampai 1 735 μs/cm (moderat sampai salin).
Iklim
TINJAUAN PUSTAKA
Salinitas Tanah dan Beberapa Sifat Tanah lain
Tanah yang disebut tanah salin ialah tanah yang mempunyai nilai salinitas lebih dari 4 mmho/cm. Secara alternatif, jika tanah dinyatakan dalam konteks konsentrasi garam, tanah bergaram/salin ialah tanah yang mengandung garam lebih dari 0.1% (1000 ppm) (Tan 1991). Sedangkan salinitas air untuk irigasi lahan pertanian dibagi menjadi 4 kelas, yaitu salinitas rendah (0 - 0.4 dS/m), salinitas moderat ( 0.4 - 1.2 dS/m), salinitas tinggi (1.2 - 2.25 dS/m) dan salinitas sangat tinggi ( 2.25 - 5 dS/m) (Sumner 2000).
Di daerah pantai, limpasan air laut ke daratan/tanah akan menyebabkan tanah tersebut mempunyai salinitas tinggi. Selain itu tanah salin dapat juga terdapat pada tanah yang secara alami memang mempunyai deposit garam. Air tanah salin biasanya mengandung 500 – 30 000 mg/l bahan terlarut (EC 0.7 - 42 dS/m), sementara air laut konsentrasi bahan terlarutnya ialah 33 000 mg/l dan Laut Mati di Israel mengandung kadar garam 270 000 mg/l (hipersalin) (Sumner 2000). Membandingkan hasil-hasil penelitian tentang salinitas tanah cukup rumit karena adanya perbedaan dalam metode pengukuran salinitas tersebut (Bernstein 1981).
Salinitas tanah akan menjadi masalah jika konsentrasi natrium klorida (NaCl), natrium karbonat (NaCO3), natrium sulfat (Na2SO4) atau garam-garam
Al-Karaki (2000) melaporkan bahwa apabila terjadi cekaman salinitas yang tinggi pada tumbuhan pertanian, Lycopersicum esculentum Mill cv. Pello, Arachis hypogaea cv. JL 24 (Gupta & Krisnamurthy 1996), dan Lactuca sativa (Ruiz-Lazano et al. 1996) akan menyebabkan terjadi penurunan kandungan P. Namun pemberian CMA pada lahan tersebut dapat meningkatkan pertumbuhan dan hasil panennya (Al-Karaki et al. 2001; Ruiz-Lazano et al. 1996).
Salinitas tanah tidak hanya bergantung pada konsentrasi garam dalam tanah, tetapi juga pada volume air dalam tanah. Salinitas pada tanah bertekstur kasar akan meningkat sampai 5 kali lipat dibandingkan dengan tanah yang bertekstur halus (Bernstein 1981, Hardjowigeno 1995).
Tumbuhan yang hidup di tanah salin dapat mengalami berbagai macam tekanan fisiologis karena pengaruh racun dari ion sodium dan klorida yang terdapat dalam tanah salin. Ion tersebut dalam jumlah tinggi pada tumbuhan akan menghancurkan struktur enzim dan makromolekul lainnya, merusak organel sel, mengganggu fotosintesis dan respirasi, dan menghambat sintesis protein. Selain itu, tingginya salinitas akan menurunkan permeabilitas akar terhadap air dan mengakibatkan penurunan laju masuknya air ke dalam tumbuhan (Marschner 1995), sehingga produktivitasnya menurun.
unsur hara mudah larut dalam air. Pada tanah alkalis, unsur P difiksasi oleh Ca sehingga tidak dapat diserap tumbuhan, sedangkan pada tanah masam unsur P diikat (difiksasi) oleh Al (Sumner 2000). Menurut Leiwakabessy et al. (2003), reaksi tanah (pH) mempunyai pengaruh yang sangat besar terhadap retensi P. Ketersediaan fosfat tanah tertinggi terjadi pada selang pH 6.0 - 6.6. Pada pH yang lebih rendah aktifitas fosfat berkurang karena mengalami retensi oleh R2O3
sedangkan pH di atasnya retensi terjadi oleh ion-ion Ca dan Mg. Namun apabila ion Na yang dominan terdapat pada tanah tersebut seperti pada tanah-tanah salin atau alkalin, maka retensi fosfat rendah.
Simbiosis Mikoriza Arbuskula (MA)
Mikoriza adalah suatu bentuk hubungan simbiosis mutualisma antara cendawan dengan perakaran tumbuhan. Istilah mikoriza berasal dari bahasa Yunani yaitu kata “Myces” yang berarti cendawan dan “Rhiza” yang berarti akar (Smith & Read 1997). Dalam simbiosis ini cendawan mendapatkan unsur karbon dari tumbuhan, sedangkan tumbuhan mendapatkan air dan nutrisi terutama P dari cendawan.
arbuskula dijumpai pada hampir semua jenis tumbuhan yang termasuk dalam
Angiospermae, Gymnospermae, Pteridophyta dan Bryophyta kecuali pada famili tumbuhan dari Cruciferae, Cyperaceae, Chenopodiaceae, Brasicaceae dan tumbuhan bermikoriza lain (Smith & Read 1997).
Mikoriza arbuskula mempunyai karakteristik yang khas yaitu: a). perakaran yang terkena infeksi tidak membesar, b). cendawan tidak membentuk struktur lapisan hifa pada permukaan akar, c). hifa menginfeksi sel korteks secara intra dan interseluler, d). membentuk struktur khusus berupa sistem percabangan hifa di dalam akar yang disebut arbuskula, e). beberapa membentuk struktur vesikula (Harley & Smith 1983).
Nama vesikula arbuskula merupakan derivat dari karakter stukturnya, arbuskula dan vesikula. Namun karena tidak semua CMA mempunyai struktur vesikula maka sebagian ahli mikoriza menyebutnya sebagai mikoriza arbuskula (MA). Mayoritas, sekitar 80% MA membentuk arbuskula dan vesikula, sedangkan lainnya hanya membentuk arbuskula tetapi tidak membentuk vesikula (Smith & Read 1997). Mikoriza arbuskula mempunyai tiga komponen penting, yaitu: akar, struktur cendawan dalam sel akar dan miselium ekstraradikal dalam tanah.
ialah organ istirahat karena jumlah selnya akan meningkat pada saat tumbuhan menua atau saat tumbuhan akan mati (Abbot & Robson 1982; Bonfante-Fosolo 1984).
Adanya simbiosis mutualisma antara CMA dengan perakaran tumbuhan dapat membantu pertumbuhan tumbuhan menjadi lebih baik, terutama pada tanah-tanah marjinal. Hal ini disebabkan MA efektif dalam meningkatkan penyerapan unsur hara makro dan mikro (Karagiannidis et al. 1995), kitinase berperan dalam melawan cendawan patogen yaitu dengan reaksi hidrolisis (Pozo et al. 2002). Sebagai contoh Phythoptora parasitica menurun pada akar yang diinokulasi cendawan mikoriza (Cordier 1998), MA juga dapat meningkatkan ketahanan tumbuhan inang dari kekeringan (Munyanziza et al. 1997; Kling & Jakobsen 1998) dan membantu pertumbuhan tumbuhan inang pada lahan yang tercemar logam berat (Munyanziza et al. 1997). Mikoriza arbuskula dapat pula menghasilkan zat pengatur tumbuh seperti auksin, sitokinin dan giberelin yang ditransfer kepada tumbuhan inangnya (Marschner 1995). Ludwig-Muller (2000) melaporkan bahwa peningkatan persentase kolonisasi akar jagung setelah diinokulasi oleh Glomus intraradices berkaitan dengan sintesis IBA.
Kolonisasi
miselium pada permukaan akar paling baik diantara 28 - 34 oC, suhu 20 - 30 oC merupakan kondisi yang terbaik bagi CMA untuk meningkatkan kolonisasi pada akar tumbuhan (Estaun, Camprubi & Calvet 1996).
Cahaya juga menunjukkan pengaruh yang nyata terhadap kolonisasi CMA (Hayman 1974). Kolonisasi cendawan pada akar anakan ramin sangat rendah jika anakan ramin disemaikan pada intensitas cahaya kurang dari 5 570 lux atau lebih dari 16 300 lux (Muin 2003).
Munculnya kolonisasi dapat berubah-ubah menurut musim, tipe tanah, kandungan air tanah, konsentrasi P, komposisi komunitas dan spesies tumbuhan (Allosopp 1998). Smith & Read (1997) menyatakan bahwa kolonisasi akar oleh CMA dapat berasal dari tiga sumber inokulum yaitu spora, potongan akar yang terinfeksi dan hifa secara keseluruhan yang disebut propagul.
Proses Infeksi CMA
Secara umum proses infeksi CMA pada akar tumbuhan melewati empat tahap, yaitu 1) induksi perkecambahan propagul dan pertumbuhan hifa, 2) kontak antara hifa dan permukaan akar serta pembentukan apresorium, 3) penetrasi hifa ke dalam akar, dan 4) perkembangan struktur arbuskula internal untuk selanjutnya terjadi simbiosis yang fungsional (Bonfante & Perotto 1995).
spesifik seperti epidermis dan korteks yang dapat dikolonisasi. Hal ini disebabkan CMA tidak mempunyai enzim yang dapat mendegradasi lignin dan suberin yang merupakan penyusun meristem. (Bonfante & Perotto 1995).
Senyawa seperti CO2, eksudat akar tumbuhan dan faktor lingkungan lainnya
dapat menstimulasi perkecambahan propagul CMA. Eksudat akar tumbuhan inang berupa flavonoid dapat menstimulir perkecambahan spora CMA dan pertumbuhan hifa (Giovanneti et al. 1993a; 1993b). Isoflavon dapat menginduksi pertumbuhan hifa, percabangan, dan diferensiasi serta penetrasi sel ke inang. Propagul cendawan akan berkecambah pada saat spora, molekul lipid, protein, glikogen yang terkandung di dalam spora terhidrolisis membentuk senyawa yang kaya akan energi, sehingga dapat digunakan untuk aktivitas metabolisme dan sintesis DNA (Becard et al. 1995).
Penetrasi CMA ke akar, tumbuhan melakukan serangkaian mekanisme pertahanan yang hampir sama dengan mekanisme apabila terjadi infeksi patogen pada tumbuhan. Mekanisme ini melewati beberapa tahapan yaitu pengenalan signal, transduksi signal dan aktivasi gen pertahanan (Garcia-Garrido & Ocampo 2002). Tumbuhan mempunyai pertahanan diri dengan mengkodekan gen yang dapat menghasilkan enzim pendegradasi dinding cendawan seperti kitinase dan β -1,3 glukanase, serta enzim yang terlibat dalam biosintesis fitoaleksin seperti fenilalanin ammonia liase. Namun pertahanan tumbuhan sebagai inang dapat ditekan dilawan dengan penyebaran hifa cendawan sehingga CMA tetap dapat berinteraksi dengan tumbuhan. Serangkaian mekanisme pertahanan tumbuhan yang lainnya yaitu mengakumulasi protein yang dapat menekan aktivitas patogen. Enzim yang berkaitan dengan pertahanan tumbuhan seperti kitinase dapat dilawan dengan penyebaran hifa cendawan tersebut. Kitinase berperan dalam melawan cendawan patogen yaitu dengan reaksi hidrolisis (Pozo et al. 2002)
Apresorium adakalanya bercabang sebelum penetrasi ke dalam epidermis. Setelah melewati epidermis umumnya hifa bercabang dalam berbagai arah, kadang-kadang tumbuh secara lateral (intraseluler) diantara sel-sel lapisan berikutnya sebelum masuk ke dalam sel korteks (Widden 1996). Penetrasi CMA melalui apresoria kemudian membentuk hifa koil di dalam sel-sel korteks dan berkembang menuju sel-sel korteks didekatnya menjadi arbuskula (Cooke, Widden & O’Halloran 1993).
berbutir-butir yang bercampur dengan protoplas sel inang. Analisa morfologi dan morfometrik menunjukkan bahwa arbuskula melengkapi perkembangannya dalam 2.5 sampai 4 hari (Alexander et al. 1989). Struktur hidup arbuskula relatif singkat berkisar antara 4 sampai 15 hari, bahkan dalam tumbuhan legum hanya 2-5 hari (Cooke, Widden & O’Hallora 1993). Di dalam arbuskula terjadi pertukaran metabolisme antara tumbuhan inang dan cendawan simbion.
Pembentukan arbuskula diawali dari suatu struktur batang, kemudian mengalami percabangan hifa dan tipe percabangannya dipengaruhi oleh tumbuhan inang (Cooke, Widden & O’Halloran 1993). Balestrini et al. (1992) melaporkan bahwa dengan terbentuknya arbuskula dalam sel korteks inang maka terjadi invaginasi plasmalema tumbuhan, fragmentasi vakuola, hilangnya amiloplas dan kenaikan jumlah organel seperti aparatus golgi. Adanya cendawan juga mempengaruhi posisi nukleus tumbuhan yang bergerak dari posisi tepi ke posisi sentral pada sel yang terinfeksi. Pada saat akar dikolonisasi CMA dan membentuk hifa koil, hifa interseluler atau arbuskula, plasmalema inang mengalami invaginasi dan proliferasi di sekitar perkembangan cendawan. Material apoplas yang terletak antara membran plasma tumbuhan yang mengalami invaginasi dan permukaan sel cendawan memberikan suatu ruang baru yang disebut ruang interfase.
H+/ATPase mencirikan simbiosis mutualisma sebagaimana juga dijumpai pada membran tumbuhan yang berbintil akar dan hal ini tidak dijumpai pada membran sekitar haustoria, tempat interaksi patogen tumbuhan (Bonfante & Perotto 1995).
Vesikula merupakan organ penyimpanan karbon untuk cendawan, dan vesikula ini juga menunjukkan awal infeksi baru. Vesikula merupakan struktur berbentuk bulat atau lonjong yang terbentuk dari hifa internal yang membengkak (Bonfante-Fosolo 1984).
Pertumbuhan Hifa Eksternal dan Produksi Spora
Adanya hifa eksternal memungkinkan tumbuhan mengeksploitasi volume tanah lebih besar. Friese & Allen (1991) melaporkan bahwa eksploitasi fisik pada absorbsi unsur hara difasilitasi oleh kecilnya diameter hifa yaitu berkisar 2-15 µm dengan rata-rata 3-4 µm. Tipisnya hifa dengan diameter kurang dari 10 µm lebih cocok untuk memanfaatkan P yang terdapat di ruang pori mikro tanah yang tidak dapat dicapai oleh akar dan rambut akar, sehingga dapat mengatasi keterbatasan difusi Pi yang lambat dalam tanah. Selain itu, hifa juga dapat menyerap air. Sehingga hifa eksternal dapat meningkatkan potensi sistem perakaran untuk mengabsorbsi unsur hara dan air. Beberapa studi menunjukkan bahwa daerah deplesi sekitar akar tumbuhan, lebih besar pada tumbuhan yang bermikoriza daripada tumbuhan yang tidak bermikoriza (Smith & Read 1997).
miselium eksternal ini dapat tumbuh dan menuju ke permukaan akar untuk membentuk unit kolonisasi.
Semua organisme merespon pengaruh temperatur. Hifa ekstraradikal miselia dari Glomus caledonium semuanya tidak dapat tumbuh pada suhu 10 oC namun dapat tumbuh baik pada suhu 25 oC (Gavito 2003). Pada suhu 0 oC G. intraradices
tidak mampu mengabsorbsi P namun dapat aktif kembali mengangkut P pada suhu 15 oC (Wang et al. 2002)
Banyaknya miselium eksternal ini sangat bervariasi, dapat sangat banyak pada beberapa contoh tanah atau bahkan menutupi akar namun tidak sampai membentuk selubung cendawan seperti pada ektomikoriza (Harley & Smith 1983). Selain miselium, CMA membentuk struktur lain yang dikenal sebagai spora, dapat dibentuk secara tunggal, berkelompok, atau di dalam suatu sporokarp tergantung jenis cendawannya. Miselium eksternal merupakan hal yang penting dalam produksi spora CMA, karbohidrat banyak di translokasikan ke tempat tersebut. Spora CMA dalam tanah merupakan bagian dari cendawan di luar akar penyusun biomassa (Smith & Read 1997).
Menurut Sieverding (1991) sporulasi CMA di dalam tanah terjadi pada pH sekitar 3.8 sampai 8.0. Toleransi dan kemampuan tumbuhan untuk tumbuh pada kondisi pH yang tidak menguntungkan ada kemungkinan karena asosiasi kolonisasi CMA dengan akar dan kemampuan CMA beradapatasi terhadap kondisi tersebut.
banyak, dapat menghasilkan cendawan dengan sporulasi yang lebih cepat, dan lebih banyak. (Brundrett 1996).
Cendawan Mikoriza Arbuskula
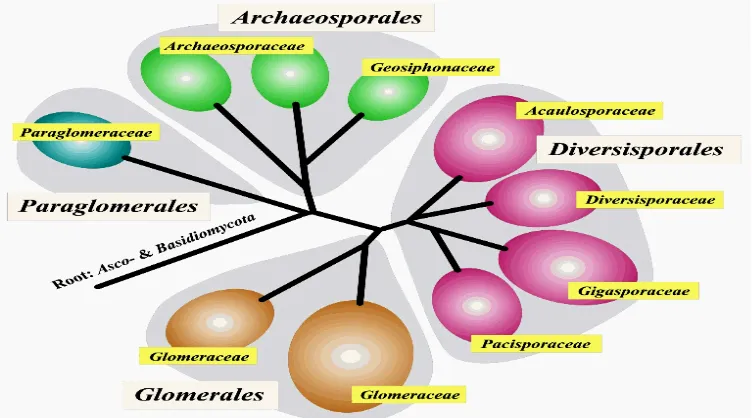
Menurut klasifikasi dari Morton & Benny (1990) cendawan mikoriza arbuskula (CMA) merupakan cendawan simbion obligat, mempunyai hifa aseptat, dan reproduksinya dilakukan secara aseksual. CMA digolongkan dalam filum
[image:39.612.134.511.458.667.2]Zygomycota kelas Zygomycetes dengan ordo Glomales. Dasar utama pembedaan adalah perkembangan, morfologi, dan struktur dinding pada globos zigospora, azigospora, klamidospora dan sporangianya. Cendawan-cendawan yang dimasukkan ke dalam ordo Glomales, ialah anggota genus Gigaspora, Scutellospora, Glomus, Sclerocystis, Acaulospora, dan Entrophospora. Menurut laporan Schüβler et al. (2001), cendawan mikoriza arbuskula (CMA) tidak digolongkan ke dalam Zygomycota lagi berdasarkan urutan gen SSU rRNA tapi digolongkan dalam filum baru, yaitu Glomeromycota (Gambar 1). Filum
ini mempunyai Kelas Glomeromycetes dengan 4 ordo, yaitu Glomerales, Diversisporales, Archaeosporales, dan Paraglomerales. Ordo Glomerales
dengan famili Glomeraceae. Ordo Diversisporales terdiri dari 4 famili, yaitu
Gigasporaceae, Diversisporaceae, Acaulosporaceae dan Pacisporaceae. Ordo
Archaeosporales terdiri dari 2 famili, yaitu Archaeosporaceae dan
Geosiphonaceae. Ordo Paraglomerales mempunyai famili Paraglomeraceae
(Walker & Schüßler 2004). Pacisporaceae pada Schüβler et al. (2001) belum tercantum.
Ekologi dan Fisiologi CMA pada Kondisi Salin
Hetrick (1984) mengatakan CMA dapat dijumpai di hampir semua jenis tanah, dari tanah masam sampai alkalin. Namun demikian komposisi kelimpahan spesies CMA dan derajat kolonisasinya berubah dengan adanya peningkatan salinitas tanah. Kim & Weber (1985) menyatakan bahwa CMA ditemukan pada vegetasi halofit pada tanah yang memiliki salinitas tinggi, namun kolonisasi akar oleh CMA menurun sejalan dengan kenaikan salinitas berdasarkan kandungan sodium tanah. Pendapat yang sama juga dilaporkan oleh Delvian (2003) bahwa keberadaan dan kelimpahan CMA di hutan pantai berhubungan negatif dengan tingkat salinitas tanah. Penurunan tingkat salinitas tanah akan meningkatkan kepadatan spora dan persentase kolonisasi CMA pada akar tumbuhan.
Pada percobaan yang dilakukan oleh Juniper & Abbot (2004) diperoleh bahwa, pengaruh NaCl pada pertumbuhan hifa G. decipiens dan S. calospora
bersifat dapat balik. Hifa yang berasal dari perkecambahan spora pada pasir dengan kandungan NaCl 300 mmol/l NaCl pertumbuhannya bertambah setelah dipindahkan ke lingkungan yang kadar salinnya lebih rendah. Tetapi hifa dari perkecambahan spora yang di tumbuhkan pada pasir nonsalin pertumbuhannya melambat setelah dipindahkan ke lingkungan salin. Morfologi hifa antara G. decipiens yang ditumbuhkan di substrat salin dan nonsalin berbeda. Panjang hifa yang dihasilkan oleh A. laevis, G. decipiens dan S. calospora berkurang dengan bertambahnya konsentrasi NaCl. Diameter hifa G. decipiens bertambah dengan semakin bertambahnya kadar NaCl. Hifa G. decipiens menebal dengan adanya NaCl, sitoplasmanya kosong dan septatnya banyak.
Coperman et al. (1996) menduga bahwa terdapat perbedaan sifat dan efisiensi pada setiap spesies CMA yang disebabkan oleh faktor genetik CMA itu sendiri. Menurut Ruiz-Lozano & Azcon (2000) bahwa perbedaan spesies CMA dan ekosistem asalnya akan menghasilkan respon pertumbuhan yang berbeda. Contohnya pada keadaan ekosistem yang salin dua jenis CMA dibandingkan, ternyata mempunyai respon yang berbeda. Glomus sp. melindungi tumbuhan dari cekaman salinitas didasarkan pada perkembangan akar, sedangkan Glomus deserticola berdasarkan perbaikan nutrisi tumbuhan.
Peranan CMA dalam Penyerapan Unsur Hara
Cendawan mikoriza arbuskula mampu memperbaiki nutrisi dan meningkatkan pertumbuhan tumbuhan inang. Seperti dijelaskan oleh Marschner (1995) simbiosis mikoriza arbuskula membentuk jalinan hifa secara intensif sehingga tumbuhan bermikoriza akan mampu meningkatkan kapasitasnya dalam menyerap unsur hara dan air. Fosfat adalah salah satu unsur hara utama yang dapat diserap oleh tumbuhan bermikoriza. Fosfat adalah salah satu unsur hara esensial yang diperlukan dalam jumlah relatif banyak oleh tumbuhan, tetapi ketersediaannya pada tanah-tanah tertentu terbatas, sehingga seringkali menjadi salah satu pembatas utama dalam peningkatan produktivitas tumbuhan. Cara yang umum untuk mengatasi hal ini salah satunya yaitu memberikan input energi yang tinggi berupa pemupukan fosfat. Untuk mengurangi input kimia tersebut yang relatif mahal, maka aplikasi inokulum CMA dapat dijadikan sebagai salah satu alternatif yang dapat digunakan dan dikembangkan. Unsur makro lainnya yang dapat diserap oleh MA ialah N dan K. Pada tumbuhan bermikoriza NH4+ dan mungkin NO3- meningkat. Unsur K konsentrasinya pada tumbuhan bermikoriza
Selain unsur makro, CMA juga dapat membantu penyerapan unsur hara mikro seperti Cu, Zn, S, Mo dan B. (Sieverding 1991; Read & Smith 1997). Mg konsentrasinya pada tumbuhan bermikoriza lebih tinggi dari pada tumbuhan tidak bermikoriza. Penyerapan Ca terlihat dipengaruhi oleh interaksi dengan unsur-unsur nutrisi yang lain.
Mikroelemen esensial bagi tumbuhan seperti Fe, Mn dan Cl secara umum juga ditemukan lebih tinggi konsentrasinya pada tumbuhan yang berasosiasi dengan cendawan tersebut. Muin (2003) melaporkan bahwa anakan ramin yang terkolonisasi CMA menyerap unsur hara mikro Mn, Fe dan Zn lebih banyak dibanding tumbuhan yang tidak terkolonisasi CMA. Serapan Fe meningkat sejalan dengan dosis fosfat alam yang diberikan. Na, Co dan Si tidak esensial untuk pertumbuhan tumbuhan, namun mereka cenderung kadang-kadang diserap tumbuhan yang bermikoriza. Beberapa logam berat dan beracun seperti Cd, Ni, Sr, Pb dan Cs serta beberapa non nutrisi seperti Br dan I juga mungkin dapat diserap (Read & Smith 1997; Sieverding 1991)
Secara umum unsur-unsur yang tidak mobil seperti P, Zn dan Cu meskipun berada di daerah rizosfer, sulit diserap oleh tumbuhan. Namun dengan adanya simbiosis CMA pada tumbuhan unsur-unsur tersebut dapat dengan baik diserap oleh tumbuhan (Read & Smith 1997).
ambang yang diperlukan P untuk berdifusi, dan merubah lingkungan organik rhizosfer secara kimia misalnya melalui pelepasan asam organik, dan peningkatan aktivitas fosfatase, serta meningkatkan produksi fitohormon yang dapat merubah fenotipe akar yaitu dengan pembentukan akar dengan nodus yang lebih tinggi juga dapat membuat umur akar menjadi lebih lama sehingga meningkatkan kapasitas penyerapan hara total (Orcutt & Nielsen 2000). Mikoriza arbuskula juga mengakibatkan terjadinya akumulasi asam-asam amino, protein, klorofil dan kandungan gula dibandingkan dengan tumbuhan nonmikoriza (Mathur & Vyas 2000).
Pertumbuhan dan fungsi mikoriza tergantung pada suplai karbon sebagai derivat fotosintesis dari tumbuhan inang. Cendawan mikoriza menerima karbohidrat dari tumbuhan inang sebanyak 4-14% dari total karbon hasil fotosintesis (Clark 1997). Allsopp (1998) menyatakan bahwa faktor-faktor yang menurunkan kapasitas fotosintesis tumbuhan akan berpengaruh terhadap fungsi MA, karena cendawan sebagai pasangan tumbuhan dalam bersimbiosis sangat tergantung pada karbon yang dihasilkan tumbuhan tersebut.
Keanekaragaman CMA dan Keanekaragaman Tumbuhan pada berbagai Ekosistem
Heijden (1998) hasilnya menunjukkan bahwa keragaman jenis CMA dapat mempengaruhi biomassa tumbuhan, biomassa cendawan, keanekaragaman jenis tumbuhan dan tumbuhan yang mengandung P.
Studi-studi tentang keanekaragaman CMA pada tanah salin menunjukkan bahwa Glomus merupakan genus dari CMA yang dominan (Delvian 2003, Landwehr et al. 2002, Purwanto 1999, Ragupathy dan Mahadevan 1991). Pada ekosistem gambut genus yang banyak ditemukan yaitu Glomus dan Acaulospora
(Muin 2003; Ekamawanti 1999).
Oehl et al. (2003) melaporkan bahwa intensifikasi pertanian akan mempengaruhi kelimpahan CMA dan struktur komunitasnya. Komunitas CMA berubah tidak hanya keanekaragamannya namun juga aspek fungsionalnya (kecepatan berkolonisasi dan kecepatan sporulasi pada biakan pot).
Ekosistem yang tergenang merupakan tempat dengan ketersediaan oksigen di daerah perakaran dengan jumlah sedikit. Cendawan mikoriza arbuskula merupakan organisme aerob, sehingga ketersediaan oksigen akan mempengaruhi formasi CMA di ekosistem tersebut. Contohnya pada ekosistem hutan rawa jumlah spora dan persentase kolonisasi lebih rendah dari pada ekosistem terestrial atau tanah kering (Khan 1993).
Disamping peranannya terhadap tumbuhan inang, CMA memegang peranan penting dalam ekosistem terrestrial. Tidak hanya keberadaanya, tetapi secara genetik dan diversitas fungsional sangat penting. Keragaman CMA menentukan struktur komunitas tumbuhan dan produktivitas ekosistem (Tilman et al. 2001).
tertentu mempunyai relung ekologis yang sangat khusus yang tidak umum, seperti halnya organisme lain, sehingga konsekuensinya, keanekaragaman komunitas CMA ini mungkin sangat peka terhadap perubahan lingkungan (Fitter et al. 2004).
Pada beberapa keadaan, kekayaan tumbuhan berhubungan dengan kekayaan spesies CMA sementara itu keragaman mikoriza sangat erat kaitannya dengan fungsi pada lingkungan setempat. Tumbuhan tidak banyak mempunyai pengaruh pada keanekaragaman CMA dan keanekaragam CMA mampu membantu kearah pemeliharaan sekumpulan tumbuhan yang berbeda (Johnson etal. 2005).
Peranan CMA dalam Komunitas Tumbuhan
Peranan yang paling menonjol dari CMA dibanding dengan tipe-tipe cendawan mikoriza lainnya dalam komunitas tumbuhan, yaitu kemampuannya untuk berasosiasi dengan sekitar 80% jenis tumbuhan. Sehingga CMA berperan penting dalam mempertahankan stabilitas keanekaragaman tumbuhan dengan cara transfer nutrisi dari satu akar tumbuhan ke akar tumbuhan yang lain yang berdekatan melalui struktur yang disebut dengan ‘ bridge hypha’ (Allen & Allen 1992).
menyebabkan perbedaan keanekaragaman spesies dan populasi CMA. Perubahan vegetasi dalam merespon perubahan lingkungannya mungkin mempunyai peranan yang besar dalam menentukan struktur komunitas CMA. Dan dari bukti-bukti menunjukkan bahwa CMA rentan terhadap perubahan lingkungan biotik maupun abiotik (Fitter et al. 2004). Sehingga peranan CMA dalam komunitas tumbuhan sangat erat kaitannya dengan perubahan lingkungannya.
BAHAN DAN METODE
A. Waktu dan Tempat
Penelitian di laksanakan di Ujung Genteng-Sukabumi, di rumah kaca Gunung Gede, di laboratorium Mikologi Gunung Gede dan di laboratorium Sitologi dan Mikroriza Herbarium Bogoriense-LIPI, Bogor. Penelitian dimulai dari bulan Februari 2003 sampai Juli 2005.
B. Bahan dan Alat
Bahan yang digunakan dalam penelitian ini adalah contoh tanah/pasir dan akar yang diambil dari hutan semenanjung Ujung Genteng Kabupaten Sukabumi, zeolit, PVLG, hara Johnson, sukrosa, aquades, alkohol 50 dan 70%, benih
Puerareaphaseoloides dan Sorghum sp. , KOH 2%; 2,5% dan 10%, HCl 1% dan 2%, Clorox, gliserin 50%, pewarna biru tripan.
Alat yang digunakan adalah kompas, pH tanah, pengukur tinggi pohon, plastik, koran, label, karet gelang, patok, pemukul, tali rafia, jerigen, spidol permanen, tustel, bak semai, pot, sendok tanah, cangkul, golok, saringan tanah bertingkat ukuran 750, 250, 100 dan 50 μm, sentrifus, mikroskop stereo dan mikroskop binokuler, cawan petri, gelas obyek, gelas penutup, pinset spora, pinset, erlenmeyer, pipet, botol film, sarung tangan dan tabung reaksi.
C. Metode Penelitian
Secara garis besar penelitian ini dibagi menjadi 2 bagian yaitu :
biakan pot cendawan (trapping) dan analisa sifat-sifat tanah khususnya salinitas.
2. Kegiatan di rumah kaca dan di laboratorium meliputi isolasi dan identifikasi CMA, yang menggunakan perbanyakan biakan pot dan persentase kolonisasi CMA pada akar tumbuhan.
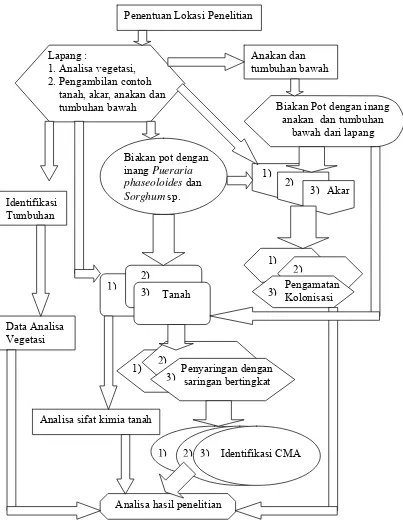
Bagan alir penelitian yang dilakukan seperti tersebut, dapat dilihat pada Gambar 2.
C.1. Di Lapang
C.1.1. Analisa Vegetasi
Analisa vegetasi dimulai dengan melakukan pengukuran terhadap fase pohon dengan keliling batas tinggi dada / 1,30 m (О > 62,8 cm) dan tiang (О : 31,4 – 62,8 cm) juga (О < 6,28 cm) anakan dan tumbuhan bawah.
Metode kuadran (Cox 1972) digunakan untuk menghitung nilai kerapatan relatif, frekuensi relatif, dominasi relatif dan indek nilai penting jenis pohon dan tiang. Metode garis menyinggung digunakan untuk analisa vegetasi fase anakan dan tumbuhan bawah. Penentuan tumbuhan dominan dilakukan berdasarkan nilai kerapatan relatif, frekuensi relatif, dominasi relatif dan indeks nilai penting jenis.
Fase sapihan tidak diamati karena fase sapihan banyak dimanfaatkan penduduk untuk kayu bakar, sehingga pada populasi setiap jenis tumbuhan, fase ini dapat dikatakan sangat sedikit dan hanya ada di tengah hutan, yaitu daerah yang jarang dijamah oleh penduduk.
Penentuan Lokasi Penelitian
Lapang :
1. Analisa vegetasi, 2. Pengambilan contoh tanah, akar, anakan dan tumbuhan bawah
Biakan pot dengan inang Pueraria phaseoloides dan
Sorghum sp.
Biakan Pot dengan inang anakan dan tumbuhan
bawah dari lapang Anakan dan tumbuhan bawah Pengamatan Kolonisasi Penyaringan dengan saringan bertingkat Tanah Akar Identifikasi CMA Analisa sifat kimia tanah
Analisa hasil penelitian Identifikasi Tumbuhan Data Analisa Vegetasi 1) 2) 3) 1) 2) 3) 1) 2) 3) 3) 3) 2) 2) 1) 1)
[image:50.612.134.537.75.601.2]Keterangan : 1) lapang, 2) biakan pot, 3) anakan
menghadap laut lepas Samudera Hindia, karang lebih menjorok ke laut, pasir lebih kasar, landai dan lebih dangkal dengan air menggenang bila air surut. Zona tengah hutan memiliki suatu spesies tumbuhan yang rapat/mendominasi zona tersebut. Selanjutnya areal penelitian dibagi menjadi 5 zona, yaitu Pantai Barat (PB), Tengah Barat (TB), Tengah Hutan (TH), Tengah Timur (TT) dan Pantai Timur (PT). Pembagian zona tersebut berlaku untuk melihat sebaran CMA dan Tumbuhan. Pembagian zona untuk melihat perbedaan komunitas tumbuhan dan hubungannya dengan CMA, areal penelitian dibagi menjadi 3 zona, yaitu 0-70 m dari pantai (P), 71-140 m dari pantai (T) dan 141 m dari pantai sampai tengah hutan (H).
C.1.2. Pengambilan Contoh Tanah, Akar untuk Pohon dan Tiang, serta Anakan dan Tumbuhan Bawah
sama pada kuadrannya, maka hanya dipilih salah satu tumbuhan untuk diambil contoh tanah dan akarnya. Contoh tumbuhan yang tidak diketahui nama ilmiahnya di ambil sebagai contoh untuk identifikasi. Beberapa tumbuhan (anakan dan tumbuhan bawah) dalam keadaan hidup diambil sebagai contoh untuk biakan pot cendawan. Akar yang diambil sebagian disimpan sementara dalam alkohol 50% setelah dicuci bersih, selama perjalanan dari lapang ke laboratorium.
Gambar 3 Peta lokasi penelitian dan letak jalur pengamatan di hutan pantai Ujung Genteng, Desa Batu, Kabupaten Sukabumi.
C.2. Percobaan di Rumah Kaca dan Laboratorium
Sebanyak 79 contoh tanah rizosfer pohon dan tiang serta akarnya berasal dari 24 titik metoda kuadran dibawa ke rumah kaca dan laboratorium. Selain itu, juga diperoleh beberapa jenis tumbuhan anakan dan tumbuhan bawah yang digunakan untuk inang dalam biakan pot anakan.
Samudera Hindia
U
Samudera Hindia
Tenda o Keterangan :
= garis transek kuadran
= garis menyinggung
Kabupaten Sukabumi
Kegiatan penelitian di rumah kaca dan laboratorium meliputi penghitungan jumlah spora, identifikasi CMA dan mengamati persentase kolonisasi. Identifikasi CMA saat ini hanya dapat dilakukan dengan menggunakan spora. Di alam pada umumnya CMA memproduksi spora dalam jumlah sangat sedikit sehingga teknik penyuburan dengan biakan pot perlu dilakukan. Contoh tanah, akar dan tumbuhan bawah yang didapat dari lapang digunakan untuk biakan pot cendawan. Tumbuhan inang yang digunakan untuk biakan potialah Pueraria phaseoloides
dan Sorghum sp. Biakan pot dipelihara di rumah kaca kurang lebih 12 bulan, dengan pemberian pupuk Johnson tanpa P. Setelah 12 bulan dilakukan analisa biakan pot dengan menghitung jumlah spora, identifikasi spora dan persentase kolonisasi CMA. Untuk mendapatkan spora, tanah disaring dengan saringan bertingkat (Brundett et al. 1994). Selain dari biakan pot, spora dari tanah yang diambil langsung dari lapang juga disaring, untuk dihitung jumlah spora dan diidentifikasi CMAnya. Spora yang baik dipilih dan diletakkan pada kaca obyek dengan media PVLG untuk selanjutnya diidentifikasi (Schenck & Perez. 1990).
persentase kolonisasi. Teknik pewarnaan akar tumbuhan yang berasal dari kultur pot yaitu Pueraria phaseoloides dan Sorghum sp. dilakukan dengan menggunakan metoda Koske dan Gemma (1989). Metode tersebut dilakukan untuk mencegah kerusakan akar, karena akar tumbuhan inang tersebut lebih kecil dan halus daripada akar tumbuhan dari hutan. Teknik pewarnaan akar tersebut dilakukan dengan cara : akar dicuci, dimasukkan dalam KOH 2,5% dan direbus pada suhu 60-90o C sampai akar menjadi bersih kurang lebih 10-30 menit. Selanjutnya KOH dibuang, dicuci air mengalir, kemudian direndam dalam HCl 1% selama 12 jam. Setelah itu HCl dibuang, dan terakhir akar diwarnai dengan pewarna biru tripan. Akar selanjutnya disimpan dalam asam gliserol 50% sampai dilakukan pengamatan dan penghitungan persentase kolonisasi.
Penghitungan kolonisasi CMA pada akar menggunakan Metoda Visual Assay, dan dilanjutkan dengan Metoda slide (Giovanneti dan Mosse 1980). Kolonisasi ditandai adanya minimal salah satu struktur berikut ini, yaitu entry point, hifa internal, arbuskula, vesikula atau koil. Persen kolonisasi CMA dihitung dengan rumus :
Jumlah akar yang terkolonisasi
% Kolonisasi CMA = --- X 100%
total panjang akar
Data persen kolonisasi, keanekaragaman dan jumlah spora CMA di lapang dan biakan pot dibandingkan dan dihubungkan dengan data lainnya.
D. Analisa Data
Σ titik jenis A ditemukan
Frekuensi jenis A = --- x 100% Σ titik keseluruhan
Frekuensi jenis A
Frekuensi relatif jenis A = --- x 100% Σ frekuensi keseluruhan
Selain kerapatan spora CMA setiap 100 g-1
tanah kering, dihitung kekayaan (R),
dimana R = jumlah jenis yang ditemukan pada sampel), dan keragaman CMA (D). Keragaman CMA (D) dihitung dengan rumus :
S
D = ---
Log N dimana : S = jumlah total spesies pada sampel
N = jumlah total spora pada sampel
E. Analisa Pendukung
Analisa pendukung terdiri dari 2, yaitu Identifikasi Tumbuhan dan analisa beberapa sifat tanah. Tumbuhan yang belum teridentifikasi nama ilmiahnya diidentifikasi di Herbarium Bogoriense, LIPI, Bogor. Analisa sifat tanah yaitu pH, jenis tanah, salinitas, P total, P tersedia dan tekstur tanah dilakukan di Laboratorium Tanah Fakultas Pertanian IPB, Bogor.
F. Penyimpanan Spesimen
HASIL DAN PEMBAHASAN
A. Analisa Vegetasi
A.1. Hasil
Dari keseluruhan hasil analisa vegetasi di hutan pantai Semenanjung Ujung Genteng,ditemukan 68 spesies tumbuhan yang terdiri dari 40 famili. Ke 68 spesies tumbuhan tersebut terdiri dari 32 spesies fase pohon dan tiang dari 20 famili, serta 51 spesies anakan dan tumbuhan bawah dari 35 famili. Hasil analisa vegetasi hutan secara keseluruhan disajikan pada Lampiran 1 dan 2.
Tumbuhan fase pohon dan tiang secara keseluruhan pada Lampiran 1 terdiri dari 32 spesies tumbuhan. Tiga spesies tumbuhan diantaranya merupakan tumbuhan yang mempunyai indeks nilai penting (NP) tertinggi, yaitu Dysoxylum parasiticum (Meliaceae), Syzygium javanicum (Myrtaceae), dan Guettarda speciosa (Rubiaceae) dengan nilai masing-masing 38.35%, 23.08% dan 17.40%. Frekuensi ke 3 spesies tersebut juga merupakan spesies yang tercatat mempunyai nilai lebih tinggi dari tumbuhan lain ialah D. parasiticum, S. javanicum, G. speciosa dengan nilai masing-masing 13.7%, 9.59 % dan 8.22%.
Tumbuhan fase anakan dan tumbuhan bawah pada Lampiran 2 terdiri dari 51 spesies tumbuhan. Terdapat 3 spesies tumbuhan yang mempunyai indeks nilai penting tertinggi (NP), yaitu Piper cf. baccatum (Piperaceae), Nephrolepis radicans (Davaliaceae), dan Syzygium javanicum (Myrtaceae) yang masing-masing mempunyai nilai 57.64%, 31.78% dan 31.78%. Sedangkan frekuensi spesies yang tercatat mempunyai nilai lebih tinggi dari tumbuhan lain ialah Piper cf. bacatum (F=20.65%) dan S. javanicum ( F=12.08%).
Pandanus tectorius (Pandanaceae) merupakan spesies tumbuhan yang relatif banyak di semenanjung Ujung Genteng, walaupun tidak termasuk kategori pohon/tiang/anakan/tumbuhan bawah. Tumbuhan tersebut tercatat sebanyak 15 pada 6 titik pada metoda kuadran, lebih banyak dari S. javanicum yang mempunyai frekuensi dan NP tertinggi setelah D. parasiticum (Lampiran 1).
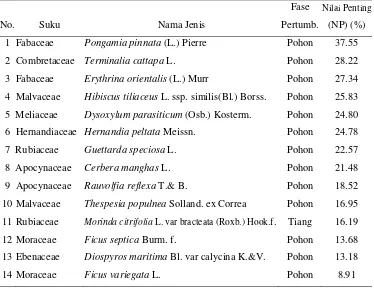
Berdasarkan komunitas tumbuhan yang dibagi dalam 3 zona, pada zona 0-70 m dp (P) (Tabel 1) terdapat 14 spesies tumbuhan fase pohon dan tiang dengan 3 spesies tumbuhan yang memiliki peran besar ialah Pongamia pinnata
(Fabaceae), Terminalia cattapa (Combretaceae), Erythrina orientalis (Fabaceae) dengan nlai masing-masing 37.55%, 28.22% dan 27.34%. Dari ke 3 tumbuhan tersebut P. pinnata dan E. orientalis memilikisebaran lebih luas (frekuensi lebih tinggi) dengan nilai yang sama yaitu 12% dari pada T. cattapa (8%) atau tumbuhan lain di zona P. Hasil analisa vegetasi selengkapnya disajikan pada Lampiran 3.
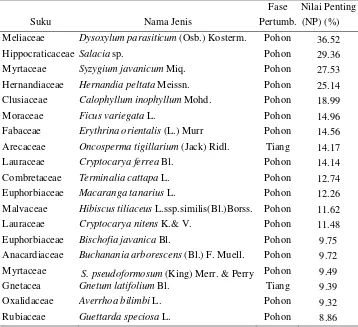
dan S. javanicum (Myrtaceae). Ketiga tumbuhan tersebut masing-masing memiliki indeks nilai penting (NP) 36.52%, 29.36% dan 27.53%. Sebaran ke 3 tumbuhan tersebut D. parasiticum merupakan tumbuhan yang memiliki sebaran paling luas dengan frekuensi 13.04 % di zona T (Lampiran 4).
Tabel 1 Nilai penting jenis tumbuhan fase pohon dan tiang pada zona 0-70 m dari pantai (TH) daerah Ujung Genteng
Fase Nilai Penting
No. Suku Nama Jenis Pertumb. (NP) (%)
1 Fabaceae Pongamia pinnata (L.) Pierre Pohon 37.55 2 Combretaceae Terminalia cattapa L. Pohon 28.22 3 Fabaceae Erythrina orientalis (L.) Murr Pohon 27.34 4 Malvaceae Hibiscus tiliaceus L. ssp. similis(Bl.) Borss. Pohon 25.83 5 Meliaceae Dysoxylum parasiticum (Osb.) Kosterm. Pohon 24.80 6 Hernandiaceae Hernandia peltata Meissn. Pohon 24.78 7 Rubiaceae Guettarda speciosa L. Pohon 22.57 8 Apocynaceae Cerbera manghas L. Pohon 21.48 9 Apocynaceae Rauvolfia reflexa T.& B. Pohon 18.52 10 Malvaceae Thespesia populnea Solland. ex Correa Pohon 16.95 11 Rubiaceae Morinda citrifolia L. var bracteata (Roxb.) Hook.f. Tiang 16.19 12 Moraceae Ficus septica Burm. f. Pohon 13.68 13 Ebenaceae Diospyros maritima Bl. var calycina K.&V. Pohon 13.18
14 Moraceae Ficus variegata L. Pohon 8.91
Pada zona 141 mdp sampai tengah hutan (H) (Tabel 3) terdapat 13 spesies tumbuhan fase pohon dan tiang yang memiliki peran besar dalam ekosistem ini ialah D. parasiticum (Meliaceae), S. javanicum (Myrtaceae) dan
dengan nilai frekuensi 21.82% di zona H. Hasil analisa vegetasi selengkapnya disajikan pada Lampiran 5.
Tabel 2 Nilai penting jenis tumbuhan fase pohon dan tiang zona di 71-140 m dari pantai (T) daerah Ujung Genteng
Fase Nilai Penting
No. Suku Nama Jenis Pertumb. (NP) (%)
1 Meliaceae Dysoxylum parasiticum (Osb.) Kosterm. Pohon 36.52
2 Hippocraticaceae Salacia sp. Pohon 29.36
3 Myrtaceae Syzygium javanicum Miq. Pohon 27.53 4 Hernandiaceae Hernandia peltata Meissn. Pohon 25.14 5 Clusiaceae Calophyllum inophyllum Mohd. Pohon 18.99
6 Moraceae Ficus variegata L. Pohon 14.96
7 Fabaceae Erythrina orientalis (L.) Murr Pohon 14.56 8 Arecaceae Oncosperma tigillarium (Jack) Ridl. Tiang 14.17 9 Lauraceae Cryptocarya ferrea Bl. Pohon 14.14 10 Combretaceae Terminalia cattapa L. Pohon 12.74 11 Euphorbiaceae Macaranga tanarius L. Pohon 12.26 12 Malvaceae Hibiscus tiliaceus L.ssp.similis(Bl.)Borss. Pohon 11.62 13 Lauraceae Cryptocarya nitens K.& V. Pohon 11.48 14 Euphorbiaceae Bischofia javanica Bl. Pohon 9.75 15 Anacardiaceae Buchanania arborescens (Bl.) F. Muell. Pohon 9.72 16 Myrtaceae S. pseudoformosum (King) Merr. & Perry Pohon 9.49 17 Gnetacea Gnetum latifolium Bl. Tiang 9.39 18 Oxalidaceae Averrhoa bilimbi L. Pohon 9.32 19 Rubiaceae Guettarda speciosa L. Pohon 8.86
Dari zona T, zona H maupun secara keseluruhan Semenanjung D. parasiticum mempunyai nilai NP dan frekuensi yang tertinggi. Jadi tumbuhan inilah yang mempunyai peranan paling besar di Hutan pantai Semenanjung Ujung Genteng.
Hasil perhitungan indeks nilai kemiripan komunitas (IS) pada ke 3 zona
tersebut di atas. menunjukkan bahwa zona P dan T mempunyai nilai IS 35.42%.
Karena nilai IS kurang dari 50% maka secara umum jenis tumbuhan fase pohon
[image:60.612.135.524.234.460.2]dan tiang pada ke 3 zona tersebut tidak menunjukkan nilai kemiripan komunitas satu dengan yang lain. Sebaran tumbuhan diseluruh semenanjung disajikan pada Gambar 9. bersama dengan sebaran CMA.
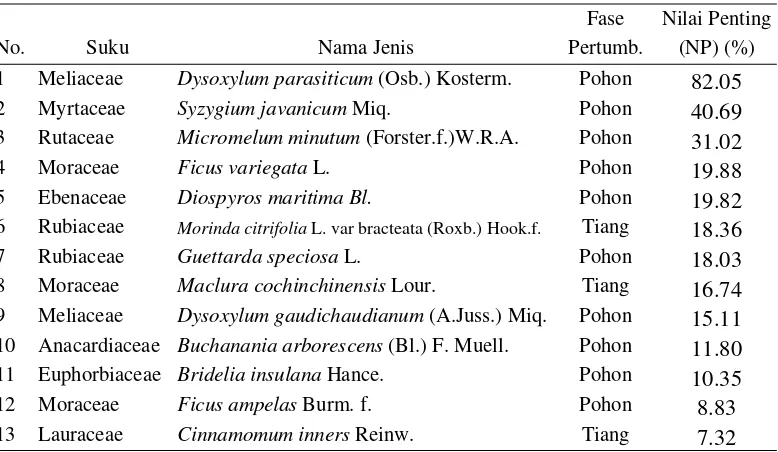
Tabel 3 Nilai penting jenis tumbuhan fase pohon dan tiang zona di 141 m dari dari pantai sampai tengah hutan (H) daerah Ujung Genteng
Fase Nilai Penting
No. Suku Nama Jenis Pertumb. (NP) (%)
1 Meliaceae Dysoxylum parasiticum (Osb.) Kosterm. Pohon 82.05 2 Myrtaceae Syzygium javanicum Miq.