KULTIVAR SLAMET DAN LUMUT
SIH HARTINI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul ”Induksi Mutasi dengan Irradiasi Sinar Gamma pada Kedelai (Glycine max (L.) Merrill) Kultivar Slamet dan Lumut” merupakan gagasan dan karya saya bersama pembimbing yang belum pernah dipublikasikan dalam bentuk apapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan oleh penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir tesis ini.
Bogor, Agustus 2008
SIH HARTINI. (Induction of Mutation with Gamma Ray Irradiation at Slamet and Lumut Cultivars of Soybean (Glycine max (L.) Merrill)).Under direction of
UTUT WIDYASTUTI and MUHAMMAD JUSUF.
The aim of this research was to produce Gα mutants of two cultivars of Soybean using 60Co gamma irradiation with 0.0; 0.1; 0.2; 0.3; 0.4; and 0.5 kGy dosage. Two Soybean cultivars tested were Slamet and Lumut. The mutated Soybean were detected at M-1 and M-2 filials. M-1 was used for screening general mutation based on plant hight variable, and M-2 for Gα and stability mutations based on stomatal opening characteristic. The results indicated that irradiation at dosage of 0.1 – 0.5 kGy produced plant mutation in different characteristics such as big seeds, flower’s colour, leaf’s form and leaflets mutation of the two cultivars. In addition, 60Co gamma irradiation at the dosage of 0.1 dan 0.3 kGy produced 10 Gα mutants putatives. The mutant had 24-40 cm hight and closed stomata at both cultivars. Molecular analysis was run in one Gα gene mutant putative only. Based on molecular analysis showed that gamma irradiation at 0.3 kGy in Slamet cultivar caused 1 Gαgene mutant.
RINGKASAN
SIH HARTINI. (Induksi Mutasi dengan Irradiasi Sinar Gamma pada Kedelai (Glycine max (L.) Merrill) kultivar Slamet dan Lumut). Dibimbing oleh UTUT WIDYASTUTI dan MUHAMMAD JUSUF.
Kedelai merupakan bahan makanan penting karena dapat dikonsumsi secara langsung maupun digunakan sebagai bahan baku agroindustri, namun untuk memenuhi tingginya kebutuhan kedelai masih tergantung impor karena produksi nasional masih sangat rendah. Untuk memenuhi kebutuhan tersebut perlu dilakukan upaya peningkatan produksi. Peningkatan produksi bisa dilakukan dengan berbagai macam cara antara lain melalui usaha pemuliaan tanaman. Pemuliaan tanaman dapat dilakukan dengan merakit kultivar baru, dan keragaman merupakan modal dasar untuk merakit kultivar baru. Salah satu upaya peningkatan keragaman yaitu dengan induksi mutasi. Mutasi bisa dihasilkan oleh beberapa agen mutagenik seperti radiasi, non radiasi maupun kimia. Sumber irradiasi yang sering digunakan adalah sinar X, sinar gamma, ultra-violet, sinar beta dari radioisotop dan sinar neutron dari reaktor atom. Penelitian ini menggunakan sinar gamma dari 60Co.
Kedelai kultivar Lumut dan Slamet memiliki perbedaan dalam sistem toleransi terhadap aluminium. Kultivar Lumut merupakan kultivar yang peka terhadap cekaman aluminium sedangkan kultivar Slamet toleran. Gen Gα diduga berperan dalam sistem toleransi terhadap aluminium pada kedua kultivar tersebut. Untuk melihat keterlibatan fungsi gen Gα secara langsung dalam sistem pertahanan terhadap cekaman Al dapat dilakukan dengan pendekatan reverse genetic yaitu menonaktifkan gen Gα. Teknik yang digunakan untuk menonaktifkan gen tersebut adalah dengan induksi mutasi menggunakan irradiasi sinar gamma karena radiasi gamma mempunyai energi yang sangat besar sehingga mampu menembus jaringan tanaman dengan maksimal.
Tujuan penelitian ini adalah untuk memperoleh berbagai macam mutan termasuk mutan gen Gα pada kedelai kultivar Slamet dan Lumut yang diinduksi irradiasi sinar gamma. Untuk mendapatkan mutan tersebut dilakukan skrining, dengan menanam kedelai dalam dua periode tanam. Periode tanam I (M-1) mengikuti pola Rancangan Acak Kelompok (RAK) dengan 2 faktor (dosis irradiasi dan kultivar) dan 3 kelompok ulangan, merupakan tahapan seleksi terhadap berbagai macam mutan. Dosis irradiasi yang digunakan adalah 0; 0.1; 0.2; 0.3; 0.4; dan 0.5 kGy, kultivar yang digunakan adalah Lumut dan Slamet. Kedelai ditanam dalam petakan, tiap petak terdapat 100 lubang, masing-masing lubang berisi 2 biji kedelai. Mutan yang diperoleh selanjutnya ditanam kembali sebagai generasi M-2. Rancangan yang digunakan dalam periode tanam II adalah Rancangan Acak Lengkap (RAL). Benih ditanam dengan membentuk barisan yang bersambungan antara satu induk dengan induk yang lain. Tiap induk diambil 30 biji, namun untuk induk yang memiliki jumlah biji kurang dari 30, biji ditanam semua. Hasil seleksi periode tanam I, dari 657 induk ditanam sebanyak 10.216 tanaman.
Selanjutnya mutan tersebut diuji stabilitasnya pada generasi M-2. Mutan stabil ditunjukkan oleh tidak adanya segregasi pada generasi M-2. Mutan warna biji dan mutan motif daun belum stabil karena mengalami segregasi pada generasi M-2, sedangkan mutan lainnya sudah stabil tidak bersegregasi. Mutan pendek merupakan prioritas dalam penelitian ini karena merupakan indikator tanaman tersebut mengalami mutasi Gα. Selanjutnya mutan pendek tersebut diperiksa perilaku stomatanya. Mutan Gα ditunjukkan oleh perubahan pembukaan stomata dimana stomata cenderung menutup. Untuk melihat perilaku stomata diambil tigapuluh sampel mutan pendek stabil, dari tigapuluh sampel tersebut diperoleh sepuluh tanaman dengan kondisi stomata menutup. Selanjutnya tanaman yang memiliki stomata menutup dilihat ekspresi gen Gα nya untuk memastikan bahwa tanaman tersebut telah mengalami mutasi. Sepuluh tanaman yang diduga mutan
Gα berdasarkan perilaku stomata, baru diperiksa satu mutan Gα berdasarkan ekspresi gennya. Mutan tersebut berasal dari kultivar Slamet dosis irradiasi 0.3 kGy. Tanaman wild type (kontrol) menghasilkan pita dengan ukuran 1380 pb, sedangkan tanaman mutan tidak menghasilkan pita tersebut. Tidak munculnya pita pada mutan menunjukkan bahwa gen Gα tidak diekspresikan karena telah mengalami mutasi.
© Hak Cipta milik IPB, tahun 2008
Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah.
b. Pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini dalam bentuk apapun tanpa izin IPB.
KULTIVAR SLAMET DAN LUMUT
SIH HARTINI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Departemen Biologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Induksi Mutasi dengan Irradiasi Sinar Gamma pada Kedelai (Glycine max (L.) Merrill) Kultivar Slamet dan Lumut
Nama : Sih Hartini
NRP : G 351060081
Disetujui Komisi Pembimbing
Dr. Ir. Utut Widyastuti, MSi. Dr. Ir. Muhammad Jusuf Ketua Anggota
Diketahui
Ketua Program Studi Biologi Dekan Sekolah Pascasarjana
Dr. Ir. Dedy Duryadi S, DEA Prof. Dr. Ir. Khairil Anwar Notodiputro, MS.
limpahan rahmat-Nya sehingga penulis dapat menyelesaikan penelitian serta penyusunan tesis ini.
Penulis menyadari sepenuhnya bahwa sangatlah sulit untuk menyelesaikan penelitian dan penyusunan tesis ini tanpa bantuan dari berbagai pihak. Oleh karena itu dalam kesempatan ini penulis ingin menyampaikan ucapan terimakasih serta penghargaan yang setinggi-tingginya kepada:
1. Dr. Ir. Utut Widyastuti, MSi. selaku ketua komisi pembimbing atas semua jerih payah dan waktu yang telah diluangkan untuk memberikan bimbingan, arahan dan kemudahan kepada penulis dari mulai penelitian hingga terselesaikannya tesis ini.
2. Dr. Ir. Muhammad Jusuf selaku anggota komisi pembimbing atas ilmu, waktu dan bimbingan yang diberikan kepada penulis dengan penuh kesabaran sehingga tesis ini dapat terselesaikan.
3. Dr. Ir. Hajrial Aswidinnoor, MSc. selaku dosen penguji atas kritik dan sarannya untuk kesempurnaan tesis ini.
4. Dekan Sekolah Pascasarjana IPB dan Ketua Program Studi Biologi atas kesempatan yang diberikan kepada penulis untuk mengikuti pendidikan Pascasarjana di IPB.
5. Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) dan Laboratorium Anatomi Tumbuhan Departemen Biologi IPB yang telah menyediakan fasilitas untuk melakukan penelitian.
6. Departemen Agama RI dan Proyek Hibah Bersaing XII atas nama Dr. Ir. Utut Widyastuti, MSi yang telah membiayai penelitian ini.
7. Pak Adi, pak Erfan, Jaya, mbak Pepi, pak Mulya, pak Muzuni, mbak Niken dan rekan-rekan di laboratorium BIORIN PPSHB IPB serta rekan-rekan BUD DEPAG 2006 yang tidak dapat penulis sebutkan satu persatu, penulis mengucapkan terimakasih yang sebesar-besarnya atas bantuan, kerjasama dan dukungannya selama ini.
Secara khusus, penulis sampaikan rasa terimakasih yang tulus kepada suami tercinta Drs. Priyono, MSi atas segala pengorbanan baik moril maupun materiil, dorongan, kesabaran, pengertian dan doanya. Kepada ibunda tercinta dan bapak ibu mertua yang tiada mengenal lelah selalu mendoakan penulis sampai saat ini, penulis sampaikan terimakasih yang tiada batas. Untuk ananda Hanif Alfian Aliefananda dan Hafidz Fadlila Akbar, terimakasih atas pengorbanan dan perjuangannya untuk melewati hari-hari tanpa kehadiran penulis. Semoga Allah SWT senantiasa melimpahkan rahmat dan kasih sayang-Nya kepada mereka, Amien.
Akhirnya, penulis berharap semoga tulisan ini dapat bermanfaat dan dapat memberikan sumbangan bagi perkembangan kedelai di Indonesia.
Bogor, Agustus 2008
RIWAYAT HIDUP
Penulis dilahirkan di Sragen, Jawa Tengah pada tanggal 17 Agustus 1971, sebagai anak keempat dari empat bersaudara pasangan Ayah Darmopitoyo (Alm) dan Ibu Sayem. Penulis menikah dengan Drs. Priyono, MSi. dan dikaruniai dua putra Hanif Alfian Aliefananda dan Hafidz Fadlila Akbar.
Penulis menyelesaikan pendidikan sarjana di Universitas Muhammadiyah Surakarta, Fakultas Pendidikan Matematika dan Ilmu Pengetahuan Alam (FPMIPA) jurusan Biologi tahun 1994.
KULTIVAR SLAMET DAN LUMUT
SIH HARTINI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul ”Induksi Mutasi dengan Irradiasi Sinar Gamma pada Kedelai (Glycine max (L.) Merrill) Kultivar Slamet dan Lumut” merupakan gagasan dan karya saya bersama pembimbing yang belum pernah dipublikasikan dalam bentuk apapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan oleh penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir tesis ini.
Bogor, Agustus 2008
SIH HARTINI. (Induction of Mutation with Gamma Ray Irradiation at Slamet and Lumut Cultivars of Soybean (Glycine max (L.) Merrill)).Under direction of
UTUT WIDYASTUTI and MUHAMMAD JUSUF.
The aim of this research was to produce Gα mutants of two cultivars of Soybean using 60Co gamma irradiation with 0.0; 0.1; 0.2; 0.3; 0.4; and 0.5 kGy dosage. Two Soybean cultivars tested were Slamet and Lumut. The mutated Soybean were detected at M-1 and M-2 filials. M-1 was used for screening general mutation based on plant hight variable, and M-2 for Gα and stability mutations based on stomatal opening characteristic. The results indicated that irradiation at dosage of 0.1 – 0.5 kGy produced plant mutation in different characteristics such as big seeds, flower’s colour, leaf’s form and leaflets mutation of the two cultivars. In addition, 60Co gamma irradiation at the dosage of 0.1 dan 0.3 kGy produced 10 Gα mutants putatives. The mutant had 24-40 cm hight and closed stomata at both cultivars. Molecular analysis was run in one Gα gene mutant putative only. Based on molecular analysis showed that gamma irradiation at 0.3 kGy in Slamet cultivar caused 1 Gαgene mutant.
RINGKASAN
SIH HARTINI. (Induksi Mutasi dengan Irradiasi Sinar Gamma pada Kedelai (Glycine max (L.) Merrill) kultivar Slamet dan Lumut). Dibimbing oleh UTUT WIDYASTUTI dan MUHAMMAD JUSUF.
Kedelai merupakan bahan makanan penting karena dapat dikonsumsi secara langsung maupun digunakan sebagai bahan baku agroindustri, namun untuk memenuhi tingginya kebutuhan kedelai masih tergantung impor karena produksi nasional masih sangat rendah. Untuk memenuhi kebutuhan tersebut perlu dilakukan upaya peningkatan produksi. Peningkatan produksi bisa dilakukan dengan berbagai macam cara antara lain melalui usaha pemuliaan tanaman. Pemuliaan tanaman dapat dilakukan dengan merakit kultivar baru, dan keragaman merupakan modal dasar untuk merakit kultivar baru. Salah satu upaya peningkatan keragaman yaitu dengan induksi mutasi. Mutasi bisa dihasilkan oleh beberapa agen mutagenik seperti radiasi, non radiasi maupun kimia. Sumber irradiasi yang sering digunakan adalah sinar X, sinar gamma, ultra-violet, sinar beta dari radioisotop dan sinar neutron dari reaktor atom. Penelitian ini menggunakan sinar gamma dari 60Co.
Kedelai kultivar Lumut dan Slamet memiliki perbedaan dalam sistem toleransi terhadap aluminium. Kultivar Lumut merupakan kultivar yang peka terhadap cekaman aluminium sedangkan kultivar Slamet toleran. Gen Gα diduga berperan dalam sistem toleransi terhadap aluminium pada kedua kultivar tersebut. Untuk melihat keterlibatan fungsi gen Gα secara langsung dalam sistem pertahanan terhadap cekaman Al dapat dilakukan dengan pendekatan reverse genetic yaitu menonaktifkan gen Gα. Teknik yang digunakan untuk menonaktifkan gen tersebut adalah dengan induksi mutasi menggunakan irradiasi sinar gamma karena radiasi gamma mempunyai energi yang sangat besar sehingga mampu menembus jaringan tanaman dengan maksimal.
Tujuan penelitian ini adalah untuk memperoleh berbagai macam mutan termasuk mutan gen Gα pada kedelai kultivar Slamet dan Lumut yang diinduksi irradiasi sinar gamma. Untuk mendapatkan mutan tersebut dilakukan skrining, dengan menanam kedelai dalam dua periode tanam. Periode tanam I (M-1) mengikuti pola Rancangan Acak Kelompok (RAK) dengan 2 faktor (dosis irradiasi dan kultivar) dan 3 kelompok ulangan, merupakan tahapan seleksi terhadap berbagai macam mutan. Dosis irradiasi yang digunakan adalah 0; 0.1; 0.2; 0.3; 0.4; dan 0.5 kGy, kultivar yang digunakan adalah Lumut dan Slamet. Kedelai ditanam dalam petakan, tiap petak terdapat 100 lubang, masing-masing lubang berisi 2 biji kedelai. Mutan yang diperoleh selanjutnya ditanam kembali sebagai generasi M-2. Rancangan yang digunakan dalam periode tanam II adalah Rancangan Acak Lengkap (RAL). Benih ditanam dengan membentuk barisan yang bersambungan antara satu induk dengan induk yang lain. Tiap induk diambil 30 biji, namun untuk induk yang memiliki jumlah biji kurang dari 30, biji ditanam semua. Hasil seleksi periode tanam I, dari 657 induk ditanam sebanyak 10.216 tanaman.
Selanjutnya mutan tersebut diuji stabilitasnya pada generasi M-2. Mutan stabil ditunjukkan oleh tidak adanya segregasi pada generasi M-2. Mutan warna biji dan mutan motif daun belum stabil karena mengalami segregasi pada generasi M-2, sedangkan mutan lainnya sudah stabil tidak bersegregasi. Mutan pendek merupakan prioritas dalam penelitian ini karena merupakan indikator tanaman tersebut mengalami mutasi Gα. Selanjutnya mutan pendek tersebut diperiksa perilaku stomatanya. Mutan Gα ditunjukkan oleh perubahan pembukaan stomata dimana stomata cenderung menutup. Untuk melihat perilaku stomata diambil tigapuluh sampel mutan pendek stabil, dari tigapuluh sampel tersebut diperoleh sepuluh tanaman dengan kondisi stomata menutup. Selanjutnya tanaman yang memiliki stomata menutup dilihat ekspresi gen Gα nya untuk memastikan bahwa tanaman tersebut telah mengalami mutasi. Sepuluh tanaman yang diduga mutan
Gα berdasarkan perilaku stomata, baru diperiksa satu mutan Gα berdasarkan ekspresi gennya. Mutan tersebut berasal dari kultivar Slamet dosis irradiasi 0.3 kGy. Tanaman wild type (kontrol) menghasilkan pita dengan ukuran 1380 pb, sedangkan tanaman mutan tidak menghasilkan pita tersebut. Tidak munculnya pita pada mutan menunjukkan bahwa gen Gα tidak diekspresikan karena telah mengalami mutasi.
© Hak Cipta milik IPB, tahun 2008
Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah.
b. Pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini dalam bentuk apapun tanpa izin IPB.
KULTIVAR SLAMET DAN LUMUT
SIH HARTINI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Departemen Biologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Induksi Mutasi dengan Irradiasi Sinar Gamma pada Kedelai (Glycine max (L.) Merrill) Kultivar Slamet dan Lumut
Nama : Sih Hartini
NRP : G 351060081
Disetujui Komisi Pembimbing
Dr. Ir. Utut Widyastuti, MSi. Dr. Ir. Muhammad Jusuf Ketua Anggota
Diketahui
Ketua Program Studi Biologi Dekan Sekolah Pascasarjana
Dr. Ir. Dedy Duryadi S, DEA Prof. Dr. Ir. Khairil Anwar Notodiputro, MS.
limpahan rahmat-Nya sehingga penulis dapat menyelesaikan penelitian serta penyusunan tesis ini.
Penulis menyadari sepenuhnya bahwa sangatlah sulit untuk menyelesaikan penelitian dan penyusunan tesis ini tanpa bantuan dari berbagai pihak. Oleh karena itu dalam kesempatan ini penulis ingin menyampaikan ucapan terimakasih serta penghargaan yang setinggi-tingginya kepada:
1. Dr. Ir. Utut Widyastuti, MSi. selaku ketua komisi pembimbing atas semua jerih payah dan waktu yang telah diluangkan untuk memberikan bimbingan, arahan dan kemudahan kepada penulis dari mulai penelitian hingga terselesaikannya tesis ini.
2. Dr. Ir. Muhammad Jusuf selaku anggota komisi pembimbing atas ilmu, waktu dan bimbingan yang diberikan kepada penulis dengan penuh kesabaran sehingga tesis ini dapat terselesaikan.
3. Dr. Ir. Hajrial Aswidinnoor, MSc. selaku dosen penguji atas kritik dan sarannya untuk kesempurnaan tesis ini.
4. Dekan Sekolah Pascasarjana IPB dan Ketua Program Studi Biologi atas kesempatan yang diberikan kepada penulis untuk mengikuti pendidikan Pascasarjana di IPB.
5. Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) dan Laboratorium Anatomi Tumbuhan Departemen Biologi IPB yang telah menyediakan fasilitas untuk melakukan penelitian.
6. Departemen Agama RI dan Proyek Hibah Bersaing XII atas nama Dr. Ir. Utut Widyastuti, MSi yang telah membiayai penelitian ini.
7. Pak Adi, pak Erfan, Jaya, mbak Pepi, pak Mulya, pak Muzuni, mbak Niken dan rekan-rekan di laboratorium BIORIN PPSHB IPB serta rekan-rekan BUD DEPAG 2006 yang tidak dapat penulis sebutkan satu persatu, penulis mengucapkan terimakasih yang sebesar-besarnya atas bantuan, kerjasama dan dukungannya selama ini.
Secara khusus, penulis sampaikan rasa terimakasih yang tulus kepada suami tercinta Drs. Priyono, MSi atas segala pengorbanan baik moril maupun materiil, dorongan, kesabaran, pengertian dan doanya. Kepada ibunda tercinta dan bapak ibu mertua yang tiada mengenal lelah selalu mendoakan penulis sampai saat ini, penulis sampaikan terimakasih yang tiada batas. Untuk ananda Hanif Alfian Aliefananda dan Hafidz Fadlila Akbar, terimakasih atas pengorbanan dan perjuangannya untuk melewati hari-hari tanpa kehadiran penulis. Semoga Allah SWT senantiasa melimpahkan rahmat dan kasih sayang-Nya kepada mereka, Amien.
Akhirnya, penulis berharap semoga tulisan ini dapat bermanfaat dan dapat memberikan sumbangan bagi perkembangan kedelai di Indonesia.
Bogor, Agustus 2008
RIWAYAT HIDUP
Penulis dilahirkan di Sragen, Jawa Tengah pada tanggal 17 Agustus 1971, sebagai anak keempat dari empat bersaudara pasangan Ayah Darmopitoyo (Alm) dan Ibu Sayem. Penulis menikah dengan Drs. Priyono, MSi. dan dikaruniai dua putra Hanif Alfian Aliefananda dan Hafidz Fadlila Akbar.
Penulis menyelesaikan pendidikan sarjana di Universitas Muhammadiyah Surakarta, Fakultas Pendidikan Matematika dan Ilmu Pengetahuan Alam (FPMIPA) jurusan Biologi tahun 1994.
DAFTAR TABEL ... xii
DAFTAR GAMBAR ... xiii
DAFTAR LAMPIRAN ... xiv
PENDAHULUAN Latar Belakang ... 1
Tujuan Penelitian ... 3
Hipotesis Penelitian ... 3
TINJAUAN PUSTAKA Mutasi ... 4
Protein Heterotrimerik-G Subunit α... 7
BAHAN DAN METODE Waktu dan Tempat ... 10
Bahan Tanaman ... 10
Metode Penelitian ... 10
Radiasi benih... 11
Penanaman di Lapang ... 11
Observasi Lapang dan Deteksi Mutasi... 12
Preparasi Struktur Anatomi Stomata... 12
Ekspresi Gen Gα dengan Teknik PCR ... 12
Analisis Data ... 15
HASIL Analisis Diskriminan terhadap Tanaman M-1 ... 16
Tipe-Tipe Mutan pada Tanaman M-1 ... 19
Uji Stabilitas M-2 ... 25
Konfirmasi Mutasi Gα dengan Observasi Stomata ... 29
Studi Ekspresi Gen Gα ... 30
PEMBAHASAN Analisis Diskriminan terhadap Tanaman M-1 ... 31
Tipe-Tipe Mutan pada Tanaman M-1 dan Uji Stabilitas pada Tanaman M-2 ... 33
Konfirmasi Mutasi Gα dengan Observasi Stomata dan Ekspresi Gen ... 37
SIMPULAN DAN SARAN Simpulan ... 38
Saran ... 38
DAFTAR PUSTAKA ... 39
DAFTAR TABEL
Halaman
1 Hasil pengelompokan dua belas populasi dari
Analisis Diskriminan ... 16 2 Asal populasi dan karakter enam kelompok baru hasil
Analisis Diskriminan ... 18 3 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan
warna biji per seribu tanaman M-1 pada lima dosis irradiasi
kultivar Lumut dan Slamet ... 19 4 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan warna
bunga per seribu tanaman M-1 pada lima dosis irradiasi
kultivar Lumut dan Slamet ... 20 5 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan jumlah
anak daun per seribu tanaman M-1 pada lima dosis irradiasi
kultivar Lumut dan Slamet ... 21 6 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan
bentuk daun per seribu tanaman M-1 pada lima dosis irradiasi
kultivar Lumut dan Slamet ... 22 7 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan
motif daun per seribu tanaman M-1 pada lima dosis irradiasi
kultivar Lumut dan Slamet ... 23 8 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan pendek
per seribu tanaman M-1 pada lima dosis irradiasi kultivar Lumut
dan Slamet ... 25 9 Warna bunga M-1, jumlah keturunan dan segregasi warna bunga
M-2 dari tujuh induk mutan kultivar Lumut dan Slamet ... 26 10 Jumlah anak daun M-1, jumlah keturunan dan segregasi jumlah
anak daun M-2 dari sembilan induk kultivar Lumut dan Slamet ... 27 11 Bentuk daun M-1, jumlah keturunan dan segregasi bentuk daun
M-2 dari enam induk mutan kultivar Lumut dan Slamet ... 27 12 Jumlah tanaman M-1, frekuensi mutan pendek stabil terhadap
wild type dan induk per seribu tanaman M-1 ... 28 13 Tanaman yang diduga mutan gen Gα berdasarkan
1 Diagram alir tahapan penelitian ... 10 2 Variasi warna biji kedelai kultivar Lumut dan Slamet ... 19 3 Variasi warna bunga kedelai kultivar Lumut dan Slamet ... 20 4 Variasi jumlah anak daun kedelai kultivar Lumut dan Slamet ... 21 5 Variasi bentuk daun kedelai kultivar Lumut dan Slamet ... 22 6 Variasi motif daun kedelai kultivar Lumut dan Slamet ... 23 7 Perbandingan ukuran biji pada kultivar Lumut wild type
DAFTAR LAMPIRAN
Halaman
1 Uji keragaman antar populasi untuk tiap-tiap variabel ... 45 2 Segregasi jumlah anak daun pada M-2 dari 60 induk mutan... 46 3 Hasil uji beda nyata tanaman M-2 stabil pendek
Latar Belakang
Kedelai merupakan tanaman pangan yang sangat penting karena dapat dikonsumsi secara langsung maupun digunakan sebagai bahan baku agroindustri. Kebutuhan kedelai nasional terus meningkat seiring dengan peningkatan jumlah penduduk di Indonesia. Pada tahun 2007 total kebutuhan kedelai sebesar 2.01 juta ton. Namun produksi kedelai nasional masih sangat rendah. Produksi kedelai pada tahun 2006 sekitar 749.04 ribu ton biji kering, mengalami penurunan sekitar 59.32 ribu ton (7.34 persen) dibandingkan dengan produksi tahun 2005 sebesar 808.35 ribu ton (BPS 2006). Bahkan pada tahun 2007 produksi kedelai nasional hanya 0.61 juta ton. Berarti terdapat kekurangan jumlah produksi yang cukup besar untuk memenuhi permintaan pasar, sehingga pada tahun 2007 Indonesia mengimpor kedelai sampai 1.42 juta ton atau sekitar 70% dari total kebutuhan pada tahun tersebut.
diharapkan dapat meningkatkan keragaman genetik pada kedua kultivar kedelai Slamet dan Lumut.
Kedelai kultivar Slamet dan Lumut memiliki perbedaan dalam sistem toleransi terhadap aluminium (Al). Kedelai kultivar Lumut merupakan kultivar yang peka terhadap cekaman Al sedangkan kultivar Slamet toleran terhadap cekaman Al (Anwar 1999). Menurut Sunarlim dan Titis (2001), keracunan Al merupakan masalah utama yang sering dijumpai pada kedelai di lahan asam dan menyebabkan penurunan produktivitas kedelai. Sehingga perlu dilakukan pemuliaan untuk memperbaiki karakter tanaman yang bersifat peka terhadap Al. Pemuliaan untuk memperoleh tanaman kedelai yang mampu beradaptasi terhadap aluminium tinggi memerlukan informasi genetik tentang adaptasi terhadap masalah tersebut. Menurut Mashuda (2006) cekaman Al dapat meningkatkan ekspresi gen Gα pada kultivar Slamet, sedangkan Lumut tidak mengalami peningkatan (Sawitri 2007). Gen Gα diduga berperan dalam sistem toleransi terhadap Al pada kedua kultivar tersebut (Suharsono & Suharsono 2006).
Untuk melihat keterlibatan fungsi gen Gα secara langsung dalam sistem pertahanan terhadap cekaman Al dapat dilakukan dengan pendekatan reverse genetic yaitu menonaktifkan gen Gα. Sinar gamma memiliki energi yang sangat besar sehingga diharapkan dapat menonaktifkan gen Gα. Menurut Fujisawa et al. (1999) kehilangan subunit α pada tanaman padi (mutan daikoku d1) menyebabkan tanaman menjadi kerdil, daun pendek menebal berwarna gelap dan biji kecil, juga terjadi perubahan pembukaan stomata (Assmann 1996). Hal ini dapat digunakan sebagai indikator awal untuk menyeleksi tanaman yang diduga mengalami mutasi Gα.
(plantlet) serta menyebabkan penurunan tinggi plantlet krisan (Kendarini 2006). Irradiasi dengan dosis 1000 - 4000 rad menyebabkan nekrotik dan pertumbuhan kalus Costus speciosus KOEN SM. terhambat ( Toruan & Mathius 1991). Ratma (1988) juga melaporkan bahwa irradiasi gamma dosis 0.2 dan 0.4 kGy dapat menghasilkan mutan kedelai pendek.
Irradiasi gamma juga banyak diaplikasikan pada berbagai tanaman untuk memperoleh karakter-karakter yang diinginkan. Pada kacang tanah irradiasi gamma dapat menghasilkan keragaman jumlah polong dan biji (Dewi et al. 1993) dan pada dosis 0.3 kGy dapat meningkatkan keragaman tinggi tanaman kacang tanah serta keragaman genetik ketahanan terhadap penyakit layu (Dewi & Mugiono 1997). Pada tanaman kedelai, dengan dosis 0.10 dan 0.20 kGy dapat meningkatkan mutasi klorofil (Ratma & Sumanggono 1998). Irradiasi sinar gamma juga mengakibatkan perubahan bentuk bunga, warna bunga, kandungan klorofil dan anthosianin bunga Gerbera (Prasetyorini 1991).
Penelitian ini menggunakan induksi mutasi dengan irradiasi sinar gamma, selanjutnya diikuti dengan seleksi terhadap tanaman kerdil dan tanaman yang memiliki karakter berbeda dengan wild type (tipe tetua). Karakter tanaman kerdil menjadi prioritas dalam penelitian ini karena karakter tersebut merupakan salah satu indikator tanaman mengalami mutasi Gα. Tanaman yang memiliki karakter kerdil selanjutnya diperiksa perilaku stomata dan ekspresi gen Gα untuk memastikan bahwa tanaman tersebut mengalami mutasi pada gen Gα.
Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk memperoleh berbagai macam mutan termasuk mutan gen Gα pada kedelai kultivar Slamet dan Lumut yang diinduksi irradiasi sinar gamma.
Hipotesis Penelitian
TINJAUAN PUSTAKA
Mutasi
Mutasi adalah perubahan yang terjadi pada materi genetik sehingga menyebabkan perubahan ekspresi. Perubahan dapat terjadi pada tingkat pasangan basa, tingkat satu ruas DNA, bahkan pada tingkat kromosom (Jusuf 2001). Mutasi dapat terjadi pada setiap bagian tanaman, namun lebih banyak terjadi pada bagian yang sedang aktif mengadakan pembelahan sel. Jika mutasi terjadi pada sel somatik, maka perubahan hanya pada bagian itu dan tidak diwariskan. Sedang bila mutasi terjadi pada sel generatif, maka akan diwariskan pada generasi berikutnya (Poespodarsono 1986). Organisme baru hasil mutasi disebut mutan.
Mutasi atau perubahan materi genetik dapat dideteksi dengan melihat perubahan pada tingkat struktur gen atau perubahan pada tingkat ekspresinya. Untuk melihat perubahan tersebut dapat dilakukan dengan membandingkan antara mutan dan tipe liarnya. Perubahan dapat terlihat pada tingkat morfologi yang terlihat oleh mata telanjang, atau pada tingkat lain yang tidak nampak oleh mata. Secara garis besar penampilan mutan dapat dilihat dari liarnya dengan tiga cara; perbedaan morfologi, perbedaan tingkat kimia, dan perbedaan tingkat adaptasi terhadap lingkungan tumbuh. Hasil mutasi yang paling mudah dilihat ialah bila terjadi perubahan morfologi seperti bentuk, ukuran atau warna (Jusuf 2001).
lain. Insersi dan delesi merupakan penambahan atau pengurangan satu atau lebih pasangan nukleotida pada suatu gen (Griffiths et al. 2005; Campbell et al. 2002).
Mutasi kromosom dapat terjadi karena perubahan jumlah kromosom atau perubahan struktur kromosom. Perubahan struktur kromosom adalah perubahan dimana jumlah kromosom tetap tetapi terjadi perubahan komposisi dan susunan bahan kromosom, yaitu delesi, duplikasi, inversi dan translokasi. Sedangkan perubahan jumlah kromosom adalah adanya penambahan atau pengurangan kromosom-kromosom utuh atau satu set kromosom lengkap (genom), perubahan ini dapat menyebabkan keragaman genetik yang akan nampak pada keragaman fenotipe seperti sifat morfologi dan fisiologi (Crowder 1997).
Mutasi dapat terjadi dengan spontan (Djojosoebagio 1988) atau akibat adanya rangsangan dari luar. Mutasi spontan merupakan bagian dari proses kehidupan suatu spesies. Selama proses kehidupan akan terus terjadi perubahan spontan pada gen walaupun dengan tingkat kecepatan yang rendah (Poespodarsono 1986).
Mutasi spontan memiliki frekuensi yang sangat kecil sekitar 10-9 sampai 10-7. Rendahnya frekuensi mutasi spontan karena pada organisme ada sistem pemeliharaan ketepatan pemasangan basa-basa DNA yang melekat pada proses replikasi. Selain itu juga terdapat sejumlah mekanisme koreksi kerusakan basa. Mutasi spontan dapat disebabkan oleh perubahan tautomerik basa-basa DNA yaitu perubahan konfigurasi suatu molekul akibat perpindahan proton atau inti hidrogen dari satu posisi ke posisi lain. Selain perubahan tautomerik ada proses kimia lain yang dapat menyebabkan terjadinya mutasi spontan yaitu depurinasi dan deaminasi. Depurinasi adalah proses pemutusan ikatan antara basa purin dengan gula deoksiribosa, sedangkan deaminasi adalah penghilangan suatu gugus amino dari suatu basa (Jusuf 2001).
mutasi yang baik, atau perubahan genetik yang tidak baik (Broertjes & Harten 1988).
Induksi terhadap mutasi dapat terjadi secara alami maupun buatan. Mutasi buatan terjadi bila digunakan mutagen dengan dosis dan waktu tertentu (Poespodarsono 1986). Mutagen merupakan faktor penyebab terjadinya mutasi. Menurut Allard (1960) mutasi dihasilkan oleh beberapa agen mutagenik, yaitu proses mekanik murni, proses kimia murni atau kombinasi antara keduanya. Mutagen tersebut dapat menghasilkan berbagai macam mutasi seperti mutasi klorofil maupun mutasi kerdil. Sedangkan Poespodarsono (1986) membagi mutagen ke dalam tiga kelompok yakni radiasi, non radiasi dan kimia. Jusuf (2001) menyatakan bahwa bahan yang dapat merangsang mutasi dapat berupa bahan yang bersifat fisik, kimia atau proses biologis. Bahan fisik yang dikenal sebagai perangsang mutasi antara lain sinar ultraviolet, sinar X dan sinar gamma. Bahan kimia yang dapat merangsang terjadinya mutasi antara lain etilmetan sulfonat (EMS), etiletan sulfonat (EES), dan hidroksilamin (HA). Sedangkan bahan biologis, yang merupakan bahan mutakhir digunakan adalah elemen loncat.
Sinar gamma merupakan salah satu bahan fisik yang banyak digunakan sebagai agen mutasi. Radiasi sinar gamma merupakan radiasi ionisasi. Bentuk radiasi ini dapat menembus sel-sel dan jaringan dengan mudah (Pai 1999). Radiasi dengan sinar gamma dapat menghasilkan dua macam efek yaitu aberasi kromosom dan hambatan mitosis (Whitson 1972). Sinar gamma diperoleh dari peluruhan zat radioaktif yang dipancarkan dari atom dengan kecepatan tinggi karena kelebihan energi. Panjang gelombang sinar gamma lebih pendek dari sinar X tetapi energinya lebih besar. Radiasi sinar gamma dapat dipancarkan oleh 60Co,
137
Cs dan lain-lain (Soeminto 1985).
acute dapat menyebabkan sel mati atau mengalami perubahan sifat (Wiryosimin 1995).
Dosis irradiasi yang diterapkan tergantung pada sensitivitas dari spesies dan bagian tanaman. Sensitivitas tergantung pada volume inti (DNA yang lebih besar lebih sensitif), jumlah kromosom (tanaman dengan kromosom lebih sedikit dengan volume inti tertentu, lebih sensitif dari tanaman dengan kromosom yang lebih banyak), dan tingkat ploidi ( lebih tinggi, sensitivitasnya lebih sedikit) (Broertjes & Harten 1988).
Efektivitas irradiasi yang diberikan pada tanaman dipengaruhi oleh faktor lingkungan dan faktor biologi. Faktor lingkungan terdiri atas oksigen, kadar air, suhu, sedangkan faktor biologi meliputi volume inti, kromosom interfase, dan faktor genetik yaitu adanya perbedaan kepekaan terhadap irradiasi (Ismachin 1988).
Protein Heterotrimerik-G subunit α
Protein G merupakan anggota dari sebagian besar kelompok protein yang ditemukan pada semua eukariot. Protein ini terikat secara lemah pada sisi sitoplasmik membran, dan berfungsi sebagai saklar yang dapat dihidupkan dan dimatikan. Pada keadaan tidak aktif protein heterotrimerik berada dalam bentuk ikatan GDP dan akan aktif dengan merubah GDP ke bentuk GTP yang berikatan
pada subunit α (Campbell et al. 2002). GTP-subunit α akan terpisah dari subunit βγ dan keduanya akan berinteraksi dengan efektor yang berada di down stream
(bagian hilir). Penurunan aktivitas GTP-subunit α menjadi GDP-subunit α melalui aktivitas GTPase menyebabkan pembentukan kembali bentuk heterotrimerik tidak aktif (Kolle 1997; Gutkins 1998).
Protein G dibagi menjadi dua yaitu protein G kecil dan protein heterotrimerik-G (Ma et al. 1991; Patrick & Gilman 1998). Protein heterotrimerik G terdiri dari tiga sub unit yang berbeda yaitu sub unit α (40-45 kDa), sub unit β ( 34-36 kDa) dan sub unit γ (7-10 kDa) (Gotor et al. 1996).
Subunit Gα terdapat pada membran plasma tanaman Arabidopsis (Weiss et al. 1997) dan padi (Iwasaki et al. 1997), juga ditemukan di dalam retikulum endoplasma Arabidopsis dan tomat (Weiss et al. 1997; Aharon et al. 1998). Protein heterotrimerik-G diketahui berperan dalam regulasi dari influk kanal ion K+ pada sel penjaga (Wu & Assmann 1994). Subunit Gα diketahui dapat mengaktifkan kanal kalsium (Ca2+) pada membran plasma sehingga meningkatkan level Ca2+ di sitoplasma pada tomat (Aharon et al. 1998) juga meningkatkan level IP3 pada tanaman kedelai (Legendre et al. 1993) serta peningkatan Reactive
oxygene Species (ROS) H2O2 pada kultur sel kedelai (Legendre et al. 1992).
Berdasarkan analisis mutasi pada gen Gα (dwarf1), ternyata Gα terlibat di dalam perpanjangan batang dan pembentukan biji padi (Fujisawa et al. 2001). Regulasi pembukaan stomata (Assmann 1996) dan pemanjangan tabung polen pada bunga lily (Ma et al. 1999) juga melibatkan protein G. Fungsi protein Gα yang lain pada tanaman diantaranya dalam transduksi sinyal auksin (Fairley-Grenot & Assmann 1991) serta terlibat dalam induksi giberelin dari gen
α-amylase pada sel aleuron oat (Jones et al. 1998).
Protein heterotrimerik-G subunit α berperan penting dalam transduksi sinyal terhadap berbagai stimulus dari luar yang diterima oleh organisme. Transduksi sinyal merupakan tanda atau pesan yang mengubah stimulus atau sinyal menjadi bentuk lain dengan melibatkan urutan reaksi biokimia tertentu di dalam sel yang dilakukan oleh enzim dan berhubungan melalui second messenger
(Voet & Donald 1995).
Proses pensinyalan sel meliputi 3 tahapan yaitu : 1) Penerimaan sinyal; 2) Transduksi sinyal; 3) Respon seluler. Penerimaan sinyal merupakan pendeteksian sinyal yang datang dari luar sel oleh sel target. Sinyal dapat terdeteksi apabila terikat pada protein seluler, biasanya pada permukaan sel yang bersangkutan. Salah satu reseptor transmembran yang menerima sinyal dan meneruskannya ke sel adalah protein G. Pengikatan molekul sinyal mengubah protein reseptor, dan selanjutnya mengawali proses transduksi. Tahap transduksi ini mengubah sinyal menjadi bentuk yang dapat menimbulkan respon seluler spesifik (Campbell et al. 2002).
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan mulai bulan Maret 2007 sampai Mei 2008, bertempat di Pusat Aplikasi Isotop dan Radiasi BATAN Jakarta, Kebun Percobaan IPB Pagentongan Sindangbarang Bogor, Laboratorium Anatomi Tumbuhan Departemen Biologi dan Laboratorium BIORIN Pusat Penelitian Sumberdaya Hayati dan Bioteknologi, IPB.
Bahan Tanaman
Bahan tanaman yang digunakan adalah benih kedelai kultivar Slamet dan Lumut.
Metode Penelitian
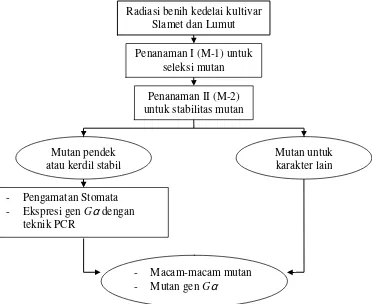
[image:34.612.151.523.383.687.2]Penelitian ini dilakukan dengan tahapan seperti pada diagram alir berikut (Gambar 1).
Gambar 1 Diagram alir tahapan penelitian. Radiasi benih kedelai kultivar
Slamet dan Lumut
Penanaman I (M-1) untuk seleksi mutan
Penanaman II (M-2) untuk stabilitas mutan
Mutan pendek atau kerdil stabil
Mutan untuk karakter lain
- Pengamatan Stomata - Ekspresi gen Gαdengan
teknik PCR
Radiasi Benih
Benih kedelai kultivar Slamet dan Lumut diradiasi dengan sinar gamma (60Co) dengan 6 taraf dosis yaitu : 0; 0.1; 0.2; 0.3; 0.4 dan 0.5 kGy. Pada setiap perlakuan diradiasi 200 biji kedelai. Kombinasi dari dosis irradiasi dengan kultivar membentuk 12 populasi yaitu populasi 1 (Lumut kontrol), populasi 2 (Lumut dosis irradiasi 0.1 kGy), populasi 3 (Lumut 0.2 kGy), populasi 4 (Lumut 0.3 kGy), populasi 5 (Lumut 0.4 kGy), populasi 6 (Lumut 0.5 kGy), populasi 7 (Slamet kontrol), populasi 8 (Slamet 0.1 kGy), populasi 9 (Slamet 0.2 kGy), populasi 10 (Slamet 0.3 kGy), populasi 11 (Slamet 0.4 kGy), dan populasi 12 (Slamet 0.5 kGy).
Penanaman di Lapang
Observasi Lapang dan Deteksi Mutasi
Karakter yang diamati pada tanaman M-1 meliputi tinggi tanaman, jumlah cabang, jumlah buku subur, jumlah buku total, jumlah polong isi, jumlah polong total, umur mulai berbunga, umur polong masak, warna polong, jumlah biji, berat total biji, berat 100 biji, warna biji, jumlah anak daun, motif daun, bentuk daun dan warna bunga. Mutasi dideteksi dari adanya perubahan pada karakter yang diamati. Mutasi pada gen Gα ditunjukkan oleh adanya batang yang pendek. Observasi pada tanaman M-2 diarahkan untuk melihat kestabilan karakter yang dianggap bermutasi yaitu melalui ada tidaknya segregasi pada karakter tersebut. Tanaman yang diduga mutan yang sudah stabil (kerdil) selanjutnya diperiksa struktur stomata dan ekspresi gen Gα nya.
Preparasi Struktur Anatomi Stomata
Pengamatan struktur anatomi stomata dengan menggunakan sediaan paradermal. Sediaan paradermal dibuat dalam bentuk sediaan semipermanen dengan metode Wholemount (Johansenn 1940). Pengamatan struktur anatomi stomata dilakukan terhadap tigapuluh sampel tanaman pendek atau kerdil dan wild type (kontrol). Pengambilan sampel dilakukan pada waktu dan kondisi yang sama (pukul 12.00 WIB). Bahan untuk pembuatan sediaan paradermal stomata adalah alkohol 70% (v/v), safranin 1% (b/v), pemutih (bayclin 5.25%) (v/v), gliserin 30% (v/v), cat kuku. Langkah-langkahnya; daun difiksasi dengan alkohol 70% (v/v), dicuci, kemudian disayat dengan silet. Hasil sayatan direndam dalam larutan pemutih (bayclin 5.25%) (v/v) selama sepuluh menit, sayatan dicuci kembali kemudian dilakukan pewarnaan menggunakan safranin 1% (b/v) selama sepuluh menit, sediaan diletakkan dalam gelas obyek, diberi media gliserin 30% (v/v) lalu ditutup dengan gelas penutup.
Ekspresi Gen Gα dengan Teknik PCR
Isolasi RNA Total. Isolasi RNA total dilakukan pada tanaman M-2 yang telah diseleksi. RNA diambil dari tanaman mutan dan wild type yang tidak diradiasi sebagai kontrol. Isolasi RNA total dengan menggunakan Kit Trizol (Invitrogen). Langkah isolasi RNA sebagai berikut : Daun sebanyak 500 – 1000 mg dari masing-masing perlakuan secara terpisah digerus di mortar dengan bantuan N2
cair sampai berbentuk tepung halus. Kemudian dicampur dengan 800 µl kit trizol. Campuran selanjutnya diinkubasi selama 5 menit pada suhu ruang kemudian ditambah dengan 200 µl chloroform dan dikocok selama 30 detik dan diinkubasi selama 3 menit dalam suhu ruang. Campuran disentrifugasi pada 9000 rpm (Jouan BR4i) selama 15 menit pada suhu 6 ºC. Cairan bagian atas diambil dan dipindahkan ke tabung baru. Selanjutnya ditambah dengan 500 µl isopropil alkohol dan diinkubasi 10 menit pada suhu ruang. Kemudian disentrifugasi pada 9000 rpm selama 10 menit pada 6 ºC. Endapan yang dihasilkan dicuci dengan penambahan 500 µl ethanol-DEPC 75% dan diikuti dengan sentrifugasi pada 5700 rpm selama 5 menit pada suhu 6 ºC. Endapan dikeringkan dengan vakum dryer selama 6 menit dan dilarutkan dalam 30 µl ddH2O-DEPC 0.1%.
Kuantifikasi dilakukan dengan melarutkan 1 µl larutan RNA total dalam 700 µl ddH2O-DEPC 0.1%, selanjutnya dibaca dengan spektrofotometer pada panjang
gelombang 260 nm dan 280 nm. Untuk mengetahui kemurnian RNA dari kontaminasi protein dilakukan dengan perhitungan nilai rasio OD260/280 (Sambrook et al. 1989). Keutuhan RNA dianalisis dengan memigrasikan RNA pada gel agarosa 1% dengan menggunakan buffer MOPS 1 X (4.2 g/l 3-Morpholinopropanesulfonic acid (C7H15NO4), 0.41 g/l Na-asetat, 0.37 g/l
Na2EDTA.H2O). 10 µg RNA total ditambah dengan 12 µl premix (20 X MOPS,
6 µl formamide, 2.1 µl formaldehide, 3.3 µl ddH2O-DEPC 0.1%) dipanaskan pada
suhu 65 ºC selama 10 menit. Selanjutnya diinkubasi dalam es selama 5 menit, kemudian ditambahkan 2 µl loading dye dengan komposisi bromofenol biru 0.25% (b/v), xylene cyanol 0.25% (b/v) dan sukrosa 15% (b/v). Campuran tersebut dimasukkan ke dalam sumur gel dan dimigrasikan selama 30 menit pada bak elektroforesis dengan tegangan 100 volt. Selanjutnya gel direndam di
dalam larutan ethidium bromida 0.5 µg/ml selama 10 menit, dibilas dengan akuades, kemudian pita RNA dilihat melalui UV transiluminator.
Sintesis cDNA. Sintesis cDNA dilakukan dengan mengikuti metode Suharsono
volume akhir 20 µl. Reaksi transkripsi balik (RT) dilakukan pada suhu 30 ºC 10 menit, 42 ºC 50 menit, 95 ºC 5 menit sebanyak satu siklus. Untuk mengetahui keberhasilan sintesis cDNA dilakukan amplifikasi cDNA melalui PCR dengan menggunakan primer aktin yang didesain dari kedelai (Accession V00450) dengan primer forward tepat pada kodon awal (5’ATGGCAGATGCCGAGG
ATAT 3’) dari ekson 1 dan primer reverse tepat pada daerah ekson 2 (5’ CAGTTGTGCGACCACTTGCA 3’) dan menggunakan cDNA sebagai
cetakan. PCR dilakukan dengan mencampur 1 µl hasil RT dengan 1 µl buffer Taq 10 X, 0.08 dNTPmix 25 mM, 0.4 µl DMSO, 1 unit enzim Taq polymerase, 10 pmol primer forward, 10 pmol primer reverse dan ddH2O sampai volume akhir 10 µl. Kondisi PCR untuk aktin adalah: denaturasi pra-PCR 95 ºC 5 menit, denaturasi pada 94 ºC 30 detik, penempelan primer pada 55 ºC 1 menit, pemanjangan DNA pada 72 ºC 1 menit 30 detik, siklus diulang sebanyak 35 kali, pemanjangan pasca PCR pada 72 ºC 5 menit dan proses pendinginan dilakukan pada 15 ºC 5 menit.
Analisis Ekspresi Gen Gαααα. Analisis ekspresi gen Gα dilakukan dengan RT-PCR menggunakan primer spesifik Gα. Primer didesain dari Soybean G protein α
subunit, SGA1 (Accession: L27418) dengan primer terletak pada 111 nukleotida sebelum kodon awal (5’ GCTTCACACTTCACACTTAACACT 3’) dan 114 sesudah stop kodon (5’ ATATTGTTGTATACCTGACCTC 3’) digunakan untuk mengamplifikasi cDNA dari gen Gα. PCR dilakukan dengan mencampur 1 µl hasil RT dengan 1 µl buffer Taq 10 X, 0.08 dNTPmix 25 mM, 0.4 µl DMSO, 1 unit enzim Taq polymerase, 10 pmol primer forward, 10 pmol primer reverse dan ddH2O sampai volume akhir 10 µl. Kondisi PCR untuk gen Gα adalah denaturasi pra-PCR 95 ºC 5 menit, denaturasi pada 94 ºC 30 detik, penempelan primer pada 58 ºC 1 menit, pemanjangan DNA pada 72 ºC 2 menit, siklus diulang sebanyak 37 kali, pemanjangan pasca PCR pada 72 ºC 5 menit dan proses pendinginan dilakukan pada 15 ºC 5 menit. Kehilangan gen Gα dilihat dengan membandingkan ekspresi antara mutan, tanaman wild type dan kontrol positif (Plasmid membawa gen Gα), yang ditandai dari ada tidaknya pita atau jumlahnya berubah menjadi lebih dari satu pita.
Analisis Data
Data morfologi dianalisis dengan Analisis Diskriminan untuk melihat kelompok mutan yang terbentuk. Kelompok yang dianggap mutan ialah yang memperlihatkan perbedaan dari wild type. Untuk melihat kestabilan mutan, populasi masing-masing mutan dibandingkan dengan populasi wild type dan induk, selanjutnya dilakukan uji beda nyata dengan uji T pada taraf nyata 5%.
HASIL
Analisis Diskriminan terhadap Tanaman M-1
Uji keragaman pada dua belas populasi yang diteliti menunjukkan adanya perbedaan yang nyata antar populasi (Lampiran 1). Keragaman tersebut terlihat pada semua karakter yang diamati, selain karakter warna polong.
Hasil analisis diskriminan menunjukkan adanya keragaman individu intra populasi yang cukup besar pada keduabelas populasi awal, sehingga terjadi tumpang tindih antar populasi. Masing-masing populasi menyebar ke kelompok populasi lain yang memiliki persamaan karakter (Tabel 1). Angka 1 sampai 12 pada tabel 1 merupakan kelompok hasil analisis diskriminan dengan jumlah anggota masing-masing. Populasi 1 (Lumut wild type) memiliki 464 anggota kelompok yang mirip Lumut dan 66 mirip populasi 2 (Lumut 0.1 kGy). Jika dilihat sebaran dari populasi 1, tampak bahwa populasi 1 terbagi menjadi dua kelompok. Populasi Lumut yang dikategorikan mutan adalah yang berada pada kelompok 3 sampai 12. Populasi 2 sampai 6 yang tergabung dalam kelompok 1 dan 2 tidak dikategorikan mutan karena masih mirip dengan populasi 1.
[image:40.612.124.517.466.696.2]
Tabel 1 Hasil pengelompokan dua belas populasi dari analisis diskriminan
Prediksi jumlah anggota kelompok hasil analisis diskriminan Populasi
Awal 1 2 3 4 5 6 7 8 9 10 11 12
Lumut kontrol (1) 464 66 0 0 0 0 0 0 0 0 0 0
Lumut 0,1 kGy (2) 48 267 179 0 0 0 1 3 0 3 7 0
Lumut 0,2 kGy (3) 22 102 278 0 0 1 0 2 1 0 2 0
Lumut 0,3 kGy (4) 1 12 4 112 44 1 0 0 0 0 1 0
Lumut 0,4 kGy (5) 0 5 5 41 44 0 0 0 0 0 3 0
Lumut 0,5 kGy (6) 0 0 1 1 7 1 0 0 0 0 0 0
Slamet kontrol (7) 1 0 0 0 0 0 512 11 2 0 0 0
Slamet 0,1 kGy (8) 0 10 2 0 0 3 68 193 160 21 0 8
Slamet 0,2 kGy (9) 0 2 0 0 0 1 55 144 254 18 4 8
Slamet 0,3 kGy (10) 0 3 1 10 2 2 1 23 49 88 42 54
Slamet 0,4 kGy (11) 0 0 1 0 0 0 0 0 0 1 9 1
Populasi 7 (Slamet wild type) tampak lebih seragam, hampir semua anggotanya mengelompok dalam kelompok 7 (97.3%). Populasi Slamet yang berada di luar kelompok 7 dikategorikan mutan. Populasi 8 sampai 12 yang masuk ke dalam kelompok 7 tidak dikategorikan mutan karena masih mirip dengan populasi 7.
Berdasarkan hasil analisis diskriminan terhadap dua belas populasi awal diperoleh enam kelompok baru, yaitu kelompok A, B, C, D, E dan F (Tabel 2). Masing-masing kelompok memiliki karakter yang berbeda-beda. Kelompok A dan kelompok D merupakan kelompok tanaman yang mempunyai sifat mirip dengan tanaman wild type Lumut dan Slamet dan sebagian besar berisi tanaman dari kedua kultivar tersebut. Kelompok B, C, E dan F merupakan kelompok yang menunjukkan perubahan karakter dari kelompok wild type, A dan D.
Tabel 2 Asal populasi dan karakter enam kelompok baru hasil analisis diskriminan
Kelompok Asal populasi Karakter
A (kontrol), 2 (62%), Populasi 1 dan 3 (30.4%)
Rata-rata: tinggi 67 cm, jumlah cabang 4, buku subur 13, buku total 14, polong isi 66, polong total 67, umur polong masak 94 hari, umur mulai bunga 42 hari, jumlah biji 134, berat total 9.9 g, berat 100 biji 7.8 g, warna biji hijau kekuningan, jumlah anak daun 3 (trifoliate), motif daun polos, bentuk daun bulat telur, warna bunga ungu.
B
Populasi 2 (32.5%), populasi
3 (68.1%) dan sebagian kecil populasi lainnya
Tanaman lebih pendek (rata-rata 58.9 cm).Terjadi penurunan pada jumlah cabang, buku subur, buku total, polong isi, polong total, jumlah biji serta berat total. Umur mulai bunga dan umur polong masak lebih lama, dan terjadi perubahan pada karakter jumlah anak daun. Ukuran biji relatif lebih besar (9.1 g/100 biji).
C (89.7%), 5 (86.7%) Populasi 4 dan 6 (90%)
Tanaman pendek (rata-rata 42.5 cm). Terjadi penurunan pada jumlah cabang, buku subur dan buku total, polong isi dan polong total, jumlah biji serta berat total,umur mulai bunga dan umur polong masak menjadi lebih lama, dan terjadi perubahan karakter warna biji, jumlah anak daun, dan bentuk daun. Rata-rata ukuran biji lebih besar ( 8.8 g/100 biji). Namun pada kelompok ini prosentase kematian sangat tinggi, terutama pada populasi 5 dan 6.
D Populasi 7 (kontrol), 8 (14.6%) dan populasi 9 (11.3%).
Rata-rata: tinggi 60.5 cm, jumlah cabang 4, buku subur 12, buku total 14, polong isi 55, polong total 56, umur polong masak 88 hari, umur mulai bunga 37 hari, jumlah biji 115, berat total 10.5 g, berat 100 biji 9.2 g, warna biji kuning, jumlah anak daun 3 (trifoliate), motif daun polos, bentuk daun bulat telur, warna bunga ungu.
E Populasi 8 (80.4%), 9 (85.6%), 10 (58.2%), dan populasi 12 (66.7%) serta sebagian kecil populasi 11
Tanaman lebih pendek (rata-rata 52.7 cm), Terjadi penurunan pada jumlah cabang, buku subur dan buku total, polong isi dan polong total, jumlah biji serta berat total. Rata-rata ukuran biji lebih besar (11.1 g/100 biji). Umur mulai bunga dan umur polong masak menjadi lebih lama, dan terjadi perubahan karakter warna biji, jumlah anak daun dan warna bunga.
F Populasi 11 (83.3%), 10 (34.9%), 12 (33.3%), serta sebagian kecil populasi 8 dan 9.
Tanaman pendek (rata-rata 39.3 cm), terjadi penurunan pada jumlah cabang, buku subur dan buku total, polong isi dan polong total, jumlah biji serta berat total. Rata-rata ukuran biji lebih besar (9.9 g/100 biji). Umur mulai bunga dan umur polong masak menjadi lebih lama, terjadi perubahan pada warna biji, jumlah anak daun, motif daun, dan warna bunga.
[image:42.612.131.530.103.646.2]
Tipe-Tipe Mutan pada Tanaman M-1
Mutan Warna Biji
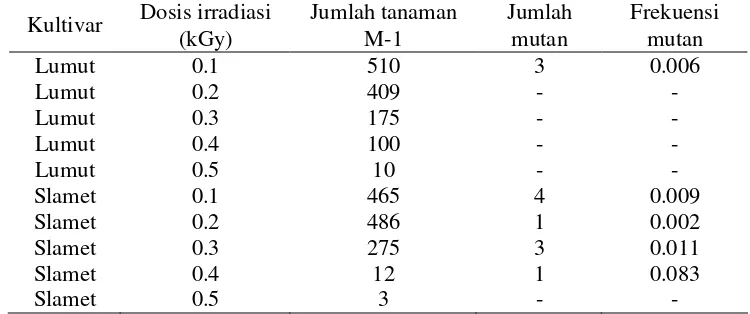
[image:43.612.181.464.317.443.2]Kulit biji kedelai umumnya berwarna coklat, kuning, atau hitam atau kombinasi dari warna tersebut. Kedelai kultivar Lumut memiliki warna biji hijau kekuningan, sedangkan Slamet memiliki warna biji kuning. Irradiasi gamma menyebabkan perubahan warna biji kedua kultivar ini. Pada kultivar Lumut warna biji menjadi adalah kuning, kuning kecoklatan dan hijau kehitaman, sedangkan kultivar Slamet meliputi coklat, krem dan kuning kehijauan (Gambar 2). Mutasi warna biji pada kultivar lumut hanya dihasilkan pada dosis 0.1 kGy, sedangkan Slamet pada dosis 0.1 sampai 0.4 kGy dengan frekuensi yang berbeda-beda (Tabel 3).
Lumut Slamet
Gambar 2 Variasi warna biji kedelai kultivar Lumut dan Slamet. (1) Kuning kehijauan, (2) Kuning, (3) Kuning kecoklatan, (4) Hijau kehitaman, (5) Kuning, (6) Coklat, (7) Krem, (8) Kuning kehijauan.
Tabel 3 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan warna biji per seribu tanaman M-1 pada lima dosis irradiasi kultivar Lumut dan Slamet
Kultivar Dosis irradiasi (kGy)
Jumlah tanaman M-1
Jumlah mutan
Frekuensi mutan
Lumut 0.1 510 3 0.006
Lumut 0.2 409 - -
Lumut 0.3 175 - -
Lumut 0.4 100 - -
Lumut 0.5 10 - -
Slamet 0.1 465 4 0.009
Slamet 0.2 486 1 0.002
Slamet 0.3 275 3 0.011
Slamet 0.4 12 1 0.083
Slamet 0.5 3 - -
1 2
4 3
5 6
8 7
[image:43.612.132.507.555.712.2]Mutan Warna Bunga
Kedelai umumnya memiliki warna bunga ungu atau putih. Kedelai kultivar Lumut dan Slamet memiliki warna bunga ungu. Irradiasi sinar gamma menyebabkan perubahan warna bunga kedelai pada kedua kultivar tersebut. Perubahan warna yang dihasilkan adalah ungu muda dan putih (Gambar 3). Kultivar Slamet memiliki frekuensi mutasi warna bunga lebih tinggi dibandingkan Lumut (Tabel 4). Mutan warna bunga hanya dihasilkan pada Lumut dosis 0.1, 0.2 dan 0.4 kGy, sedangkan Slamet dihasilkan pada dosis 0.1 sampai 0.4 kGy.
1 2 3
Gambar 3 Variasi warna bunga kedelai kultivar Lumut dan Slamet. (1) Warna bunga tipe liar Lumut dan Slamet, (2) Bunga ungu muda pada kultivar Lumut dan Slamet, (3) Bunga putih pada kultivar Lumut.
Tabel 4 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan warna bunga per seribu tanaman M-1 pada lima dosis irradiasi kultivar Lumut dan Slamet
Kultivar Dosis irradiasi (kGy)
Jumlah tanaman M-1
Jumlah mutan
Frekuensi mutan
Lumut 0.1 510 1 0.002
Lumut 0.2 409 1 0.002
Lumut 0.3 175 - -
Lumut 0.4 100 1 0.010
Lumut 0.5 10 - -
Slamet 0.1 465 1 0.002
Slamet 0.2 486 1 0.002
Slamet 0.3 275 1 0.004
Slamet 0.4 12 1 0.083
Slamet 0.5 3 - -
[image:44.612.130.509.242.376.2] [image:44.612.141.498.474.665.2]Mutan Jumlah Anak Daun
Kedelai umumnya memiliki jumlah anak daun 3 (trifoliate). Kedelai kultivar Lumut dan Slamet yang diradiasi gamma memiliki jumlah anak daun bervariasi. Variasi jumlah anak daun antara lain: kombinasi antara jumlah anak daun 2 dan 3, kombinasi 3 dan 4, kombinasi 3 dan 5, kombinasi 1, 2 dan 3, kombinasi 2, 3 dan 4, kombinasi 2, 3 dan 5, kombinasi 3, 4 dan 5 serta kombinasi 2, 3, 4 dan 5 (Gambar 4).
Gambar 4 Variasi jumlah anak daun kedelai kultivar Lumut dan Slamet. (1) Jumlah anak daun satu, (2) Jumlah anak daun dua, (3) Jumlah anak daun tiga (wild type), (4) Jumlah anak daun empat, (5) Jumlah anak daun lima.
Tabel 5 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan jumlah anak daun per seribu tanaman M-1 pada lima dosis irradiasi kultivar Lumut dan Slamet
Kultivar Dosis irradiasi (kGy)
Jumlah tanaman M-1
Jumlah mutan
Frekuensi mutan
Lumut 0.1 510 10 0.020
Lumut 0.2 409 12 0.029
Lumut 0.3 175 13 0.074
Lumut 0.4 100 4 0.040
Lumut 0.5 10 - -
Slamet 0.1 465 7 0.015
Slamet 0.2 486 5 0.010
Slamet 0.3 275 14 0.051
Slamet 0.4 12 - -
Slamet 0.5 3 - -
[image:45.612.242.406.225.334.2]Frekuensi mutan jumlah anak daun tertinggi terjadi pada kedua kultivar untuk dosis 0.3 kGy. Kultivar Lumut dosis 0.5 kGy tidak menghasilkan mutan jumlah anak daun, demikian pula pada Slamet dosis 0.4 dan 0.5 kGy (Tabel 5).
Mutan Bentuk Daun
Kedelai secara umum memiliki bentuk daun bulat telur sampai lancip. Irradiasi gamma menyebabkan variasi bentuk daun bulat dan memanjang (Gambar 5). Frekuensi mutan bentuk daun tertinggi terdapat pada kedelai kultivar Lumut dosis irradiasi 0.5 kGy. Pada kultivar ini, mutasi bentuk daun hanya terjadi pada dosis 0.3 dan 0.5 kGy. Mutasi bentuk daun pada kultivar Slamet dihasilkan oleh dosis 0.1 sampai 0.3 kGy, sedangkan pada dosis 0.4 dan 0.5 kGy tidak menghasilkan mutan bentuk daun (Tabel 6).
Gambar 5 Variasi bentuk daun kedelai kultivar Lumut dan Slamet. (1) Bentuk daun bulat telur (wild type), (2) Daun memanjang, (3) Daun bulat.
Tabel 6 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan bentuk daun per seribu tanaman M-1 pada lima dosis irradiasi kultivar Lumut dan Slamet
Kultivar Dosis irradiasi (kGy)
Jumlah
tanaman M-1 Jumlah mutan
Frekuensi mutan
Lumut 0.1 510 - -
Lumut 0.2 409 - -
Lumut 0.3 175 1 0.006
Lumut 0.4 100 - -
Lumut 0.5 10 1 0.100
Slamet 0.1 465 2 0.004
Slamet 0.2 486 1 0.002
Slamet 0.3 275 2 0.007
Slamet 0.4 12 - -
Slamet 0.5 3 - -
1 2 3
5 cm
[image:46.612.251.393.318.418.2]Mutan Motif Daun
Daun kedelai memiliki motif atau tekstur polos. Induksi dengan irradiasi sinar gamma menyebabkan daun kedelai menjadi bervariasi motifnya. Variasi motif daun tersebut antara lain : pinggiran putih, totol-totol tua muda, keriput, dan tebal gelap (Gambar 6).
Gambar 6 Variasi motif daun kedelai kultivar Lumut dan Slamet. (1) Polos (wild type), (2) Daun keriput, (3) Daun totol tua muda, (4) Daun pinggiran putih, dan (5) Daun tebal gelap.
Tabel 7 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan motif daun per seribu tanaman M-1 pada lima dosis irradiasi kultivar Lumut dan Slamet
Kultivar Dosis irradiasi (kGy)
Jumlah tanaman M-1
Jumlah mutan
Frekuensi mutan
Lumut 0.1 510 6 0.012
Lumut 0.2 409 1 0.002
Lumut 0.3 175 1 0.006
Lumut 0.4 100 1 0.010
Lumut 0.5 10 1 0.100
Slamet 0.1 465 - -
Slamet 0.2 486 - -
Slamet 0.3 275 2 0.007
Slamet 0.4 12 - -
Slamet 0.5 3 - -
1 3
4 5
[image:47.612.183.473.167.401.2]Frekuensi mutan motif daun bervariasi pada berbagai dosis irradiasi yang diberikan. Mutasi tipe ini lebih banyak terjadi pada kultivar Lumut dibandingkan Slamet. Pada kultivar Lumut dihasilkan mutan motif daun pada kelima dosis irradiasi, namun kultivar Slamet hanya dihasilkan pada dosis 0.3 kGy. Frekuensi mutan tertinggi terjadi pada kedelai kultivar Lumut dosis 0.5 kGy (Tabel 7).
Mutan Biji Besar
[image:48.612.260.388.333.440.2]Ukuran kedelai kultivar Lumut tergolong kecil bobot 100 biji kurang dari 10 gram, kultivar Slamet memiliki ukuran biji sedang (12.5 g/100 biji). Pada tanaman M-1 dari kedua kultivar diperoleh sebelas mutan biji besar dengan bobot 100 biji antara 16 gram sampai 19 gram. Pada kultivar Lumut terdapat mutan yang menghasilkan ukuran biji yang besar (Gambar 7).
Gambar 7 Perbandingan ukuran biji pada kultivar Lumut wild type dengan mutan.
Mutan Pendek
Kultivar Lumut memiliki rata-rata tinggi 75 cm, sedangkan Slamet rata-rata 60 cm. Akibat irradiasi gamma terjadi perubahan karakter pada tinggi tanaman dimana tanaman menjadi lebih pendek (Gambar 8). Jumlah dan frekuensi mutan pada masing-masing kultivar tidak sama (Tabel 8). Mutan pendek lebih banyak dihasilkan pada dosis irradiasi 0.3 sampai 0.5 kGy pada kedua kultivar, namun frekuensi tertinggi dihasilkan pada dosis 0.4 kGy untuk kultivar Lumut dan 0.3 kGy untuk kultivar Slamet.
Lumut biji besar Lumut wild type
Lumut Slamet
Gambar 8 Mutan pendek pada kedelai kultivar Lumut dan Slamet. (1) Lumut
wild type, (2) Lumut pendek, (3) Slamet wild type, (4) Slamet pendek.
Tabel 8 Jumlah tanaman M-1, jumlah mutan dan frekuensi mutan pendek per seribu tanaman M-1 pada lima dosis irradiasi kultivar Lumut dan Slamet
Kultivar Dosis irradiasi (kGy)
Jumlah tanaman M-1
Jumlah mutan
Frekuensi mutan
Lumut 0.1 510 79 0.155
Lumut 0.2 409 103 0.252
Lumut 0.3 175 91 0.520
Lumut 0.4 100 60 0.600
Lumut 0.5 10 5 0.500
Slamet 0.1 465 90 0.194
Slamet 0.2 486 78 0.160
Slamet 0.3 275 120 0.436
Slamet 0.4 12 5 0.417
Slamet 0.5 3 1 0.333
Uji Stabilitas M-2
Untuk uji stabilitas mutan dilakukan penanaman kembali tanaman mutan sebagai generasi M-2. Selanjutnya diperiksa apakah terjadi segregasi atau tidak. Mutan yang stabil diindikasikan oleh tidak adanya segregasi pada sifat yang
1 2 3 4
5 cm
termutasi. Pada tipe mutan M-1, karakter yang mengalami segregasi pada semua generasi M-2 adalah tipe mutan warna biji dan motif daun. Pada tipe mutan lainnya yaitu mutan warna bunga, mutan jumlah anak daun, mutan bentuk daun, mutan biji besar dan mutan pendek ada yang mengalami segregasi ada yang tidak.
Perubahan Warna Bunga
Hasil pengamatan segregasi terhadap mutan warna bunga menunjukkan bahwa sifat bunga ungu muda mengalami segregasi sedangkan bunga putih tidak (Tabel 9). Adanya segregasi menunjukkan bahwa tidak terjadi mutasi. Pada kedelai yang memiliki warna bunga putih menunjukkan bahwa telah terjadi mutasi pada tanaman tersebut dan mutasinya stabil sehingga generasi berikutnya akan memiliki karakter yang tetap.
Tabel 9 Warna bunga M-1, jumlah keturunan dan segregasi warna bunga M-2 dari tujuh induk mutan kultivar Lumut dan Slamet
Segregasi warna bunga M-2 Induk Asal induk Warna bunga
M-1
Jumlah
keturunan Ungu Ungu muda Putih
1 Lumut 0.1 kGy Putih 3 - - 3
2 Lumut 0.2 kGy Ungu muda 6 5 1 -
3 Lumut 0.4 kGy Putih 16 - - 16
4 Slamet 0.1 kGy Ungu muda 25 6 19 -
5 Slamet 0.2 kGy Ungu muda 2 1 1 -
6 Slamet 0.3 kGy Ungu muda 29 11 18 -
7 Slamet 0.4 kGy Ungu muda 9 8 1 -
Perubahan Jumlah Anak Daun
Setelah dilakukan pengamatan segregasi terhadap enam puluh mutan jumlah anak daun (Lampiran 2) diperoleh hasil bahwa terdapat sembilan tanaman yang stabil mengalami perubahan karakter jumlah anak daun (Tabel 10). Hal ini ditunjukkan oleh tidak adanya segregasi pada karakter tersebut. Tanaman wild type (kontrol) memiliki jumlah anak daun tiga (trifoliate), sedangkan pada tanaman mutan ditemukan adanya variasi jumlah anak daun 1, 2, 3, 4, dan 5.
Tabel 10 Jumlah anak daun M-1, jumlah keturunan dan segregasi jumlah anak daun M-2 dari sembilan induk mutan kultivar Lumut dan Slamet
Segregasi jumlah anak daun M-2 Induk Asal induk Jumlah anak daun
M-1
Jumlah keturunan
(M-2) Trifoliat
(3) Variasi
1 Lumut 0.1 kGy Variasi 2 dan 3 1 - 1
2 Lumut 0.1 kGy Variasi 3 dan 4 2 - 2
3 Lumut 0.2 kGy Variasi 3 dan 4 1 - 1
4 Lumut 0.2 kGy Variasi 2 dan 3 1 - 1
5 Lumut 0.2 kGy Variasi 4 dan 5 1 - 1
6 Lumut 0.3 kGy Variasi 3 dan 4 1 - 1
7 Lumut 0.3 kGy Variasi 3 dan 4 1 - 1
8 Lumut 0.3 kGy Variasi 3, 4 dan 5 1 - 1
9 Slamet 0.2 kGy Variasi 3 dan 5 12 - 12
Perubahan Bentuk Daun
Hasil pengamatan segregasi terhadap mutan bentuk daun diperoleh hasil bahwa pada kultivar Lumut dosis irradiasi 0.3 dan 0.5 kGy tidak terjadi segregasi, sehingga karakter bentuk daun pada tanaman ini sudah stabil. Semua mutan pada M-1 kultivar Slamet mengalami segregasi antara bentuk bulat telur dengan bentuk bulat dan memanjang (Tabel 11), jadi perubahan bentuk daun pada kultivar Slamet tersebut bukan mutasi.
Tabel 11 Bentuk daun M-1, jumlah keturunan dan segregasi bentuk daun M-2 dari enam induk mutan kultivar Lumut dan Slamet
Segregasi bentuk daun M-2 Induk Asal induk Bentuk daun
M-1
Jumlah keturunan
(M-2)
Bulat
telur Bulat Memanjang
1 Lumut 0.3 kGy Bulat 1 - 1 -
2 Lumut 0.5 kGy Memanjang 1 - - 1
3 Slamet 0.1 kGy Memanjang 12 10 - 2
4 Slamet 0.1 kGy Bulat 25 1 24 -
5 Slamet 0.3 kGy Memanjang 7 4 - 3
Perubahan Ukuran Biji
Hasil pengamatan segregasi terhadap mutan biji besar diperoleh hasil dua mutan biji besar dengan bobot 15.92 gram/100 biji dan 19.64 gram/100 biji pada kultivar Lumut. Mutan ini dihasilkan dari dosis irradiasi 0.1 dan 0.2 kGy. Perubahan karakter ukuran biji pada kultivar Slamet belum stabil sehingga tidak terwariskan ke generasi M-2.
Perubahan Tinggi Batang
Dari pengamatan segregasi terhadap mutan pendek, ternyata tidak semua karakter pendek bersifat stabil. Untuk melihat kestabilan mutan pendek dilakukan uji T terhadap masing-masing populasi tanaman pendek dibandingkan dengan populasi wild type yang ditanam pada periode tanam dua dan induknya (M-1). Untuk mengetahui ada tidaknya segregasi diperlukan perbandingan nilai tengah antara mutan dan induk. Hasil uji T untuk kontrol Slamet terhadap induknya menunjukkan bahwa nilai tengah tanaman berbeda nyata terhadap induk dimana tanaman menjadi lebih tinggi (Lampiran 3). Oleh karena itu pengambilan sampel mutan pendek untuk deteksi stomata dan ekspresi gen diambil dari tanaman M-2 yang berbeda nyata terhadap kontrolnya saja. Frekuensi mutan pendek stabil dijumpai lebih banyak pada kultivar Lumut dar