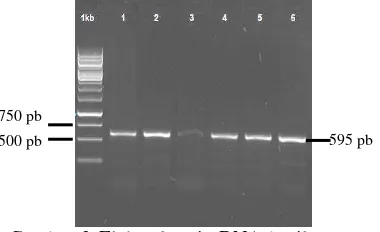
Karakteristik Gen Cytochrome Oxidase Subunit I (COI) pada Ulat Sutera Liar (Attacus atlas L.) asal Jawa Barat
Bebas
30
0
0
Teks penuh
Gambar
Dokumen terkait
Penelitian lebih lanjut mengenai penggunaan gen GH sebagai marker genetik untuk kualitas karkas masih sangat perlu dilakukan, disarankan juga untuk melakukan sekuen dengan sampel
Tujuan dari penelitian ini adalah untuk mengeksplorasi urutan basa nukleotida gen Cytochrome Oxidase Subunit I (COI) dari kerang mutiara Pinctada fucata yang berasal
