SKRIPSI
KAJIAN SENYAWA ANTIMIKROBA
BAKTERI ASAM LAKTAT ISOLAT ASI YANG BERPOTENSI SEBAGAI PROBIOTIK
Oleh:
IKE ROHMAWATI F 24053031
2010
DEPARTEMEN ILMU DAN TEKNOLOGI PANGAN FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR FAKULTAS TEKNOLOGI PERTANIAN
KAJIAN SENYAWA ANTIMIKROBA
BAKTERI ASAM LAKTAT ISOLAT ASI YANG BERPOTENSI SEBAGAI PROBIOTIK
SKRIPSI
Sebagai salah satu syarat untuk memperoleh gelar SARJANA TEKNOLOGI PERTANIAN Pada Departemen Ilmu dan Teknologi Pangan
Fakultas Teknologi Pertanian Institut Pertanian Bogor
Oleh:
IKE ROHMAWATI F 24053031
2010
DEPARTEMEN ILMU DAN TEKNOLOGI PANGAN FAKULTAS TEKNOLOGI PERTANIAN
Judul Skripsi : Kajian Senyawa Antimikroba Bakteri Asam Laktat Isolat ASI yang Berpotensi sebagai Probiotik
Nama : Ike Rohmawati NRP : F24053031
Menyetujui,
Dosen Pembimbing I Dosen Pembimbing II
(Dr. Ir. Lilis Nuraida, M.Sc.) (Siti Nurjanah, STP, M.Si.) NIP. 19621009.198703.2.002 NIP. 19760131.200501.2.001
Mengetahui, Ketua Departemen
(Dr. Ir. Dahrul Syah) NIP. 19650814.199002.1.001
Ike Rohmawati. F24053031. Kajian Senyawa Antimikroba Bakteri Asam Laktat Isolat ASI yang Berpotensi sebagai Probiotik. Di bawah bimbingan Lilis Nuraida dan Siti Nurjanah. 2010.
RINGKASAN
Senyawa antimikroba yang berasal dari bakteri asam laktat (BAL) berpotensi dapat menggantikan penggunaan pengawet sintetik yang digunakan dalam makanan. BAL dapat menghasilkan senyawa antimikroba seperti asam organik (laktat, asetat, dan propionat), hidrogen peroksida, diasetil, bakteriosin, maupun senyawa antimikroba lainnya. Diantara berbagai metabolit tersebut, bakteriosin merupakan senyawa antimikroba yang sangat menjanjikan untuk digunakan sebagai pengawet alami makanan.
Tujuan penelitian ini adalah melakukan seleksi bakteri asam laktat isolat ASI yang memiliki aktivitas penghambatan tinggi terhadap bakteri uji (Listeria monocytogenes), mengkaji senyawa antimikroba yang dihasilkan, mengetahui potensinya dalam menghasilkan senyawa bakteriosin, dan mengkaji pengaruh waktu inkubasi terhadap aktivitas antimikroba.
Penelitian ini terdiri dari dua tahap, yaitu : (1) seleksi awal isolat bakteri asam laktat (BAL) potensi probiotik yang memiliki aktivitas penghambatan tinggi terhadap bakteri Listeria monocytogenes dan (2) seleksi BAL yang berpotensi menghasilkan bakteriosin. Tahap kedua terbagi menjadi 4 bagian yaitu : (a) pengujian aktivitas antimikroba dengan metode kontak, (b) penentuan fase stasioner, (c) mempelajari pengaruh waktu inkubasi terhadap aktivitas antimikroba (inkubasi BAL sampai kira-kira awal fase stasioner dan membandingkannya dengan inkubasi BAL selama 24 jam), dan (d) pengujian aktivitas antimikroba protein terendapkan (presipitasi dengan ammonium sulfat).
Bakteri asam laktat (BAL) yang digunakan adalah BAL isolat ASI yang berpotensi sebagai probiotik sebanyak 24 isolat. Isolat-isolat tersebut antara lain : Lactobacillus pentosus (A7), Lactobacillus rhamnosus (A15, A23, A24, A29, A31, R21, R23, R24, R26, R34, B10, B16), Lactobacillus acidophillus1(A8, A22), Pediococcus pentosaceus2 (A16), Lactobacillus rhamnosus2 (B11) dan beberapa isolat yang belum diketahui genusnya (A14, A20, A27, R22, R25, R27, B13). Bakteri asam laktat (BAL) tersebut bersifat homofermentatif, kecuali isolat A20 yang bersifat heterofermentatif. Kultur bakteri uji yang digunakan adalah Listeria monocytogenes, Bacillus cereus, dan Staphylococcus aureus. Bakteri uji L. monocytogenes digunakan pada setiap tahapan sedangkan bakteri uji Bacillus cereus dan Staphylococcus aureus digunakan pada saat mempelajari pengaruh waktu inkubasi terhadap aktivitas antimikroba dan pengujian aktivitas protein terendapkan (presipitasi dengan ammonium sulfat).
Dari 12 isolat BAL yang diuji, terdapat 8 isolat BAL (A7, A15, A22, A24, R21, R24, R26, dan R27) yang berpotensi menghasilkan senyawa bakteriosin dengan waktu inkubasi BAL selama 24 jam. Supernatan bebas sel kedelapan isolat BAL tersebut memiliki aktivitas penghambatan terhadap L.monocytogenes dibandingkan kontrol (MRSB), namun tidak menginaktivasi atau menurunkan jumlah L. monocytogenes. L. monocytogenes dapat tumbuh pada MRSB atau kontrol sebesar 1.3 log sedangkan pertumbuhan L. monocytogenes pada kedelapan supernatan BAL tersebut sekitar 0.4-1.0 log. Senyawa antimikroba pada supernatan bebas sel yang tidak dinetralisasi dapat menurunkan jumlah L. monocytogenes berkisar antara 2.0-4.9 log.
Masing-masing isolat BAL (A7, A22, A24, R21, R24, dan R27) membutuhkan waktu yang berbeda untuk mencapai awal fase stasioner. Awal fase stasioner isolat BAL (A7, A22, A24, R21, R24, dan R27) diperkirakan masing-masing pada jam ke-15, ke-10, ke-13, ke-13, ke-17, dan ke-13.
RIWAYAT PENULIS
Penulis dilahirkan di Bojonegoro pada tanggal 22 April 1987 sebagai anak pertama dari pasangan Yadirun dan Tri Handayani. Bangku sekolah penulis dimulai dari TK dan SDN Sukosewu, SLTP Negeri 2 Kapas, SLTP Negeri 2 Bojonegoro, SMA Negeri I Bojonegoro dan kemudian penulis diterima menjadi mahasiswi IPB melalui jalur Undangan Seleksi Masuk IPB (USMI) pada tahun 2005. Pada tahun 2006, penulis diterima menjadi mahasiswi departemen Ilmu dan Teknologi Pangan (ITP).
Selama belajar di Departemen Ilmu dan Teknologi Pangan IPB, penulis mengikuti beberapa kegiatan dan organisasi kemahasiswaan. Diantaranya, penulis pernah menjadi panitia Masa Perkenalan Fakultas (MPF) tahun 2007, panitia Indonesian Food Expo (Ifoodex) tahun 2007, Forum Bina Islami FATETA tahun 2006-2007, dan BINA DESA tahun 2007-2009. Penulis juga pernah mengikuti PKM (Pekan Kreativitas Mahasiswa) yang didanai dengan judul “Pengembangan Telogurt Sinbiotik sebagai Inovasi Pangan Fungsional Indigenous Kaya Antosianin”. Tahun 2008, penulis melakukan praktik lapang di pabrik gula Gondang Baru, Klaten. Bulan Agustus 2009 - Januari 2010, penulis menjadi asisten praktikum mikrobiologi pangan. Saat kuliah penulis menerima beasiswa POM dan Karya Salemba 4 serta Mien R. Uno Foundation.
KATA PENGANTAR
Alhamdulillaahirobbilaalamiin. Puji dan syukur penulis panjatkan kehadirat Allah SWT atas segala karunia, hidayah, dan rahmat-Nya sehingga penulis dapat menyelesaikan penelitian yang berjudul Kajian Senyawa Antimikroba Bakteri Asam Laktat Isolat ASI yang Berpotensi sebagai Probiotik.
Ucapan terima kasih penulis sampaikan kepada sejumlah pihak yang telah berkontribusi secara langsung maupun tidak langsung dalam penyelesaian tugas akhir ini. Perkenankanlah penulis untuk mengucapkan terima kasih kepada:
1. Bapak, Ibu, Adi, pakdhe Pur, bulek Nik, pakdhe Dwi, mas Anip, mas Imam, mas Lubus, dan semua kelurga besar di Bojonegoro yang telah memberikan begitu banyak kasih sayang, dukungan moril maupun materiil sehingga penulis dapat menyelesaikan tugas akhir ini.
2. Dr. Ir. Lilis Nuraida, MSc. dan Siti Nurjanah, STP, MSi. selaku pembimbing akademik yang telah memberikan dukungan, motivasi, bimbingan, dan pengarahan kepada penulis sehingga tugas akhir ini dapat diselesaikan dengan baik.
3. Didah Nur Faridah, STP, MSi. terima kasih atas waktu dan kesediaannya sebagai dosen penguji serta masukan-masukan berarti demi perbaikan skripsi ini.
4. SEAFAST CENTER atas segala bantuan dana dan fasilitas untuk penelitian ini.
5. Dosen-dosen ITP dan TPB serta guru-guru TK, SD, SMP, SMA atas segala ilmu dan pelajaran hidup.
6. Teman satu bimbingan: Fitri dan Siska atas kerjasama, ide, dan bantuannya. 7. Teman-teman ITP khususnya angkatan 42 atas persahabatan dan kebersamaan
dalam mencari ilmu.
8. Sahabat terbaik : Ika, Tyas, Dewi, Galih, Retno, Mike, Reriel, dan Zizie. 9. Mbak Hana, mbak Sofah, mbak April, mas Yeris, mbak Denok, mbak Ria, dan
10.Pak Udin, Abah, Bu Entin, dan Kak Arif atas bantuannya dalam menemani penulis ketika lembur.
11.Serta seluruh pihak yang tidak dapat disebutkan satu persatu. Terima kasih atas segala bantuan yang diberikan kepada penulis sehingga tugas akhir ini dapat diselesaikan dengan baik.
Akhir kata, penulis berharap semoga karya tulis ini bermanfaat dan berkontribusi terhadap perkembangan ilmu pengetahuan dan teknologi, khususnya ilmu dan teknologi pangan. Amiin.
Bogor, Mei 2010
DAFTAR ISI
B. Penggunaan BAL sebagai Pengawet Biologi ... 5
C. Senyawa Antimikroba ... 7
a. Karakteristik Bakteriosin ... 10
b. Klasifikasi Bakteriosin ... 13
c. Faktor-faktor yang Mempengaruhi Produksi Bakteriosin ... 14
D. Bakteri Patogen ... 17
1. L. monocytogenes ... 17
2. S. aureus ... 18
3. B. cereus ... 19
III. METODOLOGI PENELITIAN ... 20
A. Bahan dan Alat ... 20
B. Metode Penelitian... 20
1. Seleksi Awal Isolat Bakteri Asam Laktat (BAL) Potensi Probiotik yang Memiliki Aktivitas Penghambatan Tinggi terhadap Bakteri Listeria monocytogenes ... …22
2. Seleksi BAL yang Berpotensi Menghasilkan Bakteriosin ... 22
a. Pengujian Aktivitas Antimikroba dengan Metode Kontak ... .. 22
b. Penentuan Fase Stasioner ... 23
c. Mempelajari Pengaruh Waktu Inkubasi terhadap Aktivitas Antimikroba ... 24
IV. HASIL DAN PEMBAHASAN ... 26
A. Seleksi Awal Isolat Bakteri Asam Laktat (BAL) Potensi Probiotik yang Memiliki Aktivitas Penghambatan Tinggi terhadap Bakteri Listeria monocytogenes ... .. 26
B. Seleksi BAL yang Berpotensi Menghasilkan Bakteriosin ... 28
1. Pengujian Aktivitas Antimikroba dengan Metode Kontak ... .. 28
2. Penentuan Fase Stasioner ... 32
3. Mempelajari Pengaruh Waktu Inkubasi terhadap Aktivitas Antimikroba ... 37
4. Pengujian Aktivitas Antimikroba Protein Terendapkan (Presipitasi dengan Ammonium sulfat) ... 39
V. KESIMPULAN DAN SARAN ... 43
A. KESIMPULAN ... 43
B. SARAN ... 43
DAFTAR PUSTAKA ... 45
DAFTAR TABEL
Tabel 1. Pengelompokan bakteriosin beserta contoh bakteri penghasil,
DAFTAR GAMBAR
Gambar 1. Struktur Asam Laktat (C3H6O6) ... 8
Gambar 2. Diagram Alir Penelitian ... 21
Gambar 3. Aktivitas Antimikroba BAL Potensi Probiotik terhadap L. monocytogenes ... 26
Gambar 4. Perubahan Jumlah L. monocytogenes pada Supernatan Bebas Sel yang Dinetralisasi dan tidak Dinetralisasi ... 29
Gambar 5. Hubungan pH dan Perubahan Jumlah L. monocytogenes pada Supernatan Bebas Sel yang tidak Dinetralisasi dengan Waktu inkubasi BAL selama 24 jam ... 31
Gambar 6. Grafik pertumbuhan isolat BAL A7 ... 34
Gambar 7. Grafik pertumbuhan isolat BAL A22 ... 34
Gambar 8. Grafik pertumbuhan isolat BAL A24 ... 35
Gambar 9. Grafik pertumbuhan isolat BAL R21 ... 35
Gambar 10. Grafik pertumbuhan isolat BAL R24 ... 36
Gambar 11. Grafik pertumbuhan isolat BAL R27 ... 36
DAFTAR LAMPIRAN
Lampiran 1. Jalur Fermentasi BAL Homofermentatif ... 51 Lampiran 2. Jalur Fermentasi BAL Heterofermentatif ... 52 Lampiran 3. Data Lengkap Aktivitas Penghambatan BAL terhadap
L. monocytogenes dengan Metode Sumur Agar ... 53 Lampiran 4. Data Metode Kontak Perubahan Jumlah L. monocytogenes
pada Supernatan Bebas Sel yang Dinetralisasi
dan tidak Dinetralisasi serta MRSB (Kontrol) ... 58 Lampiran 5. Uji Statistik Pertumbuhan L. monocytogenes pada Berbagai
Supernatan BAL Bebas Sel yang Dinetralisasi ... 62 Lampiran 6. Hubungan antara Nilai pH dan Perubahan Jumlah
L. monocytogenes pada Supernatan BAL Bebas Sel
yang tidak Dinetralisasi ... 63 Lampiran 7. Hasil Uji Statistik Hubungan pH dan Perubahan Jumlah
L. monocytogenes pada Supernatan Bebas Sel yang tidak
Dinetralisasi... 64 Lampiran 8. Data Pengukuran Absorbansi dan Optical Density (OD)
Terhitung ... 65 Lampiran 9. Hasil Pengujian Aktivitas Antimikroba Beberapa Supernatan Bebas Sel yang Dinetralisasi (isolat A22, A24, R22, dan R27) terhadap L. monocytogenes, B. cereus, dan S. aureus ... 67 Lampiran 10. Uji Statistik Perubahan Jumlah L. monocytogenes pada
Supernatan Bebas Sel yang Dinetralisasi dengan Waktu Inkubasi BAL selama 24 jam dan sampai
Awal Fase Stasioner ... 68 Lampiran 11. Kegiatan Presipitasi Protein... 69 Lampiran 12. Hasil Pengujian Protein Terendapkan Isolat R21
(Inkubasi Sampai Awal Fase Stasioner) terhadap
I. PENDAHULUAN
A. LATAR BELAKANG
Kesadaran masyarakat akan hubungan diet dan kesehatan semakin meningkat. Makanan yang mengandung bahan-bahan alami dianggap lebih menyehatkan daripada makanan yang mengandung bahan sintetik. Demikian juga dengan pengawet makanan, pengawet alami lebih disukai daripada pengawet sintetik.
Bakteri asam laktat (BAL) sudah lama digunakan sebagai pengawet biologi atau biopreservatif. Hal ini terlihat dari banyaknya makanan hasil fermentasi oleh BAL seperti yogurt, sauerkraut, pikel, sosis fermentasi, dan lain-lain. Selama fermentasi, BAL menghasilkan metabolit antimikroba sehingga dapat meningkatkan keamanan dan memperpanjang umur simpan makanan hasil fermentasi. Penggunaan BAL sebagai pengawet biologi tidak hanya menggunakan bakterinya tetapi dapat juga menggunakan metabolit antimikroba yang dihasilkan atau keduanya (bakteri dan metabolit antimikroba).
Bakteri asam laktat dapat menghasilkan beberapa metabolit antimikroba seperti asam organik (asam laktat, asam asetat, dan asam propionat), hidrogen peroksida (H2O2), bakteriosin, diasetil, dan lain-lain.
Senyawa-senyawa antimikroba tersebut sangat penting dalam melawan mikroba patogen di dalam makanan maupun di dalam tubuh. Salah satu sifat penting dari BAL adalah kemampuannya dalam menghasilkan bakteriosin.
Bakteriosin adalah senyawa protein yang bersifat bakterisidal atau bakteriostatik terhadap bakteri sensitif. Ketertarikan terhadap senyawa bakteriosin meningkat seiring dengan potensinya sebagai pengganti pengawet sintetik pada makanan. Meningkatnya kesadaran masyarakat akan pentingnya kesehatan serta semakin berkembangnya ilmu pengetahuan dan teknologi mendorong semakin banyak penelitian tentang bakteriosin.
pada konsentrasi rendah sehingga sangat berpotensi sebagai pengawet biologi. Banyak bakteriosin yang memiliki sifat tahan panas sehingga applicable ketika dikombinasikan dengan perlakuan panas (De Vuyst dan Vandamme, 1994). Beberapa bakteriosin memiliki ketahanan terhadap suasana asam maupun basa sehingga memungkinkan untuk dapat diaplikasikan pada berbagai macam makanan.
Melihat besarnya potensi bakteriosin sebagai pengawet makanan yang bersifat alami, eksplorasi terus dilakukan untuk mencari bakteri-bakteri baru penghasil bakteriosin. Bakteri asam laktat isolat ASI yang memiliki potensi sebagai probiotik diketahui dapat menghambat pertumbuhan beberapa bakteri patogen seperti Staphylococcus aureus, Bacillus cereus, Eschericia coli, dan Salmonella typhimurium (Nuraida et al., 2008). Hal ini menandakan bahwa BAL isolat ASI tersebut dapat menghasilkan senyawa antimikroba. Oleh karena itu, sangat penting untuk mengetahui potensi bakteri asam laktat tersebut dalam menghasilkan senyawa bakteriosin.
B. TUJUAN
Tujuan penelitian ini adalah melakukan seleksi bakteri asam laktat isolat ASI yang memiliki aktivitas penghambatan tinggi terhadap bakteri uji L. monocytogenes, mengkaji senyawa antimikroba yang dihasilkan, mengetahui potensinya dalam menghasilkan senyawa bakteriosin, dan mengkaji pengaruh waktu inkubasi terhadap aktivitas antimikroba.
C. MANFAAT
II. TINJAUAN PUSTAKA
A. BAKTERI ASAM LAKTAT
Bakteri Asam Laktat (BAL) umumnya diasosiasikan dengan habitat kaya nutrisi seperti produk susu, daging, beverages, dan sayuran. Akan tetapi, beberapa diantaranya adalah flora normal pada mulut, usus, dan vagina mamalia. Istilah BAL juga dihubungkan dengan bakteri yang berperan dalam fermentasi makanan dan pakan, serta bakteri yang berhubungan dengan kesehatan permukaan mukosa hewan dan manusia (Axelsson, 2004).
Bakteri asam laktat adalah bakteri gram positif, tidak berspora, mikroaerofilik, produk fermentasi utama dari karbohidratnya adalah asam laktat. BAL terdiri dari dua bentuk, kokus (Lactococcus, Vagococcus, Leuconostoc, Pediococcus, Aerococcus, Tetragenococcus, Streptococcus, Enterococcus) dan batang (Lactobacillus, Carnobacterium, Bifidobacterium). Bakteri asam laktat, khususnya genus Lactococcus, Lactobacillus, Leuconostoc, Pediococcus, dan Streptococcus secara tradisional digunakan sebagai kultur starter untuk fermentasi makanan dan minuman karena berkontribusi terhadap flavor dan aroma serta menghambat kerusakan (De Vuyst dan Vandamme, 1994).
Klasifikasi bakteri asam laktat menjadi beberapa genus didasarkan pada perbedaan morfologi, jenis fermentasi glukosa, perbedaan suhu pertumbuhan, produksi asam laktat, kemampuan untuk tumbuh pada konsentrasi garam tinggi, dan toleransi terhadap asam, alkali, serta garam yang berbeda-beda. Pada pengklasifikasian beberapa genus baru, penambahan karakteristik seperti komposisi asam lemak dan sifat motil juga digunakan sebagai dasar. Klasifikasi terbaru menggolongkan BAL kira-kira ke dalam 20 genus, namun dari sudut pandang teknologi pangan hanya terdapat 12 genus BAL yang utama, yaitu : Aerococcus, Carnobacterium, Enterococcus, Lactobacillus, Lactococcus, Leuconostoc, Pediococcus, Streptococcus, Tetragenococcus, Oenococcus, Weisella dan Vagococcus (Axelsson, 2004).
Embden-Meyerhof-Parnas pathway) menghasilkan sejumlah besar asam laktat pada kondisi standar. Metabolisme ini dikenal sebagai fermentasi homolaktat atau homofermentatif. Pada jalur 6-fosfoglukonat atau fosfoketolase, selain asam laktat dihasilkan juga produk lain seperti etanol, asetat dan CO2 sehingga
metabolisme ini dikenal sebagai fermentasi heterolaktat atau heterofermentatif. Leuconostoc, Oenococci, Weissela dan beberapa Lactobacilli termasuk kelompok bakteri heterofermentatif dan selainnya adalah bakteri homofermentatif (Axelsson, 2004). Menurut De Vuyst dan Vandamme (1994), yang termasuk BAL homofermentatif adalah Lactococci, Pediococci, Streptococci dan beberapa Lactobacilli. Jalur fermentasi BAL homofermentatif dan heterofermentatif dapat dilihat pada Lampiran 1 dan Lampiran 2.
Pada kondisi glukosa berlebih dan oksigen yang terbatas, BAL homofermentatif mengubah 1 mol glukosa menjadi 2 mol piruvat. Keseimbangan redoks intraseluler dipertahankan melalui oksidasi NADH seiring dengan reduksi piruvat menjadi asam laktat. Proses ini menghasilkan 2 mol ATP per glukosa yang dikonsumsi (Todar, 2009). Fermentasi homofermentatif menghasilkan asam laktat > 90 % (Bogaert dan Naidu, 2000).
BAL heterofermentatif menggunakan jalur fosfoketolase (pentosa fosfat). Pada awalnya satu mol glukosa-fosfat di dehidrogenasi menjadi 6-fosfoglukonat lalu di dekarboksilasi menghasilkan 1 mol CO2.
Pentosa-5-fosfat dipecah menjadi 1 mol gliseraldehid Pentosa-5-fosfat (GAP) dan 1 mol asetil fosfat. GAP dimetabolisme lebih jauh menjadi asam laktat seperti pada jalur homofermentatif. Asetil fosfat direduksi menjadi etanol melalui acetyl-CoA dan intermediet asetaldehid. Secara teoritis, produk akhir (CO2, laktat, dan
etanol) diproduksi dari 1 mol glukosa sesuai dengan keseimbangan molar (Todar, 2009). Fermentasi heterolaktat dari glukosa melalui jalur 6-fosfoglukonat menghasilkan 1 mol asam laktat, 1 mol etanol, 1 mol CO2, dan
1 mol ATP/mol glukosa (Axelsson, 2004).
pentosaceus, dan beberapa BAL yang belum diketahui genusnya. Lactobacilli merupakan genus terbesar BAL (Axelsson, 2004). Ciri-ciri genus Lactobacilli antara lain : Gram positif, tidak berspora, fakultatif anaerob, berbentuk batang, biasanya membentuk rantai pendek, membutuhkan media kompleks atau kaya nutrisi, katalase dan sitokrom negatif. Pediococci memiliki ciri-ciri yang sama dengan Lactobacilli (Gram positif, tidak berspora, fakultatif anaerob, katalase negatif, dan membutuhkan media yang kaya nutrisi), namun Pediococci berbentuk bulat, tidak memanjang, dan jarang berupa sel tunggal (Holt et al., 1994).
B. PENGGUNAAN BAL SEBAGAI PENGAWET BIOLOGI
Pengawetan biologi berarti penggunaan BAL, produk metabolitnya, atau keduanya untuk meningkatkan keamanan dan kualitas makanan yang pada umumnya tidak harus difermentasi (Montville dan Winkowski, 1997). BAL memiliki potensi besar untuk digunakan sebagai pengawet biologi. Hal ini dikarenakan BAL bersifat aman untuk dikonsumsi dan dapat mendominasi mikroflora pada makanan selama penyimpanan (Stiles, 1996).
Ada beberapa cara penggunaan mikroorganisme menguntungkan pada makanan. Cara tersebut meliputi penggunaan sel mikroba yang aktif tumbuh, sel mikroba yang tidak tumbuh, dan by-products serta komponen seluler dari mikroorganisme. Contoh penggunaan mikroba yang aktif tumbuh adalah penggunaan bakteri dalam mengubah susu menjadi yogurt. Sel yang tidak tumbuh dari beberapa bakteri digunakan untuk meningkatkan masa simpan susu atau daging yang disimpan di refrigerator (Ray, 2001). BAL merupakan populasi dominan pada daging maupun ikan yang disimpan pada kondisi dingin dan vakum (Stiles, 1996). Banyak by-products seperti asam laktat, asam asetat, beberapa asam amino esensial, dan bakteriosin digunakan pada berbagai makanan (Ray, 2001).
tinggi dan produk fermentasinya dapat membantu meningkatkan kesehatan manusia (Soomro et al., 2002).
BAL berperan penting dalam menghasilkan produk fermentasi berbasis susu (yogurt, kefir, susu asam, keju, dadih, dan sebagainya), daging (sosis fermentasi, urutan, dan sebagainya), buah dan sayuran (tempoyak, sauerkraut, pikel, dan sebagainya), serta minuman. Pada proses fermentasi, terjadi proses pengasaman yang cepat pada bahan mentah oleh BAL yaitu dengan menghasilkan asam organik terutama asam laktat. Produk penting lainnya adalah asam asetat, etanol, senyawa volatil, bakteriosin, eksopolisakarida, dan beberapa enzim. Dengan menghasilkan senyawa-senyawa tersebut, BAL dapat meningkatkan masa simpan dan safety, memperbaiki tekstur dan berkontribusi terhadap sensori produk fermentasi (De Vuyst dan Leroy, 2007).
Beberapa penelitian menunjukkan bahwa bermacam-macam BAL dapat melawan patogen dalam usus dan makanan. Sifat antagonis BAL terjadi melalui : (a) penurunan pH usus halus dengan memproduksi SCFA (Short Chain Fatty Acid) seperti asam laktat, asetat dan propionat; (b) membuat nutrisi spesifik tidak tersedia bagi patogen; (c) menurunkan potensial redoks usus halus; (d) menghasilkan hidrogen peroksida pada kondisi anaerobik; dan (e) menghasilkan senyawa penghambat spesifik seperti bakteriosin (Naidu dan Clemens, 2000).
C. SENYAWA ANTIMIKROBA
Senyawa antimikroba adalah senyawa biologis atau kimia yang dapat menghambat pertumbuhan dan aktivitas mikroba. Senyawa antimikroba dapat bersifat bakterisidal (membunuh bakteri), bakteriostatik (menghambat pertumbuhan bakteri), fungisidal (membunuh kapang), fungistatik (menghambat pertumbuhan kapang), germisidal (menghambat germinasi spora bakteri), dan lain sebagainya (Fardiaz, 1992).
Senyawa antimikroba pada pangan dapat dibedakan atas 3 golongan berdasarkan sumbernya : (1) senyawa antimikroba yang terdapat secara alami di dalam bahan makanan, (2) bahan pengawet yang ditambahkan dengan sengaja ke dalam makanan, dan (3) senyawa antimikroba yang terbentuk oleh mikroba selama proses fermentasi pangan (Fardiaz, 1992).
Mekanisme penghambatan mikroba oleh senyawa antimikroba antara lain disebabkan oleh : (1) perusakan dinding sel mikroba, (2) mengubah permeabilitas membran sitoplasma sehingga menyebabkan terjadinya kebocoran nutrien dari dalam sel, (3) denaturasi protein sel, dan (4) menghambat kerja enzim intraseluler (Pelczar dan Chan, 1986).
Kemampuan suatu senyawa antimikroba dalam menghambat pertumbuhan mikroba dipengaruhi oleh beberapa faktor, antara lain : konsentrasi zat pengawet, waktu penyimpanan, suhu lingkungan, sifat-sifat mikroba (jenis, konsentrasi, umur, dan keadaan mikroba), serta sifat-sifat fisik dan kimia makanan termasuk kadar air, pH, jenis dan jumlah senyawa di dalamnya (Fardiaz, 1992).
Senyawa antimikroba yang dihasilkan oleh BAL menurut Ouwehand dan Vesterlund (2004) antara lain :
1. ASAM ORGANIK
Dalam fermentasi heksosa, fermentasi homofermentatif menghasilkan asam laktat, sedangkan fermentasi heterofermentatif menghasilkan asam laktat, asetat, etanol, dan CO2. Asam lemah seperti
bakteri. Asam
am propionat memiliki aktivitas hambat kuat t (Ouwehand dan Vesterlund, 2004).
ar 1. Struktur asam laktat (C3H6O3)
nurut Naidu dan Clemens (2000), asam lipofil sam asetat dapat berpenetrasi kedalam membr m bentuk tidak terdisosiasi. Pada pH intrasel
akan terdisosiasi menghasilkan ion hidrog u kegiatan metabolisme penting seperti transloka oksidatif. Menurut Doores (1993), penggunaan
antung pada konstanta disosiasi (pKa) atau pH dalam keadaan terdisosiasi.
m laktat adalah asam lemah dengan pKa 3.86 pa yang sama, nilai pKa asam asetat dan asam u 4.73 dan 4.87. Nilai pKa yang lebih besa olekul asam propionat yang terdisosiasi kan jumlah molekul asam asetat dan asam
pada pH yang sama (Bogaert dan Naidu, da pH 4, hanya 11 % asam laktat dalam bentuk t
asam asetat dan asam propionat yang tidak ter 92 %. Campuran asam-asam ini sangat disuka kontribusi terhadap penurunan pH sementara
sam propionat dalam keadaan tidak terdisosiasi bagai antimikroba. Campuran asam asetat da nurunkan pertumbuhan Salmonella enterit
Manfaat asam laktat pada industri pangan antara lain : (1) potensi pengasaman dari asam laktat, (2) pengaturan pH oleh sodium dan potassium laktat, (3) penurunan Aw oleh sodium laktat, (4) sinergisitas dengan antioksidan seperti asam askorbat, dan (5) aktivitas antimikrobial (Bogaert dan Naidu, 2000).
2. HIDROGEN PEROKSIDA (H2O2)
BAL dapat membentuk H2O2 jika terdapat oksigen melalui
oksidasi NADH, oksidasi flavoprotein dan superoksida dismutase. Tidak adanya sumber heme, BAL tidak dapat menghasilkan katalase sehingga terjadi akumulasi H2O2 (Ouwehand dan Vesterlund, 2004).
Efek bakterisidal H2O2 dikarenakan efek oksidasi kuat terhadap sel
bakteri, sel protein kelompok sulfhydryl, dan lipid membran dapat teroksidasi. Selain itu, beberapa H2O2 menghasilkan reaksi penangkapan
O2 (oxygen scavenger) sehingga menciptakan kondisi anaerobik yang
tidak diinginkan oleh organisme tertentu. Penghambatan terhadap bakteri gram positif termasuk BAL bersifat bakteriostatik sedangkan bakteri gram negatif dapat terbunuh dengan cepat (Ouwehand dan Vesterlund, 2004). 3. DIASETIL
Diasetil (2,3-butanedione) diidentifikasi oleh Van Niel dan rekannya sebagai komponen aroma dan flavor pada mentega. Diasetil diproduksi oleh spesies dan strain dari genus Lactobacillus, Leuconostoc, Pediococcus, dan Streptococcus, dan juga organisme yang lain. Pembentukan diasetil ditekan ketika metabolisme heksosa. Diasetil dapat diproduksi berlebih jika terjadi metabolisme sitrat. Sitrat dapat diubah menjadi diasetil melalui piruvat (Ouwehand dan Vesterlund, 2004).
Seratus ppm diasetil bersifat bakterisidal terhadap E. coli dan S. aureus sedangkan terhadap L. monocytogenes bersifat bakteriostatik (Lanciotti et al., 2003). Medium yang mengandung glukosa dan sitrat menghasilkan diasetil dua kali lipat dibandingan medium yang hanya mengandung sitrat (Jyoti et al., 2003).
4. KARBONDIOKSIDA (CO2)
Karbondioksida pada umumnya terbentuk selama fermentasi heksosa oleh BAL heterofermentatif, tetapi banyak jalur metabolisme lain yang menghasilkan CO2 selama fermentasi. CO2 memiliki efek
antimikroba ganda. Pembentukan CO2 menghasilkan lingkungan
anaerobik dan CO2 sendiri memiliki aktivitas antimikroba. Mekanisme
penghambatan oleh CO2 belum diketahui, tetapi diperkirakan karena
terhambatnya dekarboksilasi enzimatik dan akumulasi CO2 pada lipid 2 lapis menyebabkan tidak berfungsinya permeabilitas membran (Ouwehand dan Vesterlund, 2004).
Pada konsentrasi rendah, CO2 dapat menstimulasi pertumbuhan
beberapa organisme sedangkan pada konsentrasi lebih tinggi dapat mencegah pertumbuhannya. Karena sifat antimikrobanya, saat ini CO2 digunakan sebagai komponen utama modified atmosphere packages (MAP). Bakteri gram negatif lebih sensitif terhadap CO2 pada MAP
daripada bakteri gram positif (Ouwehand dan Vesterlund, 2004). 5. BAKTERIOSIN
a. Karakteristik Bakteriosin
Bakteriosin dapat dihasilkan oleh bakteri gram positif dan negatif. Bakteri gram positif menghasilkan bakteriosin yang bersifat : (a) disintesis dalam ribosom, terdiri dari 30-60 asam amino; (b) aktivitas penghambatan dari sempit hingga luas terhadap bakteri gram positif; (c) tahan panas; dan (d) terdapat sistem perlindungan diri yang spesifik pada strain penghasil terhadap bakteriosin yang dihasilkannya (Jack et al., 1995).
Salah satu sifat penting bakteri asam laktat adalah kemampuanya dalam menghasilkan senyawa antimikroba yang dinamakan bakteriosin (Savadogo et al., 2006). Bakteriosin yang dihasilkan oleh BAL didefinisikan sebagai protein aktif atau protein kompleks (agregat protein, protein lipokarbohidrat, glikoprotein dan lain-lain) yang menunjukkan aktivitas bakterisidal terutama terhadap bakteri gram positif dan khususnya terhadap spesies yang berhubungan dekat dengannya. Beberapa bakteriosin yang dihasilkan BAL dapat menghambat mikroba patogen dan pembusuk seperti Bacillus cereus, Clostridium botulinum, Clostridium perfringens, Listeria monocytogenes, Staphhylococcus aureus dan lain-lain (De Vuyst dan Vandamme, 1994).
aktif pada makanan dan aktif pada pH tinggi sangat diinginkan (Martirani et al., 2002).
Bakteriosin memiliki perbedaan dalam hal spektrum aktivitas, bobot molekul, tipe aksinya, genetik dan sifat biokimianya. Bakteriosin dapat dihasilkan secara spontan atau dengan induksi. Faktor genetik bakteriosin kebanyakan terletak pada plasmid, sedikit pengecualian yaitu pada kromosom. Pelepasan bakteriosin membutuhkan ekspresi dan aktivitas protein pelepas bakteriosin serta keberadaan fosfolipase A yang tahan deterjen pada membran luar BAL. Penghambatan oleh bakteriosin bersifat spesifik spesies dan aktivitas letal terjadi melalui penjerapan (adsorpsi) pada reseptor spesifik yang terletak pada permukaan bakteri sensitif, kemudian diikuti dengan perubahan metabolisme, biologi dan morfologi yang mengakibatkan terbunuhnya bakteri sensitif (Naidu dan Clemens, 2000).
Target utama bakteriosin yang dihasilkan BAL adalah membran sitoplasma. Bakteriosin menginisiasi reaksi yang mengubah permeabilitas membran sehingga mengganggu transpor membran atau menghilangkan proton motive force (PMF) sehingga menghambat produksi energi dan biosintesis protein atau asam nukleat (Nissen-Meyer et al., 1992).
b. Klasifikasi Bakteriosin
Penggolongan bakteriosin yang dihasilkan oleh BAL menurut Ouwehand dan Vesterlund (2004) antara lain :
(1) Kelas I (Lantibiotik)
Bakteriosin kelas I dinamakan lantibiotik, peptida kecil (<5 kDa). Lantibiotik mengandung asam amino yang secara alami tidak ada di alam (seperti: lantionin dan β-metil lantionin) dengan penambahan sejumlah asam amino terdehidrasi. Asam amino tidak biasa ini disintesis melalui modifikasi setelah translasi. Contoh lantibiotik adalah nisin A, nisin Z, Lacticin 481, Lactocin S dan lain-lain.
(2) Kelas II
Bakteriosin kelas II juga kecil (<10 kDa), pada umumnya stabil terhadap panas, tidak mengandung lantionin, dan merupakan peptida aktif membran. Kelompok kedua ini dibagi lagi dalam tiga sub kelas. Kelas IIa merupakan kelompok terbesar yang mempunyai peptide listeria-active dengan sekumpulan sekuen amino-terminal. Kelas IIb adalah bakteriosin dengan dua peptida. Kelas IIc adalah bakteriosin teraktifasi-tiol.
(3) Kelas III
Bakteriosin kelas III berukuran besar (<30 kDa) dan tidak tahan panas. Bakteriosin kelas III yang sudah diisolasi hanya berasal dari genus Lactobacillus. Bakteriosin yang dikelompokkan dalam kelas ini tidak terlalu banyak karena sedikitnya pengetahuan tentang jenis ini.
(4) Kelas IV
Bakteriosin kelas IV merupakan kompleks dengan karbohidrat atau lipida. Informasi tentang bakteriosin ini sangat terbatas.
Tabel 1. Pengelompokan bakteriosin beserta contoh bakteri penghasil, nama bakteriosin dan bobot molekulnya
Jenis Bakteriosin dan Strain BAL Bakteriosin Bobot Molekul Lantibiotik (Kelas I)
Lactococcus lactis ssp. lactis ATCC114 Lactococcus lactis ssp. lactis NIZ022186 Lactobacillus sake L45
Lactococcus lactis ssp. lactis CNRZ481 Carnobacterium ssp. U149 Bakteriosin kecil tahan panas (Kelas II)
Lactococcus lactis ssp. lactis LMG2130 Lactococcus lactis ssp. cremoris 9B4 Lactococcus lactis ssp. lactis
Lactobacillus acidophilus 11088
Sumber : Naidu dan Clemens (2000)
c. Faktor-Faktor yang Mempengaruhi Produksi Bakteriosin
Produksi bakteriosin ST712BZ oleh Lactobacillus pentosus dalam medium MRSB dipengaruhi oleh pH media. Media yang memiliki pH 5.5, 6.0, dan 6.5 menghasilkan bakteriosin ST712BZ dalam jumlah besar (12.800 AU/ml) sedangkan media yang memiliki pH 5.0 dan 4.5 menghasilkan bakteriosin lebih sedikit (6.400 AU/ml dan 3.200 AU/ml) (Todorov dan Dicks, 2007). Produksi bakteriosin ST194BZ oleh Lactobacillus plantarum juga maksimum pada media yang memiliki pH 5.5, 6.0, dan 6.5 (Todorov dan Dicks, 2005)a sedangkan bakteriosin yang dihasilkan oleh L. acidophilus mencapai maksimum pada media yang memiliki pH 5.0 (Karthikeyan dan Santhosh, 2009).
Media pertumbuhan juga mempengaruhi produksi bakteriosin oleh Lactobacillus plantarum ST194BZ. Produksi optimum (12.800 AU/ml) terjadi karena penambahan tripton (20 gram/L), kombinasi tripton dan ekstrak daging (1:0.6), atau tripton dan yeast ekstrak (1:0.6). Konsentrasi KH2PO4 sebesar 10, 20, 50 g/L menghasilkan aktivitas ganda (25.600
AU/ml). Penambahan MRS dengan 1 g/L gliserol dapat menghambat produksi bakteriosin sedangkan penambahan vitamin tidak menstimulasi produksi bakteriosin (Todorov dan Dicks, 2005)a. Berbeda dengan bakteriosin ST194BZ; penambahan tripton (20 g/L), D-glukosa (20 g/L), dan sukrosa (20 g/L) pada media MRS tidak meningkatkan produksi bakteriosin ST461BZ dan ST462BZ yang dihasilkan oleh L. rhamnosus. Penambahan K2HPO4 sebanyak 20 g/L atau 50 g/L dapat meningkatkan
produksi bakteriosin menjadi dua kali lipat (25.600 AU/ml) sedangkan penambahan KH2PO4 sebanyak 20 g/L-100 g/L dapat meningkatkan
produksi bakteriosin ST462BZ sebanyak empat kali lipat (51.200 AU/ml) (Todorov dan Dicks, 2005)b. Penambahan vitamin seperti tiamin dan DL-6,8-thioctic acid dapat menghasilkan bakteriosin ST712BZ dua kali lebih banyak dibandingkan dengan kontrol (Todorov dan Dicks, 2007).
dan Dicks, 2005)a. Produksi maksimum bakteriosin yang dihasilkan oleh Lactobacillus acidophilus terjadi pada suhu 40˚C (Karthikeyan dan Santhosh, 2009). Masing-masing bakteri memiliki suhu optimum yang berbeda dalam memproduksi bakteriosin.
Menurut Parente et al. (1997) dan Lejeune et al. (1998), produksi bakteriosin pada BAL berkaitan dengan pertumbuhanya: produksi bakteriosin biasanya terjadi sepanjang fase pertumbuhan dan berhenti pada akhir fase eksponensial (kadang-kadang sebelum akhir fase eksponensial). Aktivitas bakteriosin L. plantarum N014 tidak terdeteksi sampai kultur L. plantarum N014 memasuki fase eksponensial dan aktivitas bakteriosin mencapai maksimum pada awal fase stasioner. Setelah mencapai level tertinggi, aktivitas bakteriosin menurun dengan cepat (Rattanachaikunsopon dan Phumkhachorn, 2006). Berbeda dengan L plantarum N014, bakteriosin yang dihasilkan oleh L.acidophilus memiliki aktivitas maksimum pada akhir fase stasioner (Karthikeyan dan Santhosh, 2009). Produksi senyawa mirip bakteriosin oleh Lactococcus lactis CWBI-B1410 dimulai pada fase eksponensial dan mencapai maksimum selama fase stasioner (Diop et al., 2008). Hal ini memperlihatkan bahwa produksi bakteriosin masing-masing bakteri sangat spesifik.
D. BAKTERI PATOGEN
Bakteri patogen adalah bakteri penyebab penyakit. Bakteriosin yang dihasilkan oleh BAL didefinisikan sebagai protein aktif atau protein kompleks yang menunjukkan aktivitas bakterisidal terutama terhadap bakteri Gram positif dan khususnya terhadap spesies yang berhubungan dekat denganya (De Vuyst dan Vandamme, 1994). Bakteri patogen yang memiliki hubungan dekat dengan BAL karena bersifat Gram positif misalnya Listeria monocytogenes, Bacillus cereus, Staphylococcus aureus, dan lain-lain.
1. Listeria monocytogenes
Listeria monocytogenes adalah bakteri Gram positif, berbentuk batang pendek, berukuran 0.4-0.5 x 0.5-2 μm, tidak membentuk spora, motil dengan flagella peritrichous ketika ditumbuhkan pada suhu 20-25˚C, anaerob fakultatif, kemoorganotrof, fermentasi glukosa menghasilkan L(+)-laktat, katalase positif dan oksidase negatif, serta suhu optimumnya 30-37˚C (Holt et al., 1994). L. monocytogenes tahan terhadap pH rendah dan konsentrasi NaCl tinggi, mikroaerobik, dan psikotrof (Rocourt dan Cossart, 1997).
L. monocytogenes terdapat dimana-mana dan dapat tumbuh pada suhu 0˚C. Pada suhu ini pertumbuhannya sangat lambat (Rocourt et al., 2000). Beberapa studi memperkirakan bahwa 1-10 % manusia adalah pembawa L. monocytogenes pada ususnya. Bakteri L. monocytogenes dapat ditemukan pada mamalia, burung, dan kemungkinan pada ikan serta kerang. Bakteri ini dapat diisolasi dari tanah, pakan ternak, dan sumber lingkungan lainnya (FDA, 2009a).
Listeriosis adalah infeksi serius yang disebabkan oleh L. monocytogenes. L. monocytogenes dapat menyebabkan infeksi yang serius pada wanita hamil, bayi yang baru lahir, anak-anak, dan orang dewasa yang sistem imunnya lemah (Rocourt et al., 2000). Ciri-ciri listeriosis adalah demam, nyeri otot, dan kadang-kadang muntah atau diare. Jika infeksi menyebar pada sistem saraf, muncul gejala seperti sakit kepala, leher kaku, kekacauan, hilang keseimbangan, dan sawan. Infeksi pada wanita hamil dapat menyebabkan keguguran, prematur , atau infeksi pada bayi (CDC, 2009).
Beberapa bakteriosin yang dihasilkan oleh BAL memiliki aktivitas antagonis terhadap L. monocytogenes. Nisin memiliki aktivitas antagonis terhadap L. monocytogenes dan aktivitasnya tergantung pada komposisi kimia makanan yang ditambahkan nisin. Pediocins (dari Pediococcus pentosaceus dan P. acidilactici) dapat menghambat pertumbuhan L. monocytogenes dan bakteriosin dari Lactobacillus bavaricus dapat mempengaruhi pertumbuhan L. monocytogenes pada daging sapi yang disimpan di suhu rendah (Rocourt dan Cossart, 1997).
2. Staphylococcus aureus
Staphylococcus aureus berbentuk bulat; diameternya 0.5-1.5 μm; koloni tunggal, berpasangan, atau bergerombol; Gram positif; tidak motil; tidak membentuk spora; anaerob fakultatif; kemoorganotrof; dan katalase positif. Suhu optimum S. aureus adalah 30-37˚C. Bakteri ini umumnya diasosiasikan dengan kulit dan membran mukosa vertebrata berdarah hangat, tetapi sering diisolasi dari makanan, debu dan air (Holt et al., 1994).
umumnya karena makanan tidak dijaga cukup panas (60˚C atau lebih) atau cukup dingin (7.2˚C atau kurang) (FDA, 2009)b.
Gejala keracunan oleh Staphylococci biasanya cepat dan pada banyak kasus bersifat akut. Hal ini tergantung pada tingkat kerentanan terhadap toksin, jumlah makanan terkontaminasi yang dimakan, jumlah racun pada makanan, dan kondisi kesehatan korban. Gejala umumnya adalah muak, muntah, keram perut, dan lemah atau lesu. Pada kasus yang lebih berat gejalanya adalah sakit kepala, keram otot, dan perubahan sementara pada tekanan darah. Dosis toksin kurang dari 1.0 μg pada makanan yang terkontaminasi akan menimbulkan gejala intoksikasi. Toksin dihasilkan ketika populasi S. aureus melebihi 105cfu/g (FDA, 2009)b.
3. Bacillus cereus
Bacillus cereus adalah bakteri Gram positif dengan flagella peritrichous, berbentuk batang, membentuk spora, endospora berbentuk oval, kemoorganotrof, dan umumnya katalase positif (Holt et al., 1994). B. cereus sudah ditentukan sebagai pembawa racun pada makanan sejak tahun 1955 (Todar, 2008)a.
Terdapat 2 jenis penyakit yang disebabkan oleh B cereus : (1) muntah-muntah (emetic) yang disebabkan oleh peptida tahan panas dan berbobot molekul rendah, dan (2) diare yang disebabkan oleh protein berbobot molekul besar (FDA, 2009)c. Waktu inkubasi untuk tipe emetic adalah 1-6 jam sedangkan waktu inkubasi untuk tipe diare adalah 8-16 jam. Penyebab penyakit tipe emetic adalah toksin emetic (ETE) dan mekanisme aksinya belum diketahui, sedangkan penyebab penyakit tipe diare adalah enterotoksin Nhe atau enterotoksin hemolitik (HBL) yang mekanisme aksinya dengan membentuk lubang dan aktivasi enzim adenylate cyclase (Todar, 2008)a.
III. METODOLOGI PENELITIAN
A. BAHAN DAN ALAT
Bahan-bahan yang digunakan dalam penelitian ini adalah kultur bakteri asam laktat isolat ASI yang berpotensi sebagai probiotik sebanyak 24 isolat (23 isolat bersifat homofermentatif dan 1 isolat bersifat heterofermentatif). BAL homofermentatif yang digunakan adalah Lactobacillus pentosus (A7), Lactobacillus rhamnosus (A15, A23, A24, A29, A31, R21, R23, R24, R26, R34, B10, B16), Lactobacillus acidophillus1(A8, A22), Pediococcus pentosaceus2 (A16), Lactobacillus rhamnosus2 (B11), beberapa isolat yang belum diketahui genusnya (A14, A27, R22, R25, R27, B13), sedangkan BAL heterofermentatif yang digunakan adalah isolat A20 (belum diketahui genusnya). Bahan-bahan lainnya adalah kultur bakteri uji (Listeria monocytogenes, Bacillus cereus, dan Staphylococcus aureus), medium Man-Rogosa-Sharpe Agar (MRSA) (Oxoid), Man-Rogosa-Sharpe-Broth (MRSB) (Oxoid), medium Nutrien Agar (NA) (Oxoid), medium Nutrien Broth (NB) (Oxoid), akuades, NaOH 1 N, alkohol, NaCl, buffer sitrat fosfat, dan ammonium sulfat.
Alat-alat yang digunakan adalah neraca analitik, neraca digital, hot plate, autoklaf, vortex, pH meter, sentrifuse berpendingin (Hermle Z 383 K), syringe, membran filter 0.22 m, inkubator, refrigerator, jangka sorong, spektrofotometer (UV-2450, UV-VIS Spektrofotometer), mikropipet, tip berbagai ukuran dan alat-alat laboratorium berbahan gelas.
B. METODE PENELITIAN
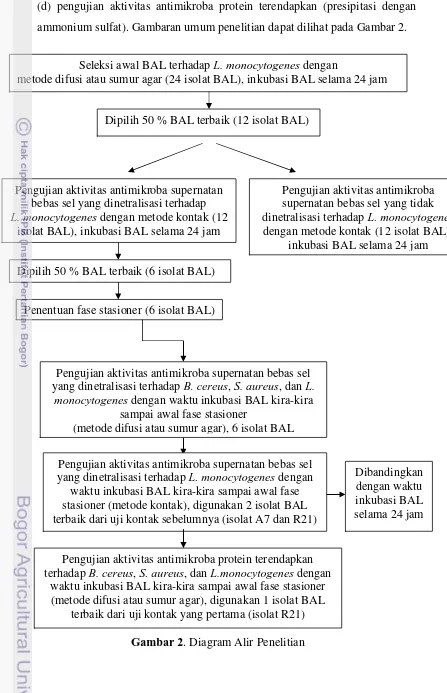
(d) pengujian aktivitas antimikroba protein terendapkan (presipitasi dengan ammonium sulfat). Gambaran umum penelitian dapat dilihat pada Gambar 2.
Gambar 2. Diagram Alir Penelitian Seleksi awal BAL terhadap L. monocytogenes dengan
metode difusi atau sumur agar (24 isolat BAL), inkubasi BAL selama 24 jam
Dipilih 50 % BAL terbaik (12 isolat BAL)
Pengujian aktivitas antimikroba supernatan bebas sel yang dinetralisasi terhadap L.monocytogenes dengan metode kontak (12
isolat BAL), inkubasi BAL selama 24 jam
Pengujian aktivitas antimikroba supernatan bebas sel yang tidak dinetralisasi terhadap L. monocytogenes dengan metode kontak (12 isolat BAL),
inkubasi BAL selama 24 jam Dipilih 50 % BAL terbaik (6 isolat BAL)
Penentuan fase stasioner (6 isolat BAL)
Pengujian aktivitas antimikroba supernatan bebas sel yang dinetralisasi terhadap B. cereus, S. aureus, dan L.
monocytogenes dengan waktu inkubasi BAL kira-kira sampai awal fase stasioner
(metode difusi atau sumur agar), 6 isolat BAL
Pengujian aktivitas antimikroba supernatan bebas sel yang dinetralisasi terhadap L. monocytogenes dengan
waktu inkubasi BAL kira-kira sampai awal fase stasioner (metode kontak), digunakan 2 isolat BAL terbaik dari uji kontak sebelumnya (isolat A7 dan R21)
Pengujian aktivitas antimikroba protein terendapkan terhadap B. cereus, S. aureus, dan L.monocytogenes dengan
waktu inkubasi BAL kira-kira sampai awal fase stasioner (metode difusi atau sumur agar), digunakan 1 isolat BAL
terbaik dari uji kontak yang pertama (isolat R21)
1. Seleksi Awal Isolat Bakteri Asam Laktat (BAL) Potensi Probiotik yang Memiliki Aktivitas Penghambatan Tinggi terhadap Bakteri Listeria
monocytogenes (Garriga et al., 1993).
Kultur bakteri uji L. monocytogenes yang telah disegarkan selama 24 jam diinokulasikan sebanyak 0.2 ml kedalam 100 ml media NA (konsentrasi 0.2 %). Campuran media dan bakteri indikator digoyang-goyang supaya merata. Sebanyak ± 20 ml media NA yang telah terisi kultur uji dituang kedalam cawan dan dibiarkan sampai memadat. Setelah memadat, dibuat sumur-sumur dengan diameter 6 mm. Sebanyak 30 l kultur bakteri asam laktat yang telah disegarkan dalam MRSB selama 24 jam dimasukkan ke dalam sumur yang sudah dibuat, lalu cawan diinkubasi pada suhu 37˚C selama 1 hari.
Zona penghambatan / zona hambat = a - b (mm) Keterangan :
a = diameter zona bening yang terbentuk disekitar sumur (mm) b = diameter sumur (6 mm)
Dipilih 50 % isolat BAL yang memiliki aktivitas penghambatan tinggi terhadap bakteri uji (Listeria monocytogenes). Senyawa antimikroba yang mungkin dihasilkan pada tahap ini adalah asam organik, hidrogen peroksida atau bakteriosin.
2. Seleksi BAL yang Berpotensi Menghasilkan Bakteriosin
a. Pengujian Aktivitas Antimikroba dengan Metode Kontak (Fardiaz, 1989 dan Rahayu, 2000)
Tahap ini bertujuan untuk menguji pertumbuhan L. monocytogenes pada supernatan bebas sel yang dinetralisasi dan tidak dinetralisasi. Penambahan NaOH sampai pH 6.5 bertujuan untuk mengeliminasi penghambatan oleh asam organik.
supernatan bebas sel yang dinetralisasi dan tidak dinetralisasi masing-masing sebanyak 2 ulangan. Dilakukan pemupukan cawan pada jam 0 dan jam ke-8 lalu cawan diinkubasi pada suhu 37˚C selama 2 hari. Inkubasi supernatan bebas sel yang dinetralisasi dan tidak dinetralisasi antara jam 0 dan jam ke-8 dilakukan pada suhu 37˚C. Dilakukan perhitungan jumlah koloni per ml sesuai dengan Standard Plate Count (SPC) dan dihitung jumlah log [Nt/No] untuk setiap perlakuan dan ulangan. Cawan yang dipilih dan dihitung adalah cawan yang mengandung jumlah koloni antara 25-250 (FDA, 2001). Jumlah koloni/ml dihitung dengan rumus : jumlah koloni pada cawan
(n1 + 0.1n2) x d Keterangan :
n1 = jumlah cawan pada pengenceran pertama n2 = jumlah cawan pada pengenceran kedua d = pengenceran pada cawan pertama.
Nilai logaritma = Log Nt No Keterangan:
Nt = jumlah koloni pada waktu t (jam ke-8) No = jumlah koloni pada waktu awal (jam ke-0)
Dipilih isolat yang aktivitas penghambatan terhadap L. monocytogenes pada supernatan yang dinetralisasinya tinggi (50 % terbaik). Nilai logaritma yang rendah dan negatif menandakan penghambatan L. monocytogenes yang tinggi.
b. Penentuan Fase Stasioner (Rattanachaikunsopon dan Phumkhachorn, 2006; Widdel, 2007).
menggunakan MRSB steril jika larutan terlalu pekat. Pengukuran absorbansi dilakukan pada panjang gelombang 660 nm sampai fase stasioner. Kurva pertumbuhan terbentuk dengan memplotkan waktu inkubasi pada sumbu x dengan Optical Density (OD) terhitung pada sumbu y. Nilai OD terhitung diperoleh dengan mengalikan absorbansi (A) dengan faktor pengenceran (FP). Nilai OD terhitung pada sumbu y dibuat dalam skala logaritmik. Fase stasioner ditandai dengan garis yang mulai mendatar pada kurva pertumbuhan. c. Mempelajari Pengaruh Waktu Inkubasi terhadap Aktivitas Antimikroba
(Garriga et al., 1993; Fardiaz, 1989 dan Rahayu, 2000).
Tahap ini bertujuan untuk mengetahui aktivitas antimikroba supernatan bebas sel yang dinetralisasi dengan waktu inkubasi BAL sampai kira-kira awal fase stasioner serta membandingkannya dengan aktivitas antimikroba supernatan bebas sel yang dinetralisasi dengan waktu inkubasi BAL selama 24 jam. BAL yang berbeda menghasilkan bakteriosin pada fase pertumbuhan yang berbeda. Beberapa BAL menghasilkan bakteriosin pada fase logaritmik dan beberapa BAL yang lain menghasilkan bakteriosin pada fase stasioner. Pada pengujian ini, dipilih awal fase stasioner untuk mengetahui aktivitas antimikroba pada fase ini. Metode yang digunakan adalah metode difusi atau sumur agar dan metode kontak.
Pada metode kontak, digunakan 2 isolat BAL terbaik dari uji kontak sebelumnya (isolat A7 dan R21). Bakteri uji yang digunakan adalah L. monocytogenes. Isolat A7 dan R21 diinkubasi sampai kira-kira awal fase stasioner (isolat A7 selama 15 jam dan isolat R21 selama 13 jam). Cara kerja yang dilakukan sama dengan uji kontak sebelumnya.
d. Pengujian Aktivitas Antimikroba Protein Terendapkan (Presipitasi dengan Ammonium Sulfat) (modifikasi Jimenez-Diaz et al., 1995)
A. Seleksi Awal Isol ivitas Penghambatan Tinggi terhadap B
dari tahap ini adalah memilih isolat BAL ambatan tinggi terhadap bakteri uji (Listeria m ivitas antimikroba BAL potensi probiotik menggunakan metode difusi atau sumur agar etode yang umum digunakan dalam mende
metode difusi agar adalah bakteri uji (L. ke dalam agar dan supernatan yang potensi ikroba dimasukkan kedalam sumur agar. Hasi g yang terbentuk disekitar sumur agar setelah ca ehand dan Vesterlund, 2004). Zona bening mena monocytogenes) tidak bisa tumbuh disekitar sum antimikroba.
batan L. monocytogenes oleh BAL potensi pr terlihat dari rata-rata zona penghambatan ata dengan satuan millimeter (mm). Zona hambat di
diameter zona bening dengan diameter sumur monocytogenes oleh BAL dapat dilihat pada G at dilihat pada Lampiran 3.
ktivitas antimikroba BAL potensi probiot
Berdasarkan Gambar 3, isolat BAL potensi probiotik memiliki kemampuan yang berbeda dalam menghambat pertumbuhan L. monocytogenes. Zona hambat menggambarkan aktivitas antimikroba BAL dalam menghambat pertumbuhan bakteri uji (L. monocytogenes). Semakin panjang zona hambat maka aktivitas antimikroba semakin tinggi.
Zona hambat L. monocytogenes oleh BAL potensi probiotik berkisar antara 0.0-9.8 mm. Zona hambat L. monocytogenes oleh isolat A16, A20, A27, A29, dan A31 sebesar 0.0 mm. Isolat A16 dan A31 sebenarnya menunjukkan aktivitas penghambatan (terdapat lingkaran bening tetapi tidak dekat sumur agar), daerah sekitar sumur agar ditumbuhi L. monocytogenes. Isolat A29 menunjukkan aktivitas penghambatan terhadap L. monocytogenes pada salah satu ulangan sedangkan ulangan yang lain tidak menunjukkan aktivitas penghambatan (Lampiran 3). Terdapat 7 isolat BAL yang memiliki zona hambat terhadap L. monocytogenes antara 4.2-6.9 mm, yaitu : isolat A8, A14, A23, R34, B10, B11, dan B13. Dua belas isolat BAL (50 % terbaik) memiliki diameter penghambatan ≥ 7.7 mm, yaitu: isolat A7, A15, A22, A24, R21, R22, R23, R24, R25, R26, R27, dan B16. Aktivitas penghambatan ini dapat disebabkan oleh BAL, asam organik, atau bakteriosin. Dua belas isolat BAL yang memiliki kemampuan menghambat L. monocytogenes terbaik dilanjutkan pada tahap selanjutnya.
Isolat BAL potensi probiotik memiliki aktivitas penghambatan yang baik terhadap bakteri Gram positif. Isolat BAL potensi probiotik yang memiliki aktivitas penghambatan terbaik terhadap L. monocytogenes juga memiliki aktivitas penghambatan terhadap S. aureus dan B. cereus. Berdasarkan penelitian Hartanti (2007), zona hambat S. aureus oleh isolat BAL (R21, R22, R23, R24, R25, R26, dan R27) berkisar antara 1.9-3.5 mm sedangkan zona hambat B. cereus berkisar antara 4.6-6.0 mm.
positif dan khususnya terhadap spesies yang berhubungan dekat dengannya (De Vuyst dan Vandamme, 1994). Pengujian pada tahap selanjutnya adalah untuk memperoleh isolat BAL yang berpotensi menghasilkan bakteriosin.
B. Seleksi BAL yang Berpotensi Menghasilkan Bakteriosin 1. Pengujian Aktivitas Antimikroba dengan Metode Kontak
Metode kontak adalah metode yang mengevaluasi aktivitas antimikroba berdasarkan perkembangan atau kematian bakteri dengan mengukur jumlah bakteri setelah diberi sejumlah zat antimikroba dan dikontakkan pada waktu tertentu (Fardiaz, 1989). Tujuan dari metode ini adalah untuk mengetahui tingkat perkembangan atau kematian bakteri uji (L. monocytogenes) setelah dikontakkan dengan supernatan bebas sel yang dinetralisasi dan tidak dinetralisasi selama waktu tertentu (8 jam). Perkembangan atau kematian bakteri dinyatakan dalam nilai logaritma. Nilai logaritma dihitung dengan rumus log [Nt/No], dengan Nt adalah jumlah bakteri setelah waktu tertentu dan No adalah jumlah bakteri awal. Nilai logaritma negatif berarti bakteri uji (L. monocytogenes) mengalami kematian setelah dikontakkan dengan supernatan yang diduga mengandung senyawa antimikroba dan sebaliknya.
Dua belas isolat BAL dari tahap sebelumnya (A7, A15, A22, A24, R21, R22, R23, R24, R25, R26, R27, dan B16) diinkubasi selama 24 jam. Bakteri L. monocytogenes dikontakkan dengan supernatan BAL bebas sel yang dinetralisasi dan tidak dinetralisasi. Tujuan penetralan adalah untuk menghilangkan pengaruh asam organik. Jika terdapat penghambatan L. monocytogenes pada supernatan bebas sel yang dinetralisasi maka senyawa yang diduga berperan adalah bakteriosin.
Gambar 4. Perubahan jumlah L. monocytogenes pada supernatan bebas sel yang dinetralisasi dan tidak dinetralisasi.
Berdasarkan Gambar 4, terdapat 8 isolat BAL yang menunjukkan aktivitas penghambatan terbaik terhadap L. monocytogenes pada supernatan bebas sel yang dinetralisasi yaitu : isolat A7, A15, A22, A24, R21, R24, R26 dan R27. Secara statistik, pertumbuhan L. monocytogenes pada kedelapan supernatan BAL tersebut berbeda nyata dengan pertumbuhan L. monocytogenes pada MRSB atau kontrol (Lampiran 5). L. monocytogenes dapat tumbuh pada MRSB (kontrol) sebanyak 1.3 log sedangkan pertumbuhan L. monocytogenes pada kedelapan supernatan isolat BAL tersebut lebih rendah yaitu berkisar antara 0.4-1.0 log. Hal ini menandakan masih terdapat senyawa antimikroba pada supernatan bebas sel yang dinetralisasi kedelapan BAL tersebut. Senyawa antimikroba yang mungkin menghambat L. monocytogenes pada kedelapan supernatan bebas sel tersebut adalah bakteriosin. Bakteriosin yang dihasilkan oleh BAL dapat membunuh atau menghambat bakteri yang berhubungan dekat dengannya yaitu bakteri gram positif seperti L. monocytogenes. Kemungkinan terdapatnya senyawa antimikroba lain seperti CO2, diasetil, dan hidrogen peroksida sangat kecil. Kedelapan BAL yang
menunjukkan aktivitas penghambatan terhadap L. monocytogenes merupakan bakteri homofermentatif sehingga kemungkinan terdapatnya CO2 sangat kecil,
CO2 umumnya dibentuk oleh bakteri heterofermentatif. Kemungkinan
terdapatnya senyawa diasetil juga sangat kecil karena produksi diasetil ditekan pada fermentasi heksosa (Ouwehand dan Vesterlund, 2004). Dari delapan
isolat BAL yang menunjukkan aktivitas penghambatan terhadap L. monocytogenes, enam diantaranya adalah L. rhamnosus yaitu isolat A15, A24, R21, R24, R26 dan R27. L. rhamnosus kemungkinan tidak menghasilkan H2O2 seperti hasil penelitian Felten et al. (1999) yang menyatakan bahwa
delapan belas isolat L. rhamnosus diketahui tidak dapat menghasilkan hidrogen peroksida. Oleh karena itu, senyawa antimikroba yang diduga kuat menghambat L. monocytogenes pada supernatan bebas sel yang dinetralisasi adalah bakteriosin.
Aktivitas antimikroba pada supernatan bebas sel isolat BAL potensi probiotik yang dinetralisasi tidak bersifat bakterisidal terhadap L. monocytogenes. Hal ini kemungkinan disebabkan oleh penggunaan hasil penyegaran BAL secara langsung sehingga senyawa antimikroba yang dihasilkan tidak maksimal. BAL hasil penyegaran seharusnya ditumbuhkan kembali pada media baru agar pertumbuhan BAL dan senyawa antimikroba yang dihasilkan maksimal. Selain itu, setiap BAL memiliki waktu inkubasi optimum serta kondisi pertumbuhan yang berbeda dalam menghasilkan senyawa bakteriosin.
Berdasarkan Gambar 4, aktivitas senyawa antimikroba pada supernatan bebas sel yang tidak dinetralisasi bersifat bakterisidal terhadap L. monocytogenes. Aktivitas bakterisidal terhadap L. monocytogenes disebabkan pada supernatan bebas sel yang tidak dinetralisasi masih terdapat senyawa asam organik. Asam organik yang menghambat adalah asam laktat karena isolat BAL yang diuji bersifat homofermentatif. Fermentasi homofermentatif menghasilkan asam laktat > 90 % (Bogaert dan Naidu, 2000).
Senyawa antimikroba pada supernatan bebas sel yang tidak dinetralisasi memiliki kemampuan bakterisidal berbeda terhadap L. monocytogenes. Penurunan jumlah L. monocytogenes berkisar antara 2.0-4.9 log. Kemampuan bakterisidal yang berbeda ini dapat disebabkan oleh banyak sedikitnya asam laktat yang tidak terdisosiasi atau keberadaan senyawa antimikroba lain seperti bakteriosin.
terdisosiasi. Pada translokasi substrat dan fosforilasi oksidatif.
sedikitnya asam laktat yang tidak terdi eh pH. Hubungan pH dan kematian L. monoc g tidak dinetralisasi dapat dilihat pada Gambar
da Lampiran 6.
ubungan pH dan perubahan jumlah L. monoc as sel yang tidak dinetralisasi dengan waktu
rkan Gambar 5, tidak ada korelasi yang signifi ya kematian L. monocytogenes (lampiran 7). belum tentu dapat membunuh L. monocytogene
R25 yang memiliki nilai pH 4.0 mampu menur nes sebesar 4.3 log sedangkan supernatan isola
nilai pH lebih besar masing-masing yaitu 4.1 umlah L. monocytogenes lebih banyak yaitu 4.5
supernatan isolat R27 dan A7 yang memiliki nilai pH lebih kecil yaitu 4.4. Supernatan isolat R27 dan A7 masing-masing dapat menurunkan jumlah L. monocytogenes sebesar 2.0 dan 2.6 log. Hal ini memperkuat dugaan kemungkinan terdapat senyawa antimikroba selain asam organik pada supernatan A24. Nilai pH rendah dan dapat membunuh L. monocytogenes lebih banyak terdapat pada supernatan R23, R25, dan B16. Hal ini kemungkinan disebabkan oleh banyaknya asam tidak terdisosiasi pada supernatan tersebut dan adanya senyawa antimikroba selain asam organik. Nilai pH lebih tinggi dan kemampuan membunuh L. monocytogenes yang rendah terdapat pada supernatan A7 dan R27. Hal ini kemungkinan disebabkan oleh sedikitnya asam yang tidak terdisosiasi pada supernatan tersebut.
L. monocytogenes merupakan bakteri patogen penyebab listeriosis. Bakteri ini menjadi perhatian karena fatalnya akibat yang ditimbulkan dan kemampuannya untuk tumbuh pada suhu rendah. Menurut FDA (2009)a, L. monocytogenes cukup tahan terhadap pembekuan, pengeringan dan pemanasan. Sifatnya yang demikian menyebabkan industri pangan memberi perhatian khusus terhadap bakteri ini. Senyawa antimikroba yang dihasilkan oleh BAL potensi probiotik diharapkan dapat melawan L. monocytogenes sehingga dapat mengurangi terjadinya listeriosis.
2. Penentuan Fase Stasioner
acidophilus memiliki aktivitas maksimum pada akhir fase stasioner (Karthikeyan dan Santhosh, 2009). Produksi senyawa mirip bakteriosin oleh Lactococcus lactis CWBI-B1410 dimulai pada fase eksponensial dan mencapai maksimum selama fase stasioner (Diop et al., 2008).
Awal fase stasioner dapat ditentukan setelah mengetahui kurva pertumbuhan bakteri. Kurva pertumbuhan bakteri terbentuk dengan memplotkan nilai Optical Density (OD) terhitung dan waktu inkubasi. Nilai Optical Density terhitung diperoleh dengan mengalikan nilai absorbansi (A) dan faktor pengenceran (FP). Nilai Optical Density terhitung pada sumbu y dinyatakan dalam skala logaritmik. Pengukuran absorbansi dilakukan dengan menggunakan spektrofotometer pada panjang gelombang 660 nm. Menurut Sutton (2006), metode spektrofotometri mengukur kekeruhan secara langsung. Kekeruhan berhubungan dengan jumlah bakteri dalam suspensi (Todar, 2008)b. Semakin pekat atau semakin banyak populasi mikroba maka cahaya yang diserap (nilai absorbansi) akan semakin besar.
Kurva pertumbuhan menggambarkan fase-fase pertumbuhan bakteri. Fase-fase pertumbuhan bakteri antara lain fase adaptasi (lag phase), fase logaritmik (logarithmic phase), fase stasioner (stationary phase), dan fase kematian (death phase). Fase adaptasi yaitu fase untuk menyesuaikan dengan substrat dan kondisi lingkungan di sekitarnya. Jumlah sel pada tahap ini mungkin tetap, tetapi kadang-kadang menurun (Fardiaz, 1992). Pada fase eksponensial, bakteri tumbuh dan membelah dengan kecepatan maksimum. Pada fase stasioner, jumlah mikroorganisme hidup akan konstan dan pada fase kematian jumlah mikroorganisme hidup menurun jumlahnya (Prescott et al., 2003).
Gambar 6.
Gambar 7.
ar 6. Kurva pertumbuhan isolat A7
Gambar 8.
Gambar 9.
ar 8. Kurva pertumbuhan isolat A24
Gambar 10.
Gambar 11.
ar 10. Kurva pertumbuhan isolat R24
Berdasarkan kurva diatas, dapat diperkirakan awal fase stasioner dari masing-masing bakteri. Selama fase logaritmik, Optical Density terhitung akan terus naik karena jumlah bakteri terus meningkat. Pada fase stasioner, Optical Density terhitung cenderung konstan atau sedikit mengalami kenaikan karena jumlah bakteri yang hidup juga konstan. Isolat A7, A22, A24, R21, R24 dan R27 diperkirakan memiliki awal fase stasioner masing-masing pada jam ke-15, jam ke-10, jam ke-13, jam ke-13, jam ke-17 dan jam ke-13. Data lengkap pengukuran absorbansi dan nilai Optical Density terhitung dapat dilihat pada Lampiran 8.
3. Mempelajari Pengaruh Waktu Inkubasi terhadap Aktivitas Antimikroba Tahap ini bertujuan untuk mengetahui aktivitas antimikroba supernatan bebas sel yang dinetralisasi dengan waktu inkubasi BAL sampai kira-kira awal fase stasioner serta membandingkannya dengan aktivitas antimikroba supernatan bebas sel yang dinetralisasi dengan waktu inkubasi BAL selama 24 jam. BAL yang berbeda dapat menghasilkan bakteriosin pada fase berbeda dan pada tahap ini dipilih awal fase stasioner untuk mengetahui aktivitas antimikroba supernatan bebas sel yang dinetralisasi pada fase ini.
Metode yang digunakan pada tahap ini adalah metode difusi atau sumur agar dan metode kontak. Supernatan bebas sel isolat BAL (A7, A22, A24, R21, R24 dan R27) yang dinetralisasi diuji aktivitas hambatnya terhadap bakteri gram positif (B. cereus, S. aureus dan L. monocytogenes) dengan metode difusi atau sumur agar. Selain itu, supernatan bebas sel isolat A7 dan R21 yang dinetralisasi juga diuji aktivitas hambatnya terhadap L. monocytogenes dengan metode kontak. Menurut De Vuyst dan Vandamme (1994), bakteriosin yang dihasilkan oleh BAL didefinisikan sebagai protein aktif atau protein kompleks yang menunjukkan aktivitas bakterisidal terutama terhadap bakteri gram positif dan khususnya terhadap spesies yang berhubungan dekat dengannya.
aureus) pada sumur agar. Hal ini terlihat dari tidak adanya zona bening disekitar sumur agar (Lampiran 9). Tidak terlihatnya zona bening kemungkinan disebabkan oleh sedikitnya senyawa antimikroba yang dihasilkan atau sedikitnya dosis yang digunakan (50μl).
Pengujian bakteriosin dengan metode difusi atau sumur agar sering tidak terdeteksi dibandingkan dengan metode lainnya. Hal ini dapat dihindari dengan memperbesar ukuran sumur, menyediakan waktu sebelum inkubasi agar larutan bakteriosin dapat berdifusi kedalam agar, meningkatkan volum sampel, menggunakan agar murni, dan lain-lain (De Vuyst dan Vandamme, 1994).
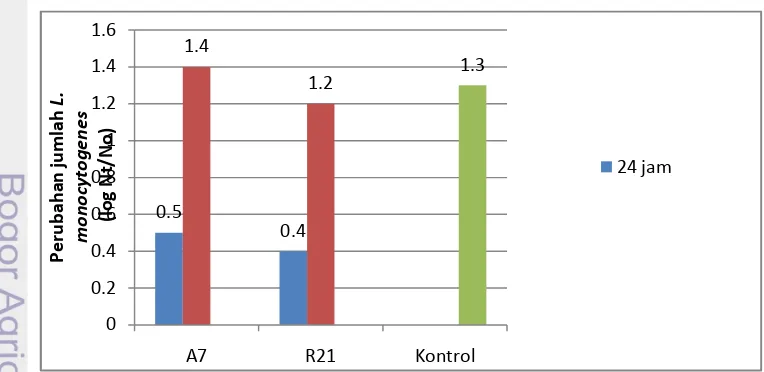
Pada metode kontak sebelumnya, BAL yang diinkubasi selama 24 jam menunjukkan aktivitas penghambatan terhadap L. monocytogenes. Dilakukan uji kontak kembali untuk mengetahui perubahan jumlah L. monocytogenes pada supernatan bebas sel yang dinetralisasi dengan inkubasi BAL sampai awal fase stasioner. Isolat BAL yang digunakan adalah isolat yang menunjukkan aktivitas penghambatan terbaik pada metode kontak sebelumnya yaitu isolat A7 dan R21. Hasil uji penghambatan L. monocytogenes oleh supernatan bebas sel isolat A7 dan R21 yang dinetralisasi dapat dilihat pada Gambar 12.
Gambar 12. Perubahan jumlah L. monocytogenes pada supernatan bebas sel yang dinetralisasi dengan waktu inkubasi BAL selama 24 jam dan sampai awal fase stasioner serta perbandingannya dengan kontrol.