PISIOLOGi
HARA FOSFOR
PADA TANAMAN JAGUNG (Zea
mays
L,)
DALAM KONDISI
CEKAMAN ALUMINIUM
Oleh
SYAFRUDDIN
f
ROGRAM PASCASARJANA
INSTITUT
PERTANIAN BOGOR
FISIOLOGI HARA FOSFOR
PADA
TANAMAN JAGUNG
(Zea
mays L.)
DALAM KONDISI CEKAMAN ALUIWNIUM
SYAFRUDDIN
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Agronomi
PROGRAM PASCASARJANA
INSTITUT P E R T A N W
BOGOR
Judul Tesis : FISIOLOGr HARA FOSFOR
PADA TANAMAN JAGUNG (Zea mays L.) DALAM KONDISI CEKAMAN AL
-N a m a : Syafiuddin
N R P : 99068
hogram Studi : Agronomi
Dr. Ir. Didv Sovaudie. M . .
Ketua Anggota
2. Ketua Program Studi Agronomi
s
Dr. Ir. Ha'gial Aswidinnoor. MSc.ABSTRACT
SYAFRUDDM. Physiological of Phosphorus Nutrient on Maim (Zea mays L.) under Stress Atwninum Condition Under the direction of Didy Sopandie and Trikoesoexnaningtyas.
An alternative to overcome soil problems in acid soil is by growing varieties that tolerant to aluminum toxicity and efficient in nutrient absorbtion and utilization, P
nutrient. Therefore, it is necessary to evaluate genotypes under Al stress condition and efficiency of P nutrient in maize. This study consisted of 3 experheas: 1). Screening of maize genotypes for At tolerance, 2). Screening of maize genotypes for P efficiency and,
3). Physiological of P d e n t under A1 toxicity.
These
experiments were conducted by growing maize genotypesin
culture solution,The results of the experiment showed that Al-tolerant maize genotypes are not always efficient in P nutrient absorbtion and utilization. On the other
hand,
some maize genotypes that are AI-sensitive are efficient in P nutrient. The AMATL-(HS 1)-C243- 1, SATP-I (S2)-C6-SO, BK-(HS)-C2- 1 13-1, BK-(HS)-C2-55-1, and Bisma genotypes is Al-tolerant and efficient and responsive to P under low P level, the CML364 genotype is Al-sensitive which efftcient and responsive to P under low level P.The growth of Al-tolerant genotypes under low concentration of At (2.5 ppm Al) combined with low P (6 ppm P) was stimulated. The concentration of P and A$-P concentration ratio in root were identified as an important mechanism of A1 tolerance in maize. The aluminum tolerant genotype (AMATL-HS-C2-SO) had higher P
concentration in root, and lower At concentration and AI-P absorbtion ratio in both root and shoot than the sensitive genotype (CML364), but was not different in the utilization efficiency and efficiency ratio of P under aluminum stress condition.
SYAFRUDDIN. Fisiologi Hara Fosfor pada Tanaman Jagung (Zea mays L.) dalam kondisi Cekaman Al-um. Dibimbing oleh Didy Sopandie dan Trikoesoemanhgtyas.
Penggunaan kultivar yang tenggang dan efisien hara tenrtama ham P merupakan pilihan sangat penting untuk mengatasi kendala cekaman Al di lahan masam. Penetitian ini bertujuan untuk mengidentifii tingkat ketenggangan genotipe jagung terhadap cekaman aluminium, ef~siensidya terhadap hara P dan fisiologi hara P dalam
dalam kondisi cebman
AL
Peneliiian terdiri dari 3 percobaan: 1) penapisan genotipe jagung untuk ketenggangan aluminium, 2) penapisan genotipe jagunguntuk
efisiensi hara P,dan
3) fisiologi hara P dalam kondisi cekaman aluminium. Ketiga percobaan in dilakdcan dengan menumbuhkan tanamsn jagung di -tan haraHasil Penelitian menunjukkan bahwa jagung genotipe yang tenggang Al belum tentu efkien terhadap P, sebaliiya genotipe yang peka Al belum tentu tidak efisien P. Genotipe AMATL-(HS1)-C243-1, SATP- 1 (S2)-C6-SO, BK-(HS)-C2-113-1, BK-(ITS)- C2-55-1, dan Bisma adalah genotipe tenggang A1 yang efisien dan responsif P pada konsentrasi P rend& Genotipe CML364 clan SATP-2(S2)-C6-SO adalah genotipe peka Al yang juga efisien dan responsif P pada konsentrasi P rendah
.
Genotipe jagung yang tenggang Al jika ditumbuhkan pada larutan hara dengan konsentrasi A1 rendah (2.5 ppm) dan d i k o m b i i i dengan konsenttasi P yang rendah (6 ppm) distimulasi pertumbuhamya temtama W b u h a n akar yang disebabkan oleh meningkatnya kadar P di akar.
Kadax P dan ratio serapan Al-P di akar sangat penting dalam mekanisme ketenggangan tanamrtn jagung terhadap cekarnan Al, karena kadar P akar yang tinggi dan disertai dengan ratio Serapan AI-P di akar yang rendah menyebabkan kadar Al di tajuk rendah pula. Genotipe tenggang (AMATL-HS-C2-SO) mempunyai
kadar
clan serapan P diakztr
retatif lebih tinggi serta kadar Al di akar dan tajuk, ratio serapanAl-P
di akar dan tajuk yang lebih rendah dibanding genotipe peka (CML364). Akan tetapi mempunyai efisiensi penggunaan dan ratio efsiensi P yang tidak k b e d a pada kondisi cekaman aiuminiumDengan ini saya menyatakan bahwa tesis yang bequdul:
FISIOLOGI HARA FOSFOR PADA TANAMAN JAGUNG (Zea mays L.) DALAM KONDISI CEKAMAN AL-
Penulis dilahirkan di Makassar pada tanggal 17 Oktober 1965 dari pasangan ayah Drs. H.Abd. Latief dan ibu Hj. Indo Sakka. Pendidikan sarjana ditempuh di Program Studi Ilmu Tanah Fakultas Pertanian Universitas Hasanuddin Makassar dan lulus tahun 1987. Pada tahun 1999, penulis memperoleh beasiswa dari dana PAATP Badan Litbang Pertanian untuk melanjutkan pendidikan di Program Studi Agronomi pada Program
Pascasarjana ZPB.
Penulis bekerja sebagai Peneliti pada Balai Penelitian Jagung dan Serealia di
D A F T A R
I S 1
Habman
DAFTAR TASEL
...
viiiDAFTAR GAMBAR
...
xDAFTAR LAMPIRAN
...
xi
PENDAHI_rLUAN
...
.
.
...
1Latar Belakang
...
1Tujuan
...
3Hipotesis
...
3TINJAUAN PUSTAKA
...
4Kernasaman dan Aluminium dalam Tanah
...
4Pengaruh Cekarnan Aluminium terhadap Tanaman
...
5Adaptasi Tanaaan terhadap Cekaman Aluminium
...
6Interaksi Fosfor dengan Aluminium
...
10Efisiensi Hara Fosfor
... .
.
...
1 1 BAHAN DANMETODE
...
.
.
...
14Waktu dan Ternpat
...
14Bahan dm Alat
...
14Metode Penelitian
...
.
.
...
14HASIL DAN PEMBAHASAN
...
19Penapisan Genotipe Jagung
unNc
CekamanAl
...
19Penapisan Genotipe jagung untuk Efisiensi hara P
...
30...
Fisiologi Hara P pada Tanaman Jagung dalam Kondisi CekamanAl
35 PEMBAHASAN UMUM...
47...
Halaman
Keterkaitan Aluminium Fospor
dan
Mekanisme KetahananLatar Belakang
S u m h daya labm yang berpotensi untuk p ' di Monesia adalah
sekitar 188.2 juta hektar, kurang lebii 36.37% diantamnya adalah jenis tanah Ultisol
dan Oxisol yang tersebar di Suiawesi, Kalbantao, Sswnatra, Irian, Jawa
dan
Nusa Tengara (Subagyo et al. 2000). Kendala utama pengembagan tanaman pangan khusunya jagung di kedua jenis tanah karena disamping mempunyai kandungan haraN, P, K, Ca, Mg clan Mo yang rendah, juga mempunyai tingkat kemasaman clan kandungan A1 tinggi yang menyebabkan produktivitasnya sangat rendah,
Kandungan A1 yang tinggi menyebabkan tanaman akan keracunan A1 clan f i i P yang tinggi (Sanches dan Salinas 1981; Marschna 1995; Subagyo et al. 2000). Umumnya pada tanah dengan pH (4,s dan kejenuhan A1 >40% sudah berac~n bagi tanaman jagung (Fathan et at. 1988). Aluminium beriIIte~&si langsung dengan ketersedian P bagi tanaman, dimana P &pat difiksasi oleh A1 baik di dalam larutan tanah rnaupun di daiam jaringan sehingga tanaman mengalami defisiensi P
(Adams 1980; Marschner 1995).
Upaya untuk mengatasi kendala kimiia
dalam
tanah, terutarna pengaruh buruk kemasaman d m Al yang tinggi antara lain dapat dilakukan dengan ameliorasimenggunakan kapur dan bakan organik atau dengan pemupukan yang tinggi.
Pendekatan ini memerlukan biaya tinggi clan
sarana
pmduksi tersebut b gtenggang terhadap cekaman tanah masam. Gemtip jagung yang tenggang mampu
memberikan hasil45% lebih tinggi dibanding yang peka apabila ditanarn pada tanah
UItisol (Kasim dan Yasin 1999).
Tanaman yang tenggang terhadap kadar Al tinggi yang &harapkcan adalah tanaman yang memiliki kemampuan menekan kelanrtan
dan
absorbsi Al dalamtanah serta memiliki kemampuan untuk menetrallran pengaruh keracunan A1 setelah disemp ke dalam jaringan tanaman Dari beberapa hasil penelitian temyata tanaman yang tenggang A1 juga lebih efisiensi dalarn menyerap ham. Efisiensi hara pada tanaman yang tenggang Al berbeda antara spesies bahkan antara setiap genotipe dalam satu spesies, Tanaman yang tenggang Al s e r h g diassosiasikan dengan tanaman yang efrsien me-aatkan P dan mampu tumbuh dengan baik pada kondisi kekurangan P di tanah masam (Gupta 1997). Tidak semua tanaman yang tenggang A1 efisiensi memanfaatkan P. Hasii peneiitian Baligar et al. (1997) pada
22 genotipe tanaman jagung yang ditumbuhkan pada media tanah dengan kejenuhan
Al40Y0 menujukkan
bahwa
genotipe yang tenggangA1
efisiensinya terhadap unsurhara bebeda-beda. Dua kemungkinam tanaman tenggang terhadap P rendah adalah:
(1) mampu mengambil hara P leg& banyak pad- kondisi suplai P yang rendah atau pada fiksasi P yang tinggi, atau (2) mampu menmdbahm P yang diserap secara efisien (PoLle d m Konzak 1990). Kemampuan tanaman untuk memanfaatkan P yang rendah pada kelarutan
Ai
yang tinggi dapat rnerupakan salah satu indikator adaptasi tanaman terhadap cekaman A1 (Delhaize et al. 1993).Al
dan fisioiogi efaiensi
hara
Pd a b
kondisi cekamsn Al. Oleh karena itudiperlukan penelitian tingkat ketenggangan jagung terhadap
AI
dan efisiensinya terhadap ham P.Tujuan Tujuan perelitian
ini
ialah :1 . Mcmperoleh ketenggangan genotipe jagung terhadap cekaman aluminium dan efisiensinya terhadap hara P.
2. Memperoleh inforrnasi efisiensi hara P pada tanaman jagung dalam kondisi
ce- ahtminium.
Hipotesis Hipotesis yang diajukan dalam penelitian ini ad* :
1 Terdapat perbedaan tingkat ketenggangan terhadap cekaman aluminium diantara genotipe tanaman jagung.
2. Terdapat perbedaan tingkat efisiensi ham P diantara genotipe tanaman jagung dalam kondisi tanpa cekarnan Aluminium
3. Gemtipe tenggang
AI
lebih efisien P dibanding yang peka dalam kondisiTINJAUAN PUSTAKA
Kemasaman dan Aluminium dalam Tanah
Tingkat kemasarnan (pH) tanah merupakan salah satu tolok ukur sifat kimia
tanah yang sangat mernpengaruhi pertumbuhan dan produktivitas tanaman baik
secara langsung melalui pengaruh konsentrasi ion H' maupun tidak langsung melalui
ketersediaan hara. Perturnbuhan tanaman pada tanah masam (pH ( 5 ) dihambat oleh
pengaruh langsung konsentrasi ion J f , toksisitas Al, Mn atau Fe yang tinggi dan
kekurangan hara
Ca,
Mg, P, dan Mo (Sanches dan Salinas 1981 ; Gupta 1997;Alam et a / . 1999).
Bentuk Al di dalam tanah sangat tergantung pada pH tanah. Pada pH (5 Al mengalami hidrolisis dalam larutan dan membentuk Al+3, dan apabila t e j a d i
kenaikan pH bentuk Af akan berubah menjadi Al(0H)z dan AI(OH)~+. Pada kondisi sekitar netral Al menjadi bentuk yang tidak farut yaitu Al(OH)3, dan pada kondisi alkalin bentuknya akan menjadi AI(0H)i . Kebanyakan kation-kation monomerik ini
terikat pada berbagai ligan organik seperti
PO^'-,
~ 0 4 % , F-, asam-asam organik,protein, dan iemak (Delhaize dan Ryan 1995). Bentuk A13+ merupakan bentuk yang
paling beracun bagi tanaman (Marschner 1995; Delhaize dan Ryan 1995).
Aluminium dalam tanah merupakan salah satu penyebab utarna peningkatan
ion pada tanah sangat masam, ha1 ini karena aiuminium menjadi larut dan terdirpat bentuk kation aluminium hidroksi atau kation Ai3*. Kation ini teradsorbsi
dan berada dalam keseimbangan dengan ion A13+ di dalam tarutan tanah, ion AI3"
(Buckman clan Brady 1982). Reaksi sederhana digambarkan sebagai berikut:
Pengaruh Cekaman Aluminium terhadap Tanaman
Kelarutan Ai yang tinggi berpengaruh langsung terhadap proses-proses fisiologis
dan
metabolis tanaman dan tidak langsung terhadap ketersedian unsur hara sehingga prtumbuhan tanaman tertekan. Menurut Alam et al. (1999), secara umum pengaruhAl pada tanaman yang ditumbuhkan pada t d masam adalah: (1)mengurangi kation betvalensi dua yang diserap oleh akar tanaman (khususnya Ca), (2) menghambat fungsi sel-sel pada jaringan meristem
akar
melalui penetrasi A1 kedalam
protoplasmaakar
dan menghasilkan morfologi akar yang tidak n o d , (3) menunukin adsorpsi anion(sod2-,
~ 0 4 % ~dan
Cl-) oleh akar k a n a meningkatnyasitus adsorpsi positif pada rizosfir
d m
apopiasakar.
Cekaman Al terbadap tanaman pertarm-tama menekan pertumbuhan akar, sehingga akar memerdek, tebal clan rapuh. Terhambatnya pertumbuhan akar tersebut karena pada inti sel, Al berasosiasi dengan DNA dan menghentikan proses pembelahan sel meristem apikal (PoHe clan Konzak 1990; Maschner 1995). Daerah
Sasaran utama cekaman
AI
pada akar addah tudung akaz, dimana rusaknya tudungakar akan mengakibatkan berkurangnya sekresi mucilage (Marschner 1995).
Keracunan AI dapat menghambat pertumbuhan tajuk dengan cara menghambat
pasokan hara, air dan sitokinin dari akar karena buruknya penetrasi akar ke sub-soil atau konduksi hidrolik akar rendah (Marschner 1995; Sopandie 1998). Rusaknya akar oleh A1 menyebabkan terganggunya penyerapan dan transpor hara Ca, K, P, Mg dan N (Polle dan Konzak 1990; Mhmchner 1995), serta peka terhadap kekeringan, yang pada akhirnya mempengaruhi pertumbuhan dan produktivitas tanaman (Gupta
1997). Akumulasi
Al
akan menyebabkan keboooran membran, disintegrasi struktur dan berkurangnya kandungan K daIam jaringan ujung akar, serta menurunkanviabititas protoplasma (Wagaisuma et al. 1995). Terbentuknya ikatan polimer Al
dengan mernbran plasma akar akibat cekaman A1 akan menyebabkan kerusakan pada membran dan kebocoran K dari sef akar (Matsumoto et al. 1992).
Toksisitas
Al
selain mempengaruhi efluks K, juga mempengaruhi efluks Ca.Gangguan
Al
terhadap Ca pada ujung akar menyebabkan defisiensi Ca pada selapikal akar atau mengubah homeostatis Ca. Perubahan ini akan memicu penyimpangan h n g s i metabolisme dalam sel ujung akar yang selanjutnya
rnenghambat pemanjangan akar (Huang et al. 1992).
Adaptasi Tanaman terhadap Cekaman Aluminium
Suatu tanaman yang tenggang terhadap keracunan
Al
mempunyai kriteria antara lain; (1) akar tidak rusak dan sanggup terns berkembang, (2) mampudan sedikit ditranslokasikan ke bagian atas tanam (4) ion A1 tidak dapat menghambat serapan dan translokasi Ca, Mg, dan K serta P (Kochian 1995).
Pengamh cekaman AI3+ tidak sama pada setiap spesies, bahkan pada tanaman
dalam satu spesiespun. Adanya perbedaan tersebut menunjukkan adanya mekanisme
toleransi yang berbeda pada setiap tanaman dalam mengatasi cekaman. Mekanisme
ketenggangan terhadap A1 nienurut Marschner (1 995) antara lain adalah: (1 ) eksklusi aluminium di membran sel, (2) KTK diding sel rendah, (3) alkalisasi di daerah perakaran, (4) proteksi ujung akar oleh mucilage, dan (5) efluks
At.
Mekanisme ketenggangan tanaman secara umum dihipatesiskan oleh Taylor
(1991) terbagi daIam dua kelompok, yaitu: (1) mekanisme penolakan secara eksternal
(external tolerance exclusion mechanism), dimana A1 dicegah masuk ke dalam simplas dan bagian metabolik yang sensitif melalui immobilisasi dinding sel,
permeabilitas selektif dari membran plasma, barier pH di rizosfir, eksudasi ligan
pengkelat (eksudasi asam organik), eksudasi fosfat dan efluks Al, dan (2) mekanisme toleransi secara internal (internal tolerance mechanism), dimana Al mengalami pengkelatan oleh asam organik di dalam sitosol, kompartementasi
AI
di vakuola,protein pengikat aluminium, enzim yang tahan
At,
d m peningkatan aktivitas enzim.Betierapa hipotesis tersebut di atas telah dibuktikan dalam beberapa penelitian antara
lain y ang sangat penting adalah:
1. Kelatisasi A1 oleh Eksudasi Asam Organik Tanaman
Menurut Delhaize dan Ryan (1995) salah satu penyebab terjadinya
yang dieksudasikan sehingga A1 terkelatisasi oleh asam organik baik di dalam jrtringan maupun di media/larutan.
Asarn organik yang dieksudasikan oleh akar tanaman umumnya d a b asam malat, asam sitrat, dan
asam
oksalat. Jumlab dan jenis yang dieksudasikan tergantung dari spesies dan kultivar tanaman. Asam malat m e wasam organik yang paling banyak dieskresikan oleh ujung akar pada tanaman gandum yang tenggang A1 @baize et a/. 1993; Ryan et al. 1995a dan 1995b; Pellet et al. 1996). TanamanCassiatora. L, tritikale, dan jagung yang tenggang A1 jika diinduksi dengan AI akan mengeksudasikan asam sitrat lebih banyak dibanding yang peka (Ma et a[. 1997; Pellet et al. 1995).Tanaman kedefai yang tenggang (Yellow Biloxi) mengeksudasi asarn oksdat d m asam sitrat lebih tinggi dibanding dengan yang sensitif (Lumut) (Sopandie 1999).
2. Kemampuan Menaikkan pH Rizosfir
Peningkatan pH rizosfir pada lanrtan/media merupakan salah satu indikator ketenggangan tanarnan terhadap cekaman Al. Penin- pH rizosfir a h meningkatkan ketersediaan unsur hara seperti P, dam menurunkan k e t e r s e d i Zn, Cu, Fe, Mn dan Al (Haynes 1990). Genotipe t w a n g pada tanaman gandum, barley,
pad< clan jagung yang d i b u h k a n pada Iarutan hara meningkatkan pH larutan d m rnenurunkan kelarutan clan toksisitas Al. Di pihak lain genotipe yang sensitif lebih rendah atau tidak mempunyai pengaruh terhadap pH clan konsentarsi k e h t a n Al
Tanaman Arabidopsis yang diinkubasi dengan 300 phi AlCb selama 12 jam
menunjukkan bahwa genotipe tenggang mampu medaikkan pH hutan dari 4.3- 4.4
rnenjadi 4.53, sedangkan pada genotipe peka kenaikan pH hanya 4.39. Kenaikan pH rizosfir sebesar 0.1 unit pada kondisi cekaman Al menyebbkan kenaikan laju pertumbuhan akar 2 kali lipat pada genotipe tenggang dibanding genotipe peka
(Degenhardt et al. 1998). HBsil penehtian Anwar et al. (1996) pada tanaman kedelai
menunjukkan bahwa genotipe yang tenggang mempunyai kemampuan dua kali lipat
dalam meningkatkan pH rizosfir dibandmg dengan yang peka.
3. Eksudasi Fosfai
Ujung akar tanaman jagung dan gandum di samping &pat mengeksudasi asam oraganik juga mampu mengeksudasikan fos&t organik. Eksudasi P dari akar tanaman adalah mempakan proses alami dari hara P dan bagiaa dari keseimbangan P
dalarn taarnan (Pellet et a1.1996). Eksudasi fosfat organik oleh akar tmiunan
merupakan mekanisme sekunder yang membuat kemampuan tanaman tenggang
terhadap Al. Detoksifikasi
ILU
o b h fosfat karena terbentuknya kornpleks P-Al (DeIhaize er al. 1993; Pellet et a1.1995 clan 1996). Pada kondisi suplai P terbatas, secara absolute eksudasi P b e r m,
tetapi secara relatif meningkat dan rnenjadi komponen utama &lam penyerapan hara P. Dilain pihak, jika tanaman disuplai P dengan baik akan menmgkatkan eksudasi dibandmg dengan tanaman stress PInteraksi Fosfor dengan AIuminium
Taoaman menyerap P dalam bentuk ion HzP04- atau ~ C P O ~ ~ , bentuk ion yang diserap tersebut tergtmtung pH tanah. Pada pH
asam
bentuk yang dominan diserap adalah H2PO4-. Pada pH sekitar netral (pH 6-7) kedua bentuk tersebut dapat diserap, sedangkan pada alkalis bentuk yang dominan adalah HPOI-~ (Buckman dan Brady1982). Pada tanah sangat masam & H e ) keianitan A13* sangat tinggi Hal in menyekbkan P tidak larut dan b a n g tersedia bagi tanam (hanya sebagian kecil H2P04' yang tersedia bagi tanaman). Aluminium tidak hanya menghambat ketersediaan P, tetapi juga mengambat transpor dan penggunaan P (Buckman dan
Brady 1989; Rao et al. 1999). Ion A1 bermuatan positif &pat berassosiasi dengan
gugus fosfat dari ATP atau fosfolipid pada membrane sehingga mempengaruhi efektifitas transport proton Hal ini akan mengakibatkan penyerapan hara yang dikatalis oleh pompa proton menurun (h4atsumoto 1988). Aluminium secara langsung berinteraksi dan memiliki kapasitas fiksasi yang tinggi terhadap unsur hara
P, baik di dalam larutan tanah maupun jaringan tanaman membentuk kompleks Al-P
yang tidak larut sehingga tidak tersedia bagi tanarnan (h4arschner 1995). Reaksi sederhana pengendapan P oleh
AI
sebagai berikut:~ 1 ~ '
+
H2P0L(larut)
+
2H20 2H2+
A1(0H)zHzP04 (Tidak larut)Keracunan aluminium kerapkali diassosiasikan dengan gejala defisiensi P,
sebdiknya P efektif sebagai agen detoksifikasi Al. Interaksi Al-P di dalam tanarnan
antara
Iain
nelapsis pada ujung daun, penghambatan pertumbuhan, daun berwarna hijau tua, dan kadang-kadang daun atau batang berwarna ungu. Defisiensi P ;disebabkan menurunnya penyerapan dan transpor P oleh pengendapan Al-P pada a k a tanaman, clan gangguan A1 dalam metabolisme P yang sudah ada pada pucuk (Foy 1996). Tanaman yang peka A1 di akar akan bergabung dengan P membentuk suatu kompleks sehingga menurunkan akumulasi P di dam (Sivaguru
dan
Paliwalr
993).Efisiensi Hara Fosfor
Efisiensi hara suatu tanaman adalah kemampuan tanaman tumbuh dan menghasilkan biomaslhasil ekonomik serta menyerap hara secara optimum baii
pada kondisi yang optimum maupun kondisi cekaman atau defisiensi hara d i dalam
tanah. Efmiensi hara setiap tanaman dapat ditingkatkan dengan cam: (1) meningkatkan kontak akar dengan tanah, terutama unsur ham yang tidak mobil
seperti P, (2) memperbesar penyerapan per unit akar, dengan memperbaiki mekanisme penyerapan, atau (3) meningkatkan kemampuan tanaman menggunakan
hara yang h a n g larut (Caradusl990).
Efisiensi hara P dapat dibedakan dalam : (I) efisiensi penyerapan P
.
yaitujumlah hara P yang diserap di &lam tanaman per unit hara P yang ditambzthkan, (2) efisiensi penggunaan P, yaitu hasil bijilbiomas yang dihasilkan per unit hara P dalam tanaman dan (3) efisiensi rasio P, yaitu perbandingan antara biomasbsil tanaman
unit P yang diperoleh, (b) tipe dan
masa
tumbuh tanaman, (c) pcmbagian P di antaraorgsn-organ tanaman, (d) translokasi
dan pembagian
P di dalam tanamm, (e) redistribusi sebelum asimilasi P, ( f ) laju kematian daun, dan (g) pembagian proporsi yang lebih besar pa& biomassa dengan hasil y m g dapat dipanen (&XI etat.
1999).Secara umum mekanisme efisiensi P pada tanaman dalam meningkatkan
ketersedhn P
dan
penyerapannnya pada kondisi kekumngan P adalah (1) eksudasibahan
kimia
ke dalam tizosfir, (2)perubahan
pada geometri atau bentuk system perakaran, dan (3) berassosiasi dengan mikroorganisme (Rengel 2000). Lebii jauh dijelaskan oleh Rao er al. (1999) b a h w adaptasi tanaman terbadap pasokan P yang a d a h berupa: (1) mekanisme tanaman yang meningJcatkan akuisisi P yang terdiri dari karakteristik morfologi akar m e b a r a n , pertumbuhan clan diameter akar, perkembangan akar rambutdan
asosiasinya dengan m i k o h ) dan karakteristikfisiologi (sistern penyerapan P dan mobiiisasi P di rizosfir), (2) mekanisme yang
meningkatkan penggunaan P terdiri dari pembagian P dalam tanaman (remobilisasi P dalam tanaman clan status P pa& organ yang dipanen) dan efisiensi penggunaan P
pada tingkat sehder (kompartementasi P pada intraseluler
dan
penggunaanmetabolisme P).
Kamkteristik akar yang berper~nan pent* ddam mekanisme efisiensi P
adalah akar rambut clan panjmg akar (Rengel2000). Akar rambut berkorelasi dengan tingkat efisiensi hara, makin banyak
aka.
rambut dan percabangan akar pada(Raghothama 1999). Genotipe gandum yang efisien P mempunyai akar rambut lebih
banyak dan akar lebih banjang di-ing yang tidak efisien P mengel 2000).
Sistem penyerapan P pada tanaaan jagung oleh plasma membran pada sel tanaman adatah melaIui kotransport ZH+/HZPO~' (Rao et al. 2000). Penyerapan P
dapat ditingkatkan jika panjang akar per unit berat tanaman dan maksimum influks per sentimeter panjang akar (I,*) meningkat serta konstanta Michaelis-Menten (K,)
dan konsentrasi minimum (C,i,) menurun.
Peningkatan eksudasi asam organik di rizosfu oleh tanaman yang d i i b u l k m pada tanah masam akan meningkatkan ketersediaan P dari kompleks Al-P dan
Fe-P
(Robert et al. 2000dan
Marcshner 1995). Secara umum tanamandikotil khususnya tanaman legume lebih efisien dibanding monokotil ddarn
memproduksi d m mengeksudasi asam organik ke rizosfir untuk rneningkatkan kelarutan P pada kondisi P defisien (Raghothama 1999). Genotipe bean dan barley
yang efisien P mempunyai aktXtas pospatase ekstraseluler di rizosfk yang lebih
besar dibanding dengan yang tidak efisien (Rengel2000).
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dikkamakan pada bulan Juli sampai November 2001 di
Rumah
Kaca dan Laboratorium Balai Penelitian Bioteknologi (Balitbio) clan di Pusat
Penelitian Tanah dan Agroklimat (PusItittanak) Bogor.
Bahan dan Alat
Bahan tanaman yang dig& adalah 22 genotipe jagung bersari bebas (Tabel Lampiran 1). Hara kultur air yang digunakan sesuai dengan Magnavaca 1982 ( d i i u dalam Urrea-Gomes et a1.1996) yang dimodifkasi dengan mengganti Fe<N03)~-4HzO menjadi Fe-EDTA seperti pada Lampiran 2. Sebagai sumber A1 digumkan AlC13 6 H20, dan untuk mengatur pH larutan digunakan NaOH IN dan HCL 1N. Bahan untuk d i s i s kandungan A1 dan P pada jaringan, antara lain adalah diethhyl ether, M-403, HC104, HzS04, KH2P04, CH3COO-Na anhidrat, NazSO3, cadmiun, H3B03. W O ,
dan
aquades.Metode Penelitian
Penelitian terdm atas tiga percobaq yaitu: (1) penapisan genotipe jagung untuk ketenggangan aluminium, (2) penapisan genotipe jagung untuk efisietlsi hara P,
dan
(3) efisiensi hara P dalam kondisi cekaman aluminiuraPercobann I: Penapisan Genoh'pe Jagung rurtuk Kdenggungan Al
Percobaan ini dimaksudkan untuk mengetahui tingkat ketahamm genotipe
jagung terbadap cekaman Al. Rancangan yang digunakan adalah Rancangan Acak Kelompok yang disusun secara petak terpisah. Sebagai petak utama adalah 6 t d
cekaman
Al, yaitu : 0 ppm (tanpa Al), 2,5 ppm, 5 ppm, 10 ppm, 20 ppm, dan 40 ppmA1
clan sebagai anak petakadalah
25 genotipe jagung. Setiap unit percobaan diulang 3 kali.Sebanyak 1 kecambah jagung tiap genotipe yang berumur 6 hari ditumbubkan selama 14 hari pada bak-bak berisi larutan hara 1.5 ZEtanaman. Setiap 2 hari larutan hara diukur vohune dan pH. Volume larutan dipertahadcan sesuai dengan
volume awal dengan penamba.hau aquades, sedan- pH dipertahankan pada skala
4.0
+
0.1 dengan menggunakan larutan NaOH 1 N atau HCl 1 N.Peubah yang d i d i s i s adalah panjang
akar,
indeks ketenggangan berupapanjang akar relatif (PAR), pertambaban panjang akar (PPAR), bobot akar relatif @AR), bobot pucuk relatif (BPR) menurut Baligar
et
al. (1989) sebagai berikut:IT=
(Pertumbuhan dengun ALp,rtw&&an trurpcr A t ) x 100%Apabila terdapat i n t d i antam genotipe dengan konsentrasi Al, maka
y a k (I) tenggapg - mempunyai nilai Xi
>
x
+
SE t(tabe1) , (2) moderat-tenggang-
nilai ant-
k
-
SE t(tabe1) dengan X+
SE t(tabe1) , dan (3) peka dengan nilaiXi 5
%
-
SE t(tabe1).PembaPn
AT:
Penapkm Genotijx JagwtguniuR
E f f i WaraP
Percobaan ini dimaksudkan untuk mendapatkan genotipe jagun% yang efisien
terhdap hrva P tanpa cekaman aluminium. Percobaan menggunakan Rancangan Acak Kebmpok yang disusun secara petak t e r p i . Sebagai petak utama d a h h 6 tarafpemberian P, yaitu :O (tanpa P), 1.4 ppm, 5 ppm 10 ppm, 15 ppm, dan 20 ppm P dan sebagai anak petak adalah 22 genotipe j a m , genotipe yang sama pada percobaan I (Tabel Lampiran 1). Setiap unit percobaan diulang 3 kali.
Satu kecambah jagung tiap genotipe yang berumur 6
hari
ditumbuhkan selarna 14 hari pada bak-bak berisi larutan hara 1.5 //tanaman. Setiap 2 hari larutan hara diulcur volume dan pHnya. Volume larutan dipertahankan sesuai dengan volume awal dengan penambahan aquades sedangkan pH dipertahankan pada skala 6.0 $: 0.1dengan menggunakan hutan NaOH 1 N atau HCl1 N.
Peubah yang dianaLisis adalah-.
1. Bo bot kering biomas dengan menggunakan uji regresi
2. Efisiensi hara antara setizrp genotipe dinilai berdasarkan:
a. Efisiensi agronornik (EAGR), yaitu bobot kering maksimal yang
dihasillcan per hara P maksimal, yang diperoleh dengan mengkorelasikan
b. Efisiensi serapan P (ESP) , yaitu jumlah hara P yang diserap di dalam tanaman per unit hara P yang ditambabkan
ESP = m g P jaringanlmg P yang ditambahkan
C. Efwiensi penggunaan P (EPP), yaitu hasil biomas yang dihstsilkan
persatuan
bara
P dalarn tanamanEPP = mg bobot kenhg tanamad konsentrasi P jaringan (mg/mg)
d. Rasio efisiensi P (REP)
,
yaitu perbandingan antara biomadhasil tanaman dan unit unsm ham P tanaman.REP = m g bobot kering tanamadmg P yang d i p
Penggolongan genotipe efisien dilakukan untuk EGAR ESP, EPP, dan REP
dengan menggunakan kriteria Xi 1
2
+
SE x t(tabe1)P ~ ~ O ~ U M
HI:
Efsiemi HaraP
dalam Kondisi C C ~ M I M AiPercobaan ini dimaksudkan untuk mempelajari efisiensi hara P tanaman
jagung dalam kondisi cekaman aluminium antara genotipe yang tenggang, moderat dan peka. Percobaan menggunakan Rancangan Acak KeIompok d a k n bentuk faktorial yang terdiri
dari
2 faktor. Faktor 1 adalah 2 genotipe jagung hasil percobaan tahap I danII,
yaitu gewtipe taggang WTL-HS-C2-SO dan peka Al (CML364) kedua genotype ini efisien dalam menyerap hara P tanpa cekaman Al. Faktor I1 terdiri dari 6 kombinasi A1-
P, yaitu : 1) tanpa Al + Pkurang
(6 pprn P), 2)Al rendah (2.5ppm
Al)
+
P kurang (6 ppm P), 3)Al tinggi (7 pp*Al)
+
P kurang (6 pprn P), 4)f 11.5 ppm P), dan 6)
Al
tinggi (7 ppm Al)+
P cukup (1 1.5 ppm P). Konsentrasi A1 dan P yang digunakan berdasarkan basil percobaan tahap I dan 11. Setiap unit perwbaan diulang 3 kali.Sebanyak 2 kecambah jagung yang telah lxxumur 6 hari ditumbuhkan pada pot-pot beski lamtan ham 3 [/pot selama 14 hari Di awal percobaan pH larutan hara diseragamkan untuk seluruh perlakuan pada skala 4.0. Dilakukan penyesuaian
votume dan pH larutan seperti pada p e r c o b I dan
II.
Setiap unit percobaan diulang 4 kali.Peubah yang dianalisis adalah panjang akar, PAR, PPAR, laju tumbuh akar,
bobot kering akar, bobot kering tajuk, ratio bobot kering akar-tajuk, kadar P akar clan
tajuk, ratio kadar P akar-tajuk, serapan P akar dan tajuk, EPP, ESP, REP, kadar Ai
HAS&
DAN
PEMBAWASAN
Penapisan Genotipe Jagnng
untuk
CekamanA1
Tingkot Kefenggungnn AI antar GenoZipe JogungFaktor konsentrasi A1 dan genotipe berpengaruh nyata terhadap panjang akar, sedangkan interaksi antara konselatasi A1 dengan genotipe tidak berpengaruh nyata (Lampiran
2).
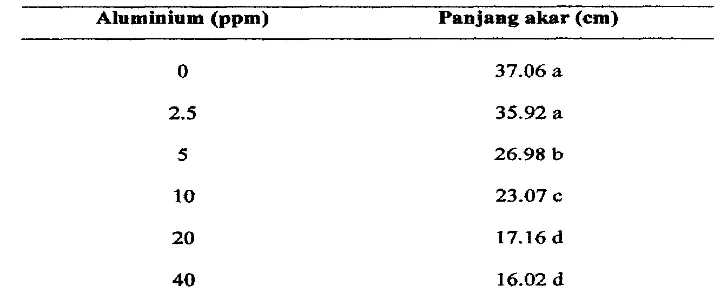
Pemberian A1 sampai pada taraf 2.5 ppm tidak beqengaruh nyata terhadap panjang aka, pengaruh nyata terlihat pada 5 ppm atau lebih yang menyebabkan parijang akar&in
pendek
(Tabel 1). Panjang akar setiap genotipe bervariasi antara 18.72-
31.39 cm. Akar terpanjang ditunjukkan pada genotipe BK- (HS)-(22-129-1, BK-@IS)-C2-113-1, SATP-l(S2)-C6-S0,dan
Bisma (Tabel 2).Tabel I
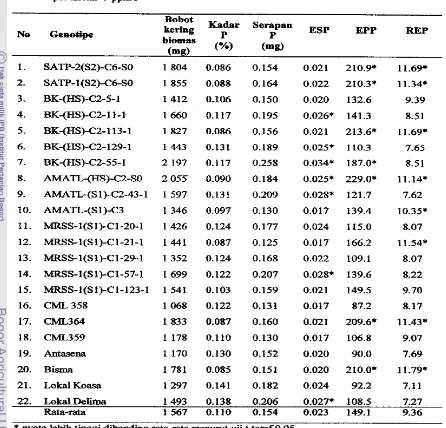
.
Rata- raia Panjang Akar Tanrtman Jagung set+ Konsentrasi A1 Aluminium (ppm) Panjang akar (em) [image:38.543.96.454.311.463.2]Tabel 2. Rata-rata pa4ang Aka Tanaman Jagung Setiap Genotipe
No Genotipe Panjang akar (cm)
1. SATP-2(SZ)-C6-S0 24.84 g-j
2. SATP-1 (S2)-C6-SO 29.14 abc
3. BK-(HS)-C2-5-1 25.33 f-i
4. BK-@IS)-C2- 1 1
-
1 5. BK-fHS)-C2-113-16. BK-(HS)-C2-129- 1
7. BK-@IS)-C2-55-1
8. AMATL-@is)-C2-SO
9. AMATL-(S 1)-C243-1
10. AMATL-(S1)-C3
11. MRSS-I(S1)-C1-20-1
12. MRSS-l(S1)-Cl-21-1
13. MRSS-l(S1)-Cl-29-1
14. MRSS-I(S I)-C1-57-1
15. MRSS-l(S1)-C1-123-1
16. CML 358
17. CML364
18. CML359
19. Antasena
20. Bisrna
2 1. Lokal Koasa
22. Lokal Delima
28.78 bcd
30.55 ab
31.38 a
28.44 b-e
26.19 e-i
22.56 ij
27.75 cde
27.08 C-f
26.78 d-h
24.72 hij 25.31 f-i 26.81 d-h 18.72k 20.67 jk 24.50 hij 25.25 f-i 29.14 abc
23.94 ij 24.86 g-j
FaMor konsentrasi Al, genotipe rnaupun interaksi antara konsentrasi A1 dengan
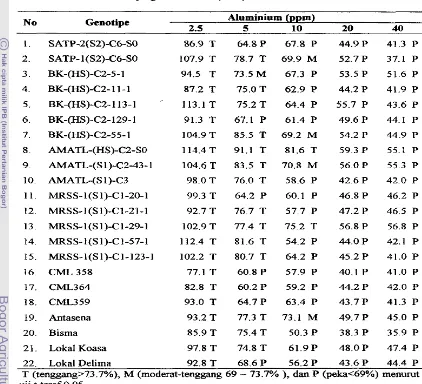
genotipe menunjukkan pengaruh nyata-terhadap panjang akar relatif (PAR) seperti ditunjukkan pada Lampiran 3. Adany8 interaksi tersebut menunjukkan ba.hwa tingkat ketenggangan
AI
antar genotipe berdasarkan nilai PAR berbeda pada setiap kollsentrasi Tingkat ketenggangan Al berdasarkan nilai PAR &pat dikelompokkansebagai
berikut:
tenggang (PAR >73.7%), moderat-tenggang (PAR 69 - 73.7 %), dan peka (169%)). PengeIompokan ini berbeda dengan yangumum
digunakan dimam PAR >50% dianggap tenggang baik pada tanaman jagung @is 1997 dan Baligar et at. 1997) maupun pada tanaman lainnnya seperti kedelai (Soepandi ef al. 2000)
dan
padi (Jagau 2000).Secara umum &pat ditunjukkan bahwa makin tinggi konsentrasi Al, tingkat
ketenggangan tanaman bedasarkan nilai PAR makin peka. Pada pemberian 2.5 pprn Al beberapa genotipe mempunyai nilai PAR yang cederung lebih tinggi dibanding tanpa
A1 (Tabel 3). Pada konsentrasi 5 pprn &n 10 ppm A1 ketenggangan setiap genotipe
berbeda-beda, sedangkan pada konsentrasi 20 pprn clan 40 pprn
A1
semua genotipetergolong
peka
Genotipe AMATL-(HS)-C2-SO clan MRSS- 1 (S 1)-C 1-29- 1 tengganghingga 10 ppm AL Genotipe SAW-1 (S2)-C6-SO, BK-(HS)-C2-55-1, AMATL-(SI)-C2- 43-1, dan Antasena tenggang pada 5 ppm A1 tetapi moderat tenggang pada 10 ppm
Al.
Genotipe BK-@IS)-CZ-1 1-1, BK-(HS)-C2-113-1, AMATL-(S1)-C3, MRSS-l(S 1)-Cl-
21 -1, MRSS-l(SI)-Cl-57-1 tenggang -a 5 ppm A1 dan genotipe BK-@IS)-C2-5- 1 moderat tenggang hingga pada 5 pprn
Al.
Genotipe lainnya peka pada 5 ppm A1 (TabelTabel 3. Tingkat Ketenggangan Genotipe Jagung pada Berbagai Konsentrasi A1
Berdasarkan Panjang A k x Relatif (PAR)
No Genotipe Aluminium (ppm)
2.5 5 10 20 40
SATP-2(S2)-C6-SO
SATP- l(S2)-C6-SO
BK-@IS)-C2-5-1
BK-(HS)-C2- 1 1
-
1BK-(HS)-C2- 1 13-1 .-
BK-@IS)-C2-129- 1
BK-(HS)-C2-55- 1
AMATL-(HS)-C2-SO
AMATL-(S 1)-C2-43- 1
AMATL-(S 1)-C3
MRSS-l(S1)-C 1-20-1
MRSS-1(si)-c1-21-1
MRSS-l(S1)-C1-29-1
MRSS-l(S1)-C1-57-1
MRSS-l(S1)-C1-123-1
16. CML. 3 5 8 77.1 T 60.8 P 57.9 P 40.1 P 41.0 P
19. Antasena 93.2 T 77.3 T 73.1 M 49.7 P 45.0 P
20. Bisma 85.9 T 75.4 T 50.3 P 38.3 P 35.9 P
2 1 . Lokal Koasa 97.8 T 74.8 T 61.9P 48.0P 47.4 P
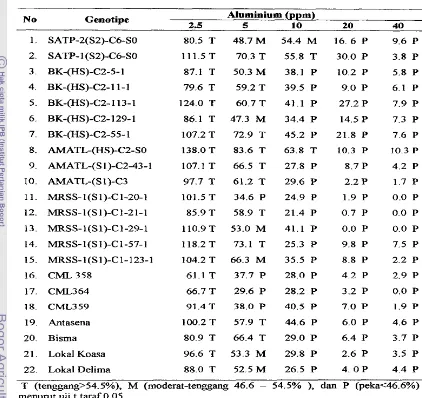
Pertarnbahan panjang akar relatif (PPAR) berbeda nyata antar setiap konsentrasi
Al,
gemtipe maupun intmaksi antara A1 dengan genotipe (Lampiran 3). Nil& peubah - PPAR menurun dengan meningkatnya konsentrasi Al, kecuali pada konsentrasi 2.5 pprnA1 beberapa genotipe justru rnempunyai PPAR yang cenderung lebih tinggi dibanding dengan tanpa Al, bahkan genotipe BK-@IS)-C2-f 13-1, AMATL-(HS1)-C2-SO, dan MRSS-l(S1)-C1-57-1 rnempunyai PPAR pada konsentrasi 2.5 pprn A1 nyata lebih tinggi
dibanding dengan tanpa Al. Hasil uji t terhadap PPAR menunjukkan bahwa ketenggangan terhadap Al setiap genotipe jagung dapat dikelompokan ke dalam: 1) tenggang @PAR >54.5%). 2) moderat-tenggang (PPAR 46.6 - 54.5%), dan 3) peka
(<46.60/0).
Berdasarkan pengelompokan ketenggangan tampak semua genotipe tergolong tenggang pada konsetrasi A1 2.5 ppm. Tingkat ketenggangan terhadap A1 antara genotipe akan beragam pada konsentarasi 5 -10 pprn Al, akan tetapi jika konsentrasi Al ditingkatkan menjadi 20 pprn dan 40 ppm Al maka semua genotipe tergolong peka. Genotipe SATP- 1 (S2)-C6-SO dan AMATL-HS-C2-SO tenggang hingga konsentrasi 10 pprn Al. Genotipe SATP-I (S2)-C6-SO, BK-(HS)-C2-113-1, BK-(HS)-C2-11-1, BK- @S)-C2-55-1, Ah4ATL-(HS)-C2-43-1, AMATL-(S1)-C3, MRSS-I(S1)-Cl-21-1,
MRSS-l(S1)-Cl-57-1, Antasena, d m Bisma hanya tenggang psda konsentrasi 5 pprn A1 dan peka pada 10
-
40 ppm Al. Genotipe SATP-2(S2)-C6-SO tergolong moderat-tenggang pada konsentrasi 5 dan 10 pprn Al, genotipe SATP-2(SZ)-C6-S0, BK-(HS)-
C2-129-1, MRSS-l(S 1)-C1-123-1, Lokal Koasa clan Lokal Delima moderat tenggang pada konsentrasi 5 pprn Al dan peka pada 10 ppm Al. Genotipe lainnya tergolong peka
Tabel 4. Tingkat Ketenggangan Genotipe Jagung pada Berbagai Konsentrasi A1
Berdasarkan Pertambahan Panjang Akar Relatif PPAR)
No Genotipe Aluminium (ppm)
2.5 5 10 20 40
1. SATP-2(S2)-C6-S0 80.5 T 48.7 M 54.4 M 16. 6 P 9.6 P
2. SAW-1 (S2)-C6-SO 111.5 T 70.3 T 55.8 T 30.0 P 3.8 P 3. BK-(HS)-C2-5-1 87.1 T 50.3 M 38.1 P 10.2 P 5.8 P
4. BK-(HS)-C2- 1 1
-
1 79.6 T 59.2 T 39.5 P 9.0 P 6.1 P5. BK-@IS)-C2-113- 1 124.0 T 60.7 T 41.1 P 27.2 P 7.9 P
6. BK-@IS)-C2- 129-1 86.1 T 47.3 M 34.4 P 14.5 P 7.3 P
7. BK-@IS)-C2-55-1 107.2 T 72.9 T 45.2 P 21.8 P 7.6 P
8. AMATL-(HS)-C2-SO 138.0 T 83.6 T 63.8 T 10.3 P 10.3 P
9. AMATL-(S 1)-C2-43-1 107.1 T 66.5 T 27.8 P 8.7 P 4.2 P 10. AMATL-(S1)-C3 97.7 T 61.2 T 29.6 P 2.2 P 1.7 P
11. mSS-l(S1)-Cl-20-1 101.5 T 34.6 P 24.9 P 1.9 P 0.0 P
12. MRSS-l(S1)-Cl-21-1 85.9 T 58.9 T 21.4 P 0.7 P 0.0 P
13. h4RSS-l(S1)-Cl-29-1 110.9T 53.0 M 41.1 P 0.0 P 0.0 P
14. ~ ~ ~ ~ - 1 ( ~ 1 ) - ~ 1 - 5 7 - 1 118.2 T 73.1 T 25.3 P 9.8 P 7.5 P
15. MRSS-l(S1)-C1-123-1 104.2 T 66.3 M 35.5 P 8.8 P 2.2 P
16. CML.358 61.1 T 37.7 P 28.0 P 4.2 P 2.9 P
17. CML364 66.7 T 29.6 P 28.2 P 3.2 P 0.0 P
18. CML359 91.4 T 38.0 P 40.5 P 7.0 P 1.9 P
19. Antasena 100.2 T 57.9 T 44.6 P 6.0 P 4.6 P
20. Bisma 80.9 T 66.4 T 29.0 P 6.4 P 3.7 P
2 1. Lokal Koasa 96.6 T 53.3 M 29.8 P 2.6 P 3.5 P
Peubah bobot akar relatif (BAR) nyata d i p e n g a d oleh falctor konsentrasi Al,
genotipe maupun interaksi antara konsentrasi A1 dan gemtipe (Lampiran 3). Pemberian
Al
pada konsentrasi 2.5 pprn menstirnuhi pertumbuhan akar beberapa genotipe terutama pa& genotipe BK-@IS)-C2-113- I , AMATL-(HS)-C2-SO, AMATL-(S I)-C2-43-1, dan MRSS-l(S1)-Cl-57-1. HaI ini terlihat pada nilai BAR yang nyata lebih tinggi dibanding tanpa Al. Secara umum penjngkatan konsentrasi Al di atas 2.5 pprn akan menyebabkan semakin rendahnya
nilai
BAR dari semua genotipe.Pengelornpokan ketenggangan Al setiap genotipe berdasarkan nilai BAR dari hasil uji t sebagai berikut: 1) tenggang
(BAR
>71.2 %), 2) moderat-tenpgang P A R 65.2 - 71.2%), dan 3) peka ( 6 5 . 2 % ) . Semua genotipe tergolong tenggang pada konsentrasi 2.5 pprn A1 dan peka pada konsentrasi >20 pprnA4
kecuali genotipeMRSS-1 (S1)-CI -57-1 yang mash tenggang hingga pada konsentrasi 20 pprn Al. Pada
konsentrasi 5-10 pprn terjadi pemisahan antara genotipe tenggang dengan genotipe peka (Tabel 5). Genotipe MRSS-l(S1)-C1-29-1 tenggang pada konsentrasi 5 ppm d m 10 pprn
Al. Genotipe AMATL-(HS)-C2-SO dan AMATL-(SI)-C2-43-1 tengang pada konsentrasi 5 pprn A1 dan moderat-tenggang pada 10 pprn Al. Genotipe SATP-I(S2)- C6-SO, BK-S)-CZ-5-1, BK-@IS)-C2-113-1, BK-@IS)-C2-55-1, BK-(HS)-C2-113-1, AMATL-(S1)-C3, MRSS-l(S1)-Cl-20-1, MRSS-l(S1)-CI-123-1, CML364, dan lokal
Tabel 5. Tingkat Ketenggangan Genotipe Jagung pada Berbagai Konsentrasi Al Berdasarkan Bobot Akar Relatif (BAR)
No Genotipe Aluminium @pm)
2.5 5 10 20 40
1. SATP-2(S2)-C6-SO 70.8 T 55.6 P 49.3 P 45.3 P 33.6 P 2. SATP- l(S2)-C6-SO 104.8 T 85.7 T 60.0 P 59.4 P 44.0 P
3. BK-(HS)-C2-5-1 89.4 T 72.6 T 47.2 P 42.5 P 33.3 P
9. AMATL-(S 1)-C2-43- 1 127.4 T 90.3 T 66.4 M 53.7 P 51.0 P
19. Antasena
20. Bisma
2 1, Lokal Koasa 111.1 T 61.2 P 33.5 P 30.3 P 28.0 P
22. Lokal Delima 73.7 T 74.6 T 43.1 P 29.3 P 27.1 P
[image:45.543.58.481.73.494.2]Bobot tajuk relatif (BTR) nyata dipengaruhi oleh faktor konsentrasi
Al,
genotipe,dan
interaksi antara konsentrasi A1 dengan genotipe (Lampiran 3). Pemberian 2.5 pprn A1 juga cenderung meningkatkan bobot tajuk pada beberap genotipe, terutama pada genotipe BK-@IS)-C2-113-1 yang mempunyai nilai BTR 36% lebii tinggi dibandii tanpaAt
Nilai BTR semakin menurun dengan meningkatnya konsentrasi A1 di atas 2.5 ppm, tingkat penurunan pada setiap kenaikan konsentrasi bewariasi antar genotipe (Tabel 6).Hasil uji t terhadap niIai BTR menunjukkan bahwa tingkat ketenggangan antara
genotipe berbeda pada setiap konsentrasi Al. Ketenggangan setiap genotipe berdasarkan nilai
BTR
dapat dikelornpokkan ke ddarn: 1) tenggang adalah yang mempunyai nilai BTR >69.3%, 2) moderat-tenggang dengan nilai BTR 62.9 - 69.3%, dan 3) peka dengannilai BTR (62.9%. Berdasarkan pengelompokan ini, maka semua genotipe tergolong
tenggang jika pemberian A1 hanya 2.5 ppm clan semuanya peka jika pemberian A1 s e h y a k 40 pprn Al. Genotipe MRSS-l(S1)-Cl-57-1 dan AMATL-(S1)-C2-43-1 masing-masing tergolong tenggang sampai pada konsentrasi 20 pprn A t Sebanyak 6 genotipe yang tenggang pada konsentrasi 5 pprn tetapi peka pada konsentrasi >10 pprn
AI,
yaitu : SATP-l(S2)-C6-SO, AMATL-(S1)-C3, MRSS-l(S1)-Cl-21-1, MRSS-l(S1)-CI -29-1, MRSS-1 (S1)-CI-123-1, dan CML.359. Genotipe BK-(HS)-C2-I 13-1, clan BK-
(HS)-C2-55-1. Genotipe AMATL-(HS)-C2-SO tergolong genotipe yang moderat-
tenggang hingga pemberian 10 pprn Al, sedangkan genotipe lainnya sudah tergobng
Tabel 6. Tingkat Ketenggangan Genotipe Jagung pada Berbagai Konsentrasi A1 Berdasarkan Bobot Tajuk Relatif (ESTR)
N o Genotipe Aluminium (ppm)
2.5 5 10 20 40
10. AMATL-(S 1)-C3 111.7 T 71.3 T 35.7 P 43.6 P 44.8 P
11. MRSS-l(S1)-CI-20-1 83.3 T 69.2 M 45.2 P 37.3 P 38.2 P
12. MRSS-I(S1)-CI-21-1 118.1 T 92.1 T 55.0 P 61.9 P 52.5 P 13. MRSS-I(S1)-CI-29-1 104.9 T 86.6 T 53.4 P 51.2 P 55.4 P
14. MRSS-l(S1)-Cl-57-1 102.3 T 70.4 T 58.2 M 72.4 T 45.9 P
15. MRSS-l(S1)-Cl-123-1 82.0 T 82.0 T 49.7 P 43.5 P 43.4 P 16. CML 358 71.8 T 83.7 T 71.1 T 64.0M 59.4 P 17. CML364 93.8 T 69.0 M 44.8 P 38.2 P - 35.2 P 18. CML359 85.3 T 81.9 T 47.8 P 57.6 P 51.6 P
19. Antasena 71.4 T 49.6 P 39.4 P 37.8 P 24.9 P
20. Bisma 98.7 T 67.3 M 29.6 P 43.3 P 35.7 P
21. Lokal Koasa 100.8 T 42.6 P 25.3 P 31.8 P 29.6 P 22. Lokal Delima 84.9 T 68.4M 36.7 P 32.5 P 29.5 P
Konse-i
Al
nntuk Pengujian Ketenggangan pa& Tanaman JaglrngHasil analisis terhadap beberapa peubah, terlihat bahwa penentuan konsentrasi
pengujian ketenggangan Al pada tanaman jagung dapat ddakukan dengan menggunakan peu- panjang &ar relatif (PAR), pertmbahm panjang akar relatif (PPAR), bobot kering akm relatif (BAR), maupun bobot tajuk relatatif (BTR). Hal ini karena peubah-
peubah tersebut dipengaruhi o k h adanya interaksi antara konsentrasi A1 dengan
genoiipe, dimana genotipe beragam ketenggangnya akibat adanya perbedaan konsentrasi
Al. Sebaran data dan ~ngelompokan tingkat ketenggangan tanaman terhadap cekarnan A1 bedasarkan nilai PPAR tenrtama pada konsentrasi 5 ppm dan 10 ppm A1 lebih
beragam dibading peubah lainnya, oleh karena itu penggunaan peubah pertambahan panjang akar relatif lebih baik digunakan untuk rnenentukan taraf konsentrasi pengujian
ketenggangan A1 pada tanaman jagung dibanding dengan peubah lainnya. Pengunaan peubah pertambahan panjang akar dapat menghindari variasi peubah akibat perbedaan
genetik clan perbedaan vigor awal kecambah setiap genotipe.
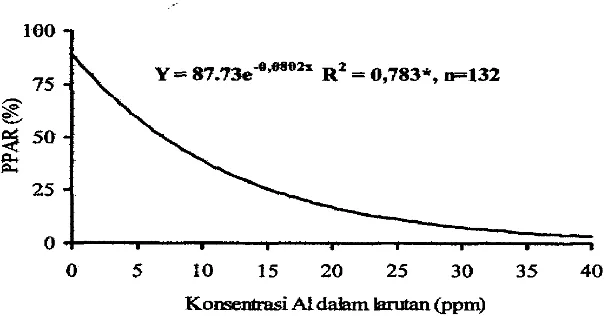
Pertambahan panjang akar relatif (PPAR) menunjukkan adanya hubungan dengan konsentrasi Al, hubungan tersebut bersifat 10% yaitu Y = 87.73e4.0802X dengan
nilai R' = 0.783* (Gambar I). Apabila nilai rata-rata PPAR pada seluruh genotipe (50.03%) dianggap sebagai pemisah antar setiap galur, maka taraf konsentrasi untuk pengujian ketenggangan Al pada tanaman jagung adalah 7 ppm Taraf ini mendekati konsentrasi yang digunakan oleh Parentoni et aL(1997) yaitu pada konsentrasi 6 ppm Af, tetapi agak berbeda dengan yang disarankan oleh Kasim
dan
Wassom (1990) dengan0
5
10 15 20 25 30 35 40 [image:49.543.106.409.61.220.2]Ko~lsentrasi
At
dalam hutan (ppm)Gambar 1 . Hubungan antara pertambahan panjang akar relatif (PPAR) dengan konsentrasi At dalam larutan
Penapisan Genotipe Jagung untuk Etisiensi Hara P
Ehsil sidik ragam terhadap b b o t kering biimas munjkkan pengaruh nyata dari Eaktor konsentmsi AI, gemtip, dan interaksi antara konsentrasi A1 dengan genotipe ( L a m p a
3-
Konsentrasi A1 khubungaxi hadratik dengan berat kering biomas Sifatkmdmtik dari hubungan antara konsentrasi At dengan bobot kering Somas masing-
masing genotipe disajikan pada Tabel 7. Sebanyak 8 genotipe yang mempunyai b o w kering biomrts r d d i (yang &pat dicaprti vleh suatu gewtipe) aatara 1887.41
-
2104.5 mg/tanamau yang nyata
kb&
tiaggi d a h n c h g rata-rats genotipe (1743.4mg/tanztmm), kedelapan genotipe tersebut adalah :
SATP-2(S2)-C&SO, SATP-l(S2)-
c6-SO,
BK-(HS)-C~-~
13-1, BK+S)-C~-SS- I, AMATL-(HS)-CZSO,m n ( s
ir
agronomik antara 152.9 - 200.7Yo yang nyata lebh tin& dibanding de.ngan rata-rah
Tabel 7. Model mtematik (Y) Persuman Hubungan antara P dengan Bdmt KerJng Riomas, Robot Kering Etiornas Mahima1 Dugaan,
dan
Nilai E3isiensi Agronomik PAGIZ) dari Genotipe JagungSATP-2(S2)-C6-SO
SATP-1 (S2)-C6-SO
BK-(HS)-C2-5-1
BK-@IS)-C2-11-1
BK-(HSfC2- 1 13- 1
BK-(HS)-C2-I 29-1
BK-(HS)-C2-55-1
AMATL-(HS)-C2SO
AMATL-(S1)-C2-43-1
AMATL-(S1)-C3
MRSS-1 (S 1 )-C 1-20-1
MRSS-l(SI)-Cl-21-1
MRSS-l(S1)-C 1-29-1
MRSS-l(S1)-Cl-57-1
MRSS-l(Sl)-Cl-123-1
CML 358
CML364
CML359
22. Ld1t1 Delitmr 0.59 853 103.4 -5.1 10.1 1377.1 135.8
Ra%-~ata 12.0 1603.9 137.3
[image:50.543.57.485.105.517.2]E f i h Sempan, E m iPenggunuan, dun Raib
Eflriensi
P
Analisis kadar P jariagan biomas dilakukan terhadap trutaman jagwig yang diberi perlakuan 5 ppm A1 untuk menentukan efisiensi P (serapan, pengg- dan ratio efisiensi) antam genotipe. KonsentTasi 5 ppm P dianggap sebagai kondisi P kurang bagi pemmbuhan tanaman secara mid. Menurut Polle
dan
Konzak (1990) suatu tanaman yang tenggag dan efisien menyerap P adalah tanaman yang mampu mengambil P lebih banyak dalam kondisi suplai P rendah. Efisiensi hara P suatutanarnan ditunjukkan dengan tingginya bobot kering yang dihasilkan d a b kondisi P rendah (Rao et al. 1999).
Sebanyak 22 genotipe yang diuji ternyata mempunyai tingkat efisiensi b r a P
yang beragam dan 14 diantaranya tergolong efisien. Dari 14 genotipe tersebut hanya genotipe AMATL-(HS)-C2-SO yang efisien hara P baik berdasarkan efisiensi serapan, efisiensi penggunaan, maupun rasio efisiensi. Genotipe SATP-Z(S2)-C6-S0, SATP-
1 (S2)-C6-SO, BK-@S)-C2- 1 13- 1, CML364, dan Bisma efisien P dalam penggunaan dan ratio efisiensi. Genotipe AMATL-(S 1)-C3 dan MRSS- 1 (S 1 )-C 1 -2 1
-
1 efisien P dari segi ratio efisiensi. Genotipe lainnya seperti BK-(HS)-C2-11-1, BK-@IS)-C2-129-1,Tabel 8- Robot kering biornas, kadar P, v a n
P,
efisieii serapan P (ESP), e&iensi penggunaan P(EPF),
dan r a s h efisiensi P (REP) setiap genotipe pada-
5 m PNe Geeotipe
P
EPP
REP
b h s
(mg)
e"')
img) SATP-2(S2)-C6-S0SATP-1 (SZ)-C6-SO BK-(HS)-C2-5- 1 BK-@IS)-C2-11 -I BK-(HS)-C2-113-1 BK-(HS)-C2-129-1 BK-(HS)-C2-55- 1 AMATyRs)-C2SO AMATL-(S 1)-C2-43- 1 AMATL-(S 1)-C3 MRSS-l(S1)-C1-20-1 MRSS-l(S1)-C1-21-1 MRSS-l(S1)-C1-29-1 MRSS-I(Sl)-cI-57-l MRSS-l(S1)-CI-123-1 CML 358
[image:52.539.57.503.87.515.2]Xonsentrnsi
P
untuk Pengujian Efisiemi Hara pada Tanaman JagungHubungan antara konsentrasi P dengan bobot kering biomas menunjukkitn sifat
yang kuadratik (Garnbar 2.) dengan model maternatiknya sebagai berikut: Y= 0.9328 +
0.1175X
-
0.005 X' (R' = 0.62*). Berdasarkan model ini maka pemberian P sampai pada 11.5 pprn akan rneningkatkan bobot kering biomas, akan tetapi jika konsentrasi Pdinaikkan di atas 11.5 ppm P akan menurunkan bobot kcring biomas. Menurut Marschner (1995) pemunbuhan optimal adalah pada kandungan hara antara taraf kritikal
defisiensi dengan kritikal toksisitas, dimana secara praktis titik kritikal defisiensi adalah
90-95% dari pertumbuhan maksimal. Berdasarkan kriteria ini, maka titik kritis
defisisiensi P adalah pa& taraf 6.0 pprn P, sedangkan titik kntis toksisitas adalah 11.5 pprn P. Taraf kritis ini mendekati yang digunakan oleh Robert et ai. (2000), yaitu pada konsentrasi 5 dan 10 pprn P. Oleh karena itu konsentrasi 6 dan 11.5 pprn P digunakan
dalam pengujian lanjutan.
2.0
-
-
Y = -0.005~' + 0 . 1 1 7 5 ~ + 0.9328 ~ % . 6 2 " ==I32b4 1.6
-
E
3 1 . 2 -
3
.
-
8 0.8-
Y
!
Kritikal toksisitas+-.
0
%
0.4-
!a Krifikul 3efisien
0.0
.
d0 5 10 15 20
Konsentrasi P d a r n larutan (pprn)
Fisiologi Hsra P Tanaman Jagung dalam Kondisi Cekaman
A1
Untuk mengetahui efisiensi hara P pada tanaman jagung dalam keadaan cekaman Al digunakan 2 genotipe, yaitu genotipe tenggang (WTL-(HS)-C2-SO) dan peka
(CML364), yang efisien terhadap hara P. Genotipe ini ditumbuhkan pada larutan hara
yang mengandung kombinasi
AI-P,
yaitu 1) Iarutan 2.5 ppmAl
+
6 ppm P, 2) 2.5 pprnA l + 11.5ppmP,3)larutan7ppmAl+ 6ppmP,dan4)7ppinAl+ l i . 5 p p m P .
Penampiha Agronomi
Faktor genotipe dan larutan (Al-P) menunjukkan pengaruh nyata terhadap semua
peubah komponen agronomik, kecuali pengaruh %or genotipe terhadap ratio bobot
kering tajuk-akar. Interaksi antara genotipe dengan larutan (AI-P) berpenganth nyata
terhadap peubah panjang akar relatif (PAR), pertam- panjang akar (PPAR), bobot
akar, bobot tajuk, dan bobot total biomas, sedangkan terhadap panjang akar dan ratio
bobot tajuk-akar tidak berpengaruh nyata (Lampiaran 4.).
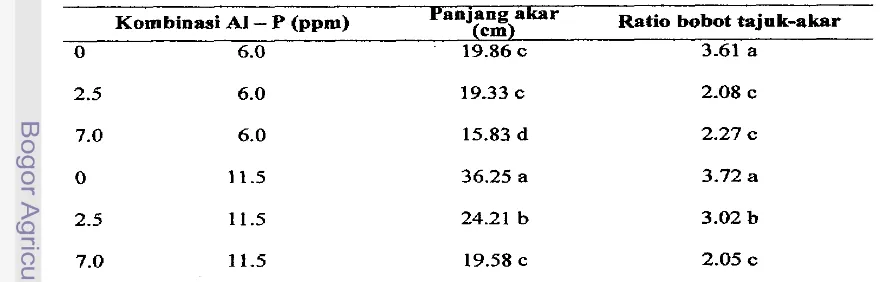
Akar genotipe AMATL-HS-C2-SO lebii panjang dibanding CML364, sedan&= ratio bobot kering tajuk-akar tidak berbeda nyata (Tabel 9). Pemberian A1 pada larutan hara m e n g b b a t pemanjangan akar maupun ratio bobot kering tajuk-akar. Makin tinggi konsentrasi A1 makin terhambat pemanjangan akar dan bobot kering akar (Tabel 10).
Genotipe AMATL-HSC2-SO yang ditumbuhkan pada konsentasi A1 rendah (2.5 ppm) mempunyai nilai PAR lebih tinggi 15% jika dikombinasi dengan P kurang (6
konsentrasi A1 tinggi (7 ppm), maka nilai PAR lebii rendah 15% dan 40% dibandiig tanpa A1 masing-masing pada kombinasi 6
dan
1 1.5 pprn P. Genotipe peka CML364 meskipun ditumbuhkan pada konsentrasi Al rendah (2.5 ppm) tetap mempunyai d a i PAR yang lebih rendah dibanding tanpa A1 dan semakin rendah pada konsentrasi 7 pprnAL Genotipe AMATL-HS-C2-SO mempunyai nilai PAR yang lebih tinggi d i b d i g
Tabel 9. Rata-rata Panjang Akar dan Ratio Bobot Kering Tajuk-Akar Tanaman Jagung Genotipe Tenggang AMATL-HS-C2-SO dan Peka CML364
Genotipe Panjang akar (cm) Ratio bobot tajuk-akar
AMATL-HS-C2-SO 25.38 a 2.80 a
CML 364 19.64 b 2.78 a
i~ uti oleh hurut yang sarna menunjukkan tidak bebeda nyata menurut D W T
T,"d
$2
Tabel 10. Rata-rata Panjang Akar dan Ratio Bobot Kering Tajuk-Akar Tanaman J-g setiap Kornbinasi Larutan Hara AI-P
Kombinasi A1
-
P (ppm) Panjang akar (em) Ratio bobot tajok-akar0 6.0 19.86 c 3.61 a
[image:55.546.45.488.345.486.2]Gambar 3. Panjang akar relatif (PAR) genotipe tenggang (AMATL-HS-CZ) dan peka (CML364) di larutan hara ALP
Genotipe tenggang (AMATL-HS-C2-SO) juga mempunyai nilai pertambahan
panjang akar relatif (PPAR) yang lebih tinggi (57%) jika ditumbuhkan pada larutan hara
dengan konsentrasi 2.5 ppm A1 dibanding tanpa Al pada dikombinasi P kurang (6 ppm), akan tetapi lebih rendah 49% apabila larutan hara dengan konsentrasi 2.5 ppm A1
dibanding tanpa Al apabila dikombinasi dengan cukup P (1 1.5 ppm). Genotipe yang
peka C E 3 6 4 , meskipun ditumbuhkan pada Iarutan yang mengandung A1 rendah (2.5 ppm Al) baik kurang P maupun cukup P mempunyai nilai PPAR lebih rendah dibanding
.tanpa Al, penurunannya 60% jika kurang P dan 58% jika cukup P. Penurunan nilai
PPAR lebih tinggi jika AI ditingkatkan konsentrasinya menjadi 7 ppm. Penurunan nilai
PPAR genotipe AMATL-HS-C2-SO lebih rendah dibanding CML364 jika ditumbuhkan
6 0 11.5 6.0 11,5 Konsenbasi P
daiam
lamtan (ppm)Gambar 4. Pertambdun panjang akar relatif (PPAR) gemtipe tenggang (AMATL-HS-C2) dan peka (CML364) pada larutan h a Al-P
Pemberian 2.5 pprn A1 dikombinasi dengan 6 pprn P pada genotip AMATL-HS- C2-SO cenderung meningkatkan laju tumbuh akar. Apabila konsentrasi A1 ditingkatkan
menjadi 7 pprn maka Iaju tumbuh akar cenderung l e b i terhambat. Pada konsentrasi 1 1.5 pprn P, hanya dengan pemberian 2.5 pprn A1 laju tumbuh akar nyata bbih lambat dibanding tanpa Al. Pemberian A1 pada genotipe CML364 yang peka dapat menekan
laju tumbuh akar h i k pada kondisi kurang P maupun cukup P. Penghambatan laju
tumbuh akar oleh A1 pada genotipe AMATL-HS-C2-SO lebii rendah dibanding CMI.364, kecuali pada cekaman 7 pprn A1 dengan larutan yang diberi cukup P. Laju tumbuh akar lebih lambat pada larutan yang kurang P dibanding dengan cukup P baik
Tabel 11. Laju Tumbuh Akar, Bobot Kering Akar, dan Bobot Kering Tajuk Genotipe Tenggang AMATL-HS-C2-SO dan Peka CML364 yang Ditumbuhkan pada Larutan hara AI-P
xornbinasi Laju tumbuh akar Bobot kering akar Bobot kering tajuk
A I - P (mm/hari) (mdtanaman) (mg/tanaman)
AMATG AMATL
AMATL CML364 ( P P ~ ) HS-C2SO CML364 HS-CZSO CML364 HS-C2-SO
0 6.0 0.023 ef 0.032 ed 477.3 bc 480.3 bc 1676.8bc 1778.3 b
2.5 6.0 0.032 de 0.015 fg 514.5 b 372.5 d 1124.2ed 745.0 ef
7.0 6.0 0.013 fg 0.013 fg 265.5 e 140.8 f 623.7 fg 306.28 0 11.5 0.074 a 0.061 b 699.5 a 525.0 b 2747.7 a 1849.3 b
2.5 11.5 0.049 bc 0.032 de 548.5 b 409.5 cd 1334.2cd 1536.2bc
7.0 11.5 0.040 cd 0.009 g 364.7 d 209.3 ef 743.3 ef 430.3 g
Rata-rata 0.038 0.027 478.3 356.2 1375.0 1107.5
Angka yang diikuti oleh huruf yang sarna menunjukkan tidak bebeda nyata menurut DMRT taraf 0.05
Pada keadaan tercekam
AI,
pembentukan bahan kering (akar dan tajuk) tanamanterhambat. Pada konsentrasi 2.5 pprn A1 dan P kurang genotipe AMATL-HS-C2-SO mempunyai bobot kering akar cenderung lebih tinggi dibanding tanpa Al. Makin tinggi
konsentrasi A1 menyebabkan pembentukan bahan kering semakin rendah. Pembentukan
bahan kering akar genotipe AMATL-HS-C2-SO lebih banyak dibanding CML,364
apabila ditumbuhkan di larutan yang mengandung
Al
dan P yang sama, sedangkan bobotkering tajuk genotipe AMATL-HS-C2-SO lebih banyak dibanding CML364 jika
Kadar P tajuk, efisiensi pengunaau P, dan ratio A1 pada akar-tajuk dipengaruhi secara nyata oleh perbedaan konsentrasi Iarutan (Al-P), tetapi tidak dipengaruhi secara nyata oleh perbedaan genotipe maupun interaksi antara genotipe dengan konsentrasi
larutan ALP. Peubah kadar P akar, ratio kadar P pada akar-tajuk, total serapan P,
efisiensi serapan P, ratio efisiensi P, kadar ahuninium pada akar dan tajuk dipengaruhi secara nyata
oleh
interaksi antara genotipe dengan konsentrasiIarutan
ALP @.ampiran6 dan 7).
Kadar P tajuk dan efisiensi penggunaan P (EPP) genotipe AhG4TL-HS-C2-SO nyata lebih tinggi dibanding Ch4L364, sedangkan ratio A1 pada aka.-tajuk antara genotipe AMATL-HS-C2-SO dengan CML364 tidak menunjukkan perbedaan yang
nyata (Tabel 12). S e m umurn makin tinggi konsentrasi A1 menyebabkan semakin rnenurunnya kadar P &lam tajuk, etkiensi p e n g u m P @PP) clan ratio kadar A1 pada akar-tajuk. Apabila konsentrasi Al sarna, maka peningkatan P di larutan meningkatkan
kadar
P &lam tajuk tetapi tetapi tidak terhadap EPP clan ratio A1 dalam akar-tajukTabel 12. Kadar P dalam Tajuk, Efisiensi Penggunaan P (EPP) dan Ratio Kadm A1
dalam Akar-Tajuk Tanaman Jagung Genotipe Tenggang AMATL-HS-C2- SO dan Peka CML364
Genotipe P tajuk (ppm) EPP Ratio
A1
Akar-TajukAh4ATL-HS-C2-SO
4027 a 4183 a 14.1 aCML 364 3463 b 3191
b
12.7 aTabel 13.