UNTUK MERAKIT KEDELAI TOLERAN
INTENSITAS CAHAYA RENDAH
DESTA WIRNAS
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRACT
DESTA WIRNAS. Selection Character for Low-Light Tolerance in Soybean Based
on Biometric and Molecular Analysis under the direction of Didy Sopandie, Trikoesoemaningtyas and Sobir.
Soybean is a high value commodity because it is the primary source of protein for the people of Indonesia. The increase of the domestic soybean production is important to fulfill soybean consumption. The increase of the domestic production could be reached by extensification of harvesting area and improvement of productivity. One of the alternatives for extensification of soybean production is growing soybean as intercrop with estate crops. The main problem in growing soybean as intercrop with estate crops is low light intensity. The problem of low light intensity in the intercropping of soybean with estate crops should be overcome by developing tolerant soybean varieties. Developing tolerant soybean varieties to low light intensity needs genetic markers as spesific selection criteria or marker assisted selection.
The objective of this study was to develop selection criteria for low light intensity tolerance in soybean based on quantitative and molecular analysis. Quantitative analysis was conducted to estimate the inheritance pattern of agronomic traits under low light intensity which was conducted in a diallel matting design; whereas, the molecular analysis was conducted to identify RAPD markers linked to QTL related to agronomic traits under low-light intensity condition. Four parental lines were used in the diallel crossing, namely, Ceneng, Godek (tolerant parents), Slamet and Pangrango (sensitive parents). The genetic materials used in the QTL analysis based on RAPD markers were Ceneng, Godek and F6 Recombinant Inbred Lines derived from the hybridization between Ceneng and Godek.
The diallel analysis revealed that the additive-dominant gene action was sufficient to explain the variability of agronomic traits of soybean under low light intensity condition except for the number of total pod per plant. The allelic interaction for plant height, number of productive branch, number of total node and number of filled pod was over dominance; whereas, number of unfilled pod, number of total pod, percentage of filled fod and seed weight were controlled by partial-dominant gene action. The broad-sense heritabilities were high for all of the traits (0.75-0.96); whereas, narrow sense heritabilities were high for the number of unfilled pod, percentage of filled fod and seed weight. The results suggested that seed weight is the selection criteria for yield improvement of soybean under low light intensity condition.
The results of molecular analysis show that 9 primers were polymorphic and linked to the tolerant parent (Ceneng). The Primers only produced 14 RAPD markers which were polymorphic and linked to the tolerant parent. The markers are distributed into a linkage group containing seven markers. RAPD markers (OPE15-800, OPM20-800) is linked to two QTLs controlling number of productive node and seed weight, respectively. The Marker linked to QTL controlling seed weight could be used in a marker assisted selection for tolerant soybean lines to low light intensity.
ABSTRAK
DESTA WIRNAS. Pemilihan Karakter Seleksi Berdasarkan Analisis Biometrik dan
Molekuler untuk Merakit Kedelai Toleran Intensitas Cahaya Rendah. Dibimbing oleh Didy Sopandie, Trikoesoemaningtyas dan Sobir.
Kedelai adalah komoditi yang bernilai ekonomi tinggi karena merupakan sumber utama protein nabati bagi bangsa Indonesia. Peningkatan produksi kedelai sangat penting dilakukan untuk memenuhi kebutuhan kedelai nasional melalui perluasan areal panen dan peningkatan produktivitas. Perluasan areal panen dapat dilakukan dengan memanfaatkan lahan yang ada di bawah tegakan tanaman perkebunan ( kondisi intensitas cahaya rendah). Kendala utama budidaya kedelai di bawah tegakan adalah berkurangnya intensitas cahaya yang diterima oleh tanaman sehingga perlu tersedia varietas yang adaptif dan berdaya hasil tinggi dalam kondisi intensitas cahaya rendah.
Tujuan penelitian ini adalah memilih karakter seleksi bagi kedelai toleran intensitas cahaya rendah melalui analisis kuantitatif dan molekuler. Analisis kuantitatif dilakukan berdasarkan analisis dialel yang bertujuan untuk mengetahui pola pewarisan karakter agronomi kedelai pada kondisi intensitas cahaya rendah. Bahan tanaman yang digunakan dalam analisis dialel adalah empat tetua yaitu Ceneng dan Pangrango (tetua toleran) serta Slamet dan Godek (tetua peka), dan F1 hasil persilangan di antara keempat tetua. Analisis molekuler dilakukan melalui analisis QTL yang bertujuan untuk mendapatkan marka molekuler yang terpaut dengan QTL yang mengendalikan daya hasil pada kondisi intensitas cahaya rendah. Bahan tanaman yang digunakan dalam analisis QTL adalah Ceneng, Godek dan
rekombinant inbreed lines generasi F6 hasil persilangan kedua tetua.
Hasil penelitian menunjukkan bahwa model genetik aditif-dominan mampu menjelaskan keragaman karakter agronomi kedelai pada kondisi intensitas cahaya rendah kecuali karakter jumlah polong total. Ragam aditif berpengaruh nyata untuk karakter jumlah polong hampa, persentase polong isi dan daya hasil sedangkan ragam dominan berpengaruh nyata untuk semua karakter yang diamati. Nilai heritabilitas arti luas tergolong tinggi untuk semua karakter yaitu berkisar antara 75.0–96.0%, sedangkan nilai heritabilitas arti sempit tergolong tinggi hanya pada karakter jumlah polong hampa, persentase polong isi dan daya hasil Berdasarkan hasil penelitian ini maka karakter seleksi untuk perbaikan daya hasil kedelai pada kondisi intensitas cahaya rendah adalah daya hasil.
Hasil yang diperoleh dari analisis molekuler adalah 9 primer RAPD yang menghasilkan 14 marka polimorfik dan terpaut dengan tetua toleran terhadap intensitas cahaya rendah. Konstruksi peta pautan dibuat dengan menggunakan 14 marka RAPD tersebut menghasilkan satu kelompok pautan yang mengandung tujuh marka. Dalam penelitian ini diperoleh dua QTL yang masing-masing mengendalikan karakter jumlah buku total dan daya hasil. Marka yang terpaut dengan QTL yang mengendalikan karakter jumlah buku total adalah OPE15-800, sedangkan marka RAPD yang terpaut dengan QTL yang mengendalikan karakter daya hasil adalah OPM20-800. Berdasarkan hasil penelitian ini maka disarankan untuk menggunakan marker yang terpaut dengan QTL yang mengendalikan daya hasil sebagai alat bantu seleksi bagi kedelai toleran terhadap intensitas cahaya rendah.
SURAT PERNYATAAN
Saya menyatakan dengan sebenar-benarnya, bahwa segala pernyataan dalam disertasi yang berjudul :
PEMILIHAN KARAKTER SELEKSI BERDASARKAN ANALISIS BIOMETRIK DAN MOLEKULER UNTUK MERAKIT KEDELAI
TOLERAN INTENSITAS CAHAYA RENDAH
adalah benar merupakan hasil karya saya sendiri dengan arahan komisi pembimbing dan belum pernah dipublikasikan. Semua data dan informasi yang dikutip dari karya yang diterbitkan oleh penulis lain telah disebutkan dalam teks dan dicantumkan
dalam daftar pustaka.
Bogor, 23 Agustus 2007
©Hak cipta milik
Institut Pertanian Bogor
, tahun 2007
Hak cipta dilindungi
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari
Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk
PEMILIHAN KARAKTER SELEKSI BERDASARKAN ANALISIS
BIOMETRIK DAN MOLEKULER BAGI KEDELAI TOLERAN
INTENSITAS CAHAYA RENDAH
OLEH:
DESTA WIRNAS
A361030041
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor
pada Program Studi Agronomi
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Judul : Pemilihan Karakter Seleksi Berdasarkan Analisis Biometrik dan Molekuler untuk Merakit Varietas Kedelai Toleran Intensitas Cahaya Rendah
Nama Mahasiswa : Desta Wirnas
Nomor Pokok : A361030041
Menyetujui: Komisi Pembimbing
Prof. Dr. Ir. Didy Sopandie, MAgr Ketua
Dr. Ir. Trikoesoemaningtyas, MSc Anggota
Dr. Ir. Sobir, MS Anggota
Mengetahui:
Ketua Program Studi Agronomi
Dr. Ir. Satriyas Ilyas, MSc
Dekan Sekolah Pascasarjana IPB
Prof. Dr. Ir. Khairil A. Notodiputro, MSc
RIWAYAT HIDUP
Penulis dilahirkan di Kampung Baru, Sumatera Barat pada tanggal 28 Desember 1970 sebagai anak kedua dari pasangan M. Said Pk. Sati dan Rosanah (alm.). Penulis telah menikah dengan Ir. Saiful, MM dan telah dikaruniai tiga orang putra dan putri yaitu Muhammad Hanif Saiful (9 tahun), Ghania Sakira Saiful (5 tahun) dan Hadin Harridhy Saiful (4 tahun).
Pendidikan sarjana pertanian di bidang Ilmu dan Teknologi Benih ditempuh di Jurusan Budidaya Pertanian, Fakultas Pertanian, Institut Pertanian Bogor pada tahun 1990. Pada tahun 1996 penulis melanjutkan program master di bidang Pemuliaan Tanaman, Fakultas Pascasarjana, Institut Pertanian Bogor. Kesempatan untuk melanjutkan pendidikan program doktor di bidang Pemuliaan Tanaman penulis
peroleh tahun 2003 atas beasiswa dari Departemen Pendidikan Nasional RI. Penulis bekerja sebagai staf pengajar pada laboratorium Genetika dan Pemuliaan Tanaman,
Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor sejak 1999 sampai sekarang.
KATA PENGANTAR
Puji dan syukur ke hadirat Allah SWT, berkat rahmat dan hidayah-Nya sehingga disertasi ini dapat diselesaikan. Penelitian yang berjudul Pemilihan Karakter Seleksi Berdasarkan Analisis Biometrik dan Molekuler untuk
Merakit Kedelai Toleran Intensitas Cahaya Rendah telah diselesaikan dalam
waktu 20 bulan di bawah bimbingan dan dukungan komisi pembimbing yaitu Prof. Dr. Ir. Didy Sopandie, MAgr, Dr. Ir. Trikoesoemaningtyas, MSc dan Dr. Ir. Sobir, MS.
Penelitian dan penulisan disertasi ini dapat diselesaikan berkat bantuan dari Direktorat Jendral Pendidikan Tinggi, Departemen Pendidikan Nasional melalui beasiswa BPPS tahun 2003-2006 dan Hibah Tim Penelitian Pascasarjana tahun
2004-2005 serta bantuan biaya penelitian dari L’Oreal Indonesia-Komisi Nasional Indonesia untuk UNESCO.
Bantuan dan dukungan dari berbagai pihak berupa material maupun non material merupakan karunia Allah SWT kepada Penulis, untuk itu penulis menyampaikan terima kasih dan penghargaan yang tak terhingga kepada:
1. Tim BPPS, Dirjen Pendidikan Tinggi yang telah memberikan beasiswa kepada penulis untuk mengikuti program doktor di IPB
2. Prof. Dr. Ir. Didy Sopandie, MAgr sebagai Ketua Komisi Pembimbing serta Dr. Ir. Trikoesoemaningtyas, MSc dan Dr. Ir. Sobir, MS, sebagai Anggota Komisi Pembimbing
3. Prof. Dr. Ir. Didy Sopandie, MAgr, Dr. Ir. Trikoesoemaningtyas, MSc dan Dr. Ir. Nurul Khumaida, MS yang telah memberikan kesempatan kepada penulis untuk bergabung dalam Hibah Tim Penelitian Pascasarjana pada tahun 2004-2005
4. Dr. Yudiwanti sebagai penguji pada saat ujian tertutup serta Dr. Sriani Sutjiprihati dan Dr. Darman M.Arsyad sebagai penguji luar komisi pada saat ujian terbuka
5. L’Oreal Indonesia-Komisi Nasional Indonesia untuk UNESCO atas bantuan dana penelitian
7. Kepala dan staf Kebun Percobaan Balai Besar Bioteknologi dan Sumberdaya Genetik, Cimanggu Bogor
8. Kepala Pusat Penelitian Bioteknologi Perkebunan, Bogor serta Kepala dan staf Lab. Molekuler Tanaman khususnya Dr. Nurhaimi, Nani, Nurul dan Yadi
9. Kepala dan staf Lab. RGCI, Dept. Agronomi dan Hortikultura Fakultas Pertanian IPB
10.Rektor IPB yang telah memberikan izin kepada penulis untuk melanjutkan pendidikan doktor di IPB
11.Sekolah Pascasarjana dan PS Agronomi IPB yang telah menerima penulis sebagai mahasiswa SPs IPB
12.Ketua dan Staf Pengajar Dept. Agronomi dan Hortikultura Fakultas Pertanian IPB atas segala dukungan yang diberikan
13.Staf Bagian Genetika dan Pemuliaan Tanaman atas dukungan dan bantuan dana yang diberikan
14.Orang tua dan kerabat penulis yang selalu memberikan motivasi dan do’a selama penulis mengikuti program doktor di IPB
15.Suami dan anak-anak tercinta yang telah memberikan cinta, pengertian serta do’a dan dukungan selama menyelesaikan pendidikan
16.Teman-teman staf AGH angkatan 2003 (Dini Dinarti, Dewi Sukma dan M. Syukur) atas kebersamaan selama menjalani program S3 di IPB
17.Teman-teman di SPs PS Agronomi khususnya Tim HPTP (La Muhuria, Kisman, Imam Widodo, Ika kartika, Tri Lestari) atas dukungan dan kerjasama selama pelaksaan penelitian
18. Semua pihak yang tidak dapat penulis sebutkan satu per satu
Akhir kata semoga Allah SWT berkenan membalas budi baik Bapak dan Ibu semua. Mudah-mudahan karya ilmiah ini dapat memperkaya keilmuan dan bermanfaat bagi semua.
Bogor, 23 Agustus 2007
DAFTAR ISI
PENDAHULUAN ...Error! Bookmark not defined. Latar Belakang ...Error! Bookmark not defined.
Tujuan...Error! Bookmark not defined.
TINJAUAN PUSTAKA ...Error! Bookmark not defined. Arah Pengembangan Kedelai ...Error! Bookmark not defined.
Pertumbuhan dan Produksi Kedelai ...Error! Bookmark not defined.
Pemuliaan bagi Lingkungan Bercekaman ...Error! Bookmark not defined.
Perakitan Varietas Kedelai Toleran Intensitas Cahaya RendahError! Bookmark not defined.
Pembentukan Karakter seleksi ...Error! Bookmark not defined.
Analisis QTL ...Error! Bookmark not defined.
PEWARISAN KARAKTER AGRONOMI KEDELAI PADA KONDISI
INTENSITAS CAHAYA RENDAH ...Error! Bookmark not defined.
PENDAHULUAN...Error! Bookmark not defined.
Latar Belakang ...Error! Bookmark not defined.
Tujuan...Error! Bookmark not defined.
BAHAN DAN METODE ...Error! Bookmark not defined.
Waktu dan Tempat Penelitian ...Error! Bookmark not defined.
Metode...Error! Bookmark not defined.
Analisa Data ...Error! Bookmark not defined.
HASIL DAN PEMBAHASAN ...Error! Bookmark not defined.
KESIMPULAN DAN SARAN ...Error! Bookmark not defined.
Kesimpulan...Error! Bookmark not defined.
Saran...Error! Bookmark not defined.
ANALISIS QTL YANG MENGENDALIKAN KARAKTER AGRONOMI
KEDELAI PADA KONDISI INTENSITAS CAHAYA RENDAH ...Error!
Bookmark not defined.
PENDAHULUAN...Error! Bookmark not defined.
Latar Belakang ...Error! Bookmark not defined.
Tujuan...Error! Bookmark not defined.
BAHAN DAN METODE ...Error! Bookmark not defined.
1. Pembentukan Populasi Pemetaan: Pembentukan RILs F6Error! Bookmark not defined.
2. Analisis Fenotipe Tetua dan RILs F6...Error! Bookmark not defined.
3. Konstruksi Peta Pautan Toleransi terhadap Intensitas Cahaya RendahError! Bookmark not defined.
HASIL DAN PEMBAHASAN ...Error! Bookmark not defined.
1. Pembentukan RILs F6 ...Error! Bookmark not defined.
2. Analisis Fenotipe Rils F6 pada Kondisi Intensitas Cahaya Rendah Error! Bookmark not defined.
3. Konstruksi Peta Pautan Toleransi terhadap Intensitas Cahaya RendahError! Bookmark not defined.
Kesimpulan...Error! Bookmark not defined.
Saran...Error! Bookmark not defined.
PEMBAHASAN UMUM ...Error! Bookmark not defined.
KESIMPULAN DAN SARAN ...Error! Bookmark not defined.
DAFTAR PUSTAKA ...Error! Bookmark not defined.
DAFTAR TABEL
No Halaman
1. Rekapitulasi hasil analisis ragam pengaruh genotipe terhadap karakter
agronomi kedelai pada kondisi intensitas cahaya rendahError! Bookmark not defined.
2. Hasil pengujian kesesuaian model aditif-dominan bagi karakter agronomi kedelai kondisi intensitas cahaya rendah...Error! Bookmark not defined.
3. Ragam dan peragam array karakter tinggi tanaman saat panen pada kondisi intensitas cahaya rendah ...Error! Bookmark not defined.
4. Ragam dan peragam array karakter jumlah cabang produktif pada kondisi intensitas cahaya rendah...Error! Bookmark not defined.
5. Ragam dan peragam array karakter jumlah buku total pada kondisi intensitas cahaya rendah...Error! Bookmark not defined.
6. Ragam dan peragam array karakter jumlah polong isi pada kondisi intensitas cahaya rendah...Error! Bookmark not defined.
7. Ragam dan peragam array karakter jumlah polong hampa pada kondisi
intensitas cahaya rendah...Error! Bookmark not defined.
8. Ragam dan peragam array karakter jumlah polong total pada kondisi intensitas cahaya rendah...Error! Bookmark not defined.
9. Ragam dan peragam array karakter persentase polong isi pada kondisi intensitas cahaya rendah...Error! Bookmark not defined.
10. Ragam dan peragam array karakter daya hasil pada kondisi intensitas cahaya rendah ...Error! Bookmark not defined.
11. Nilai pengaruh ragam aditif, ragam dominan dan rata-rata peragam aditif dan dominan terhadap karakter agronomi kedelai pada kondisi intensitas cahaya rendah ...Error! Bookmark not defined.
12. Distribusi gen pada tetua dan pengaruh dominansi (h2) terhadap karakter
agronomi kedelai pada kondisi intensitas cahaya rendahError! Bookmark not defined.
resesif dalam tetua (KD/KR) serta jumlah kelompok gen yang memberikan efek dominansi (h2/H2) ...Error! Bookmark not defined.
14. Nilai heritabilitas arti luas dan heritabilitas arti sempit karakter agronomi kedelai pada kondisi intensitas cahaya rendahError! Bookmark not defined.
15. Respon seleksi karakter agronomi kedelai pada kondisi intensitas cahaya rendah ...Error! Bookmark not defined.
16. Hasil pengukuran efektivitas seleksi untuk daya hasil kedelai pada kondisi intensitas cahaya rendah...Error! Bookmark not defined.
17. Sumber keragaman pengaruh genotipe terhadap karakter agronomi kedelai pada kondisi intensitas cahaya rendahError! Bookmark not defined.
18. Keragaan karakter tinggi saat panen, jumlah cabang produktif, jumlah buku total, jumlah polong isi, dan jumlah polong hampa generasi F5 pada kondisi intensitas cahaya penuh...Error! Bookmark not defined.
19. Keragaan karakter jumlah polong isi, jumlah polong hampa, persen polong isi, dan daya hasil Generasi F5 pada kondisi intensitas cahaya penuh ...Error! Bookmark not defined.
20. Nilai parameter genetik kedelai generasi F5 hasil persilangan tetua Ceneng dan Godek ...Error! Bookmark not defined.
21. Hasil uji nilai tengah antara tetua Ceneng dan GodekError! Bookmark not defined.
22. Uji kenormalan shapiro-wilk untuk populasi F6 hasil persilangan Ceneng dan Godek pada kondisi intenistas cahaya rendahError! Bookmark not defined.
23. Rekapitulasi analisis sidik ragam pengaruh genotipe terhadap karakter
agronomi kedelai generasi F7 pada kondisi intensitas cahaya rendahError! Bookmark not defi
24. Nilai ragam fenotipe, ragam lingkungan, ragam genotipe, dan nilai
heritabilitas kedelai generasi F6 pada kondisi intensitas cahaya rendahError! Bookmark not d
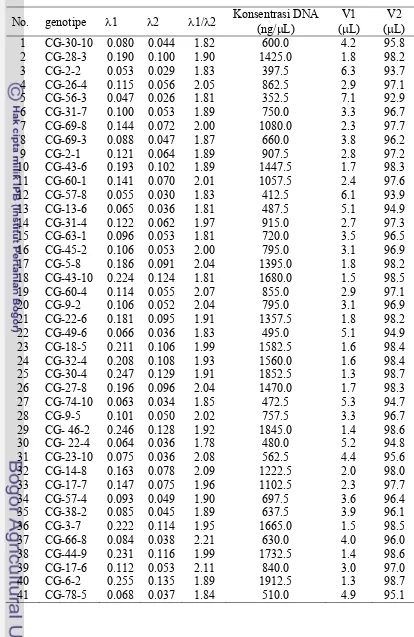
25. Hasil pengujian DNA tetua Ceneng dan Godek menggunakan spekrofotometer...Error! Bookmark not defined.
26. Acuan penyiapan konsentrasi DNA tetua yang diperlukan dalam seleksi primer ...Error! Bookmark not defined.
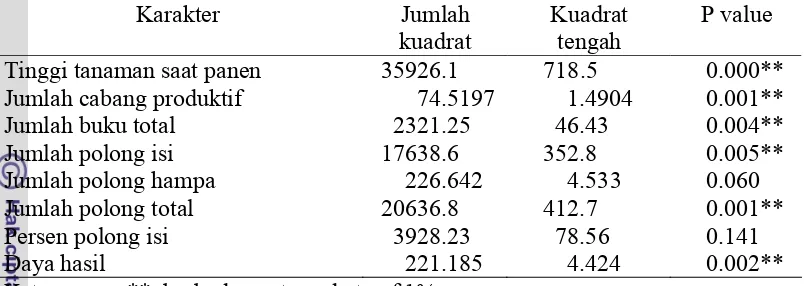
27. Daftar primer dan marka yang digunakan analisis QTL yang
mengendalikan toleransi terhadap intensitas cahaya rendah pada kedelaiError! Bookmark not d
28. Rekapitulasi hasil seleksi primer ...Error! Bookmark not defined.
30. Perbandingan nilai tengah dan nilai diferensial seleksi yang diperoleh dengan menggunakan marker assisted selectionError! Bookmark not defined.
LAMPIRAN
No Halaman
1. Daftar primer yang diseleksi untuk mendapatkan primer yang polimorfik dan terpaut dengan tetua toleran...Error! Bookmark not defined.
DAFTAR GAMBAR
No Halaman
Gambar 1. Bagan alir rencana penelitian ...Error! Bookmark not defined.
Gambar 2. Hubungan ragam (Vr) dan peragam (Wr) serta sebaran array karakter tinggi tanaman saat panen pada kondisi intensitas cahaya rendah ...Error! Bookmark not defined.
Gambar 3. Hubungan ragam (Vr) dan peragam (Wr) serta sebaran array
karakter jumlah cabang total pada kondisi intensitas cahaya rendahError! Bookmark not
Gambar 4. Hubungan ragam (Vr) dan peragam (Wr) serta sebaran array
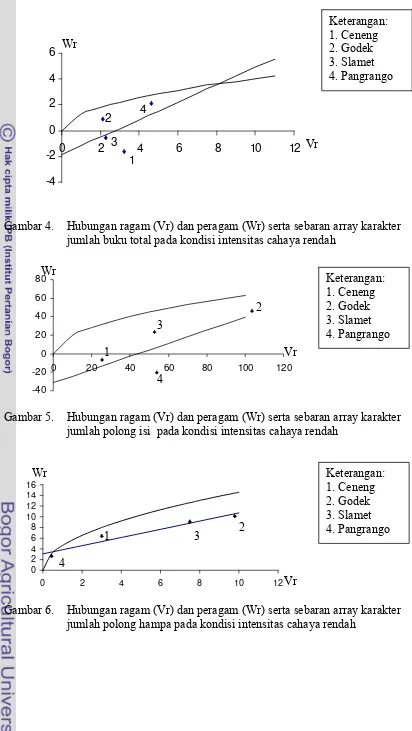
karakter jumlah buku total pada kondisi intensitas cahaya rendahError! Bookmark not de
Gambar 5. Hubungan ragam (Vr) dan peragam (Wr) serta sebaran array
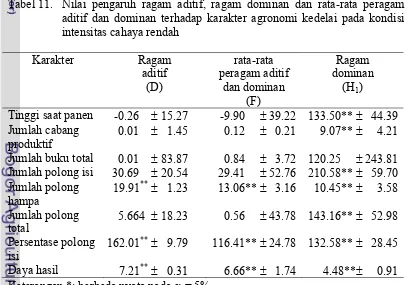
karakter jumlah polong isi pada kondisi intensitas cahaya rendahError! Bookmark not d
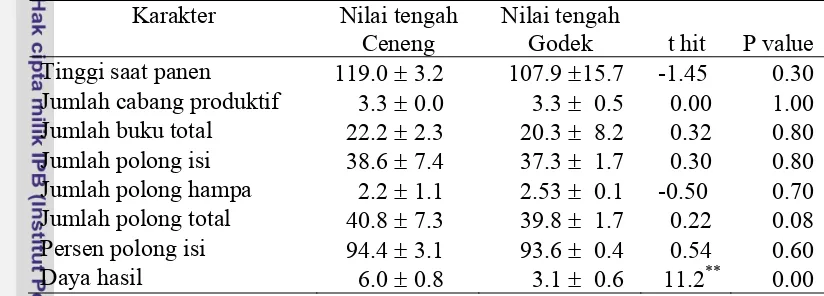
Gambar 6. Hubungan ragam (Vr) dan peragam (Wr) serta sebaran array karakter jumlah polong hampa pada kondisi intensitas cahaya rendah ...Error! Bookmark not defined.
Gambar 7. Hubungan ragam (Vr) dan peragam (Wr) serta sebaran array
karakter jumlah polong total pada kondisi intensitas cahaya rendahError! Bookmark not
Gambar 8. Hubungan ragam (Vr) dan peragam (Wr) serta sebaran array karakter persentase polong isi pada kondisi intensitas cahaya rendah ...Error! Bookmark not defined.
Gambar 9. Hubungan ragam (Vr) dan peragam (Wr) serta sebaran array karakter daya hasil per tanama pada kondisi intensitas cahaya rendah ...Error! Bookmark not defined.
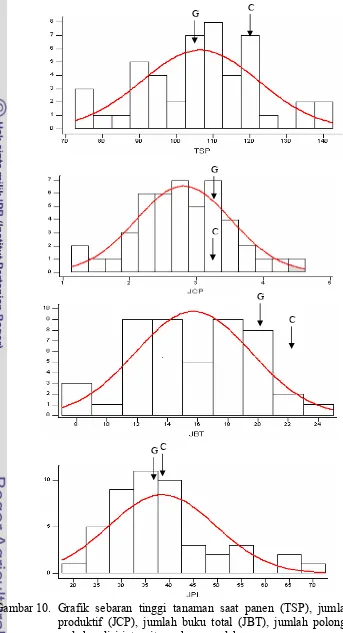
Gambar 10. Grafik sebaran tinggi tanaman saat panen (TSP), jumlah cabang produktif (JCP), jumlah buku total (JBT), jumlah polong isi (JPI) pada kondisi intensitas cahaya rendah.Error! Bookmark not defined.
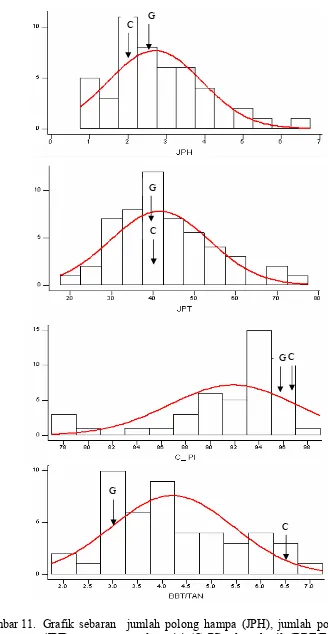
Gambar 11. Grafik sebaran jumlah polong hampa (JPH), jumlah polong total (JPT), persentase polong isi (C_PI), daya hasil (BBT/TAN) pada kondisi intensitas cahaya rendah.Error! Bookmark not defined.
Kedelai adalah komoditi yang bernilai ekonomi tinggi karena merupakan
sumber utama protein nabati bagi bangsa Indonesia (Nugraha et al. 2002).
Permintaan pasar terhadap kedelai terus meningkat sejalan dengan pertambahan
jumlah penduduk yang ditandai dengan peningkatan impor kedelai dari tahun ke
tahun (Balitbang Deptan 2005a). Menurut Balitbang Deptan (2005b), konsumsi
kedelai tahun 2004 adalah 2.015 juta ton, sedangkan produksi kedelai nasional
hanya 0.707 juta ton sehingga Indonesia harus impor kedelai sebanyak 1.307 juta
ton.
Peningkatan produksi kedelai merupakan langkah strategis yang harus
dilakukan untuk memenuhi kebutuhan kedelai nasional melalui perluasan areal
panen dan peningkatan produktivitas. Salah satu alternatif untuk perluasan areal
panen adalah dengan memanfaatkan lahan yang ada di bawah tegakan karet yang
belum menghasilkan sehingga perlu tersedia varietas adaptif dan berdaya hasil
tinggi di bawah tegakan karet. Pengembangan kedelai sebagai tanaman sela di
bawah tegakan tanaman karet sangat potensial mengingat lahan yang tersedia
cukup luas. Menurut Balitbang Deptan (2005a), potensi lahan di bawah tegakan
tanaman perkebunan adalah sebesar 12.1 juta hektar, 3-4 % diantaranya
merupakan areal tanaman baru yang belum menghasilkan (TBM) yang dapat
dimanfaatkan untuk pertanaman kedelai dengan pola tumpang sari.
Kendala utama budidaya kedelai di bawah tegakan karet adalah
berkurangnya intensitas cahaya yang diterima oleh tanaman. Pengurangan
intensitas cahaya dapat mencapai 75% pada tegakan karet yang berumur 4 tahun.
Agar dapat memanfaatkan lahan yang ada di bawah tegakan tanaman perkebunan
maka diperlukan pengembangan varietas yang mampu tumbuh sehingga dapat
berproduksi dengan baik pada kondisi intensitas cahaya rendah (Sopandie et al.
2003a; Sopandie et al. 2003b; Sopandie et al. 2003c).
Peningkatan produktivitas merupakan tujuan utama pemuliaan kedelai yang
dapat dicapai melalui seleksi untuk perbaikan daya hasil. Menurut Wricke dan
untuk perbaikan potensi hasil dapat dilakukan secara langsung terhadap daya hasil
atau tidak langsung melalui karakter yang lain.
Masalah yang dihadapi dalam pengembangan kedelai untuk toleransi
terhadap intensitas cahaya rendah adalah belum ditentukan karakter seleksi yang
tepat bagi perbaikan daya hasil kedelai dalam kondisi intensitas cahaya rendah.
Penentuan karakter seleksi dapat dilakukan berdasarkan pola pewarisan dari
karakter yang ingin diperbaiki. Berdasarkan hasil studi pola pewarisan yang
dilakukan oleh peneliti sebelumnya ternyata belum dapat dipastikan karakter
seleksi yang tepat bagi kedelai toleran intensitas cahaya rendah. Hal ini
disebabkan oleh hasil studi pola pewarisan berbagai karakter kedelai pada kondisi
intensitas cahaya rendah menunjukkan hasil yang berbeda-beda, namun terdapat
kecenderungan bahwa daya hasil mempunyai nilai heritabilitas lebih tinggi
dibandingkan dengan karakter lainnya (Sopandie et al. 2003c; Handayani 2003;
La Muhuria 2007; Kisman 2007).
Berdasarkan informasi yang diperoleh dari hasil penelitian terdahulu maka
terdapat peluang untuk menggunakan karakter komponen hasil maupun hasil
sebagai karakter seleksi. Menurut Austin (1993), seleksi berdasarkan daya hasil
pada lingkungan bercekaman menghadapi kendala karena sulit memisahkan
faktor-faktor yang menyebabkan penurunan daya hasil. Hal ini didukung oleh
Tester dan Bacic (2005) serta Ceccareli et al. (2007) bahwa kesulitan seleksi pada
kondisi bercekaman adalah adanya interaksi genotipe dan lingkungan yang
bersifat kualitatif.
Selain itu, seleksi berdasarkan daya hasil untuk perbaikan produktivitas
kedelai pada kondisi intensitas cahaya rendah harus dilakukan di lingkungan
target. Seleksi untuk perbaikan produktivitas kedelai pada kondisi intensitas
cahaya rendah harus dilakukan di bawah tegakan karet atau lingkungan buatan
yang menyerupai lingkungan di bawah tegakan yaitu di bawah naungan paranet.
Seleksi pada lingkungan target tidak mudah karena sulit mendapatkan lingkungan
yang dapat memberikan tingkat dan waktu cekaman yang sesuai.
Guna mengatasi kendala seleksi perbaikan produktivitas kedelai dalam
kondisi intensitas cahaya rendah maka diperlukan pembentukan marka seleksi
diinginkan adalah marka yang tidak dipengaruhi oleh lingkungan sehingga seleksi
tidak perlu dilakukan di bawah naungan paranet atau tegakan karet. Menurut
Mohan, et al. (1997) serta Farroq dan Azam (2002), salah satu marka yang dapat
dimanfaatkan sebagai marka seleksi adalah marka molekuler yang terpaut dengan
QTL yang mengendalikan daya hasil. Melalui pemanfaatan marka molekuler
sebagai alat bantu seleksi maka sangat memungkinkan melakukan seleksi secara
tidak langsung tanpa dipengaruhi oleh lingkungan dan seleksi tidak perlu
dilakukan di lingkungan target.
Tujuan umum penelitian ini adalah menentukan karakter seleksi untuk
meningkatkan akurasi seleksi sehingga dapat memaksimalkan kemajuan genetik
yang diperoleh dalam pengembangan kedelai toleran intensitas cahaya rendah.
Kegiatan yang dilakukan dalam penelitian ini terdiri dari studi pola pewarisan dan
identifikasi marka RAPD yang terpaut dengan toleransi terhadap intensitas cahaya
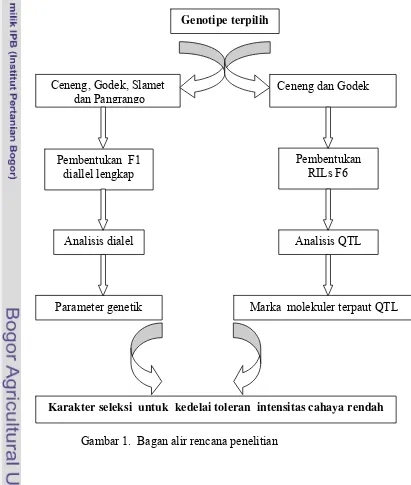
rendah (Gambar 1).
Hasil yang diperoleh dalam penelitian ini menunjukkan bahwa di antara
karakter hasil dan komponen hasil yang dipelajari, ternyata daya hasil dapat
digunakan sebagai karakter seleksi bagi perakitan varietas kedelai toleran
intensitas cahaya rendah karena daya hasil mempunyai nilai heritabilitas arti
sempit yang cukup tinggi. Hasil penelitian ini didukung oleh hasil penelitian
sebelumnya yang menunjukkan bahwa daya hasil mempunyai nilai heritabilitas
arti luas maupun arti sempit lebih tinggi dibandingkan dengan karakter lainnya.
Dalam pengembangan marka seleksi bagi daya hasil tinggi pada kedelai dalam
kondisi intensitas cahaya rendah diperoleh dua marka RAPD yang terpaut
masing-masing dengan QTL yang mengendalikan jumlah buku produktif dan daya hasil.
Hasil penelitian ini sangat bermanfaat untuk merencanakan dan
melaksanakan kegiatan pemuliaan yang efisien untuk mengembangkan kedelai
toleran intensitas cahaya rendah terutama yang berhubungan dengan kegiatan
seleksi. Dengan mengintegrasikan teknik konvensional dengan teknik molekuler
maka kegiatan pemuliaan kedelai toleran intensitas cahaya rendah bisa dipercepat.
Diharapkan kegiatan pemuliaan untuk perakitan varietas kedelai toleran intensitas
Tujuan
Tujuan umum penelitian ini adalah mengembangkan karakter seleksi yang
sesuai bagi kedelai toleran intensitas cahaya rendah sehingga akan
memaksimalkan kemajuan genetik yang diperoleh, namun secara khusus
penelitian ini bertujuan untuk:
1. Memperoleh informasi tentang pola pewarisan karakter agronomi kedelai pada
kondisi intensitas cahaya rendah
2. Mengidentifikasi QTL yang mengendalikan karakter agronomi kedelai pada
kondisi intensitas cahaya rendah.
Gambar 1. Bagan alir rencana penelitian Ceneng, Godek, Slamet
dan Pangrango
Analisis dialel
Parameter genetik
Analisis QTL
Karakter seleksi untuk kedelai toleran intensitas cahaya rendah Genotipe terpilih
Marka molekuler terpaut QTL Pembentukan
RILs F6 Pembentukan F1
diallel lengkap
TINJAUAN PUSTAKA
Arah Pengembangan Kedelai
Tujuan pemuliaan adalah meningkatkan nilai ekonomi tanaman melalui
peningkatan produktivitas atau kualitas produk yang dihasilkan sehingga menjadi
lebih bermanfaat bagi kehidupan manusia. Seringkali produktivitas tinggi tidak
cukup untuk meningkatkan peneriman petani atau konsumen terhadap varietas
yang dihasilkan sehingga peningkatan produktivitas harus diikuti dengan
peningkatan kualitas seperti komposisi gizi atau bentuk dan warna (Chahal dan
Gosal 2003).
Prioritas arah pemuliaan kedelai di Indonesia pada masa yang akan datang
adalah peningkatan produktivitas tanaman untuk mendukung pengembangan
kedelai yang sesuai bagi agroekosistem lahan kering beriklim basah dan lahan
kering beriklim sedang (Sumarno et al. 2000; Arsyad 2000). Menurut Pinem
(2000), pengembangan kedelai diarahkan untuk peningkatan produktivitas lahan
beririgasi intensif, lahan kering bereaksi masam, lahan gambut, lahan pasang
surut, serta lahan perkebunan rakyat, lahan perkebunan swasta atau BUMN, dan
lahan hutan sosial.
Menurut Balitbang Deptan (2005b), peningkatan produksi kedelai dilakukan
melalui peningkatan produktivitas dan perluasan areal panen. Peningkatan
produktivitas dapat dicapai dengan pemanfaatan varietas unggul berdaya hasil
tinggi terutama diarahkan untuk daerah yang telah menjadi sentra produksi
kedelai, tetapi produktivitas masih rendah seperti Jawa Timur, Jawa Tengah,
NTB, Jawa Barat, Lampung, Sumatra Utara, dan Sulawesi Selatan.
Potensi lahan untuk pengembangan kedelai adalah 1.7 juta hektar berupa
lahan sawah, lahan kering, lahan pasang surut, lahan di bawah tegakan tanaman
perkebunan dan lahan bukaan baru (Balitbang Deptan 2005b. Lahan di bawah
tegakan perkebunan yang belum menghasilkan adalah salah satu lahan yang
berpotensi untuk perluasan areal tanam kedelai melalui pola tumpangsari
(Sopandie et al. 2003c). Menurut Balitbang Deptan (2005a), potensi lahan di
bawah tegakan tanaman perkebunan adalah sebesar 12.1 juta hektar, 3-4 % di
antaranya merupakan areal tanaman baru yang belum menghasilkan (TBM) yang
Kendala yang dihadapi dalam pemanfaatan lahan di bawah tegakan tanaman
perkebunan adalah rendahnya intensitas cahaya yang diterima oleh tanaman
sehingga mengganggu pertumbuhan dan produksi tanaman. Pemanfaatan lahan
yang ada di bawah tegakan perkebunan perlu didukung oleh ketersediaan varietas
yang toleran dan berdaya hasil baik pada kondisi di bawah tegakan tanaman
perkebunan (Sopandie et al. 2003c; Sopandie et al. 2006; Sopandie 2006).
Pertumbuhan dan Produksi Kedelai pada Kondisi Intensitas Cahaya Rendah
Cahaya matahari mempunyai peran penting dalam proses fisiologi tanaman
seperti fotosintesis, menutup dan membukanya stomata, pertumbuhan dan
perkembangan, serta perkecambahan tanaman. Ketersediaan cahaya matahari
sangat menentukan tingkat produksi tanaman. Tanaman menangkap cahaya
matahari yang selanjutnya digunakan dalam proses fotosintesis sehingga intensitas
cahaya matahari merupakan salah faktor yang membatasi proses fotosintesis
(Levitt 1980; Taiz dan Zeiger 1991; Salisbury dan Ross 1992).
Kebutuhan cahaya bagi tanaman kedelai untuk mencapai fotosintesis
maksimal adalah berkisar antara 0.3-0.8 kal/cm2/menit atau setara dengan 432-1152 kal/cm2/hari (Salisbury dan Ross 1992). Nilai rata-rata intensitas cahaya matahari pada areal terbuka adalah 398.4 kal/cm2/hari, sedangkan nilai rata-rata intensitas cahaya matahari di bawah tegakan karet berumur 1, 2, 3 dan 4 tahun
masing-masing adalah 326.7 kal/cm2/hari, 237.6 kal/cm2/hari dan 109.2 kal/cm2/hari, dan 38.2 kal/cm2/hari (Sopandie et al. 2002). Nilai intensitas cahaya di bawah tegakan karet umur 2 tahun setara dengan intensitas cahaya di bawah
naungan paranet 25% dan umur 3 tahun setara dengan intensitas cahaya di bawah
paranet 50%, sedangkan umur 4 tahun sudah kurang dari intensitas cahaya di
bawah naungan paranet 75% (Chozin et al. 1998; Chozin et al. 1999).
Penurunan intensitas cahaya akan mempengaruhi pertumbuhan dan hasil
tanaman. Beberapa hasil penelitian tentang pengaruh intensitas cahaya matahari
menunjukkan bahwa penurunan intensitas cahaya mengakibatkan gangguan
pertumbuhan dan penurunan hasil (Daubenmire 1974; Baharsjah 1980; Anderson
akan memberikan respon terhadap intensitas cahaya rendah. Hasil penelitian
Baharsjah (1980), Asadi et al. (1997), Elfarisna (2000), Khumaida (2002),
Handayani (2003), La Muhuria (2007), dan Kisman (2007) menunjukkan bahwa
pada kedelai terjadi perubahan karakter agronomi, morfologi, anatomi, fisiologi,
dan molekuler sebagai akibat penurunan intensitas cahaya.
Perubahan morfologi dan agronomi kedelai akibat penurunan intensitas
cahaya sampai 40% setelah perkecambahan mengakibatkan penurunan jumlah
buku, jumlah cabang, diameter batang, jumlah polong, serta hasil biji pada
tanaman kedelai. Penurunan intensitas cahaya menjadi 40% sejak pengisian
polong menyebabkan penurunan jumlah polong, hasil biji, dan kadar protein
kedelai (Baharsjah 1980). Hasil pengujian pada 28 galur kedelai di bawah
naungan paranet 33% menunjukkan penurunan daya hasil berkisar antara 2-45%
dibandingkan dengan hasil kondisi tanpa naungan atau intensitas cahaya penuh
(Asadi et al. 1997). Intensitas cahaya rendah menyebabkan peningkatan tinggi
tanaman serta penurunan jumlah cabang, jumlah buku, jumlah polong isi dan daya
hasil (Handayani 2003). Sopandie et al (2002) melaporkan bahwa kedelai yang
ditanam di bawah naungan paranet 50% mengalami penurunan hasil biji sampai
60% dibandingkan dengan kondisi tanpa naungan (Sopandie et al. 2002,
Sopandie et al. 2003c).
Perubahan anatomi meliputi peningkatan luas daun, kandungan klorofil a,
kandungan klorofil b, namun terjadi penurunan kerapatan trikoma, tebal daun,
lapisan palisade dan rasio klorofil a dan b (Elfarisna 2000; Handayani 2003; La
Muhuria 2007; Kisman 2007). Perubahan fisiologi meliputi penurunan aktivitas
berbagai enzim yang berhubungan dengan fotosintesis seperti ribulaose
biphosphate carboxylase/oxygenase (Rubisco), sucrose phosphate synthase (SPS),
malatedehydrogenase (MDH), dan asam invertase (AI). Penurunan intensitas
cahaya juga mengakibatkan penurunan laju fotosintesis maksimum dan laju
transpor elektron maksimum, laju respirasi gelap dan titik kompensasi cahaya
serta penurunan kandungan sukrosa dan pati (La Muhuria 2007).
Perubahan molekuler berhubungan dengan peningkatan ekspresi gen-gen
yang mengendalikan fotosíntesis kedelai dalam kondisi intensitas cahaya rendah.
(light harvesting complex binding protein) dan gen Rubisco meningkat sejalan
dengan bertambah lamanya perlakuan naungan meskipun kedua gen ini belum
dapat membedakan genotipe toleran dengan genotipe peka terhadap intensitas
cahaya rendah. Khumaida (2002) juga melaporkan adanya kandidat gen-gen
fotosíntesis pada kedelai seperti gen JJ3, CAB-3, phyB dan ATHB-2. Hasil
analisis ekspresi kandidat gen ini yang dilakukan oleh Kisman (2007)
menunjukkan bahwa pada genotipe kedelai yang toleran intensitas cahaya rendah,
kandidat gen JJ3, phyB dan ATHB-2 terekspresi lebih kuat dibandingkan dengan
genotipe peka.
Perubahan-perubahan spesifik yang terjadi pada kedelai dalam kondisi
intensitas cahaya rendah pada berbagai tingkatan merupakan bentuk adaptasi
kedelai terhadap intensitas cahaya rendah. Secara umum kemampuan tanaman
untuk tumbuh dan berproduksi dengan baik pada kondisi intensitas cahaya rendah
tergantung pada kemampuan tanaman untuk beradaptasi pada kondisi intensitas
cahaya rendah. Adaptasi tanaman terhadap intensitas cahaya rendah ditentukan
oleh kemampuan tanaman untuk melakukan fotosintesis pada kondisi defisit
cahaya (Sopandie 2006).
Menurut Hale dan Orcut (1987), adaptasi tanaman terhadap intensitas
cahaya rendah pada dasarnya dilakukan dengan dua cara melalui peningkatan luas
daun untuk meningkatkan jumlah cahaya yang ditangkap dan mengurangi jumlah
cahaya yang ditransmisikan dan direfleksikan. Levitt (1980) menyatakan bahwa
adaptasi tanaman terhadap intensitas cahaya rendah dicapai melalui mekanisme
penghindaran (avoidance), berkaitan dengan perubahan anatomi dan morfologi
yang dilakukan oleh daun guna meningkatkan efisiensi fotosintesis dan
mekanisme toleran, berkaitan dengan penurunan titik kompensasi cahaya dan
respirasi.
Mekanisme penghindaran dilakukan melalui peningkatan luas daun untuk
meningkatkan jumlah cahaya yang ditangkap serta mengurangi jumlah cahaya
yang ditransmisikan dan refleksikan sehingga jumlah cahaya yang digunakan
oleh tanaman untuk melakukan fotosintesis meningkat. Struktur anatomi yang
mendukung mekanisme penghindaranadalah kutikula, lapisan lilin dan bulu daun
Mekanisme toleransi tanaman terhadap intensitas cahaya rendah
berhubungan dengan penurunan laju respirasi dan titik kompensasi cahaya.
Titik kompensasi cahaya adalah kondisi yang menunjukkan laju asimilasi CO2
sama dengan laju evolusi O2 dalam proses respirasi. Tanaman yang toleran
terhadap intensitas cahaya rendah akan menurunkan titik kompensasi cahaya dan
laju respirasi sehingga produk fotosintesis dapat terakumulasi (Levitt 1980).
Mekanisme penghindaran pada kedelai merupakan mekanisme untuk
meningkatkan jumlah cahaya yang ditangkap meliputi peningkatan luas daun,
pengurangan ketebalan daun, pengurangan kepadatan trikoma dan peningkatan
kandungan pigmen fotosintesis. Sampai saat ini mekanisme untuk mengurangi
cahaya yang ditransmisikan dan direfleksikan belum dilaporkan pada kedelai
(Khumaida 2002; Sopandie et al. 2002; La Muhuria 2007; Kisman 2007).
Mekanisme toleransi terhadap intensitas cahaya rendah pada kedelai
meliputi peningkatan laju fotosintesis maksimum serta penurunan laju respirasi
gelap dan titik kompensasi cahaya. Dalam kondisi intensitas cahaya rendah
genotipe toleran memiliki laju fotosintesis maksimum lebih tinggi serta laju
respirasi gelap dan titik kompensasi cahaya lebih rendah. Peningkatan laju
fotosintesis serta penurunan laju respirasi gelap dan titik kompensasi cahaya
ditunjukkan oleh kandungan sukrosa dan pati yang lebih tinggi pada genotipe
toleran dibandingkan genotipe dalam kondisi intensitas cahaya rendah (La
Muhuria 2007).
Berdasarkan hasil studi fisiologi terhadap respon dan mekanisme adaptasi
kedelai terhadap intensitas cahaya rendah sangat memungkinkan melakukan
perbaikan daya hasil kedelai pada kondisi intensitas cahaya rendah. Perbaikan
tanaman untuk meningkatkan produktivitas di lahan marjinal dapat diupayakan
melalui perbaikan adaptasi terhadap cekaman abiotik untuk mencegah penurunan
hasil (Sopandie 2006).
Pemuliaan bagi Lingkungan Bercekaman
Lingkungan bercekaman adalah lingkungan suboptimum bagi pertumbuhan
dan produksi tanaman. Lingkungan suboptimum merupakan lingkungan yang
produktivitas tanaman pada lingkungan bercekaman ditentukan oleh tingkat
cekaman pada lingkungan target (Sopandie et al. 2004; Sopandie 2006).
Perbaikan produktivitas tanaman di lahan bercekaman dapat dilakukan melalui
perbaikan potensi hasil untuk mendapatkan varietas berdaya hasil tinggi dan
perbaikan daya adaptasi tanaman untuk mendapatkan varietas yang toleran
(Accevedo dan Fereres 1993). Menurut Baker (1993), produktivitas tanaman
pada kondisi bercekaman ditentukan oleh potensi hasil tanaman serta daya
adaptasi tanaman terhadap cekaman.
Perbaikan potensi hasil merupakan upaya untuk meningkatkan produktivitas
tanaman pada kondisi cekaman ringan atau sedang. Adanya interaksi genotipe
dengan lingkungan dalam kondisi cekaman ringan atau sedang tidak
menyebabkan perubahan rangking genotipe. Perbaikan potensi hasil dapat
dilakukan melalui perbaikan kapasitas fotosintesis dan respirasi untuk
meningkatkan biomassa tanaman dan perbaikan sink capacity dan partisi
fotosintat (Accevedo dan Fereres 1993).
Upaya lain untuk perbaikan produktivitas tanaman pada tingkat cekaman
ringan adalah pembentukan idiotype breeding (Romagosa dan Fox 1993).
Idiotype breeding adalah perbaikan tanaman dengan mengembangkan kombinasi
karakter yang mendukung fotosintesis, pertumbuhan dan produksi tanaman.
Keuntungan dari idiotype breeding adalah pemulia akan mempunyai gambaran
yang jelas dalam menyeleksi karakter tanaman yang mendukung peningkatan
potensi hasil (Romagosa dan Fox 1993;Sopandie 2006).
Perbaikan adaptasi tanaman merupakan upaya untuk perbaikan produktivitas
tanaman pada kondisi cekaman berat (Romagosa dan Fox 1993). Kendala yang
dihadapi dalam perbaikan produktivitas tanaman untuk lingkungan bercekaman
berat adalah adanya interaksi genotipe dengan lingkungan. Interaksi genotipe
dengan lingkungan terjadi jika genotipe memperlihatkan respon yang berbeda
pada kondisi lingkungan yang berbeda.
Interaksi genotipe dengan lingkungan dikelompokkan menjadi dua yaitu
interaksi yang bersifat kuantitatif dan interaksi yang bersifat kualitatif. Interaksi
genotipe dengan lingkungan yang bersifat kuantitatif tidak menyebabkan
unggul pada lingkungan yang berbeda. Interaksi genotipe lingkungan yang
bersifat kualitatif merupakan kendala dalam pemuliaan bagi lingkungan
bercekaman berat karena mengakibatkan perubahan rangking genotipe. Genotipe
berdaya hasil tinggi pada satu lingkungan bisa mengalami penurunan hasil yang
cukup tajam pada lingkungan berbeda akibat pengaruh cekaman (Romagosa dan
Fox 1993; Roy 2000; Bernardo 2002; Chahal dan Gosal 2003).
Menurut Ceccareli (1996), interaksi genotipe dengan lingkungan yang
bersifat kualitatif merupakan salah satu kesulitan dalam pemuliaan untuk
lingkungan bercekaman terutama pada saat melakukan seleksi. Menurut
Ceccareli et al. (2007), seleksi pada lingkungan bercekaman harus dilakukan di
lingkungan target sehingga dapat memaksimalkan ekspresi gen-gen yang
mengendalikan daya hasil maupun daya adaptasi tanaman terhadap cekaman.
Interaksi genotipe lingkungan menyebabkan seleksi untuk perbaikan daya
hasil pada kondisi bercekaman tidak mudah karena daya hasil sangat dipengaruhi
oleh lingkungan sehingga daya hasil mempunyai nilai heritabilitas yang rendah
(Ceccareli 1994). Agar seleksi menjadi lebih efisien maka diperlukan penetapan karakter seleksi yang tepat untuk memilih genotipe yang dapat mempertahankan
daya hasil pada lingkungan bercekaman.
Seleksi pada kondisi bercekaman dapat dilakukan berdasarkan fenotipe,
marka molekular, dan gabungan antara fenotipe dan marka molekuler (Bernardo
2002). Seleksi berdasarkan fenotipe menjadi sulit karena nilai heritabilitas yang
rendah dan adanya interaksi antara genotipe dan lingkungan yang bersifat
kualitatif. Untuk meningkatkan efisiensi seleksi maka seleksi dapat dilakukan
menggunakan marka molekuler. Marka molekuler yang terpaut dengan QTL atau
yang mengendalikan daya hasil pada kondisi bercekaman serta QTL yang
mengendalikan toleransi terhadap cekaman merupakan salah satu marka yang
dapat dijadikan alat bantu seleksi (Forster et al. 2000; Hussain 2006).
Perakitan Varietas Kedelai Toleran Intensitas Cahaya Rendah
Upaya peningkatan produksi kedelai sebagai tanaman sela di bawah tegakan
perkebunan menghadapi kendala yaitu rendahnya intensitas cahaya akibat
adaptif dan berproduksi tinggi pada kondisi intensitas cahaya rendah (naungan).
Upaya pengembangan kedelai toleran intensitas cahaya rendah telah berhasil
dilakukan, tetapi masih terbatas pada tingkat cekaman ringan (naungan 33%)
yaitu pola tumpangsari kedelai dengan jagung (Asadi et al. 1997). Upaya
pengembangan kedelai toleran intensitas cahaya rendah dengan tingkat cekaman
berat yaitu dalam pola tumpangsari dengan tanaman perkebunan telah dilakukan
oleh Sopandie et al. (2002), Sopandie et al. (2003c) serta Sopandie et al. (2006).
Pengembangan kedelai toleran intensitas cahaya rendah dimulai dengan
melakukan karakterisasi pada plasma nutfah yang tersedia untuk mendapatkan
informasi tentang keragaman dari karakter yang akan diperbaiki. Menurut
Makmur (1992), keberhasilan program pemuliaan sangat ditentukan oleh
ketersediaan keragaman genetik.
Asadi et al. (1997) melaporkan bahwa terdapat keragaman yang cukup
tinggi dalam plasma nutfah kedelai ketika ditanam pada kondisi naungan ringan
yaitu pola tumpangsari dengan jagung. Sampai saat ini varietas Pangrango adalah
salah satu varietas kedelai yang toleran untuk kondisi naungan ringan (naungan
33%) yaitu pola tumpangsari dengan jagung.
Sopandie et al. (2002) juga melaporkan bahwa terdapatnya perbedaan
respon antara berbagai genotipe kedelai yang menunjukkan adanya keragaman
genotipe kedelai pada kondisi intensitas cahaya rendah. Dalam kondisi naungan
berat terdapat beberapa genotipe seperti Godek A dan Klungkung mengalami
penurunan hasil sampai 50%, sedangkan genotipe Wilis hanya mengalami
penurunan hasil sekitar 20%. Genotipe Ceneng memiliki daya hasil yang lebih
tinggi dibandingkan dengan genotipe lainnya dan tergolong genotipe yang
mempunyai daya adaptasi baik karena memiliki daya hasil lebih tinggi
dibandingkan dengan daya hasil pada kondisi intensitas cahaya penuh.
Selain oleh ketersediaan keragaman genetik maka keberhasilan pemuliaan
juga ditentukan oleh pemilihan metode seleksi yang tepat. Pemilihan metode
seleksi dilakukan berdasarkan sistem penyerbukan tanaman dan pola pewarisan
dari karakter yang akan diperbaiki. Berdasarkan informasi tentang pola pewarisan
maka dapat juga ditentukan karakter seleksi yang tepat (Roy, 2000; Chahal dan
memilih metode dan karakter seleksi yang tepat maka perencanan dan
pelaksanaan program pemuliaan akan menjadi lebih efisien.
Menurut Chahal dan Ghosal (2003), beberapa teori dasar tentang seleksi
adalah seleksi hanya efektif pada karakter yang dikendalikan oleh faktor genetik.
Seleksi dilakukan untuk memilih genotipe, bukan untuk memilih gen-gen yang
diinginkan. Seleksi akan menghasilkan kombinasi gen-gen yang baru bukan
menghasilkan gen-gen baru.
Secara umum pemuliaan tanaman menyerbuk sendiri diarahkan untuk
membentuk galur murni. Dasar genetik pembentukan galur murni pada tanaman
menyerbuk sendiri adalah bahwa penyerbukan sendiri pada tanaman homozigot
akan menghasilkan genotipe homozigot dan segregasi pada tanaman heterozigot
akan meningkatkan proporsi genotipe homozigot dan menurunkan proporsi
heterozigot (Chalal dan Gosal 2003).
Metode seleksi yang digunakan untuk tanaman menyerbuk sendiri seperti
kedelai adalah pedigree, bulk, silang balik, single seed descent dan back cross.
Berdasarkan pola pewarisannya maka metode pedigree banyak digunakan untuk
seleksi pada karakter kualitatif atau karakter kuantitatif yang mempunyai nilai
heritabilitas yang tergolong tinggi sehingga seleksi dapat dilakukan pada generasi
awal (Moreno-Gonzales dan Cubero 1993; Roy 2000; Chahal dan Gosal, 2003).
Metode silang balik diterapkan dengan tujuan memasukkan satu atau dua
karakter pada varietas atau genotipe yang sudah mempunyai daya adaptasi atau
karakter agronomi yang sudah baik. Metode bulk dan single seed descent umum
digunakan untuk seleksi pada karakter kuantitatif atau karakter yang mempunyai
nilai heritabilitas yang tergolong rendah atau sedang sehingga seleksi dilakukan
pada generasi lanjut yaitu F5 atau F6. Dalam metode single seed descent hanya
satu benih yang diambil secara acak dari setiap tanaman untuk ditanam pada
generasi berikutnya (Poespodarsono 1988; Stoskopf et al. 1993; Roy 2000;
Chalal dan Ghosal 2003 ).
Di samping untuk menentukan metode seleksi, maka informasi tentang pola
pewarisan sangat diperlukan dalam memilih karakter seleksi karena tercapainya
tujuan pemuliaan ditentukan metode dan karakter seleksi yang digunakan. Seleksi
mendapatkan genotipe yang membawa gen-gen yang mengendalikan karakter
yang diinginkan sehingga seleksi mengakibatkan peningkatan frekuensi gen pada
generasi berikutnya. Seleksi akan efektif jika dilakukan pada populasi yang
memiliki keragaman genetik (Roy 2000).
Berbagai hasil penelitian tentang pola pewarisan kedelai pada kondisi
intensitas cahaya rendah telah dilaporkan oleh Wijayanti (2002), Rostini et al.
(2002), Wibowo (2002), Handayani (2003), La Muhuria (2007), dan Kisman
(2007). Hasil penelitian Wijayanti (2002) dan Wibowo (2002) menyimpulkan
bahwa tidak terdapat pengaruh tetua betina dalam pewarisan beberapa karakter
agronomi seperti tinggi tanaman, jumlah polong hampa dan persentase polong isi.
Hal yang sama juga dilaporkan pada kedelai bahwa tidak terdapat pengaruh tetua
betina dalam pewarisan karakter sifat kandungan klorofil (Rostini et al. 2002;
Handayani 2003). La Muhuria (2007) melaporkan bahwa tidak terdapat pengaruh
tetua betina dalam pewarisan karakter daya hasil pada kedelai pada kondisi
intensitas cahaya rendah. Dengan demikian dalam kondisi intensitas cahaya
rendah tidak terdapat pengaruh tetua betina dalam pewarisan karakter kandungan
klorofil dan daya hasil. Hal ini menunjukkan bahwa karakter tersebut
dikendalikan oleh gen-gen yang terdapat di inti.
Hasil penelitian La Muhuria (2007) pada karakter anatomi serta agronomi
menunjukkan bahwa karakter kerapatan trikoma, klorofil a, dan klorofil b
diwariskan dengan nilai heritabilitas arti sempit lebih rendah dibandingkan
dengan karakter daya hasil. Kisman (2007) menyatakan bahwa karakter daya
hasil diwariskan dengan nilai heritabilitas yang lebih tinggi dibandingkan dengan
karakter luas daun.
Berbagai karakter morfologi dan agronomi kedelai pada kondisi intensitas
cahaya rendah diwariskan dengan nilai heritabilitas arti luas tergolong rendah
sampai sedang. Karakter agronomi kedelai seperti tinggi tanaman, jumlah cabang,
jumlah polong, persentase polong isi dan hasil pada populasi persilangan antara
tetua Pangrango dan Ceneng dalam kondisi intensitas cahaya rendah memiliki
nilai heritabilitas yang rendah yaitu berkisar antara 0.21-0.34 (Wibowo 2002).
Handayani (2003) melaporkan nilai heritabilitas kandungan klorofil dan jumlah
melaporkan bahwa karakter hasil dan komponen hasil diwariskan dengan nilai
heritabilitas antara 2.57 sampai 55.56%.
Adanya keragaman pada karakter morfologi dan agronomi kedelai memberi
peluang untuk menggunakan karakter morfologi dan agronomi sebagai krtiteria
seleksi. Nilai heritabilitas daya hasil yang lebih tinggi memungkinkan
menggunakan daya hasil sebagai karakter seleksi untuk perbaikan potensi hasil
kedelai pada kondisi intensitas cahaya rendah. Sampai sejauh ini informasi yang
lebih lengkap tentang dasar genetik dan pola pewarisan sifat karakter morfologi
dan agronomi pada kondisi intensitas cahaya rendah belum banyak dilaporkan.
Hal ini mennyebabkan belum ada metode pemuliaan yang tepat untuk
mengembangkan kedelai toleran intensitas cahaya rendah.
Pemilihan Karakter Seleksi
Selain menentukan metode seleksi yang tepat, keberhasilan program
pemuliaan dapat dipercepat dengan pemilihan karakter seleksi yang tepat. Seleksi
dapat dilakukan secara langsung atau tidak langsung. Seleksi langsung hanya
efisien jika karakter yang ingin diperbaiki mempunyai nilai heritabilitas yang
tinggi. Namun jika karakter yang ingin diperbaiki mempunyai nilai heritabilitas
yang rendah maka seleksi tidak langsung menggunakan satu atau beberapa
karakter akan lebih efisien.
Pemilihan karakter yang akan digunakan sebagai karakter seleksi tidak
langsung memerlukan informasi tentang pola pewarisan dan keeratan
hubungannya dengan karakter yang ingin diperbaiki dengan daya hasil. Pola
pewarisan dapat diketahui dengan mempelajari aksi gen yang mengendalikan
karakter yang ingin diperbaiki. Aksi gen tersebut diduga berdasarkan model
genetik yang menyertakan parameter aksi gen dan interaksinya. Besarnya
pengaruh gen dan lingkungan terhadap suatu karakter diduga berdasarkan nilai
heritabilitas (Trikoesoemaningtyas 2002).
Heritabilitas arti luas adalah proporsi ragam genetik total yang
mempengaruhi keragaman fenotipe. Ragam genetik terdiri dari ragam aditif,
ragam dominan, dan ragam epistasis yang masing-masing disebabkan oleh aksi
yang disebabkan oleh alel yang terdapat dalam lokus, ragam dominan adalah
ragam yang disebabkan oleh interaksi antar alel dalam satu lokus, sedangkan
ragam epistasis adalah ragam yang disebabkan oleh interaksi antar alel dari lokus
yang berbeda (Falconer dan Mackay 1996).
Terdapat dua tipe heritabilitas yaitu heritabilitas arti luas dan heritabilitas
arti sempit. Heritabilitas arti luas merupakan proporsi ragam genetik total
terhadap ragam fenotipe, sedangkan heritabilitas arti sempit merupakan proporsi
ragam aditif terhadap ragam fenotipe. Nilai heritabilitas dapat diduga secara
langsung melalui pendugaan komponen ragam serta secara tidak langsung melalui
regresi antara tetua dengan turunannya dan respon seleksi. Pendugaan komponen
ragam dapat dilakukan dengan menggunakan populasi dari berbagai rancangan
persilangan (Roy 2000).
Salah satu rancangan persilangan yang dapat digunakan adalah persilangan
diallel. Persilangan diallel adalah semua kemungkinan kombinasi persilangan di
antara beberapa genotipe (Hayman 1954; Roy 2000). Dengan membuat
persilangan diallel maka sangat memungkinkan melakukan analisis genetik yang
lebih luas untuk menduga nilai parameter genetik yang meliputi komponen ragam,
aksi gen, jumlah gen dan nilai heritabilitas yang mengendalikan suatu karakter
berdasarkan pendekatan yang dikembangkan oleh Hayman. Disamping itu
persilangan diallel memungkinkan juga analisa daya gabung antar tetua baik daya
gabung umum maupun daya gabung khusus menggunakan rumus yang
dikembangkan oleh Griffing (Griffing 1956). Asumsi yang harus dipenuhi dalam
analisis diallel adalah tetua homozigot, segregasi merupakan segregasi diploid,
FI dan resiproknya tidak berbeda atau tidak ada efek maternal, tidak terdapat
interaksi antara gen dari alel yang berbeda (epistasis), tidak ada peristiwa
multialel, dan gen-gen tersebar secara merata di antara tetua (Hayman 1954;
Griffing 1956; Singh dan Choudhary 1976).
Pemilihan metode seleksi dan karakter seleksi yang tepat merupakan kunci
keberhasilan pemuliaan tanaman. Informasi yang diperoleh dari analisis silang
diallel dapat digunakan untuk menentukan metode dan karakter seleksi.
Informasi penting yang diperoleh dalam analisis diallel adalah nilai heritabilitas
digunakan untuk populasi yang diduga, sedangkan nilai heritabilitas arti sempit
dapat digunakan untuk seleksi pada populasi progeninya. Dengan demikian
hanya nilai heritabilitas arti sempit yang dapat digunakan untuk menduga
kemajuan genetik yang diperoleh pada progeninya (Griffing 1956).
Dibandingkan dengan rancangan persilangan yang lain, maka disain
persilangan diallel dapat diterapkan lebih luas karena dalam persilangan diallel
memungkinkan terjadinya semua kombinasi persilangan seperti yang terjadi pada
populasi kawin acak di alam. Dengan demikian informasi yang diperoleh dapat
diterapkan untuk berbagai genotipe, tidak terbatas pada genotipe yang dianalisis.
Selain informasi yang diperoleh lebih banyak dan penerapannya juga lebih luas,
keuntungan lain menggunakan analisis silang diallel adalah parameter genetik
dapat diduga pada generasi awal. Kelemahan analisis silang diallel pada tanaman
menyerbuk sendiri adalah jumlah genotipe yang dapat digunakan sangat
tergantung pada kemampuan melakukan hibridisasi (Hayman 1954; Roy 2000;
Saleem et al. 2005).
Beberapa hasil penelitian tentang analisis silang diallel pada kedelai untuk
karakter kandungan nutrisi maupun ketahanan terhadap cekaman biotik seperti
penyakit sudah banyak dilaporkan (Cho dan Scott 2000; Tukamuhabwa et al.
2002; Fronza et al. 2004; Gesteira et al. 2003), namun ketahanan terhadap
cekaman abiotik, khususnya intensitas cahaya rendah belum banyak dilaporkan.
Produksi dan vigor benih pada kedelai dikendalikan oleh aksi gen aditif dan
non aditif. Pengaruh ragam aditif lebih besar dari ragam non aditif terhadap
produksi dan vigor benih (Pantalone et al. 1996; Cho dan Scott 2000).
Tukamuhabwa et al. (2002) melaporkan hasil analisis silang diallel dengan
menggunakan populasi setengah diallel untuk mempelajari karakter pecah polong
pada kedelai. Hasilnya menunjukkan bahwa terdapat interaksi antar lokus
(epistasis) pada gen yang mengendalikan karakter pecah polong. Disamping itu
disimpulkan juga bahwa gen yang mengendalikan karakter pecah polong bersifat
dominan parsial.
Ketahanan terhadap penyakit akar merah yang disebabkan oleh Fusarium
solani f.sp. glycines bersifat over dominance serta dikendalikan paling tidak oleh
penyakit akar merah berkisar antara 48.0-62.0%, sedangkan heritabilitas arti luas
berkisar antara 90.0-95.0% (Vello dan Luis 2004).
Analisis diallel dilakukan berdasarkan dua pendekatan yaitu pendekatan
Griffing dan pendekatan Hayman (Hayman 1954; Griffing 1956). Dalam analisis diallel aksi gen yang mengendalikan suatu karakter diduga dari model genetik
aditif dominan serta ada atau tidak epistasis, tetapi jenis interaksi tidak dapat
diketahui. Jika dibandingkan dengan pendekatan Griffing, maka pendekatan
Hayman lebih menguntungkan dalam pendugaan parameter genetik karena
terdapat uji validitas model genetik yang diajukan. Pendekatan Griffing lebih
banyak digunakan untuk menduga daya gabung umum dan daya gabung khusus
di antara tetua.
Informasi yang diperoleh dari hasil analisis diallel sangat berguna untuk
memilih satu atau beberapa karakter untuk dijadikan karakter seleksi. Nilai
parameter genetik yang diperhatikan dalam memilih karakter seleksi adalah nilai
komponen ragam dan heritabilitas. Karakter seleksi yang baik untuk seleksi
progeni adalah karakter yang lebih dipengaruhi oleh ragam aditif atau yang
mempunyai nilai heritabilitas arti sempit yang tergolong tinggi. Setelah karakter
seleksi ditetapkan maka dapat diduga kemajuan seleksi atau respon seleksi dari
karakter yang diinginkan (Falconer dan Mackay 1996).
Keberhasilan seleksi dengan menggunakan karakter kuantitatif diduga
dengan menghitung nilai diferensial seleksi dan kemajuan seleksi. Diferensial
seleksi adalah selisih nilai tengah genotipe terseleksi dengan nilai tengah populasi
asal genotipe terseleksi, sedangkan kemajuan seleksi adalah selisih nilai tengah
turunan hasil seleksi dengan nilai tengah dari populasi yang telah diseleksi. Jika
seleksi dilakukan secara tidak langsung maka efektivitas seleksi dapat diukur
dengan menghitung respon terkorelasi masing-masing karakter terhadap daya
hasil (Falconer dan Mackay 1996).
Kelemahan seleksi menggunakan karakter kuantitatif adalah pengaruh
lingkungan tidak bisa dihindari terutama pada kondisi bercekaman sehingga
kemajuan seleksi yang diperoleh menjadi lebih kecil. Selain itu seleksi juga
tergantung pada umur tanaman, misal untuk daya hasil, seleksi baru bisa
berdasarkan karakter kuantitatif juga memiliki beberapa keuntungan seperti lebih
murah bahkan lebih mudah dari segi teknik pengerjaan.
Analisis QTL
Umumnya sifat agronomi yang bernilai ekonomi tinggi yang dimiliki oleh
tanaman, seperti daya hasil, ketahanan terhadap cekaman biotik seperti hama dan
penyakit, maupun ketahanan terhadap cekaman abiotik seperti kekeringan,
salinitas, suhu rendah, dan pH dikendalikan oleh banyak gen sehingga disebut
karakter kuantitatif (Falconer dan Mackay 1996). Karakter kuantitatif
diterjemahkan sebagai karakter dengan nilai yang mempunyai distribusi kontinyu
yang diperoleh dari hasil pengukuran atau penghitungan. Karakter kuantitatif
dikontrol oleh banyak gen dan masing-masing gen bersifat aditif terhadap karakter
tersebut. Ekspresi gen-gen yang mengendalikan karakter kuantitatif sangat
dipengaruhi oleh lingkungan (Bernardo 2002). Pengaruh lingkungan yang besar terhadap keragaman karakter kuantitatif merupakan salah satu kesulitan yang
dihadapi pemulia dalam melakukan seleksi.
Pada kondisi lingkungan bercekaman nilai heritabilitas daya hasil lebih
rendah dibandingkan dengan kondisi optimum karena lingkungan bercekaman
sangat mempengaruhi daya hasil. Hal ini mengakibatkan seleksi terhadap daya
hasil akan memberikan kemajuan genetik yang rendah untuk meningkatkan daya
hasil (Ceccareli 1994). Dengan demikian diperlukan karakter seleksi selain daya
hasil yang dapat digunakan sebagai alat bantu seleksi untuk memperbaiki daya
hasil pada kondisi bercekaman.
Alat bantu seleksi yang diharapkan adalah berupa marka yang mempunyai
nilai heritabilitas tinggi atau pengaruh lingkungan kecil. Alat bantu seleksi yang
tidak dipengaruhi oleh lingkungan adalah marka molekuler. Pemanfaatan marka
molekuler sebagai alat bantu didasarkan pada polimorfik yang ditemui pada
tingkat DNA. Menurut Brar (2002), penggunaan marka molekuler sebagai alat
bantu seleksi dapat meningkatkan akurasi seleksi karena marka molekuler tidak
dipengaruhi oleh lingkungan, stabil dan terekspresi pada semua stadia
pertumbuhan tanaman. Di samping nilai heritabilitas yang tinggi, keuntungan
terbatas (Gupta et al. 2002). Teknik yang merupakan kombinasi antara pemuliaan
konvensional dan bioteknologi yang berbasis marka molekuler dapat
meningkatkan akurasi seleksi atau mempersingkat waktu pencapaian tujuan
pemuliaan. Strategi potensial pemanfaatan bioteknologi dalam bidang pemuliaan
adalah melalui marka assisted selection (MAS) (Bernardo 2002).
Sampai sejauh ini sudah tersedia banyak jenis marka molekuler yang dapat
digunakan. Masing-masing marka molekuler mempunyai keuntungan dan
kesulitan yang berbeda dari segi teknik, fasilitas dan biaya yang diperlukan.
Salah satu marka molekuler yang murah dan mudah secara teknik adalah RAPD
(Random Amplified Polymorphic DNA). Random Amplified Polymorphic DNA
adalah teknik untuk mengamplifikasi DNA dengan mesin PCR menggunakan
primer tunggal berukuran 10 nukleotida. Primer yang digunakan merupakan
primer acak yang mengamplifikasi genom target secara acak. Primer acak yang
dicampurkan dengan reaksi amplifikasi akan berikatan dengan sekuens
komplemen di sepanjang genom target. Selanjutnya sekuens target akan
teramplifikasi yang dapat divisualisasikan pada gel agarose yang diwarnai dengan
ethidium bromida.
Dibandingkan dengan teknik molekuler yang lain, teknik RAPD lebih
sederhana karena tidak memerlukan informasi tentang genom, DNA tidak perlu
dipotong dengan enzim restriksi, tidak perlu pelabelan dengan radioisotope,
jumlah DNA yang diperlukan lebih sedikit dan jumlah lokus yang diketahui lebih
banyak yaitu 1-10 lokus. Kelemahan teknik RAPD adalah tingkat
reprodusibilitasnya sangat rendah sehingga sering memberikan hasil yang tidak
konsisten (Gupta et al. 2002; Toruan-Mathius dan Hutabarat 2003).
Dalam marka RAPD, alel yang berbeda pada lokus yang sama ditandai
dengan ada atau tidaknya pita hasil amplifikasi. Individu tanaman dibedakan
berdasarkan ada atau tidak pita hasil amplikasi. Jika muncul pita maka genotipe
individu adalah homozigot dominan atau heterozigot. Jika pita tidak muncul
untuk lokus yang sama maka individu tanaman adalah homozigot resesif. Marka
RAPD merupakan marka dominan sehingga individu homozigot dan heterozigot
tidak dapat dibedakan, tetapi dapat membedakan individu homozigot dominan
bersegregasi. Individu homozigot dan heterozigot akan menghasilkan pita
amplifikasi yang sama (Liu 1998).
Marka RAPD merupakan salah satu marka molekuler yang banyak
digunakan dalam membuat peta genetik. Pemilihan marka sangat ditentukan oleh
komposisi genotipe dalam populasi atau tujuan penelitian. Marka RAPD sangat
sesuai untuk diaplikasikan pada populasi double haploid (DH) dan recombinant
inbreed lines (RILs). Dalam populasi DH atau RILs sebagian besar genotipe
terdiri dari genotipe homozigot dominan atau homozigot resesif (Liu 1998; Gupta
et al. 2002).
Beberapa tahun terakhir pengembangan alat bantu seleksi dalam pemuliaan
tanaman banyak dilakukan melalui analisis molekuler dengan pemetaan marka
molekuler yang terpaut dengan QTL (Quantitative Trait Loci) yang
mengendalikan karakter yang akan diperbaiki. Penggunaan marka molekuler
yang terpaut dengan QTL yang mengendalikan karakter yang akan diperbaiki
dikenal dengan marker assisted selection (MAS) atau marker assisted breeding
(MAB) (Arus dan Moreno-Gonzales 1993).
Agar dapat memanfaatkan marka molekuler sebagai MAS dalam program
seleksi terhadap karakter yang diinginkan maka marka yang berasosiasi dengan
QTL yang mengendalikan karakter tersebut harus diidentifikasi terlebih dahulu.
Indentifikasi marka yang berasosiasi dengan QTL dapat dilakukan melalui
analisis dan pemetaan QTL (Azrai et al. 2002; Ruswandi et al. 2002).
Quantitative Trait Loci (QTL) adalah lokus yang mengendalikan karakter
kuantitatif. Pemetaan QTL merupakan kombinasi antara analisis pautan kualitatif
dengan analisis genetika kuantitatif. Pemetaan QTL meliputi konstruksi pemetaan
genom dan penelusuran hubungan antara karakter kuantitatif dengan marka
polimorfik. Hasil pemetaan QTL dapat menyediakan informasi tentang jumlah
dan aksi gen yang mengontrol suatu karakter serta lokasinya pada kromosom.
Peta QTL dapat juga dijadikan informasi awal bagi kegiatan kloning yaitu
kloning berbasis pemetaan dari suatu gen yang berasosiasi dengan karakter
tertentu (Liu 1998; Surahman 2002; Winicov 2002).
Dengan memanfaatkan marka molekuler yang terpaut dengan QTL sebagai