KUANTlFlKASl METABOLISME BENlH PADA PERIODE I,
PERIODE KONSERVASI
DAN
AWAL SIMPAN PERIODE II
UNTUK DETEKSI VIGOR GENETIK JAGUNG
Oleh
FAlZA
CHAlRANl SUWARNO
AGR.
89511PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
SUMMARY
FAIZA CHAIRANI SUWARNO. Quantification of Seed Metabo-
lism During Periode I, Conservation Periode and Early
Storage of Periode I1 for Detecting Genetic Vigor in Corn
(Under supervision of SJAMSOE' OED SADJAD as the leader,
MAGGY T. SUHARTONO, MOH. ANWAR NUR, AHW4D ANSORI MATTJIK,
and ISWANDI ANAS as the committee members).
The study consisted of three experiments, i.e. (1)
determination of initial seed vigor (Va)
,
which was afactorial experiment with variety and phosphate fertili-
zer as the factors; (2) determination of conservation vi-
gor before storage (vKSS) of seed lots harvested from ex-
periment (1) ; and (3) determination of initial vigor be- fore storage (vaS) of seed lots used in experiments (1)
and (2).
The experiments were aimed to detect the genetic vi-
gor of corn through quantification of seed metabolism ba-
sed on absolute viability at the morphological maturity
(MM) periode up to early storage periode.
Seed vigor reached the highest value at the physio-
logical maturity (MF) indicated by maximum dry weight and
the appearance of black layer. The maximum seed vigor
was significantly determined by the seed vigor at the MM
based on the delta values (D), and the duration of ma-
turing periode between MM and MF ( R ~ = 63.6%*)
.
The high-er seed vigor at the MM and the longer duration of ma-
turing periode between MM and MF, the higher Va of the
seed.
The D lines derived from the Ptotal and Pphytate re-
present a quadratic function, which is in agreement with
the viability concept, and they can be used for deter-
mining the morphological maturity. According to the
Steinbauer concept, MM is characterized by the maximum D l
when the germination speed is minimum while the germina-
tion capacity is at maximum level. Steinbauer-Sadjad
concept stated that at the stage, the potensial viabili-
ty (V P ) is reaching the maximum value and the vigor (Vg) is just starting to increase.
It is important to consider the seed coat develop-
ment for determining the MM stage technologically. In
corn, seed coat which developed from suberized outer la-
yer of nucellus is semipermeable, whereas the seed coat
developed from the integuments is desintegrated. This
process takes 1-2 weeks after seeds embryo axis complete-
ly formed in 3 weeks after anthesis, if the technological
considered, assuming that the energy is needed for the
development of suberin layer. After the process is com-
pleted, the V reaches maximum value and the V is star-
P
gting to increase, that indicating MM.
The seed vigor before storage periode Was) which
is one of the important factors determining the seed abi-
lity to maintain its vigor during storage periode, was
closely correlated to the seed vigor at MM based on D va-
**
lues (r = 72.9% )
.
The higher seed vigor at MM, thehigher its vas.
Genetic vigor of corn reflected by the yield of the S
varieties could be detected by the Vat VKS
,
and vaS.The coefficients of Mallow (Cp) and determination (R2)
which were respectively 2.8 and 59.9% (highly
significant), resulted from the regression analysis indi-
cated that the genetic vigor could be well detected by Va
and vaS. The genetic vigor could also be better detected
by Vat
vaS
and vKSS based on either VKS or inversed vigorarea (BV-I) with R~ 61.1% significantly and Cp 4.0, R2
61.5% significantly and Cp 4.0 respectively.
Those experiments indicated that genetic vigor could
be detected through the quantification of seed metabo-
lism. For further studies more viability parameters at
and critical periode have to be considered besides more
genotypes should also be used to increase the genetic va-
RINGKASAN
FAIZA CEAIRANI SWARNO. Kuantifikasi Metabolisme Benih
pada Periode I, Periode Konservasi dan Awal Simpan Perio-
de I1 Untuk Deteksi Vigor Genetik Jagung (Dibawah bim-
bingan SJAMSOEcOED SADJAD sebagai ketua, MAGGY T.
SOHARTONO, MOB. ANWAR NUR, AHMAD ANSORI MATTJIK, dan
ISWANDI ANAS masing-masing sebagai anggota).
Penelitian terdiri dari 3 percobaan, yaitu (1) pe-
nentuan vigor awal benih (Val yang merupakan suatu perco-
baan faktorial dengan menggunakan faktor varietas dan
faktor pemupukan fosfat; (2) penentuan vigor konservasi
sebelum simpan (vKSS) lot-lot benih yang diperoleh dari
percobaan (1) ; dan ( 3 ) penentuan vigor awal sebelum sim-
pan (vas) lot-lot benih yang digunakan pada percobaan (1)
dan (2).
Tujuan penelitian adalah mempelajari kemungkinan
mendeteksi vigor genetik jagung melalui kuantifikasi me-
tabolisme benih berdasarkan penilaian viabilitas absolut
mulai dari saat matang morfologi (MM) sampai dengan awal
periode simpan.
Vigor benih mencapai maksimum pada saat masak fisio-
logi (MF) yang ditandai oleh bobot kering benih maksimum dan munculnya b l a c k l a y e r . Status vigor benih pada saat
benih mengakumulasi bobot kering, nyata ditentukan oleh
status vigor benih pada saat MM berdasarkan tolok ukur
nilai delta (D) dan lamanya waktu pemasakan benih antara
*
MM-MF ( R ~ = 63 - 6 % )
.
Semakin tinggi vigor benih pada sa-at MM dan semakin lama waktu pemasakan benih antara MM-
MF, semakin tinggi pula Va benih tersebut.
Garis-garis nilai D yang diperoleh dari tolok ukur
Ptotal dan Pfitat pada umumnya menunjukkan fungsi kuadra-
tik yang memenuhi kaidah viabilitas benih, dan dapat di-
gunakan untuk menentukan saat matang morfologi. Dalam
konsepsi Steinbauer, MM dicirikan oleh nilai D maksimum
karena pada saat itu vigor (Vg) masih minimum sedangkan
viabilitas potensial (Vp) maksimum. Dalam konsepsi
Steinbauer-Sadjad pada saat itu Vp sudah mendekati maksi-
mum sedangkan V baru nyata membesar. g
Untuk mencapai stadia itu perkembangan kulit benih
perlu diamati dan dijadikan ciri MM dipandang dari segi
teknologi benih. Kulit benih jagung pada hakekatnya me-
rupakan lapisan luar nucellus yang bergabus (suberized)
bersifat semipermeabel sedangkan kulit benih yang berasal
dari integumen mengalami desintegrasi. Proses ini memer-
lukan waktu 1-2 minggu sesudah poros embrio terbentuk
sempurna 3 minggu setelah antesis, bila saat MM didasar-
tukan lapisan suberin sebagai struktur benih diperlukan
energi. Setelah itu baru mulai ada Vg, bersamaan dengan
saat V mencapai maksimal, yang mengindikasikan saat MM.
P
Vigor benih pada awal periode simpan (vaS) yang me-
rupakan salah satu parameter penting yang menentukan ke-
mampuan benih mempertahankan vigornya selama di periode
simpan ternyata berkorelasi sangat erat dengan vigor be-
nih pada saat MM berdasarkan nilai D (r = 72.9%**)
.
Se- makin tinggi vigor benih pada saat MM, semakin tinggivaSnya
.
Vigor genetik jagung dengan tolok ukur produksi va-
rietasnya dapat dideteksi melalui Val
vKSS
dan vaS. Ber-dasarkan nilai koefisien Mallow (Cp) dan koefisien deter-
minasi (R2) dari persamaan regresi yang diperoleh, vigor
genetik dapat dideteksi melalui parameter Va dan
vaS,
de-ngan nilai R2 59.9% yang sangat nyata dan C p 2.8. Vigor
genetik dapat dideteksi lebih baik lagi bila didasarkan
pada parameter Val
vas
dan VKS dengan tolok ukur VKSatau kebalikan luas bidang vigor (BV-'1
,
dengan koefisiendeterminasi dan koef isien Mallow masing-masing R2 61.1%
yang nyata dan Cp 4.0, serta R~ 61.5% yang nyata dan Cp
Penelitian ini menunjukkan bahwa vigor genetik dapat
penelitian selanjutnya perlu ditambahkan parameter-
parameter viabilitas pada periode simpan, periode
konservasi sebelum tanam dan periode kritikal, disamping
penggunaan varietas yang lebih banyak agar diperoleh
KUANTlFlKASl METABOLISME BENIH PADA PERIODE I, PERIODE KONSERVASI DAN AWAL SIMPAN PERIODE II
UNTUK DETEKSI VIGOR GENETIK JAGUNG
Oleh
:
FAlZA CHAlRANl SUWARNO AGR. 8951 1
Disertasi sebagai salah satu syarat untuk memperoleh gelar Doktor dalam ilmu-ilmu pertanian
pada
Program Pascasarjana lnstitut Pertanian Bogor
PROGRAM PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Disertasi : KUANTIFIKASI METABOLISME BENIH PADA PERIODE I, PERIODE KONSERVASI DAN AWAL SIMPAN PERIODE I1 UNTUK DETEKSI
VIGOR
GENETIK JAGUNGNama Mahasiswa : Faiza Chairani Suwarno Nomor Pokok : 895 1 1
Meny etuj ui 1. Komisi Pembimbing
Prof. Dr Ir Sjarnsoe'oed Sadjad, MA. Ketua
.
Moh. Anwar NurAnggota AIQXota
Dr Ir H. Ahmad Ansori Mattjik
\
2. Ketua Program Studi Agronomi
Prof. Dr Ir H. A. Surkati Abidin
Tanggal lulus :
2.1
-F
EB
1995
Dr Ir Iswandi Anas Anggota
Pascasarj a
a
RIWAYAT HIDUP
Penulis dilahirkan di Bogor pada tanggal 8 Oktober
1952, dari Bapak Almarhum H. Chairani Achmad dan Ibu
Almarhumah Hj. Masturah.
Pada tahun 1971 penulis lulus dari SMA Negeri I1 Bo- gor. Tahun 1976 lulus dari Jurusan Agronomi Fakultas Pertanian IPB. Tahun 1989 memperoleh gelar Magister
Sains pada Program Studi Agronomi Fakul tas Pascasarj ana
IPB.
Penulis bekerja sebagai research assistant di Bio-
trop pada tahun 1977-1978. Sejak tahun 1981 hingga seka-
rang bekerja sebagai staf pengajar pada Fakultas Pertani-
KATA PENGANTAR
Pu j i dan syukur penulis pan j atkan kehadirat Allah
SWT, karena hanya atas perkenan-Nyalah penulis dapat
menyelesaikan disertasi ini.
Pada kesempatan ini penulis menyampaikan terima ka-
sih yang sebesar-besarnya kepada :
1. Rektor IPB dan Direktur Program Pascasarjana yang te-
lah memberikan kesempatan kepada penulis untuk mengi-
kuti program S3 di Institut Pertanian Bogor; serta
Team Managemen Program Doktor (TMPD) atas beasiswa
yang diberikan.
2. Bapak Prof. Dr Ir Sjamsoe'oed Sadjad, MA. sebagai ke-
tua komisi pembimbing atas pengarahan dan bimbingannya
mulai dari perencanaan penelitian sampai dengan sele-
sainya penulisan disertasi ini.
3. Ibu Dr Ir Maggy T. Suhartono, Bapak Dr Ir H. Moh. Anwar
Nur, Bapak Dr Ir H. Ahmad Ansori Mattjik dan Bapak Dr
Ir Iswandi Anas masing-masing sebagai anggota komisi
pembimbing, atas saran dan koreksi yang telah diberi-
kan mulai dari perencanaan penelitian sampai dengan
penulisan disertasi ini.
4. Bapak Dr Subandi dari Kelti Genetika dan Pemuliaan
5. Ketua Kelti Biokimia Balittan Bogor yang telah memban-
tu dalam analisis asam fitat, dan Ibu Sutriani di La-
boratorium Ilmu Tanah IPB atas bantuannya dalam anali-
sis fosfor.
6. Rekan-rekan staf pengajar dan karyawan/karyawati Labo-
ratorium Ilmu dan Teknologi Benih IPB atas segala ban-
tuannya
.
7. Almarhum Ayah dan almarhumah Ibu tercinta, suami dan
anak-anak tersayang (Dr Ir Suwarno, MS., Willy dan
Jimmy), serta seluruh keluarga, atas dorongan moril
serta doa yang tiada henti-hentinya dan juga atas pe-
ngertian dan bantuan yang diberikan hingga terwujudnya
disertasi ini.
8. Semua pihak yarig tidak dapat disebutkan satu per satu,
Semoga Allah SWT mernberikan rahmat-Nya kepada semua
yang telah membantu penulis hingga selesainya disertasi
ini
.
Bogor, Februari 1995
DAFTAR IS1
Halaman
DAFTAR IS1
. . .
iv DAFTAR TABEL. . .
viDAFTAR GAMBAR
. . .
xiDAFTAR SIMBOL
. . .
xiiiGLOSARI
. . .
xvPENDAKULUAN
Latar Belakang
...
1Tujuan
. . .
4 Hipotesis. . .
4 TINJAUAN PUSTAKAKuantifikasi Metabolisme Benih
. . .
6 Vigor Genetik. . .
11Akumulasi dan Pengaruh Fosfat
Terhadap Viabilitas Benih
. . .
19Devigorasi Benih
. . .
26 BAHAN DAN METODETempat dan Waktu
. . .
35 Bahan dan Alat. . .
35Metode Penelitian
. . .
37Pelaksanaan Penelitian
. . .
41HASIL DAN PEMBAHASAN
I
.
Vigor Awal (Va). . .
52 I1.
Garis Nilai Delta. . .
60111
.
Vigor Konservasi Sebelum Simpan (vKSS)...
64IV
.
Vigor Awal Sebelum Simpan (Va)...
66V
.
Vigor Genetik (Vg,, 1. . .
69KESIMPULAN
...
77SARAN
. . .
78 DAFTAR PUSTAKA...
79DAFTAR TABEL
Nomor Halaman
Teks
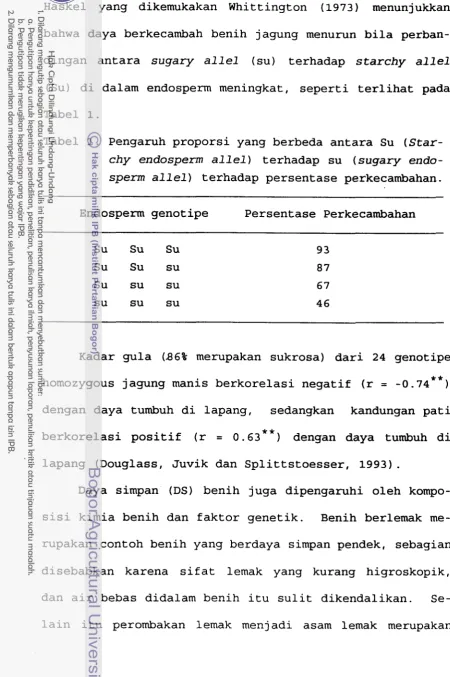
1. Pengaruh proporsi yang berbeda antara Su (starchy endosperm a1 1 el ) terhadap su (sugary endosperm allel) terhadap persentase perkecambahan. 16
2. Nilai
vKST
galur - galur dan varietas kedelai(Kartika dalam Sadjad, 1993) 32
3. Produksi, bobot kering benih dan jumlah benih/ tongkol varietas Genj ah Kertas, Arjuna,
Harapan Baru dan Hawai Super Sweet 52
4 . Va benih, kandungan Photal dan Pf ita6 benih
saat MM, vigor ber asarkan ni a1 MPVMM,
dan waktu antara MM
-
MF 555. Persamaan regresi antara bobot kering benih (mg/benih) (Y) dengan waktu (rninggu) (XI,
kondisi optimum dan sub optimum. 57
6. Persamaan regresi nilai delta (P ) (ug/benih) (Y) terhadap waktu (rninggu)
(RPP
pada perla- kuan kondisi optimum, sub optimum dan rata-ratanya 61
7. Nilai VKS, luas bidang vigor (BV) dan luas
BV-I 64
8. Vigor awal sebelum simpan (vaS) dengan tolok
ukur kebal ikan Daya Hantar Listrik (DHL-l) 67
9. Koefisien determinasi dan koefisien Mallow dari semua kemungkinan persamaan re resi antara V
gen dengan Val VKS
, dan Va
8
10. Koefisien determinasi dan koefisien Mallowdari semua kemungkinan per amaan regresi
antara V gen dengan Val BV-' dan
vaS
7 0 11. Persamaan regresi ntara VNomor
Lam~iran
1. Data Hasil Analisis Tanah.
2. Deskripsi Varietas Jagung
3. Persamaan regresi antara nilai DTZ (Y) dan waktu deraan pada varietas Genjah Kertas
(Vl) , Arjuna (V21, Harapan (V3) dan Hawai Super Sweet (V4) pada kondisi optimum (PI) dan sub optimum (PO). 4. Nilai intersep DTZl ZPO dan VKS
5. Sidik ragam pengaruh faktor-faktor varietas (V) dan pemupukan (P) serta interaksinya terhadap produksi (g/pot )
.
6. Sidik ragam pengaruh faktor-faktor varietas (V) dan pemupukan (P) serta interaksinya terhadap bobot benih (g/1000 butir)
.
7. Sidik ragam pengaruh faktor-faktor varietas (V) dan pemupukan (P) serta interaksinya terhadap jumlah benih/tongkol.
Hal aman
8. Sidik ragam pengaruh faktor-faktor varietas (V) dan pemupukan (P) serta interaksinya
terhadap kecepatan tumbuh benih (%/etmal)
.
92 9. Sidik ragam pengaruh faktor-faktor varietas(V) dan pemupukan (P) serta interaksinya terhadap P total benih (ppm) saat MM
(berdasarkan sempurnanya embryo). 93 10. Sidik ragam pengaruh faktor-faktor varietas
(V) dan pemupukan (P) serta interaksinya terhadap asam fitat benih (ppm) saat MM
(berdasarkan sempurnanya embryo). 93 11. Sidik ragam pengaruh faktor-faktor varietas
(V) dan pemupukan (P) serta interaksinya
terhadap vigor berdasarkan nilai DMPVMM. 94 12. Sidik ragam pengaruh faktor-faktor varietas
(V) dan pemupukan (P) serta interaksinya terhadap bobot kering benih (g/tongkol)
.
13. Sidik ragam pengaruh faktor-faktor varietasviii
14. Sidik ragam pengaruh faktor-faktor varietas (V) dan pemupukan (PI serta interaksinya terhadap luas bidang vigor (BV).
15. Sidik ragam pengaruh faktor-faktor varietas (V) dan pemupukan (P) serta interaksinya terhadap daya hantar listrik air rendaman benih (DHL).
16. Sidik ragam regresi bobot kering benih (Y) terhadap waktu (XI) dan kuadrat waktu (X2) untuk varietas Genjah Kertas pada kondisi
optimum. 96
17. Sidik ragam regresi bobot kering benih (Y) terhadap waktu (XI) dan kuadrat waktu (XZ) untuk varietas Arjuna pada kondisi optimum. 18. Sidik ragam regresi bobot kering benih (Y)
terhadap waktu (XI) dan kuadrat waktu (X2) untuk varietas Harapan Baru pada kondisi optimum
.
1 9 . Sidik ragam regresi bobot kering benih (Y) terhadap waktu (Xl) dan kuadrat waktu (X2) untuk varietas Hawai Super Sweet pada
kondisi optimum. 98
20. Sidik ragam regresi bobot kering benih (Y) terhadap waktu (Xl) dan kuadrat waktu (X2) untuk varietas Genjah Kertas pada kondisi
sub optimum. 98
21. Sidik ragam regresi bobot kering benih (Y) terhadap waktu (XI) dan kuadrat waktu (X2) untuk varietas Arjuna pada kondisi sub
optimum. 99
22. Sidik ragam regresi bobot kering benih (Y) terhadap waktu (Xl) dan kuadrat waktu (X2) untuk varietas Harapan Baru pada kondisi
sub optimum. 99
23. Sidik ragam regresi bobot kering benih (Y) terhadap waktu (XI) dan kuadrat waktu (X2) untuk varietas Hawai Super Sweet pada
kondisi sub optimum. 100
24. Sidik ragam regresi bobot kering benih (Y) ter- hadap vigor benih berdasarkan nilai DMPVMM
(X1) dan lamanya MM-MF berdasarkan puncak
Sidik ragam regresi nilai D (P = Ptot- 'fi at ) (Y) terhadap waktu ff?) dan
kuahrat waktu (X2) untuk varietas Genjah Kertas pada kondisi optimum.
Sidik ragam regresi nilai D (P = Ptot- Pfi at) (Y) terhadap waktu fa?) dan kuahrat waktu (X2) untuk varietas Arjuna pada kondisi optimum.
Sidik ragam regresi nilai D (P = Ptot- Pfi at) (Y) terhadap waktu ff?) dan
kuagrat waktu (X2) untuk varietas Harapan Baru pada kondisi optimum.
Sidik ragam regresi nilai D (P = Ptot- Pfi at) (Y) terhadap waktu fg?) dan kuasrat waktu (X2) untuk varietas Hawai Super Sweet pada kondisi optimum.
Sidik ragam regresi nilai D (P = Ptot- Pfi at) (Y) terhadap waktu
fa?)
dankuahrat waktu (X2) untuk varietas Genjah Kertas pada kondisi sub optimum.
Sidik ragam regresi nilai D (P = Ptot- Pfi at) (Y) terhadap waktu ($?f dan
kuahrat waktu (X2) untuk varietas Arjuna pada kondisi sub optimum.
Sidik ragam regresi nilai D (P = Ptot- 'fi at ) (Y) terhadap waktu (%?f dan
kuahrat waktu (X2 ) untuk varietas Harapan
Baru pada kondisi sub optimum.
Sidik ragam regresi nilai D (P = Ptot- Pfi at) (Y) terhadap waktu fa?) dan kuahrat waktu (X2) untuk varietas Hawai Super Sweet pada kondisi sub optimum.
Sidik ragam regresi nilai D (P = Ptot- 'fi at (Y) terhadap waktu (by? dan
kuahrat waktu (X2) untuk varietas Genjah Kertas pada rata-rata kondisi optimum dan sub optimum.
Sidik ragam regresi nilai D (P = Ptot- Pfi at) (Y) terhadap waktu (RPP dan
35. Sidik ragam regresi nilai D (P = Ptot- Pfi at) (Y) terhadap waktu 799) dan
kuahrat waktu (X2) untuk varietas Harapan Baru pada rata-rata kondisi optimum dan sub optimum.
36. Sidik ragam regresi nilai D (P = Ptot- 'fi at ) (Y) terhadap waktu B f ) dan
kuahrat waktu (X2) untuk varietas Hawai Super Sweet pada rata-rata kondisi
optimum dan sub optimum.
37. Sidik ragam regresi vigor awal sebelum simpan (vaS) relatif (Y) terhadap nilai
DMPVMM relatif (X)
.
38. Sidik ragam regresi vigor genetik (V en) (Y) terhadap vigor awal (Va) (XI) 8an vigor awal sebelum simpan (vaS) (X2)
.
39. Sidik ragam regresi vigor genetik (V )(Y) terhadap vigor awal (Va) (XI)
,
g$?gor awal sebelum simpan (vaS) (X2) dan vigor konservasi sebelum simpan (vKSS) dengan tolok ukur nilai VKS (X3).
40. Sidik ragam regresi vigor genetik (V )
Nomor
DAFTAR GAMBAR
Teks
Halaman
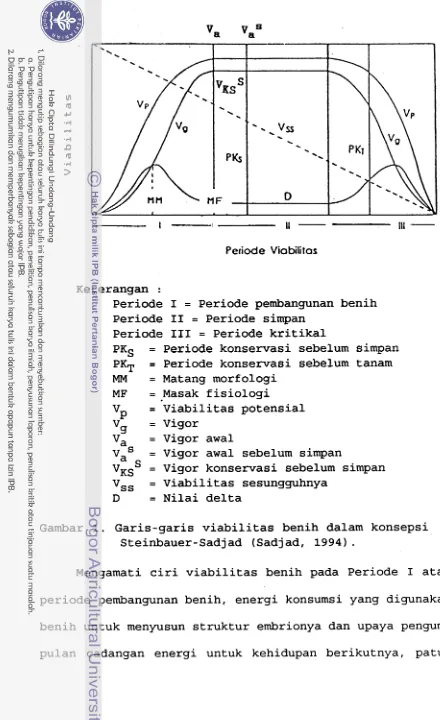
1. Garis-garis viabilitas benih dalam konsepsi
Steinbauer-Sadjad (Sadjad, 1994)
.
72. Struktur molekul asam fitat 19 3. Bobot kumulatif Pt al, Pti in dan Pnon
fitin (mg/tongko?F mulal &ari pollnasi
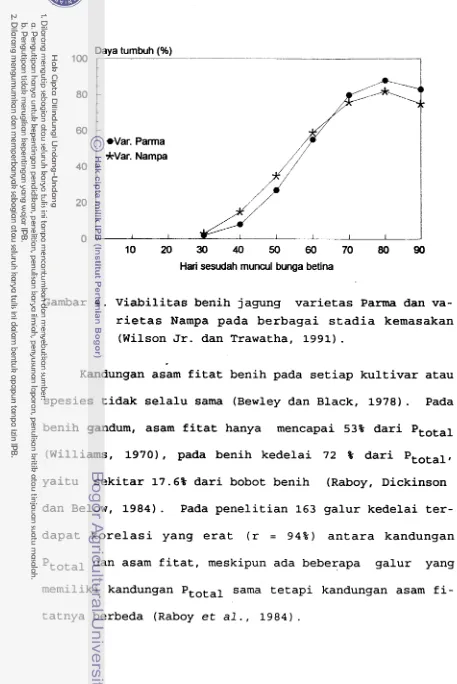
sampai masak (Earley dan DeTurk, 1944). 20 4. Viabilitas benih jagung varietas Parma dan
varietas Nampa pada berbagai stadia
kemasakan (Wilson dan Trawatha, 1991). 21
5. Perubahan ATP selama perkembangan benih
A. Benih Brassica napus var. Gorczanski dan var. Victor (Ching et al. 1974)
B. Benih gandum (Williams, 1970)
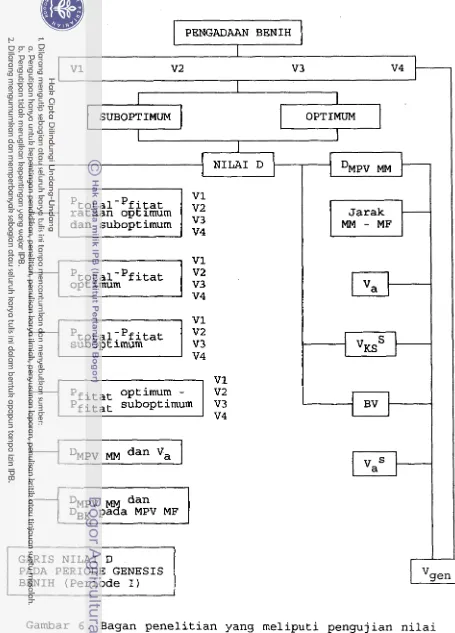
6 . Bagan penelitian yang meliputi pengujiaan
nilai D (kiri) dan hubungan antara V
dengan berbagai parameter viabilitasg%enih
lainnya (kanan) 36
7. Tanaman jagung dalam pot percobaan 42 8. Kecambah normal (atas) dan kecambah abnormal
(bawah) yang tumbuh dari embrio 3 minggu
setelah antesis 49
9. Pertumbuhan jagung varietas Hawai Super Sweet (baris kedua) tampak lemah dibandingkan varietas Harapan Baru (baris pertama) dan
varietas Arjuna (baris ketiga) 53 10. Pertumbuhan jagung pada tanah optimum
(kanan) dan sub optimum (kiri)
11. Kurva akumulasi bobot kering benih pada kondisi tanah optimum
12. Kurva akumulasi bobot kering benih pada kondisi tanah sub optimum
13. Garis nilai delta (Pkon) pada kondisi
optimum 63
15. Hasil uji tetrazolium benih varietas
Harapan Baru ( H ) , Hawai Super Sweet (S) ,
DAFTAR SIMBOL BV D DHL DMPVMM D~~ Ekon Eres Etim MPC MPV SMD
= Bidang vigor
= Delta
= Daya hantar listrik
= Delta pada momen periode viabilitas matang
morfologi
= Delta pada uji tetrazolium
= Energi yang dikonsumsi
= Energi yang sisa
= Energi yang ditimbun
= Masak fisiologi
= Matang morfologi
= Mesin pengusangan cepat
= Momen periode viabilitas
= Periode konservasi sebelum simpan
= Periode konservasi sebelum tanam
= Sistem multiplikasi devigorasi
= Vigor awal
= Vigor awal sebelum simpan
= Vigor daya simpan
= Viabilitas (pada tingkat) deteriorasi
= Vigor enforced
= Vigor genetik
= Vigor konservasi
= Vigor konservasi sebelum simpan
= Viabilitas potensial
= Viabilitas sesungguhnya
= Panjang periode viabilitas (Y=O) diukur
dari titik (0,O) sampai garis fungsi nilai
GLOSARI
Antesis
Bidang vigor
= Saat sampainya tepung sari pada kepala putik dalam proses pe- nyerbukan.
= Tolok ukur viabilitas benih dalam
dimensi waktu atas dasar perhi- tungan luas garis fungsi Nilai D.
Masak fisiolagi = Stadium akhir periode pembangunan
atau genesis benih yang mencapai bobot kering dan vigor maksimum
Matang morfologi = Suatu stadium dalam periode pem- bangunan atau genesis benih di mana benih mencapai pembangunan struk- turnya yang secara anatomis sudah lengkap, yang disusul kemudian oleh stadium pengisian bahan cadangan energinya.
Nilai delta
Parameter
Periode I
Periode I1
= Nilai selisih V dan V atau ja-
P
gbarannya yang digunakan untuk meng- ukur vigor benih dalam dimensi
waktu
.
= Sifat suatu populasi. Misalnya pa- da benih, viabilitas potensial me- rupakan parameter lot benih karena merupakan salah satu sifat lot
sebagai populasi.
= Fragmen periode viabilitas benih yang pertama, yang merupakan pe- riode pembangunan benih atau pe- riode genesis. Ditandai disaat antesis sampai benih mencapai ma- sak fisiologi.
= Fragmen periode viabilitas benih
xvi
de penyimpanan benih. Ditandai dari saat benih mencapai masak fi- siologi, sampai benih tidak bisa disimpan lagi dan h a m s segera di- t anam.
Periode konservasi = Periode simpan yang temporer, dima- na viabilitas benih dipertahankan
(konservasi) baik sebelum benih disimpan sesudah masak fisiologi selama panen dan pengolahan, atau sebelum benih ditanam sesudah me- lampaui periode simpan.
Periode simpan = Kurun waktu periode viabilitas
fragmen kedua atau disebut Periode 11.
Sistem multiplikasi
devigorasi = Dalam konteks kuantifikasi metabo- lisme benih ialah sistem untuk men- simulasi viabilitas benih dalam bentuk garis dengan membuat jabaran periode viabilitas yang pendek me- '
lalui proses devigorasi yang dimul- tiplikasi.
Uj i tetrazolium = Metode uji viabilitas benih dengan mendasarkan pewarnaan topografis benih akibat pengendapan garam For- mazan oleh terjadinya reaksi larut- an 2-3-5 Triphenyl tetrazolium
khlorida dengan ion H+ pada sel-sel yang hidup.
Viabilitas absolut = Viabilitas senyatanya. Informasi- nya bersifat simulatif yang memper- kirakan status viabilitas suatu lot benih dalam suatu kurun waktu.
xvi i
kromosom, atau garis viabilitas.
Viabilitas potensial = Parameter viabilitas lot benih yang menunjukkan kemampuan benih menumbuhkan tanaman normal yang berproduksi normalpada kondisi la- pang produksi yang optimum.
Viabilitas sesungguhnya = viabilitas yang digambarkan oleh Sadjad linear dengan periode viabilitas
Vigor awal = Parameter vigor lot benih pada sa- at benih mencapai masak fisiologi.
Vigor benih
Vigor genetik
= Kemampuan benih menumbuhkan tanaman normal pada kondisi suboptimum di lapang produksi atau sesudah disim- pan dalam kondisi simpan yang sub optimum dan ditanam dalam kondisi lapang yang optimum.
= Suatu parameter viabilitas yang mernbedakan antara keunggulan va- rietas satu dengan lainnya dalam ketahanan hidupnya terhadap ber- bagai kondisi suboptimum (umum), maupun suatu kondisi sub optimum tertentu (spesif ik)
.
Latar Belakang
Penilaian viabilitas suatu lot benih umumnya dituju-
kan untuk satu fragmen bahkan satu momen periode viabili-
tas saja, yang tidak menggambarkan status viabilitasnya
pada seluruh periode hidup benih. Kuantifikasi metabo-
lisme benih yang berlandaskan pada konsepsi Steinbauer-
Sadjad menjabarkan viabilitas benih yang diindikasikan
oleh gejala metabolisme atau pertumbuhan ke dalam garis
viabilitas mulai antesis sampai benih mati.
Ge j ala metabolisme pada periode pembangunan benih
mencakup gejala anabolisme dan katabolisme yang berkaitan
dengan pembentukan struktur benih dan pengisian cadangan
makanan serta senyawa-senyawa penting lainnya yang meng-
indikasikan energi yang digunakan ( Ekon) dan energi yang
diakumulasi (Etim) benih tersebut. Dari periode konser-
vasi sampai dengan periode kritikal (Periode 111) proses katabolisme berhubungan dengan kemunduran benih yang di-
indikasikan antara lain oleh perubahan integritas membran
sel dan aktifitas enzim.
Parameter viabilitas benih di dalam konsepsi Stein-
bauer-Sadj ad dinyatakan dalam dua keadaan, yaitu optimum
falsafatinya adalah bahwa semua kehidupan merupakan hasil
manifestasi mereka dalam mengatur diri di dalam kondisi
optimum maupun sub optimum. Kriteria vigor benih menurut
Heydecker, yang kemudian dilengkapi oleh Sadjad (1975), adalah :
1. Tahan disimpan,
2. Berkecambah cepat dan merata,
3. Bebas dari penyakit,
4. Tahan terhadap berbagai gangguan mikro organismme,
5. Bibit tumbuh kuat, baik di tanah yang basah maupun
kering,
6. Bibit dapat memanfaatkan persediaan makanan dalam be-
nih semaksimum mungkin sehingga daripadanya dapat
tumbuh j aringan- j aringan yang b a n ,
7. La ju tumbuhnya t inggi ,
8. Menghasilkan produksi yang tinggi dalam waktu ter-
tentu,
9. Antara pertumbuhan di lapangan dan daya berkecambah
di laboratorium tidak menunjukkan perbedaan,
10. Tahan terhadap saingan baik terhadap tumbuhan-
tumbuhan lain atau tanaman lain, baik spesiesnya sen-
diri maupun spesies lain dalam pertanaman tersebut.
Produksi sebagai salah satu kriteria vigor benih di-
bidang ilmu dan teknologi benih, vigor genetik (TIgen) me-
rupakan parameter viabilitas yang membedakan keunggulan
varietas yang satu dengan lainnya dalam ketahanan hidup-
nya terhadap berbagai kondisi sub optimum (umum) atau
kondisi sub optimum tertentu (spesif ik) (Sadjad, 1994)
.
Deteksi Vgen dilakukan dengan mengamati keragaman
fenotipe berbagai varietas pada lingkungan yang sama.
Vgen yang didasarkan pada tolok ukur produksi, ditentu-
kan oleh banyak sifat, yang sebagian berhubungan dengan
pembentukan dan pengisian benih serta status vigor benih
setelah masak fisiologi.
Pada periode pembangunan benih, akumulasi Ptotal dan
'fitat menunjukkan kecenderungan yang sama dengan garis viabilitas potensial dan vigor benih dalam konsepsi
Steinbauer-Sadjad. Dari hasil penelitian diketahui bahwa
kandungan Ptotal didalam benih mempengaruhi vigor awal
(Rusdi, 1988) dan daya simpan benih (Kurnia, 1989).
Untuk periode konservasi dan awal periode simpan, status
vigor benih dapat diindikasikan antara lain oleh aktifi-
tas enzim dehidrogenase dan kemampuan benih mempertahan-
kan integritas mernbran sel.
Hubungan antara masing-masing parameter viabilitas
benih terhadap parameter V gen serta besarnya kontribusi
terhadap V
gen hingga saat ini belum diketahui. Untuk itu perlu dilakukan penelitian mengenai deteksi V gen melalui
kuantifikasi metabolisme benih berdasarkan kaidah-kaidah
viabilitas benih.
Tu j uan
Penelitian ini bertujuan mempelajari kuantifikasi
metabolisme benih mulai periode pembangunan benih (Perio-
de I) sampai dengan awal simpan Periode I1 untuk mende- teksi Vgen jagung.
Hypotesis
1.
vgen
jagung merupakan fungsi vigor benih pada saatmasak fisiologi di periode I (vigor awal, Va)
,
vigorkonservasi sebelum simpan (vKSS) dan vigor awal sebe-
lum simpan Was)
.
Semakin tinggi ValvKSS
danvaS
semakin tinggi Vgen. S
Vgen w f(Vat VKS
, vaS)
2. Pada periode I, vigor awal benih (Va) dapat dideteksi
melalui :
a. Vigor yang dinilai dari DMPVMM pada saat matang
morfologi. Semakin besar nilai vigor atas dasar
DMPVMM, semakin tinggi Va benih.
gi. Semakin efisien, semakin tinggi Va benih.
c. Bobot kering benih pada saat masak fisiologi. Se-
makin tinggi bobot kering benih, semakin tinggi Va
benih.
3. Pada periode I, garis nilai delta yang diamati berda-
sarkan tolok ukur P total dan P fitat menunjukkan ke-
cenderungan kuadratik sesuai dengan kaidah viabilitas
benih dan dapat digunakan menentukan saat matang mor-
f ologi
.
4.
vKSS
dapat dideteksi melalui SMD (Sistem MultiplikasiDevigorasi)
,
dengan tolok ukur ZPo, DTZ-I danBV-l .
Semakin tinggi ZPo, D T Z - ~ dan BV-', semakin tinggi vi-
gor konservasi benih.
5.
vaS
dapat diindikasikan berdasarkan integritas membransel benih yang diamati melalui tolok ukur daya hantar
listrik rembesan benih. Semakin tinggi daya hantar
11. TINJAUAN PUSTAKA
Kuantifikasi Metabolisme Benih
Viabilitas benih sebagai fokus ilmu benih dapat di-
telaah melalui pendekatan sitologi, fisiologi, biokimiawi
dan matematik. Kuantifikasi metabolisme menurut Sadjad
(1994) pada hakikatnya memanfaatkan ciri-ciri garis yang
menunjukkan fenomena pertumbuhan ataupun gejala metabo-
lisme benih dari awal periode pembentukan benih sampai
periode pertumbuhan di lapang untuk mendeteksi viabilitas
benih dalam dimensi waktu, dan pada akhirnya dapat me-
ngembangkan simulasi viabilitas absolut benih.
Konsepsi Steinbauer-Sadjad yang merupakan landasan
dari kuantifikasi metabolisme benih, membagi periode via-
bilitas menjadi tiga fragmen, yaitu fragmen pembangunan
benih pada Periode I, fragmen penyimpanan benih pada Pe-
riode I1 dan fragmen penanaman pada Periode I11 atau pe-
riode kritikal. Garis-garis viabilitas yang digambarkan
dalam masing-masing fragmen tidak sama (Gambar 1). Garis
viabilitas potensial (V ) dan vigor (V ) pada Periode I
P
9dan I11 berbentuk sigmoid dengan arah berlawanan, sedang-
7..
Petiode Viobilitos
Keterangan :
Periode I = Periode pembangunan benih Periode I1 = Periode simpan
Periode I11 = Periode kritikal
PKS = Periode konservasi sebelum simpan PKT = Periode konservasi sebelum tanam MM = Matang morfologi
MF = Masak fisiologi
v~
= Viabilitas potensialvg = Vigor
Va = Vigor awal
= Vigor awal sebelum simpan Va
v~~
= Vigor konservasi sebelum simpan vss = Viabilitas sesungguhnya [image:184.563.46.486.10.730.2]D = Nilai delta
Gambar 1. Garis-garis viabilitas benih dalam konsepsi Steinbauer-Sadjad (Sadjad, 1994)
.
Mengamati ciri viabilitas benih pada Periode I atau
periode pembangunan benih, energi konsumsi yang digunakan
benih untuk menyusun struktur ernbrionya dan upaya pengum-
di j adikan j abaran nilai D. Bertolak dari pengamatan
Early dan DeTurk (1944) terdapat garis-garis Ptotal dan
Pfitinl yang kalau diamati dari saat antesis sampai ma-
sak fisiologi, selisih keduanya memiliki kecenderungan
garis yang sama dengan garis nilai D pada konsepsi Stein-
bauer-Sadjad (Sadjad, 1994).
Bila dikaitkan dengan akumulasi P di dalam benih,
energi konsumsi (Elion) dapat diasosiasikan sebagai P yang
digunakan untuk pertumbuhan dan perkembangan benih. Ni-
lai Ekon dapat diperoleh dari selisih energi timbunan
(Etim) dengan sisa energi (Ere,
,
yaitu selisih jumlahPtotal dan jumlah Psimpanan (Pfifat) Dasar pernikirannya adalah semakin kecil nilai D semakin tinggi vigor benih.
Kecilnya nilai D dapat diperoleh dari Etim yang besar dan
Ekon yang besar, tetapi dapat pula berasal dari Etim yang
kecil dan Ekon yang kecil. Agar dapat dibedakan antara
kemungkinan pertama dengan yang kedua, perlu diberikan
faktor koreksi dengan mengalikannya dengan Eres/Ekon, se-
hingga pada kemungkinan pertama nilai D makin diperkecil
sedangkan pada kemungkinan kedua nilai D makin diperbesar
(Sadjad, 1994).
Nilai D = (Etim
-
Ekon) x FKFK = Eres/Ekon
Nilai D = Ere, Eres/Ekon = 2 IEkon
Seperti terlihat pada Gambar 1, nilai D meningkat
terus mulai saat antesis sampai saat matang morfologi,
kemudian menurun kembali sampai saat masak f isiologi
.
Titik maksimum bagi nilai D berada di sekitar saat matang
morf ologi, karena pada momen itu terjadi banyak pembela-
han dan dif erensiasi sel
.
Selanjutnya terj adi pengisiancadangan makanan ke dalam benih, sampai saat masak fisio-
logi, dimana nilai D minimum sedangkan bobot kering benih
dan vigor benih maksimum. Saat matang morfologi dan ma-
sak fisiologi merupakan momen yang penting untuk dipakai
sebagai titik kritikal dalam penentuan viabilitas benih
pada Periode I.
Menurut Sadjgd (1989) benih j agung mencapai saat ma-
tang morfologi 20 hari setelah antesis dan saat masak fi-
siologi 30 hari setelah itu, khususnya untuk jagung yang
biasa dipanen 56 hari setelah antesis. Frey (1981) me-
nyatakan bahwa periode kritikal bagi perkembangan benih
jagung adalah 2 sampai 3 minggu setelah keluar rambut
50%. Saat masak fisiologi oleh Daynard dan Duncan
(1969) dicirikan dengan adanya b l a c k l a y e r yang menun-
jukkan bobot kering benih telah mencapai maksimum
.
Setelah benih mencapai vigor maksimum pada momen pe-
perode konservasi (PK). PK meliputi waktu sesudah MPV MF
sebelum panen, waktu panen, waktu pengolahan dan pena-
nganan benih (seed handling). Pada waktu itu vigor benih
harus dikonservasi, sehingga disebut vigor konservasi se-
belum simpan (vKSS) (Sadjad, 1994)
.
Parameter VKS memiliki tolok ukur spesif ik. Hingga
saat ini
vKSS
selalu dideteksi melalui sistem multiplika-si devigorasi (SMD) dengan tolok ukur nilai VKS dan luas
bidang vigor (BV) (Sadjad, 1993).
Apabila selama PK benih dapat mempertahankan vigor-
nya, maka benih akan mempunyai
vaS
(vigor awal sebelumsimpan) tinggi sehingga didapatkan daya simpan (DS ) yang
lama dan vigor daya simpan (VDS) yang tinggi pula.
Dalam hasil. penelitian Saenong (1986), hubungan an-
tara vigor awal (Va) benih dengan DS benih jagung - dan
kedelai adalah sebagai berikut :
1. Benih jagung
DS = 24.265
-
0.131 Va + 0.197 Ve-
0.006 VaVeVa =
-
16.45 + 2.60 XI Ve = - 29.31+
1.44 X22. Benih kedelai
DS = 4.67 - 0.105 Va + 0.290 V,
-
0.003 VaVe V, = - 45.97 + 0.69 X1DS = Daya simpan benih yang dinyatakan dalam kurun waktu
penurunan viabilitas benih sebesar 40% (P40) (minggu)
Va = Vigor awal benih
Ve = Vigor enforced benih = vigor benih yang dipengaruhi
faktor lingkungan simpan
X 1 = Daya hantar listrik (DHL) air rendaman benih
(umhos/g benih)
X2 = Viabilitas benih setelah didera etanol selama (15 +
45) menit untuk benih jagung dan (15
+
15) menit un-tuk benih kedelai
in-'^%)
.
Seperti telah dikemukakan sebelumnya, benih yang
mempunyai Va yang tinggi diperkirakan akan mempunyai DS
panjang dan VDS yang tinggi. Dengan status vigor yang
demikian diharapk-an benih akan mampu melewati periode hi-
dup selanjutnya (periode konservasi sebelum tanam, PKT)
tanpa mengalami deteriorasi yang besar, sehingga pada sa-
at ditanam dapat tumbuh menjadi tanaman yang normal dan
berproduksi normal pada kondisi lapang yang beragam.
Vigor Genetik
Menurut International Seed Testing Association
(ISTA), vigor benih adalah semua sifat-sifat yang menen-
tukan tingkat aktifitas dan penampakan potensial dari be-
bit; sedangkan Association of Official Seed Analysts
(AOSA) memberikan definisi vigor lebih singkat dan lang-
sung (meskipun hampir sama) yaitu semua sifat yang menen-
tukan potensi kecepatan dan keserempakan tumbuh, dan po-
tensi untuk dapat berkembang menjadi bibit yang normal
pada kondisi lingkungan yang beragam (AOSA, 1983)
.
Mc.
Daniel (1973) membuat batasan vigor benih sebagai penam-
pakan superior suatu genotipe, setelah ditanam, diban-
dingkan dengan genotipe yang sama atau genotipe yang ber-
beda pada kondisi tertentu. Sadjad (1972) mendefinisikan
vigor benih sebagai kemampuan benih untuk tumbuh dan ber-
kembang menjadi bibit yang normal dalam kondisi lingkung-
an tumbuh yang sub optimum, sehingga diharapkan benih itu
dapat kuat tumbuh menjadi tanaman normal meskipun kondisi
alam tidak ideal. Selain itu, benih mampu berproduksi
normal pada kondisi sub optimum dan diatas normal pada
kondisi optimum (Sadj ad, 1980 )
.
Sebab- sebab perbedaanvigor benih dapat bersifat genetik maupun fisiologi
(Sadjad, 1972)
.
Perry dalam AOSA (1983) mendef inisikanvigor benih sebagai sifat fisiologi yang ditentukan oleh
genotipe dan dimodifikasi oleh lingkungan, yang mengatur
kemampuan benih untuk tumbuh cepat menjadi bibit yang ta-
han terhadap faktor-faktor lingkungannya yang beragam.
bit; sedangkan Association of Official Seed Analysts
(AOSA) memberikan definisi vigor lebih singkat dan lang-
sung (meskipun hampir sama) yaitu semua sifat yang menen-
tukan potensi kecepatan dan keserempakan tumbuh, dan po-
tensi untuk dapat berkembang menjadi bibit yang normal
pada kondisi lingkungan yang beragam (AOSA, 1983)
.
Mc.
Daniel (1973) membuat batasan vigor benih sebagai penam-
pakan superior suatu genotipe, setelah ditanam, diban-
dingkan dengan genotipe yang sama atau genotipe yang ber-
beda pada kondisi tertentu. Sadjad (1972) mendefinisikan
vigor benih sebagai kemampuan benih untuk tumbuh dan ber-
kembang menjadi bibit yang normal dalam kondisi lingkung-
an tumbuh yang sub optimum, sehingga diharapkan benih itu
dapat kuat tumbuh menjadi tanaman normal meskipun kondisi
alam tidak ideal. Selain itu, benih mampu berproduksi
normal pada kondisi sub optimum dan diatas normal pada
kondisi optimum (Sadjad, 1980). Sebab-sebab perbedaan
vigor benih dapat bersifat genetik maupun fisiologi
(Sadjad, 1972)
.
Perry dalam AOSA (1983) mendef inisikanvigor benih sebagai sifat fisiologi yang ditentukan oleh
genotipe dan dimodifikasi oleh lingkungan, yang mengatur
kemampuan benih untuk tumbuh cepat menjadi bibit yang ta-
han terhadap faktor-faktor lingkungannya yang beragam.
dan mempengaruhi hasil tanaman tersebut.
Produksi adalah parameter vigor penting dan umum di-
gunakan untuk menentukan perbedaan genetik. Untuk dapat
menilai perbedaan hasil diantara berbagai varietas, va-
rietas-varietas tersebut biasanya ditanam pada lingkungan
yang sama dan kemudian hasilnya dibandingkan (Pollock dan
Roos, 1972)
.
Menurut Sadjad (1994) vigor genetik (Vgen)merupakan parameter viabilitas yang membedakan antara ke-
unggulan varietas satu dengan lainnya dalam ketahanan hi-
dupnya terhadap berbagai kondisi sub optimum (umum), m u -
pun suatu kondisi sub optimum tertentu (spesifik).
Whittington (1973) menyatakan bahwa mempelajari ke-
ragaman kuantitatif yang dikendalikan secara genetik pada
perkecambahan benih bukanlah ha1 yang mudah. Faktor-
faktor lingkungan selama pembentukan benih dan penyimpan-
an benih memberikan pengaruh yang besar terhadap perke-
cambahan benih selanjutnya, sementara dari segi genetik
testa dan bagian luar benih merupakan genotipe maternal,
endosperm biasanya 2/3 maternal dan 1/3 paterna1,dan em-
brio setengah maternal dan setengah lagi paternal genoti-
Pe
Dari beberapa penelitian tentang keragaman genotipe
terhadap vigor benih, sering di jumpai adanya interaksi
(Whittington, 1973; Buxton dan Sprenger, 1976; Tomer dan
Maguire, 1990)
.
Melalui tehnik regresi Finlay danWilkinson (1963), perbedaan kecenderungan respons masing-
masing genotipe terhadap lingkungan sub optimum sampai
dengan optimum dapat diketahui dari koefisien regresi
yang diperoleh. Pada penelitian benih bit gula, jagung
dan kacang polong perbedaan genotipe terlihat paling je-
las pada kondisi sub optimum (Whittington, 1973), sedang-
kan pada kondisi optimum kurang terlihat perbedaannya.
Pada benih kapas, penelitian dengan 18 galur Gossypium
h i r s u t u m dan 12 galur G. barbadens menunjukkan adanya ko-
relasi positif antara parameter viabilitas dengan suhu
udara (optimum dan rendah), namun nilainya kecil sehingga
evaluasi galur-galur tersebut h a m s dilakukan pada ke-
dua kondisi tersebut (Buxton dan Sprenger, 1976).
Hibrida barley dan hibrida j agung menunjukkan per-
tumbuhan dan perkecambahan yang lebih cepat serta laju
respirasi yang lebih tinggi dibandingkan dengan tetuanya.
Perbedaan tersebut sebagian dicirikan dari ukuran benih-
nya (Mc. Daniel, 1973)
,
dan jumlah benih/tongkol yanglebih banyak khususnya pada benih jagung hibrida
(Poneleit dan Egli, 1979)
.
Mc. Daniel (1973) mempelaja-ri faktor-faktor genetik yang mempengaruhi vigor benih
faktor genetik menentukan potensi tumbuh dan vigor mela-
lui proses biokimiawi. Mitokhondria yang berfungsi seca-
ra superior pada hibrida heterotik (yang dinamakannya mi-
tokhondrial heterosis) diukur berdasarkan antara lain
laju respirasi dan efisiensi oxidative fosf orilasi. Mi-
tokhondrial heterosis ini berkorelasi sangat erat dengan
vigor dan potensi produksi hibrida. Hibrida yang tidak
heterotik (yaitu yang tidak lebih unggul dari tetuanya
dalam ha1 laju pertumbuhan dan hasil) tidak menunjukkan
fungsi mitokhondria yang superior. Jadi mitokhondrial
heterosis dapat dijadikan sebagai tolok ukur untuk menge-
valuasi vigor hibrida secara biokimiawi.
Copeland (1976) menyatakan bahwa selain tanaman hi-
brida, tanaman poliploid juga menunjukkan Vgen yang lebih
tinggi dibandingkan dengan tanaman inbred dan diploid.
Pada benih kacang polong, Vgen yang diamati berda-
sarkan persentase perkecambahan berhubungan dengan kompo-
sisi kimia benih tersebut. Varietas Gradus yang Vgennya
rendah, mengandung lebih banyak stahiosa daripada pati,
dibandingkan varietas Pilot yang mempunyai Vgen tinggi
(Haldane dalam Whittington, 1973), dan mengandung lebih
banyak sukrosa (Koostra, 1962).
Perbedaan satu gen tynggal di bagian endosperm (tri-
Haskel yang dikemukakan Whittington (1973) menunjukkan
bahwa daya berkecambah benih jagung menurun bila perban-
dingan antara sugary allel (su) terhadap starchy allel
(Su) di dalam endosperm meningkat
,
seperti terlihat padaTabel 1. Pengaruh proporsi yang berbeda antara Su (Star- chy endosperm allel) terhadap su (sugary enda- sperm allel) terhadap persentase perkecambahan.
Endosperm genotipe Persentase Perkecambahan
Su Su Su 93
Su Su su 87
Su su su 67
su su su 46
Kadar gula (86% merupakan sukrosa) dari 24 genotipe
* *
homozygous jagung manis berkorelasi negatif (r = -0.74dengan daya tumbuh di lapang, sedangkan kandungan pati
berkorelasi positif (r = 0.63**) dengan daya tumbuh di
lapang (Douglass, Juvik dan Splittstoesser, 1993).
Daya simpan (DS benih j uga dipengaruhi oleh kompo-
sisi kimia benih dan faktor genetik. Benih berlemak me-
rupakan contoh benih yang berdaya simpan pendek, sebagian
disebabkan karena sifat lemak yang kurang higroskopik,
dan air bebas didalam benih itu sulit dikendalikan. Se-
[image:194.566.46.497.49.727.2]proses yang mempercepat kemunduran benih karena asam le-
mak dapat meningkatkan aktifitas cendawan dan juga bersi-
fat toksik bagi benih. Meskipun demikian, benih dari
spesies tertentu ada yang mempunyai kesamaan komposisi
kimia tetapi daya simpannya berbeda akibat perbedaan po-
tensi genetik. Contohnya benih c h e w i n g f e s c u e dan L o l i u m
m u l t i f l o r u m Lam. mempunyai komposisi kimia yang sama, te-
tapi benih L o l i u m m u l t i f l o r u m Lam.mempunyai DS yang lebih
lama (Copeland, 1976)
.
Pada 31 varietas dan galur padi, DS tidak berkorelasi dengan kandungan amilosa benih dandormansi benih. Meskipun demikian, benih-benih yang ter-
golong w a x y rice daya simpannya lebih pendek dari pada benih non waxy r i c e (Juliana, e t al., 1990).
Selain komposisi kimia benih, perbedaan konsistensi
struktural benih pada varietas-varietas jagung sangat be-
sar pengaruhnya terhadap vigor benih selama di penyimpan-
an. Berdasarkan hasil penelitian selama 20 tahun, ter- nyata varietas-varietas jagung yang termasuk ke dalam ti-
pe mutiara dan gigi kuda mempunyai DS yang lebih panjang
dari pada benih jagung manis (Priestley, 1986).
Seleksi sifat genetik DS benih melalui metode peng-
pengusangan cepat berkurang dengan sangat nyata. Ini me-
nun j ukkan bahwa perbaikan genet ik untuk DS benih memung-
kinkan untuk dilakukan melalui seleksi dengan metode
pengusangan cepat. Penelitian 235 genotipe kedelai yang
disimpan dalam kondisi simulasi tropika dengan suhu 30°c
dan RH 80%, menunjukkan bahwa genotipe yang mempunyai DS
pan j ang mempunyai viabilitas awal yang tinggi, mempunyai
persentase hard seed yang tinggi, ukuran benih yang kecil
dan waktu pemasakan benih yang lebih cepat (Minor dan
Paschal, 1982)
.
Dari penelitian Minor dan Paschal (1982) tersebut,
terlihat bahwa waktu pemasakan benih dan viabilitas awal
benih di periode pembangunan benih (Periode I Stein-
bauer-Sadj ad) yang dipengaruhi oleh faktor genetik, me-
nentukan vigor benih pada periode simpan. Hasil peneliti-
an Faizah (1994) juga memperlihatkan bahwa Vgen pada be-
nih kedelai dapat dideteksi melalui parameter vigor awal
V
,
vigor konservasi sebelum simpan (vKSS) dengan tolokukur nilai VKS dan kebalikan luas bidang vigor (BV) -I,
serta parameter vigor awal sebelum simpan (vas)
.
Deteksi Vgen benih mulai dari momen berlakunya kai-
dah-kaidah viabilitas benih (saat matang morfologi) Sam-
pai dengan titik anomali di periode kritikal (Periode
Akumulasi dan pengaruh P terhadap viabilitas benih
Unsur P adalah salah satu unsur yang banyak diperlu-
kan dalam biosintesis makromolekul, antara lain fosfoli-
pid, gula fosfat, nukleotida dan koenzim. Di dalam be-
nih, P disimpan dalam bentuk fitin, yaitu garam (Ca,Mg)
dari asam fitat (myoinositol hexafosfat) (Epstein, 1972).
Asam fitat (Gambar 2) merupakan sumber P bagi proses me-
tabolisme selama perkecambahan (Gardner et al., 19851,
sebelum dapat menyerap P dari dalam tanah.
H OPO ,H2
I I
Gambar 2. Struktur molekul asam fitat
Selama perkembangan benih j agung, akumulasi P
(Ptotal dan Pf itin) menunjukkan kecenderungan garis sig-
moid, seperti terlihat pada Gambar 3 (Early dan DeTurk,
1944). Demikian pula dengan akumulasi P pada benih kede-
lai (Hanway dan Weber, 1971).
H inggu
Gambar 3. Bobot kumulatif Ptotal, Pfitin dan Pnon fitin (mg/tongkol) mulai dari polinasi sampai masak (Earley dan DeTurk, 1944)
.
but ternyata sama dengan kecenderungan garis akumulasi
bobot kering benih jagung (Frey, 1981) dan garis viabili-
tas benih jagung manis varietas P a m dan Nampa (Wilson
Jr. dan Trawatha (1991) seperti terlihat pada Gambar 4.
Tidak berbeda dengan jagung, akumulasi bobot kering pada
benih gandum juga cenderung sigmoid, tetapi pada barley
cenderung kuadratik (Lingle dan Chevalier, 1985). Hasil
penelitian Agrawal dan Kaur (1977) pada benih gandum me-
nunjukkan bahwa pola akumulasi bobot kering cenderung sa-
Daya tumbuh (%)
loo
7
I
1 0 2 0 3 0 4 0 5 0 6 0 70 80 90
[image:199.563.43.500.45.729.2]Hari sesudah muncul bunga betina
Gambar 4 . Viabilitas benih jagung varietas Parma dan va- rietas Nampa pada berbagai stadia kemasakan
(Wilson Jr. dan Trawatha, 1991)
.
Kandungan asam fitat benih pada setiap kultivar atau
spesies tidak selalu sama (Bewley dan Black, 1978). Pada benih gandum, asam fitat hanya mencapai 53% dari Ptotal (Williams, 1970), pada benih kedelai 72 % dari Ptotall
yaitu sekitar 17.6% dari bobot benih (Raboy, Dickinson dan Below, 1984). Pada penelitian 163 galur kedelai ter-
dapat korelasi yang erat ( r = 94%) antara kandungan Ptotal dan asam fitat, meskipun ada beberapa galur yang
memiliki kandungan Ptotal sama tetapi kandungan asam fi-
Dalam benih serealia, fitin bergabung dengan butir-
butir aleuron di lapisan aleuron dan jarang sekali dite-
mukan dalam protein body di endosperm. Hasil penelitian
Williams (1970) menun jukkan bahwa pada benih gandum asam
fitat hanya ditemukan di dalam lapisan aleuron, poros em-
brio dan skutelum.
Menurut Ching et al. (1974), benih pada stadia masak
hampir tidak mengandung P dalam bentuk ATP. ATP sebagai
sumber energi, hanya dibentuk selama proses pembelahan
sel dan pengisian cadangan makanan. Kandungan ATP menca-
pai maksimum pada saat bobot basah benih meficapai maksi-
mum, kemudian turun terus dan mencapai minimum pada saat
benih masak dimana benih berada da1,am keadaan kering dan
istirahat (quiesce'nce)
.
Perubahan kandungan ATP selamaperkembangan benih dapat dilihat pada Gambar 5.
Fitase adalah enzim yang menghidrolisis fitin untuk
melepaskan P, kation-kation (Ca, Mg) dan myoinositol.
Fosfat digunakan dalam proses respirasi dan pembentukan
makro molekul di bagian axis. Myoinositol diduga diguna-
kan untuk membentuk dinding sell karena senyawa tersebut
adalah prekursor gula pentosyl dan uranosyl yang biasa
berasosiasi dengan pektin dan polisakarida-polisakarida
dinding sel lainnya (Bewley dan Black, 1985).