PENDAHULUAN
Salah satu kendala dalam pengembangan peternakan khususnya sapi perah di Indonesia adalah ketersediaan pakan baik itu kualitas maupun kuantitasnya. Kondisi tersebut juga disebabkan oleh semakin menurunnya daya dukung lingkungan terhadap penyediaan pakan. Konversi lahan hutan dan lahan pertanian untuk kebutuhan nirpertanian berdampak pada vegetasi dan resapan air. Dampaknya adalah mineral esensial yang bervalensi rendah dan berbobot atom kecil hanyut dan mineral toksik polivalen dan berbobot atom besar tinggal dalam tanah. Situasi ini menyebabkan terjadinya distorsi status mineral pada tanah, tanaman, dan kemungkinan ternak.
Suplementasi mineral esensial merupakan upaya alternatif untuk mengatasi distorsi status mineral khususnya defisiensi mineral. Pemakaian mineral organik menjadi pilihan karena lebih mudah diserap oleh tubuh dan resiko antagonis mineral lebih kecil. Defisiensi status mineral Zn dilaporkan oleh Little (1986) bahwa kandungan Zn pakan ternak ruminansia berkisar antara 20 dan 38 mg kg-1bahan kering. Jika defisiensi terus terjadi, dapat menyebabkan parakeratosis pada jaringan usus dan kelenjar ambing sehingga menurunkan produktivitas ternak. Tembaga dibutuhkan sebagai komponen seruloplasmin, dismutase superoksida, oksida lisil, dan oksidase sitokrom (NRC 2001). Sutrisno (1983) melaporkan bahwa pada ruminansia status Cu adalah mulai dari marjinal sampai defisien. Sementara pada kebanyakan ternak, Cu sangat sulit diserap, pada ternak ruminansia hanya 1 sampai 3% (McDowell 1992). Selen merupakan bagian integral dari enzim glutation peroksidase. Selen kurang dapat diabsorbsi oleh ternak ruminansia terutama dalam bentuk selenit. Penggunaan bentuk organik lebih efektif dibandingkan dengan anorganik. Kromium adalah mineral esensial yang berhubungan dengan kerja insulin. Kromium yang esensial adalah Cr3+, namun sulit diserap, sedangkan Cr6+ mudah larut tetapi bersifat toksik. Oleh karena itu, satu-satunya bentuk pasokan kromium ke dalam tubuh ternak adalah dalam bentuk ikatan ligand organik (Sutardi 2002).
tinggi, seringkali menjadi tidak efisien bagi ternak ruminansia karena protein tersebut didegradasi dalam rumen, sementara lemaknya tidak tersedia bagi mikrob rumen karena terikat oleh struktur organik lainnya. Proses pemanasan (sangrai) akan meningkatkan efisiensi pakan kualitas tinggi sehingga protein akan diproteksi dari degradasi rumen dan lemak menjadi tersedia bagi mikrob rumen. Oleh karena itu kedelai sangrai dapat meningkatkan mutu ransum ternak ruminansia. Kacang kedelai juga merupakan sumber asam linoleat yang merupakan asam lemak esensial.
Suplementasi lemak dalam ransum ternak ruminansia sering digunakan untuk meningkatkan produktivitas ternak. Penggunaan minyak dalam ransum ternak ruminansia dapat mengganggu pertumbuhan mikrob rumen, karena asam lemak tak jenuh toksik bagi mikrob rumen bahkan asam linoleat adalah racun bagi protozoa. Oleh karena itu, suplementasi minyak dalam bentuk lemak yang diproteksi bertujuan supaya lemak tidak mengganggu fermentasi dalam rumen dan menyediakan asam lemak bagi induk semangnya. Penggunaan minyak jagung dan minyak ikan dalam ransum ternak ruminansia selain memasok asam lemak esensial juga diharapkan menjadi prekursor bagi sintesis asam lemak yang bermanfaat bagi kesehatan.
Sebagian masyarakat masih mempunyai persepsi yang kurang baik terhadap produk peternakan karena dianggap sebagai pemicu terjadinya penyakit jantung dan aterosklerosis. Pada kenyataannya conjugated linoleic acid (CLA) pada produk ternak ruminansia, termasuk susu, sangat bermanfaat bagi kesehatan manusia. Peranan CLA di antaranya adalah mengurangi aterosklerosis, antidiabetes, meningkatkan mineralisasi tulang, modulasi sistem kekebalan tubuh, mengurangi kejadian kanker payudara, lambung, kolon dan kulit (Lee et al. 1994; Bellury 1995; Nicolosi et al. 1997; Banni dan Martin 1998; Houseknecht et al. 1998).
produksi leukotrien (LT4) yang merupakan komponen sel darah putih dan mediator dalam sistem kekebalan tubuh (Sinclair 1993). Asam lemak omega-3 khususnya EPA dan DHA adalah komponen penting otak dan organ vital lain sehingga penting bagi kecerdasan. Minyak ikan ini juga dapat memperkaya CLA susu (Chilliard et al. 1999; Chouinard et al. 2001).
Tujuan Penelitian
1. Meningkatkan produktivitas ternak melalui suplementasi kacang kedelai sangrai, mineral organik, dan sabun mineral.
2. Mengevaluasi penggunaan suplemen pakan (feed suplement) sabun mineral dari minyak jagung dan minyak ikan, kacang kedelai sangrai dan mineral proteinat dalam upaya meningkatkan kadar CLA dan memasok omega-3 di dalam susu.
3. Memproteksi asam-asam lemak esensial dari degradasi mikrob rumen melalui pembuatan sabun mineral.
Manfaat Penelitian
1. Mendapatkan teknologi pembuatan mineral organik atau proteinat mineral untuk mengatasi defisiensi mineral.
2. Memperoleh teknologi pembuatan sabun mineral sebagai feed suplement
dalam memperkaya susu dengan asam-asam lemak yang bermanfaat bagi kesehatan.
3. Meningkatkan nilai gizi susu.
Hipotesis
TINJAUAN PUSTAKA
Sistem Pencernaan dan Penyerapan Nutrien pada Ternak Ruminansia
Proses pencernaan pada ternak ruminansia terjadi secara mekanis di mulut, fermentatif oleh mikrob di rumen, dan hidrolitis oleh enzim pencernaan di abomasum dan duodenum hewan induk semang. Sistem fermentasi dalam perut ruminansia terjadi pada sepertiga dari alat pencernaannya. Hal tersebut memberikan beberapa keuntungan yaitu produk fermentasi dapat disajikan ke usus dalam bentuk yang lebih mudah diserap, makan cepat, menampung makanan dalam jumlah banyak, mencerna serat kasar, dan menggunakan nitrogen nonprotein. Di samping keuntungan tersebut, terdapat kerugian karena banyak energi yang terbuang sebagai CH4 (6 sampai 8%) dan sebagai panas fermentasi (4 sampai 6%), protein bernilai hayati tinggi mengalami degradasi menjadi NH3, dan mudah menderita ketosis (Sutardi 1977).
Perut ruminansia terdiri atas empat bagian yaitu retikulum, rumen, omasum, dan abomasum. Retikulum mempunyai tiga katub penghubung, pertama menuju rumen, kedua menghubungkan dengan oesofagus, dan retikuloomasal. Fungsi utama retikulum adalah mengontrol perintah aliran pakan dan membentuk jalan pakan kembali ke oesofagus selama proses ruminasi. Rumen merupakan bagian terbesar perut ruminansia yang merupakan tempat terjadinya proses fermentasi. Omasum berperan dalam penyerapan air dan beberapa asam lemak. Omasum memiliki penghubung bagian depan dengan retikulum dan bagian belakang dengan abomasum. Digesta dipompa dari omasum langsung ke abomasum. Abomasum merupakan perut sederhana seperti pada nonruminansia. Bagian depan abomasum berhubungan dengan omasum dan usus halus bagian belakang. Abomasum memproduksi asam dan merupakan bagian saluran pencernaan tempat awal proteolisis. Hasil pencernaan tersebut akhirnya masuk ke dalam sistem peredaran darah (Collier 1985).
Pencernaan dan penyerapan karbohidrat
bahan makanan utama ruminan yang rata-rata mengandung 75% karbohidrat. Karbohidrat terutama dalam bentuk karbohidrat kompleks (selulosa, hemiselulosa), di samping yang mudah larut (pati, gula dan yang sejenis) (Parakkasi 1999).
Karbohidrat didefinisikan sebagai polihidroksi aldehida dan keton serta turunannya. Karbohidrat diklasifikasikan dalam tiga kelompok utama yaitu monosakarida (gula sederhana), oligosakarida (yang paling banyak terdapat di alam adalah disakarida), dan polisakarida (bentuk karbohidrat yang paling kompleks) (Pike dan Brown 1984).
Monosakarida, sesuai dengan namanya adalah bentuk karbohidrat yang paling sederhana. Monosakarida diklasifikasikan dalam bentuk aldehid dan keton, dan dikelompokkan berdasarkan jumlah atom karbon. Monosakarida dalam bentuk aldehid, berdasarkan jumlah atam karbon adalah triosa (gliseradehid); tetrosa (eritrosa, triosa); pentosa (xilosa, ribosa, arabinosa); heksosa (glukosa, galaktosa dan mannosa); dan heptosa. Glukosa dan fruktosa terdapat dalam bentuk bebas pada buah-buahan dan madu. Pentosa dan heksosa mempunyai peranan yang sangat penting dalam metabolisme sel. Pentosa siap disintesis dalam sel. Ribosa adalah pentosa yang sangat penting dalam sistem biologi dan dikonversi menjadi deoksiribosa dan ribitol. Ribosa dan deoksiribosa adalah komponen asam nukleat (RNA dan DNA). Ribosa juga adalah komponen nukleotida (ATP, ADP, dan AMP) (Pike dan Brown 1984).
Oligosakarida yang meliputi disakarida (sukrosa, maltosa, laktosa) dan trisakarida. Secara umum oligosakarida adalah gula-gula yang mengandung 2 sampai 10 unit monosakarida. Setiap gula diikat oleh hidroksil dengan melepaskan satu molekul air. Sukrosa dihidrolisis menjadi glukosa dan fruktosa. Laktosa ditemukan dalam susu dan dibentuk dari glukosa dan galaktosa. Maltosa mengandung dua molekul glukosa dan dibentuk dari hidrolisis pati (Pike dan Brown 1984).
dinding sel tanaman, bersama dengan lignin memberikan kekuatan pada dinding sel. Selulosa, hemiselulosa, pektin, gum juga disebut sebagai serat. Serat merupakan komponen tanaman yang tahan enzim pencernaan manusia (Pike dan Brown 1984). Selulosa dapat dicerna oleh enzim yang dihasilkan oleh mikrob rumen (Frandson 1996).
Karbohidrat yang memasuki rumen seperti selulosa, hemiselulosa, pati, dan karbohidrat yang larut air seperti fruktosa akan dipecah menjadi gula sederhana (Gambar1). Selulosa pertama dirombak menjadi selobiosa oleh enzim
-1,4 glukosidase, selanjutnya dikonversi menjadi glukosa. Hemiselulosa dikonversi menjadi pentosa juga oleh enzim -1,4-glukosidase, selanjutnya menjadi fruktosa-6-fosfat. Pati dikonversi menjadi maltosa dan selanjutnya menjadi glukosa atau glukosa-1-fosfat. Hasil pencernaan tersebut memasuki siklus glikolisis Embden-Meyerhoff dan menghasilkan piruvat sebagai produk akhirnya. Piruvat oleh bakteri rumen difermentasi menghasilkan asetat, butirat dan propionat (Collier 1985).
propionat. Kebanyakan propionat yang sampai di hati selanjutnya dioksidasi atau dikonversi menjadi glukosa. Asam butirat kebanyakan dikonversi menjadi keton selama penyerapan di epitelium rumen, sehingga kadar butirat sangat rendah dalam darah portal.
Pencernaan dan penyerapan protein
Protein adalah bahan organik esensial untuk semua sel dan menyusun hampir 18% berat tubuh ternak. Protein adalah polimer kompleks dengan berat molekul antara 5000 sampai 1 juta. Berat molekul yang besar terjadi karena terdiri atas asam-asam amino yang mengalami polimerisasi menjadi suatu rantai polipeptida. Penggabungan asam-asam amino tersebut terbentuk dari ikatan antara gugus amino (NH2) dari suatu asam amino dengan gugus karboksil dari asam amino yang lain dengan membebaskan satu molekul air (H2O) (Frandson 1996).
Ternak ruminansia mempunyai kemampuan unik untuk bertahan dan berproduksi tanpa sumber protein pakan karena adanya sintesis protein mikrob dalam rumen. Mikrob rumen dimanfaatkan oleh ternak bersama protein pakan yang bebas dari degradasi dalam rumen, memasok protein ke usus halus untuk dicerna dan diserap (Zinn dan Owens 1988).
Mikrob rumen menggunakan sumber N untuk sintesis protein yang berasal dari protein pakan dan N nonprotein (NPN). Sapi dapat tumbuh, bereproduksi, dan laktasi walau pakan hanya mengandung NPN sebagai sumber N. Secara umum mikrob rumen mengandung antara 20 dan 60% protein kasar dari bahan keringnya. Protein kasar bakteri rumen cenderung bervariasi dengan rataan 50% (±5%), di lain pihak protein kasar protozoa lebih bervariasi lagi dengan rataan 40% dengan kisaran 20 sampai 60% (Zinn dan Owen 1988).
tubuhnya. Lebih kurang mikrob rumen dapat menggunakan N amonia, karena itu mikrob lebih suka merombak AA tersebut menjadi amonia (Sutardi 1997).
Kebanyakan bakteri rumen dapat menggunakan N-NH3 sebagai sumber N walaupun beberapa spesies membutuhkan tambahan senyawa N (protein atau karbon dari asam amino tertentu) untuk pertumbuhan paling cepat atau efisien. Bakteri aktif menyerap N-NH3 sementara protozoa tidak. Penyerapan amonia meningkat jika konsentrasi amonia rumen meningkat. Keracunan amonia sering terjadi jika konsentrasi amonia melebihi 100 mg dl-1 (Zinn dan Owen 1988).
Amonia merupakan sumber N utama untuk sintesis de novo asam amino mikrob rumen. Konsentrasi N-NH3 5 mg% atau 3.57 mM dalam rumen sudah cukup untuk memenuhi kebutuhan N mikrob (Sutardi 1977). Sementara Agustin
et al. 1992; dan Erwanto et al. 1993 menyatakan bahwa kadar NH3 optimal untuk pertumbuhan mikrob rumen yang relevan dengan produksi ternak adalah 8 mM. Dengan demikian batasan 5 mg% (± 4 mM) adalah batas minimal dan 200 mg l-1 (± 14 mM) merupakan batas maksimal (Sutardi 1997). Sintesis MCP (microbial crude protein) sekitar 20 g 100g-1 total bahan organik, yang dicerna dalam rumen sekitar 9.6 sampai 33.2 g 100g-1 atau 14.5 g 100g-1 bahan organik, yang benar-benar difermentasi dalam rumen 7.6 sampai 20.3 g 100g-1 bahan organik (Zinn dan Owen 1988).
fenilalanina (Phe) dan triptofan (Trp), dan sejumlah asam amino yang bersifat semiesensial atau koesensial (Merchen dan Titgemeyer 1992).
Peptida atau asam amino bercabang sebagai sumber asam lemak rantai bercabang (branched chain fatty acid=BCFA) penting bagi pertumbuhan bakteri selulolitik. Pencernaan serat bergantung pada pasokan BCFA dari pakan atau mikrob lain dalam rumen. Defisiensi BCFA, amonia dan nutrien lain dapat menyebabkan energi (ATP) tidak seimbang. Proses fermentasi berlanjut tapi ATP yang diproduksi tidak digunakan mikrob untuk pertumbuhan. Sebagian bakteri dapat tumbuh tanpa sumber karbohidrat untuk energi. Strain bakteri tertentu membutuhkan struktur karbon dari asam amino esensial dan asam amino tersebut dapat diinkorporasikan ke dalam protein mikrob. Bakteri tertentu lebih menyukai peptida sebagai sumber N. Kemampuan untuk menggunakan asam amino atau peptida dapat mengurangi kebutuhan energi (Zinn dan Owen 1988).
Sebagian besar pencernaan dan penyerapan protein pascarumen, prosesnya sama dengan ternak nonruminansia. Ruminansia memiliki pH lambung dan duodenum yang lebih rendah sehingga dapat meningkatkan pencernaan protein. Semua protein larut oleh pepsin dan HCl dalam lambung, kemudian dicerna dalam usus halus.
Pencernaan dan penyerapan lemak
Rumen memodifikasi lemak dalam beberapa cara. Asam lemak ditemukan dalam bentuk yang sudah diesterifikasi dalam pakan konvensional, dan mikrob rumen menghidrolisis triasil gliserol ke dalam bentuk asam lemak bebas dan gliserol atau komponen lain, bergantung pada bentuk lemak pakan. Setelah proses lipolisis terjadi proses biohidrogenasi. Karena proses biohidrogenasi bergantung pada kehadiran karboksil bebas, lipolisis adalah obligator pertama dalam modifikasi lemak yang diesterifikasi dalam pakan. Tidak semua bakteri mampu melakukan lipolisis, dan protozoa tidak memiliki aktivitas lipolitik. Fraksi pakan yang mengalami lipolisis dan biohidrogenasi lebih rendah pada pakan biji-bijian, dengan demikian lebih banyak yang lewat ke lambung. Meskipun terjadi dengan cepat, lipolisis tetap dibatasi untuk mencegah kelebihan asam-asam lemak polyunsaturated bebas yang mengganggu pencernaan serat dan menghambat proses biohidrogenasi. Proses hidrolisis bergantung pada bentuk alami lemak pakan. Minyak tumbuhan seperti linseed oil dihidrolisis lebih sempurna (lebih 90%), sementara minyak ikan kurang dari 50% (Byers dan Schelling 1988).
Proses biohidrogenasi terjadi dalam rumen dan dilakukan oleh mikrob. Proses ini menghasilkan penambahan H+ pada asam-asam lemak ikatan rangkap. Biohidrogenasi asam lemak tidak jenuh terjadi melalui mekanisme penting, yaitu pemindahan atom H+ oleh mikrob. Jika proses tersebut sempurna maka semua ikatan rangkap dikonversi menjadi ikatan satu dan asam lemak menjadi jenuh
(saturated). Hampir semua asam lemak tidak jenuh tanaman terdapat dalam
bentuk konfigurasi cis sehingga depot lemak pada ternak nonruminansia semuanya dalam bentuk cis. Mikrob rumen menghasilkan isomer-isomer trans, perubahan panjang rantai, perubahan posisi ikatan rangkap, dan asam lemak rantai bercabang. Semua proses ini menghasilkan depot lemak yang unik pada ruminansia yang berbeda dari lemak pakan (Byers dan Schelling 1988).
lemak tidak jenuh C18 dihidrolisis melalui proses lipolisis kemudian dihidrogenasi oleh bakteri yang berbeda. Produk akhir dari asam lemak ikatan rangkap ganda C18 adalah asam stearat. Meskipun banyak posisi dan isomer-isomer dari monoenoik dan dieonoik, asam lemak berakumulasi dalam rumen khususnya jika rumen kelebihan lemak. Di antara itu jumlah trans-vaccenic acid (C18:1 n=7) sangat penting. Hidrogenasi PUFA C20-22 masih kontroversial. Menurut Ashes et
al. (1992) dan Palmquist dan Kinsey (1994) tidak terjadi biohidrogenasi pada rumen yang ditambahkan minyak ikan dengan konsentrasi 5 mg ml-1 in vitro,
namun Van Nevel dan Demeyer et al. (1995) melaporkan proses hidrogenasi in
vitro maupun in vivo dan Gulati et al. (1999) menunjukkan adanya
biohidrogenasi saat level minyak ikan < 2 mg ml-1 cairan rumen. Penambahan lemak menurunkan konsentrasi protozoa. Penurunan ini bergantung pada sumber lemak. Linseed oil sangat kuat menurunkan protozoa (Ikwuegbu dan Sutton 1982). Lemak lain yang kaya PUFA seperti minyak kedelai mengurangi protozoa lebih sedikit (Doreau et al. 1997). Asam linolenat memiliki daya toksisitas tinggi (Doreau et al. 1997).
Kebutuhan dan Peranan Mineral pada Ternak
Zink (Zn)
Lebih dari 100 tahun yang lalu, Zn telah diketahui penting untuk pertumbuhan jamur Aspergillus niger. Selanjutnya pada tahun 1934, Zn pertama kali diketahui sebagai nutrien esensial pada tikus. Pada tahun 1940 Keilin dan Mann mengisolasi dan memurnikan enzim karbonik anhidrase yang mengandung 0.33% Zn (McDowell 1992). Konsentrasi Zn pada kebanyakan jaringan mamalia adalah sekitar 10-100µg g-1 bobot basah (30 sampai 250µg g-1 berat kering), dengan variasi kecil untuk setiap spesies. Konsentrasi tertinggi ditemukan dalam jaringan seperti kulit, rambut, bulu, dan wool (McDowell 1992), sedangkan menurut Georgievskii (1982) konsentrasi tertinggi terdapat dalam tulang, hati, kulit, dan rambut. Di samping itu, Zn juga terdapat dalam pankreas, ginjal, kelenjar pituitari, kelenjar adrenal, testes, kelenjar asesoris kelamin jantan, dan mukosa dengan konsentrasi relatif lebih tinggi. Serapan utama Zn adalah tulang (15%), otot (45%), dan wool (27%). Penyerapan kembali dari tulang kemungkinan terjadi selama masa laktasi pada ternak (Lee et al. 1999).
Kandungan total Zn pada jaringan dan organ sapi berada di bawah kontrol mekanisme homeostasis. Kadarnya berkurang sedikit jika ransum defisien Zn. Kandungan Zn hati, pankreas, dan tulang berkurang 30 sampai 60% ketika gejala defisiensi klinis sudah mulai tampak. Konsumsi Zn melebihi 600 ppm meningkatkan kandungan Zn jaringan, yang mengindikasikan kegagalan mekanisme kontrol homeostasis pada level tinggi (Miller et al. 1988).
tinggi dibandingkan Zn susu. Konsentrasi Zn susu meningkat dengan tingginya Zn pakan dan menurun jika konsumsi Zn rendah. Kebanyakan dari Zn dalam susu sapi berikatan dengan fraksi protein dengan berat molekul tinggi (Miller et al. 1988). Kadar Zn normal susu adalah 3 sampai 5 mg l-1. Kadar Zn kolostrum mencapai 14 mg l-1 (Underwood dan Suttle 2001).
Lebih dari 200 enzim merupakan metaloenzim Zn. Peranan utama Zn dalam tubuh ternak berhubungan dengan fungsi enzim. Enzim-enzim tersebut antara lain adalah karbonat anhidrase, alkohol dehidrogenase, laktat dehidrogenase, glutamat dehidrogenase, dan alkaline fosfatase. Superoksida dismutase mengandung Cu-Zn, Zn berperan dalam perlindungan jaringan melawan peroksidase lemak. Peranan Zn juga dikaitkan dengan sintesis DNA, RNA, protein, ekspresi potensi gen, pembelahan, pertumbuhan, dan perbaikan sel. Di samping itu Zn juga berperan dalam stabilisasi membran, sistem kekebalan sel, kerja insulin, glukagon, kortikotropin, dan hormon lainnya. Kerja follicle
stimulating dan lutenizing hormones meningkat oleh Zn dan berperan dalam
keratinisasi dan kalsifikasi (Miller et al. 1988).
Little (1986) melaporkan bahwa kandungan Zn pada pakan ruminansia berkisar antara 20 dan 38 mg kg-1 bahan kering. Padahal kebutuhan Zn bagi ternak ruminansia adalah 40 ppm untuk sapi perah, 20 sampai 30 ppm untuk pertumbuhan dan finishing sapi, dan 35 sampai 50 untuk domba (NRC 1980). Pada sapi laktasi produksi tinggi (35 kg) membutuhkan 73 mg kg-1 Zn (NRC 2001). Kandungan Zn bakteri rumen 130 sampai 220 ppm, merupakan kebutuhan Zn mikrob rumen (Hungate 1966).
bulan (90.1 kg) (Adriani 2003). Suplementasi Zn(Lys)2 pada sapi dara dengan ransum limbah industri dapat memacu pertumbuhannya yaitu 1.24 kg hr-1 (Tanuwiria 2004).
Gejala awal defisiensi Zn pada sapi adalah penurunan konsumsi pakan, pertumbuhan bobot badan rendah, dan kelebihan salivasi. Efek lainnya adalah penggunaan nutrien setelah pencernaan rendah, di antaranya keseimbangan nitrogen dan sulfur rendah. Peningkatan ekskresi Zn dalam urin yang mengindikasikan penurunan penggunaan protein dan parakeratosis kulit (McDowell 1992).
Penyerapan Zn pada ternak ruminansia merupakan proses dinamis yang dipengaruhi oleh faktor pakan dan fisiologi. Persentase penyerapan meningkat apabila konsumsi Zn menurun dan akan berkurang jika konsumsi tinggi. Persentase penyerapan Zn lebih tinggi pada sapi muda dari pada yang tua, hal ini mungkin menunjukkan deposisi Zn dalam jumlah besar pada jaringan tubuh. Kemampuan penyerapan akan berkurang dengan meningkatnya umur (Miller et al. 1988). Sifat antagonis mineral Ca dan fitat serta EDTA (ethylenediamine tetraacetic acid) terhadap penggunaan Zn pada babi dan unggas, tidak menjadi masalah pada ruminansia. Kandungan Zn serum darah, rib, hati, otak, dan wool domba tidak dipengaruhi oleh level Ca pakan 1 sampai 4% bahan kering. Penyerapan Zn menurun jika Cu, Cd pakan tinggi karena Cu atau Cd dapat menggantikan ikatan Zn pada metalotionin (Church 1988). Sementara itu kasein, ekstrak hati, minyak jagung, tepung darah, EDTA, vitamin D, sitrat, pikolinat, dan asam amino (histidina, glutamina, sisteina) meningkatkan penyerapan Zn (McDowell 1992).
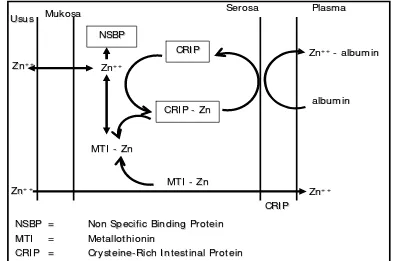
Usus halus merupakan tempat utama penyerapan dan ekskresi Zn pada ruminansia. Penyerapan Zn membutuhkan kondisi aerob. Pankreas menyekresikan suatu ligand (asam pikolinat) masuk ke dalam duodenum mengikat Zn. Ligand-Zn kompleks diangkut ke dalam sel epitel (Miller et al.
Zink dalam plasma terdiri atas 77% terikat longgar dengan albumin, 20% terikat kuat dengan α-2 makroglobulin, dan 2 sampai 8% bebas yang akan dikeluarkan melalui urin (0.5 sampai 0.8 mg hr-1) atau feses (Berdanier 1998). Penyerapan Zn dipengaruhi oleh kolekalsiferol, leukosit endogenus, prostaglandin E2. Metalotionin dalam mukosa sel berperan dalam mengontrol ketersediaan jumlah Zn untuk diangkut ke dalam sistem sirkulasi (Miller et al. 1988)
Suplementasi mineral organik terutama mineral kompleks atau khelate dengan asam amino atau peptida dapat meningkatkan penyerapan dan penggunaan Zn (Lee et al. 1999). Mineral diikat oleh karboksil dan kelompok amino dari asam amino tersebut. Mineral tersebut dipercaya dapat melindungi asam amino dari perombakan dalam rumen dan juga menyediakan mineral pada ternak dalam bentuk organik (Lee at al. 1999). Beberapa penelitian menunjukkan ketersediaan biologis Zn organik dibandingkan bentuk anorganik, termasuk ZnO dan ZnSO4 (Spears 1989; Wedekind et al. 1992). Mineral proteinat dimanfaatkan oleh ternak mengikuti jalur penyerapan peptida atau asam amino (Close 2000).
Usus
NSBP == Non Specific Binding ProteinNon Specific Binding Protein
MTI
MTI == MetallothioninMetallothionin
CRI P
CRI P == CrysteineCrysteine--Rich I ntestinal ProteinRich I ntestinal Protein
Zn
Tembaga (Cu)
Pada Tahun 1928 Cu pertama kali ditemukan sebagai mineral esensial bagi pembentukan hemoglobin tikus. Penemuan tersebut diikuti dengan kejadian bahwa Cu esensial bagi pertumbuhan dan pencegahan gangguan fisiologi dan klinis pada semua tipe ternak yang digembalakan. Pada tahun 1931, pertama kali ditemukan Cu sebagai mineral esensial bagi ternak ruminansia. Pada saat itu, ternak memperlihatkan gejala defisiensi Cu seperti anemia, diare, dan hilangnya nafsu makan, dan keadaan ternak kembali membaik setelah terapi Cu (McDowell 1992).
Konsentrasi Cu dalam jaringan hati, otak, ginjal, jantung, rambut atau wool pada hampir semua spesies adalah tinggi. Pankreas, otot kulit, dan tulang memiliki konsentrasi yang lebih rendah. Kandungan terendah terdapat dalam tiroid, pituitari, prostat dan timus. Jaringan ternak ruminansia yang dianalisis menunjukkan hati memiliki konsentrasi Cu terbesar sekitar 100 sampai 600 ppm bahan kering pada ternak dewasa normal. Kadar tersebut bisa menurun hingga di bawah 10 ppm jika defisiensi Cu, dan lebih 600 ppm jika kelebihan (Miller et al.
1988).
Tembaga dalam plasma atau darah utuh domba mendekati 100 µg dl-1 tetapi dapat meningkat sampai 165 µg dl-1 jika Cu, Mo dan S pakan tinggi (Miller
et al. 1988). Konsentrasi Cu susu sapi dan kambing adalah 0.15 mg l-1
(Underwood dan Suttle 2001).
seperti gastrin, (6) Cu-Zn superoksida dismutase (Cu-ZnSOD) berperan dalam dismutasi O2 dan H2O2, dan (7) tirosinase untuk mengubah tiroksin menjadi melanin.
Fungsi lain mineral Cu adalah sebagai komponen ikatan nukleotida adenin pada membran mitokondria, erythrocuprein yang berfungsi melindungi sel dari radikal bebas yang sangat reaktif akibat metabolisme sel, dan cerobrocuprein
yaitu protein larut air (0.33% Cu) juga sebagai superoksida dismutase (Church 1988). Tembaga esensial bagi sistem kekebalan normal ternak ruminansia (Suttle dan Jones 1986).
Mineral-mineral Fe, Mo, S, Zn, Pb dan Cd mempengaruhi kebutuhan mineral Cu, demikian pula dengan protein (McDowell 1992). Perkiraan kebutuhan Cu bergantung pada level minimum mineral antagonisnya. Level Zn dan Fe yang tinggi akan menurunkan penyerapan mineral Cu dan cenderung meningkatkan kebutuhan Cu (Underwood 1977). Kebutuhan Cu pada ternak ruminansia berkisar mulai dari 8 sampai 10 ppm (Underwood 1977). Kebutuhan Cu pada sapi perah adalah 10 ppm (NRC 1989). Kandungan Cu hijauan kurang dari 3 ppm bahan kering, sehingga sering terjadi defisiensi Cu pada ternak yang digembalakan, oleh karena itu suplementasi Cu harus dilakukan (Miller et al. 1988). Defisiensi Cu pada ternak ruminansia menghasilkan gejala yang bergantung pada spesies, umur, dan jenis kelamin serta kualitas dan lama defisiensinya.
Mineral Cu dapat diserap dalam semua segmen saluran pencernaan, walaupun tempat penyerapan utama berada di usus halus. Tembaga diserap oleh enterosit, Cu diikat baik oleh albumin atau transkuprein serta asam amino tertentu khususnya histidina. Transkuprein berkompetisi dengan albumin mengikat Cu dalam usus halus. Waktu paruh ikatan albumin dan Cu adalah 10 menit. Tembaga dikirim ke hati dan berinkorporasi ke dalam protein transpor α-globulin disebut seruloplasmin. Seruloplasmin membawa enam atom Cu dan diperkirakan 60 sampai 95% Cu plasma diangkut oleh protein ini. Namun seruloplasmin tidak hanya berguna untuk mengangkut Cu ke semua bagian tubuh tapi juga memiliki aktivitas enzim seperti ferroksidase, amida oksidase, dan superoksida dismutase (Berdanier 1998).
Ikatan metalotionin dalam mukosa usus mempunyai arti penting dalam menghambat translokasi Cu (Cousins 1985). Pengangkutan Cu dalam darah sebagian besar melalui pengikatan dengan albumin. Suplemen Cu dapat disediakan dalam bentuk Cu-sulfat, Cu-oksit, Cu-karbonat, Cu-klorit, Cu-khelat, dan Cu-proteinat. Cu-sulfat dan Cu-oksit adalah bentuk umum yang paling sering digunakan. Cu-oksit lebih tidak efektif dan tidak tersedia dari pada Cu-sulfat (Cromwell et al. 1989). Suplementasi Cu dengan ketersediaan biologis tertinggi hingga terendah adalah CuSO4, CuCO3, CuO. Ketersediaan biologis Cu-proteinat lebih besar dari Cu-sulfat pada sapi yang diberi pakan mengandung Mo (Kincaid
et al. 1986).
Toleransi spesies terhadap toksisitas Cu berbeda. Ternak ruminansia sangat sensitif terhadap toksisitas Cu, nonruminansia sangat toleran terhadap Cu. Perbedaan tersebut akibat adanya perbedaan dalam metabolisme S. Sapi toleran hingga level 100 ppm Cu, untuk domba 25 ppm (NRC 1980). Toksisitas Cu terjadi pada ruminansia, tapi tidak pada nonrumiansia. Toksisitas Cu terjadi pada kondisi penggembalaan, Cu tinggi namum rendah Mo dan S (0.1 sampai 0.2 ppm). Konsentrasi Zn pakan yang tinggi mencegah toksisitas Cu. Kandungan 100 ppm Zn bahan kering pakan mengurangi penyimpanan Cu hati (Pope 1971)
250 ppm tidak mempengaruhi asam lemak jenuh serum babi. Minyak kedelai dan minyak hewan menurunkan asam lemak jenuh serum. Lemak rantai sedang
(medium chain triglycerides=MCT) tidak mempengaruhi asam lemak jenuh
dibandingkan dengan kontrol (tanpa suplementasi lemak). Ada interaksi antara Cu dan lemak, peningkatan level Cu meningkatkan monounsaturated fatty acid
dan menurunkan polyunsaturated fatty acid dengan pemberian 5% triasil gliserol rantai sedang, dan kebalikan pada saat babi diberi pakan tanpa lemak. Penambahan Cu 125 dan 250 ppm tidak mempengaruhi monounsaturated dan
polyunsaturated fatty acid serum yang diberi minyak kedelai dan minyak hewan. Namun minyak kedelai menghasilkan monounsaturated yang paling rendah dan
polyunsaturated tertinggi di serum, dan MCT dan minyak hewan tidak
mempengaruhi monounsaturated dan polyunsaturated serum babi (Dove 1993). Profil asam lemak fosfolipid membran dipengaruhi oleh ransum (cukup dan defisien Cu). Asam lemak stearat (C18:0), docosahexaenoic acid (C22:6n-3) dan
docosadienoic acid (C22:2n-3) meningkat pada ransum yang defisien Cu, dan di sisi lain asam linolenat (C18:3n-3) dan oleat (C18:1n-9) menurun. Polyunsaturated fatty
acid menurun pada tikus yang mendapat ransum defisien Cu dibandingkan dengan ransum yang cukup Cu. Namun asam lemak dengan C22:6 tidak dipengaruhi oleh ransum (Abu Salah 1991).
Kromium (Cr)
yang dipengaruhi oleh kromium di antaranya adalah glisina, serina, dan metionina. Kromium berperan dalam kofaktor untuk insulin dalam metabolisme asam amino yang berbeda dengan insulin penggunaan glukosa (Anderson 1987).
Kebutuhan kromium pada manusia relatif lebih diketahui dibanding pada ternak. Konsentrasi kromium dalam jaringan dan organ, kecuali paru-paru, menurun dengan bertambahnya umur. Konsentrasi kromium jaringan pada bayi lebih besar dibandingkan dengan orang dewasa (Anderson 1987). Kebutuhan kromium cenderung meningkat pada kondisi stress. Kromium diserap terutama pada usus halus. Bentuk anorganik dari kromium seperti kromium klorit dan kromium oksit sangat sedikit diserap sekitar 0.5%. Rendahnya penyerapan kromium disebabkan pembentukan senyawa yang tidak larut dan adanya ikatan antara kromium bebas dengan agen kompleks dalam pakan ternak. Di samping itu penyerapan Cr dihambat oleh mineral lain seperti Zn dan Fe. Sekali Cr masuk dalam darah dan diangkut ke jaringan, akan diikat sebagai komponen dari GTF. Setelah itu dapat diakumulasi dalam jaringan pada konsentrasi relatif rendah. Kromium dikeluarkan terutama dalam urin dan sebagian pada rambut, keringat, dan empedu. Pada kondisi stress jumlah kromium yang dikeluarkan meningkat 10 sampai 300 kali (Ohh dan Lee 2005).
Kromium membentuk kompleks antara insulin dan reseptor insulin yang memasilitasi interaksi jaringan dan insulin (Mertz et al. 1974). Ruminansia menggunakan asetat dari pada glukosa sebagai sumber karbon untuk lipogenesis dan hal ini perlu sensitifitas insulin (Gardner et al. 1998). Suplementasi 1 mg. kg -1
Cr (khelate dengan asam amino) pada domba dewasa meningkatkan 30% potensi glukosa untuk digunakan dalam sintesis lemak melalui peningkatan aktivitas ATP-sitrat liase. Penelitian lain menunjukkan penambahan Cr 5 mg kg-1 (khelate dengan asam amino) bahan kering terhadap pakan alami yang mengandung 0.8 sampai 1.6 mg kg-1 bahan kering meningkatkan produksi susu (Yang et al. 1996). Chang dan Mowat (1992) melaporkan bahwa suplementasi Cr dalam bentuk GTF meningkatkan efisiensi pakan pada sapi stress.
protein terhadap pertumbuhan, konsumsi dan rasio pertumbuhan terhadap konsumsi. Kromium meningkatkan ketiga variabel tersebut pada domba yang diberi ransum tinggi protein dan menurunkan ketiga komponen tersebut pada domba yang mendapat ransum rendah protein. Pada fase kedua yaitu minggu keenam hingga 12 tidak mempengaruhi pertumbuhan, konsumsi dan rasio pertumbuhan terhadap konsumsi.
Kromium juga berperan dalam membangun sistem kekebalan tubuh dan konversi hormon tiroksin (T4) menjadi triodotironin (T3) (Burton 1995). Kromium juga berpengaruh pada pembentukan sistem kekebalan humoral (HI) maupun kekebalan yang diperantarai sel (CMI). Dalam HI suplemen kromium meningkatkan produksi antibodi atau immunoglobulin (Igs), sedangkan suplementasi kromium meningkatkan respons blastogenik (blastogenesis limfosit) terhadap immunostimulan (Spears 1999). Produksi antibodi akan meningkat akibat penurunan konsentrasi kortisol. Hormon ini berperan dalam peningkatan glukoneogenesis pada saat ternak stress. Proses glukoneogenesis akan menekan sintesis protein dalam hati sehingga sintesis antibodi juga menurun (Sohn et al. 2000).
Kebutuhan Cr pada ternak belum diketahui dengan pasti. Suplementasi mineral organik (Cr-Rhizopus sp.) dan anorganik (CrCl3.6H2O) 1, 2, 3 dan 4 ppm
in vitro menunjukkan bahwa Cr organik 1 ppm menghasilkan kecernaan bahan
organik tertinggi (34.7%), dan produksi VFA serta NH3 adalah 86 mM, dan 11.01 mM. Sedangkan suplementasi mineral Cr anorganik, kecernaan bahan organik tertinggi pada suplementasi 4 ppm (33.2%), VFA (87 mM) dan NH3 (10.13 mM). Hasil ini memperlihatkan bahwa penggunaan mineral kromium organik lebih efisien dibandingkan kromium anorganik (Jayanegara 2003).
Suplementasi kromium organik 1.9 ppm dalam ransum mengandung hidrolisat bulu ayam dapat meningkatkan produksi, laktosa, protein, dan lemak susu secara berturut-turut sebesar 17.3%, 19.0%, 27.6 dan 31.8% (Muktiani 2002).
ternak. Konsentrasi normal dalam jaringan adalah rendah yaitu kurang dari 0.1 mg kg-1 bobot segar, namun akan menjadi kelebihan jika kromium lebih dari 10 mg kg-1 (Puls 1994).
Starich dan Blincoe (1983) menyatakan bahwa kromium organik dapat diserap 20 sampai 30 kali lebih efisien dari sumber anorganik. Senyawa kromium seperti (Cr2O3) telah lama dikenal sebagai marker untuk mengetahui konsumsi pakan, kecernaan nutrien dan mineral yang dikeluarkan. Kromium ini hampir semua dikeluarkan dalam feses. Penggunaan Cr-EDTA (Cr-ethylenediamine
tetraacetic acid) sebagai marker menunjukkan khelate mineral tidak cukup
meningkatkan penyerapan dan ketersediannya (Downes dan McDonald 1964). Salah satu alasan penggunaan kromium sebagai marker dalam pakan ruminansia adalah penyerapannnya yang sangat sedikit. Namun penggunaan kromium organik mungkin berbeda karena dapat diserap dengan baik. Peningkatan konsentrasi kromium pakan dari 2.6 sampai 62.5µg kg-1 bahan kering dengan kromium pikolinat (CrP) meningkatkan kromium hati 50% tetapi tidak mempengaruhi otot domba (Olsen et al. 1996).
Selen (Se)
Selen pertama kali ditemukan dan dinamakan oleh J.J. Berzelius pada tahun 1818. Selen terdapat di alam dalam berbagai bentuk dan warna serta sering ditemukan dalam kombinasi dengan mineral Pb, Cu, Hg dan Ag. Kombinasi ini disebut selenit. Secara kimia sangat mirip dengan sulfur. Selen mempunyai tiga valensi yaitu Se2+, Se4+, dan Se6+, dan bisa berada dalam bentuk selenit dan selenat. Selen dapat bereaksi dengan sulfur dan oksigen membentuk selenmetionina, selensistina, metilselensisteina dan dimetilselenit, dan senyawa-senyawa ini mudah menguap (Berdanier 1998). Saat ini diketahui selen dan vitamin E berperan dalam detoksifikasi peroksida dan radikal-radikal bebas (Berdanier 1998).
sampai 75 ppb tetapi level tersebut hanya bertahan kurang dari 20 sampai 40 hari. Kandungan Se susu akan meningkat dari 10 sampai 37 µg l-1jika Se pakan ditingkatkan dari 47 sampai 770 ppb, akan tetapi respons ini tidak linier karena terjadi penurunan kandungan Se susu bila konsumsi Se meningkat (Miller et al.
1988). Level Se darah bervariasi bergantung pada konsumsi pakan dan kemungkinan faktor lain. Mayoritas Se plasma dan eritrosit domba berikatan dengan selenoenzim glutation peroxidase (GSH-Px).
Selen adalah komponen esensial dari GSH-Px. Glutation peroxidase
mengkatalis pengurangan berbagai macam peroksida seperti hidrogen peroksida. Vitamin E berperan dalam penekanan produksi radikal bebas (Berdanier 1998). Sedangkan GSH-Px menghancurkan peroksida sebelum sampai di membran sel. Penelitian terakhir menunjukkan bahwa vitamin E dan selen bekerja sinergis (Awad et al. 1994; Levander et al. 1995).
Beberapa enzim yang terkait dengan fungsi mineral selen adalah cytosolic glutathione peroxidase, phospholipid hydroperoxide glutathione peroxidase, gastrointestinal glutathione peroxidase, extracellular glutathione peroxidase, selenoprotein W, selenoprotein P, iodothyronine deiodinase, dan sperm capsule seleprotein. Cytosolic peroxidase, GPX1, banyak terdapat di sel darah merah, hati dan ginjal. Cytosolic peroxidase memproteksi sel darah merah atau jaringan dari serangan peroksida selama defisien selen, sehingga GPX1 ini berperan dalam penyimpanan selenoprotein (Berdanier 1998). Ternak dapat bertahan hidup tanpa gen GPX1 sehingga enzim ini tidak esensial (Cheng et al. 1997). Gastrointestinal
peroxidase (GPX2) terutama ditemukan dalam saluran pencernaan dan
melindungi mukosa usus dari hidroperoksida pakan (Chu et al. 1993; Berdanier 1998). Plasma atau extracelluler peroxidase (GPX3) terutama terdapat di ginjal, paru-paru, payudara dan plasenta. Peroksidase ini berperan sebagai antioksidan pada jaringan ekstraseluler. Phospholipid hidrogenperoxide glutathione peroxidase (GPX4) banyak terdapat dalam testes, berperan sebagai antioksidan intraseluler (Berdanier 1998). Deiodinase mengubah T4 (tetraiodothyronine) ke bentuk aktif T3 (triiodothyronine). Deiodinase berada terutama di hati dan ginjal.
Thioredoxin reductase (TRR), dalam beberapa jaringan sebagai GPX1 dan
Sistem TRR mempengaruhi aktivitas glutathion transferase (GST) walaupun GST tidak mengandung selen (Underwood dan Suttle 2001). Selenoprotein (Sel-P) memiliki potensi mengikat logam berat seperti Cd, Hg (Hill 1972) dan Pb (Rastocci et al. 1970).
Kebutuhan minimal Se pakan untuk ternak belum dapat ditentukan secara akurat. Namun kemungkinan kebutuhan Se untuk ternak mulai dari 0.05 sampai 0.3 ppm. Kebutuhan Se untuk sapi perah adalah 0.3 ppm (NRC 1989). Jumlah Se yang disimpan dalam jaringan tubuh tinggi, jika konsumsi pakan cukup atau tinggi. Oleh karena itu, untuk menentukan kebutuhan Se ternak harus memperhatikan sejarah konsumsi Se, jika konsumsi tinggi maka kebutuhan minimal harus rendah.
Penyerapan Se terutama terjadi antara duodenum dan ileum serta dikeluarkan dalam duodenum. Jumlah Se yang dikeluarkan melalui empedu kecil, rata-rata kurang dari 2% dosis Se intravena. Pada ruminansia, Se dikeluarkan terutama dalam feses, dan pada nonruminansia dalam urin. Pola penyerapan dan retensi Se pada sapi perah yang diberi Se pakan dengan level 50 sampai 260 ppb adalah linier, tetapi retensi Se pada sapi menjadi mengikuti pola terbalik jika pakan mengandung Se 300 sampai 1300 ppb (Miller et al. 1988).
Mikrob rumen dapat menggabungkan Se ke dalam seleno asam amino tetapi Se lebih terikat kuat dalam protein mikrob dalam selenometionina dibandingkan selenit atau selenat sebagai sumber Se (Miller et al. 1988). Echevarria et al. (1988) menyatakan tidak ada efek keracunan pada domba yang diberi pakan yang mengandung 9 ppm selenit selama 30 hari.
Biosintesis Nutrien Susu pada Sapi Laktasi
Bahan utama yang diserap dari darah oleh kelenjar ambing adalah, glukosa, asam amino, asam lemak, asetat, -hidroksibutirat (BHBA), dan komponen mineral. Glukosa adalah prekursor laktosa, asam sitrat, dan kebanyakan dari gliserol dari triasil gliserol yang disintesis dalam sel. Semua asam amino esensial dan beberapa asam amino nonesensial yang dibutuhkan dalam sintesis protein susu berasal dari darah. Semua purin dan pirimidin nukleotida yang dibutuhkan untuk sintesis DNA dan RNA dan laktosa serta fungsi sel lain disintesis dalam sel. Sumber energi untuk proses sintesis dalam sel adalah oksidasi dari glukosa dan asetat dalam siklus asam sitrat dan oksidasi fosforilatif.
Biosintesis lemak susu
Kandungan lemak susu menunjukkan keragaman yang besar mulai dari 1 atau 2 sampai 50%. Kandungan lemak pada susu sapi berbeda dalam satu bangsa, bergantung pakan dan lingkungan (Collier 1985).
Triasil gliserol merupakan komponen utama dari lemak susu (97 sampai 98%) sisanya adalah fosfolipid serta komponen lipid kecil lainnya. Biosintesis beberapa prekursor asam lemak terjadi dalam mitokondria khususnya nonruminansia. Biosintesis asam lemak, gliserol, dan intermediate lainnya terjadi di sitosol dan biosintesis triasil gliserol di retikulum endoplasmik. Asam lemak yang masuk ke kelenjar ambing dari darah berasal dari lemak pakan, lemak hasil sintesis hati dan lemak yang dimobilisasi dari jaringan adiposa (Collier 1985).
bahwa lebih dari 50% asam lemak susu berasal dari darah, dan sepertiga C16 dan kebanyakan asam C18. Sumber ketiga adalah asam lemak hasil sintesis dari kelenjar ambing yang berasal dari asetat dan BHBA. BHBA terutama digunakan untuk karbon empat dari asam lemak yang disintesis dalam kelenjar dan sebagian untuk asetat yang digunakan sebagai asetil Co-A untuk sintesis asam lemak. Asetat menyumbangkan semua asam lemak pendek (C4-14) dan sebagian C16 (Larson 1985).
Asam lemak rantai panjang dalam tanaman umumnya dalam bentuk asam lemak tak jenuh. Kemudian diubah oleh mikrob rumen melalui hidrogenasi ikatan rangkap menjadi asam lemak jenuh. Untuk meningkatkan kandungan asam lemak tak jenuh susu, asam lemak tak jenuh pakan tersebut dienkapsulasi (Larson 1985).
Biosintesis laktosa susu
Laktosa atau gula susu, adalah karbohidrat yang ditemukan dalam susu. Pertama kali diisolasi oleh Bartolettus 1633 dan strukturnya ditemukan sebagai 4-O- -galaktopiranosil-D-glukopiranos oleh Haworth dan Long 1927 (Ebner dan Schanbacher 1974). Laktosa disusun dari satu molekul glukosa dan satu molekul galaktosa yang bersatu dalam ikatan karbon 1-4galaktosida (Larson 1985). Kadar laktosa susu sapi adalah sekitar 4.6%, yang relatif konstan dan berkaitan dengan pengaturan tekanan osmotik susu dalam pembentukan dan proses sekresi susu (Larson 1985).
Tahapan pembentukan laktosa dan enzim yang bekerja dalam proses tersebut: Glukosa + ATP Glukosa-6-fosfat + ADP
Glukosa-6-fosfat Glukosa -1-fosfat
Uridin trifosfat + glukosa-1-P Uridindifosfat-glukosa-1+1-pirofosfat Uridin difosfat glukosa Uridin difosfat-galaktosa
Uridin-galaktosa + glukosa-1-fosfat Laktosa-1-fosfat + Uridin difosfat Laktosa-1-fosfat Laktosa + fosfat
(1) Heksokinase (2) Fosfoglukomutase
(3) Uridin difosforil glukosa pirofosforilase (1)
(2) (3) (4)
(4) Uridin difosforil galaktosa-4-epimerase
(5) Laktosa sintase (galaktosil transferase dan α-laktalbumin).
Biosintesis protein susu
Protein susu sebagian besar berupa kasein (α-kasein, -kasein, -kasein, dan χ-kasein), -laktoglobulin, dan α-laktalbumin. Selain itu juga terdapat protein-protein seperti serum albumin, immunoglobulin, enzim-enzim dan struktur fungsional lainnya yang secara kolektif menyumbangkan sistem protein kompleks dalam susu (Larson 1985).
Mekanisme sintesis protein dimulai dengan transkripsi mRNA. RNA polimerasi yang bergantung pada DNA dan mengambil prekursor-prekursor nukleotida yaitu guanosine triphosphate (GTP), adenosine triphosphate (ATP), cytidine triphosphate (CTP) dan uridine triphosphate (UTP). Proses ini menghasilkan formasi mRNA. mRNA yang dibentuk membawa pesan kode genetik. mRNA bergerak dari nukleus ke ribosom (lokasi rough endoplasmic retikulum dan cytoplasma). Proses berikutnya adalah aktivasi asam amino. Asam amino pertama diaktifkan oleh reaksi ATP dan bergabung dengan tRNA, membentuk senyawa aminoacyl-tRNA. tRNA spesifik untuk setiap asam amino dan disintesis di bawah perintah DNA. Setelah aktivasi asam amino, proses berikutnya adalah translasi yaitu ikatan polipeptida protein disintesis dalam kompleks ribosomal dalam proses translasi. Setiap trinukleotida dalam ikatan mRNA mengandung kode asam amino yang disebut kodon. Tempat dalam tRNA adalah nukleotida antikodon. Selanjutnya asam amino yang telah aktif diikat oleh tRNA yang mempunyai antikodon dan ditempatkan pada mRNA sesuai dengan kodonnya. Asam-asam amino tetap terikat pada tRNA sampai terbentuk polimerasi asam amino dan tRNA dilepas kembali ke sitoplasma (Larson 1985). Tahapan sintesis protein sebagai berikut.
1. Transkripsi DNA
2. Aktivasi asam amino
AA + ATP + tRNA AA –tRNA+AMP + PP
3. Translasi
(AA – tRNA)n AA1-AA2-AA3---AAn
Biosintesis dan Manfaat Conjugated Lenoleic Acid
Conjugated linoleic acid memiliki aktivitas anti kanker yang
ditunjukkan dalam in vitro dan ternak model. Penemuan akhir-akhir ini menunjukkan bahwa CLA dapat melokalisasi pertumbuhan dan penyebaran sel kanker payudara pada manusia. CLA dalam produk susu pada level tertentu mempunyai aktivitas anti kanker in vivo (Fritsche et al. 1997). Hunter (1998) menyatakan bahwa CLA dapat memperlambat perkembangan beberapa jenis penyakit kanker dan jantung. Meski merupakan komponen lemak, namun CLA dapat mengurangi lemak tubuh dan meningkatkan massa otot dengan lemak yang sedikit. Kandungan CLA tinggi dapat melawan cachexia, kanker, dan malaria. Di samping itu, CLA dapat menghambat perkembangan kanker kulit, dan kanker lambung pada tikus, mencegah aterosklerosis dan menormalkan toleransi glukosa yang lemah pada penderita diabetes melitus.
Conjugated linoleic acid ditemukan dalam lemak susu dan daging ternak ruminansia melalui dua sumber (Fritsche dan Steinhart 1998; Griinarii dan Bauman 1999). Pertama, CLA dibentuk melalui biohidrogenasi asam linoleat dalam rumen oleh aktivitas bakteri Butyrivibrio fibrisolvens. Kedua, CLA disintesis oleh jaringan hewan dari trans-11 C18-1 (intermediate lain dalam biohidrogenasi asam lemak tak jenuh). CLA adalah isomer asam linoleat C18:2 (9.11) dan C18:2 (10.12), yang setiap posisi memiliki ikatan ganda yaitu konfigurasi cis trans, cis cis, dan trans trans. Konfigurasi cis-9, trans-11
octadecadienoic acid (C18:2 c-9 t-11) secara biologis diduga merupakan isomer aktif. Menurut Martin dan Jenkins (2002) bahwa pH rumen di bawah 6 sangat mempengaruhi produksi trans-C18:1 dan isomer-isomer CLA oleh campuran bakteri rumen yang ditumbuhkan dalam continous culture. Konsentrasi trans C18:1 menurun pada kultur pH 5.5. Trans C18:1 merupakan prekursor sintesis CLA dalam
kelenjar ambing, maka ransum yang dapat menurunkan pH rumen dapat mengurangi ketersediaan trans-C18:1 dan konsekuensinya mengurangi sintesis CLA oleh jaringan ternak ruminansia. Oleh karena itu, untuk memaksimalkan sintesis CLA dalam jaringan ternak ruminansia membutuhkan pakan yang mampu mempertahankan pH rumen di atas enam
Secara umum, minyak tumbuhan mengandung asam linoleat tinggi dan memberikan respons terbesar dalam peningkatan kadar CLA lemak susu (Kelly et al. 1998). Minyak tumbuhan tidak biasa digunakan dalam pakan ternak ruminansia karena menghambat pertumbuhan mikrob rumen (Jenkins 1993). Suatu metode untuk mengurangi efek tersebut adalah membuat garam Ca dari asam lemak sehingga sebagian besar asam lemak luput dari perombakan oleh mikrob rumen, dan hanya sebagian yang dihidrogenasi. Konsentrasi CLA dalam lemak susu juga meningkat dengan pemberian minyak ikan atau tepung ikan, bahkan minyak ikan lebih banyak meningkatkan kadar CLA lemak susu dibandingkan dengan minyak tumbuhan dalam jumlah yang sama (Chouinard et al. 1998).
Peranan Minyak Jagung dan Minyak Ikan
Penggunaan minyak jagung dalam ransum menghasilkan gas CH4 sebesar 20.8% dan efisiensi penggunaan energi (VFA) sebesar 81%. Selanjutnya penggunaan minyak jagung relatif lebih banyak memberi keuntungan daripada kerugian (Sutardi 1997). Lemak dalam ransum akan mempengaruhi fermentasi rumen. Lemak sebagai senyawa nonpolar, tidak mudah atau segera akan larut dalam medium cairan rumen, karena itu lemak cenderung berasosiasi dengan partikel pakan dan mikrob rumen dan bentuk asosiasinya berupa penutupan permukaan secara fisik oleh lemak (Pantoja et al. 1995)
tinggi menyebabkan aktivitas metabolik protozoa menjadi terganggu dan banyak protozoa yang mati.
Minyak jagung mengandung asam linoleat yang cukup tinggi dibandingkan minyak lainnya. Di dalam rumen terjadi dua proses metabolisme pada asam linoleat. Pertama adalah linoleat kemungkinan dihidrogenasi membentuk asam asetat dan butirat. Kedua adalah linoleat dicerna oleh aktivitas bakteri anaerob Butyrivibrio fibrisolven melalui proses isomerasi asam linoleat menjadi asam linoleat terkonjugasi.
Minyak ikan merupakan produk limbah pengalengan ikan. Minyak ikan kaya asam lemak arakhidonat yang merupakan pembentuk hormon prostaglandin-E2. Selain itu minyak ikan kaya akan omega-3. Menurut Sinclair (1993), asam lemak omega-3 diduga berperan dalam produksi leukotriena (LT4) yang merupakan komponen sel darah putih dan merupakan mediator dalam sistem pembentukan kekebalan tubuh.
Minyak ikan sebagai sumber asam lemak tidak jenuh dalam ransum sapi perah mampu meningkatkan kadar omega-3 susu. Kammerlehner (1995) menyatakan bahwa asam lemak omega-3 dapat mencegah dan membantu proses penyembuhan penyakit jantung. Omega-3 merangsang pertumbuhan dan perkembangan sel otak pada anak balita dan dapat mengurangi gangguan penyakit arterosklerosis pada manusia dewasa.
Sabun Mineral
Sabun kalsium termasuk sabun yang tidak larut dalam air (Soedarmo et al. 1988). Sabun kalsium ini merupakan bentuk lemak terlindungi dan merupakan sumber lemak yang efektif dalam pakan ternak ruminansia. Menurut Jenkins dan Palmquist (1984), sabun dapat dengan mudah dicampur dengan beberapa jenis pakan, dan tidak mengganggu sistem fermentasi rumen. Peningkatan kadar kalsium pakan berasam lemak dapat menurunkan pengaruh negatif lemak dalam rumen seperti penurunan kecernaan serat. Sabun kalsium tidak bersifat toksik terhadap bakteri rumen (Palmquist et al. 1986).
Mekanisme proteksi produk sabun kalsium ini tidak berdasarkan titik cair asam lemak, tetapi berdasarkan tingkat keasaman atau pH. Sabun kalsium ini tetap utuh pada suasana keasaman netral, dan terpisah pada tingkat keasaman tinggi pH 3 (Fernandez 1999). Penelitian terakhir menunjukkan bahwa penambahan sabun kalsium asam lemak (Ca-Pufa) dalam ransum sapi menghasilkan susu 3.5 kg hr-1 dan 4 kg hr-1 FCM lebih tinggi dari pada sapi yang diberi ransum kontrol (Moallem et al. 1997).
Tinjauan Umum Literatur
Sistem pencernaan pada ternak ruminansia memiliki keunikan karena adanya sistem pencernaan fermentatif oleh mikrob rumen. Keunikan tersebut menghasilkan nutrien produk ternak ruminansia berbeda dengan ternak lainnya. Oleh karena itu, pola pemberian pakan dan nutrien harus memperhatikan sistem pencernaan dalam rumen. Salah satu nutrien yang khas pada ternak ruminansia adalah CLA. Conjugated linoleic acid memiliki efek positif pada kesehatan manusia. Asam lemak ini disintesis di rumen dan kelenjar ambing. Conjugated linoleic acid dapat ditingkatkan melalui manipulasi pakan seperti suplementasi sumber asam linoleat dalam pakan.
pascarumen, sehingga dapat meningkatkan asam lemak tak jenuh dan produktivitas pada ternak ruminansia.
MATERI DAN METODE
Penelitian dilaksanakan di Laboratorium Ilmu Nutrisi Ternak Perah, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian ini terdiri atas tiga tahap. Tahap pertama adalah pembuatan suplemen sabun kalsium, zink minyak ikan dan minyak jagung, zink organik (Zn-organik), tembaga organik (Cu-organik), kromium organik (Cr-(Cu-organik), selen organik (Se-organik) dan kedelai sangrai. Tahap kedua adalah pengujian suplememen dengan percobaan in vivo
pada domba. Tahap ketiga adalah aplikasi suplemen terbaik dengan percobaan in vivo pada sapi perah.
Percobaan in vivo pada domba dilakukan di Peternakan Domba Garut Lesan Putra Ciomas, Bogor dan percobaan in vivo pada sapi perah dilakukan di Peternakan Swadaya Sapi Perah Pondok Ranggon, Jakarta.
Pembuatan Suplemen
Sebelum membuat sabun kalsium atau sabun zink, bilangan penyabunan minyak yang digunakan ditentukan terlebih dahulu. Bilangan penyabunan digunakan untuk mengetahui KOH yang dibutuhkan untuk mengikat gugus karboksil dari asam lemak yang terdapat dalam minyak tersebut.
Bilangan penyabunan
Lima mililiter minyak dimasukkan ke dalam tabung erlenmyer 250 ml, kemudian ditambahkan 50 ml KOH 0.5 N. Erlenmeyer tersebut dihubungkan dengan refluks selama 30 menit sampai tidak ada lapisan minyak. Selanjutnya ditambahkan 1 ml fenolftalein dalam erlenmeyer kemudian dititrasi dengan HCl 0.5 N.
Bilangan Penyabunan = (Vb – Vs) x 0.02805 g/5 x BJ; Vb: volume titer blanko; Vs : volume titer sampel; Bj: berat jenis minyak.
Pembuatan sabun minyak ikan dan minyak jagung
dipanaskan dengan menggunakan hot plate. Labu erlenmeyer dihubungkan dengan pendingin tegak hingga semua minyak larut. Setelah itu ditambahkan mineral CaCl2 atau ZnCl2 sampai jenuh, sehingga larutan tersebut mengendap. Endapan dipisahkan dan ditambahkan onggok sebagai carrier dengan perbandingan 1:1 dan siap dicampurkan ke dalam konsentrat.
Pembuatan mineral organik
Enam ratus gram singkong segar dicampurkan dengan 400 ml larutan mineral (Zn, Cu, Se dan Cr) dalam plastik tahan panas. Mineral yang digunakan adalah ZnCl2, CuCl2.2H2O, CrCl3.6H2O, SeO2. Konsentrasi larutan mineral untuk mineral Zn, Se dan Cr adalah 1000 ppm, sementara untuk Cu digunakan 500 ppm. Singkong bermineral tersebut dikukus atau diautoklaf hingga matang. Setelah itu singkong diangkat dan disimpan dalam wadah plastik. Sebanyak 0.5 gram ragi tape komersial ditaburkan di atas singkong tersebut. Wadah dibungkus kertas dan diinkubasikan selama 3 hari. Pada hari ke-3 mineral organik dipanen lalu ditambahkan pollard sebagai karrier dengan rasio 2:1 dan dikeringkan dengan oven pada suhu 42ºC. Setelah kering kemudian digiling dan siap untuk digunakan dalam ransum. Khusus pada pembuatan kromium organik, sebanyak 100 ppm asam amino triptofan ditambahkan ke dalam media tumbuh yeast.
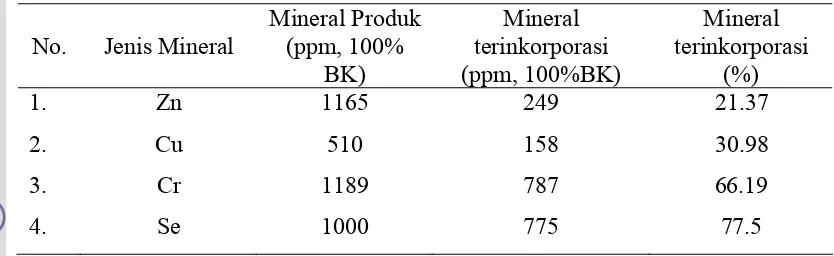
Sampel produk yang dihasilkan, diambil untuk penentuan kadar mineral yang terinkorporasi ke dalam protein yeast dengan menggunakan AAS (Tabel 1). Satu gram sampel dimasukkan ke dalam tabung dan ditambahkan 10 ml larutan TCA 20%. Tabung disentrifuse dengan kecepatan 3000 rpm selama 10 menit. Supernatan dibuang, endapan yang didapatkan ditimbang sebanyak 0.8 gram dan dimasukkan ke dalam labu destruksi lalu ditambahkan HNO3 pekat sebanyak 10 ml. Labu dipanaskan mula-mula dengan api kecil kemudian ditingkatkan dan larutan dibiarkan mendidih selama 5 menit. Setelah larutan dingin ditambah 2 ml H2SO4 pekat, 2 ml HClO4 70% dan 0.2 ml AgNO3 10% dan pemanasan kembali dilakukan sampai larutan jernih. Kadar mineral dibaca dengan menggunakan AAS sebagai mineral yang terinkorporasi dalam ragi. Suplementasi ke dalam ransum sesuai dengan kebutuhan ternak, didasarkan pada kadar tersebut.
Tabel 1. Kadar Mineral total dan mineral proteinat produk
Kacang kedelai sangrai dipanaskan dengan menggunakan kompor minyak selama 20 menit pada suhu mencapai 100°C, kemudian didinginkan dan digiling untuk dicampurkan ke dalam ransum.
Evaluasi Suplemen Minyak, Sabun Mineral, Mineral Organik dan Kedelai Sangrai pada Domba
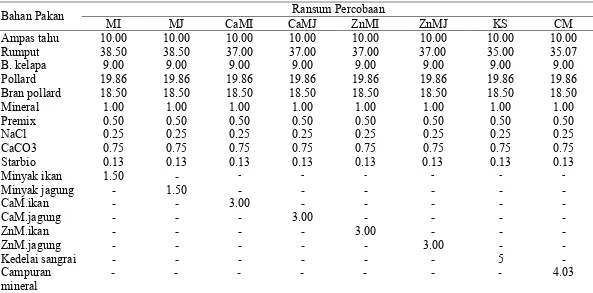
Percobaan pada domba bertujuan untuk menguji dan membandingkan suplemen minyak tanpa proteksi, minyak terproteksi dalam bentuk sabun mineral, mineral organik serta kedelai sangrai pada produktivitas ternak. Rancangan yang digunakan adalah Rancangan Acak Kelompok yang terdiri atas delapan perlakuan dan empat kelompok sebagai ulangan. Pengelompokan didasarkan pada bobot awal ternak. Percobaan ini menggunakan 32 ekor domba Priangan betina dengan bobot badan awal 22.38±3.56 kg, umur antara 6 dan 12 bulan. Pemberian ransum adalah rumput 2 kg hr-1 ekor-1, konsentrat 560 g hr-1 ekor-1, dan ampas tahu 510 g hr-1 ekor-1. Air minum diberikan ad libitum.
Perlakuan pada percobaan ini adalah: MI = Ransum basal + 1.5% Minyak ikan MJ = Ransum basal + 1.5% Minyak jagung
KS = Ransum basal + 5% Kedelai sangrai
CM = Ransum basal + Mineral Zn, Cu, Se, dan Cr organik
Komposisi bahan pakan dan nutrien ransum yang digunakan, pada Tabel 2 dan 3. Peubah yang diamati adalah konsumsi bahan kering, bahan organik, konsumsi nutrien, kecernaan nutrien dengan metode koleksi total, retensi N, pertambahan bobot badan, VFA total rumen (Sutardi 1994), NH3 rumen (Sutardi 1994), Populasi bakteri rumen (Ogimoto dan Imai 1981, dengan metode colony
count), populasi protozoa rumen (Ogimoto dan Imai 1981), kolesterol,
Ampas tahu 10.00 10.00 10.00 10.00 10.00 10.00 10.00 10.00
Rumput 38.50 38.50 37.00 37.00 37.00 37.00 35.00 35.07
B. kelapa 9.00 9.00 9.00 9.00 9.00 9.00 9.00 9.00
Pollard 19.86 19.86 19.86 19.86 19.86 19.86 19.86 19.86
Bran pollard 18.50 18.50 18.50 18.50 18.50 18.50 18.50 18.50
Mineral 1.00 1.00 1.00 1.00 1.00 1.00 1.00 1.00
Premix 0.50 0.50 0.50 0.50 0.50 0.50 0.50 0.50
NaCl 0.25 0.25 0.25 0.25 0.25 0.25 0.25 0.25
CaCO3 0.75 0.75 0.75 0.75 0.75 0.75 0.75 0.75
Starbio 0.13 0.13 0.13 0.13 0.13 0.13 0.13 0.13
Minyak ikan 1.50 - - - - - - -
Minyak jagung - 1.50 - - - - - -
CaM.ikan - - 3.00 - - - - -
CaM.jagung - - - 3.00 - - - -
ZnM.ikan - - - - 3.00 - - -
ZnM.jagung - - - 3.00 - -
Kedelai sangrai - - - - 5 -
Campuran mineral
- - - - - 4.03
Bahan BK R= Rumput; MI= Minyak ikan; MJ= Minyak jagung; CaMI= Ca-Minyak ikan; CaMJ= Ca-minyak jagung; ZnMI= Zn-minyak ikan; ZnMJ= Zn-minyak jagung; KS= Kedelai sangrai; CM= Campuran mineral (Zn, Cu, Se, Cr-organik), SK=Serat Kasar, TDN=Total digestible nutrient,
ME=Metabolizable energy
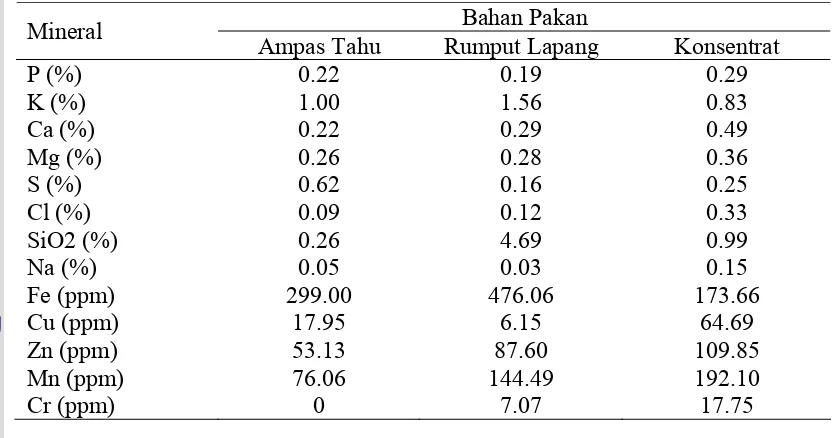
Tabel 4. Kandungan mineral bahan pakan komponen ransum domba
Bahan Pakan Mineral
Ampas Tahu Rumput Lapang Konsentrat
Suplementasi Sabun Kalsium, Mineral Organik dan Kedelai Sangrai pada Sapi Perah laktasi
Percobaan pada sapi perah laktasi ini bertujuan untuk mengetahui kombinasi ransum perlakuan dengan pemberian berbagai suplemen untuk mendukung produksi dan kualitas susu sapi. Penelitian ini dilakukan di Peternakan Swadaya Sapi Perah Pondok Ranggon Jakarta.
Percobaan menggunakan Rancangan Acak Kelompok 5 x 4. Pengelompokan dilakukan berdasarkan kepemilikan ternak. Penelitian dilakukan dalam waktu dua bulan dengan masa preliminari dua minggu. Bobot badan awal sapi perah laktasi percobaan adalah 361.4 ± 40.39 kg. Ransum basal yang digunakan disesuaikan dengan ransum yang diberikan pada peternakan tersebut. Pemberian pakan adalah konsentrat 1.4 kg hr-1 ekor-1, ampas tahu 36 kg hr-1 ekor-1 dan rumput 10 kg hr-1 ekor-1. Pakan diberikan dua kali sehari yaitu pada waktu pagi dan sore. Air minum diberikan ad libitum. Perlakuan pada sapi perah laktasi adalah:
B = Ransum Basal KS = B + Kedelai sangrai
CM = KS + Zn, Cu, Se, Cr-organik
CaMJ = CM + Sabun kalsium minyak jagung CaMI = CM + Sabun kalsium minyak ikan
Tabel 5. Komposisi bahan pakan komponen ransum sapi perah laktasi. Ransum
Bahan/Nutrien
Basal KS CM CaMJ CaMI
Rumput lapangan (%) 55 50 47.37 44.37 44.37
Ampas tahu (%) 35 35 35 35 35
Konsentrat (%) 10 10 10 10 10
Kedelai sangrai (%) - 5 5 5 5
Mineral (%) - - 2.63 2.63 2.63
CaMJ (%) - - - 3 -
CAMI (%) - - - - 3
KS= Kedelai sangrai; CM= Campuran mineral Zn, Cu, Cr dan Se organik; CaMJ= Sabun kalsium minyak jagung; CaMI= Sabun kalsium minyak ikan;
Tabel 6. Nutrien bahan pakan komponen ransum sapi perah laktasi Bahan pakan
Nutrien
Konsentrat yang diberi suplemen
B KS CM CaMJ CaMI AT RL
Bahan
kering(%) 87.39 87.55 87.36 88.30 87.74 32.45 37.37 Abu (%) 11.01 7.93 7.07 7.68 7.78 1.16 3.97 Protein (%) 10.26 12.06 14.62 10.81 13.74 8.24 2.83 Lemak (%) 5.71 4.57 2.65 6.10 5.92 3.01 0.42 Serat Kasar
(%) 11.76 9.98 11.13 8.60 9.00 7.13 11.79 TDN (%) 65.0 74.9 76.4 77.6 78.0 80.7 69.0 ME (Mkal/kg) 2.44 2.89 2.95 3.01 3.02 3.15 2.62
Zn (ppm) 56 226 280 170 253 27* 53
Cu (ppm) 6 5 28 14 24 5* 7
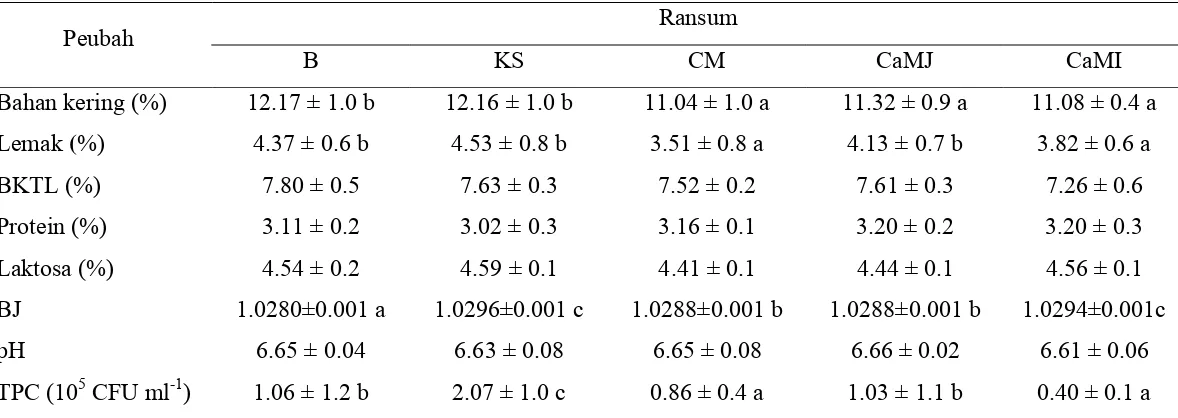
HASIL DAN PEMBAHASAN
Pertumbuhan dan Konsumsi serta Kecernaan Nutrien pada Domba yang Diberi Ransum yang Disuplementasi Minyak, Kedelai Sangrai, Sabun Mineral, dan Mineral Organik
Pertambahan bobot badan domba yang diberi ransum yang disuplementasi minyak ikan, sabun kalsium minyak ikan, sabun kalsium minyak jagung, kedelai sangrai, dan campuran mineral lebih tinggi (p<0.01) dibandingkan dengan ransum yang disuplementasi minyak jagung, sabun zink minyak ikan, dan sabun zink minyak jagung (Tabel 7). Peningkatan bobot badan tersebut disebabkan oleh peningkatan retensi N pada domba dan efisiensi penggunaan lemak.
Konsumsi bahan kering domba yang diberi ransum yang disuplementasi minyak jagung, sabun kalsium minyak jagung, dan kedelai sangrai lebih tinggi (p<0.01) dibandingkan dengan domba yang diberi ransum yang disuplementasi minyak ikan, sabun kalsium minyak ikan, sabun zink minyak ikan, sabun zink minyak jagung, dan campuran mineral. Kecernaan bahan kering ransum yang disuplementasi sabun kalsium minyak ikan lebih tinggi (p<0.01) dibandingkan ransum lainnya. Retensi N (g hr-1) domba yang diberi ransum yang disuplementasi sabun kalsium minyak ikan, kedelai sangrai, dan campuran mineral lebih tinggi (p<0.01) dibandingkan dengan domba yang diberi ransum yang disuplementasi minyak ikan, minyak jagung, sabun kalsium minyak jagung, sabun zink minyak ikan, dan sabun zink minyak jagung. Total digestible nutrient
Ransum Peubah
MI MJ CaMI CaMJ ZnMI ZnMJ KS CM
Pbb (g hr-1) 99 ± 20 c 86 ± 16 b 114 ± 35 c 103 ± 30 c 88 ± 41 b 53 ± 37 a 105 ± 25 c 103 ± 27 c Konsumsi BK
(g hr-1) 820±120a 903 ± 56 b 856±105 a 947±25 b 847±110 a 785±57 a 933±42 b 805±85 a Kecernaan
BK (%) 68 ± 2.4 a 70 ± 0.8 a 76 ± 5.6 b 68 ± 3.7 a 60 ± 4.7 a 65 ± 7.0 a 68 ± 3.0 a 68 ± 4.9 a Retensi N
(g hr-1) 10± 0.5 a 10.0±1.5 a 11.8± 1.0b 9.8 ± 0.8 a 9.0 ± 0.9 a 9.0 ± 0.4 a 12.2 ±1.2b 13.2±1.3 b Retensi N/N
konsumsi (%) 42± 1.4 a 45 ± 7.4 b 48 ± 5.7 b 39 ± 8.3 a 35 ± 10.7 a 46 ± 5.6 b 46 ± 6.1 b 55 ± 4.1 b Retensi N/N
tercerna (%) 64± 7.9 b 58 ± 6.2 a 61 ± 5.6 a 56±10.3 a 60 ± 8.0 a 68 ± 3.9 c 64 ± 8.0 b 76 ± 2.9 d TDN (%) 70 ± 4.0 b 70 ± 0.8 b 75 ± 4.5 b 65 ± 3.7 a 69 ± 4.8 a 65 ± 6.3 a 67 ± 2.5 a 66 ± 5.0 a
yang diberi ransum yang disuplementasi sabun zink (108.5 g hr vs 70 g hr ). Pengaruh suplementasi minyak ikan dalam bentuk sabun kalsium tidak berbeda dengan minyak ikan pada pertambahan bobot badan dan konsumsi bahan kering. Asam lemak rantai panjang (> C20 ) pada minyak ikan kemungkinan masih ada yang lolos dari degradasi rumen sehingga asam lemak tersebut dapat dimanfaatkan oleh induk semang. Minyak ikan kaya akan asam lemak arakhidonat sebagai prekursor prostaglandin yang merupakan substan seperti hormon yang berperan dalam penyerapan nutrien. Kenyataan ini juga dapat dilihat pada TDN ransum yang disuplementasi minyak ikan dan sabun kalsium minyak ikan tidak berbeda (70 vs 76%). Penggunaan 3% minyak ikan tuna yang diproteksi (rumen protected tuna oil) dan minyak hewan (tallow) 3% dalam ransum domba menghasilkan pertambahan bobot badan yang tidak berbeda di antara keduanya yaitu 196 g hr-1 dan 218 g hr-1 (Kitessa et al. 2001).
Suplementasi kacang kedelai sangrai menghasilkan pertambahan bobot badan domba cukup tinggi (105 g hr-1), sama dengan ransum yang diberi sabun kalsium minyak ikan dan kalsium minyak jagung. Pertambahan bobot badan yang tinggi terjadi karena kacang kedelai sangrai menyediakan protein dan energi sekaligus, sehingga responsnya bisa menyamai ransum yang disuplementasi minyak ikan dan kalsium minyak jagung yang merupakan sumber energi. Peningkatan konsumsi bahan kering domba yang diberi kedelai sangrai terjadi karena suplemen tersebut dapat memperbaiki palatabilitas.
Pertambahan bobot badan domba yang diberi ransum yang disuplementasi mineral organik (Zn, Cu, Cr dan Se-organik) mencapai 103 g hr-1. Suplementasi mineral organik pada penelitian ini dapat meningkatkan produktivitas domba. Peningkatan pertumbuhan tersebut terjadi karena mineral yang disuplementasi merupakan mineral yang mempunyai peranan yang sangat besar dalam pertumbuhan. Mineral Zn berperan dalam pertumbuhan sel. Mineral Zn merupakan bagian dari enzim yang terkait dengan pertumbuhan. Mineral Cr adalah bagian dari GTF yang berfungsi membantu kinerja insulin untuk memasok glukosa dalam sel. Oleh karena itu mineral tersebut mampu meningkatkan pertumbuhan ternak. Di samping itu Cr juga berperan dalam metabolisme protein khususnya inkorporasi asam amino. Mineral Se juga berperan sangat besar dalam pertumbuhan karena mampu mengubah hormon T4 menjadi T3 yang memacu metabolisme umum sehingga produktivitas ternak meningkat dengan suplementasi Se. Langlands et al. (1991) melaporkan suplementasi Se dengan konsentrasi 60 ηmol l-1 sampai 400 ηmol l-1 darah meningkatkan pertumbuhan wool, diameter serat, dan pertambahan bobot badan domba.
Menurut Rojas et al. (1995) ketersediaan Zn-lys lebih besar sebagai sumber mineral Zn dibanding lainnya, hal ini berdasarkan akumulasi Zn di dalam hati, ginjal dan pankreas domba lebih besar. Domba yang mendapat ransum limbah agroindustri dan ransum standar yang disuplementasi dengan Zn, Cu lisinat serta Zn, Cu proteinat menghasilkan pertumbuhan sebesar 46 sampai 157 g hr-1. Pertumbuhan domba tertinggi dihasilkan dari ransum hasil samping yang disuplementasi dengan Zn, Cu proteinat (Irawan 2002).
Konsumsi pakan merupakan respons dari interaksi beberapa faktor yaitu penginderaan, gastrointestinal dan sistem syaraf. Penelitian ini menunjukan bahwa konsumsi ransum yang mendapat suplementasi minyak baik yang diproteksi dalam bentuk kalsium minyak ikan atau kalsium minyak jagung maupun tanpa proteksi tidak berbeda. Konsumsi yang lebih rendah pada minyak ikan dibandingkan dengan kalsium minyak jagung kemungkinan disebabkan oleh bau amis minyak ikan. Kitessa et al. (2001) melaporkan bahwa pemberian 3% minyak ikan yang diproteksi (PTO = protected tuna oil) dan minyak ikan tanpa proteksi (UTO = unprotected tuna oil ) mempengaruhi konsumsi bahan kering (847 g hr-1 vs 736 g hr-1) tetapi tidak mempengaruhi konsumsi lemak domba (59.3 g hr-1 vs 52.3 g hr-1).
Konsumsi bahan kering domba yang diberi ransum yang disuplementasi kedelai sangrai paling tinggi di antara semua perlakuan (933 g hr-1). Hal ini disebabkan aroma dan rasa yang mungkin disukai oleh ternak. Pada ternak ruminansia indera penciuman memegang peranan penting dalam konsumsi pakan. Konsumsi bahan kering domba yang diberi ransum dengan suplementasi campuran mineral hanya mencapai 805 g hr-1. Rendahnya konsumsi bahan kering ini disebabkan suplemen tersebut merupakan hasil biofermentasi sehingga menghasilkan mineral proteinat (yeast). Produk tersebut memproduksi protein
yeast sehingga kadar protein pakan juga meningkat. Protein pakan yang tinggi mengakibatkan konsumsi menjadi menurun karena SDA=specific dynamic action
yang disuplementasi dengan Zn, Cu proteinat, Zn-(Lys)2 +Cu-(Lys)2 lebih rendah dari Zn-(Lys)2 (584 g hr-1, 506 g hr-1 Vs 658 g hr-1).
Kecernaan bahan kering yang tinggi pada sabun kalsium minyak ikan dapat disebabkan oleh konsumsi yang rendah sehingga laju aliran pakan dalam saluran pencernaan lebih lambat. Proteksi minyak ikan dapat memasok asam lemak arakhidonat sebagai prekursor prostaglandin. Bahan itu berfungsi dalam penyerapan nutrien di usus sehingga meningkatkan kecernaan.
lemak tinggi dalam ransum dapat digunakan dan tidak mengganggu sistem fermentabilitas dalam rumen jika lemak tersebut diproteksi.
Retensi N pada percobaan ini berkisar mulai dari 8.99 sampai 13.12 g hr-1. Kalsium minyak ikan menghasilkan retensi N tinggi (11.78 g hr-1) karena minyak ikan kemungkinan masih mengandung protein. Pembuatan CaMI menggunakan KOH dalam alkohol yang disertai pemanasan, sehingga diduga protein yang masih terdapat dalam minyak ikan tersebut mengalami perubahan struktur protein dan dapat lolos dari degradasi mikrob rumen. Hal tersebut menyebabkan pasokan nitrogen meningkat sehingga retensi N juga meningkat.
Beberapa metode yang dapat menurunkan laju dan tingkat degradasi rumen terhadap protein pakan yaitu pemanasan, kimia, atau kombinasi kimia dan pemanasan (Kaufmann dan Lupping 1982; Satter 1986; Broderick et al. 1991; Schwab 1995). Perlakuan kimia pada protein pakan dapat dibagi ke dalam tiga kategori yaitu kimia yang mengombinasikan dengan ikatan protein seperti aldehid; proses kimia yang merubah struktur protein melalui denaturasi seperti asam, alkali dan etanol; proses kimia yang mengikat protein tetapi sedikit atau tidak mengubah struktur protein seperti tanin (Broderick et al. 1991; Schwab 1995).