Trop. For. Manage. J. V (1) : 11-21 (1999)
PATOGENISITAS Rhizoctonia solani PADA SEMAI Pinus merkusii
DAN Acacia mangium
Pathogenicity of Rhizoctonia solani on Pinus merkusii and Acacia
mangium Seedlings
ACHMAD1),SOETRISNO HADI1),ELIS N.HERLINAYANA1) danAGUS SETIAWAN2)
ABSTRACT
Pathogenicity of Rhizoctonia solani was studied by evaluating the ability of the pathogen to attack several ages of Pinus merkusii and Acacia mangium seedlings. Results showed that R. solani attacked P. merkusii from seed stage up to seven week-old seedlings, while eight week-old ones were
free from the pathogen’s attack. On A. mangium, 16 day-old seedlings were uninfected by R. solani, while 12 day-old ones were still attacked by the pathogen.
Pathogenesis of R. solani was also studied by evaluating the activities of cellulolytic and pectolytic enzymes produced by the fungi and compared them with the activities of the same enzymes
produced by Fusarium oxysporum. Result showed that R. solani’s cellulolytic enzymes activity,
reflected by the activity of C1-cellulase, was lower compared with that of F. oxysporum. On the other hand, R. solani was more intensive degrading pectin medium than F. oxysporum, such phenomenon reflected higher activity of pectolytic enzymes activity of R. solani compared with that of F. oxysporum.
PENDAHULUAN
Hutan Tanaman Industri (HTI) yang dicanangkan pembangunannya sejak tahun 1984, menjadi tumpuan harapan kesinambungan pasokan bahan baku industri perkayuan, sekaligus dimaksudkan untuk merehabilitasi areal hutan produksi yang sudah tidak produktif, serta untuk membangun lahan-lahan kritis agar menjadi produktif kembali. Pinus merkusii dan Acacia mangium adalah dua di antara jenis-jenis pohon yang disarankan untuk ditanam di areal HTI. Keduanya merupakan jenis asli Indonesia yang cepat tumbuh.
Pembangunan HTI berimplikasi dengan penanaman pohon sejenis, seumur, pada areal pertanaman yang luas. Keberhasilan pembangunan HTI sangat ditentukan antara lain oleh tersedianya semai berkualitas tinggi dalam jumlah yang cukup. Penyakit lodoh merupakan salah satu penyebab utama berkurangnya jumlah semai di pesemaian. Semai P. merkusii dan A. mangium keduanya rentan terhadap penyakit tersebut.
Penyakit lodoh disebabkan oleh beberapa jenis fungi patogen penghuni tanah, salah satu di antaranya adalah Rhizoctonia solani. Fungi patogen ini mempunyai daya adaptasi yang tinggi, memiliki banyak strain, kisaran inangnya luas, dan mampu bertahan hidup lama di dalam tanah (Baker dan Snyder, 1970; Ogoshi, 1975). Perpaduan antara sifatnya sebagai saprob dan sifat patogeniknya yang tidak hanya terbatas pada inang tertentu,
jadikan R. solani sebagai patogen yang penting secara ekonomi, serta sulit dikendalikan di lapangan.
Tujuan penelitian ini adalah untuk mengetahui patogenisitas R. solani pada semai P. merkusii dan A. mangium, yaitu dengan menguji periode serangannya. Di samping itu tujuannya juga juga untuk mempelajari patogenesis fungi tersebut dengan menguji aktivitas enzim-enzim selulolitik dan pektolitiknya. Kegunaan mengetahui periode serangan tersebut adalah untuk menentukan sampai umur berapa semai masih memerlukan perlindungan terhadap serangan R. solani, sedang melalui uji patogenesis dapat diketahui mekanisme serangan patogen sebagai informasi untuk memanfaatkan mekanisme pertahanan inang dalam rangka pengembangan cara perlindungan semai terhadap patogen bersangkutan.
BAHAN DAN METODE
Isolat R. solani yang digunakan dalam percobaan diperoleh melalui isolasi lang-sung dari pangkal batang semai P. merkusii yang terserang lodoh. Isolat yang digunakan memiliki karakteristik: hifa berukuran 6-9 m, bersepta, percabangannya tegak lurus, berwarna putih kecoklatan, tidak menghasilkan konidia, menghasilkan sklerotia, multinukleat, membentuk sel-sel monilioid. Karakteristik tersebut sesuai dengan karak-teristik R. solani yang dikemukakan Ogoshi (1975) serta Sneh dkk. (1991).
Patogenisitas R. solani pada Semai P. merkusii dan A. mangium
Inokulum R. solani disiapkan dengan menanam tiga potongan koloni ( 6 mm) berumur lima hari pada 100 g media CMS ('Corn Meal Sand'), yang terdiri atas campuran pasir, hancuran biji jagung dan air (96:4:20 g/g/ml), dalam labu Erlenmeyer volume 250 ml yang diotoklaf (121 oC, 1 atmosfer) selama 60 menit. Labu beserta isinya ditempatkan pada suhu kamar selama dua minggu. Untuk kontrol media patogen, labu Erlenmeyer berisi jenis media campuran yang sama tanpa diinokulasi fungi ditempatkan pada suhu kamar selama dua minggu.
Benih P. merkusii dan A. mangium yang akan digunakan direndam dalam air destilata selama 24 jam. Benih yang tenggelam kemudian disterilkan permukaannya dengan merendam dalam larutan NaOCl 0.5% selama 10 menit, dan selanjutnya benih dibilas beberapa kali dengan air steril. Benih kemudian dikeringanginkan dengan mele-takkannya dalam wadah yang telah dialasi kertas saring steril, selanjutnya benih ditabur pada bak pengecambah berisi media tanam steril. Penaburan benih dilakukan secara bertahap dengan hitungan mundur untuk memperoleh semai dengan umur yang dikehendaki. Semai dipelihara dengan penyiraman tiap sore hari hingga mencapai umur yang diperlukan.
dan 16 hari untuk A. mangium. Taraf 0 pada umur semai menunjukkan bahwa bahan pertanamannya adalah benih. Satuan percobaannya adalah 20 semai atau benih yang masing-masing ditanam dalam wadah tanam terpisah. Media tanam yang digunakan berupa campuran tanah Latosol Darmaga bukan dari pertanaman dan pasir ( 1:1 b/b) yang sudah diotoklaf (121oC, 1 atmosfer) selama 60 menit.
Infestasi dilakukan dengan mencampur merata 2.5 g inokulum dengan 100 g media tanam dalam politub ('polytube'), kemudian ditambah 10 ml air steril. Untuk kontrol ditambah media patogen, ke dalam tiap 100 g media tanam ditambahkan 2.5 g media patogen dan 10 ml air steril, sedang untuk kontrol tanpa infestasi patogen dan penambahan media patogen media hanya ditambah 10 ml air steril. Wadah tanam selanjutnya ditutup dengan plastik, kemudian didiamkan selama empat hari.
Setelah empat hari, dilakukan penanaman semai P. merkusii atau A. mangium dengan umur yang dikehendaki. Sebelum ditanam, semai yang diambil dari bak penge-cambah terlebih dahulu akarnya dicuci dan disemprot dengan air steril. Wadah tanam kemudian ditempatkan di bawah rumah plastik yang dinaungi paranet 70 % untuk P. mer-kusii, atau di rumah kaca untuk A. mangium. Penyiraman dilakukan tiap sore hari meng-gunakan air kran.
Pengamatan terhadap semai yang mati dilakukan tiap hari hingga 14 hari setelah tanam (hst) untuk P. merkusii dan 12 hst untuk A. mangium. Untuk umur 0 minggu, pengamatan dilakukan sejak benih berkecambah sampai dengan 14 atau 12 hari kemudian berturut-turut untuk P. merkusii dan A. mangium
Patogenesis R. solani
Studi patogenesis R. solani dilakukan dengan menguji aktivitas enzim-enzim selulolitik dan pektolitik fungi patogen tersebut. Sebagai pembanding, pengujian juga dilakukan terhadap Fusarium oxysporum yang juga merupakan salah satu jenis patogen lodoh. Untuk mewakili aktivitas enzim selulolitik, dilakukan pengujian aktivitas FP-ase ('Filter Paper'-ase) atau selulase-C1, sedang untuk enzim pektolitik diwakili oleh aktivitas poligalakturonase. Aktivitas selulase-C1 ditentukan dengan teknik spektrofotometri menggunakan glukosa sebagai standar, sedang aktivitas poligalaktoronase ditentukan dengan teknik iodometri menggunakan asam galakturonat monohidrat sebagai standar.
Enzim diekstrak dari isolat yang ditumbuhkan pada beberapa macam 'media'. Media pertama adalah CMS steril dalam tabung reaksi yang diinokulasi dengan satu potongan koloni fungi patogen ( 6 mm) dan diinkubasi selama tujuh hari, dan sebagai kontrol adalah media CMS yang tidak diinokulasi fungi patogen. Media ke dua adalah CMS steril dalam tabung reaksi yang di atasnya ditempatkan 0.1 g potongan batang semai steril (panjang potongan 1 cm) berumur 2 atau 8 minggu, kemudian media diinokulasi dengan satu potongan koloni fungi patogen 6 mm dan selanjutnya diinkubasi selama tujuh hari. Kontrol disiapkan dengan prosedur sama, hanya saja tidak dilakukan inokulasi fungi patogen. Sterilisasi media CMS dilakukan dalam otoklaf (121oC, 1 atmosfer) selama 60 menit, sedang sterilisasi permukaan potongan batang semai P. merkusii dilakukan dengan cara merendamnya dalam HgCl2 0.5 % selama 3 menit
Uji lain untuk mendeteksi produksi pektinase dilakukan dengan menumbuhkan fungi pada media pektin (Atlas, 1993) dan diinkubasi selama 10 hari. Produksi pektinase ditandai antara lain oleh berubahnya warna media dari hijau kecoklatan menjadi merah.
HASIL PENELITIAN
Patogenisitas R. solani pada Semai P. merkusii dan A. mangium
Hasil pengamatan pada P. merkusii menunjukkan bahwa R. solani mengakibatkan lodoh benih, lodoh dalam tanah, dan lodoh pangkal batang. Untuk lodoh pangkal batang, patogen menyerang hipokotil semai yang masih sukulen. Gejala khas penyakit lodoh tipe tersebut adalah terjadinya penggentingan hipokotil akibat maserasi jaringan dan diikuti pembusukan hipokotil dekat permukaan tanah secara cepat, sehingga semai yang masih segar menjadi rebah.
Infestasi R. solani nyata mengakibatkan persentase semai mati yang tinggi dibanding kontrol maupun yang ditambah media patogen, baik pada P. merkusii maupun A. ma-ngium. Meskipun demikian rata-rata persentase semai mati tersebut memperlihatkan nilai simpangan baku yang tinggi (Tabel 1). Hal tersebut menunjukkan bahwa persentase semai mati akibat serangan R. solani dipengaruhi oleh umur semai.
Tabel 1. Persentase Semai P. merkusii dan A. mangium yang Mati pada Beberapa Perlakuan Media Tanam dan Tingkat Umur
Perlakuan Semai mati
*)
(%)
P. merkusii A. mangium Media tanam
Kontrol 6.2(b) 6.78 0.0(b) 0.00
Ditambah media patogen 5.8(b) 6.68 0.0(b) 0.00
Diinfestasi R. solani 61.7(a) 43.65 69.0(a) 43.32 Umur semai
P. merkusii (minggu)
A. mangium (hari)
0 0 59.0(a) 40.94 33.3(a) 51.64
2 4 65.0 (a) 45.21 33.3(a) 51.64
4 8 60.0 (a) 43.84 33.3(a) 51.64
6 12 25.0(b) 23.33 15.0(b) 25.10
7 16 8.0 (c) 7.88 0.0(c) 0.00
8 0.0 (d) 0.00
*) - nilai tiap kolom: rataan simpangan baku dalam persen
- nilai rataan pada kolom yang sama yang diikuti huruf yang sama tidak berbeda pada taraf nyata 5 % berdasarkan uji jarak berganda Duncan
sedang pada tingkat umur lainnya tidak terjadi kematian semai. Untuk A. ma-ngium, tidak ditemukan benih/semai mati pada kedua perlakuan kontrol pada semua tingkat umur semai.
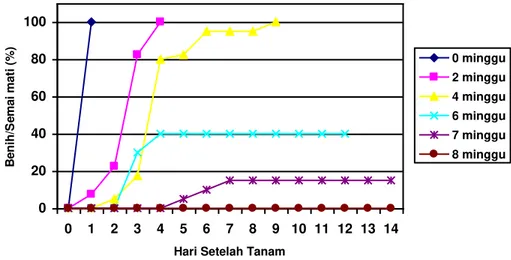
Serangan R. solani pada semai P. merkusii terjadi dari stadia benih hingga semai berumur tujuh minggu, sedang semai berumur delapan minggu telah bebas dari serangan. Semai mati hingga 100 % terjadi hingga umur semai empat minggu, dan sesudah umur tersebut persentase semai mati nyata berkurang. Serangan tercepat terjadi pada semai umur 2 minggu karena pada 4 hst semai mati telah mencapai 100% (Gambar 1).
0 20 40 60 80 100
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Hari Setelah Tanam
B
e
ni
h/
S
e
m
a
i
m
a
ti
(
%
)
0 minggu 2 minggu 4 minggu 6 minggu 7 minggu 8 minggu
Gambar 1. Serangan R. solani pada Beberapa Tingkat Umur Semai P. merkusii
Serangan R. solani pada semai A. mangium terjadi dari stadia benih hingga semai berumur 12 hari, sedang semai berumur 16 hari telah bebas dari serangan. Semai mati hingga 100% terjadi hingga umur semai delapan hari, dan pada umur semai 12 hari serangan R. solani nyata berkurang (Gambar 2).
Patogenesis R. solani
0 20 40 60 80 100
0 1 2 3 4 5 6 7 8 9 10 11 12
Hari Setelah Tanam
B
e
n
ih
/S
e
m
a
i
Ma
ti
(
%
)
0 hari 4 hari 8 hari 12 hari 16 hari
Gambar 2. Serangan R. solani pada Beberapa Tingkat Umur Semai A. mangium
Tabel 2. Kadar Glukosa yang Ditimbulkan oleh Filtrat Biakan R. solani dan F. oxysporum pada Beberapa Macam Media Beserta Kontrolnya
Perlakuan*) Kadar glukosa
**)
(mg/ml)
Kadar asam galakturonat monohidrat2) (mg/ml) R. solani - CMS 0.078 (b) 0.0035 1.378 0.0700 F. oxysporum – CMS 0.171 (d) 0.0155 1.353 0.0346
Kontrol CMS 0.013 (a) 0.0000 1.323 0.0438
R. solani - CMS - 2m 0.157 (cd) 0.0056 1.311 0.0608 F. oxysporum - CMS - 2m 0.213 (e) 0.0163 1.323 0.0084
Kontrol CMS - 2m 0.013 (a) 0.0014 1.354 0.0169
R. solani - CMS - 8m 0.202 (e) 0.0056 1.311 0.0777 F. oxysporum - CMS - 8m 0.323 (f) 0.0120 1.329 0.0176
Kontrol CMS - 8m 0.015 (a) 0.0014 1.323 0.0438
1) *- CMS-2m atau -8m: media CMS ditambah potongan batang semai P. merkusii umur 2minggu atau 8 minggu; kontrol: media tanpa fungi patogen
2) nilai rataan yang diikuti huruf yang sama tidak berbeda pada taraf 5 % berdasarkan uji jarak berganda Duncan
keti-ga macam media tersebut, CMS tanpa potonketi-gan batang semai diduketi-ga menketi-gandung selu-losa terendah, yaitu yang terdapat pada hancuran biji jagung, dan kandungan seluselu-losa media makin meningkat berturut-turut oleh penambahan potongan batang semai P. mer-kusii umur dua minggu dan delapan minggu.
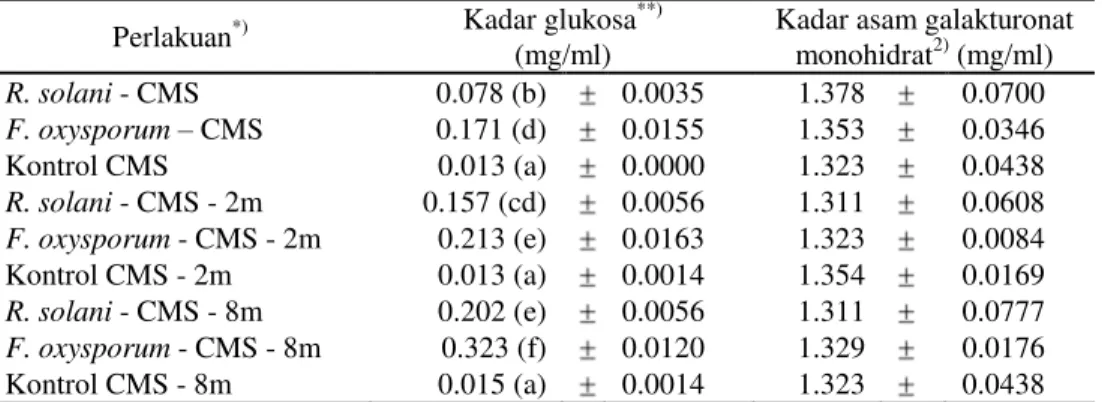
Kadar glukosa yang ditimbulkan oleh filtrat biakan F. oxysporum pada CMS dan CMS yang ditambah potongan batang semai P. merkusii umur dua minggu atau delapan minggu berturut-turut sebesar 0.171, 0.213, dan 0.323 mg/ml. Kadar glukosa yang ditimbulkan oleh filtrat biakan R. solani pada urutan media yang sama juga meningkat, yaitu sebesar 0.078, 0.157, dan 0.202 mg/ml. Hasil tersebut menunjukkan bahwa aktivitas selulase-C1 kedua jenis fungi patogen makin meningkat dengan makin banyaknya selulosa yang terdapat pada media.
Berlainan dari aktivitas selulase-C1 kedua jenis fungi patogen yang dapat dide-teksi pada semua media yang mengandung selulosa, aktivitas poligalakturonase ternyata tidak dapat dideteksi berdasarkan teknik iodometri yang telah dilakukan. Hal tersebut ditunjukkan oleh kadar asam galakturonat monohidrat yang ditimbulkan oleh filtrat biakan fungi pada semua media maupun kontrolnya yang tidak berbeda nyata (Tabel 2).
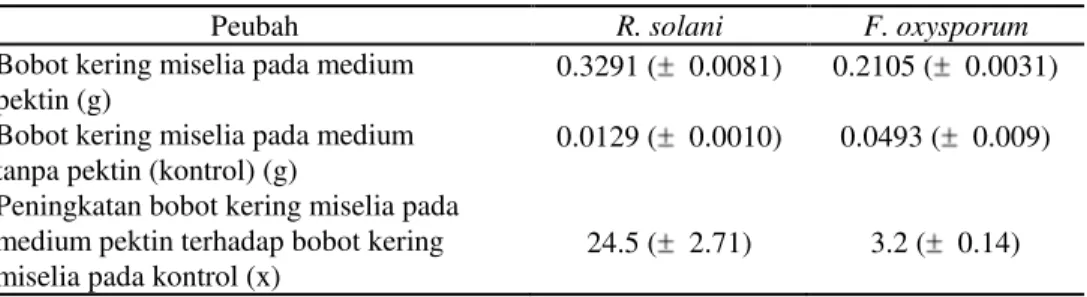
Meskipun aktivitas poligalakturonase kedua fungi patogen tidak berhasil dideteksi dengan teknik iodometri yang diterapkan, pembiakan kedua jenis fungi patogen pada medium pektin mengindikasikan dihasilkannya pektinase. Pertumbuhan kedua jenis fungi pada medium yang mengandung pektin lebih intensif dibanding pertumbuhan pada medium tanpa pektin. Bobot kering miselia R. solani dan F. oxysporum pada medium pektin lebih tinggi berturut-turut hingga 24.5 dan 3.2 kali dibanding bobot kering miselia kedua jenis fungi pada medium tanpa pektin (Tabel 3).
Tabel 3. Rata-rata Bobot Kering*) Miselia R. solani dan F. oxysporum pada Medium Pektin Beserta Kontrolnya
Peubah R. solani F. oxysporum
Bobot kering miselia pada medium pektin (g)
0.3291 ( 0.0081) 0.2105 ( 0.0031)
Bobot kering miselia pada medium tanpa pektin (kontrol) (g)
0.0129 ( 0.0010) 0.0493 ( 0.009)
Peningkatan bobot kering miselia pada medium pektin terhadap bobot kering miselia pada kontrol (x)
24.5 ( 2.71) 3.2 ( 0.14)
*)
bobot kering miselia setelah diinkubasi pada suhu 60oC selama 24 jam
PEMBAHASAN
Patogenisitas R. solani pada Semai
P. merkusii dan A. mangium
R. solani, dapat menyerang P. merkusii dari benih hingga semai berumur tujuh minggu. Rentang periode serangan ini lebih panjang dari yang dilaporkan Hodge dan Luchle (1959), yaitu hingga semai berumur empat atau lima minggu. Terhadap A. ma-ngium, R. solani menyerang benih hingga semai umur 12 hari. Oleh karena perlindungan semai terhadap R. solani dan patogen lodoh secara umum hendaknya tetap dilakukan hingga semai berumur 7 minggu untuk P. merkusii atau hingga semai berumur 12 hari untuk A. mangium.
R. solani sangat intensif menyerang semai P. merkusii dan A. mangium muda. Persentase semai mati makin berkurang dengan makin tuanya kedua jenis semai. Hal tersebut menunjukkan bahwa makin tua semai, ketahanannya terhadap serangan patogen lodoh makin meningkat. Boyce (1961) mengaitkan serangan patogen dengan kondisi jaringan semai yang masih sukulen. Achmad (1997) mempelajari anatomi batang semai P. merkusii berumur 2 hingga 8 minggu. Ia melaporkan bahwa semua bagian batang semai P. merkusii berumur dua minggu masih berupa jaringan sukulen, dan sifat sukulen tersebut makin berkurang dengan bertambah tuanya semai. Batang semai berumur lima minggu mulai berkayu, dan dengan makin bertambahnya umur semai maka batangnya makin keras.
Patogenesis R. solani
R. solani merupakan salah satu jenis patogen lodoh. Agrios (1988) menguraikan proses infeksi penyakit lodoh tipe lodoh benih yang melibatkan enzim-enzim pendegra-dasi lamela tengah dan dinding sel, meliputi pektinase, selulase, dan protease. Pada lodoh pangkal batang, terjadi gejala maserasi dan pembusukan jaringan terinfeksi secara cepat. Gejala demikian memperlihatkan terlibatnya aktivitas enzim-enzim patogen pen-degradasi lamela tengah dan dinding sel inang, yaitu pektinase dan selulase, dalam patogenesis (Bateman dan Basham, 1976).
Penambahan filtrat kedua jenis fungi yang dibiakkan pada medium CMS pada kertas saring, menyebabkan terdeteksinya glukosa. Hal tersebut menunjukkan aktivitas selulase-C1 kedua jenis fungi patogen untuk mendegradasi selulosa sebagai sumber karbon yang terdapat pada hancuran biji jagung. Penambahan potongan batang semai P. merkusii pada media CMS mengakibatkan aktivitas selulase-C1 kedua jenis fungi yang nyata lebih besar. Hal ini menunjukkan bahwa potongan batang semai P. merkusii menjadi substrat selulosa tambahan di samping selulosa yang terdapat pada hancuran biji jagung. Hasil ini sejalan dengan hasil penelitian Bateman (1964b) dan Yoshida dkk. (1989) yang berturut-turut melaporkan produksi selulase oleh R. solani dan F. oxysporum.
minggu. Hal ini menunjukkan bahwa kadar selulosa batang semai berumur delapan minggu lebih besar dibanding kadar selulosa batang semai yang berumur dua minggu, dengan asumsi bahwa perbedaan aktivitas selulase-C1 tersebut semata-mata disebabkan oleh faktor substrat.
Secara umum hasil penelitian memperlihatkan bahwa produksi glukosa F. oxyspo-rum nyata lebih tinggi dibanding R. solani pada semua media yang mengandung selulosa sebagai sumber karbon. Hal tersebut menunjukkan bahwa aktivitas selulase-C1 F. oxys-porum lebih tinggi dibanding R. solani.
Bila aktivitas selulase-C1 nyata terdeteksi pada filtrat biakan kedua jenis fungi patogen pada media yang mengandung selulosa, maka aktivitas pektinase, dalam hal ini adalah poligalakturonase (PG), tidak berhasil dideteksi menggunakan teknik iodometri yang diterapkan. Hal tersebut diduga berkaitan dengan kuantitas bahan tanaman sebagai sumber pektat dan teknik deteksinya. Dalam penelitian ini digunakan 0.1 g potongan batang semai P. merkusii sebagai sumber pektat dan deteksi dilakukan dengan teknik iodometri. Bateman (1963 a&b) juga menggunakan teknik deteksi iodometri, akan tetapi contoh sumber pektatnya jauh lebih banyak, yaitu 100 g hipokotil semai Phaseolus vul-garis. Ayers, Papavizas, dan Diem (1966) mendeteksi aktivitas PG dan PATE ('pectic acid transeliminase' atau asam poligalakturonat transeliminase) dengan teknik spektro-fotometri. Aktivitas PG dideteksi pada panjang gelombang 515 nm, dan teknik tersebut mampu mendeteksi asam galakturonat yang merupakan produk aktivitas PG hingga 0.05 mg/ml.
Meskipun dengan teknik iodometri aktivitas poligalakturonase tidak berhasil dideteksi, pembiakan pada medium pektin mengindikasikan bahwa kedua jenis fungi patogen menghasilkan pektinase. Dibanding aktivitas pektinase F. oxysporum, aktivitas pektinase R. solani diduga lebih tinggi. Hal tersebut ditunjukkan oleh lebih tingginya peningkatan bobot kering miselia R. solani pada medium pektin yang menunjukkan bahwa pektinase fungi patogen tersebut lebih efektif mendegradasi pektin dan selanjut-nya memanfaatkanselanjut-nya sebagai sumber karbon dibanding F. oxysporum. Dugaan lebih tingginya aktivitas pektinase R. solani tersebut juga didukung oleh perubahan warna medium yang lebih tajam, di samping itu fungi patogen tersebut juga mengakibatkan medium menjadi transparan.
Goodman dkk. (1986) mengemukakan bahwa pektinase terdiri atas enzim-enzim pektin metil esterase (PME), poligalakturonase (PG), pektin metil galakturonase (PMG), pektin transeliminase (PTE) dan asam poligalakturonat transeliminase (PATE). Pembiakan fungi pada medium pektin dapat memberikan indikasi dihasilkannya pektinase, akan tetapi tidak dapat diketahui macam enzim yang dihasilkan oleh tiap jenis fungi patogen tersebut. Meskipun demikian secara umum hasil ini mendukung hasil pene-litian Bateman (1963a&b) yang berhasil mendeteksi aktivitas PG dalam filtrat biakan R. solani dengan menggunakan hipokotil P. vulgaris sebagai sumber pektat. Ayers, dkk. (1966) juga berhasil mendeteksi aktivitas PG dan PATE dari hipokotil jenis tanaman yang sama yang terinfeksi R. solani.
sinergistik berperan dalam maserasi jaringan. Keberadaan PG dalam maserasi jaringan terinfeksi sifatnya esensial. Selulase komersial bila tanpa PG tidak mengakibatkan mase-rasi jaringan, sedang PG bekerja lebih baik bila terdapat selulase. Hasil penelitian ini menunjukkan dugaan lebih tingginya aktivitas pektolitik R. solani dibanding F. oxyspo-rum, sebaliknya aktivitas selulolitik F. oxysporum lebih tinggi dibanding aktivitas selulo-litik R. solani. Hasil tersebut dengan didukung oleh pernyataan Bateman (1963b) dapat menerangkan fenomena lebih tingginya patogenisitas R. solani dibanding F. oxysporum terhadap semai P. merkusii sebagaimana dilaporkan oleh Achmad (1991 & 1997).
KESIMPULAN
R. .solani menyerang P. merkusii dan A. mangium dari stadia benih hingga semai umur tertentu. Pada P. merkusii, serangan terjadi hingga semai berumur tujuh minggu, sedang semai berumur delapan minggu telah bebas dari serangan R. solani. Pada A. mangium, semai berumur 16 hari baru terbebas dari serangan, sedang semai hingga berumur 12 hari masih terserang.
Aktivitas enzim-enzim selulolitik R. solani, yang diwakili oleh enzim selulase-C1, lebih rendah dibanding F. oxysorum. Di lain pihak, R.solani lebih intensif mendegradasi medium pektin dibanding F. oxysporum, dan hal tersebut mencerminkan lebih tingginya aktivitas enzim pektolitik R. solani dibanding F. oxysporum. Tingginya aktivitas enzim pektolitik R. solani mendukung kenyataan tingginya patogenisitas fungi tersebut di lapangan, karena gejala maserasi jaringan terutama ditentukan oleh aktivitas enzim-enzim pektolitik.
DAFTAR PUSTAKA
Achmad., 1991. Kemampuan Rhizopogon sp. untuk Perlindungan Hayati Terhadap Penyebab Penyakit Lodoh pada Pinus merkusii. Tesis Magister Sains. Program Pascasarjana IPB, Bogor. 68 hlm.
Achmad., 1997. Mekanisme Serangan Patogen dan Pertahanan Inang serta Pengendalian Hayati Penyakit Lodoh pada Pinus merkusii. Disertasi Doktor. Program Pascasarjana IPB, Bogor. 164 hlm.
Agrios, G. N., 1988. Plant Pathology, 3rd ed. Academic Press Inc., San Diego, New York, Berkeley, Boston, London, Sydney, Tokyo, Toronto. 803 hlm.
Atlas, R.M., 1993. Handbook of Microbiological Media. CRC Press, Boca Raton, Ann Arbor, London, Tokyo. 1079 p.
Ayers, W.A., G.C. Papavizas, dan A.F. Diem., 1966. Polygalacturonate Transeliminase and Polygalacturonase Production by Rhizoctonia solani. Phytopathology, 56:1006-1011.
Baker, K.F. and W.C. Snyder. 1970. Ecology of Soil-borne Plant Pathogens, Prelude to Biological Control. Univ. California Press, California. 571 p.
Bateman, D.F., 1963b. The "Macerating Enzymes" of Rhizoctonia solani. Phyto-pathology, 53:1178-1186.
Bateman, D.F., 1964b. Cellulase and the Rhizoctonia Disease of Bean. Phyto-pathology 54:1372-1377.
Bateman, D.F. and H.G. Basham. 1976. Degradation of Plant Cell Walls and Membranes by Microbial Enzymes, pp. 316-355. In Physiological Plant Pathology, vol. 4. (R. Heitefuss dan P.H. Williams, (eds.). Springer-Verlag, Berlin, Heidelberg, New York.
Boyce, J.S. 1961. Forest Pathology. McGraw-Hill Co. Inc., New York, Toronto, London. 572 p.
Goodman, R.N., Z. Kiraly, dan K.R. Wood. 1986. The Physiology and Biochemistry of Plant Disease. Univ. Missouri Press, Columbia. 433 p.
Hodge, C.S. dan J.L. Luchle. 1959. Nursery Diseases of Southern-pines. Forest Pest Leaflet No. 32. US Department of Agriculture Forest Service, Washington. Ogoshi, A. 1975. Grouping of Rhizoctonia solani Kahn and Their Perfect Stages. Rev.
Plant Protection Res., 8:93-103.
Sneh, B., L. Burpee, dan A. Ogoshi. 1991. Identifcation of Rhizoctonia Species. The Amer. Phytopathol. Soc. Press, St. Paul - Minnesota. 135 p.
Trop. For. Manage. J. V(2) : 1-12 (1999)
KOMPOSISI JENIS HAYATI DI EKOSISTEM TAMBAK
TUMPANGSARI POLA EMPANG PARIT: Studi Kasus di RPH Tegal
Tangkil, BKPH Ciasem, KPH Purwakarta, Jawa Barat
Biological Species Composition of Silvofishery Ecosystem Using
Cannal-pond Pattern: A case Study in RPH Tegal Tangkil, BKPH Ciasem, KPH
Purwakarta, West Java
JENI RIVIANA1),CECEP KUSMANA2) dan AGUS PRIYONO3)
ABSTRACT
Biotic component composition of silvofishery using cannal-pond pattern was elucidated in this study. This pattern is the combination between fishpond canal and mangrove vegetation growing in the center.
Sampling method was conducted in the four fish ponds with different variation of mangrove vegetation composition, such as Avicennia marina monoculture, Rhizophora mucronata monoculture, regularly mixed Rhizophora mucronata and Avicennia marina, and randomly mixed Rhizophora mucronata and Avicennia marina.
The research results showed that empang parit (fish pond) having a randomly mixed R. mucronata and A. marina give the most suitable habitat for the terrestrial fauna and aquatic biota as a feeding and breeding areas.
PENDAHULUAN
Hutan mangrove merupakan sumberdaya alam yang berperan ganda, baik ditinjau dari aspek ekonomi maupun ekologi. Secara fisik hutan mangrove dapat berfungsi untuk mempercepat perluasan lahan, melindungi pantai dan tebing sungai dan mengolah bahan limbah. Fungsi biologis hutan mangrove adalah sebagai tempat mencari makan, memijah dan membesarkan anak berbagai jenis biota air serta habitat berbagai jenis satwa liar, terutama burung.
Selain itu, secara ekonomi hutan mangrove dapat digunakan sebagai sumber bahan bakar, bahan bangunan, perikanan, pertanian dan keperluan produksi lainnya.. Sejalan dengan meningkatnya pertumbuhan penduduk, semakin meningkat pula berbagai kebutuhan yang bertumpu pada sumberdaya alam, sehingga diantaranya meluaslah pengkonversian hutan mangrove menjadi penggunaan lain seperti perikanan, pertanian, pemukiman dan lain-lain. Salah satu pemanfaatan hutan mangrove yang saat ini banyak terjadi di pantai utara pulau Jawa adalah tambak tumpangsari. Yang merupakan kombinasi antara tambak dengan vegetasi mangrove. Namun demikian semakin meluasnya petak-petak tambak tumpangsari yang kurang memperhatikan komposisi jenis pohon mangrove,
1) Alumni Fakultas Kehutanan IPB
2) Staf Pengajar Jurusan Manajemen Hutan, Fakultas Kehutanan IPB
akan mempengaruhi kenekaragaman hayati terutama penurunan keanekaragaman jenis fauna. Padahal fauna ditambak tumpangsari merupakan salah satu sumberdaya alam hayati yang mendukung proses-proses ekologis di dalam ekosistem mangrove, dimana fauna tersebut memerlukan mangrove sebagai habitat hidupnya. Dengan terganggunya kehidupan fauna di suatu tambak, akan terganggu pula proses-proses ekologis yang berlangsung di dalam ekosistem yang bersangkutan.
Penelitian ini bertujuan untuk mengetahui komposisi jenis pohon mangrove yang memberikan habitat yang cukup baik terhadap berbagai jenis fauna di tambak tumpangsari pola empang parit.
METODE
Lokasi Penelitian
Penelitian ini dilaksanakan di Blanakan, RPH Tegal Tangkil, BKPH Ciasem, KPH Purwakarta. Secara geografis, Blanakan terletak antara 107031’-107039’ BT dan 6011’ LU-6020’LS. Blanakan mempunyai luas areal sekitar 8.581 ha, antara lain 2.304,50 ha merupakan kelompok hutan yang telah dikukuhkan, yang mana 1.391,50 ha dari kelompok hutan tersebut telah dikonversi menjadi tambak tumpangsari.
Jenis tanah terdiri dari tanah aluvial di sebelah selatan dan tanah berpasir di sebelah utara. Seluruh areal merupakan dataran rendah payau dengan ketinggian 0 - 2 m di atas permukaan laut.
Data yang diambil adalah data primer dan data sekunder. Pengambilan sampel dilakukan pada empat pola tambak empang parit dengan komposisi jenis pohon yang berbeda, yaitu kombinasi antara Avicennia marina dengan Rhizophora mucronata berpola teratur, kombinasi antara A. marina dengan R. mucronata berpola acak, A. marina monokultur dan R. mucronata monokultur.
Analisis vegetasi
Analisis vegetasi dilakukan dengan cara kuadrat. Bagian tambak yang berhutan dibagi habis kedalam plot-plot berukuran 10 m x 10 m untuk risalah pohon. Kemudian plot-plot tersebut dibagi lagi kedalam sub-sub plot yang lebih kecil secara nested sampling dengan ukuran 5 m x 5 m dan 2 m x 2 m masing-masing untuk risalah pohon, pancang dan semai.
Pengamatan fauna
Pengamatan fauna diurnal dan nokturnal
Pengamatan fauna dilakukan dengan metode titik pengamatan yaitu dengan mengamati fauna di sepanjang jalur pengamatan. Panjang jalur dan jumlah titik pengamatan fauna disesuaikan dengan lokasi analisis vegetasi, luas petak tambak dan waktu aktif satwa.
Adapun serangga nokturnal dilakukan pengamatan pada pukul 19.00 – 21.10 WIB dengan lama pengamatan satu jam (dua kali ulangan).
Pengamatan terhadap parameter biologi, fisika dan kimia perairan.
Pengambilan contoh air dilakukan di tiga stasiun pengamatan yaitu stasiun pada tempat masuknya air tambak, 85 m dari tempat masuknya air tambak dan di tengah tambak di bawah penutupan vegetasi pada setiap petak tambak yang terpilih. Parameter yang diamati terdiri dari fisik-kimia (suhu, salinitas, pH, DO, nitrat dan fosfat) dan biologi perairan tambak (plankton, benthos, udang dan ikan).
Analisis data dilakukan dengan rumus sebagai berikut :
Analisis vegetasi dihitung dengan rumus sebagai berikut (Soerianegara dan Indrawan, 1985) :
(1) Tingkat dominasi jenis tumbuhan untuk tingkat pohon dan INP=KR+FR untuk tingkat permudaan
INP = KR + FR + DR
dimana :
KR : Kerapatan Relatif DR : Dominansi Relatif FR : Frekuensi Relatif INP : Indeks Nilai Penting
(2) Keragaman jenis tumbuhan dan fauna ditentukan dengan rumus Shanon Weiner Index of General Diversity :
H’ = - pi ln pi dimana:
H’ : Keragaman jenis
pi : Jumlah individu spesies ke-i dibagi dengan jumlah total individu semua jenis.
(3) Kelimpahan plankton dan kepadatan benthos ditentukan dengan rumus (Yulianda dan Damar, 1994) :
P
n
Vs
1
Vo
Vr
Op
Oi
N
dimana:
N : Kelimpahan plankton (ind/l) Oi : Luas gelas penutup (mm2)
Op : Luas satu lapangan pandang (mm2) Vr : Volume botol contoh plankton hasil saringan (ml) Vo : Volume satu tetes air contoh (ml) Vs : Volume air yang disaring oleh jaring plankton (l) n : Jumlah plankton yang tercacah p : Jumlah ulangan
(4) Kepadatan benthos dihitung dengan rumus :
dimana :
N : Kepadatan benthos (ind/ m2) x : Jumlah individu per satu satuan alat m : Luas bukaan mulut alat (m2)
HASIL DAN PEMBAHASAN
Hasil
Komposisi Jenis dan Struktur Tegakan
Indeks nilai penting jenis tumbuhan pada tegakan mangrove di setiap tipe tambak disajikan pada Tabel 1.
Tabel 1. Tingkat Nilai Penting pada Tegakan Mangrove pada Berbagai Tingkat Pertumbuhan.
Tingkat Pertumbuha
Tipe Tambak
A B C D
Species Species Species Species
Pohon R. mucronata (17,76 %)
Pancang R. mucronata (29,74 %) A. marina (149,15 %)
Keragaman jenis tumbuhan di setiap tipe tambak termasuk rendah seperti tertera pada Tabel 2.
Tabel 2. Indeks Keragaman Jenis Tumbuhan pada Setiap Tipe Tambak
Komposisi Jenis Fauna
Ditambak B ditemukan jenis-jenis fauna yang dapat dibedakan ke dalam tujuh golongan. Banyaknya jenis serangga dan Crustacea tertinggi ditemukan pada tambak B, sedangkan jenis burung dan ikan yang terbanyak terdapat pada tambak C. Adapun jenis reptilia, amphibia dan mamalia relatif sedikit di setiap tipe tambak atau bahkan tidak ditemukan (Tabel 3).
Tabel 3. Keragaman Jenis Fauna di Setiap Tipe Tambak
Klasifikasi Nilai Keragaman Jenis
A B C D
Burung 16 18 19 12
Mamalia 1 1 1 -
Reptilia 2 2 1 1
Crustacea 4 6 5 5
Amphibia - 1 - 1
Ikan 3 5 6 3
Serangga 17 32 19 14
Fauna Darat
Pengamatan fauna pada pagi hari (Tabel 4) menunjukkan bahwa tambak B memiliki keragaman jenis burung tertinggi, sedangkan nilai keragaman jenis yang terendah terdapat di tambak D. Kelimpahan jenis burung tertinggi terdapat pada tambak C yang mempunyai keragaman jenis yang relatif sama dengan tambak A. Kelimpahan, jumlah jenis dan keragaman jenis serangga tertinggi terdapat pada tambak B, sedangkan yang terendah di tambak D. Pengamatan fauna pada siang hari (Tabel 4), terinventarisir mamalia jenis garangan jawa dan reptilia jenis kadal kebun di tambak A, B dan C dengan kelimpahan yang relatif rendah.
Sedangkan pengamatan fauna pada sore hari menunjukan bahwa reptilia jenis ular kadut belang ditemukan pada tambak A dan B, dan ular tambak pada tambak D. Tambak B mempunyai keragaman, jumlah dan kelimpahan jenis serangga tertinggi, sedangkan tambak D yang terendah.
Tabel 4. Keragaman, Kelimpahan dan Jumlah Jenis Fauna Darat di Setiap Tipe Tambak pada Pengamatan Pagi, Siang, Sore dan Malam Hari.
Waktu Penga- matan
Klasifi- kasi
Tipe Tambak
A B C D
Jlh n H’ Jlh n H’ Jlh n H’ Jlh n H’
Pagi Burung 13 77 213 18 89 2,51 14 117 2.,13 11 47 2,06 Serangga 8 53 204 12 127 2,40 7 49 1,87 7 84 1,83
Siang Mamalia 1 3 0 1 5 0 1 2 0 - - -
reptilia 1 3 0 1 1 0 1 1 0 - - -
Sore Burung 11 22 226 8 22 1,92 12 35 2,38 8 15 1,93
Malam Reptilia 1 2 0 1 4 0 - - - 1 6 0
Biota Perairan
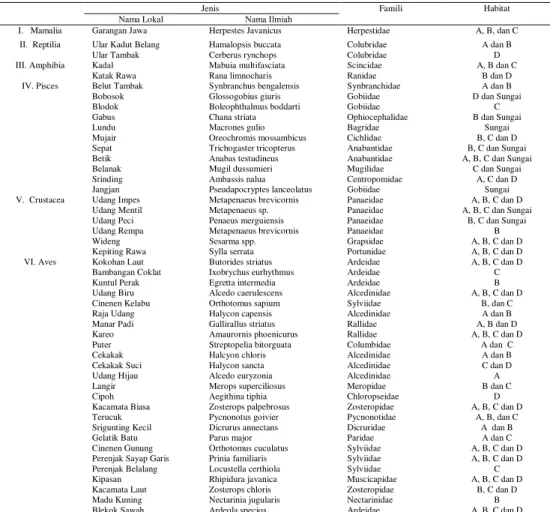
Keragaman jenis ikan tertinggi dijumpai pada tambak C dan terendah pada tambak A. Tambak B mempunyai keragaman, jumlah dan kelimpahan jenis udang tertinggi. Keragaman jenis plankton tertinggi dijumpai pada tambak D dan terendah pada tambak C. Tambak A memiliki kelimpahan benthos tertinggi, sedangkan kelimpahan yang terendah terdapat pada tambak D. (Tabel 5).
Tabel 5. Keragaman Kelimpahan dan Jumlah Jenis Fauna Perairan di Setiap Tipe Tambak
Klasifikasi Tipe Tambak
A B C D
Jlh n H’ Jlh n H’ Jlh n H’ Jlh n H’
Ikan 3 32 0,81 5 16 1,51 6 54 1,72 3 18 0,91
Udang 2 35 0,47 4 134 1,09 3 118 0,89 3 84 0,88
Plankton 16 7332 2,36 17 14017 2,12 10 17467 1,64 24 11968 2,83
Benthos 3 2417 1,09 1 1272 0 1 1527 0 1 636 0
Keterangan = Jlh : Jumlah jenis; n : Kelimpahan; H’ : Keragaman jenis
Fauna Yang Hidup di Darat dan Air
Pada tambak B dan D ditemukan amphibia jenis katak rawa. Keragaman jenis Crustacea tertinggi dijumpai pada tambak B dan terendah pada tambak A (Tabel 6). Jenis Crustacea yang ditemukan, antara lain kepiting rawa dan wideng.
Tabel 6. Keragaman, Kelimpahan dan Jumlah Jenis Fauna yang Hidup di Darat dan Air.
Klasifikasi
Tipe Tambak
A B C D
Jlh n H’ Jlh n H’ Jlh n H’ Jlh n H’
Rana limnocharis - - - 1 1 0 - - - 1 4 0 Crustacea 2 9 0,53 2 23 0,69 2 21 0.66 2 23 0,54 Keterangan = Jlh : Jumlah jenis; H' : Keragaman Jenis; n : Kelimpahan
Tabel 7. Jenis-jenis Fauna beserta Habitatnya yang terinventarisir selama periode Penelitian .
Jenis Famili Habitat
Nama Lokal Nama Ilmiah
I. Mamalia Garangan Jawa Herpestes Javanicus Herpestidae A, B, dan C II. Reptilia Ular Kadut Belang
Tabel 8. Jenis-Jenis Serangga beserta Habitatnya yang Terinventarisir selama Periode Penelitian.
Klasifikasi Ordo Famili Jenis Habitat
Araneae Araneidae A, B, C dan D
Coleoptera Carabidae B
Languriidae B, dan C
Noteridae A, B, C dan D
Ostomidae C
Scarabaeidae B, dan C
Staphylinidae B dan D
Diptera Anisopodidae B dan D
Acroiceridae A, B, dan C
Culicidae A, B, C dan D
Simulidae B, dan C
Tanyderidae Xylomyd sp. A, B, C dan D
Xylomyidae A dan B
Hemiptera Gerridae A
Homoptera Aleyrodidae Cicada sp. C
Cicadidae B
Cicadellidae A, B, C dan D
Cercopidae B
Psyllidae Apis sp. A, B, C dan D
Hymenoptera Apidae A, B, C dan D
Braconidae Formica sp. A, B, dan C
Formicidae A, B, C dan D
Ichneumonidae B
Pompilidae A, B, C dan D
Isoptera Rhinotermitidae Danaus B Lepidoptera Danaidae plexippus B, dan C Geometridae Alsopnila A, B, C dan D
Notodontidae pometaris B
Odonata Coenagrionidae A, B, dan C
Libellulidae A dan B
Orthoptera Acrididae A dan B
Grylidae B
Kualitas Perairan
Tabel 9. Parameter Fisika dan Kimia Perairan di Setiap Tipe Tambak.
Parameter Tipe Tambak
A B C D
Fisika
Salinitas ‰ 10,37 6,44 6,47 5,75
Suhu (0C) 32,20 32,44 31,00 30,00
Kimia
PH 7,50 6,55 6,50 7,55
DO (ppm) 8,55 8,13 11,83 10,46
Nitraty (ppm) 0,06 0,10 0,06 0,07
Orthofosfat (ppm) 0,06 0,04 0,02 0,04
Pembahasan
Komposisi dan Struktur Tegakan
Tambak A, B dan C didominasi oleh jenis A. marina. Tambak D mempunyai keragaman jenis tertinggi pada tingkat pohon, sedangkan pada tingkat pancang dan semai tergolong rendah, yang didominasi oleh jenis R. mucronata dan cenderung membentuk tegakan monokultur. Tambak A mempunyai pola tegakan yang lebih teratur daripada tambak B. Jenis A. marina pada tambak A lebih terpusat di tengah secara berkelompok dan dikelilingi oleh R. mucronata, sedangkan A. marina dan R. mucronata di tambak B tersebar secara acak. Keragaman jenis tegakan di setiap tipe tambak termasuk rendah. Tambak C mempunyai keragaman jenis terendah pada tingkat pertumbuhan pohon, pancang dan semai yang cenderung membentuk tegakan monokultur jenis A. marina. Kecilnya nilai indeks keragaman ini disebabkan oleh jumlah jenis yang sedikit dengan tingginya kelimpahan jenis tertentu, tetapi mempunyai ketersebaran jenis yang rendah. Jenis A. marina mempunyai diameter rata-rata, tinggi rata-rata dan kerapatan pohon terbanyak daripada jenis lain. Hal ini disebabkan karena jenis tersebut termasuk jenis intoleran yang cepat tumbuh dan mampu beradaptasi dengan kondisi habitat setempat. Secara keseluruhan tiap jenis mempunyai tinggi rata-rata dan diamater rata-rata relatif sama. Hal ini didukung oleh pernyataan Samingan (1972) bahwa hutan mangrove merupakan vegetasi yang agak seragam.
Komposisi jenis Fauna
Fauna Darat
serangga yang lebih rendah daripada tambak B, diduga adanya komposisi tegakan campuran berpola teratur, menyebabkan rendahnya ketersebaran sumber makanan, sehingga terjadi kompetisi dan pemangsaan interjenis maupun intrajenis yang tinggi. Hal ini akan berpengaruh terhadap keragaman jenis fauna lainnya. Tambak C memiliki keragaman jenis burung yang sama dengan tambak A dan memiliki kelimpahan yang lebih tinggi daripada tambak A, tetapi tingkat ketersebarannya rendah sehingga terjadi dominasi jenis tertentu yang mengakibatkan kompetisi antarjenis lebih tinggi dalam memperoleh sumber makanan yang sama. Tambak D mempunyai keragaman jenis serangga terendah. Hal ini disebabkan oleh komposisi tegakan yang cenderung didominasi oleh jenis R. mucronata sehingga ketersediaan sumber makanan kurang beraneka dan hanya disukai oleh jenis serangga tertentu, selanjutnya akan mempengaruhi keragaman jenis burung. Pengamatan fauna pada siang hari, ditemukan satu jenis fauna Herpestes javanicus yang bersifat karnivora. Kelimpahan jenis tersebut terbanyak dijumpai pada tambak B. Hal ini disebabkan adanya jenis ikan tertentu pada perairan tersebut seperti ikan gabus, sepat rawa dan mujair yang lebih disukai sebagai sumber makanannya. Disamping itu, letak tambak B berdekatan dengan areal semak belukar yang cenderung disukai sebagai tempat tinggalnya. Di tambak A, B dan C ditemukan jenis reptilia kadal kebun (Mabulia multifasciata) yang cenderung rendah kelimpahannya, diduga karena jenis ini kurang mampu beradaptasi terhadap kondisi habitat mangrove.
Pengamatan fauna pada sore hari menunjukkan bahwa tambak C memiliki keragaman jenis burung tertinggi. Aktivitas burung dalam mencari makanan pada sore hari mulai menurun sedangkan persaingan dalam mencari tempat beristirahat mulai naik. Tambak C merupakan habitat yang paling disukai burung sebagai tempat beristirahat. Jenis A. marina sebagai tempat bertengger yang mendominasi petak tersebut cenderung berdiameter besar, bertajuk lebat dan bercabang kokoh, sehingga mampu menopang jenis burung terutama burung air. Adapun tambak B mempunyai keragaman jenis burung terendah, disebabkan petak tersebut kurang disukai sebagai tempat beristirahat dan lebih sesuai sebagai tempat mencari makan yang ditunjukan oleh tingginya keragaman jenis burung pada pagi hari daripada sore hari di petak tersebut.
Pengamatan fauna pada malam hari menunjukkan bahwa kelimpahan jenis reptilia tertinggi ditemukan pada tambak D. Hal ini dipengaruhi oleh jenis tumbuhan yang cenderung didominasi oleh jenis R. mucronata. Jenis ini cenderung mempunyai ciri yang khas, terdiri dari beberapa batang dalam satu rumpun dan bentuk tajuk cenderung membulat. Kondisi ini lebih disukai jenis reptilia sebagai tempat berlindung dari predator. Keragaman jenis serangga tertinggi terdapat pada tambak B. Hal ini sesuai dengan komposisi jenis tumbuhan yang merupakan tegakan campuran berpola acak, memberikan sumber makanan yang lebih beraneka. Tambak A mempunyai keragaman jenis serangga yang lebih rendah daripada tambak C. Hal ini diduga adanya pengaruh faktor lingkungan yang kurang sesuai seperti kecepatan angin, suhu dan kelembaban udara.
Biota Perairan