SKRIPSI
PENGARUH BAKTERI ASAM LAKTAT TERHADAP PERUBAHAN
RAFFINOSEFAMILYOLIGOSACCHARIDES (RFO) PADA PROSES
PEMBUATAN TEPUNG LABU KUNING (Cucurbitamoschata)
Oleh
HURRY ZAMHOOR PRATAMA F24052173
2010
FAKULTAS TEKNOLOGI PERTANIAN INSTITUT PERTANIAN BOGOR
PENGARUH BAKTERI ASAM LAKTAT TERHADAP PERUBAHAN
RAFFINOSEFAMILYOLIGOSACCHARIDES (RFO) PADA PROSES
PEMBUATAN TEPUNG LABU KUNING (Cucurbitamoschata)
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar SARJANA TEKNOLOGI PERTANIAN Pada Departemen Ilmu dan Teknologi Pangan
Fakultas Teknologi Pertanian Institut Pertanian Bogor
Oleh
HURRY ZAMHOOR PRATAMA F24052173
2010
Judul Skripsi : Pengaruh Bakteri Asam Laktat Terhadap Perubahan Raffinose Family Oligosaccharides (RFO) Pada Proses Pembuatan Tepung Labu Kuning (Cucurbita moschata)
Nama : Hurry Zamhoor Pratama
NIM : F24052173
Menyetujui, Pembimbing I
(Prof. Dr. Ir. Rizal Sjarief SN, DESS) NIP: 19480409 197302 1 001
Pembimbing II
(Sri Usmiati, S.Pt, M.Si) NIP: 19681123 199803 2 001
Mengetahui, Ketua Departemen
(Dr. Ir. Dahrul Syah) NIP: 19650814 199002 1 001
Hurry Zamhoor Pratama. F24052173. PENGARUH BAKTERI ASAM
LAKTAT TERHADAP PERUBAHAN RAFFINOSE FAMILY
OLIGOSACCHARIDES (RFO) PADA PROSES PEMBUATAN TEPUNG LABU KUNING (Cucurbita moschata). Di bawah bimbingan Rizal Sjarief SN dan Sri Usmiati. 2010
RINGKASAN
Buah labu kuning (Cucurbita moschata) kaya akan pro-vitamin A dan di Indonesia dapat diolah menjadi berbagai panganan tradisional. Buah labu kuning diduga mengandung senyawa penyebab flatulensi. Senyawa tersebut dikenal dengan Raffinose Family Oligosaccharides (RFO), yaitu rafinosa, stakiosa dan verbaskosa. Senyawa RFO dapat direduksi dengan bantuan enzim α-galaktosidase yang berasal dari mikroba.
Penelitian ini dimaksudkan untuk menurunkan kadar RFO menggunakan bakteri asam laktat (BAL) pada proses pembuatan tepung labu kuning. Empat spesies BAL yang digunakan yaitu L. plantarum, L. brevis, B. longum dan L. mesenteroides. Perlakuan diberikan dengan merendam irisan buah labu kuning dalam suspensi BAL dengan populasi 106 dan 107 CFU/ml. Dari penelitian ini dapat dilihat besar penurunan kadar RFO dan pengaruh perendaman terhadap kadar β-karoten dan rendemen.
Hasil penelitian menunjukkan buah labu kuning segar mengandung rafinosa (612,0 mg/100 g bk), stakiosa (36,3 mg/100 g bk), dan verbaskosa (6,0 mg/100 g bk). Pemberian perlakuan perendaman dalam suspensi bakteri asam laktat dan perbedaan jumlah populasinya menghasilkan reduksi RFO yang relatif sama. Kadar RFO tepung labu kuning terendah adalah perlakuan perendaman dengan bakteri B. longum dengan populasi 107 CFU/ml sebesar 550,54 ppm atau turun sebesar 8,75% selama perendaman. Perlakuan perendaman dengan bakteri B. longum dengan populasi 107 CFU/ml juga menghasilkan kadar rafinosa dan stakiosa terendah masing-masing sebesar 389,25 ppm (turun 7,77%) dan 149,82 ppm (turun 18,47%). Kadar verbaskosa terendah diperoleh tepung labu kuning perlakuan perendaman dengan bakteri L. plantarum dengan populasi 107 CFU/ml sebesar 7,53 ppm atau turun sebesar 56,75% selama perendaman.
Kadar β-karoten buah labu kuning segar adalah 0,74 mg/100 g bb. Kadar akhir β-karoten tepung labu kuning relatif sama antara tepung labu perlakuan dan kontrol. Berdasarkan besaran nilai, kadar β-karoten tepung labu kontrol tanpa rendam adalah 269,62 ppm, perendaman dengan air destilata adalah 230,65 ppm, dan perendaman dengan bakteri rata-rata adalah 238,44 ppm. Perlakuan populasi bakteri 107 CFU/ml menghasilkan kadar β-karoten 239,50 ppm lebih tinggi dari perlakuan populasi 106 CFU/ml sebesar 237,39 ppm.
RIWAYAT HIDUP PENULIS
Penulis yang dilahirkan di Jakarta pada tanggal 12 Oktober 1987 merupakan putra pertama dari pasangan Bapak M. Zamzam dan Ibu Dwi Setyawati. Penulis menyelesaikan pendidikan di TK Pertiwi Pamulang, SD Negeri Pamulang I, SLTP Negeri 1 Pamulang dan SMA Negeri 1 Ciputat. Penulis diterima di Institut Pertanian Bogor (IPB) melalui undangan seleksi masuk pada tahun 2005 tanpa memiliki jurusan. Setelah setahun kuliah, penulis diterima di Departemen Ilmu dan Teknologi Pangan (ITP) dengan Mayor Teknologi Pangan.
Penulis pernah mengikuti acara-acara seminar atau pelatihan, diantaranya Seminar Mahasiswa Teknologi Pangan dan Ilmu Gizi Tingkat Nasional, yang diselenggarakan oleh HIMITEPA IPB, Pelatihan Sistem Manajemen Halal, Pelatihan ISO 9001 dan 22000, dan lain-lain.
KATA PENGANTAR
Segala puji dan syukur penulis panjatkan kehadirat Alloh subhanahu wa ta’ala dan sholawat serta salam tercurah kepada Nabi Muhammad
b
. Atas kehendak dan karunia-Nya, penelitian dan skripsi yang berjudul “Pengaruh Perendaman Menggunakan Bakteri Asam Laktat Pada Proses Pembuatan Tepung Labu Kuning (Cucurbita moschata)” dapat diselesaikan.Penulis menyadari bahwa skripsi ini diselesaikan dengan bantuan banyak pihak. Oleh karena itu, penulis hendak mengucapkan terima kasih kepada:
1. Ibu dan Ayah di rumah yang senantiasa mendo‟akan penulis. 2. Prof. Dr. Ir. Rizal Sjarief SN, DESS selaku pembimbing akademik. 3. Sri Usmiati, S.Pt, M.Si selaku pembimbing di Balai Litbang Pascapanen. 4. Dr. Ir. Yadi Haryadi, M.Sc yang telah bersedia menjadi penguji.
5. Dra. Sri Yuliani, Apt. selaku ketua tim penelitian di Balai Litbang Pascapanen.
6. Pak Heru, Pak Ato, Pak Yudi, Pak Tri, Pak Sis, Mba Ika, Bu Ning, staf dan teknisi di Balai Litbang Pascapanen yang telah banyak membantu selama penelitian.
7. Rino Hidayat, teman seperjuangan dan teman satu tim: Sri dan Tri yang telah membantu selama penelitian.
8. Siyam yang membantu editing.
9. Yusup, Ela, dan Hera atas konsultasi statistiknya.
10. Para moderator forum Pustakaku.net yang membantu menyediakan jurnal-jurnal.
11. Para penghuni Wisma At-Tauhid: Bombay, Angga, Dimas, Hasan, Mas Eko dan teman-teman terbaik: Achid, Asep, Adit, Agung Ridwan, Sunanto yang selalu menyemangati.
Semoga Alloh membalas seluruh kebaikan yang telah diberikan. Terakhir, penulis berharap semoga hasil penelitian dan skripsi ini bermanfaat bagi dunia teknologi pangan dan bidang-bidang lain yang terkait dengan penelitian ini.
DAFTAR ISI
Halaman
LEMBAR PENGESAHAN ... ii
RINGKASAN ... iv
RIWAYAT HIDUP PENULIS ... v
KATA PENGANTAR ... vi
DAFTAR ISI ... viii
DAFTAR GAMBAR ... x
DAFTAR TABEL ... xi
DAFTAR LAMPIRAN ... xii
I. PENDAHULUAN ... 1
A.LATAR BELAKANG ... 1
B.TUJUAN PENELITIAN ... 2
C.MANFAAT PENELITIAN ... 2
II. TINJAUAN PUSTAKA ... 3
A.LABU KUNING ... 3
B.OLIGOSAKARIDA KELUARGA RAFINOSA (RAFFINOSE FAMILY OLIGOSACCHARIDES/ RFO) ... 6
C.KAROTENOID DAN ß-KAROTEN ... 8
D.BAKTERI ASAM LAKTAT ... 10
1. Lactobacillusplantarum ... 11
2. Lactobacillusbrevis ... 11
3. Bifidobacteriumlongum ... 12
4. Leuconostocmesenteroides ... 13
III. METODOLOGI PENELITIAN ... 15
A.BAHAN DAN ALAT ... 15
1. Bahan ... 15
2. Alat ... 15
B.TEMPAT DAN WAKTU PENELITIAN ... 15
C.METODE PENELITIAN ... 15
a. Aktivasi dan propagasi ... 16
b. Pembuatan kultur kerja ... 16
c. Penentuan lama perendaman ... 17
2. Penelitian utama ... 17
3. Pembuatan tepung labu kuning ... 18
a. Persiapan bahan ... 18
b. Pengeringan menggunakan pengering kabinet ... 19
4. Perlakuan dan rancangan percobaan ... 19
5. Metode Analisis ... 20
a. Analisis Angka Lempeng Total/ Total Plate Count (TPC) (BAM, 2009) ... 20
b. Analisis Kadar β-karoten ... 21
c. Analisis Kadar Oligosakarida ... 21
d. Analisis Kadar Air (AOAC, 1995)... 22
IV. HASIL DAN PEMBAHASAN ... 23
A.PENELITIAN PENDAHULUAN ... 23
1. Pembuatan kultur kerja ... 23
2. Pemilihan lama perendaman ... 24
B.PENELITIAN UTAMA ... 24
1. Kadar Oligosakarida ... 24
a. Rafinosa ... 27
b. Stakiosa ... 31
c. Verbaskosa ... 33
2. Kadar β-karoten ... 37
3. Rendemen ... 40
V. SIMPULAN DAN SARAN ... 42
A.SIMPULAN ... 42
B.SARAN ... 42
DAFTAR PUSTAKA ... 43
DAFTAR GAMBAR
Halaman
Gambar 1 Buah labu kuning ... 3
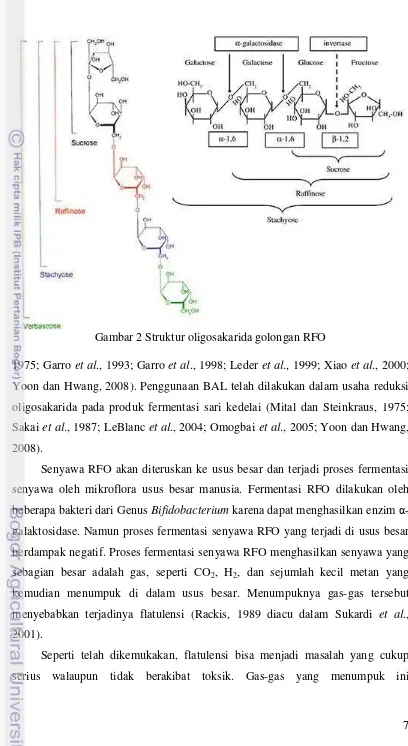
Gambar 2 Struktur oligosakarida golongan RFO... 7
Gambar 3 Struktur β-karoten ... 9
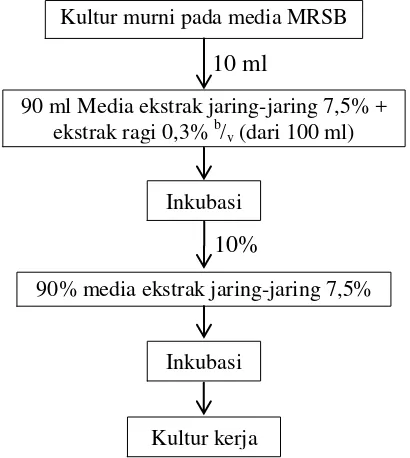
Gambar 4 Diagram alir pembuatan kultur kerja ... 16
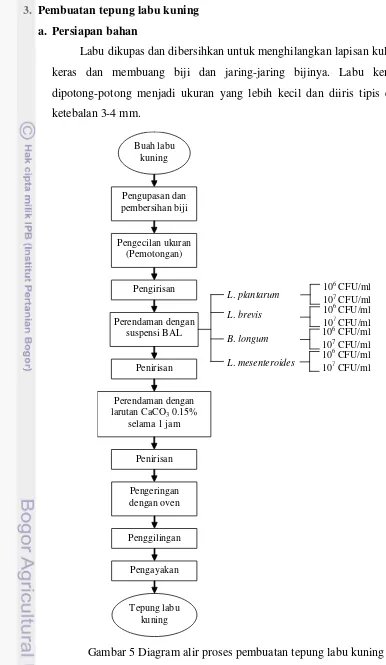
Gambar 5 Diagram alir proses pembuatan tepung labu kuning ... 18
Gambar 6 Pengering kabinet ... 19
Gambar 7 Kultur kerja dengan media ekstrak jaring-jaring... 23
DAFTAR TABEL
Halaman
Tabel 1 Komposisi kimia buah labu kuning per 100 gram ... 5
Tabel 2 TPC (CFU/ml) kultur kerja dua konsentrasi media ekstrak jaring-jaring . 23 Tabel 3 Enumerasi bakteri asam laktat pada kultur kerja ... 24
Tabel 4 Pengaruh perlakuan terhadap kadar senyawa total RFO (ppm) ... 25
Tabel 5 Reduksi total RFO (%) tepung labu kuning setelah perlakuan ... 27
Tabel 6 Pengaruh perlakuan terhadap kadar rafinosa (ppm) ... 28
Tabel 7 Reduksi rafinosa (%) tepung labu kuning setelah perlakuan ... 29
Tabel 8 Pengaruh perlakuan terhadap kadar stakiosa (ppm) ... 31
Tabel 9 Reduksi stakiosa (%) tepung labu kuning setelah perlakuan ... 32
Tabel 10 Pengaruh perlakuan terhadap reduksi verbaskosa (ppm) ... 34
Tabel 11 Reduksi verbaskosa (%) tepung labu kuning setelah perlakuan ... 35
Tabel 12 Pengaruh perlakuan terhadap kadar β-karoten (ppm) ... 38
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Foto-foto proses pengolahan ... 49
Lampiran 2. Foto-foto hasil pengeringan ... 51
Lampiran 3. Rekapitulasi data perlakuan ... 52
Lampiran 4. Data TPC kultur kerja ... 53
Lampiran 5. Peak HPLC standar oligosakarida dan β-karoten ... 54
Lampiran 6. Peak HPLC analisis oligosakarida ... 55
Lampiran 7. Peak HPLC analisis b-karoten ... 56
I. PENDAHULUAN
A. LATAR BELAKANG
Labu kuning (Cucurbita moschata) merupakan salah satu jenis tanaman yang tersebar di beberapa wilayah di Indonesia. Buah labu kuning selama ini telah populer dibuat kolak dan disayur padahal pemanfaatannya bisa diperluas dengan menerapkan teknologi pengolahan lebih lanjut misalnya menjadi tepung. Dalam bentuk tepung, labu kuning dapat diolah menjadi produk turunan lainnya yang lebih beragam atau dapat dengan mudah dikombinasikan (komposit) dengan tepung lainnya. Tepung komposit labu kuning telah diaplikasikan dalam pembuatan roti dan bubur bayi.
Pembuatan tepung dari labu kuning sebenarnya bukan hal yang baru. Beberapa negara maju di Amerika dan Eropa telah menjadikan tepung labu kuning sebagai salah satu komoditi industri lokal. Di Indonesia, industri tepung labu kuning belum banyak berkembang karena kebutuhan pasar akan tepung labu kuning masih relatif kurang.
Labu kuning mengandung zat gizi penting. Salah satu komponen terpenting dalam labu kuning adalah β-karoten dan karotenoid lainnya. Hal ini bisa diketahui dari warna daging buah labu tersebut yang menunjukkan tingginya kandungan karotenoid pada buah labu kuning. Senyawa β-karoten merupakan pro-vitamin A dan di dalam tubuh akan diubah menjadi vitamin A. Labu kuning diharapkan bisa menjadi solusi dalam permasalahan defisiensi vitamin A. Defisiensi vitamin A cukup menjadi masalah utama di Indonesia. Sebagian besar balita di Indonesia mengalami defisiensi vitamin A subklinis.
tidak memiliki enzim α-galaktosidase untuk memutus ikatan α-galaktosida pada RFO. Oligosakarida yang berada di usus besar akan difermentasi oleh mikroba. Flatulensi bisa menjadi masalah yang cukup serius walaupun tidak berakibat toksik. Flatulensi terjadi karena terjadi penumpukan gas-gas yang menyebabkan mual, keram perut, diare, dan ketidaknyamanan pada perut. Efek flatulensi menjadi salah satu penyebab dari kurang termanfaatkannya buah labu kuning.
Pada proses penepungan labu kuning, oligosakarida terutama RFO dapat direduksi sebelum proses pengeringan. Untuk dapat mereduksi RFO diperlukan enzim α-galaktosidase yang dapat diperoleh dari mikroba seperti bakteri, kapang, dan khamir. Permasalahan lain dalam pengolahan labu kuning menjadi tepung adalah kadar β-karoten pada produk akhirnya mengalami penurunan akibat ketidakstabilannya terhadap oksidasi dan suhu yang tinggi. Proses pengeringan labu kuning perlu memperhatikan suhu dan lama waktu proses yang efektif dalam mengurangi kadar air.
Pada penelitian ini, digunakan beberapa bakteri jenis BAL untuk tujuan mereduksi jumlah RFO pada buah labu kuning dan mendapatkan jenis bakteri asam laktat dan populasi terbaik dan efektif mereduksi RFO. Pada akhir penelitian, tepung labu kuning yang dihasilkan diharapkan memiliki kadar RFO yang rendah.
B. TUJUAN PENELITIAN
Penelitian ini bertujuan untuk menurunkan kadar RFO dalam pembuatan tepung labu kuning dengan jumlah maksimal 0,3 g/kg bk tepung dengan menggunakan proses perendaman menggunakan bakteri asam laktat.
C. MANFAAT PENELITIAN
II. TINJAUAN PUSTAKA
A. LABU KUNING
Labu kuning sudah dikenal masyarakat sejak berabad-abad yang lalu. Saat ini pemanfaatan labu kuning sudah semakin luas. Labu kuning yang berwarna jingga ketika telah matang digunakan sebagai bahan baku pembuat kue atau sebagai dekorasi. Daging buahnya yang kesat dan aroma yang kuat maka penyajian labu kuning dilakukan dengan dipanggang (Carew, 1977). Labu kuning telah diolah menjadi tepung yaitu dengan mengeringkan irisan daging buahnya hingga kadar air kurang dari 6% kemudian digiling. Di beberapa negara maju, labu kuning telah diolah menjadi selai, jeli, kue, dan produk kalengan (Nurhayati, 2005). Di Indonesia, pemanfaatan labu kuning belum optimal. Labu kuning yang dipanen muda biasanya untuk disayur dan yang dipanen tua dibuat menjadi panganan tradisional seperti kolak dan dodol, sementara bijinya diolah menjadi kwaci (Primasari, 2006).
Labu kuning atau dikenal dengan nama lain labu parang atau waluh merupakan buah dari tanaman menjalar yang termasuk ke dalam Kelas Dicotyledone dan Familia Cucurbitaceae. Genus Cucurbita terdiri atas lima spesies yaitu Cucurbita argyrosperma Huber, C. ficifolia Bouché, C. moschata (Duchesne ex Lam.) Duchesne ex Poiret, C. maxima Duchesne ex Poiret, dan C. pepo L. (Saade dan Hernández, undate). Tanaman labu kuning berasal dan awal penyebaran adalah dari benua Amerika (Bisognin, 2002). Spesies C. moschata
telah dibudidayakan di India, Angola, Jepang, dan Pulau Jawa sejak sepuluh tahun terakhir abad XIV (Saade dan Hernández, undate). Tanaman labu kuning berbentuk semak yang tumbuh merambat dengan bentuk batang segilima. Bagian tanaman yang dimanfaatkan adalah buahnya. Buah labu kuning berbentuk bulat, berukuran besar dan berwarna kuning kecoklatan. Berat rata-rata 3-5 kg tetapi ada yang mencapai 15 kg (Novary, 1999).
Tanaman labu kuning dapat tumbuh pada dataran rendah maupun tinggi. Ketinggian tempat ideal adalah antara 0-1500 m dpl dengan kondisi suhu 18-27°C dan pH tanah 5,5-7,0. Tanaman labu kuning toleran terhadap kekeringan, relatif sedikit membutuhkan air dan sensitif terhadap genangan air. Kelembaban yang tinggi berbahaya karena dapat menyebabkan perkembangan penyakit daun sehingga tidak ada satupun spesies Cucurbita yang tumbuh baik pada daerah tropis yang lembab (Widjaya dan Sukprakarn, 1994).
Pemanenan labu dilakukan dengan cara memotong buah dari batang dan meninggalkan sebagian tangkainya (Thompson et al., 1957). Widjaya dan Sukprakarn (1994) menyatakan labu yang dipanen dalam keadaan matang dipotong dari tangkainya setelah ditanam selama 90-120 hari. Mutu buah labu dan daya awetnya selama penyimpanan ditentukan oleh tingkat kematangan buah pada waktu pemetikan. Tingkat kematangan yang tepat akan mengurangi kerusakan dan mempunyai umur kesegaran yang lebih panjang. Buah labu yang dipetik muda segera mengalami perubahan sifat fisiko-kimia dan menyebabkan kerusakan buah (Budiman et al., 1984).
Menurut Middleton (1977), labu yang ditanam di daerah tropis memiliki aroma yang khas, berkadar air tinggi (sekitar 95%) dan unsur lain yang jumlahnya kecil. Labu kuning mengandung beberapa vitamin dan mineral yang dibutuhkan oleh tubuh. Buah labu kuning mengandung air, karbohidrat, vitamin dan mineral. Karbohidrat labu kuning terdiri atas selulosa, pati, hemiselulosa, dan pektin. Hampir semua jenis buah mengandung kadar pati yang cukup tinggi namun beberapa di antaranya menurun setelah pematangan (Meyer, 1982). Komposisi kimia dari buah labu kuning disajikan pada Tabel 1.
Tabel 1 Komposisi kimia buah labu kuning per 100 gram
Komponen Satuan Jumlah
merupakan pigmen warna alami yang banyak tersebar pada tanaman. Sebagian besar karotenoid adalah prekursor vitamin A (pro-vitamin A) yang berarti bila dikonsumsi akan dimetobolisme oleh tubuh menjadi vitamin A. Labu kuning dapat menjadi sumber pro-vitamin A yang baik. Besarnya kadar karotenoid buah labu kuning dipengaruhi oleh perbedaan varietas dan tingkat kematangannya (Gross, 1991).
B. OLIGOSAKARIDA KELUARGA RAFINOSA (RAFFINOSE FAMILY
OLIGOSACCHARIDES/ RFO)
Oligosakarida merupakan gula dengan tiga hingga dua puluh unit sakarida. Oligosakarida merupakan rantai pendek polisakarida (Manning et al., 2004). Karakteristik senyawa oligosakarida (Manning dan Gibson, 2004) adalah a) terdiri atas susunan monosakarida antara lain glukosa, galaktosa, xylosa, dan fruktosa, b) memiliki ikatan glikosidik yang terdiri dari ikatan ß-(1,4), α-(1,4), ß-(1,6), dan α -(1,6) (Wilbraham dan Matta, 1992), dan c) memiliki berat molekul yang rendah di bawah polisakarida.
Umumnya, senyawa-senyawa oligosakarida tidak dapat dicerna oleh sistem pencernaan manusia. Oligosakarida tersebut memiliki ikatan glikosidik tertentu yang memang mukosa usus manusia tidak menghasilkan enzim yang memutus ikatan tersebut. Contoh oligosakarida adalah rafinosa dan keluarganya (RFO): stakiosa dan verbaskosa, yang merupakan turunan dari α-galaktosil sukrosa. Struktur dari senyawa RFO dapat dilihat pada Gambar 2.
1975; Garro et al., 1993; Garro et al., 1998; Leder et al., 1999; Xiao et al., 2000; Yoon dan Hwang, 2008). Penggunaan BAL telah dilakukan dalam usaha reduksi oligosakarida pada produk fermentasi sari kedelai (Mital dan Steinkraus, 1975; Sakai et al., 1987; LeBlanc et al., 2004; Omogbai et al., 2005; Yoon dan Hwang, 2008).
Senyawa RFO akan diteruskan ke usus besar dan terjadi proses fermentasi senyawa oleh mikroflora usus besar manusia. Fermentasi RFO dilakukan oleh beberapa bakteri dari Genus Bifidobacterium karena dapat menghasilkan enzim α -galaktosidase. Namun proses fermentasi senyawa RFO yang terjadi di usus besar berdampak negatif. Proses fermentasi senyawa RFO menghasilkan senyawa yang sebagian besar adalah gas, seperti CO2, H2, dan sejumlah kecil metan yang
kemudian menumpuk di dalam usus besar. Menumpuknya gas-gas tersebut menyebabkan terjadinya flatulensi (Rackis, 1989 diacu dalam Sukardi et al., 2001).
Seperti telah dikemukakan, flatulensi bisa menjadi masalah yang cukup serius walaupun tidak berakibat toksik. Gas-gas yang menumpuk ini
menyebabkan tanda-tanda seperti mual, keram perut, diare, dan ketidaknyamanan pada perut (Rackis et al., 1970 diacu dalam Tanaka et al., 1975). Karena itulah senyawa RFO disebut sebagai faktor anti-nutrisi yang perlu direduksi jumlahnya pada bahan pangan. Oligosakarida umumnya terdapat pada biji-bijian, kacang-kacangan, dan ubi-ubian. Jumlah oligosakarida maksimal terkonsumsi hingga diasumsikan tidak menimbulkan efek anti-nutrisi adalah sebesar 0,3 g/kg berat badan/hari (Oku, 1995 diacu dalam Lianawati, 1997).
C. KAROTENOID DAN ß-KAROTEN
Karotenoid merupakan salah satu di antara pigmen-pigmen yang paling penting dan tersebar luas di alam. Karoten menyebabkan warna kuning, jingga, merah dan ungu, dapat ditemukan pada tanaman tingkat tinggi, alga, fungi, dan bakteri, baik dalam jaringan nonfotosintetik maupun jaringan fotosintetik bersama klorofil. Karoten dikenal sebagai pewarna alami yang tidak bersifat racun dalam bahan pangan dan telah dikenali sebagai substansi kimia sebelum dikenal vitamin A (Bauernfeind, 1981). Karotenoid mendapatkan namanya dari senyawa yang mewakili golongannya yaitu β-karoten, pigmen jingga yang pertama kali diisolasi dari wortel oleh Wackenroder pada tahun 1831 (Gross, 1991).
Karotenoid merupakan polimer isoprenoid yang terbentuk dengan bergabungnya delapan unit C5H8. Secara struktural, karotenoid dibagi ke dalam
dua golongan besar berdasarkan keberadaan gugus fungsional spesifiknya, yaitu karotenoid hidrokarbon (hydrocarbon carotenoid, C40H56) yang hanya terdiri atas
Gambar 3 Struktur β-karoten
Karotenoid merupakan lipid, oleh karena itu dapat larut dalam lipid lainnya dan dalam pelarut lemak seperti aseton, alkohol, dietil eter, dan kloroform. Golongan karoten larut dalam pelarut non-polar seperti petroleum eter dan heksan sedangkan golongan xantophil larut dengan sangat baik pada pelarut polar seperti alkohol (Gross, 1991). Senyawa karotenoid stabil di dalam sel tanaman namun isolatnya mudah mengalami perubahan molekul yaitu isomerisasi cis-trans dan degradasi oleh panas, cahaya, oksigen, sedikit asam, dan senyawa aktif permukaan seperti silika (Ball, 2005).
Sebagian besar sumber vitamin A adalah karoten yang banyak terdapat dalam bahan pangan nabati. Senyawa vitamin A aktif dipresentasikan oleh retinoid dan prekursor karotenoid vitamin A (provitamin A karotenoid). Telah jelas bahwa karotenoid membantu fungsi-fungsi selular sebagai prekursor vitamin A (Stahl et al., 1994). Aktivitas antioksidan karotenoid baik dari provitamin A maupun non-provitamin A dihasilkan dari interaksi langsung dengan spesies oksigen aktif. Karoten penting untuk penglihatan, pertumbuhan, diferensiasi jaringan, reproduksi, serta perawatan sistem kekebalan (Ball, 2000).
Penyebab utama hilangnya karoten pada sayuran adalah oksidasi sebagai akibat tingginya struktur ikatan tak jenuh pada karotenoid. Degradasi karotenoid dapat terjadi karena: a) autooksidasi yang berlangsung secara spontan dan menyebabkan reaksi berantai radikal bebas dengan adanya oksigen, b) fotooksidasi yang dihasilkan oksigen dengan adanya cahaya, c) coupled oxidation dalam sistem yang mengandung lemak (Gross, 1991 diacu dalam Kidmose et al., 2002). Kerusakan enzimatis dapat terjadi terutama disebabkan enzim lipoksigenase. Enzim lipoksigenase terdapat secara luas pada sayuran yang mengandung klorofil dan telah dilapokan bahwa kehilangan karotenoid berhubungan dengan aktivitas enzim tersebut (Hutchings, 1999). Dalam bentuk larutan maupun kristal, karotenoid mengalami autooksidasi ketika ada oksigen melalui proses radikal bebas berantai. Proses oksidasi ini distimulasi oleh suhu, cahaya, kelembaban, dan beberapa jenis logam (Gross, 1991).
D. BAKTERI ASAM LAKTAT
Bakteri asam laktat merupakan sekelompok bakteri Gram positif yang memiliki kesamaan karakteristik secara morfologi, metabolik, dan fisiologis. Golongan bakteri ini adalah tidak berspora, sel berbentuk bulat atau batang dan memproduksi asam laktat sebagai hasil akhir utama proses fermentasi karbohidrat (Axelsson, 2004).
Menurut sejarah, Genus Lactobacillus, Leuconostoc, Pediococcus dan Streptococcus merupakan awal dan inti dari kelompok BAL. Pada perkembangan selanjutnya, terjadi revisi taksonomi dan muncul genus baru hingga saat ini terdapat sekitar 20 genus. Namun, genus-genus yang penting dalam sudut pandang teknologi pangan yaitu Aerococcus, Carnobacterium, Enterococcus, Lactobacillus, Lactococcus, Lactosphaera, Leuconostoc, Oenococcus, Pediococcus, Streptococcus, Tetragenococcus, Vagococcus, dan Weissella. Genus Bifidobacterium sering dianggap memilki sifat tipikal yang sama dengan BAL asli padahal berbeda secara filogenetik dan keunikannya dalam memfermentasi gula (Axelsson, 2004).
heterofermentatif. Proses homofermentatif menghasilkan produk akhir hanya asam laktat melalui jalur glikolisis. Proses heterofermentatif menghasilkan produk akhir sampingan seperti etanol, asetat, dan CO2 selain asam laktat melalui jalur
6-fosfoglukonat atau fosfoketolase (Axelsson, 2004).
1. Lactobacillusplantarum
Lactobacillus plantarum termasuk salah satu spesies Lactobacillus yang sering ditemui pada pikel, sawi asin dan sauerkraut. Lactobacillus plantarum merupakan bakteri asam laktat yang utama dan akhir pada proses fermentasi sayuran. Hal tersebut dikarenakan bakteri ini memiliki perbedaaan metabolisme dan toleran terhadap kondisi pH rendah. Lactobacillus plantarum berbentuk batang lurus dengan kisaran lebar 0,9-1,2 µm dan panjang 3-8 µm, berukuran tunggal atau membentuk rantai pendek serta merupakan Gram-positif (Li, 2004). Lactobacillus plantarum mampu memfermentasi glukosa membentuk produk asam D-L-laktat tanpa gas atau dikatakan bersifat homofermentatif. Bakteri ini juga dapat memfermentasi amigladin, selobiosa, laktosa, manitol, sukrosa, galaktosa, maltosa, sorbitol, dan trehalosa. Kemampuan dalam memfermentasi melibiosa dan rafinosa membedakan L. plantarum dengan L. casei (Ono et al., 1992).
Koloni L. plantarum berwarna putih atau kuning dan beberapa galur bersifat motil dan pada media agar mempunyai ciri-ciri bulat, licin, padat, putih, kadang-kadang kuning terang atau gelap, berdiameter 3 mm, bersifat anaerobik fakultatif. Bakteri ini dapat tumbuh pada suhu 15°C pada umumnya dan tidak dapat tumbuh pada suhu 45°C, dengan suhu optimalnya berkisar 30-35°C (Gilliland, 1986).
2. Lactobacillusbrevis
heterofermentatif obligat menghasilkan asam laktat, asam asetat, CO2, dan
etanol dari fermentasi karbohidrat (Teixeira, 1999).
Optimal tumbuh pada suhu sekitar 30°C dan pada kondisi pH rendah yaitu 4,0-5,0 (Teixeira, 1999). Bakteri L. brevis tumbuh pada APT broth dengan pH 3,0 yang diasamkan oleh asam sitrat, hidroklorat, fosfat, atau tartarat, pH 3,7 untuk asam laktat, dan pH 4,0 untuk asam asetat (Juven, 1976 diacu dalam ICMSF, 1980). Bakteri ini dapat tumbuh hampir enam kali lebih baik dalam kondisi aerobik (diagitasi) daripada dalam kondisi anaerobik (Stamer dan Stoyla, 1967).
3. Bifidobacteriumlongum
Bakteri dari genus Bifidobacterium pertama kali diisolasi pada tahun 1899 dari bayi sehat yang minum ASI oleh Tissier dari Institut Pasteur di Prancis. Bersifat anaerobik, Gram-positif, tidak membentuk spora, batang pleomorfik, dan dulunya dinamakan Bacillus bifidus communis. Namanya menunjukkan cabang morfologi dari bakteri; bifidus merupakan bahasa Latin yang berarti „membelah menjadi dua bagian‟. Terakhir, bakteri ini disatutempatkan bersama genus Lactobacillus sebagai L. bifidus. Di tahun 1960-an, bakteri ini diterima sebagai genus tersendiri dan diklasifikasikan sebagai Bifidobacterium. Selain dari sifat-sifat yang telah disebutkan sebelumnya, karakteristik fenotip utama dari Bifidobacterium adalah memproduksi asam laktat dan asetat sebagai produk utama dari fermentasi glukosa. Bifidobacterium longum merupakan salah satu dari sekitar tiga puluh spesies bifidobakteria yang berasal dari manusia dan merupakan mikroflora usus manusia (Ishibashi et al., 1997).
Meskipun bifidobakteria bersifat anaerob obligat, terdapat toleransi terhadap keberadaan oksigen yang bergantung pada spesies dan jenis media. Selain itu dengan adanya CO2, sensitivitas terhadap O2 juga bervariasi
untuk pertumbuhan adalah 6,5-7,0 dan tidak ada pertumbuhan pada pH di bawah 5 atau di atas 8 (Ballongue, 2004). Bifidobacterium longum dan beberapa spesies Bifidobacterium lainnya kecuali B. bifidum mampu memfermentasi oligosakarida seperti rafinosa (Benno, 1995).
4. Leuconostocmesenteroides
Bakteri ini bersifat heterofermentatif. Pada kondisi mikroaerofilik, glukosa diubah menjadi asam D-laktat, etanol, dan CO2 dalam jumlah molar
yang sama. Semua genus Leuconostoc adalah fakultatif anaerob. Suhu optimum pertumbuhan adalah antara 20-30°C dan suhu minimum pertumbuhan adalah 5°C. Leuconostoc mesenteroides subsp. mesenteroides memiliki waktu generasi yang pendek dan pertumbuhan yang baik diperoleh dalam 24 jam inkubasi pada suhu 30°C (Garvie, 1986 diacu dalam Dellaglio et al., 1995). Pertumbuhan L. mesenteroides berhenti ketika pH mencapai 5,4-5,7 (McDonald et al., 1990).
Sel L. mesenteroides merupakan Gram positif, tidak membentuk spora, dan tidak motil. Bentuk selnya bervariasi tergantung kondisi pertumbuhannya. Sel yang tumbuh pada media glukosa menjadi memanjang dan bentuknya nampak seperti laktobasili ketimbang streptokoki. Beberapa galur, selnya berbentuk kokus ketika ditumbuhkan pada media susu. Selnya bisa terlihat soliter atau dalam pasangan membentuk rantai pendek hingga sedang. Ketika ditumbuhkan pada media padat, selnya memanjang dan dianggap berbentuk batang (Garvie, 1986 diacu dalam Dellaglio et al., 1995). Beberapa galur dari L. mesenteroides memproduksi dekstran ekstraselular yang membentuk jaket padat elektron pada permukaan sel (Brooker, 1977).
(Dellaglio et al., 1995). Dari semua Leuconostoc yang diisolasi dari tanaman, L. mesenteroides subsp. mesenteroides merupakan yang paling dominan dan berperan penting dalam proses fermentasi beragam produk fermentasi tanaman dan sayuran (Mundt, 1970 diacu dalam Dellaglio et al., 1995).
III. METODOLOGI PENELITIAN
A. BAHAN DAN ALAT 1. Bahan
Bahan-bahan yang digunakan dalam penelitian ini adalah buah labu kuning umur 2,5-3 bulan diperoleh dari Kaponan - Jawa Tengah, kultur bakteri asam laktat Lactobacillus plantarum, Lactobacillus brevis, dan Bifidobacterium longum koleksi Balai Besar Penelitian dan Pengembangan Pascapanen Pertanian, kultur Leuconostoc mesenteroides dari koleksi Universitas Gadjah Mada, air destilata, CaCO3, de Man Rogosa Sharpe
Agar/MRSA (Oxoid), de Man Rogosa Sharpe Broth/MRSB (Oxoid), ekstrak khamir, NaCl fisiologis, standar (rafinosa – stakiosa – verbaskosa (Sigma) dan β-karoten) dan bahan-bahan kimia untuk analisis kadar oligosakarida dan β -karoten.
2. Alat
Alat-alat yang digunakan adalah pengering kabinet, blender (Philips), blender kering (National), laminar (Esco), mikropipet (Socorex), inkubator (Harstra), lemari pendingin (Sanyo), timbangan digital, timbangan analitik (Precisa), vortex (Barnstead International), otoklaf (Hirayama, Jepang), dan sistem HPLC, serta alat-alat gelas sepeti tabung reaksi, tabung reaksi bertutup, cawan Petri, pipet serologis (Iwaki – Pyrex), dan botol gelas.
B. TEMPAT DAN WAKTU PENELITIAN
Penelitian dilaksanakan di Balai Besar Penelitian dan Pengembangan Pascapanen Pertanian Cimanggu Bogor dan berlangsung sejak Juni 2010 hingga Desember 2010.
C. METODE PENELITIAN 1. Penelitian pendahuluan
a. Aktivasi dan propagasi
Kultur dari stok (agar miring, agar tusuk, media susu, dan atau media MRSB) diaktifkan pada media MRSB 10 ml, diinkubasi pada suhu 37°C selama 2 hari, setelah itu disimpan dalam refrigerator sebagai kultur stok. Untuk propagasi memperbanyak kultur diambil 1 ml kultur stok kemudian diinokulasikan ke media MRSB 9 ml dan diinkubasi pada suhu 37°C selama 2 hari. Hasilnya merupakan kultur intermediate.
b. Pembuatan kultur kerja
Kultur kerja dibuat dengan memanfaatkan jaring-jaring biji buah labu kuning. Jaring-jaring biji ditimbang dan ditambah air hingga diperoleh konsentrasi yang diinginkan kemudian diblender hingga halus. Bubur jaring-jaring kemudian disaring untuk memisahkan potongan serat yang masih berukuran besar. Hasil saringan kemudian disebut ekstrak jaring-jaring. Percobaan awal dilakukan untuk melihat pertumbuhan bakteri asam laktat pada media ekstrak jaring-jaring. Konsentrasi ekstrak dibuat 5% dan 7,5%. Media ekstrak jaring yang telah dibuat diinokulasikan bakteri asam laktat, yaitu Lactobacillus plantarum dan Lactobacillus brevis. Aktivasi dan perbanyakan kultur pada media eksrak jaring-jaring dilakukan dalam dua tahap. Proses pembuatan kultur kerja dapat dilihat pada Gambar 4.
Kultur murni pada media MRSB
90 ml Media ekstrak jaring-jaring 7,5% + ekstrak ragi 0,3% b/v (dari 100 ml)
90% media ekstrak jaring-jaring 7,5%
10%
Kultur kerja Inkubasi
Inkubasi
Setelah inkubasi selama dua hari, enumerasi total bakteri dilakukan terhadap kultur kerja dari kedua spesies bakteri dengan metode TPC. Konsentrasi ekstrak jaring yang dipilih adalah yang memiliki nilai TPC minimal 108 CFU/ml. Pembuatan kultur kerja selanjutnya menggunakan konsentrasi yang terpilih untuk memperbanyak empat spesies BAL yang akan digunakan dalam penelitian ini.
c. Penentuan lama perendaman
Perendaman irisan labu dengan kultur dilakukan selama empat pengamatan waktu yaitu: 12, 15, 18, dan 21 jam untuk setiap perlakuan dan kontrol. Pemilihan lamanya perendaman ditentukan dari penampakan visual irisan labu kering dan juga mempertimbangkan efektifitas proses perendaman.
2. Penelitian utama
3. Pembuatan tepung labu kuning a. Persiapan bahan
Labu dikupas dan dibersihkan untuk menghilangkan lapisan kulit yang keras dan membuang biji dan jaring-jaring bijinya. Labu kemudian dipotong-potong menjadi ukuran yang lebih kecil dan diiris tipis dengan ketebalan 3-4 mm.
Gambar 5 Diagram alir proses pembuatan tepung labu kuning
b. Pengeringan menggunakan pengering kabinet
Irisan labu yang mendapat perlakuan perendaman maupun kontrol dicuci dengan air bersih lalu direndam dengan larutan CaCO3 0,15% selama
1 jam. Setelah itu, irisan labu ditiriskan atau diangin-anginkan (tempering) untuk mengurangi jumlah air pada labu setelah perendaman. Irisan labu diletakkan pada nampan-nampan dan dimasukkan ke dalam pengering kabinet (Gambar 6) dengan suhu pengeringan diatur pada 55°C. Proses pengeringan terus dilakukan hingga irisan labu „mengering‟. Irisan labu dikatakan kering bila mudah dipatahkan. Irisan labu yang sudah kering kemudian digiling lalu diayak dengan ukuran 80 mesh. Penetapan kadar RFO dan β-karoten pada tepung labu kemudian dilakukan terhadap tepung labu kuning untuk setiap jenis larutan perendam dan kontrol.
Gambar 6 Pengering kabinet
4. Perlakuan dan rancangan percobaan
Rancangan percobaan yang digunakan dalam penelitian ini adalah Rancangan Acak Lengkap (RAL) dengan variabel:
a. Spesies bakteri (A):
1. A1: Lactobacillus plantarum; 2. A2: Lactobacillus brevis;
3. A3: Bifidobacterium longum; dan 4. A4: Leuconostoc mesenteroides; b. Jumlah populasi bakteri (B):
Perlakuan yang diberikan merupakan kombinasi dari kedua variabel di atas sehingga terdapat delapan perlakuan. Sebagai pembanding terdapat perlakuan kontrol, yaitu:
A0 : Kontrol dengan perendaman air destilata; dan NON : Kontrol tanpa perendaman mikroba
Model rancangan percobaan adalah sebagai berikut: Yij = µ + Pi+ εij
Yij : Pengamatan pada perlakuan ke i dan ulangan ke j
µ : Rataan umum
Pi : Pengaruh perlakuan ke i
εij : Galat perlakuan ke i dan ulangan ke j
Setiap perlakuan dan kontrol dilakukan dengan tiga kali ulangan. Data yang diperoleh kemudian diolah menggunakan metode ANOVA (Analysis of Variance) dengan uji lanjut Duncan pada taraf 5%.
5. Metode Analisis
Keterangan:
N : Jumlah sel dalam satuan colony forming unit (CFU) per ml sampel ∑C : Total seluruh koloni pada cawan yang dapat dihitung
n1 : Jumlah cawan dari pengenceran pertama yang dihitung
n2 : Jumlah cawan dari pengenceran kedua yang dihitung
d : Nilai pengenceran dari penghitungan pertama yang digunakan
b. Analisis Kadar β-karoten
Analisis kadar β-karoten dilakukan di LIPI menggunakan metode HPLC. Sampel kering yang telah dihomogenkan ditimbang 1 gram lalu dilarutkan dalam pelarut nonpolar. Minyak diekstrak dengan pelarut aseton selama semalam. Setelah minyak terekstrak, aseton diuapkan dengan vacum evaporator. Kondisi selama pengekstrakan diusahakan agar terlindung dari cahaya. Lemak yang terekstrak dilarutkan dalam pelarut THF : Acetonitril (60 : 40). Kemudian disaring dengan miliphore 0.5 µ. Kemudian sebanyak 1 ml sampel diinjeksi ke sistem HPLC. Detektor yang digunakan adalah UV Detector pada panjang gelombang (λ) 554 nm. Kolom yang digunakan jenis C8 dengan laju alir 1 µl/menit dan suhu ambient. Perhitungan kadar β -karoten adalah sebagai berikut :
c. Analisis Kadar Oligosakarida
Analisa kadar oligosakarida dilakukan di LIPI menggunakan metode HPLC. Sampel tepung ditimbang sebanyak 1 gram kemudian diekstrak dengan metanol – air (80:20). Sebanyak 1 ml hasil ekstraksi diinjeksi ke sistem HPLC. Sistem HPLC yang digunakan memiliki kolom NH2 dengan
waktu retensi dan setiap peak menunjukkan satu jenis komponen oligosaakrida. Perhitungan kadar oligosakarida menggunakan rumus:
d. Analisis Kadar Air (AOAC, 1995)
Penetapan kadar air dilakukan dengan menggunakan oven. Cawan logam atau porselin dikeringkan dalam oven selama 30 menit dengan suhu 100 sampai 105oC. Kemudian didinginkan dalam desikator selama 30 menit dan setelah dingin segera ditimbang. Sampel sebanyak 2 gram ditimbang dan dikeringkan dalam oven pada suhu 105oC selama 6 jam sampai tercapai berat konstan, dan didinginkan dalam desikator sekitar 30 menit kemudian segera ditimbang dan dilakukan perhitungan:
Keterangan : A : Berat cawan
IV. HASIL DAN PEMBAHASAN
A. PENELITIAN PENDAHULUAN 1. Pembuatan kultur kerja
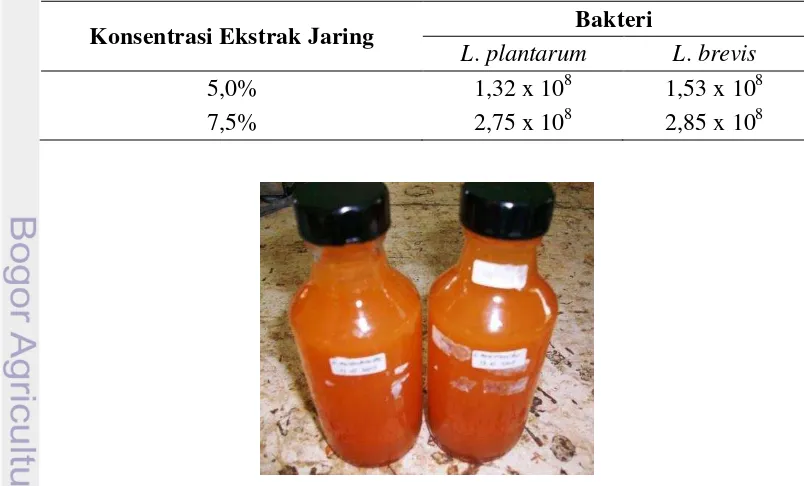
Kultur kerja menggunakan media ekstrak jaring-jaring biji labu dibuat dalam dua konsentrasi yaitu 5,0% dan 7,5%. Ekstrak khamir ditambahkan pada media ekstrak jaring-jaring untuk memberi nutrisi tambahan seperti vitamin dan protein supaya pertumbuhan bakteri optimal. Ekstrak jaring-jaring kemungkinan juga mengandung gula-gula yang terlarut dalam air sebagai nutrisi bagi bakteri. Kultur kerja yang dihasilkan tampak keruh setelah inkubasi di suhu ruang selama dua hari, terjadi pengendapan komponen yang tersuspensi pada ekstrak jaring-jaring yang tidak tersaring pada saat pembuatan ekstrak. Kultur kerja dibuat dalam botol gelas dengan volume ±150 ml (Gambar 7). Hasil enumerasi total BAL pada kultur kerja dapat dilihat pada Tabel 2. Berdasarkan nilai TPC tersebut, ekstrak jaring-jaring dengan konsentrasi 7,5% dipilih sebagai media kultur kerja untuk mempersiapkan jumlah BAL pada 108 CFU/ml (Tabel 3).
Tabel 2 TPC (CFU/ml) kultur kerja dua konsentrasi media ekstrak jaring-jaring Konsentrasi Ekstrak Jaring Bakteri
L. plantarum L. brevis
5,0% 1,32 x 108 1,53 x 108
7,5% 2,75 x 108 2,85 x 108
Tabel 3 Enumerasi bakteri asam laktat pada kultur kerja Bakteri Asam Laktat TPC (CFU/ml) L. plantarum 2,10 x 108 L. brevis 4,30 x 108 B. longum 1,40 x 1010 L. mesenteroides 3,75 x 108
2. Pemilihan lama perendaman
Irisan labu kering hasil perendaman selama 12, 15, 18, dan 21 jam masing-masing oleh empat BAL dibandingkan secara visual dengan memperhatikan warna. Hasil pengamatan visual terhadap sawut labu kuning adalah semakin lama perendaman, maka semakin pucat warnanya. Perendaman selama 18 jam kemudian dipilih dengan mempertimbangkan kenampakkan sawut labu kuning yang berwarna kuning/orange cerah.
Proses reduksi RFO kemungkinan berlangsung sejak beberapa jam pada awal perendaman menggunakan bakteri karena RFO juga merupakan sumber karbon untuk pertumbuhan bakteri dan sintesis enzim yang berperan dalam reduksi (hidrolisis) RFO. Enzim tersebut bersifat inducible (terinduksi) (Sakai et al., 1987; Roy et al., 1991; Garro et al., 1996; Xiao et al., 2000), yaitu enzim yang diproduksi karena terdapatnya substrat yang sesuai.
B. PENELITIAN UTAMA 1. Kadar Oligosakarida
Daging buah labu kuning segar yang digunakan pada penelitian ini ternyata mengandung oligosakarida rafinosa (0,6120 g/100 bk), stakiosa (0,0363 g/100 g bk), verbaskosa (0,0060 g/100 g bk) dan total oligosakarida sebesar 0,6543 g/100 g bk (0,0468 g/100 g bb) dengan kadar air sebesar 92,84%.
non-pereduksi (Mital et al., 1975). Enzim ini umumnya dihasilkan pada kondisi kultur yang ditambahkan dengan satu atau lebih kelompok gula α -D-galaktopiranosil sebagai sumber karbon, bersifat terinduksi, intraselular, dan sebagian lain berikatan dengan membran pada kebanyakan mikroba (Garro et al., 1996; Yoon dan Hwang, 2008). Beberapa galur Lactobacillus sp. dan Leuconostoc sp. dilaporkan menghasilkan enzim α-galaktosidase dengan aktivitas tertinggi pada bagian intraselular (LeBlanc et al., 2004; Yoon dan Hwang, 2008).
Hasil uji statistik menunjukkan kadar total RFO tepung labu kuning tidak berbeda nyata untuk setiap perlakuan, kontrol rendam air destilata, dan kontrol tanpa rendam. Perbedaan spesies bakteri yang digunakan dan jumlah bakteri yang disuspensikan pada perendam tidak berpengaruh nyata terhadap kadar RFO tepung labu kuning.
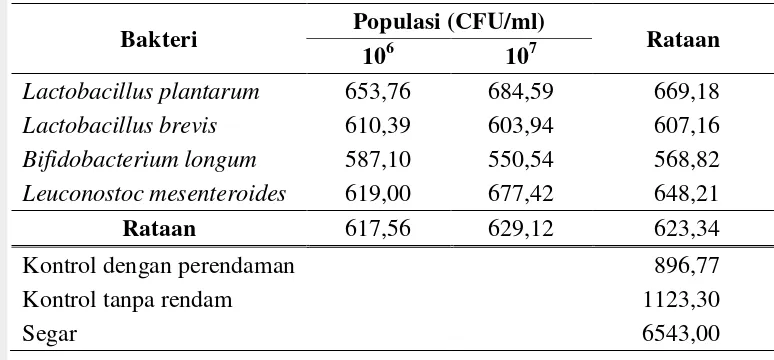
Tabel 4 Pengaruh perlakuan terhadap kadar senyawa total RFO (ppm)
Bakteri Populasi (CFU/ml) Rataan
106 107
Lactobacillus plantarum 653,76 684,59 669,18 Lactobacillus brevis 610,39 603,94 607,16 Bifidobacterium longum 587,10 550,54 568,82 Leuconostoc mesenteroides 619,00 677,42 648,21
Rataan 617,56 629,12 623,34
Kontrol dengan perendaman 896,77
Kontrol tanpa rendam 1123,30
Segar 6543,00
brevis menghasilkan kadar total RFO tepung labu kuning sebesar 610,39 ppm untuk populasi 106 CFU/ml dan turun menjadi 603,94 ppm untuk populasi 107 CFU/ml (Tabel 4).
Berdasarkan nilai rata-rata, kadar total RFO semakin rendah dengan semakin tinggi populasi bakteri yang disuspensikan ke perendam kecuali untuk perlakuan bakteri L. plantarum dan L. mesenteroides. Kadar total RFO tepung labu kuning perlakuan L. plantarum populasi 107 CFU/ml yaitu 684,59 ppm lebih tinggi dari populasi 106 CFU/ml yaitu 653,76 ppm dan untuk perlakuan L. mesenteroides populasi 107 CFU/ml yaitu 677,12 ppm lebih tinggi dari populasi 106 CFU/ml yaitu 619,00 ppm (Tabel 4). Hal tersebut kemungkinan disebabkan oleh adanya penurunan aktivitas enzim α-galaktosidase. Aktivitas enzim α-galaktosidase dapat mengalami penurunan disebabkan oleh beberapa faktor, salah satunya adalah nilai pH. Mital et al. (1973) melaporkan bahwa α -galaktosidase dari laktobasili yang diujinya memiliki aktivitas optimum pada kisaran pH 5,2-5,9. Bakteri memfermentasi gula-gula monosakarida dan disakarida lebih banyak dan lebih cepat dengan semakin banyaknya populasi bakteri sehingga menghasilkan asam laktat yang lebih banyak dan menyebabkan penurunan pH yang lebih cepat. Penurunan pH hingga lebih rendah dari pH optimum aktivitas α-galaktosidase menyebabkan fermentasi RFO menjadi lambat. Yoon dan Hwang (2008) mendapatkan aktivitas relatif α -galaktosidase bakteri L. mesenteroides turun menjadi 60% pada pH 5,5. Mital dan Steinkraus (1975) menyatakan aktivitas α-galaktosidase dari L. plantarum menurun karena pH medium (sari kedelai) telah mencapai pH 4,5 yang menyebabkan hidrolisis partial dari stakiosa.
(tertinggi 8,48% dan terendah 6,94%) sedangkan perendaman dengan air destilata menurunkan RFO sebesar 3,46%.
Tabel 5 Reduksi total RFO (%) tepung labu kuning setelah perlakuan
Bakteri Populasi (CFU/ml) Rataan
106 107
* Reduksi total. Persentase dihitung terhadap segar.
** Nilai dalam tanda kurung adalah reduksi akibat perendaman yaitu selisih antara persentase reduksi total perlakuan perendaman dan kontrol tanpa rendam.
Persentase reduksi RFO rata-rata pada tepung labu kuning untuk perlakuan populasi 107 CFU/ml sebesar 7,55%, sedikit lebih rendah dari perlakuan populasi 106 CFU/ml yaitu 7,73%. Akan tetapi, reduksi RFO menggunakan bakteri B. longum dengan populasi 107 CFU/ml sebesar 8,75%, lebih tinggi dibandingkan dengan populasi 106 CFU/ml sebesar 8,20%. Berdasarkan besarnya persentase reduksi, bakteri B longum memiliki potensi yang baik dalam mereduksi total RFO.
a. Rafinosa
kadar rafinosa tepung labu. Kadar rafinosa tiap perlakuan disajikan pada Tabel 6.
Tabel 6 Pengaruh perlakuan terhadap kadar rafinosa (ppm)
Bakteri Populasi (CFU/ml) Rataan
106 107
Lactobacillus plantarum 460,57 501,43 481,00 Lactobacillus brevis 448,03 440,14 444,08 Bifidobacterium longum 425,09 389,25 407,17 Leuconostoc mesenteroides 449,46 488,53 469,00
Rataan 445,79 454,84 450,31
Kontrol dengan perendaman 669,18
Kontrol tanpa rendam 864,87
Segar 6120,00
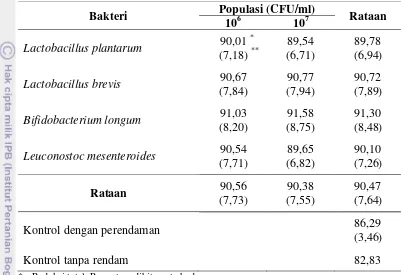
Kadar RFO terbanyak pada labu kuning adalah rafinosa yaitu sebesar 6120,00 ppm (93,5% dari total RFO). Kadar rafinosa tepung labu kuning kontrol tanpa rendam adalah 864,87 ppm. Labu kuning yang direndam dengan suspensi bakteri menghasilkan kadar rafinosa tepung labu kuning yang lebih rendah rata-rata 450,31 ppm. Kadar rafinosa paling rendah diperoleh pada tepung labu kuning perlakuan rendam bakteri B. longum dengan populasi 107 CFU/ml sebesar 389,25 ppm sedangkan populasi 106 CFU/ml sebesar 425,09 ppm. Kadar rafinosa tepung labu kuning perlakuan L. brevis menurun dari 448,03 ppm untuk populasi 106 CFU/ml menjadi 440,14 ppm untuk populasi 107 CFU/ml (Tabel 6).
disebabkan oleh penurunan aktivitas enzim α-galaktosidase akibat pH yang lebih rendah tercapai pada populasi 107 CFU/ml.
Tabel 7 Reduksi rafinosa (%) tepung labu kuning setelah perlakuan
Bakteri Populasi (CFU/ml) Rataan
106 107
* Reduksi total. Persentase dihitung terhadap segar.
** Nilai dalam tanda kurung adalah reduksi akibat perendaman yaitu selisih antara persentase reduksi total perlakuan perendaman dan kontrol tanpa rendam.
Berdasarkan Tabel 7, kadar rafinosa tepung labu kuning kontrol tanpa perendaman menurun sebesar 85,87% dari kadar rafinosa labu kuning segar. Penurunan kadar rafinosa akibat perendaman dengan air destilata sebesar 3,19% dan akibat perendaman dengan suspensi bakteri adalah sebesar rata-rata 6,78%. Penurunan kadar rafinosa terbesar terjadi untuk perlakuan dengan perendaman bakteri B. longum. Reduksi rafinosa selama perendaman mencapai 7,48%.
populasi 107 CFU/ml lebih kecil dibandingkan dengan populasi 106 CFU/ml.
Kelompok bifidobakteria dapat mengasimilasi rafinosa seefektif glukosa sebagai sumber karbon untuk pertumbuhannya (Xiao et al., 2000). Menurut Garro et al. (1999), rafinosa native maupun hasil hidrolisisnya dari stakiosa sudah tidak terdeteksi sejak awal proses fermentasi sari kedelai menggunakan B. longum. Scalabrini et al. (1998) juga melaporkan bahwa B. longum MB300 yang diujinya mereduksi rafinosa native sari kedelai hingga 56% pada proses fermentasi selama 24 jam dan tidak terdeteksi setelah fermentasi selama 48 jam. Untuk keperluan penggunaan rafinosa maka bifidobakteria menghasilkan enzim α-galaktosidase (Desjardins et al., 1990). Enzim α-galaktosidase merupakan enzim yang terinduksi dan disintesis oleh sel bakteri bila terdapat substrat yang sesuai yaitu senyawa α -galaktosida (misal rafinosa dan stakiosa).
Rendahnya bakteri L. plantarum dalam mereduksi rafinosa sejalan dengan penelitian yang dilakukan Mital dan Steinkraus (1975), bakteri L. plantarum yang diujikan hanya sedikit menghidrolisis rafinosa native sari kedelai yaitu kurang dari 40% dan sejak 12 jam fermentasi jumlah rafinosa tetap tidak berubah hingga jam ke-60. Rafinosa pada sari kedelai menjadi penyebab terinduksinya enzim α-galaktosidase pada bakteri L. plantarum (Mital dan Steinkraus, 1975).
b. Stakiosa
Hasil uji statistik menunjukkan bahwa perbedaan spesies bakteri yang digunakan dan jumlah bakteri yang disuspensikan tidak berdampak nyata terhadap kadar stakiosa tepung labu kuning, demikian pula dengan kontrol tanpa perendaman. Kadar stakiosa untuk setiap perlakuan disajikan pada Tabel 8.
Tabel 8 Pengaruh perlakuan terhadap kadar stakiosa (ppm)
Bakteri Populasi (CFU/ml) Rataan
106 107
Lactobacillus plantarum 176,70 175,63 176,16 Lactobacillus brevis 150,18 150,18 150,18 Bifidobacterium longum 153,05 149,82 151,44 Leuconostoc mesenteroides 154,48 180,65 167,56
Rataan 158,60 164,07 161,34
Kontrol dengan perendaman 193,55
Kontrol tanpa rendam 216,85
Segar 363,00
Senyawa RFO terbanyak setelah rafinosa yang terdeteksi pada buah labu kuning segar adalah stakiosa dengan jumlah sebesar 363 µg/gram bk (ppm) atau sebanyak 5,55% dari total RFO labu kuning segar. Kadar stakiosa tersebut berkurang menjadi 216,85 ppm untuk tepung labu kuning kontrol tanpa rendam dan rata-rata sebesar 161,34 ppm untuk tepung labu kuning perlakuan rendam dengan bakteri. Tepung labu kuning perlakuan bakteri B. longum dengan populasi 107 CFU/ml memberikan kadar stakiosa paling rendah yaitu 149, 82 ppm dan lebih rendah dari populasi 106 CFU/ml sebesar 153,05 ppm. Kadar stakiosa tepung labu kuning perlakuan L. brevis tidak berbeda untuk populasi 106 CFU/ml dan 107 CFU/ml yaitu 150,18 ppm (Tabel 8).
stakiosa sebesar 180,65 ppm, lebih tinggi dari populasi 106 CFU/ml sebesar 154,48 ppm. Perlakuan populasi bakteri memberikan rataan kadar stakiosa tepung labu kuning sebesar 164,07 ppm untuk populasi 107 CFU/ml lebih tinggi dibandingkan dengan populasi 106 CFU/ml sebesar 158,60 ppm. Kemungkinan pada populasi 107 CFU/ml, enzim α-galaktosidase bakteri L. plantarum dan L. mesenteroides mengalami penurunan aktivitas dan hanya mampu menyebabkan hidrolisis partial dari stakiosa yang menghasilkan komponen rafinosa dan galaktosa (Mital dan Steinkraus, 1975) akibat tercapainya nilai pH yang lebih rendah dari pH optimum aktivitas α -galaktosidase.
Tabel 9 Reduksi stakiosa (%) tepung labu kuning setelah perlakuan
Bakteri Populasi (CFU/ml) Rataan
106 107
* Reduksi total. Persentase dihitung terhadap segar.
** Nilai dalam tanda kurung adalah reduksi akibat perendaman yaitu selisih antara persentase reduksi total perlakuan perendaman dan kontrol tanpa rendam.
dengan air destilata menurunkan kadar stakiosa sebesar 6,42%. Berdasarkan persentase reduksi selama perendaman dengan suspensi bakteri, reduksi stakiosa diperkirakan dua kali lebih tinggi dibandingkan reduksi rafinosa (Tabel 9).
Penelitian reduksi stakiosa menggunakan ekstrak bebas-sel L. brevis dilakukan oleh Mital et al. (1973) yang menunjukkan bahwa L. brevis menghidrolisis stakiosa sari kedelai sebanyak 13% selama 8 jam inkubasi namun pada kondisi optimal hidrolisis stakiosa bisa mencapai 50%. Proses fermentasi sari kedelai yang dilakukan Mital dan Steinkraus (1975) menggunakan L. plantarum dan menunjukkan bahwa terjadi hidrolisis stakiosa native sebanyak 60%.
Bakteri L. mesenteroides JK55 yang digunakan Yoon dan Hwang (2008) mampu menghidrolisis habis stakiosa native sari kedelai dalam waktu 10 jam. Scalabrini et al. (1998) melaporkan bahwa B. longum MB300 yang digunakannya mampu menghidrolisis stakiosa native sari kedelai sebanyak 26% dalam waktu 24 jam dan hingga 65% dalam waktu 48 jam. Garro et al. (1999) melaporkan bahwa bakteri B. longum CRL 849 yang ditelitinya mereduksi stakiosa native sari kedelai hingga 49,3% selama 7 jam pertama fermentasi. Bifidobacterium memetabolisme empat unit sakarida lebih cepat dari L. plantarum yang membutuhkan lebih dari 30 jam fermentasi untuk mereduksi jumlah yang sama (Garro et al., 1999). Hou et al. (2000) melaporkan bahwa hasil reduksi stakiosa pada sari kedelai terjadi hingga 50,68% selama 48 jam fermentasi menggunakan B. longum B6.
c. Verbaskosa
Tabel 10 Pengaruh perlakuan terhadap reduksi verbaskosa (ppm)
Bakteri Populasi (CFU/ml) Rataan
106 107
Lactobacillus plantarum 16,49 a 7,53 a 12,01 Lactobacillus brevis 12,19 a 13,62 a 12,90 Bifidobacterium longum 8,96 a 11,47 a 10,22 Leuconostoc mesenteroides 15,05 a 8,24 a 11,64
Rataan 13,17 10,22 11,69
Kontrol dengan perendaman 34,05 b
Kontrol tanpa rendam 41,58 b menjadi rata-rata 11,69 ppm untuk perlakuan perendaman dengan bakteri. Rataan kadar verbaskosa pada perlakuan perendaman bakteri dengan populasi 107 CFU/ml sebesar 10,22 ppm, lebih rendah dibandingkan dengan populasi 106 CFU/ml sebesar 13,17 ppm.
Tepung labu kuning perlakuan bakteri L. plantarum dengan populasi 107 CFU/ml memberikan kadar verbaskosa sebesar 7,53 ppm, lebih rendah dibandingkan dengan populasi 106 CFU/ml sebesar 16,49 ppm. Perlakuan bakteri L. mesenteroides memberikan kadar verbaskosa sebesar 8,24 ppm untuk populasi 107 CFU/ml lebih rendah dibandingkan dengan populasi 106 CFU/ml sebesar 15,05 ppm. Berbeda dengan komponen RFO sebelumnya (rafinosa dan stakiosa), perlakuan bakteri L. brevis memberikan kadar verbaskosa sebesar 13,62 ppm untuk populasi 107 CFU/ml lebih tinggi dari populasi 106 CFU/ml sebesar 12,19 ppm. Hal yang sama juga diperoleh pada tepung labu kuning perlakuan bakteri B. longum yang memiliki kadar verbaskosa sebesar 11,47 ppm untuk populasi 107 CFU/ml lebih tinggi dari populasi 106 CFU/ml sebesar 8,96 ppm.
dengan air destilata sebesar 43,25% dan perlakuan perendaman dengan suspensi bakteri sebesar rata-rata 80,51%. Selama perendaman dengan suspensi bakteri, kadar verbaskosa menurun sebesar rata-rata 49,81%. Berdasarkan persentase reduksi selama perendaman dengan suspensi bakteri, reduksi verbaskosa tersebut lebih tinggi dari reduksi rafinosa dan stakiosa. Persentase reduksi verbaskosa terbesar terjadi pada perlakuan bakteri B. longum sebesar rata-rata 52,28%.
Tabel 11 Reduksi verbaskosa (%) tepung labu kuning setelah perlakuan
Bakteri Populasi (CFU/ml) Rataan
106 107
* Reduksi total. Persentase dihitung terhadap segar.
** Nilai dalam tanda kurung adalah reduksi akibat perendaman yaitu selisih antara persentase reduksi total perlakuan perendaman dan kontrol tanpa rendam.
Reduksi jumlah RFO pada tepung labu kuning kontrol tanpa perendaman menunjukkan bahwa proses pengeringan mampu mengurangi senyawa RFO dalam jumlah yang cukup besar. Senyawa RFO merupakan turunan α -(1,6)-galaktosil dari sukrosa. Sukrosa memiliki ikatan ß-fruktofuranosida yang tidak stabil dan dapat mengalami hidrolisis ketika dehidrasi berlangsung (Schebor et al., 1999). Ketiga senyawa RFO mengalami hidrolisis pada ikatan tersebut maka ketika RFO diukur, tidak lagi terdeteksi sebagai rafinosa, stakiosa, dan verbaskosa dengan jumlah yang sama dengan sebelum diukur. Kadar air yang tinggi pada irisan labu dan proses pengeringan yang lama memungkinkan terjadinya reaksi hidrolisis tersebut.
Labu kuning yang mendapat perlakuan perendaman dengan air destilata mengalami reduksi total RFO hingga sebesar 86,29%. Penurunan RFO kemungkinan disebabkan adanya aktivitas bakteri dan atau mikroba lainnya yang secara alami terdapat pada labu kuning segar dan atau pada air destilata yang digunakan. Hal ini karena labu kuning segar dan air destilata yang digunakan pada penelitian ini tidak disterilisasi terlebih dahulu. Mikroba dapat berasal dari operator/pekerja, udara, perkakas yang digunakan dan kotoran yang menempel pada kulit buah labu berpindah ke potongan labu kuning segar. Letak buah labu kuning yang dekat dan bahkan bersentuhan dengan tanah dapat menjadi penyebab utama termuatnya mikroba tanah pada permukaan kulit buah labu kuning yang kemudian selama proses pembersihan dan pengecilan ukuran dapat berpindah ke potongan labu. Beberapa BAL terdapat dalam jumlah sangat sedikit pada jaringan tanaman seperti Lactobacillus, Pediococcus, Enterococcus, dan Leuconostoc mesenteroides (Mäki, 2004). Sebagian besar dari material tanaman mengandung mikroba epifitik dalam jumlah yang banyak dan beragam, dan sejumlah kecil populasi bakteri asam laktat (Fleming dan McFeeters, 1981).
dan disakarida- daripada senyawa RFO dan jumlahnya banyak, sehingga penggunaannya menjadi lebih utama bagi bakteri untuk metabolisme dan pertumbuhannya. Perbedaan jumlah populasi bakteri yang disuspensikan ke perendam hingga satu log secara umum tidak berpengaruh nyata terhadap kadar akhir setiap senyawa RFO pada tepung labu kuning.
Pada perlakuan kontrol rendam air destilata, jumlah RFO pada labu kuning mengalami reduksi dengan kadar RFO lebih kecil dari kadar RFO kontrol tanpa perendaman. Telah diketahui sebelumnya bahwa pertumbuhan mikroba lain juga mungkin terjadi pada irisan labu yang direndam dengan air destilata yang berarti juga terjadi proses fermentasi. Penggunaan senyawa RFO oleh mikroba tersebut dapat terjadi karena enzim-enzim yang berperan dalam hidrolisis RFO bisa terinduksi pada beberapa spesies bakteri, dan dihasilkan oleh beberapa spesies kapang dan khamir.
2. Kadar β-karoten
Buah labu kuning segar pada penelitian ini memiliki kadar β-karoten sebesar 102,83 ppm (µg/g bk) atau sebesar 0,74 mg/100 g bb. Nilai tersebut lebih kecil dibandingkan kadar β-karoten berbagai varietas buah labu kuning spesies C. moschata yaitu berkisar 3,1 – 7 mg/100 g (Murkovic et al., 2002).
Kadar β-karoten tepung labu kuning yang mendapat perlakuan perendaman bakteri dan kontrol perendaman air destilata lebih rendah dari kadar β-karoten tepung labu kontrol tanpa rendam. Secara statistik, besarnya kadar β-karoten tepung labu kuning yang diperoleh tidak berbeda nyata untuk setiap perlakuan dan kontrol. Rincian kadar β-karoten tepung labu kuning dapat dilihat pada Tabel 12.
melakukan karakterisasi salah satu enzim penyebab carotenes bleaching yaitu lipoksigenase yang diisolasi dari buah labu kuning (Cucurbita moschata varietas Jepang). Enzim tersebut memiliki aktivitas optimum pada pH 6,5 dan mengalami penurunan aktivitas hingga 30% pada pH 4,0. Selama 18 jam proses perendaman, bakteri-bakteri yang digunakan pada perlakuan akan memfermentasi senyawa gula-gula (karbohidrat) yang ada di jaringan labu. Proses fermentasi tersebut menghasilkan senyawa asam organik yaitu asam laktat dan atau asam organik lainnya seperti asam asetat. Akumulasi asam-asam organik akan menyebabkan penurunan pH menjadi asam-asam (di bawah pH netral) sehingga enzim penyebab carotenes bleaching menjadi berkurang aktivitasnya. Enzim menjadi teraktivasi maksimum pada tepung labu kuning kontrol perendaman dengan air destilata sehingga kadar β-karoten lebih rendah dari tepung labu kuning perlakuan perendaman dengan bakteri.
Tabel 12 Pengaruh perlakuan terhadap kadar β-karoten (ppm)
Bakteri Populasi (CFU/ml) Rataan
106 107
Lactobacillus plantarum 236,92 228,23 232,58 Lactobacillus brevis 235,84 243,55 239,70 Bifidobacterium longum 236,20 240,50 238,35 Leuconostoc mesenteroides 240,59 245,70 243,14
Rataan 237,39 239,50 238,44
Kontrol dengan perendaman 230,65
Kontrol tanpa rendam 269,62
Hal yang sama diperoleh dari hasil pengaruh jumlah populasi bakteri pada perendam terhadap kadar β-karoten. Berdasarkan rataan kadar β-karoten, tepung labu perlakuan jumlah bakteri 107 CFU/ml memiliki β-karoten lebih tinggi dari perlakuan jumlah bakteri 106 CFU/ml (Tabel 12). Jumlah bakteri yang lebih banyak sepuluh kali lipat kemungkinan menyebabkan turunnya pH menjadi sedikit lebih asam karena jumlah asam organik yang lebih banyak.
bakteri lebih rendah dibandingkan tepung labu kuning kontrol tanpa perendaman. Beberapa penelitian menunjukkan adanya pengaruh tingkat keasaman (pH) terhadap kestabilan β-karoten. Chen et al. (1995) melaporkan bahwa jus wortel yang diasamkan hingga pH 4,0 menggunakan asam sitrat kemudian dipasteurisasi pada suhu 105°C selama 30 detik mengalami sedikit penurunan β-karoten dan adanya sedikit peningkatan isomer cis sebesar 4%. Isomerisasi cis menyebabkan perubahan warna pada sayuran karena sifat-sifat spektral karotenoid cis berbeda dari karotenoid trans (Kidmose et al., 2002). Tersisipnya satu atau lebih isomer cis pada sistem all-trans terkonjugasi menyebabkan pergeseran hipsokromik sebesar 2-5 nm. Perbedaan sifat spektral dari isomer cis menghasilkan hue (saturasi) yang lebih terang dan intensitas yang lebih rendah dibanding dengan isomer trans (Gross, 1991). Hal ini yang dapat menyebabkan kadar β-karoten yang terukur menjadi lebih rendah. Kestabilan β-karoten terhadap pH dipengaruhi oleh lamanya paparan oleh asam, konsentrasi asam, dan sistem dari keberadaan β-karoten itu sendiri.
Pada penelitian ini, pengaruh dari akumulasi asam selama perendaman terhadap β-karoten teramati secara visual. Irisan labu yang mendapat perlakuan perendaman tampak mengalami bleaching (pemucatan) ketika sekitar dua jam setelah perendaman atau saat awal proses pengeringan dalam keadaan irisan labu kuning masih basah (Gambar 8). Faktor-faktor lain yang kemungkinan dapat mempengaruhi kadar β-karoten, yaitu distribusi senyawa β-karoten pada daging buah labu itu sendiri dan keseragaman ukuran dan tingkat kematangan antar buah labu kuning yang digunakan.
3. Rendemen
Berdasarkan hasil yang diperoleh, rendemen labu kuning kering yang sebelumnya mendapat perlakuan perendaman (dengan bakteri maupun kontrol rendam air destilata) rata-rata sebesar 3,53%, lebih kecil dari rendemen tepung labu yang tidak mendapat perlakuan perendaman yaitu 5,93%. Secara umum, rendemen tidak dipengaruhi oleh perbedaan spesies bakteri dan jumlah populasinya tetapi perlakuan perendaman (baik dengan bakteri maupun kontrol rendam air destilata) berpengaruh nyata (P≤5%) terhadap penurunan rendemen pengeringan irisan labu kuning. Besarnya rendemen untuk setiap perlakuan dapat dilihat pada Tabel 13.
Tabel 13 Pengaruh perlakuan terhadap rendemen pengeringan (%)1
Bakteri Populasi (CFU/ml) Rataan
106 107
Rendemen dihitung berdasarkan prosentase rasio irisan labu kering terhadap ±200 g irisan labu basah untuk setiap perlakuan.
a-c
Nilai dengan superskrip huruf yang tidak sama berarti ada beda nyata pada taraf 5% berdasarkan test Duncan.
seperti L. mesenteroides (Juven et al., 1984), L. plantarum (Sakellaris, 1988; Kalam dan Belarbi, 1995), L. casei dan L. lactis (Kalam dan Belarbi, 1995).
Penguraian jaringan yang terjadi akibat fermentasi oleh mikroba kemungkinan menyebabkan air perendam mudah terpenetrasi ke dalam jaringan irisan labu kuning sehingga irisan labu kuning menjadi berair. Ketika proses pengeringan berlangsung, proses dehidrasi yang terjadi pada irisan labu kuning yang direndam mungkin berbeda dengan dehidrasi pada irisan labu kuning kontrol yang tidak direndam. Dehidrasi yang terjadi pada irisan labu kuning yang direndam kemungkinan berlangsung lebih optimal. Air yang terjebak pada jaringan buah labu kuning menjadi lebih mudah keluar dan lebih banyak dipindahkan.
Perbedaan dehidrasi yang terjadi pada labu kuning yang direndam dan yang tidak direndam terlihat dari perbedaan kenampakan kedua irisan labu yang telah kering. Irisan labu kering yang sebelumnya direndam terlihat lebih tipis dibandingkan irisan labu kering yang tidak direndam padahal ketebalan irisan ketika masih segar adalah relatif sama yaitu ±3 mm. Kemudian, irisan labu kering tanpa perendaman terlihat mengeriput sedangkan irisan labu kering dengan perendaman terlihat melebar dan memiliki bentuk yang hampir sama ketika masih basah namun ukuran mengecil dan menipis tetapi tidak mengkisut.
V. SIMPULAN DAN SARAN
A. SIMPULAN
Buah labu kuning segar mengandung rafinosa (612,0 mg/100 g bk), stakiosa (36,3 mg/100 g bk), dan verbaskosa (6,0 mg/100 g bk). Pemberian perlakuan perendaman dalam suspensi bakteri asam laktat dan perbedaan jumlah populasinya menghasilkan reduksi RFO yang relatif sama. Kadar RFO tepung labu kuning terendah adalah perlakuan perendaman dengan bakteri B. longum dengan populasi 107 CFU/ml sebesar 550,54 ppm atau turun sebesar 8,75% selama perendaman. Perlakuan perendaman dengan bakteri B. longum dengan populasi 107 CFU/ml juga menghasilkan kadar rafinosa dan stakiosa terendah masing-masing sebesar 389,25 ppm (turun 7,77%) dan 149,82 ppm (turun 18,47%). Kadar verbaskosa terendah diperoleh tepung labu kuning perlakuan perendaman dengan bakteri L. plantarum dengan populasi 107 CFU/ml sebesar 7,53 ppm atau turun sebesar 56,75% selama perendaman.
Kadar β-karoten buah labu kuning segar adalah 0,74 mg/100 g bb. Kadar akhir β-karoten tepung labu kuning relatif sama antara tepung labu perlakuan dan kontrol. Berdasarkan besaran nilai, kadar β-karoten tepung labu kontrol tanpa rendam adalah 269,62 ppm, perendaman dengan air destilata adalah 230,65 ppm, dan perendaman dengan bakteri rata-rata adalah 238,44 ppm. Perlakuan populasi bakteri 107 CFU/ml menghasilkan kadar β-karoten 239,50 ppm lebih tinggi dari perlakuan populasi 106 CFU/ml sebesar 237,39 ppm.
Perlakuan perendaman menyebabkan penurunan rendemen irisan labu kering tetapi perbedaan perlakuan spesies bakteri dan jumlah populasinya menghasilkan rendemen yang relatif sama. Perlakuan perendaman dengan bakteri asam laktat menghasilkan rendemen rata-rata sebesar 3,52%, kontrol perendaman dengan air destilata sebesar 3,57%, dan kontrol tanpa rendam sebesar 5,93%.
B. SARAN
Penelitian dengan topik yang sama dapat dilakukan kembali dengan mengurangi lama waktu fermentasi, penggunaan jenis bakteri asam laktat lain secara tunggal atau kombinasi, dan pemberian pra-perlakuan untuk menekan