POTENSI DAN KARAKTERISASI RUMPUT LAUT
Gracilaria
verrucosa
(
BELOW STANDARD
)
SEBAGAI BAHAN BAKU
BIOETANOL
SOPAN SABILI
DEPARTEMEN TEKNOLOGI HASIL PERAIRAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER
INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul “Potensi dan Karakterisasi Rumput Laut Gracilaria verrucosa (Below Standard) sebagai Bahan Baku Bioetanol” adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka dibagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
SOPAN SABILI. Potensi dan Karakterisasi Rumput Laut Gracilaria verrucosa (Below Standard) sebagai Bahan Baku Bioetanol. Dibimbing oleh UJU dan DESNIAR.
Potensi rumput laut G. verrucosa (below standard) sebagai sumber bahan baku bioetanol telah diteliti. Rumput laut dengan kualitas below standard secara struktur memiliki karakteristik lebih rapuh dan kristalinitas yang lebih rendah, namun rumput laut ini memiliki kadar agar tidak berbeda jauh dengan G. verrucosa yang memenuhi standard pabrik (meet standard). Hasil hidrolisis tepung rumput laut G. verrucosa (meet standard dan below standard)menghasilkan gula pereduksi masing-masing sebesar 105,47 mg/mL dan 94,78 mg/mL dengan jenis gula yaitu arabinosa, glukosa, maltosa dan xilosa. Fermentasi bioetanol dilakukan menggunakan hidrolisat rumput laut G. verrucosa (below standard)dengan khamir Saccharomyces cerevisiae AL IX. Profil pertumbuhan khamir S. cerevisiae AL IX menunjukkan penurunan pada hari ke-2 dan naik kembali pada hari ke-3. Produksi bioetanol dengan khamir S. cerevisiae AL IX menghasilkan kadar etanol optimum pada hari ke-4 mencapai 0,285%. Pengukuran gula pereduksi pada kultur fermentasi mengalami penurunan jumlah gula pereduksi hingga akhir fermentasi. Kata kunci: Below standard, bioetanol, biomassa, hidrolisis, rumput laut
ABSTRACT
SOPAN SABILI. Potency and Characterization of Seaweed Gracilaria verrucosa (Below Standard) as Bioethanol Biomass. Supervised by UJU and DESNIAR.
The potential of seaweed G. verrucosa (below standard) as a bioethanol feedstock was investigated. The seaweed of below standard quality structurally had a more fragile characteristic and lower crystallinity, nevertheless the seaweed had agar content not much different by G. verrucosa that meet industry standard (meet standard). The reducing sugar of hydrolysis G. verrucosa (meet standard and below standard) seaweed powder were 105.47 mg/mL and 94.78 mg/mL respectively, with the monosaccharides contents i.e. arabinose, glucose, maltose and xylose. Bioethanol production was conducted using hydrolysate of G. verrucosa (below standard)with fermentant yeast Saccharomyces cerevisiae AL IX. Growth profile by S. cerevisiae AL IX starter showed decreasing up to day 2nd and increasing at day 3rd. Bioethanol production by S. cerevisiae AL IX showed the optimum yield of ethanol production at day 4th which reached 0.285%. Measurement of reducing
sugar at fermentation culture by yeast starter showed decreasing yields of reducing sugar up to the end of fermentation.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah, dan pengutipan tersebut tidak merugikan kepentingan IPB
POTENSI DAN KARAKTERISASI RUMPUT LAUT
Gracilaria
verrucosa
(
BELOW STANDARD
)
SEBAGAI BAHAN BAKU
BIOETANOL
SOPAN SABILI
Skripsi
Sebagai salah satu syarat untuk memperoleh gelar Sarjana Perikanan pada
Departemen Teknologi Hasil Perairan
DEPARTEMEN TEKNOLOGI HASIL PERAIRAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
INSTITUT PERTANIAN BOGOR BOGOR
KATA PENGANTAR
Puji dan syukur penulis ucapkan kehadirat Allah SWT atas segala limpahan rahmat dan hidayah-Nya sehingga penulis dapat menyelesaikan karya ilmiah yang berjudul “Potensi dan Karakterisasi Rumput Laut Gracilaria verrucosa (Below Standard) sebagai Bahan Baku Bioetanol”, sebagai salah satu syarat untuk menyelesaikan pendidikan Sarjana di Departemen Teknologi Hasil Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor. Penulis mengucapkan terima kasih kepada semua pihak yang telah membantu dalam proses penulisan karya ilmiah ini, terutama kepada:
1 Dr Eng Uju, SPi MSi dan Dr Desniar, SPi MSi selaku dosen pembimbing yang telah memberikan penulis pengarahan agar karya ilmiah ini dapat diselesaikan.
2 Prof Dr Ir Joko Santoso, MSi selaku Ketua Departemen Teknologi Hasil Perairan dan mewakili dari Komisi Pendidikan Departemen yang telah memberikan motivasi dan saran.
3 Dr Ir Iriani Setyaningsih, MS selaku dosen penguji yang telah memberikan motivasi dan saran.
4 Dr Dwi Setyaningsih, Mbak Neli dan Kak Lia dari Surfactant and Bioenergy Research Center (SBRC), Institut Pertaninan Bogor atas segala fasilitas dan bantuannya.
5 Dr Yopi dari Lembaga Ilmu Pengetahuan Indonesia (LIPI) atas segala fasilitas dan bantuannya.
6 Orang tua atas doa dan kasih sayang serta seluruh keluarga dan teman-teman THP 48 atas kerja samanya.
Penulis menyadari bahwa masih banyak kekurangan dalam penulisan karya ilmiah ini. Penulis mengharapkan kritik dan saran yang dapat membangun selama penelitian dilakukan. Semoga tulisan ini bermanfaat bagi pihak-pihak yang membaca dan membutuhkan.
Bogor, Januari 2016
DAFTAR ISI
DAFTAR GAMBAR ... xvi
DAFTAR LAMPIRAN ... xvi
PENDAHULUAN ... 1
Latar Belakang ... 1
Perumusan Masalah ... 2
Tujuan Penelitian ... 2
Manfaat Penelitian ... 2
Ruang Lingkup Penelitian ... 2
METODE PENELITIAN ... 3
Waktu dan Tempat ... 3
Bahan dan Alat ... 3
Prosedur Penelitian ... 3
Prosedur Analisis ... 5
Analisis Data ... 9
HASIL DAN PEMBAHASAN ... 9
Karakteristik Rumput Laut Gracilaria verrucosa ... 9
Karakteristik Tepung Rumput Laut ... 10
Karakteristik Hidrolisat Tepung Rumput Laut ... 14
Produksi Bioetanol dengan S. cerevisiae AL IX ... 15
KESIMPULAN DAN SARAN ... 19
Kesimpulan ... 19
Saran ... 19
DAFTAR PUSTAKA ... 20
LAMPIRAN ... 24
DAFTAR GAMBAR
1 Diagram alir penelitian ... 4
2 Visualisasi rumput laut G. verrucosa (meet standard dan below standard) ... 10
3 Histogram komposisi kimia tepung rumput laut G. verrucosa (meet standard dan below standard) ... 10
4 Spektrum FTIR tepung rumput laut G. verrucosa (meet standard dan below standard) ... 12
5 Mikrograf rumput laut G. verrucosa (meet standard dan below standard) ... 13
6 Histogram gula pereduksi dan jenis gula hidrolisat rumput laut G. verrucosa (meet standard dan below standard) ... 14
7 Produksi bioetanol, gula pereduksi dan profil pertumbuhan S. cerevisiae AL IX ... 16
8 Jalur Embden-Meyerhoff-Parnas (EMP) ... 18
DAFTAR LAMPIRAN
1 Kurva standard galaktosa I ... 252 Kurva satndard galaktosa II ... 25
3 Komposisi pereaksi Dinitrosalisilik (DNS) ... 25
4 Komposisi media yang digunakan ... 26
1
PENDAHULUAN
Latar Belakang
Energi merupakan salah satu sumber kebutuhan yang sangat penting untuk manusia. Konsumsi terbesar energi banyak diserap untuk kegiatan industri dan transportasi. Peningkatan produksi minyak dunia pada tahun 2013 sebesar 550.000 barel/hari, jumlah ini setengah lebih kecil dari pertumbuhan konsumsi minyak dunia yang mencapai 1,4 juta barel/hari (BP 2014), berdasarkan hal tersebut diperkirakan cadangan energi fosil di dunia akan habis dalam 30-50 tahun lagi (Chang et al. 2010). Bioetanol merupakan salah satu alternatif pengganti atau subtitusi bahan bakar minyak fosil, karena memiliki kandungan oksigen mencapai 35% yang menghasilkan pembakaran lebih sempurna, nilai oktan tinggi hingga 118 dan mengandung gas emisi karbon monoksida lebih rendah hingga 19-25% (Indartono 2005). Produksi bioetanol tahun 2003 mencapai 14,682 ton, meningkat lebih dari empat kali lipat hingga tahun 2012 yang memproduksi bioetanol sebesar 61,752 ton (BP 2014).
Bioetanol selain dapat dihasilkan oleh bahan baku yang memiliki sumber gula dan pati juga dapat diproduksi dari tanaman yang mengandung selulosa. Saat ini, pengembangan bioetanol berbahan dasar lignoselulosa dari tanaman banyak mendapat perhatian karena tidak berkompetisi dengan bahan pangan. Salah satu sumber daya hayati yang potensial sebagai sumber bahan baku bioetanol adalah rumput laut. Berbeda dengan sumber material lignoselulosa terestrial, rumput laut memiliki lapisan lignin yang hampir tidak ada, karena keberadaan lignin sebagai pelindung selulosa dan hemiselulosa akan menghambat pemecahan polisakarida (Yanagisawa et al. 2011). Lapisan lignin yang tipis ini akan berpengaruh dalam menghasilkan gula yang tinggi, sehingga dapat digunakan sebagai sumber gula untuk menghasilkan bioetanol. Adams et al. (2009) melaporkan bahwa rumput laut memiliki potensi produksi bioetanol 4-23 kali lebih tinggi dibandingkan dengan umbi-umbian dan gula bit.
2
Pemanfaatan rumput laut below standard ini perlu dilakukan, karena rumput laut G. verrucosa memiliki kadar karbohidrat mencapai 60 % (Kumar et al. 2013). Bioetanol dihasilkan dari cairan hasil fermentasi gula dari sumber karbohidrat dengan bantuan mikroorganisme (Hapsari dan Pramashinta 2013). Fermentasi dengan bahan baku rumput laut umumnya menggunakan khamir S. cerevisiae. Jenis khamir ini merupakan mikroorganisme yang mampu mengkonversi gula menjadi etanol melalui fermentasi. Penggunaan rumput laut segar sebagai sumber bahan baku produksi bioetanol, sebelumnya telah dilakukan, seperti Gracilaria verrucosa (Kumar et al. 2013), Eucheuma cottonii (Setyaningsih et al. 2012) dan Kappaphycus alvarezii (Meinita et al. 2012), sedangkan penggunaan rumput laut below standard masih sangat sedikit dilaporkan. Oleh karena itu, penelitian tentang karakterisasi dan pemanfaatan rumput laut below standard sebagai sumber bahan baku produksi bioetanol penting untuk dilakukan.
Perumusan Masalah
Rumusan masalah penelitian ini adalah penggunaan dan pemanfaatan bahan baku rumput laut sebagai sumber material lignoselulosa yang melimpah dan dapat digunakan untuk produksi bioetanol serta potensi rumput laut G. verrucosa (below standard)yang kurang dimanfaatkan.
Tujuan Penelitian
Tujuan dari penelitian ini adalah membandingkan karakteristik kimia dan fisik tepung rumput laut serta hidrolisat tepung rumput laut G. verrucosa (meet standard dan below standard) dan melihat produksi bioetanol dari hidrolisat rumput laut G. verrucosa menggunakan khamir S. cerevisiae AL IX.
Manfaat Penelitian
Manfaat penelitian ini adalah memberikan informasi mengenai pemanfaatan dan karakteristik rumput laut G. verrucosa (meet standard dan below standard) sebagai bahan baku produksi bioetanol.
Ruang Lingkup Penelitian
3
METODE PENELITIAN
Waktu dan Tempat
Penelitian ini dilaksanakan pada Januari hingga Juli 2015. Penelitian dilakukan di beberapa laboratorium yaitu Laboratorium Mikrobiologi Hasil Perairan, Departemen Teknologi Hasil Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor, Laboratorium Nanoteknologi Pangan dan Pertanian, Badan Penelitian dan Pengembangan Pertanian, Laboratorium Kimia, Pusat Studi Biofarmaka, Laboratorium Kimia Analitik, Balai Penelitian Ternak, Bogor, Laboratorium Terpadu, Institut Pertanian Bogor, Laboratorium Bioetanol, Surfactant and Bioenergy Research Center, Institut Pertanian Bogor, Laboratorium Mikrobiologi, Pusat Antar Universitas, Institut Pertanian Bogor, dan Laboratorium Bioteknologi, Lembaga Ilmu Pengetahuan Indonesia, Bogor.
Bahan dan Alat
Bahan utama yang digunakan adalah rumput laut G. verrucosa (meet standard dan below standard) diperoleh dari perairan Pontang, Serang, Banten, biakan khamir yang digunakan adalah khamir S. cerevisiae AL IX diperoleh dari Surfactant and Bioenergy Research Center (SBRC). Bahan lainnya yang digunakan adalah akuades, H2SO4 3%, kertas saring, pereaksi dinitrosalisilik (DNS)
(Sigma-Aldrich®), kalsium oksida (CaO), arang aktif, media Potato Dextrose Agar (PDA) (Oxoid), media Potato Galactose Agar (PGA) (Merck®), dan media Yeast
Galactose Maltose Peptone (YGMP) (Merck®).
Alat yang digunakan untuk preparasi sampel adalah wadah, trash bag, saringan. Alat yang digunakan untuk karakterisasi bahan baku adalah timbangan, erlenmeyer, beaker glass dan oven. Alat yang digunakan untuk hidrolisis asam adalah erlenmeyer, beaker glass, kertas lakmus, saringan, botol jar, filtrasi vakum dan autoklaf (Hirayama Hiclave HVE-50, Tokyo-Japan). Alat yang digunakan untuk analisis glukosa adalah tabung reaksi, penangas air, gelas ukur, corong pisah dan Spektrofotometer UV-Vis (Hitachi U-2900, Illinois-USA). Alat yang digunakan untuk fermentasi adalah cawan, botol, distilator dan Density Meter (DMA 4500 M, Graz-Austria) untuk mengukur kadar bioetanol. Alat pendukung lain yang digunakan adalah magnetic stirrer (C-Mag HS 7, Selangor-Malaysia) Fourier Transform Infra Red (FTIR) (Bruker Tensor 37, Massachusetts-USA), Scanning Electron Microscopy (SEM) (Zeiss Evo MA-1, Jena-Germany) X-Ray Diffraction (XRD) (Bruker D8 Advance, Massachusetts-USA) dan Ultras Fast Liquid Chromatograph (UFLC) (Shimadzu LC-20AB, Kyoto-Japan).
Prosedur Penelitian
4
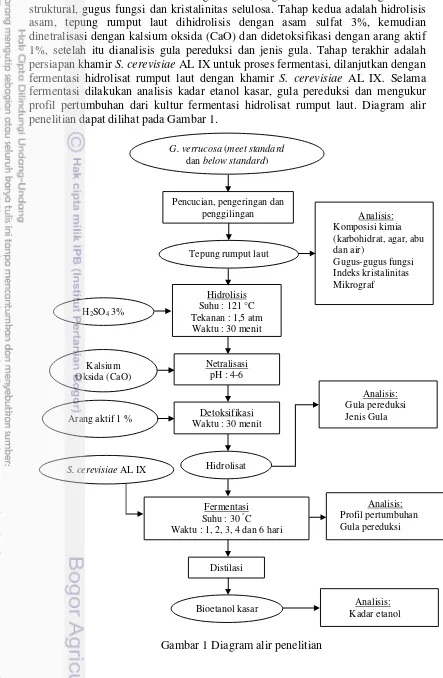
laut G. verrucosa dianalisis kadar lignoselulosa, agar, air dan abu serta gambaran struktural, gugus fungsi dan kristalinitas selulosa. Tahap kedua adalah hidrolisis asam, tepung rumput laut dihidrolisis dengan asam sulfat 3%, kemudian dinetralisasi dengan kalsium oksida (CaO) dan didetoksifikasi dengan arang aktif 1%, setelah itu dianalisis gula pereduksi dan jenis gula. Tahap terakhir adalah persiapan khamir S. cerevisiae AL IX untuk proses fermentasi, dilanjutkan dengan fermentasi hidrolisat rumput laut dengan khamir S. cerevisiae AL IX. Selama fermentasi dilakukan analisis kadar etanol kasar, gula pereduksi dan mengukur profil pertumbuhan dari kultur fermentasi hidrolisat rumput laut. Diagram alir penelitian dapat dilihat pada Gambar 1.
Gambar 1 Diagram alir penelitian
Tepung rumput laut
S. cerevisiae AL IX Hidrolisat
Fermentasi Suhu : 30 °C
Waktu : 1, 2, 3, 4 dan 6 hari G. verrucosa (meet standard
5
Pembuatan Tepung Rumput Laut
Rumput laut G. verrucosa (meet standard dan below standard)diambil dari perairan Pontang, Serang, Banten. Perlakuan awal pada rumput laut yaitu pencucian dengan air untuk membersihkan dari garam dan pengotor yang terdapat pada rumput laut. Rumput laut dijemur dengan sinar matahari (5-7 hari) hingga kering. Rumput laut yang sudah kering kemudian digiling menggunakan grinder hingga menjadi tepung.
Hidrolisis Asam (Fakhrudin et al. 2014; Setyaningsih et al. 2012 dan Meinita
et al. 2012a)
Asam yang digunakan untuk hidrolisis adalah asam sulfat (H2SO4) 3%.
Sebanyak 30 g rumput laut kering dilarutkan dalam 100 mL larutan asam sulfat 3%. Hidrolisis dilakukan melalui dua tahap, tahap pertama 15 g sampel dimasukkan ke dalam 100 mL dipanaskan dengan suhu 121 °C, tekanan 1,5 atm selama 15 menit dan dilanjutkan dengan tahap kedua yaitu penambahan 15 g sampel ke dalam botol dan dipanaskan dengan suhu 121 °C, tekanan 1,5 atm selama 15 menit. Hidrolisat dinetralisasi dengan larutan kalsium oksida jenuh (1:3 dalam akuades) hingga mencapai kisaran pH 4-6 dan disaring menggunakan saringan nylon mesh, kemudian disaring kembali dengan menggunakan filtrasi vakum. Tahap selanjutnya yaitu dilakukan detoksifikasi pada hidrolisat dengan arang aktif konsentrasi 1% (w/v), setelah ditambahkan, diaduk dengan stirrer dengan suhu 30 °C selama 30 menit dan hidrolisat dipisahkan kembali dengan menggunakan filtrasi vakum. Persiapan Biakan Saccharomyces cerevisiae AL IX (Setyaningsih et al. 2012)
Isolat khamir S. cerevisiae AL IX disegarkan menggunakan media Potato Galactose Agar (PGA) dan diinkubasi dengan suhu 30 °C selama 48-72 jam. Selanjutnya khamir S. cerevisiae AL IX dipindahkan ke media Yeast Galactose Maltose Peptone (YGMP) kemudian diinkubasi selama 24 jam dengan suhu inkubasi 30 °C. Setelah 24 jam khamir starter S. cerevisiae AL IX telah siap digunakan untuk proses fermentasi hidrolisat rumput laut.
Fermentasi dengan Saccharomyces cerevisiae AL IX (Fakhrudin et al. 2014) Proses fermentasi dilakukan dengan mengatur waktu fermentasi dalam kondisi anaerob. Mendekati proses fermentasi, hidrolisat dipasteurisasi pada suhu 70 °C selama 15 menit. Hidrolisat yang telah disiapkan kemudian ditambahkan Starter. S. cerevisiae AL IX sebanyak 10% (v/v) dimasukkan kedalam hidrolisat rumput laut dengan volume kultur 100 mL. Hidrolisat difermentasikan selama 6 hari. Pengukuran dilakukan pada hari ke-1, 2, 3, 4 dan 6 hari dengan parameter yang diuji meliputi gula pereduksi, kadar etanol kasar dan profil pertumbuhan.
Prosedur Analisis
Kadar Lignoselulosa (Van Soest et al. 1991)
6
dipanaskan. Sampel diekstrak selama 60 menit dari mulai mendidih. Sampel yang telah diekstrak disaring menggunakan cawan masir yang telah ditimbang sebelumnya. Residu dibilas menggunakan air panas dan aseton. Cawan masir beserta residu dikeringkan pada oven 105 oC selama ± 4 jam sampai beratnya konstan. Sampel diangkat dan didinginkan dalam desikator, kemudian cawan ditimbang. Kadar NDF dihitung menggunakan rumus sebagai berikut :
NDF (%) = x 100% Keterangan : A = Berat sampel awal (g)
B = Berat cawan kosong (g)
C = Berat cawan dengan sampel yang sudah dikeringkan (g) Analsisi ADF diawali dengan memasukkan sampel sebanyak 1 g ke dalam gelas piala 600 mL dan ditambahkan 100 mL larutan Acid Detergent Solution (ADS) kemudian dipanaskan. Sampel diekstrak selama 60 menit dari mulai mendidih. Sampel disaring menggunakan cawan masir yang telah ditimbang sebelumnya. Residu dibilas menggunakan air panas dan aseton. Cawan masir beserta residu dikeringkan pada oven 105 oC selama ± 4 jam hingga beratnya konstan. Sampel diangkat dan didinginkan dalam desikator, kemudian ditimbang. Kadar ADF dan hemiselulosa dihitung menggunakan rumus sebagai berikut :
ADF (%) = x 100% Keterangan : A = Berat sampel awal (g)
B = Berat cawan kosong (g)
C = Berat cawan dengan sampel yang sudah dikeringkan (g) Kadar hemiselulosa dihitung dengan mengurangi kadar NDF dengan kadar ADF. Kadar hemiselulosa dihitung menggunakan rumus sebagai berikut :
Hemiselulosa (%) = NDF - ADF
Analisis selulosa merupakan lanjutan dari analisis ADF. Sampel hasil analisis ADF yang sudah ditimbang (C) ditambah larutan asam sulfat 72% sampai terendam selama 3 jam, kemudian residu dibilas menggunakan air panas dan aseton. Residu dikeringkan pada oven 105 oC selama ± 4 jam sampai beratnya stabil, kemudian diangkat dan didinginkan dalam desikator. Cawan dikeluarkan dari desikator dan ditimbang. Kadar selulosa dihitung menggunakan rumus sebagai berikut :
Selulosa (%) = x 100% Keterangan : A = Berat sampel awal (g)
B = Berat sampel analisa ADF (g)
C = Berat cawan dengan sampel yang sudah dikeringkan (g) Analisis kadar lignin merupakan lanjutan dari analisa selulosa. Sampel hasil analisis selulosa yang sudah dikeringkan dipanaskan dalam tanur dengan suhu ±
7
600 oC. Cawan didinginkan dalam desikator dan ditimbang. Kadar lignin dihitung
menggunakan rumus sebagai berikut :
Lignin (%) = x 100% Keterangan : A = Berat sampel awal (g)
B = Berat sampel analisa selulosa (g)
C = Berat cawan dengan sampel yang sudah dikeringkan (g) Kadar Air (SNI 01-2891-1992)
Perhitungan kadar air dan kadar abu mengacu pada SNI 01-2891-1992. Penghitungan kadar air dimulai dengan menimbang sampel sebanyak 1-2 g, kemudian dimasukkan ke dalam cawan porselin yang telah dikeringkan pada suhu 105 oC selama 3 jam hingga beratnya konstan. Kemudian sampel didinginkan dalam desikator. Kadar air dihitung dengan rumus sebagai berikut:
Kadar air (%) = x 100% berat kering Keterangan : I1 = Berat sampel awal (g)
I2 = Berat sampel setelah dikeringkan (g)
Kadar Abu (SNI 01-2891-1992)
Penghitungan kadar abu dimulai dengan menimbang sampel 2-3 g pada cawan porselin yang telah diketahui beratnya. Sampel dipijarkan dalam tanur pada suhu 550 oC sampai pengabuan sempurna. Cawan didinginkan dalam desikator
kemudian ditimbang sampai bobot tetap. Kadar abu dihitung dengan rumus sebagai berikut:
Kadar abu (%) = x 100% Keterangan : I1 = Berat sampel + cawan (g)
I2 = Berat cawan kosong (g)
Ekstraksi Agar (modifikasi Guerrrero et al. 2014)
Sebanyak 10 g tepung rumput laut G. verrucosa (meet standard dan below standard) dididihkan dalam air distilasi (300 mL) selama 1 jam dan kemudian larutan dipisahkan menggunakan nylon mesh. Larutan dibiarkan menjadi gel pada suhu kamar dan dibekukan satu malam. Setelah itu, gel agar dilelehkan dengan air kran dan dikeringkan dengan sinar matahari. Rendemen agar merupakan perhitungan persentase dari sampel.
Analisis Gugus Fungsi
Rumput laut G. verrucosa (meet standard dan below standard) kering dianalisis gugus-gugus fungsi dengan menggunakan Fourier Transform Infra Red (FTIR). Sampel sebanyak 0,02 g dicampurkan dengan kalium bromida (KBr) dan ditekan hingga berbentuk film tipis. Spektrum dilakukan selama rentang 1800-600 cm-1 dengan empat pemindaian yang tercatat pada resolusi 4 cm-1.
8
Analisis Indeks Kristalinitas
Analisia indeks kristalinitas biomassa dianalisis dengan X-Ray Diffraction (XRD). Sampel yang digunakan adalah G. verrucosa (meet standard dan below standard) yang dikeringkan. Sampel sebanyak 0,3 g tepung rumput laut dimasukkan pada plat plastik dan dilakukan pada mode transmisi menggunakan Cu-Kα radiasi, dioperasikan pada 40 kV dan 35 mA, kemudian dipindai melalui difraksi sudut (2�) = 5-80o dengan ukuran tahapan 0.02o. Indeks kristalinitas (CrI) ditentukan dengan metode Segal et al. (1959):
CrI (%) = x 100%
Keterangan : I200 = Puncak intensitas kristalin dari selulosa I pada 2� = 22.5o
Iam = Intensitas amorpos pada 2� = 18.7o
Analisis Mikrograf
Analisia mikrograf dilakukan terhadap rumput laut G. verrucosa (meet standard dan below standard) yang dikeringkan. Sampel sebanyak 0,2 g disebar pada sebuah piringan metal-silinder dengan pita karbon dan dilapisi dengan emas di bawah vakum. Karakterisasi mikrograf dilakukan dengan menggunakan instrumen Scanning Electron Microscopy (SEM) yang dioperasikan pada 14 kV tegangan percepatan.
Analisis Gula Pereduksi (Miller 1959)
Sebanyak 1 mL hidrolisat (dengan pengenceran) dan 3 mL pereaksi Dinitrosalisilik (DNS) dimasukkan ke dalam tabung reaksi. Kemudian sampel dan pereaksi ditempatkan dalam penangas air mendidih selama 15 menit. Setelah itu, dibiarkan dingin pada suhu ruang. Kadar gula pereduksi diukur dengan Spektrofotometer UV-Vis pada panjang gelombang 540 nm.
Analisis Profil Gula Sederhana
Hidrolisat sebanyak 450 µL dipipet langsung ke dalam vial High Performance Liquid Chromatograph (HPLC) (tanpa pengenceran) dan dianalisis dengan instrumen alat Ultra Fast Liquid Chromatograph (UFLC) dengan kolom Aminex-HPX 87H dengan flow rate 0,6 mL/min dan volume injeksi sebesar 20 µL selama 20 menit. Standard gula yang digunakan adalah xilosa, mannosa, maltosa, glukosa, arabinosa, dan galaktosa. Perhitungan dilakukan dengan Reflective Index Detector (RID-20A) sehingga terbentuk kromatogram.
Kadar Etanol (Fakhrudin et al. 2014)
Kultur fermentasi hasil produksi bioetanol hidrolisat rumput laut dituang ke dalam labu leher kemudian dididihkan dalam distilator hingga beberapa menit dan menghasilkan uap air. Uap air yang dihasilkan diambil sebanyak 10 mL disimpan dalam wadah untuk dianalisis. Analisis kadar bioetanol kasar ditentukan dengan menggunakan Density Meter DMA 4500 M.
I200- Iam
9
Profil Pertumbuhan (FDA 2001)
Saccharomyces cerevisiae AL IX diaktivasi dengan dilakukan pengambilan 2 ose kultur kerja yang diinokulasikan ke dalam tabung reaksi yang berisi 10 mL media Yeast Maltose Galactose Peptone (YMGP), kemudian diinkubasi selama 24 jam dengan suhu 30 °C. Setelah 24 jam, hasil aktivasi masing-masing diambil sebanyak 1 mL (10%) dan diinokulasikan ke dalam tabung reaksi yang berisi 9 mL hidrolisat rumput laut G. verrucosa (below standard) yang telah dipasteurisasi, kemudian diinkubasi selama 24 jam dengan suhu 30 °C. Pembuatan profil pertumbuhan dilakukan dengan memindahkan 1 mL suspensi pengenceran 100 dengan mikropipet ke dalam larutan 9 mL Buffered Peptone Water (BPW) 0,1% untuk mendapatkan pengenceran 10-1. Pengenceran dilakukan 10-2, 10-3, 10-4,
hingga 10-5. Sebanyak 1 mL suspensi dari setiap pengenceran dimasukkan ke dalam cawan petri secara duplo dan ditambahkan 15 mL sampai 20 mL Plate Count Agar (PCA) yang sudah didinginkan pada setiap cawan yang sudah berisi suspensi, untuk khamir larutan PCA ditambahkan sebanyak 0,1% kloramfenikol. Setelah itu, diinkubasi pada temperatur 35 °C selama 48 jam sampai dengan 72 jam.
Analisis Data
Analisis data diperlukan untuk mendapatkan kesimpulan dari percobaan yang dilakukan. Analisis yang digunakan pada penelitian ini adalah analisis secara deskriptif. Data diolah dengan menggunakan aplikasi Microsoft Excel 2013 untuk mendapatkan nilai rata-rata dan standar deviasi.
HASIL DAN PEMBAHASAN
Karakteristik Rumput Laut Gracilaria verrucosa
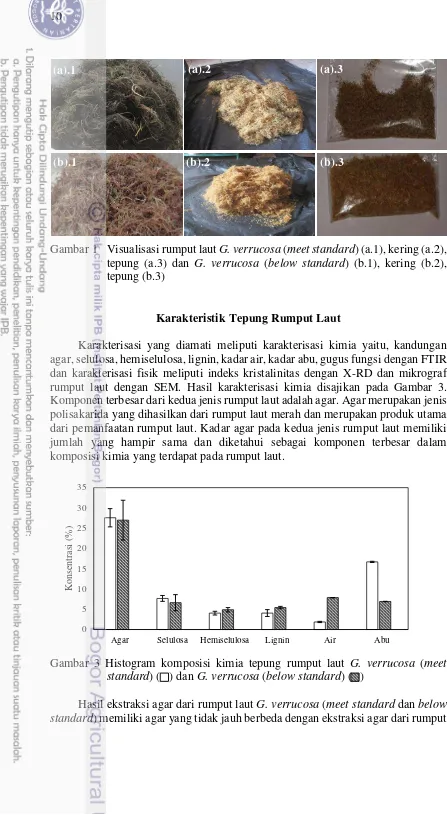
Gracilaria verrucosa merupakan rumput laut dari jenis Rhodophyta dan digunakan dalam produksi agar. Perbedaan kualitas rumput laut G. verrucosa diduga dapat mempengaruhi agar yang dihasilkan, sehingga banyak industri pengolah rumput laut memilah-memilah kualitas rumput laut yang didapatkan dari para petambak rumput laut G. verrucosa. Rumput laut yang dihasilkan petani memiliki dua jenis kualitas berbeda, yaitu meet standard dan below standard. Perbedaan dua jenis rumput laut G. verrucosa (meet standard dan below standard) dapat dilihat pada Gambar 2.
10
Gambar 1 Visualisasi rumput laut G. verrucosa (meet standard)(a.1), kering (a.2), tepung (a.3) dan G. verrucosa (below standard) (b.1), kering (b.2), tepung (b.3)
Karakteristik Tepung Rumput Laut
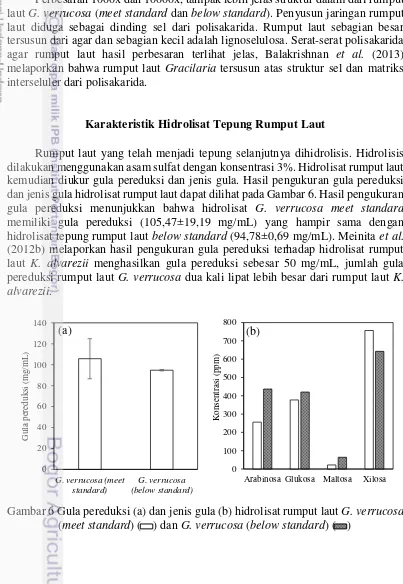
Karakterisasi yang diamati meliputi karakterisasi kimia yaitu, kandungan agar, selulosa, hemiselulosa, lignin, kadar air, kadar abu, gugus fungsi dengan FTIR dan karakterisasi fisik meliputi indeks kristalinitas dengan X-RD dan mikrograf rumput laut dengan SEM. Hasil karakterisasi kimia disajikan pada Gambar 3. Komponen terbesar dari kedua jenis rumput laut adalah agar. Agar merupakan jenis polisakarida yang dihasilkan dari rumput laut merah dan merupakan produk utama dari pemanfaatan rumput laut. Kadar agar pada kedua jenis rumput laut memiliki jumlah yang hampir sama dan diketahui sebagai komponen terbesar dalam komposisi kimia yang terdapat pada rumput laut.
Gambar 3 Histogram komposisi kimia tepung rumput laut G. verrucosa (meet standard)( ) dan G. verrucosa (below standard) ( )
Hasil ekstraksi agar dari rumput laut G. verrucosa (meet standard dan below standard)memiliki agar yang tidak jauh berbeda dengan ekstraksi agar dari rumput
0
Agar Selulosa Hemiselulosa Lignin Air Abu
11
laut G. verrucosa dengan larutan alkali yang dilakukan Kumar et al. (2013) dengan rendemen agar sebesar 32,53±1,42%. Dalam hal ini, ekstraksi dengan pelarut alkali atau tanpa alkali tidak mempengaruhi jumlah rendemen agar rumput laut G. verrucosa. Marinho-Soriano dan Bourret (2005) melakukan ekstraksi agar dari jenis rumput laut Gracilaria melaporkan bahwa selain musim dan faktor lingkungan, suhu dan salinitas dapat mempengaruhi rendemen agar yang dihasilkan. Pengaruh lain yang dapat mempengaruhi adalah umur panen rumput laut, Santika et al. (2014) melaporkan bahwa umur panen 50, 60, dan 70 hari menghasilkan rendemen agar yang relatif sama, sedangkan viskositas dan kekuatan gel tertinggi pada panen 60 hari dan menurun ketika umur panen 70 hari. Perbedaan rendemen agar diduga karena bahan baku diambil dari perairan yang sama sehingga faktor-faktor pendukung seperti lingkungan, suhu, salinitas dan umur panen tidak berpengaruh.
Kadar selulosa, hemiselulosa dan lignin rumput laut G. verrucosa (meet standard dan below standard)masing-masing memiliki kadar yang hampir sama. Santi et al. (2012) melaporkan bahwa kadar lignin dari rumput laut hijau Ulva lactuca dan Chaeteomorpha crassa masing-masing memiliki kadar lignin 2,9% dan 4%, hasil tidak jauh berbeda didapatkan dari kadar lignin G. verrucosa (meet standard dan below standard) masing-masing sebesar 4% dan 5,41%. Masalah utama sumber biomassa lignoselulosa sebagai bahan baku produksi bioetanol adalah kandungan lignin yang berperan sebagai inhibitor dalam konversi polisakarida menjadi monosakarida atau disakarida. Musatto dan Teixeira (2010) menyatakan bahwa lignin berikatan dengan selulosa dan hemiselulosa dan berfungsi memberikan tingkat kekerasan pada dinding sel. Hsu et al. (2010) melaporkan pengukuran kadar lignin jerami sebesar 14,9 %, hasil ini tiga kali lebih besar dari lignin rumput laut dan menyatakan bahwa lignin yang rendah dapat mengindikasikan bahwa sampel tidak perlu melalui proses pretreatment dan hidrolisis dalam keadaan yang ekstrim.
Hasil berbeda karakterisasi tepung rumput laut G. verrucosa (meet standard dan below standard) diketahui pada kadar air dan abu. Pengukuran menunjukkan bahwa rumput laut G. verrucosa (meet standard) memiliki kadar air lebih rendah daripada kadar abu, sedangkan rumput laut G. verrucosa (below standard) memiliki kadar air dan abu sama. Kumar et al. (2013) melaporkan bahwa rumput laut Gracilaria verrucosa memiliki kadar air lebih rendah (4,4±0,15%) dibandingkan kadar abu (12,73±0,15%) dan menyatakan bahwa perbedaan kadar air dan abu pada rumput laut dapat dipengaruhi oleh penurunan salinitas akibat dari mengalirnya air tawar yang lebih banyak pada periode tertentu. Harun et al. (2013) melaporkan kadar air yang lebih tinggi dipengaruhi oleh pengeringan yang tidak berjalan sempurna. Perbedaan kualitas rumput laut dapat mempengaruhi kadar air dan abu yang diduga disebabkan salinitas dan pengeringan rumput laut yang dilakukan.
12
Gambar 4 Spektrum FTIR tepung rumput laut G. verrucosa (meet standard) (a) dan G. verrucosa (below standard)(b)
Pita gelombang yang lemah pada kedua jenis rumput laut G. verrucosa (meet standard dan below standard) ditunjukkan pada pita gelombang 1600 cm-1,
merupakan peregangan dari cincin aromatik yang menandakan adanya lignin (Kumar et al. 2009), kemudian ditemukan juga puncak gelombang 1434 cm-1 dengan ikatan C-H dan pada 1022 cm-1 dengan ikatan gugus C-O-C yang menunjukkan karakteristik selulosa (Siddhanta et al. 2011) kemudian pita 1040-1060 cm-1 menandakan regangan gugus C-O hemiselulosa (Zhao et al. 2008).
Puncak panjang gelombang kuat ditunjukkan pada 930 cm-1, 895 cm-1 dan 775 cm-1 untuk rumput laut G. verrucosa (meet standard). Puncak 930 cm-1
menandakan getaran gugus C-O-C pada polimer subunit 3,6-anhidro-α-L-galaktosa (Barros et al. 2013), hal ini diduga rumput laut G. verrucosa (meet standard) memiliki polimer subunit 3,6-anhidro-α-L-galaktosa yang tinggi. Marinho-Soriano dan Bourret (2005) melaporkan bahwa polisakarida agar dengan subunit 3,6-anhidrogalaktosa mengindikasikan agar memiliki gel yang kuat. Pita yang muncul pada 897 cm-1 menandakan gugus β-linked glukosa yang merupakan polimer dalam selulosa (Uju et al. 2015), hal ini diduga telah terjadi pemecahan selulosa menjadi glukosa pada rumput laut G. verrucosa (meet standard).
Rumput laut G. verrucosa (below standard) menunjukkan puncak kuat pada panjang gelombang 870 cm-1, 830 cm-1 dan 750 cm-1. Daerah panjang gelombang
800-850 cm-1 merupakan polisakarida agar dari gugus sulfat, pita 830 cm-1 yang muncul menandakan grup sulfat dari 2-O-D-galaktosa (Barros et al. 2013), sedangkan puncak pada 872 cm-1 menandakan keberadaan grup sulfat dari
6-O-galaktosa (Kumar et al. 2012). Grup sulfat galaktosa yang terdeteksi pada rumput laut G. verrucosa (below standard) mengindikasikan bahwa agar mengandung sulfat yang tinggi. Marinho-Soriano dan Bourret (2005) melaporkan bahwa agar dengan sulfat yang tinggi memiliki gel yang lemah.
13
Puncak kuat pada rumput laut G. verrucosa (meet standard dan below standard)masing-masing memiliki ditandai pada panjang gelombang 775 cm-1 dan 750 cm-1. Kumar et al. (2009) menyatakan bahwa panjang gelombang tersebut mengindikasikanratio dari kristalin selulosa. Puncak 775 cm-1 diduga memiliki kristalinitas selulosa lebih tinggi dibandingkan dengan puncak 750 cm-1. Hasil
pengukuran diduga bahwa rumput laut G. verrucosa (meet standard)memiliki agar dengan gel yang kuat dan kristalinitas tinggi, sedangkan rumput laut G. verrucosa (below standard)memiliki agar dengan gel lemah dan kristalinitas rendah.
Rumput laut G. verrucosa (meet standard dan below standard) masing-masing diukur indeks kristalinitas selulosa. Kristalinitas adalah tingkat keteraturan selulosa dengan sifat seperti kristal yang dicirikan dengan derajat kristalinitas yang terdapat pada sumber bahan baku (Pari et al. 2011). Taherzadeh dan Karimi (2008) melaporkan bahwa susunan mikrofibril yang rapat membuat selulosa menjadi tidak mudah untuk dihidrolisis oleh enzim. Selulosa memiliki bentuk struktur kristalin dan polimer penyusun lainnya seperti hemiselulosa dan lignin memiliki bentuk struktur amorf. Indeks kristalinitas dihitung dengan metode Segal et al. (1959).
Pengukuran indeks kristalinitas menunjukkan bahwa kristalinitas selulosa rumput laut G. verrucosa (meet standard dan below standard) sebesar 74,5% dan 54,8% berturut-turut. Selulosa alam mempunyai kristalinitas tinggi karena ikatan inter dan intramolekuler hidrogen yang membuat selulosa tidak larut dan sulit untuk dihidrolisis (Puri 1984). Perbedaan kristalinitas rumput laut diduga karena umur panen rumput laut G. verrucosa. Kamat et al. (2002) melaporkan bahwa tanaman hemp dengan umur panen 30 hari memiliki kadar α-selulosa 37% dan kadar lignin 14%, meningkat pada umur panen 90 hari dengan kadar α-selulosa 44% dan kadar lignin 11%. Hasil ini didukung dengan analisis gugus fungsi bahwa kristalinitas rumput laut G. verrucosa (meet standard) lebih tinggi dibandingkan G. verrucosa (below standard). Indeks kristalinitas yang lebih rendah dapat mempermudah proses pemecahan menjadi gula-gula sederhana.
Karakterisasi fisik dilakukan untuk mengetahui perbedaan struktur pada masing-masing rumput laut G. verrucosa. Hasil mikrograf rumput laut G. verrucosa (meet standard dan below standard)dapat dilihat pada Gambar 5.
Gambar 5 Mikrograf rumput laut G. verrucosa (meet standard)(a) dan G. verrucosa (below standard) (b)
(a.) 100x
(b.)1000x (a.) 1000x
14
Hasil mikrograf menunjukkan bahwa permukaan pada masing-masing bahan memiliki perbedaan pada perbesaran 100x, rumput laut G. verrucosa (meet standard)memiliki struktur thallus yang lebih kompak dengan susunan epidermis melapisi thallus rumput laut, sedangkan rumput laut G. verrucosa (below standard) memiliki struktur permukaan thallus yang sudah tidak halus. Perbedaan struktur luar diduga karena rumput laut G. verrucosa (below standard) merupakan jenis kualitas rendah dengan penanganan pasca panen yang tidak baik, disebabkan penjemuran yang terkena air tawar dan menyebabkan permukaan luar jenis rumput laut rusak.
Perbesaran 1000x dan 10000x, tampak lebih jelas struktur dalam dari rumput laut G. verrucosa (meet standard dan below standard). Penyusun jaringan rumput laut diduga sebagai dinding sel dari polisakarida. Rumput laut sebagian besar tersusun dari agar dan sebagian kecil adalah lignoselulosa. Serat-serat polisakarida agar rumput laut hasil perbesaran terlihat jelas, Balakrishnan et al. (2013) melaporkan bahwa rumput laut Gracilaria tersusun atas struktur sel dan matriks interseluler dari polisakarida.
Karakteristik Hidrolisat Tepung Rumput Laut
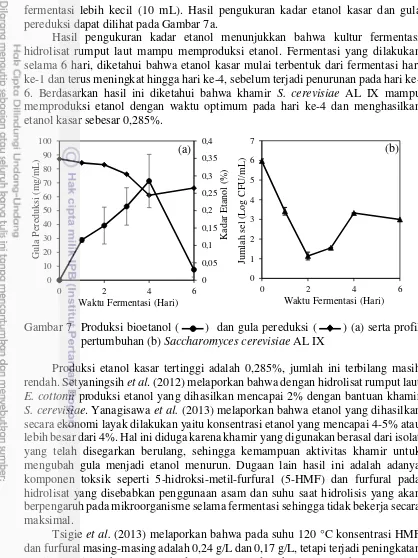
Rumput laut yang telah menjadi tepung selanjutnya dihidrolisis. Hidrolisis dilakukan menggunakan asam sulfat dengan konsentrasi 3%. Hidrolisat rumput laut kemudian diukur gula pereduksi dan jenis gula. Hasil pengukuran gula pereduksi dan jenis gula hidrolisat rumput laut dapat dilihat pada Gambar 6. Hasil pengukuran gula pereduksi menunjukkan bahwa hidrolisat G. verrucosa meet standard memiliki gula pereduksi (105,47±19,19 mg/mL) yang hampir sama dengan hidrolisat tepung rumput laut below standard (94,78±0,69 mg/mL). Meinita et al. (2012b) melaporkan hasil pengukuran gula pereduksi terhadap hidrolisat rumput laut K. alvarezii menghasilkan gula pereduksi sebesar 50 mg/mL, jumlah gula pereduksi rumput laut G. verrucosa dua kali lipat lebih besar dari rumput laut K. alvarezii.
15
Yadav et al. (2011) melaporkan bahwa hidrolisis dengan asam sulfat dapat meningkatkan konsentrasi gula pada hidrolisat mencapai 2 kali lipat. Hal ini diduga karena penggunaan konsentrasi asam sulfat dan waktu hidrolisis yang dilakukan. Penggunaan dua tahap hidrolisis dan konsentrasi asam sulfat 3% yang dipanaskan dengan suhu 121 oC mampu menghasilkan gula pereduksi yang lebih banyak.
Penggunaan asam sulfat juga dapat menghasilkan produk samping, Jonsson et al. (2013) melaporkan bahwa lignin akan membentuk komponen fenolik akibat dari hidrolisis asam, sedangkan Meinita et al. (2012b) melaporkan bahwa konsentrasi asam sulfat dan reaksi yang lebih lama dapat menurunkan komponen gula dan menghasilkan produk samping seperti 5-hidroksil-metil-furfural (5-HMF) dan asam levulinik.
Gambar 6b menunjukkan bahwa jenis gula pada hidrolisat tepung rumput laut G. verrucosa diantaranya, arabinosa, glukosa, maltosa, dan xilosa. Gula arabinosa hidrolisat tepung rumput laut G. verrucosa (meet standard)diperkirakan memiliki konsentrasi yang lebih rendah dibandingkan glukosa, sedangkan gula arabinosa dan glukosa pada G. verrucosa (below standard) diperkirakan memiliki konsentrasi yang hampir sama. Maltosa dan xilosa memiliki konsentrasi yang hampir sama pada kedua jenis rumput laut sebagai gula terendah dan tertinggi.
Rumput laut sebagai sumber biomassa lignoselulosa terdiri dari selulosa, hemiselulosa dan lignin memiliki peran dalam penyusunan jenis monosakarida. Selulosa berperan dalam kontribusi glukosa, kemudian hemiselulosa berperan dalam kontribusi gula pentosa dan heksosa yaitu xilosa, arabinosa dan glukosa (Musatto dan Teixeira 2010). Salah satu komponen terbesar dari rumput laut adalah agar, Guerrero et al. (2014) melaporkan bahwa agar yang tersusun dari 3,6 anhidrogalaktosa berkontribusi dalam menghasilkan galaktosa. Sarfat et al. (2013) menggunakan rumput laut merah (E. cottonii) sebagai bahan baku produksi bioetanol, hasil pengukuran jenis gula menunjukkan bahwa gula tertinggi adalah galaktosa, sedangkan hasil dari pengukuran jenis gula menunjukkan tidak adanya jenis galaktosa. Hal ini diperkirakan bahwa galaktosa dalam rumput laut telah mengalami perubahan dalam bentuk yang lebih sederhana akibat dari hidrolisis dengan panas dan asam, sehingga hanya jenis gula glukosa, arabinosa, xilosa dan maltosa yang terdeteksi pada hidrolisat rumput laut.
Produksi Bioetanol dengan S. cerevisiae AL IX
16
fermentasi lebih kecil (10 mL). Hasil pengukuran kadar etanol kasar dan gula pereduksi dapat dilihat pada Gambar 7a.
Hasil pengukuran kadar etanol menunjukkan bahwa kultur fermentasi hidrolisat rumput laut mampu memproduksi etanol. Fermentasi yang dilakukan selama 6 hari, diketahui bahwa etanol kasar mulai terbentuk dari fermentasi hari 1 dan terus meningkat hingga hari 4, sebelum terjadi penurunan pada hari ke-6. Berdasarkan hasil ini diketahui bahwa khamir S. cerevisiae AL IX mampu memproduksi etanol dengan waktu optimum pada hari ke-4 dan menghasilkan etanol kasar sebesar 0,285%.
Gambar 7 Produksi bioetanol ( ) dan gula pereduksi ( ) (a) serta profil pertumbuhan (b) Saccharomyces cerevisiae AL IX
Produksi etanol kasar tertinggi adalah 0,285%, jumlah ini terbilang masih rendah. Setyaningsih et al. (2012) melaporkan bahwa dengan hidrolisat rumput laut E. cottonii produksi etanol yang dihasilkan mencapai 2% dengan bantuan khamir S. cerevisiae. Yanagisawa et al. (2013) melaporkan bahwa etanol yang dihasilkan secara ekonomi layak dilakukan yaitu konsentrasi etanol yang mencapai 4-5% atau lebih besar dari 4%. Hal ini diduga karena khamir yang digunakan berasal dari isolat yang telah disegarkan berulang, sehingga kemampuan aktivitas khamir untuk mengubah gula menjadi etanol menurun. Dugaan lain hasil ini adalah adanya komponen toksik seperti 5-hidroksi-metil-furfural (5-HMF) dan furfural pada hidrolisat yang disebabkan penggunaan asam dan suhu saat hidrolisis yang akan berpengaruh pada mikroorganisme selama fermentasi sehingga tidak bekerja secara maksimal.
Tsigie et al. (2013) melaporkan bahwa pada suhu 120 °C konsentrasi HMF dan furfural masing-masing adalah 0,24 g/L dan 0,17 g/L, tetapi terjadi peningkatan masing-masing sebesar 1,12 g/L dan 0,43 g/L pada suhu 135 °C. Suhu yang terlalu tinggi akan menyebabkan gula pada substrat terdegradasi menjadi komponen lain seperti 5-hidroksi-metil-furfural (5-HMF) dan furfural yang dapat bersifat toksik untuk mikrorganisme fermentasi. Meinita et al. (2012b) melaporkan bahwa HMF dan asam levulinik mulai menghambat produksi etanol pada konsentrasi 0,05 g/L dan 0,5 g/L. Cho et al. (2011) menyatakan bahwa hidrolisat lignoselulosa seperti rumput laut tidak hanya mengandung gula-gula yang dapat difermentasi, tetapi juga
17
mengandung komponen seperti furan, asam lemah dan komponen fenolik lainnya yang dapat menghambat fermentasi etanol.
Penurunan kadar etanol yang terjadi pada hari ke-6 diduga karena semakin berkurangnya nutrisi dan pertumbuhan sel. Idral et al. (2012) melaporkan bahwa ketersediaan nutrisi dalam medium sudah mulai berkurang sehingga khamir mengubah etanol menjadi asam asetat yang menyebabkan penurunan kadar etanol. Penggunaan jenis mikroba lain dalam hidrolisat rumput laut telah banyak dilakukan seperti Sarfat et al. (2013) yang menggunakan khamir Pachysolen tannophilus, tetapi etanol yang dihasilkan dari jenis khamir ini masih lebih rendah dari jenis khamir S. cerevisiae.
Gula pereduksi yang dihasilkan hidrolisat telah memenuhi syarat untuk dilakukan fermentasi yaitu sebesar 10%, Sebayang et al. (2006) melaporkan bahwa dengan kandungan gula 10-18% etanol yang dihasilkan dari fermentasi sudah cukup baik dengan kadar etanol 12%. Hasil penelitian ini menunjukkan bahwa gula pereduksi selama proses fermentasi menunjukkan grafik penurunan seiring dengan terbentuknya etanol. Hargreaves et al. (2013) melaporkan bahwa gula pereduksi awal sebelum fermentasi sebesar 80 g/L, ketika etanol mencapai 37 g/L, gula pereduksi turun menjadi 2 g/L dengan konversi gula menjadi etanol mencapai 98%. Gula pereduksi akan menurun ketika etanol telah terbentuk. Hal ini berbeda dengan yang dihasilkan pada penelitian ini, yaitu produksi etanol maksimum hanya 0,285%. Artinya konversi gula yang terjadi hanya sebesar 24,13%, yaitu dari 87 mg/mL pada awal fermentasi menjadi 66 mg/mL pada hari ke-4. Rendahnya konversi gula ini diduga yang menyebabkan rendahnya kadar etanol yang dihasilkan.
Profil pertumbuhan khamir memberikan gambaran mengenai faktor-faktor lingkungan yang mempengaruhi pertumbuhan suatu khamir seperti substrat, suhu lingkungan dan pH (Kusumaningati et al. 2013). Pengukuran jumlah koloni sel dengan melihat profil pertumbuhan dapat dilihat pada Gambar 7b. Grafik menunjukkan bahwa koloni sel awal S. cerevisiae AL IX pada medium YMGP sebesar 7,483 Log CFU/mL, kemudian dilakukan fermentasi dan pengukuran jumlah koloni sel pada medium hidrolisat rumput, jumlah koloni sel pada hari ke-0 mengalami penurunan hingga hari ke-2, kemudian mulai terjadi kenaikan jumlah koloni hingga hari ke-4 dan terjadi penurunan kembali pada hari ke-6 dengan jumlah koloni sebesar 2,996 Log CFU/mL.
Setyaningsih et al. (2012) melakukan pengukuran jumlah sel dengan dua metode yaitu adaptasi cepat dan lambat, melaporkan bahwa jumlah sel S. cerevisiae dengan adaptasi cepat, memiliki konsentrasi sel 6,90x107 sel/mL pada hari ke-1, kemudian terjadi penurunan pada hari ke-2 dengan konsentrasi sel 5,35x107 sel/mL
dan meningkat kembali pada hari ke-3 7,55x107 sel/mL, selanjutnya terjadi penurunan hingga akhir fermentasi. Pengukuran adaptasi lambat menunjukkan konsentrasi sel 6,90x107 sel/mL pada hari ke-1, kemudian terjadi penurunan pada
18
Hasil pengukuran jumlah koloni sel jika dihubungkan dengan produksi etanol dengan S. cerevisiae AL IX, mempunyai perbedaan karena etanol kasar yang diproduksi mengalami kenaikan hingga hari ke-4, sedangkan jumlah koloni sel mengalami penurunan pada hari ke-2. Sarfat et al. (2013) melaporkan S. cerevisiae AL IX merupakan khamir yang telah teradaptasi pada media yang mengandung galaktosa, hasil pengukuran jumlah total sel dan etanol yang dilakukannya memiliki korelasi, jumlah total sel yang semakin tinggi, konsentrasi etanol yang dihasilkan meningkat. Timson (2007) melaporkan khamir menghasilkan enzim galaktosa mutarotase dan UDP-4-epimerase yang dapat memetabolisme galaktosa hingga menjadi glukosa, melalui glikolisis mengubah glukosa menjadi etanol.
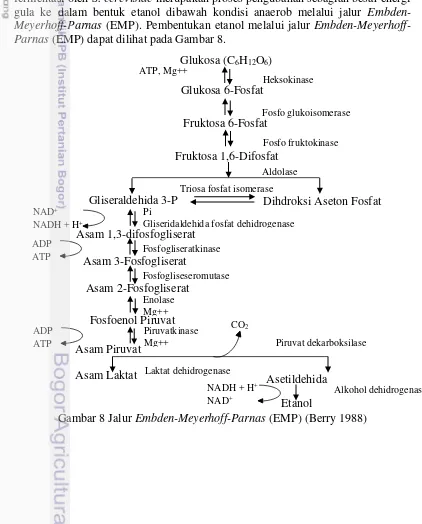
Penurunan total sel pada pengukuran profil pertumbuhan dengan hidrolisat rumput laut diduga S. cerevisiae AL IX merupakan khamir teradaptasi dengan media galaktosa, sedangkan hidrolisat tidak menghasilkan jenis galaktosa sehingga khamir diduga mengalami adaptasi dengan media glukosa, xilosa dan arabinosa. Borines et al. (2013) dan Mohagheghi et al. (2002) melaporkan bahwa proses fermentasi oleh S. cerevisiae merupakan proses pengubahan sebagian besar energi gula ke dalam bentuk etanol dibawah kondisi anaerob melalui jalur Embden-Meyerhoff-Parnas (EMP). Pembentukan etanol melalui jalur Embden-Meyerhoff-Parnas (EMP) dapat dilihat pada Gambar 8.
19
Jalur Embden-Meyerhoff-Parnas (EMP) merupakan jalur glikolisis yang dilalui oleh khamir S. cerevisiae untuk mengubah glukosa menjadi etanol. Glukosa dengan bantuan enzim-enzim mampu dipecah hingga menjadi piruvat, kemudian piruvat dipecah menjadi etanol dengan enzim piruvat dekarboksilase (PDC) dan alkohol dehidrogenase (ADH). Ndaba et al. (2014) melaporkan bahwa S. cerevisiae ketika difermentasi dalam media campuran glukosa dan xilosa, khamir tidak mampu memanfaatkan xilosa sebagai sumber karbon. Li et al. (2010) melaporkan bahwa khamir S. cerevisiae 424A (LNH-ST) mampu memanfaatkan gula-gula pentosa dan heksosa, dikarenakan jenis khamir ini merupakan hasil rekayasa genetik untuk memfermentasikan xilosa.
KESIMPULAN DAN SARAN
Kesimpulan
Rumput laut G. verrucosa (meet standard dan below standard) sebagai sumber biomassa produksi bioetanol secara karakteristik kimia dengan melihat hasil uji lignoselulosa, dan agar memiliki hasil yang hampir sama, namun berbeda untuk uji kadar air dan abu. Secara fisik rumput laut G. verrucosa (meet standard dan below standard) memiliki perbedaan struktural berdasarkan hasil mikrograf dan uji indeks kristalinitas. Gula pereduksi hidrolisat kedua jenis rumput laut memiliki jumlah yang hampir sama dan jenis gula sederhana pada hidrolisat diantaranya arabinosa, glukosa, maltosa dan xilosa. Fermentasi S. cerevisiae AL IX selama enam hari memiliki kadar etanol optimum pada hari ke-4 mencapai 0,285%.
Saran
20
DAFTAR PUSTAKA
Adams JM, Gallagher JA, Donnison IS. 2009. Fermentation study on Saccharina latissima for bioethanol production considering variable pre-treatment. Journal of Applied Phycology. 21: 569-574.
Balakrishnan CP, Jenifer P, Esakkilingam M. 2013. Algal documentation and phytochemical studies of red algae Gracilaria corticata of Manapad Coast, Tamil Nadu. Journal Pharmacognosy and Phytochemistry. 2(4): 193-197. Barros FCN, da Silva DC, Sombra VG, Maciel JS, Feitosa JPA, Freitas ALP, de
Paula RCM. 2013. Structural characterization of polysaccharide obtained from red seaweed Gracilaria caudata (J Agardh). Carbohydrate Polymers. 92: 598-603.
Berry DR. 1988. Physiology of Industrial Fungi. London (UK). Blackwell Scientific Pub;ications.
[BP] British Petroleum. 2014. BP Statictical Review of World Energy. London (UK): British Petroleum p.l.c.
Borines MG, de Leon RL, Cuello JL. 2013. Bioethanol production from the macroalgae Sargassum spp.. Bioresource Technology. 138: 22-29.
[BSN] Badan Standardisasi Nasional. 1992. Cara Uji Makanan dan Minuman SNI 01-2891-1992. Jakarta (ID): Badan Standardisasi Nasional.
Chang HN, Nag-Jong K, Kang J, Jeong CM. 2010. Biomass-derived volatile fatty acid platform for fuels and chemicals. Biotechnology and Bioprocess Engineering. 15: 1-10.
Cho DH, Soo-Jeong S, Bae Y, Park C, Yong HK. 2011. Ethanol production from acid hydrolysate based on the construction and demolition wood waste using Pichia stiptis. Bioresources Technology. 102: 4439-4443.
Fakhrudin J, Setyaningsih D, Rahayuningsih M. 2014. Bioethanol production from seaweed Eucheuma cottonii by neutralization and detoxification of acidic catalyzed hydrolysate. International Journal of Environment and Science Development. 5(5): 455-458.
[FAO] Food and Agriculture Organization. 2013. Social and Economic Dimensions of Carrageenan Seaweed Farming. Roma (IT): Food and Agriculture Organization.
[FDA] Food and Drug Administration. 2001. Biological Analytical Manual online. New Hampshire (US): Food, Drug and Administration.
Guerrero P, Etxabide A, Leceta I, Penalba M, de la Caba K. 2014. Extraction of agar from Gelidium sesquipedale (Rhodopyta) and surface characterization of agar based films. Carbohydrate Polymers. 99: 491-498.
21
mempercepat konversi minyak tanah ke bahan bakar nabati. Jurnal Teknologi Kimia dan Industri. 2(2): 240-245.
Hargreaves PI, Barcelos CA, da Costa ACA, Junior NP. 2013. Production of ethanol 3G from Kappaphycus alvarezii: evaluation of different process strategies. Bioresource Technology. 134: 257-263.
Harun M, Montolalu RI, Suwetja IK. 2013. Karakteristik fisika kimia karaginan rumput laut jenis Kappaphycus alvarezii pada umur panen yang berbeda di perairan Desa Tihengo Kabupaten Gorontalo Utara. Jurnal Media Teknologi Hasil Perairan. 1(1): 7-12.
Hsu TC, Gia-Luen G, Wen-Hua C, Wen-Song H. 2010. Effect of dilute acid pretreatment of rice straw on structural properties and enzymatic hydrolysis. Bioresource Technology. 101: 4907-4913.
Idral DD, Salim M, Mardiah E. 2012. Pembuatan bioetanol dari ampas sagu dengan proses hidrolisis asam dan menggunakan Saccharomyces cerevisiae. Jurnal Kimia Unand. 1(1): 34-39.
Indartono Y. 2005. Bioethanol, Alternatif Energi Terbarukan: Kajian Prestasi Mesin dan Implemntasi di Lapangan. Jakarta (ID): LIPI.
Jonsson LJ, Alriksson B, Nilvebrant NO. 2013. Bioconversion of lignocellulose: inhibitors and detoxification. Biotechnology for Biofuels. 6(16): 1-10. Kamat J, Roy DN, Goel K. 2002. Effect of harvesting age on the chemical
properties of hemp plants. Journal Wood Chemistry and Technology. 22(4): 285-293.
Kumar R, Mago G, Balan V, Wyman CE. 2009. Physical and chemical characterizations of corn stover and poplar solids resulting from leading pretreatment technologies. Bioresource Technology. 100: 3948-3962. Kumar S, Godiya CB, Siddhanta AK. 2012. Carrageenan from Sraconema
scinaioides (Gigartinales, Rhodophyta) of Indian waters. Carbohydrate Polymers. 87: 1657-1662.
Kumar S, Gupta R, Kumar G, Sahoo D, Kuhad RC. 2013. Biethanol production from Gracilaria verrucosa, a red alga, in a biorefinery approach. Bioresource Technology. 135: 150-156.
Kusumaningati MA, Nurhatika S, Muhibuddin A. 2013. Pengaruh konsentrasi inokulum bakteri Zymomonas mobilis dan lama fermentasi pada produksi etanol dari sampah sayur dan buah Pasar Wonokromo Surabaya. Jurnal Sains dan Semi Pomits. 2(2): 218-213.
Li BZ, Balan V, Ying-Jin Y, Dale BE. 2010. Process optimization to convert forage and sweet sorghum bagasse to ethanol based on ammonia fiber expansion (AFEX) preteatment. Bioresource Technology. 101: 1285-1292.
22
Meinita DNM, Ji-Young K, Gwi-Taek J, Koo HM, Park SM, Yong-Ki H. 2012a. Bioethanol production from the acid hydrolysate of the carrageenophyte Kappaphycus alvarezii (cottonii). Journal of Applied Phycology. 24: 857-862.
Meinita DNM, Yong-Ki H, Gwi-Taek J. 2012b. Detoxification of acidic catalyzed hydrolysate of Kappaphycus alvarezii (cottonii). Bioprocess Biosystem Engineering. 35:93-98.
Miller GL. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Journal Analytical Chemistry. 31(3): 426–428.
Mohagheghi A, Evans K, Yat-Chen C, Zhang M. 2002. Cofermentation of glucose, xylose and arabinose by genomic DNA-integrated xylose/arabinose fermenting strain of Zymomonas mobilis AX101. Journal of Applied Biochemistry and Biotechnology. 98-100: 885-899.
Musatto SI, Teixeira JA. 2010. Lignocellulose as raw material in fermentation processes. Journal of Applied Microbiology and Microbial Biotechnology. 897-907.
Ndaba B, Chiyanzu I, Marx S, Obiore G. 2014. Effect of Saccharomyces cerevisiae and Zymomonas mobilis on the co-fermentation of sweet shorgum bagasse hydrolysate pretreated under varying conditions. Biomass and Bioenergy 71: 350-356.
Pari G. 2011. Pengaruh selulosa terhadap struktur karbon arang, Bagian I: Pengaruh suhu karbonasi. Jurnal Penelitian Hasil Hutan. 29(1): 33-45.
Puri VP. 1984. Effect of crystallinity and degree of polymerization of cellulose on enzymatic saccharification. Biotechnology Bioengineering. 26:1219-1222. Santi RA, Sunarti TC, Santoso D, Triwisari DA. 2012. Komposisi kimia dan profil
polisakarida rumput laut hijau. Jurnal Akuatika. 3(2): 105-114.
Santika LG, Ma’ruf WF, Romadhon. 2014. Karakteristik agar rumput laut Gracilaria verrucosa budidaya tambak dengan perlakuan konsentrasi alkali pada umur panen yang berbeda. Jurnal Pengolahan dan Bioteknologi Hasil Perairan. 3(4): 98-105.
Sarfat MS, Rahayuningsih M, Suryani A, Setyaningsih D. 2013. Modifikasi fermentasi hidrolisat asam Eucheuma cottonii menjadi bioetanol menggunakan Saccharomyces cerevisiae dan Pachysolen tannophilus. Jurnal Teknologi Industri Pertanian. 23(3): 199-209.
Sebayang F. 2006. Pembuatan etanol dari molase secara fermentasi menggunakan sel Saccharomyces cerevisiae tang terimobilisasi pada kalsium alginat. Jurnal Teknologi Proses. 5(2): 75-80.
Segal L, Creely JJ, Martin AE, Conrad CM. 1959. An emprical method for estimating the degree of crystallinty of native cellulose using the x-ray diffractometer. Journal Textille Research. 29(10): 786-794.
23
bioethanol production. International Journal of Environment and Bioengineering. 3(2): 98-110.
Siddantha, AK, Chattbar MU, Mehta GK, Sanandiya ND, Kumar S, Oza MD, Prasad K, Meena R. 2011. The cellulose contents of Indian seaweeds. Journal of Applied Phycology. 23: 919-923.
Taherzadeh MJ, Karimi K. 2008. Pretreatment of lignoselluosic wastes to improve ethanol and biogas production: a review. International Journal Molecular Science. 9(9): 1621-1651.
Timson DJ. 2007. Galactose metabolism in Saccharomyces cerevisiae. Journal Dynamic Biochemistry, Process Biotechnology and Molecul Biology. 1(1): 63-73.
Tsigie YA, Chung-Hsan W, Huynh LH, Ismadji S, Yi-Hsu J. 2013. Bioethanol production from Yarrowia lipolytiva Po1g biomass. Bioresources Technology. 145: 210-216.
Uju, Wijayanta AT, Goto M, Kamiya N. 2015. Great potency of seaweed waste biomass from carrageenan industry for bioethanol production by peracetic acid-ionic liquid pretreatment. Biomass and Bioenergy. 81: 63-69.
Van Soest PJ, Robertson JB, Lewis BA. 1991. Methods for dietary fiber neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. Journal Dairy Science. 74: 3583-3597.
Yadav KS, Naseeruddin S, Prashanthi GS, Sateesh L, Rao LV. 2011. Bioethanol fermentation of concentrated rice straw hydrolysate using co-culture of Saccharomyces cerevisiae and Pichia stipitis. Bioresource Technology. 102: 6473-6478.
Yanagisawa M, Nakamura K, Aragi O, Nakasaki K. 2011. Production of high concentrations of bioethanol from seaweeds that contain easily hydrolyzable polysaccharides. Journal Process Biochemistry. 46: 2111-2116.
24
25
Lampiran 1 Kurva standard galaktosa I
Lampiran 2 Kurva standard galaktosa II
Lampiran 3 Komposisi pereaksi Dinitrosalisilik (DNS)
Dinitrosalisilik (DNS) SIGMA-ALDRICH® 10,6 gram
NaOH MERCK® 19,8 gram
Aquades 1416 mL
K Na Tartarat MERCK® 306 gram
Fenol MERCK® 7,6 gram
Na Metabisulfat MERCK® 8,3 gram
26
Lampiran 4 Komposisi media yang digunakan Potato Galactose Agar (PDA)
Potato 20 g/L Galactose MERCK® 10 g/L
Agar 20 g/L
Yeast Maltose Galactose Agae (YMGP) Yeast MERCK® 5 g/L
Maltose MERCK® 5 g/L Galactose MERCK® 10 g/L Peptone MERCK® 5 g/L
Lampiran 5 Contoh perhitungan gula pereduksi dan konversi gula menjadi etanol Contoh perhitungan gula pereduksi G. verrucosa (meet standard)
Konsentrasi = ((x Abs. Sampel + 0,2134) / 0,0045) x pengenceran (1000) / 1000 = (0,261 + 0,2134) / 0,0045 x 1000 / 1000
= 100,607 mg/mL
Contoh perhitungan konversi gula menjadi etanol
Konversi gula = ((gula perduksi H0 - gula pereduksi H1) / gula perduksi H0) x 100%
27
RIWAYAT HIDUP
Penulis dilahirkan di Tangerang pada tanggal 2 Juli 1993. Penulis merupakan anak ketiga dari tiga bersaudara dari pasangan Ayah bernama Dul Salim dan Ibu bernama Saidah. Pendidikan yang telah ditempuh penulis dimulai dari TK Nurul Huda pada tahun 1997 kemudian melanjutkan pendidikan di SDN 04 Pondok Aren pada tahun 1999 dan lulus pada tahun 2005. Penulis selanjutnya menempuh jenjang pendidikan menengah di SMPN 2 Pondok Aren dan lulus pada tahun 2008. Pendidikan selanjutnya ditempuh di SMAN 4 Kota Tangerang Selatan dan lulus pada tahun 2011. Penulis diterima sebagai mahasiswa di Departemen Teknologi Hasil Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor melalui jalur SNMPTN Undangan pada tahun 2011.