PENDUGAAN PARAMETER GENETIK KETAHANAN
TERHADAP PENYAKIT ANTRAKNOSA DAN BEBERAPA
KARAKTER KUANTITATIF PADA CABAI
SYAIDATUL ROSIDAH
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Pendugaan Parameter Genetik Ketahanan terhadap Penyakit Antraknosa dan Beberapa Karakter Kuantitatif pada Cabai adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
SYAIDATUL ROSIDAH. Pendugaan Parameter Genetik Ketahanan terhadap Penyakit Antraknosa dan Beberapa Karakter Kuantitatif pada Cabai. Dibimbing oleh MUHAMAD SYUKUR.
Penelitian dilaksanakan di Kebun Percobaan Leuwikopo dan Laboratorium Pemuliaan Tanaman IPB pada bulan Oktober 2012 sampai April 2013. Populasi hasil persilangan genotipe C15 dan C2 digunakan untuk mempelajari parameter genetik ketahanan terhadap penyakit antraknosa yang disebabkan oleh Colletotrichum acutatum dan beberapa karakter kuantitatif pada cabai. Sebanyak dua puluh buah matang hijau dari setiap tanaman diinokulasi dengan isolat PYK 04. Hasil penelitian menunjukkan bahwa ketahanan cabai terhadap antraknosa dikendalikan oleh gen resesif. Tidak ada efek maternal dalam penurunan karakter yang diamati kecuali karakter panjang buah. Heritabilitas arti luas tergolong tinggi untuk karakter kejadian penyakit, diameter nekrosis, tinggi tanaman, diameter batang, tinggi dikotomus, umur berbunga, dan umur panen; dan tergolong sedang untuk karakter panjang dan diameter buah. Heritabilitas arti sempit tergolong tinggi untuk karakter diameter batang, tinggi dikotomus, dan umur berbunga; sedang untuk karakter kejadian penyakit, tinggi tanaman, dan umur panen; dan rendah untuk karakter diameter nekrosis, panjang buah dan diameter buah. Rasio ragam aditif bernilai tinggi untuk karakter kejadian penyakit, tinggi tanaman, diameter batang, tinggi dikotomus, dan umur berbunga; sedang untuk karakter diameter nekrosis, umur panen, dan diameter buah; dan rendah untuk karakter panjang buah. Pembentukan varietas cabai tahan antraknosa dan berdaya hasil baik sebaiknya diarahkan pada varietas open pollinated (bersari bebas).
Kata kunci: cabai, Colletotrichum acutatum, heritabilitas, heterosis, heterobeltiosis
ABSTRACT
SYAIDATUL ROSIDAH. Estimation of Resistance Genetic Parameter for Anthracnose Disease and Quantitative Characters of Pepper. Supervised by MUHAMAD SYUKUR.
medium for fruit length and diameter character. Narrow-sence heritability were high for stem diameter, dichotomous height, days for flowering; medium for disease incidence, plant height, days for harvesting; and low for necrotic diameter, fuit length and diameter character. Additive-variance ratio were high for disease incidence, plant height, stem diameter, dichotomous height, and days for flowering; medium for necrotic diameter, days for harvesting, fruit diameter; and low for fruit length character. Producing pepper varieties which are resistance to anthracnose and good yield should be aimed to open pollinated varieties.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Agronomi dan Hortikultura
PENDUGAAN PARAMETER GENETIK KETAHANAN
TERHADAP PENYAKIT ANTRAKNOSA DAN BEBERAPA
KARAKTER KUANTITATIF PADA CABAI
SYAIDATUL ROSIDAH
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
Judul Skripsi : Pendugaan Parameter Genetik Ketahanan terhadap Penyakit Antraknosa dan Beberapa Karakter Kuantitatif pada Cabai Nama : Syaidatul Rosidah
NIM : A24090176
Disetujui oleh
Dr Muhamad Syukur, SP MSi Pembimbing
Diketahui oleh
Dr Ir Agus Purwito, MScAgr Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga penelitian ini dapat terselesaikan. Tema penelitian yang dilaksanakan pada bulan Oktober 2012 sampai April 2013 ini adalah ketahanan cabai terhadap penyakit penting, dengan judul Pendugaan Parameter Genetik Ketahanan terhadap Penyakit Antraknosa dan Beberapa Karakter Kuantitatif pada Cabai.
Terima kasih penulis ucapkan kepada:
1. Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor yang berkenan menerima penulis menjadi mahasiswa 2. Dr Muhamad Syukur, SP MSi selaku pembimbing skripsi dan Abdul
Hakim, SP selaku laboran Laboratorium Pendidikan Pemuliaan Tanaman, IPB yang telah banyak memberi saran selama penelitian
3. Prof Dr Ir HMH Bintoro MAgr selaku pembimbing akademik yang telah memberikan arahan, semangat, dan kasih sayangnya selama penulis belajar di IPB
4. Ibu Siti Kalimah, Ayah Sukardi (wali), dan Bapak Ahmad Faozi atas doa, dorongan, dan kasih sayangnya
5. Seluruh dosen Departemen Agronomi dan Hortikultura yang telah memberikan pembelajaran selama penulis kuliah
6. Direktorat Jenderal Pendidikan Tinggi, Himpunan Keluarga Rembang Bogor (HKRB), Lembaga Amil Zakat Alhurriyyah, Karya Salemba Empat, Bakti BCA, dan Ikatan Alumni Korea-Indonesia atas bantuan dan beasiswa yang diberikan kepada penulis
7. Seluruh teman-teman atas saran dan doanya Semoga hasil penelitian ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Hipotesis Penelitian 2
TINJAUAN PUSTAKA 2
Serangan Colletotrichum acutatum terhadap Capsicum annuum L. 2 Ketahanan Capsicum annuum L. terhadap Colletotrichum acutatum 3 Pemuliaan Capsicum annuum L. Resisten Colletotrichum acutatum 4
METODE 5
Tempat dan Waktu 5
Bahan dan Alat 5
Metode Pelaksanaan 5
Analisis Data 6
HASIL DAN PEMBAHASAN 7
Karakter Ketahanan Cabai terhadap Antraknosa 8
Karakter Vegetatif Cabai 12
Karakter Generatif Cabai 15
SIMPULAN DAN SARAN 18
Simpulan 18
Saran 18
UCAPAN TERIMAKASIH 18
DAFTAR PUSTAKA 18
RIWAYAT HIDUP 23
DAFTAR TABEL
1 Jumlah tanaman pada setiap populasi berdasarkan skor ketahanan
terhadap antraknosa 8
2 Rata-rata, ragam, dan simpangan baku diameter nekrosis setiap populasi 10 3 Nilai rata-rata, galat baku, dan uji nilai tengah kejadian penyakit dan
diameter nekrosis F1 dan F1R 10
4 Komponen ragam kejadian penyakit dan diameter nekrosis pada cabai
yang disebabkan oleh C. acutatum 10
5 Heterosis dan heterobeltiosis kejadian penyakit dan diameter nekrosis
antraknosa pada cabai 11
7 Nilai rata-rata, ragam baku, dan uji nilai tengah tinggi tanaman, diameter
batang, dan tinggi dikotomus F1 dan F1R 13
8 Komponen ragam tinggi tanaman, diameter batang, dan tinggi
dikotomus pada cabai 13
9 Heterosis dan heterobeltiosis tinggi tanaman, diameter batang, dan tinggi
dikotomus pada cabai 14
10 Nilai rata-rata, ragam, dan ragam baku karakter umur berbunga, umur
panen, panjang buah, dan diameter buah setiap populasi cabai 15 11 Nilai rata-rata, ragam baku, dan uji nilai tengah umur berbunga, umur
panen, panjang buah, dan diameter buah F1 dan F1R 16 12 Komponen ragam umur berbunga, umur panen, panjang buah, dan
diameter buah pada cabai 16
13 Heterosis dan heterobeltiosis umur berbunga, umur panen, panjang buah,
dan diameter buah pada cabai 17
DAFTAR GAMBAR
1 Panjang diameter nekrosis 9
PENDAHULUAN
Latar Belakang
Cabai merupakan komoditas hortikultura yang strategis. Permintaan terhadap cabai cenderung tetap sehingga produksi cabai harus dapat dilakukan dengan berkesinambungan. Produksi cabai ditentukan oleh luas areal pertanaman dan produktivitas. Luas areal pertanaman cabai tahun 2012 mencapai 120 094 ha. Namun, luasnya areal pertanaman tidak didukung dengan nilai produktivitas cabai yang tinggi. Produktivitas cabai nasional tahun 2012 hanya mencapai 7.4 ton ha-1 (BPS 2013). Kondisi ini masih jauh dari produktivitas potensial cabai yang mampu mencapai 20-30 ton ha-1 (Syukur et al. 2010).
Tingkat produktivitas cabai dipengaruhi oleh gangguan hama dan penyakit. Salah satu penyakit yang cukup besar pengaruhnya terhadap produktivitas cabai adalah antraknosa (Humaedah 2012). Antraknosa dapat menyebabkan kehilangan hasil cabai sampai 50% (Pakdeevaraporn et al. 2005). Efri (2010) menyatakan bahwa serangan antraknosa pada batang dan daun tidak menimbulkan masalah yang berarti bagi tanaman, tetapi dari bagian inilah penyakit dapat berkembang ke buah dan dapat menimbulkan kerusakan yang sangat serius. Akibatnya produktivitas cabai yang dapat dipanen menurun.
Antraknosa disebabkan oleh cendawan dari genus Colletotrichum. Beberapa spesies antraknosa telah teridentifikasi di Indonesia antara lain C. acutatum, C. gloeosporioides, dan C. capsici. Spesies C. acutatum adalah jenis yang pertama dilaporkan dan paling dominan di Indonesia (AVRDC 2009). Suatu penelitian menyatakan bahwa C acutatum lebih virulen dibanding C. gloeosporioides dan C. capsici (Mongkolporn et al. 2010). Umumnya, varietas cabai yang ada rentan terhadap antraknosa. Pengujian yang dilakukan pada lima kultivar nasional dan sepuluh kultivar koleksi IPB menunjukkan bahwa semua kultivar cabai tersebut rentan terhadap antraknosa (C. acutatum) (Marliyanti et al. 2013). Kim et al. (2010) melakukan pengujian terhadap 209 aksesi cabai dan 173 aksesi terinfeksi C. acutatum. Kim et al. (2012) menemukan bahwa dari 869 kultivar cabai yang diuji, 847 kultivar dinyatakan rentan terhadap C. acutatum.
Penggunaan pestisida telah banyak dilakukan untuk mengendalikan penyakit antraknosa pada cabai. Aplikasi pestisida yang berlebihan tidak hanya meningkatkan biaya produksi, tetapi juga berbahaya bagi petani dan konsumen. Oleh karena itu, perlu dilakukan upaya pengendalian lain yang bersifat ramah lingkungan. Salah satu hal yang dilakukan yaitu pembentukan kultivar baru dengan cara menyilangkan tanaman cabai resisten antraknosa sebagai donor dengan cabai lainnya (Prasath et al. 2007; Kaur et al. 2011).
2
2007). Dengan demikian perlu adanya studi pendugaan parameter genetik ketahanan terhadap penyakit antraknosa dan beberapa karakter kuantitatif pada cabai.
Tujuan Penelitian
Penelitian ini bertujuan menduga parameter genetik ketahanan terhadap penyakit antraknosa yang disebabkan oleh Colletotrichum acutatum dan beberapa karakter kuantitatif pada cabai.
Hipotesis Penelitian
Hipotesis yang akan dibuktikan dalam penelitian ini adalah:
1. Heritabilitas arti sempit dan arti luas pada cabai tergolong tinggi untuk karakter ketahanan terhadap penyakit antraknosa dan karakter kuantitatif. 2. Tidak ada efek maternal untuk karakter ketahanan terhadap penyakit
antraknosa dan karakter kuantitatif.
3. Ragam aditif lebih tinggi daripada ragam non aditif untuk karakter ketahanan terhadap penyakit antraknosa dan karakter kuantitatif.
4. Heterosis tergolong rendah untuk karakter ketahanan terhadap penyakit antraknosa dan karakter kuantitatif.
TINJAUAN PUSTAKA
Serangan Colletotrichum acutatum terhadap Capsicum annuum L.
Penyakit antraknosa dapat dilihat pada tiga fase, yaitu: fase pembibitan atau damping off, fase bercak daun, dan fase buah. Serangan antraknosa pada fase buah menyebabkan berkurangnya bobot kering buah dan menurunnya kandungan capcaisin serta oleoresin (Mistry et al. 2008). Capcaisin ini dikenal sebagai senyawa yang menimbulkan rasa pedas pada cabai dan diketahui mampu menghambat pertumbuhan berbagai organisme pengganggu tanaman salah satunya cendawan antraknosa (Kraikruan et al. 2008).
3 Antraknosa merupakan penyakit yang disebabkan oleh cendawan dari genus Colletotrichum. Beberapa spesies telah teridentifikasi yaitu C. gloeosporioides, C. acutatum, C. dematium, C. capsici, and C. coccodes (Park 2005). Pada tahun 2010, Golzar dan Wang melaporkan adanya spesies baru yaitu C. phormii yang menyerang tanaman Phormium tenax di Australia. Syukur et al. (2009) dan Stankova et al. (2011) menyatakan bahwa tingkat serangan antraknosa tergantung dari jenis tanaman inang dan spesies antraknosa yang menyerang. Hasil penelitian yang dilakukan oleh Wharton dan Uribeondo (2004) dan Mongkolporn et al. (2010) menunjukkan bahwa dari spesies yang diuji, C. acutatum merupakan spesies yang memiliki virulensi paling besar terhadap cabai.
Cendawan C. acutatum merupakan spesies cendawan penyebab antraknosa yang memiliki ciri warna kultur abu-abu sampai pink, tidak mempunyai fase seksual, tumbuh optimal pada suhu 24-26 0C, rata-rata tumbuh pada media PDA (suhu optimal) adalah 7.7 (4.9-9.9) mm hari-1, dan menginfeksi pada buah yang belum matang (Freeman et al. 1998). C. acutatum menghasilkan koloni oranye pucat dengan sedikit miselium aerial dan beberapa konidia berwarna oranye mengelilingi pusatnya (Than et al. 2008a).
Ketahanan Capsicum annuum L. terhadap Colletotrichum acutatum
Pengaruh antraknosa yang cukup besar dalam menurunkan produksi cabai (Kim et al. 2010; Lee et al. 2010) menjadi suatu hal yang penting untuk diperhatikan. Banyak penelitian yang telah dilakukan untuk mengetahui tentang ciri morfologi (Freeman dan Katan 1997), molekular (Roca et al. 2003), dan fisiologi (Oanh et al. 2004; Dieguez-Uribeondo et al. 2005) cendawan penyebab antraknosa. Selain itu juga telah dilakukan identifikasi mengenai gejala timbulnya penyakit antraknosa (Sergeeva et al. 2008) dan metode pengendaliannya.
Penelitian yang mempelajari hubungan senyawa kimia di dalam tanaman cabai dengan ketahanannya terhadap antraknosa telah dilakukan. Dilaporkan bahwa vilurensi C. acutatum dipengaruhi oleh konsentrasi ABA. Pemberian ABA dapat meningkatkan diameter nekrosis dan kejadian penyakit pada cabai (Hwang et al. 2008). Ada pula yang menyatakan bahwa ketahanan cabai terhadap antraknosa dipengaruhi oleh kandungan capsaicin. Cabai dengan kandungan capsaicin rendah mudah terserang antraknosa (Kaur et al. 2011) meskipun keterangan selanjutnya menyatakan bahwa kandungan capsaicin tidak berkorelasi dan tidak dapat dijadikan sebagai penanda ketahanan terhadap antraknosa (Syukur et al. 2009).
Senyawa lain yang diduga berkaitan dengan ketahanan terhadap antraknosa adalah enzim peroksidase. Peningkatan aktivitas enzim peroksidase akan meningkatkan produk toksin bagi patogen, sehingga menghasilkan tingkat ketahanan yang lebih tinggi terhadap infeksi (Agrios 1997). Penelitian Zen et al. (2002) selanjutnya menjelaskan bahwa aktivitas enzim peroksidase yang tinggi tidak berkaitan dengan intensitas penyakit antraknosa yang rendah karena perbedaan aktivitas enzim ini timbul setelah terjadi infeksi penyakit.
4
serangan antraknosa (Marvel 2003; Ivey et al. 2004). Sejalan dengan berkembangnya isu lingkungan, penggunaan biofungisida mulai dilakukan. Agen yang dapat digunakan sebagai biofungisida di antaranya Trichoderma harzianum (Warin et al. 2009) dan mengkudu (Morinda citrifolia) (Efri 2010) yang terbukti dapat menghambat serangan antraknosa. Pengendalian dari dalam dilakukan dengan melakukan rekayasa susunan genetik memalui program pemuliaan tanaman.
Pemuliaan Capsicum annuum L. Resisten Colletotricum acutatum
Pengendalian antraknosa dapat dilakukan melalui kombinasi metode yaitu pengendalian secara kimia dengan pestisida, biologi dengan agen hayati, dan molekular dengan membentuk kultivar resisten (Wharton dan Uribeondo 2004). Penggunaan kultivar resisten adalah pengendalian yang paling murah, mudah, aman, dan efektif. Pengendalian ini tidak hanya menghilangkan serangan penyakit tetapi juga mengurangi biaya pengendalian secara kimia dan mekanik, sehingga mengurangi kontaminasi senyawa kimia terhadap lingkungan (Than et al. 2008b).
Pembentukan kultivar baru dapat dilakukan melalui seleksi, hibridisasi, mutasi, atau introduksi. Hibridisasi lebih baik digunakan dalam perakitan kultivar resisten antraknosa karena umumnya kultivar yang resisten mempunyai bentuk buah yang kurang bagus dan tidak disukai konsumen. Hal yang perlu dipersiapkan sebelum dilakukan hibridisasi adalah tetua resisten antraknosa (sebagai donor) dan tetua dengan sifat lain yang diinginkan. Seleksi tetua resisten antraknosa dapat dilakukan di laboratorium melalui inokulasi cendawan ke dalam buah. Inokulasi dapat dilakukan dengan metode celup atau suntik. Syukur (2007) menyatakan bahwa metode celup digunakan untuk mengindikasikan mekanisme ketahanan cabai secara fisik sedangkan metode suntik secara biokimia . Menurut Yoon dan Park (2001) dan Park (2005) metode screening (seleksi) yang baik adalah metode suntik dengan kerapatan konidia minimal 1x105 konidia ml-1.
5
METODE
Tempat dan Waktu
Penanaman cabai dilakukan di Kebun Percobaan Leuwikopo, Institut Pertanian Bogor (IPB). Kegiatan pemurnian, perbanyakan, pemeliharaan, inokulasi cendawan, dan pengamatan terhadap gejala penyakit antraknosa dilaksanakan di Laboratorium Pendidikan Pemuliaan Tanaman Departemen Agronomi dan Hortikultura, IPB. Penelitian dilakukan pada bulan Oktober 2012 sampai April 2013.
Bahan dan Alat
Bahan tanaman yang digunakan adalah cabai genotipe IPB C-15 sebagai tetua tahan (P1), IPB C-2 sebagai tetua rentan (P2), F1(IPB C-15 X 1PB C-2), F1 Resiprokal (IPB 2 X IPB 15), BCP1(FI X 1PB 15), BCP2 (F1 X 1PB C-2), dan F2. Bahan lain yang digunakan yaitu tray, platik polietilen, tisu, plastik, spidol, dan label. Pupuk yang digunakan adalah NPK mutiara, gandasil D, gandasil B, urea, SP-36, dan KCl. Pestisida yang digunakan adalah furadan 3G, Curacron 500EC, Dithane M-45, dan Antracol. Isolat yang digunakan adalah biakan murni C. acutatum PYK 04. Peralatan yang digunakan adalah alat-alat budidaya tanaman, timbangan, meteran, jangka sorong, gunting, alat suntik, kawat, ember, dan haemocytometer.
Metode Pelaksanaan
Set populasi cabai ditanam sebanyak 13 tanaman F1, 20 tanaman P1, P2, F1R, 50 tanaman BCP1, BCP2, dan 126 tanaman F2. Penanaman dilakukan 2 bulan setelah semai (BSS). Karakter kuantitatif diamati pada semua tanaman cabai. Karakter kuantitatif yang diamati meliputi: (1) umur berbunga (HSS), diamati jumlah hari mulai dari semai sampai tanaman telah berbunga; (2) umur panen (HSS), diamati jumlah hari mulai dari semai sampai buah siap dipanen (kematangan 70%); (3) tinggi tanaman (cm), diukur dari pangkal batang sampai pucuk setelah panen pertama; (4) diameter batang (mm), diukur 5 cm dari permukaan tanah, setelah panen pertama; (5) tinggi dikotomus (cm), diukur dari pangkal batang sampai cabang dikotomus, setelah panen kedua; (6) panjang buah (cm), diukur dari pangkal hingga ujung buah dari 3 buah segar, setelah panen kedua; dan (6) diameter buah (mm), diukur pada bagian pangkal dari 3 buah segar, setelah panen kedua.
Bahan pengujian penyakit adalah buah cabai yang dipanen pada saat buah sudah tua tapi masih hijau. Setiap tanaman diambil sepuluh buah untuk dilakukan inokulasi cendawan. Pengujian penyakit dilakukan sebanyak dua ulangan. Media perbanyakan inokulum adalah potato dextrose agar (PDA). Kepadatan inokulum diatur mencapai 5x105 konidia ml-1 dengan haemacytometer.
6
berbeda. Buah ditempatkan di atas kawat dalam bak plastik. Untuk menjaga kelembaban, pada dasar bak plastik diletakkan tisu yang telah disemprot dengan air. Kemudian bak ditutup dengan plastik polietilen dan diinkubasi pada suhu 250C selama 7 hari.
Pengamatan karakter ketahanan terhadap antraknosa meliputi kejadian penyakit yang diamati pada hari kelima dan diameter nekrosis pada hari ketujuh setelah inokulasi. Kejadian penyakit (KP) dihitung menggunakan rumus: KP = (n/N) x 100%; dengan KP = kejadian penyakit, n = jumlah buah yang terserang, dan N = jumlah buah total. Skor dan kriteria ketahanan terhadap penyakit antraknosa berdasarkan kejadian penyakit diduga dengan metode Yoon (2003) dimodifikasi Syukur et al. (2007), yaitu sebagai berikut: sangat tahan: 0-10% terserang; tahan: 11-20% terserang; moderat: 21-40% terserang; rentan: 41-70% terserang; sangat rentan: lebih dari 70% terserang.
Analisis Data
1. Pendugaan nilai heritabilitas
Heritabilitas arti luas dihitung berdasarkan rumus Allard (1960):
Heritabilitas arti sempit dihitung berdasarkan rumus Warner (1952):
Keterangan:
h2bs
=
heritabilitas arti luas h2ns = heritabilitas arti sempit VF1 = ragam populasi F1 VP1 = ragam populasi P1 VP2 = ragam populasi P2VBC1= ragam populasi backcross dengan tetua C-15 VBC2= ragam populasi backcross dengan tetua C-2 VF2 = ragam populasi F2
Nilai duga heritabilitas digolongkan rendah jika h2bs< 0.2, sedang jika 0.2 ≤ h2bs ≤ 0.5, dan tinggi jika h2bs> 0.5 (Halloran et al, 1979). Ragam dihitung berdasarkan rumus perhitungan ragam populasi.
2. Pendugaan efek maternal
Efek maternal diduga dengan uji t untuk membandingkan nilai tengah F1 dan F1R. Rumus uji t yang digunakan adalah:
7 S2F1, S2F1R = ragam populasi F1, F1R
nF1, nF1R = jumlah individu dalam populasi F1, F1R
Jika nilai thitung > ttabel (0.025, n-1), nilai tengah kedua populasi berbeda nyata pada taraf 5%.
3. Rasio ragam aditif (a)
Rasio ragam aditif dihitung untuk mengetahui perbandingan agam aditif dengan ragam non aditif. Rasio ragaqm aditif diduga dengan rumus:
a = (h2ns/h2bs) x 100%,
dengan a: Rasio heritabilitas arti sempit dan arti luas, h2ns: heritbilitas arti sempit, dan h2bs: heritabilitas arti luas.
4. Heterosis
a. Heterosis tetua tengah
Heterosis tetua tengah adalah peningkatan atau penurunan penampilan F1 dibandingkan dengan nilai tengah kedua tetua. Rumus heterosis tetua tengah adalah:
Heterosis =
dengan XF1 adalah nilai rata-rata F1 dan MP adalah nilai tengah rata-rata kedua tetua.
b. Heterobeltiosis
Heterobeltiosis adalah peningkatan atau penurunan penampilah F1 dibandingkan dengan tetua terbaik. Rumus heterobeltiosis adalah:
Heterobeltiosis =
dengan XF1 adalah nilai rata-rata F1 dan HP adalah nilai tengah rata-rata tetua terbaik.
HASIL DAN PEMBAHASAN
Kondisi Umum
Penelitian dilakukan tanpa menggunakan perancangan lay out percobaan. Tanaman cabai yang digunakan merupakan populasi bersegegrasi yang setiap tanamannya mewakili genotipe tersendiri. Penanaman awal populasi cabai adalah 20 tanaman tetua (P1, P2), dan F1R; 13 tanaman F1; 80 tanaman BCP1 dan BCP2; serta 160 tanaman F2. Populasi tetua, F1, dan F1R ditanam lebih sedikit dibanding populasi lainnya karena populasi ini merupakan genotipe yang memiliki homozigositas yang tinggi. Populasi F1 hanya ditanam 13 tanaman karena dari 144 benih yang disemai hanya 13 bibit yang mampu tumbuh hingga waktu penanaman di lapang.
Selama masa pertumbuhan vegetatif, penyakit layu bakteri, penyakit kuning yang disebabkan oleh Gemini virus, serta hama tungau menyerang
F1-MP
MP X 100%,
F1-HP
8
pertanaman cabai. Serangan layu bakteri mengakibatkan banyak tanaman cabai yang mati. Untuk mengendalikan penyakit tersebut, diaplikasikan kapur pertanian.
Menjelang waktu pengamatan karakter kuantitatif dan pengujian penyakit jumlah tanaman yang dapat diamati adalah 10 tanaman P1, P2, dan F1R; 9 tanaman F1; 50 tanaman BCP1 dan BCP2; dan 126 tanaman F2. Pengujian ketahanan terhadap antraknosa dilakukan di laboratorium. Cabai yang masih hijau digunakan sebagai bahan pengujian karena antraknosa yang disebabkan oleh Colletotrichum acutatum mulai menyerang buah dari fase hijau sampai pasca panen.
Pengujian penyakit antraknosa dilakukan dengan metode suntik. Cabai yang sudah diinokulasi oleh konidia antraknosa kemudian diinkubasi. Selama inkubasi, kelembaban lingkungan mikro harus dijaga untuk memberikan kondisi optimal pertumbuhan antraknosa. Hal tersebut dilakukan dengan meletakkan tisu basah di dasar bak penyimpanan buah cabai yang telah diinokulasi dan menutup bak dengan plastik polietilen. Selain itu, selama inkubasi cabai diletakkan dalam kondisi gelap untuk memicu pertumbuhan cendawan.
Karakter Ketahanan Cabai terhadap Antraknosa
Antraknosa merupakan penyakit penting yang menyebabkan penurunan produksi yang serius pada cabai (Lee et al. 2010). Pengujian ketahanan cabai terhadap antraknosa dapat dilakukan dengan metode microinjection yang selanjutnya diamati kejadian penyakit dan diameter nekrosisnya.
Tabel 1 Jumlah tanaman pada setiap populasi berdasarkan skor ketahanan
Rata-rata 2.50 5.00 4.40 4.80 4.10 4.42 4.40
Ragam (σ2
) 93.33 91.39 168.75 132.22 317.11 294.74 369.1
Simpangan baku (σ) 9.66 9.56 12.99 11.49 17.81 17.17 19.21
a
P1: IPB C15; P2: IPB C2; F1: IPB C15 X IPB C2; F1R: IPB C2 X IPB C15; BCP1: F1X IPB C15; BCP2: F1 X IPB C2; F2: selfing F1
9 hibrida resisten terhadap antraknosa dari gen resesif membutuhkan waktu dan usaha lebih (Kim et al. 2007) sehingga pembentukan varietas cabai tahan antraknosa diarahkan pada varietas bersari bebas (open pollinated). Seleksi untuk membentuk varietas tersebut dapat dilakukan pada generasi lanjut.
Kim et al. (2008a) menemukan bahwa ketahanan C. baccatum terhadap antraknosa (C. acutatum) dikendalikan oleh gen dominan tunggal sedangkan menurut Mahasuk et al. (2009) gen tersebut adalah resesif tunggal pada buah matang hijau dan dominan tunggal pada buah matang merah. Hasil penelitian Pakdeevaraporn et al. (2005) dan Kim et al. (2008b) menyatakan bahwa ketahanan C. annuum terhadap C. capsici dikendalikan oleh gen resesif. Hal ini menunjukkan bahwa gen ketahanan terhadap antraknosa berbeda tergantung spesies cabai dan isolat antraknosanya.
P1 dan P2 merupakan genotipe yang mempunyai homozigositas yang tinggi. Hal ini ditunjukkan dengan ragam keduanya yang rendah (Tabel 1). F1 merupakan turunan pertama dari hasil persilangan P1 dengan P2 (P1XP2) sedangkan F1R merupakan turunan pertama dari hasil persilangan P2 dengan P1 (P2XP1). Gen-gen dalam F1 dan F1R merupakan gen-gen yang bersifat heterozigot yang terbentuk dari gabungan gen-gen homozigot P1 dan P2, sehingga secara genetik F1 dan F1R bersifat homogen. Ragam kejadian penyakit F1 dan F1R (168.75 dan 132.22) lebih tinggi dari pada ragam kedua tetuanya. Keragaman ini lebih dipengaruhi oleh lingkungan. Ragam BCP1 dan BCP2 lebih besar dari ragam P1, P2, F1, dan F1R. Ragam BCP1 dan BCP2 dipengaruhi oleh ragam genetik dan lingkungan. Keragaman genetik keduanya terbentuk dari persilangan F1 (homogen heterozigot) dengan tetuanya (homogen homozigot) menghasilkan progeni yang sebagian bersifat homozigot dan sebagian lagi bersifat heterozigot. Populasi F2 mempunyai ragam yang paling tinggi (369.1). Keragaman populasi F2 dipengaruhi oleh keragaman genetik, lingkungan, dan interaksi genetik dengan lingkungan. F2 dihasilkan dari selfing populasi F1. Gen-gen F1 yang mengalami selfing bersegregasi sehingga diperoleh proGen-geni dengan kombinasi gen yang bervariasi. Sebagian progeni memiliki gen yang mengarah kepada P1, sebagian mengarah kepada P2, dan sebagian mengarah kepada F1.
Karakter ketahanan cabai terhadap antraknosa yang diamati selanjutnya adalah diameter nekrosis. Diamater nekrosis ini diukur pada jarak terpanjang penyebaran penyakit antraknosa (Gambar 1). Hasil pengukuran diameter nekrosis menunjukkan bahwa P1 memiliki diameter nekrosis yang jauh lebih rendah dari pada P2. Berkaitan dengan kriteria ketahanan terhadap antraknosa bahwa P1 yang bersifat moderat memiliki diameter nekrosis yang rendah dan P2 yang bersifat sangat rentan memiliki diameter nekrosis yang lebih tinggi, begitu pula dengan F1, F1R, BCP1, BCP2, dan F2 yang mengarah ke rentan sampai sangat rentan memiliki diameter nekrosis yang lebih tinggi dari P1. Dapat dikatakan bahwa semakin rentan suatu populasi, diameter nekrosisnya akan
10
semakin tinggi. Hal ini karena cabai yang rentan akan memudahkan cendawan C. acutatum berkembangbiak dan terus memperluas jangkauan serangannya.
Nilai ragam P1, P2, F1, dan F1R relatif lebih rendah diikuti oleh ragam BCP1, BCP2, dan F2 yang lebih tinggi. Nilai ragam ini sesuai dengan komposisi genetik P1, P2, F1, dan F1R yang homogen dan BCP1, BCP2, F2 yang heterogen. Ragam F2 mempunyai nilai tertinggi karena kontitusi genetik yang menyebar dari P1 sampai mengarah ke P2 (Tabel 2).
Tabel 2 Rata-rata, ragam, dan simpangan baku diameter nekrosis setiap populasi
Parameter Populasi
a
P1 P2 F1 F1R BCP1 BCP2 F2
Rata-rata diameter nekrosis
(mm) 10.66 22.75 21.79 18.20 16.18 19.09 17.48
Ragam (σ2
) 23.39 34.23 31.17 40.42 62.74 62.38 67.76
Simpangan baku (σ) 4.77 5.85 5.58 6.36 7.92 7.89 8.23
a
P1: IPB C15; P2: IPB C2; F1: IPB C15 X IPB C2; F1R: IPB C2 X IPB C15; BCP1: F1X IPB C15; BCP2: F1X IPB C2; F2: selfing F1
Tabel 3 Nilai rata-rata, standar deviasi, dan uji nilai tengah kejadian penyakit dan diameter nekrosis F1 dan F1R
aF1:IPB C15 X IPB C2; F1R: IPB C2 X IPB C15; tn: tidak berbeda nyata
Nilai tengah kejadian penyakit dan diameter nekrosis F1 dan F1R dibandingkan dengan melakukan uji t. Hasil pengujian menunjukkan bahwa nilai tengah F1 dan F1R tidak berbeda nyata pada kedua karakter (Tabel 3). Hal ini berarti bahwa pewarisan sifat ketahanan cabai terhadap C. acutatum tidak dipengaruhi oleh efek maternal. Dengan kata lain, sifat ketahanan tersebut dikendalikan oleh gen-gen yang berada di dalam inti sel.
Tabel 4 Komponen ragam kejadian penyakit dan diameter nekrosis pada cabai yang disebabkan oleh C. acutatum
Komponen ragam Karakter
Kejadian penyakit Diameter nekrosis
Ragam lingkungan (σE) 117.82 29.60
Ragam fenotipe (σP) 369.20 67.76
Ragam genetik (σG) 251.38 38.16
Ragam aditif (σA) 126.55 10.40
Rasio ragam aditif 50.34 27.25
Heritabilitas arti luas (h2bs) 68.09 56.32
Heritabilitas arti sempit (h2ns) 34.28 15.35
Heritabilitas merupakan komponen genetik yang menunjukkan seberapa besar suatu sifat diturunkan kepada turunannya. Heritabilitas dibedakan menjadi heritabilitas arti luas dan arti sempit. Heritabilitas arti luas diduga dari perbandingan ragam genetik dengan ragam fenotipe. Ragam genetik diduga dari pengurangan ragam populasi F2 yang mewakili ragam fenotipe dengan rata-rata ragam P1, P2, dan P3 yang mewakili ragam lingkungan. Heritabilitas arti sempit diduga dari perbandingan ragam aditif dengan ragam fenotipe. Ragam aditif
Populasia Karakter
Kejadian penyakit (%) Diameter nekrosis (mm)
F1 85±12.99 21.8±5.58
F1R 86±11.49 18.2±6.36
11 ditentukan dengan mengurangkan 2 kali ragam F2 yang merupakan ragam fenotipe dengan jumlah ragam BCP1 dan BCP2 yang merupakan ragam non aditif.
Berdasarkan hasil perhitungan diperoleh bahwa nilai heritabilitas arti luas kejadian penyakit antraknosa termasuk dalam kategori tinggi yaitu 68.09. Hal ini menunjukkan bahwa keragaman kejadian penyakit lebih dipengaruhi oleh keragaman genetik. Adapun nilai heritabilitas arti sempitnya termasuk dalam kategori sedang yaitu 34.28. Sumbangan ragam aditif terhadap ragam genetik pada karakter kejadian penyakit dapat dilihat dari nilai rasio radam aditif yaitu 50.34. Nilai ini menunjukkan sumbangan ragam aditif yang cukup tinggi yaitu 50.34% dari total ragam genetik. Karakter diameter nekrosis penyakit antraknosa yang muncul mempunyai nilai heritabilitas arti luas yang tergolong tinggi dan nilai heritabilitas arti sempit yang tergolong rendah. Sumbangan ragam aditif terhadap total ragam genetik adalah 27.55%. Menurut Syukut et al. (2011) keragaman genetik dan heritabilitas sangat bermanfaat dalam proses seleksi. Seleksi akan efektif jika populasi tersebut mempunyai keragaman genetik yang luas dan heritabilitas yang tinggi.
Beberapa penelitian mengenai ketahanan antraknosa sudah banyak dilakukan. Syukur et al. (2007), menemukan bahwa ketahanan cabai (C. annuum) terhadap antraknosa (C. acutatum) mempunyai heritabilitas arti luas yang tinggi dan heritabilitas arti sempit yang sedang. Sanjaya (2003), melaporkan bahwa ketahanan cabai hasil persilangan C. annuum dengan C. chinense terhadap antraknosa (C. gloeosporioides dan C. capsici) mempunyai nilai heritabilitas arti luas yang rendah. Yustisiani et al. (2006) menyatakan ketahanan cabai hasil persilangan cabai merah dengan cabai ungu terhadap antraknosa (C. gloeosporioides) mempunyai heritabilitas arti luas dan arti sempit yang tinggi.
Tabel 5 Heterosis dan heterobeltiosis kejadian penyakit dan diameter nekrosis antraknosa pada cabai
a
P1: IPB C15; P2: IPB C2; F1: IPB C15 X IPB C2; MP: rata-rata tetua
Nilai tengah P1 dan P2 untuk karakter kejadian penyakit berbeda jauh dan nilai tengah F1 berada di antara kedua tetuanya (Tabel 5). Nilai heterosis tetua tengah 39.92% artinya tingkat kejadian penyakit populasi F1 39.92% lebih tinggi dari nilai tengah kedua tetua. Heterobeltiosis bernilai 226.92% artinya tingkat kejadian penyakit populasi F1 226.92% lebih tinggi dari kejadian penyakit tetua P1. Nilai heterosis dan heterobeltiosis tersebut menunjukkan bahwa F1 bersifat lebih rentan terhadap antraknosa. Posisi relatif nilai tengah F1 terhadap kedua tetua dan rata-ratanya (MP) untuk karaker kejadian penyakit dapat dilihat pada Gambar 2. Berdasarkan gambar, dapat dikatakan bahwa
Komponen heterosisa Karakter
Kejadian penyakit (%) Diameter nekrosis (mm)
12
populasi F1 tidak lebih unggul dari tetuanya karena lebih mengarah ke populasi yang rentan terhadap antraknosa.
Diameter nekrosis penyakit antraknosa populasi F1 berada di antara tetua P1 dan P2. Heterosis tetua tengah sebesar 30.50% artinya, diameter nekrosis populasi F1 30.50% lebih panjang dibanding nilai tengah diameter nekrosis kedua tetuanya. Heterobeltiosis 104.41%, artinya diameter nekrosis populasi F1 104.41% lebih panjang dibanding tetua P1.
Gambar 2. Skema posisi relatif nilai tengah F1 terhadap kedua tetua dan rata-ratanya
(MP) berdasarkan skor ketahanan terhadap antraknosa
Karakter Vegetatif Cabai
Nilai rata-rata tetua P1 dan P2 tidak berbeda jauh pada karakter tinggi tanaman dan diameter batang tetapi berbeda jauh untuk karakter tinggi dikotomus (Tabel 6). Rata-rata tinggi dikotomus tetua P2 lebih tinggi daripada tetua P1. Berdasarkan kisaran tinggi tanaman yang sama dan tinggi dikotomus yang berbeda, dapat dikatakan bahwa tetua P1 mempunyai tajuk yang lebih panjang dibanding tetua P2.
Tabel 6 Nilai rata-rata, ragam, dan simpangan baku karakter tinggi tanaman, diameter batang, dan tinggi dikotomus setiap populasi cabai
a
P1: IPB C15; P2: IPB C2; F1: IPB C15 X IPB C2; F1R: IPB C2 X IPB C15; BCP1: F1 X IPB C15; BCP2: F1X IPB C2; F2: selfing F1
Keragaman yang muncul pada populasi P1, P2, F1, F1R lebih dipengaruhi oleh lingkungan, keragaman populasi BCP1, BCP2, dan F2
Karakter Populasi
a
P1 P2 F1 F1R BCP1 BCP2 F2
Tinggi tanaman (cm)
Rata-rata 49.08 54.32 52.18 59.82 54.78 50.87 63.65
Ragam 136.48 95.98 89.23 87.80 185.58 146.89 214.62
Simpangan
baku 11.68 9.80 9.45 9.37 13.62 12.12 14.65
Diamater batang (mm)
Rata-rata 9.44 9.18 9.09 9.60 9.76 8.30 10.87
Ragam 3.50 2.29 2.60 3.94 5.65 4.18 6.93
Simpangan
baku 1.87 1.51 1.61 1.99 2.38 2.04 2.63
Tinggi dikotomus (cm)
Rata-rata 8.32 20.70 12.39 14.76 11.17 14.98 15.72
Ragam 2.50 4.55 5.03 7.71 8.01 17.84 19.66
Simpangan
baku 1.58 2.13 2.24 2.78 2.83 4.22 4.43
P1 (Moderat)
MP F1 P2
(Sangat rentan)
13 dipengaruhi oleh keragaman genetik dan lingkungan. Oleh karena itu, nilai ragam populasi P1, P2, F1, dan F1R mempunyai nilai yang terkecil, diikuti oleh ragam BCP1 dan BCP2, selanjutnya ragam terbesar oleh populasi F2 untuk semua karakter vegetatif yang diamati.
Berdasarkan data Tabel 7 dapat diketahui bahwa uji t untuk membandingkan nilai tengah populasi F1 dan F1R memberikan hasil yang tidak berbeda nyata pada karakter tinggi tanaman, diameter batang, dan tinggi dikotomus. Hal ini menunjukkan bahwa tidak ada efek maternal dalam pewarisan ketiga karakter tersebut. Dengan kata lain, gen-gen pengendali karakter tersebut berada di dalam inti sel.
Tabel 7 Nilai rata-rata, standar deviasi, dan uji nilai tengah tinggi tanaman, diameter batang, dan tinggi dikotomus F1 dan F1R
Populasia
Ragam lingkungan dan ragam genetik pada karakter tinggi tanaman mempunyai proporsi sumbangan yang sama (Tabel 8). Berbeda dengan karakter diameter batang dan tinggi dikotomus yang mempunyai proporsi ragam genetik yang lebih tinggi dibanding ragam lingkungannya. Proporsi ragam tersebut menentukan nilai heritabilitas arti luas. Semakin tinggi proporsi ragam genetiknya, maka heritabilitas arti luas akan bernilai semakin tinggi. Hasil penelitian Syukur et al. (2010a) karakter tinggi tanaman, diameter batang, dan tinggi dikotomus mempunyai ragam genetik yang luas.
Tabel 8 Komponen ragam tinggi tanaman, diameter batang, dan tinggi
Ragam lingkungan (σE) 107.23 2.79 4.03
Ragam fenotipe (σP) 214.62 6.93 19.66
Ragam genetik (σG) 107.39 4.14 15.63
Ragam aditif (σA) 96.77 4.03 13.47
Rasio ragam aditif 90.11 97.34 86.18
Heritabilitas arti luas (h2bs) 50.04 59.74 79.50
Heritabilitas arti sempit (h2ns) 45.09 58.15 68.51
14
mempunyai rasio ragam aditif yang tinggi, artinya gen-gen atitif mempunyai pengaruh yang cukup besar dalam penurunan karakter-karakter tersebut.
Nilai tengah tinggi tanaman tetua P1 dan P2 tidak terlihat berbeda (Tabel 9). Nilai heterosis tinggi tanaman sangat rendah yaitu 0.93% sedangkan heterobeltiosisnya bernilai negatif yaitu -3.94%. Heterosis 0.93% artinya tinggi tanaman populasi F1 0.93% lebih tinggi dibanding nilai tengah tinggi tanaman kedua tetuanya. Heterobeltiosis -3.94% artinya tinggi tanaman populasi F1 lebih rendah 3.94% dati tinggi tanaman tetua P2. Menurut Rodrigues et al. (2012), nilai heterosis tinggi tanaman bervariasi antara positif dan negatif tergantung dari genotipe yang disilangkan. Meskipun tinggi tanaman tidak dapat digunakan sebagai kriteria seleksi untuk daya hasil (Syukur et al. 2010c), karakter ini tetap harus diperhatikan karena petani menyukai tanaman cabai yang pendek dan berbuah lebat (Kusmana et al. 2009). Mengacu pada penelitian Kusmana et al. (2009), hibrida F1 termasuk dalam kategori pendek.
Tabel 9 Heterosis dan heterobeltiosis tinggi tanaman, diameter batang, dan
Heterosis tetua tengah (%) 0.93 -2.36 -14.61
Heterobeltiosis (%) -3.94 -3.71 -40.14
aP1: IPB C15; P2: IPB C2; F1: IPB C15 X IPB C2; MP: rata-rata tetua
Karakter diameter batang dan tinggi dikotomus mempunyai heterosis dan heterobeltiosis yang bernilai negatif. Berdasarkan nilai heterosis dan heterobeltiosis yang didapatkan, diameter batang populasi F1 2.36% lebih rendah dari rata-rata diameter batang kedua tetuanya dan 3.71% lebih rendah dari tetua P1. Nilai tengah tinggi dikotomus populasi F1 14.61% lebih rendah dari rata-rata nilai tengah tinggi dikotumus kedua tetuanya dan 40.14% lebih rendah dari tetua P2.
15 Karakter Generatif Cabai
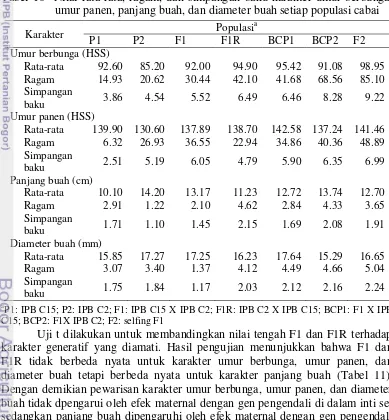
Umur berbunga dan panen tetua P1 lebih lama dibanding tetua P2. F1 mempunyai umur berbunga dan panen yang mengarah ke tetua P1. Diduga gen pengendali umur genjah tanaman bersifat resesif. Panjang dan diameter buah tetua P1 lebih pendek daripada P2, tetapi perbedaan ini tidak terlalu jauh. F1 mempunyai panjang dan diameter buah yang mengarah ke tetua P2. Diduga karakter panjang dan diameter buah dikendalikan oleh gen dominan. Penelitian Marame et al. (2008) juga menunjukkan bahwa karakter panjang buah dikendalikan oleh gen dominan. Ragam populasi P1, P2, F1, dan F1R mempunyai nilai terkecil diikuti oleh ragam BCP1, BCP2, dan selanjutnya ragam F2. Hal ini terjadi karena populai P1, P2, F1, dan F1R mempunyai susunan genetik yang homogen sehingga keragamannya hanya dipengaruhi oleh lingkungan. Sedangkan populasi BCP1, BCP2, dan F2 mempunyai keragaman susunan genetik yang lebih tinggi.
Tabel 10 Nilai rata-rata, ragam, dan simpangan baku karakter umur berbunga, umur panen, panjang buah, dan diameter buah setiap populasi cabai
Karakter Populasi
a
P1 P2 F1 F1R BCP1 BCP2 F2
Umur berbunga (HSS)
Rata-rata 92.60 85.20 92.00 94.90 95.42 91.08 98.95
Ragam 14.93 20.62 30.44 42.10 41.68 68.56 85.10
Simpangan
baku 3.86 4.54 5.52 6.49 6.46 8.28 9.22
Umur panen (HSS)
Rata-rata 139.90 130.60 137.89 138.70 142.58 137.24 141.46
Ragam 6.32 26.93 36.55 22.94 34.86 40.36 48.89
Simpangan
baku 2.51 5.19 6.05 4.79 5.90 6.35 6.99
Panjang buah (cm)
Rata-rata 10.10 14.20 13.17 11.23 12.72 13.74 12.70
Ragam 2.91 1.22 2.10 4.62 2.84 4.33 3.65
Simpangan
baku 1.71 1.10 1.45 2.15 1.69 2.08 1.91
Diameter buah (mm)
Rata-rata 15.85 17.27 17.25 16.23 17.64 15.29 16.65
Ragam 3.07 3.40 1.37 4.12 4.49 4.66 5.04
Simpangan
baku 1.75 1.84 1.17 2.03 2.12 2.16 2.24
a
P1: IPB C15; P2: IPB C2; F1: IPB C15 X IPB C2; F1R: IPB C2 X IPB C15; BCP1: F1 X IPB C15; BCP2: F1X IPB C2; F2: selfing F1
16
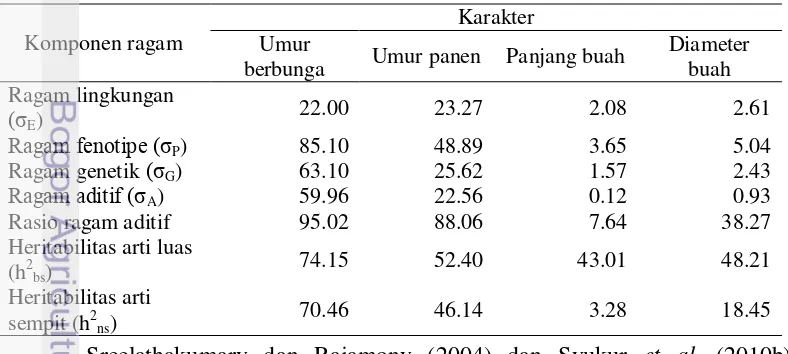
Karakter umur berbunga mempunyai ragam genetik yang lebih tinggi dibanding ragam lingkungannya (Tabel 12). Berbeda dengan karakter umur panen, panjang buah, dan diameter buah yang mempunyai ragam lingkungan dan ragam genetik dengan proporsi sumbangan sama besar. Proporsi ragam tersebut menentuntukan nilai heritabilitas arti luas. Semakin tinggi proporsi ragam genetiknya, maka heritabilitas arti luas akan bernilai semakin tinggi. Menurut Syukur et al. (2010a), karakter umur berbunga dan umur panen mempunyai ragam genetik yang sempit dan karakter panjang dan diameter buah mempunyai ragam genetik yang luas.
Tabel 11 Nilai rata-rata, standar deviasi, dan uji nilai tengah umur berbunga, umur panen, panjang buah, dan diameter buah F1 dan F1R
Populasia Tabel 12 menunjukkan heritabilitas arti luas untuk karakter umur berbunga dan umur panen tergolong tinggi dan untuk karakter panjang dan diameter buah tergolong sedang. Adapun nilai heritabilitas arti sempit ditentukan oleh besarnya ragam aditif. Heritabilitas arti sempit tergolong tinggi untuk karakter umur berbunga, sedang untuk karakter umur panen, dan rendah untuk karakter panjang dan diameter buah. Besarnya kendali ragam aditif dalam ragam genetik dapat dilihat dari rasio ragam aditif. Karakter umur berbunga dan panen mempunyai rasio ragam aditif yang tinggi, artinya gen-gen aditif mempunyai pengaruh yang cukup besar dalam penurunan karakter-karakter tersebut. Karakter panjang dan diameter buah mempunyai rasio ragam aditif kecil yang berarti bahwa kedua karakter tersebut lebih dipengaruhi oleh gen-gen non aditif. Tabel 12 Komponen ragam umur berbunga, umur panen, panjang buah, dan
Ragam fenotipe (σP) 85.10 48.89 3.65 5.04
Ragam genetik (σG) 63.10 25.62 1.57 2.43
Ragam aditif (σA) 59.96 22.56 0.12 0.93
Rasio ragam aditif 95.02 88.06 7.64 38.27
17 heritabilitas arti luas untuk karakter panjang dan diameter buah tergolong tinggi. Marame et al. (2008) menunjukkan bahwa karakter umur panen mempunyai heritabilitas arti luas yang tinggi dan heritabilitas arti sempit yang rendah.
Heterosis dan heterobeltiosis digunakan untuk mengetahui keunggulan F1 dibanding kedua tetuanya. Semua karakter yang diamati mempunyai heterosis yang bernilai positif. Karakter umur berbunga dan panen mempunyai heterobeltiosis yang bernilai positif dan karakter panjang dan diameter buah mempunyai heterobeltiosis yang bernilai negatif. Heterosis yang rendah (Tabel 13) menunjukkan bahwa F1 tidak jauh berbeda dengan rata-rata kedua tetuanya. Heterosis dan heterobeltiosis karakter panjang dan diameter buah bervariasi bisa positif dan negatif tergantung genotipe yang disilangkan (Sujiprihati et al. 2007; Daryanto et al. 2010).
Nilai heterosis dan heterobeltiosis yang positif untuk karakter umur berbunga dan panen menunjukkan bahwa F1 berumur lebih dalam dibanding umur kedua tetuanya. Menurut Syukur et al. (2010d), tanaman cabai dikategorikan genjah jika memiliki umur panen lebih genjah dari cabai varietas Hot beauty (dilaporkan umur panennya 112-118 HSS). Berdasarkan ketentuan tersebut, F1 termasuk tanaman berumur dalam. Padahal karakter unggul yang menjadi sasaran program pemuliaan (Hilmayanti et al. 2006) dan disukai oleh masyarakat adalah cabai yang berumur genjah. Dengan demikian, F1 tidak memenuhi keunggulan berumur genjah.
Tabel 13 Heterosis dan heterobeltiosis umur berbunga, umur panen, panjang buah, dan diameter buah pada cabai
Komponen heterosisa
18
SIMPULAN DAN SARAN
Simpulan
Ketahanan cabai (Capsicum annuum L.) terhadap antraknosa (Colletotrichum acutatum) dikendalikan oleh gen resesif. Tidak ada efek maternal dalam penurunan karakter yang diamati kecuali karakter panjang buah. Heritabilitas arti luas tergolong tinggi untuk karakter kejadian penyakit, diameter nekrosis, tinggi tanaman, diameter batang, tinggi dikotomus, umur berbunga, dan umur panen; dan tergolong sedang untuk karakter panjang dan diameter buah. Heritabilitas arti sempit tergolong tinggi untuk karakter diameter batang, tinggi dikotomus, dan umur berbunga; sedang untuk karakter kejadian penyakit, tinggi tanaman, dan umur panen; dan rendah untuk karakter diameter nekrosis, panjang buah dan diameter buah. Rasio agam aditif bernilai tinggi untuk karakter kejadian penyakit, tinggi tanaman, diameter batang, tinggi dikotomus, dan umur berbunga; sedang untuk karakter diameter nekrosis, umur panen, dan diameter buah; dan rendah untuk karakter panjang buah. Pembentukan varietas cabai tahan antraknosa dan berdaya hasil baik sebaiknya diarahkan pada varietas open pollinated (bersari bebas).
Saran
Perlu dilakukan penelitian mengenai ketahanan terhadap penyakit antraknosa menggunakan populasi persilangan yang baru untuk melihat konsistensi nilai duga parameter genetik dan karakter kuantitaif pada cabai.
UCAPAN TERIMA KASIH
Penulis menyampaikan ucapan terima kasih kepada Kementerian Pertanian yang telah membiayai penelitian ini melalui hibah KKP3T tahun 2012 a.n. M Syukur.
DAFTAR PUSTAKA
Agrios GN. 1997. Plant Pathology. Fourth Edition. New York (US): Academic Press.
Allard RW. 1960. Principles of Plant Breeding. New York (US): J Wiley & Sons. Arif AB, Sujiprihati S, Syukur M. 2012. Pendugaan heterosis dan heterobeltiosis
pada enam genotip cabai menggunakan analisis silang dialel penuh. J. Hort. 22(2):103:110.
19 [BPS] Badan Pusat Statistika. 2013. Laporan Bulanan Data Sosial Ekonomi.
Edisi 38. Jakarta (ID): Badan Pusat Statistik.
Daryanto A, Sujiprihati S, Syukur M. 2010. Heterosis dan daya gabung karakter agronomi cabai (Capsicum annuum L.) pada persilangan half diallel. J. Agron. Indonesia 38(2):113-121.
Dieguez-Uribeondo J, Forster H, Soto-Estrada A, Adaskaveg JE. 2005. Subcuticular-intracellular hemibiotrophic and intercellular necrotrophic development of Colletotrichum acutatum on almond. Phytopathology 95:751-758.
Efri. 2010. Pengaruh ekstrak berbagai bagian tanaman mengkudu (Morinda citrifolia) terhadap perkembangan penyakit antraknosa pada tanaman cabe (Capsicum annuum L.). J. HPT Tropika 10:52-58.
Freeman S, Katan, T. 1997. Identification of Colletotrichum species responsible for anthracnose and root necrosis of strawberry in Israel. Phytopathology 87:516-521.
Freeman S, Katan T, Shabi E. 1998. Characterizazion of Colletotrichum spesies responsible for anthracnose disease of various fruits. Plant disease 82(6):596-605.
Golzar H, Wang C. 2010. First report of Colletotrichum phormii the cause anthracnose of Phormium texas in Australia. Australian Plant Disease Notes 5:110-112.
Halloran GM, Knight R, McWhirter KS, Sparrow DHB. 1979. Plant Breeding. Brisbane (AU): Australian Vice Chancellors Committee.
Hartuti N, Asgar A. 1992. Kualitas bahan baku dan hasil olahan cabai di tingkat industri komersial dan rumah tangga di bandung. Bul. Penel. Hort. 26:142-150.
Herison C, Rustikawati, Sudarsono. 2008. Seleksi indeks kearah daya hasil dan ketahanan terhadap virus pada genotype cabai merah introduksi. Jerami 1(3):105-110.
Hilmayanti I, Dewi W, Murdaningsih, Rahardja M, Rostini N, Setiamihardja R. 2006. Pewarisan karakter umur berbunga dan ukuran buah cabai merah (Capsicum annuum L.). Zuriat 17:86-93.
Humaedah U. 2012. Penyakit tanaman cabai: antraknosa, busuk batang dan busuk daun bagaimana mengendalikannya?[internet]. [diunduh 2012 Maret 12]. Tersedia pada Http://www.deptan.go.id.
Hwang S, Kim J, Kim YH, Kim HT. 2008. ABA increases susceptibility of pepper fruits to infection of anthracnose by Collectotrichum acutatum. Plant Pathol. J. 24(4):400-406.
Ivey MLL, Mera JR, Miller SA. 2004. Evaluation of fungicides for the control of anthracnose on immature pepper fruit. Plant Pathology 126:7-8.
Kaur, N., J.S. Dhiman, and D.S. Khurana. 2011. Physiological and biochemical traits analysis of Capsicum annuum L. germplasm for resistence to Colletotrichum capsici. Journal of Cell Plant Sciences 2(3):12-21.
20
Kim SG, Ro NY, Hur OS, Ho-Cheol, Gwag JG, Huh YC. 2012. Evaluation of resistance to Colletotrichum acutatum in pepper genetic resources. Research in Plant Disease 18(2):93-100.
Kim SH, Yoon JB, Do JW, Park HG. 2007. Resistance to anthracnose caused by Colletotrichum acutatum in chili pepper (Capsicum annuum L.). J. Crop Sci. Biotech. 10(4):277-280.
Kim SH, Yoon JB, Park HG. 2008a. Inheritance of anthracnose resistance in a new genetic resource, Capsicum baccatum PI1594137. J. Crop Sci. Biotech. 11(1):13-16.
Kim SH, Yoon JB, Do JW, Park HG. 2008b. A major recessive gene associated with anthracnose resistance to Colletotrichum capsici in chili pepper (Capsicum annuum L.). Breeding Science 58:137-141.
Kirana R, Sofiari E. 2007. Heterosis dan heterobeltiosis pada persilangan lima genotip dengan metode dialel. J. Hort. 17(2):111-117.
Kraikruan W, Sangchote S, Sukprakarn S. 2008. Effect of capcaisin on germination of Colletotrichum capsici conidia. Kasetsart J. (Nat. Sci.) 42:417-422.
Kusmana, Kirana R, Hidayat IM, Kusandriani Y. 2009. Uji adaptasi beberapa galur cabai merah di dataran medium Garut dan dataran tinggi Lembang. J. Hort. 19(4):371-376.
Lee J, Hong JH, Do JW, Yoon JB. 2010. Identification of QTLs for resistance to anthracnose to two Colletotrichum species in pepper. J. Crop Sci. Biotech. 13(4):227-233.
Mahasuk P, Taylor PWJ, and Mongkolporn O. 2009. Identification of two new genes conferring resistance to Colletotrichum acutatum in Capsicum baccatum. Phytopathology 99:1100-1104.
Marame F, Desalegne L, Singh H, Fininsa C, Sigvald R. 2008. Genetic components and heritability of yield and yield related traits in hot pepper. Research Journal of Agriculture and Biological Sciences 4(6):803-809. Marliyanti L, Syukur M, Widodo. 2013. Daya hasil 15 galur cabai ipb dan
ketahanannya terhadap penyakit antraknosa yang disebabkan oleh Colletotrichum acutatum.Bul. Agrohorti 1(1):7–13.
Marvel JK. 2003. Biology and Control of Anthracnose [thesis]. Virginia (US): Virginia Polytechnic Institute and State University.
Mistry DS, Sharma IP, Patel ST. 2008. Biochemical parameters of chilli fruits as influenced by Colletotrichum capsici (Sydow) Butler and Bisby infection. Karnataka Journal of Agricultural Science 21(4):586-587.
Mongkolporn O, Montri P, Supakaew T, Taylor, PWJ. 2010. Differential reactions on mature green and ripe chili fruit infected by three Colletotrichum spp. Plant Dis. 94:306-310.
Nayaka SC, Shankar ACU, Niranjaya SR, Prakash HS, and Mortensen CN. 2009. Anthracnose disease of chilli pepper. Technical Buletin.
Oanh LTK, Korpraditskul V, Rattanakreetaku C. 2004. A pathogenicity of anthracnose fungus, Colletotrichum capsici on various Thai chilli varieties. Kasetsart J. (Nat. Sci.) 38:103–108.
21 Park. 2005. Differential Interaction Between Pepper Genotypes and Colletotrichum Isolates Causing Anthracnose [thesis]. Seoul (KR): Seoul Natl. Univ
Prasath D, Ponnuswami, Muralidharan V. 2007. Source of resistance to anthracnose (Colletotrichum capsici) disease in Capsicum species. Indian Journal of Agricultural Sciences 77(7):73-74.
Roca MG, Davide LC,Mendes-Costa MC,Wheals L. 2003. Conidial anastomosis tubes in Colletotrichum. Fungal Genetics and Biology 40:138–145. Rodrigues R, Goncalves L, Bento CS, Sundre CP, Robaina RR, Amaral JAT.
2012. Combining ability and heterosis for agronomic traits in chili pepper. Horticultura Brasileira 30:226-233.
Sanjaya L. 2003. Keterpautan marka amplified fragment length polymorphism dengan sifat resisten penyakit antraknos pada cabai berdasarkan metode bulk segregant analysis. J. Hort. 13(3):169-176.
Sergeeva V, Nair NG, Spooner-Hart R. 2008. Evidence of early flower infection in olives (Olea europaea) by Colletotrichum acutatum and C. gloeosporioides causing anthracnose disease. Australasian Plant Disease Notes 3:81-82.
Sharma VK, Semwal CS, Uniyal SP. 2010. Genetic variability and character association analysis in bell pepper (Capsicum annuum L.). Journal of Horticulture and Forestry 2(3):058-065.
Sreelathakumary I, Rajamony L. 2004. Variability, heritability and genetic advance in chilli (Capsicum annuum L.). Journal of Tropical Agriculture 42(1-2):35-37.
Stankova B, Vichova J, Pokorny R. 2011. Virulence of Colletotrichum acutatum isolates to several host plant. Acta Universitatis Agriculturae Et Silviculrurae Mendelianae Brunensis 59(3):161-170.
Sujiprihati S, Yunianti R, Syukur M, Undang. 2007. Pendugaan nilai heterosis dan daya gabung beberapa komponen hasil pada beberapa persilangan dialel penuh enam genotipe cabai (Capsicum annuum L.). Bul. Agron. 35(1):28-35.
Syukur M. 2007. Analisis Genetik dan Studi Pewarisan Sifat Ketahanan Cabai (Capsicum annuum L.) terhadap Antraknosa yang Disebabkan oleh Colletotrichum acutatum [disertasi]. Bogor (ID): Institut Pertanian Bogor. Syukur M, Sujiprihati S, Koswara J, Widodo. 2007. Pewarisan ketahanan cabai (Capsicum annuum L.) terhadap antraknosa yang disebabkan oleh Colletotrichum acutatum. Bul. Agron. 35:112-117.
Syukur M, Sujiprihati S, Koswara J, Widodo. 2009. Ketahanan terhadap antraknosa yang disebabkan oleh Colletotrichum acutatum pada beberapa genotipe cabai (Capsicum annuum L.) dan korelasinya dengan kandungan kapcaisin dan peroksidase. J. Agron. Indonesia 37(3):233-239.
Syukur M, Sujiprihati S, Siregar A. 2010a. Pendugaan parameter genetik beberapa karakter agronomi cabai F4 dan evaluasi daya hasilnya menggunakan rancangan perbesaran (augmented design). Jurnal Agrotropika 15(1):9-16.
22
pepper (Capsicum annuum L.). HAYATI Journal of Biosciences 17(4):183-188.
Syukur M, Sujiprihati S, Yunianti R, Nida K. 2010c. Pendugaan komponen ragam, heritabilitas, dan korelasi untuk menentukan kriteria seleksi cabai (Capsicum annuum L.) populasi F5. J. Hort. Indonesia 1(3);74-80.
Syukur M, Sujiprihati S, Yunianti R, Kusumah DA. 2010d. Evaluasi daya hasil cabai hibrida dan daya adaptasinya di empat lokasi dalam dua tahun. J. Agron. Indonesia 38(1):43-51.
Syukur M, Sujiprihati S, Yunianti R, Kusumah DA. 2011. Pendugaan ragam genetic dan heritabilitas karakter komponen hasil beberapa genotipe cabai. J. Agrivigor 10(2):148-156.
Than PP, Jeewon R, Hyde KD, Pongsupasamit S, Mongkolporn O, Taylor PWJ. 2008a. Characterization and pathogenicity Colletotrichum species associated with anthracnise in chilli (Capsicum spp.) in Thailand. Plant Pathology 57:562-572.
Than PP, Prihastuti H, Phoulivong S, Taylor PWJ, Hyde KD. 2008b. Chili anthracnose disease caused by Colletotrichum species. J Zhejiang Univ Sci B. 9(10): 764–778.
Warin I, Chaiyawat S, Chiradej C, Montree I, Sorwaporn K, Kan C. 2009. Bioactive compound of antifungal metabolite from Trichoderma harzianum mutant strain for the control of anthracnose of chili (Capsicum annuum L.). Philipp Agric Scientist 92(4):392-397.
Warner JN. 1952. A method of estimating heritability. Agron. J. 44:427-430. Wharton PS, Uribeondo JD. 2004. The biology of Colletotrichum acutatum.
Anales del Jardin Botanico de Madrid 61(1): 3-22.
Wusani M. 2004. Pola pewarisan Karakter Ketahanan terhadap Penyakit Antraknosa (Colletotrichum gloesporioides Penz.) pada Cabai (Capsicum annuum var Jatilaba x Capsicum chinense-27) [tesis]. Bogor (ID): Institut Pertanian Bogor.
Yoon JB, Park HG. 2001. Screening method for resistance tp pepper fruit anthracnose: pathogen sporulation, inoculation methods related to inoculums concentrations and post-inoculation environment. J. Kor. Soc. Hort. Sci. 42(4):389-393.
Yunianti R, Sastrosumarjo S, Sujiprihati S, Surahman M, Hidayat SH. 2010. Kriteria seleksi untuk perakitan varietas cabai tahan Phytophtora capsici Leonian. J. Agron. Indonesia 38(2):122-129.
Yustisiani D, Dewi W, Rachmadi M, Ruswandi D, Rostini N, Setiamihardja R. 2006. Pewarisan karakter ketahanan terhadap antraknos (Colletotrichum gloeosporioides) pada hasil persilangan tanaman cabai ungu x cabai merah genotip RS07. Zuriat 17(2):154-163.
23
RIWAYAT HIDUP
Penulis, Syaidatul Rosidah, lahir di Rembang pada tanggal 24 Januari 1991. Penulis merupakan anak kedua dari Bapak Ahmad Faozi dan Ibu Siti Kalimah. Selama masa pendidikan, penulis tinggal bersama Bapak Sukardi sebagai wali.
Penulis menempuh pendidikan menengah di SMA N 1 Pamotan. Berdasarkan hasil SNMPTN, penulis dinyatakan diterima di Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut pertanian Bogor, dan mulai aktif sebagai
mahasiswi pada tanggal 10 Agustus 2009. Selama menjadi mahasiswi IPB, penulis mengikuti organisasi Bina Desa Fakultas Pertanian tahun 2010 dan Himpunan Mahasiswa Agronomi (HIMAGRON) tahun 2011-2012. Penulis juga aktif dalam berbagai kepanitian seperti panitia Masa Perkenalan Fakultas dan Departemen, Lomba Karya Tulis Ilmiah dan Seminar Nasional cabai Nusantara tahun 2012, dan Festival Bunga dan Buah Nusantara tahun 2013.
Adapun prestasi yang pernah diraih penulis selama belajar di IPB adalah mengikuti kegiatan PKM-P didanai DIKTI pada tahun 2011 dengan judul
“Pengaruh Pengaturan Kemasan Kalium Permanganat (KMnO4) dan Plastik
Polietilen dalam Penyimpanan Buah Klimakterik”, terdaftar sebagai salah satu
inovator dalam program 104 Inovasi Indonesia dengan judul paper “Oksidator
Etilen untuk Memperpanjang Masa Simpan Buah Klimakterik” dan judul invensi
“Masak Pelan-pelan, Makan Kapan-kapan”, mengikuti program PKM-P didanai
DIKTI tahun 2012 dengan judul ” Karakterisasi Beberapa Genotipe Lokal Ubi Kayu Rendah HCN untuk Perakitan Varietas dalam Mendukung Ketahanan dan Kedaulatan Pangan Nasional”, dan PKM-K didanai DIKTI tahun 2012 dengan
judul “T-Valia: Tahu dengan Bahan Dasar Kacang Koro Pedang (Canavalia