IDENTIFIKASI BEGOMOVIRUS INDONESIA PADA TOMAT
DAN ANALISIS DIVERSITAS GENETIK GEN AV1 SERTA
PEMANFAATANNYA UNTUK PENGEMBANGAN TANAMAN
TAHAN VIRUS
TRI JOKO SANTOSO
`
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa
Identifikasi
Begomovirus
Indonesia Pada Tomat Dan Analisis Diversitas Genetik Gen
AV1
serta
Pemanfaatannya Untuk Pengembangan Tanaman Tahan Virus
adalah karya saya dengan arahan komisi pembimbing dan belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.Bogor, November 2008
Tri Joko Santoso
ABSTRACT
TRI JOKO SANTOSO. Identification of Indonesian Begomoviruses in Tomato and Genetic Diversity Analysis of AV1 Gene as well Its Use for Developing Virus Resistant Plant. Under directions of SUDARSONO, HAJRIAL ASWIDINNOOR, SRI HENDRASTUTI HIDAYAT, and MUHAMMAD HERMAN.
transgenic tobacco plants and the accumulation of the virus in the transgenic plants tissue. Result of conventional breeding showed that F1-doublecross plants (crossing between F1-TYLCV and F1-CMV plants) revealed a resistant phenotype indicating integration of both two resistance genes in one plant has been occured following effication and PCR analysis. The resistant-doublecross F1 plants then were selected for the horticultural traits and subjected to performing the advanced breeding for developing Indonesian multiple virus resistance tomatoes.
RINGKASAN
TRI JOKO SANTOSO. Identifikasi Begomovirus Indonesia Pada Tomat dan Analisis Diversitas Genetik Gen AV1 serta Pemanfaatannya Untuk Pengembangan Tanaman Tahan Virus. Dibimbing oleh SUDARSONO, HAJRIAL ASWIDINNOOR, SRI HENDRASTUTI HIDAYAT, dan MUHAMMAD HERMAN.
Tomat (Lycopersicon esculentum, Mill) merupakan salah satu tanaman sayuran yang penting di Indonesia, baik secara ekonomi atau kandungan nutrisinya. Produksi tomat sangat dipengaruhi oleh penyakit keriting daun yang disebabkan oleh Tomato (yellow) leaf curl virus (TYLCV/ToLCV), salah satu anggota dari genus Begomovirus, famili Geminiviridae. Pada saat ini, belum ada cara yang secara efektif mengendalikan penyakit ini. Penggunaan tanaman tomat tahan merupakan cara yang terbaik untuk mengendalikan Begomovirus. Teknik rekayasa genetik memberikan peluang untuk mengembangkan tomat transgenik tahan terhadap Begomovirus melalui pendekatan ketahanan yang berasal dari patogen (PDR). Gen AV1 dari Begomovirus merupakan gen yang mengekspresikan protein selubung yaitu suatu protein yang bertanggung jawab dalam enkapsidasi partikel virus dan berperan di dalam penentuan spesifisitas penularan virus dan perkembangan gejala. Tujuan penelitian ini adalah (1) untuk mendeteksi Begomovirus yang menginfeksi tanaman tomat pada beberapa daerah area produksi tomat di Jawa Timur, Jawa Tengah, Daerah Istimewa Jogjakarta dan Jawa Barat menggunakan teknik PCR, (2) mempelajari keragaman genetik isolat-isolat Begomovirus yang menginfeksi tomat dari beberapa area produksi di Indonesia berdasarkan teknik PCR-RFLP, (3) mengidentifikasi dan menganalisis diversitas Begomovirus yang berasosiasi dengan penyakit keriting daun pada tomat berdasarkan sekuen asam nukleat dan asam amino prediksi dari gen AV1, (4) untuk mendapatkan konstruksi gen AV1 pada vektor ekspresi dan transforman tembakau hasil transformasi genetik dengan gen AV1 menggunakan vektor bakteri A. tumefaciens, (5) untuk mendapatkan tanaman tembakau transgenik yang membawa gen AV1 dan tahan terhadap Begomovirus, (6) untuk mendapatkan galur-galur tanaman tomat yang tahan terhadap Begomovirus (TYLCV) yang dikombinasikan dengan ketahanan terhadap CMV.
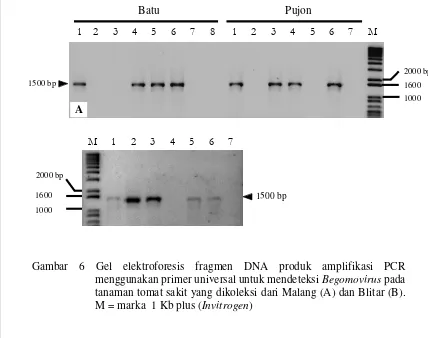
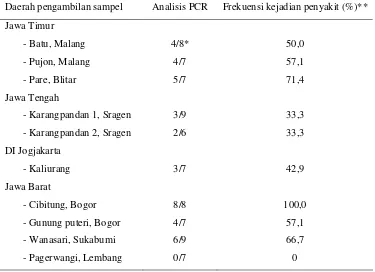
Deteksi Begomovirus dilakukan dengan mengamplifikasi genom Begomovirus dengan teknik PCR menggunakan sepasang primer degenerate yang universal (top primer) untuk Begomovirus yaitu primer PAL1v1978-F dan PAR1c715-R. Hasil percobaan menunjukkan bahwa sampel-sampel tanaman tomat sakit yang dikoleksi dari beberapa daerah di Jawa Timur, Jawa Tengah, Daerah Istimewa Jogjakarta dan Jawa Barat mengindikasikan adanya infeksi oleh Begomovirus setelah dideteksi menggunakan teknik PCR dengan primer universal. Infeksi Begomovirus ditunjukkan oleh adanya pita DNA hasil amplifikasi PCR yang berukuran 1500 bp. Frekuensi kejadian penyakit yang berasosiasi dengan Begomovirus bervariasi antara 0-100%. Frekuensi kejadian penyakit tertinggi (100%) terjadi di daerah Cibitung (Bogor) dan terendah (0%) terjadi di Pagerwangi (Lembang).
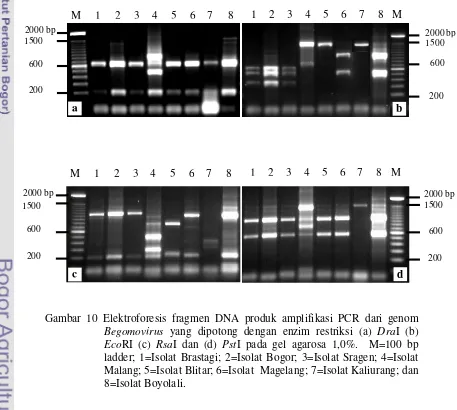
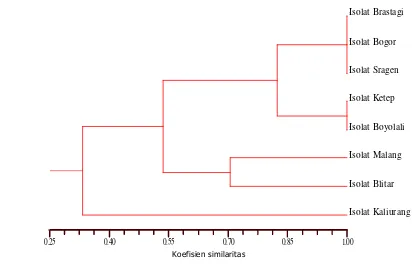
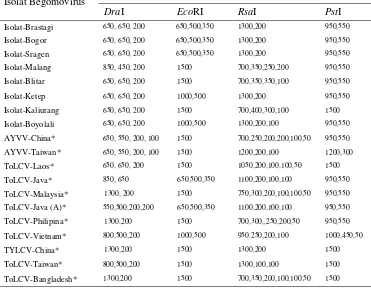
primer universal untuk Begomovirus dipotong dengan menggunakan empat macam enzim restriksi, yaitu DraI, EcoRI, RsaI dan PstI untuk melihat keragaman genetiknya. Pola pemotongan dengan enzim restriksi dari delapan isolat Begomovirus dan fragmen RFLP prediksi isolat Begomovirus dari DNA database GenBank digunakan untuk menentukan identitas genetik dan keragaman di antara isolat-isolat Begomovirus tersebut. Produk amplifikasi PCR yang dipotong dengan empat macam enzim restriksi mengindikasikan bahwa ada polimorfisme dari fragmen-fragmen DNA di antara 8 isolat Begomovirus yang berasal dari daerah-daerah di Jawa dan Sumatera tersebut. Analisis filogenetik menunjukkan bahwa isolat-isolat Begomovirus terbagi menjadi 3 kelompok yang berbeda. Isolat-isolat Brastagi, Bogor, Sragen, Ketep dan Boyolali berkerabat dekat dengan Tomato Leaf Curl Virus-Java (ToLCV-Java) atau ToLCV-Java (A), isolat Malang dan Blitar berkerabat dekat dengan Ageratum Yellow Vein Virus-China (AYVV-China), sedangkan isolat Kaliurang berkerabat dengan Tomato Yellow Leaf Curl Virus-China (TYLCV-China) atau ToLCV-Laos.
Amplifikasi PCR menggunakan asam nukleat total dan primer spesifik untuk gen AV1 Begomovirus, sekuensing secara langsung dari produk PCR, dan analisis sekuen asam nukleotida dan asam amino menggunakan BLAST telah dilakukan. Hasil dari percobaan adalah (i) adanya pita DNA hasil amplifikasi PCR membuktikan bahwa sampel-sampel tomat yang sakit terinfeksi oleh Begomovirus (ii) hasil analisis BLAST menggunakan sekuen nukelotida dan asam amino menunjukkan bahwa fragmen DNA hasil amplifikasi PCR adalah gen AV1 dari Begomovirus, (iii) identitas asam nukleat dan asam amino dari gen AV1 di antara isolat-isolat Begomovirus mengindikasikan bahwa isolat-isolat tersebut adalah isolat Ageratum yellow vein virus (AYVV) Indonesia, dan (iv) hasil analisis filogenetik mengindikasikan bahwa delapan isolat Begomovirus tersebut terbagi menjadi dua kelompok yang berbeda.
Serangkaian tahapan untuk konstruksi gen AV1 Begomovirus juga telah dilakukan diantaranya adalah amplifikasi gen AV1 menggunakan primer spesifik, transformasi ke bakteri E. coli DH5α dan kloning gen tersebut ke vektor ekspresi pBI121. Transformasi genetik dilakukan dengan cara eksplan potongan daun tanaman tembakau yang ditumbuhkan secara in vitro ditransformasi melalui ko-kultivasi dengan A. tumefaciens yang mengandung konstruksi gen AV1. Hasil percobaan menunjukan bahwa gen AV1-Begomovirus berhasil diamplifikasi dan disisipkan ke dalam vektor ekspresi pBI121. Tanaman-tanaman tembakau hasil transformasi genetik dengan gen AV1 telah dihasilkan dan diaklimatisasi di rumah kaca dan diketahui telah membawa gen ketahanan terhadap kanamisin (gen nptII) dan gen AV1.
mekanisme ketahanan yang terjadi dapat dijelaskan lebih detail.
Pemuliaan konvensional dilakukan untuk mendapatkan galur-galur tomat yang tahan TYLCV (Begomovirus) yang dikombinasikan dengan ketahanan terhadap CMV. Materi tanaman yang digunakan dalam percobaan adalah tanaman generasi F1-TYLCV (hasil persilangan galur tahan dan rentan TYLCV) dan tanaman generasi F1-CMV (hasil persilangan galur rentan dan galur transgenik tahan CMV). Hasil percobaan menunjukkan bahwa bioasai tanaman-tanaman F1-doublecross (F1DC-Intan/R8-110-11//FLA456/Intan dan F1
DC-CL6046/R8-110-11//FLA456/CL6046) dengan TYLCV diperoleh masing-masing 10 dan 9 tanaman yang menunjukkan fenotipe tahan. Hal ini mengindikasikan bahwa tanaman-tanaman F1-kombinasi tersebut telah membawa gen ketahanan terhadap TYLCV. Deteksi gen CP-CMV dengan teknik PCR mengindikasikan bahwa gen tersebut juga telah terbawa pada tanaman-tanaman F1-DC. Dengan demikian, pada penelitian ini telah diperoleh tanaman-tanaman F1-doublecross/F1-DC (hasil persilangan antara F1-TYLCV tahan dan F1-CMV tahan) yang memperlihatkan fenotipe yang tahan terhadap TYLCV dan membawa gen ketahanan terhadap CMV. Tanaman-tanaman F1-DC ini akan dijadikan sebagai materi untuk pengembangan varietas tomat tahan TYLCV dan CMV selanjutnya.
© Hak Cipta milik IPB, tahun 2008
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
IDENTIFIKASI BEGOMOVIRUS INDONESIA PADA TOMAT
DAN ANALISIS DIVERSITAS GENETIK GEN AV1 SERTA
PEMANFAATANNYA UNTUK PENGEMBANGAN TANAMAN
TAHAN VIRUS
TRI JOKO SANTOSO
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Agronomi
SEKOLAH PASCA SARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup : Dr. Ir. Endang Nurhayati, MS
Penguji pada Ujian Terbuka : Dr. Ir. Agus Purwito, MSc.Agr
PRAKATA
Puji dan syukur penulis panjatkan pada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini dapat diselesaikan. Disertasi ini berjudul “Identifikasi Begomovirus Indonesia Pada Tomat dan Analisis Diversitas Genetik Gen AV1 serta Pemanfaatannya Untuk Pengembangan Tanaman Tahan Virus”. Disertasi ini memuat dua bab yang merupakan pengembangan dari naskah artikel yang diajukan ke jurnal ilmiah. Bab 4 berjudul “Identitas dan keragaman genetik Begomovirus yang berasosiasi dengan penyakit keriting pada tomat berdasarkan teknik PCR-RFLP” telah diterbitkan (AgroBiogen 4[1]: 9-7. April 2008). Bab 5 berjudul “Identity and sequence diversity of Begomovirus associated with yellow leaf curl disease of tomato in Indonesia” juga telah diterbitkan (Microbiology Indonesia 2[1]: 1-7. April 2008).
Pada kesempatan ini, penulis menyampaikan terima kasih yang sebesar-besarnya kepada Prof Dr Ir Sudarsono MSc selaku ketua komisi pembimbing, Dr Ir Hajrial Aswidinnoor MSc, Dr Ir Sri Hendrastuti Hidayat MSc dan Dr Muhammad Herman selaku anggota komisi pembimbing yang telah banyak membimbing dan mengarahkan penulis selama menjalani pendidikan dan penelitian di Sekolah Pascasarjana, Institut Pertanian Bogor. Terima kasih juga disampaikan kepada Dr Ir Endang Nurhayati MS, Dr Ir Agus Purwito MSc dan Dr Ir Ati Srie Duriat, APU (Profesor Riset) selaku dosen penguji luar komisi pada ujian tertutup dan terbuka yang telah banyak memberi masukan dan saran. Di samping itu, penulis juga menyampaikan terima kasih yang tidak terkira kepada Kepala Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian dan Sekretaris Badan Litbang Pertanian atas ijin dan kesempatan yang diberikan kepada penulis untuk melanjutkan studi di Sekolah Pasca Sarjana, IPB. Demikian juga kepada pimpinan proyek USAID-ABSP II dan PTAAP II Badan Litbang Pertanian beserta staf yang telah membiayai sekolah ini. Penulis juga menyampaikan terima kasih kepada Rektor IPB dan Ketua Program Studi Agronomi yang telah menerima penulis untuk menjadi mahasiawa program doktor dan atas bimbingan serta dorongan yang telah diberikan selama penulis menjalani masa studi.
Penulis juga mengungkapkan terima kasih yang setulus-tulusnya kepada Bapak dan Ibu, Bapak dan Ibu mertua serta seluruh keluarga atas segala lantunan doa, jerih payah dan kasih sayangnya sehingga penulis mempunyai motivasi untuk menyelesaikan studi dan penelitian. Ungkapan terima kasih juga disampaikan untuk istri tercinta dan anak-anak tersayang atas kesabaran dan ketabahan dalam mendampingi, memberi motivasi dan inspirasi selama penulis menempuh studi.
Penulis juga menyampaikan terima kasih kepada rekan-rekan staf peneliti serta para teknisi yang tergabung dalam tim penelitian transformasi padi di Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian. Ucapan terima kasih juga penulis sampaikan kepada semua pihak yang tidak bisa penulis sebutkan satu per satu yang telah memberi dukungan moril dan membantu dalam pelaksanaan penelitian.
ini dapat bermanfaat di kemudian hari. Amien.
Bogor, November 2008
RIWAYAT HIDUP
Penulis dilahirkan di Klaten, Jawa Tengah, pada tanggal 19 Mei 1972 dari
pasangan Bapak Ngadimin Hadi Sumarto dan Ibu Sri Natun sebagai anak kedua
dari lima bersaudara. Pada tahun 2005 menikah dengan Atmitri Sisharmini MSi
dan dikaruniai dua orang anak putri dan putra, Aulia Izzati Putri (2,5 tahun) dan
Rais Arkan Nugraha (6 bulan).
Pada tahun 1984 penulis menyelesaikan pendidikan SD di SDN II Meger,
Ceper, Klaten kemudian melanjutkan ke SMPN I Ceper dan lulus pada tahun
1987. Pada tahun 1990 lulus dari SMAN I Klaten. Penulis memperoleh gelar
Sarjana Pertanian dari Fakultas Pertanian, Universitas Gadjah Mada di
Yogyakarta pada tahun 1995. Pada tahun 2004 memperoleh gelar Magister Sains
dari Program Studi Bioteknologi, Sekolah Pascasarjana, Institut Pertanian Bogor
melalui beasiswa pendidikan dan dana penelitian dari proyek ARMP II, Badan
Litbang Pertanian, Departemen Pertanian. Pada tahun 2004 melanjutkan studi
program Doktor (S3) dengan biaya dari proyek USAID-ABSP II dan PTAAP
(Departemen Pertanian) pada program Studi Agronomi Institut Pertanian Bogor.
Penulis saat ini bekerja sebagai staf peneliti di Kelompok Peneliti Biologi
Molekuler, Balai Besar Penelitian dan Pengembangan Bioteknologi dan
Sumberdaya Genetik Pertanian (BB BIOGEN) Bogor, sejak tahun 1996 sampai
DAFTAR ISI
DAFTAR TABEL ………... xviii
DAFTAR GAMBAR ... xx
DAFTAR LAMPIRAN ... xxiii
I. PENDAHULUAN Latar Belakang ... 1
Tujuan Penelitian ... 6
Strategi dan Alur Penelitian ... 6
II. TINJAUAN PUSTAKA Famili Geminiviridae ... 9
Karakterisasi molekuler dari Begomovirus ... 12
Keragaman genetik dari Begomovirus ... 14
Teknik deteksi dan identifikasi Begomovirus ... ... 15
Pemuliaan konvensional untuk ketahanan terhadap Begomovirus .. 17
Rekayasa genetik untuk ketahanan terhadap Begomovirus ... 18
III. DETEKSI BEGOMOVIRUS YANG MENGINFEKSI TOMAT MENGGUNAKAN TEKNIK POLYMERASE CHAIN REACTION (PCR) Abstrak ... 20
Abstract ... 21
Pendahuluan ... 22
Bahan dan Metode ... 23
Hasil ... 26
Pembahasan ... 29
Simpulan ... 33
Daftar Pustaka ... 34
IV. IDENTITAS DAN KERAGAMAN GENETIK BEGOMOVIRUS YANG BERASOSIASI DENGAN PENYAKIT KERITING PADA TOMAT BERDASARKAN TEKNIK PCR-RFLP Abstrak ... 36
Abstract ... 37
Pendahuluan ... 38
Bahan dan Metode ... 39
Hasil ... 43
Pembahasan .. ... 49
Simpulan ... 51
V. IDENTITY AND SEQUENCE DIVERSITY OF BEGOMOVIRUS ASSOCIATED WITH YELLOW LEAF CURL DISEASE OF TOMATO IN INDONESIA
Abstrak ... 54
Abstract ... 55
Introduction ... 56
Materials and Method ... 57
Results ... 60
Discussions ... 66
Conclusion ... 67
References ... 68
VI. KONSTRUKSI GEN AVI BEGOMOVIRUS PADA VEKTOR EKSPRESI DAN INTRODUKSINYA KE TEMBAKAU MENGGUNAKAN Agrobacterium tumefaciens Abstrak ... 70
Abstract ... 71
Pendahuluan ... 72
Bahan dan Metode ... 74
Hasil ... 79
Pembahasan ... 86
Simpulan ... 88
Daftar Pustaka ... 89
VII. ANALISIS MOLEKULER DAN UJI KEEFEKTIFAN GEN AV1 PADA TANAMAN TEMBAKAU TRANSGENIK UNTUK KETAHAHAN TERHADAP BEGOMOVIRUS Abstrak ... 91
Abstract ... 92
Pendahuluan ... 93
Bahan dan Metode ... 95
Hasil ... 97
Pembahasan ... 104
Simpulan ... 107
Daftar Pustaka ... 107
VIII. PENDEKATAN KONVENSIONALUNTUK KETAHANAN TOMAT TERHADAP BEGOMOVIRUS YANG DIKOMBINASIKAN DENGAN KETAHANAN TERHADAP CMV Abstrak ... 109
Abstract ... 110
Pendahuluan ... 111
Hasil ... 119
Pembahasan ... 126
Simpulan ... 130
Daftar Pustaka ... 130
IX. PEMBAHASAN UMUM ... 133
X. SIMPULAN UMUM DAN SARAN ... 140
DAFTAR PUSTAKA ... 142
DAFTAR TABEL
Halaman
1. Deskripsi gejala dominan pada tanaman tomat sakit yang
ditemukan di beberapa lokasi pengambilan sampel ... 27
2. Frekuensi kejadian penyakit yang disebabkan oleh infeksi
Begomovirus dari beberapa lokasi pengambilan sampel ... 29
3. Ukuran fragmen DNA yang dihasilkan dari pemotongan produk PCR dari 8 isolat Begomovirus dan prediksi RFLP isolat-isolat dari DNA database menggunakan enzim restriksi DraI, EcoRI,
RsaI dan PstI ... 47
4. Isolate identity, observed symptoms on collected tomato samples, location of collected samples, and number of determined nucleic acid and predicted amino acid sequences based on the polymerase chain reaction amplified putative AV1
gene ……….………... 61
5. Percentages of sequence identities of AV1 gene among suspected Begomoviruses isolates determined in this research and three
Begomoviruses available in the GenBank database ... 63
6. Distance matrices (%) based on predicted AV1 gene amino acid sequences of suspected Begomoviruses isolates determined in this research, Ageratum yellow vein virus (AYVV), Soybean crinkle leaf virus (SCLV), Pepper leaf curl virus (PepLCV), Tomato leaf curl virus (ToLCV), and Cassava mosaic virus
(CasMV) ... 64
7. Jumlah tunas dan planlet yang dihasilkan serta persentase tunas menjadi planlet pada transformasi genetik tembakau dengan gen
AV1Begomovirus melalui bantuan A. tumefaciens ... 83
8. Deteksi PCR gen AV1 dan bioasai tanaman tembakau transgenik putatif generasi T0 dengan Begomovirus di rumah kaca ...
99
9. Kategori respon tanaman tembakau transgenik putatif setelah
dianalisis PCR dan bioasai ... 100
10. Hubungan antara analisis PCR, bioasai, jumlah kopi dan
keberadaan virus target dalam tanaman transgenik ... 103
11. Materi tanaman yang digunakan dalam penelitian ... 113
Begomovirus ... 114
13. Skoring keparahan gejala pada tanaman yang terserang CMV ... 115
14. Konfirmasi ketahanan tetua terhadap TYLCV melalui penularan
dengan serangga vektor kutu kebul di rumah kaca ... 119
15. Skrining beberapa galur tomat terhadap TYLCV melalui penularan dengan serangga vektor kutu kebul di rumah kaca
... 120
16. Skrining beberapa tanaman tomat terhadap CMV menggunakan
penularan secara mekanis di rumah kaca ... 122
17. Berat benih yang dihasilkan dari masing-masing F1-silang ganda
... 124
18. Skrining tomat F1-IC-Intan/R8-110-11//FLA456/Intan (39) terhadap TYLCV melalui penularan dengan serangga vektor kutu
kebul di rumah kaca ... 125
19 Skrining tomat F1IC-CL6046/R8-110-11//FLA456/CL6046 (38)
terhadap TYLCV melalui penularan dengan serangga vektor kutu
DAFTAR GAMBAR
Halaman
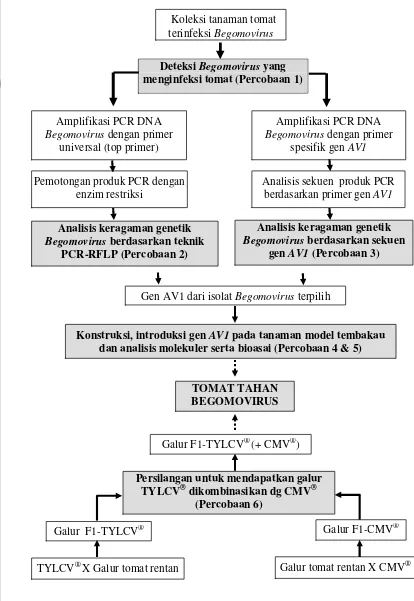
1. Diagram alur strategi penelitian dan keterkaitan antar percobaan
dari selutruh kegiatan penelitian ... 8
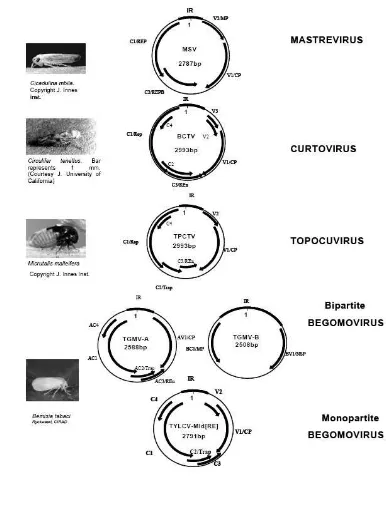
2. Taksonomi dari famili Geminiviridae: tipe spesies, organisasi
genom, tanaman inang dan vektor serangganya. ... 9
3. Organisasi genom masing-masing genus dari famili Geminiviridae
dan serangga vektor utamanya ... 11
4. Morfologi gejala pada tanaman tomat yang diduga terinfeksi oleh
Begomovirus yang ditemukan di lapang ... 26
5. Elektroforesis gel dari fragmen DNA hasil optimasi teknik amplifikasi PCR menggunakan primer universal untuk mendeteksi Begomovirus pada sampel koleksi Laboratorium Virologi, PS
Proteksi Tanaman, IPB ... 27
6. Gel elektroforesis fragmen DNA produk amplifikasi PCR menggunakan primer universal untuk mendeteksi Begomovirus pada
tanaman tomat sakit yang dikoleksi dari Malang dan Blitar ... 28
7. Tipe gejala pada tanaman tomat sakit yang ditemukan di daerah
Pagerwangi, Lembang, Jawa Barat ... 30
8. Tanaman tomat sakit yang diduga terinfeksi Begomovirus yang
ditemukan di lapang ... 43
9. Elektroforesis hasil amplifikasi PCR DNA Begomovirus pada sampel tanaman tomat menggunakan primer universal dari 8 daerah
yang berbeda pada agarosa gel 1%. ... 44
10. Elektroforesis fragmen DNA produk amplifikasi PCR dari genom Begomovirus yang dipotong dengan ensim restriksi (a) DraI (b)
EcoRI (c) RsaI dan (d) PstI pada gel agarosa 1%. ... 45
11. Dendrogram hasil analisis fragmen DNA restriksi dari isolat-isolat Begomovirus asal tomat dari 8 daerah yang berbeda menggunakan
program NTSYSpc-21 ... 46
12. Dendrogram yang dihasilkan oleh analisis keragaman genetik berdasarkan fragmen situs restriksi dari isolat-isolat Begomovirus yang terdiri dari 8 isolat lokal Indonesia dan isolat-isolat dari
database bank gen menggunakan program NTSYSpc-21. ... 48
13. Tomato plants exhibited various leaf-curl symptoms. Subsequent
14. Agarose gel electropherogram of polymerase chain reaction (PCR) amplified DNA fragments of putative AV1. The DNA fragments were amplified by PCR using AV1 specific primers and total
nucleic acid of diseased tomato sample …... 60
15. Alignment of partial amino acid sequences predicted from determine nucleotide sequences of AV1 gene of eight Begomovirus isolates determined in this research and seven Begomovirus isolates
available from GenBank DNA database... 62
16. Phylogenetic relationship based on predicted AV1 gene amino acid sequences of suspected Begomoviruses isolates determined in this research, and other Begomoviruses available in the GenBank DNA
database………... 65
17 Elektroforesis pada gel agarosa 1%. (a) produk amplifikasi gen AV1 dari dua isolat Begomovirus (CP 8 dan CP11) menggunakan primer spesifik CPPROTEIN-V1 dan CPPROTEIN-C1. (b) DNA plasmid rekombinan pCP8 (1-6) dan pCP11 (1-6) hasil isolasi dari koloni
tunggal bakteri E. coli DH5α ……… 79
18. Peta plasmid biner pBI121 yang membawa gen pelapor gus dangen
marker nptII pada struktur T-DNAnya ... 80
19. Elektroforesis fragmen gen AV1 yang dipotong dari vektor pGEM-T easy dan fragmen gen GUS dari vektor ekspresi pBI121 dengan enzim restriksi XbaI dan SacI pada gel agarosa 1%. AV1 = fragmen gen AV1 yang berukuran 780 bp; GUS = fragmen gen GUS yang
berukuran 2000 bp ... 81
20. Elektroforesis hasil verifikasi insersi fragmen gen AV1 dengan
enzim restriksi XbaI dan SacI ... 82
21. Peta konstruksi plasmid biner pBI-CP yang membawa gen AV1 Begomovirus dengan promoter 35S-CaMV dan terminator nos, dan
gen marker nptII pada struktur T-DNA ... 82
22. Transformasi genetik tembakau dengan gen AV1 melalui vektor A.
tumefaciens ... 84
23. Elektroforesis gel hasil amplifikasi gen nptII pada 46 tanaman tembakau transgenik putatif generasi T0 menggunakan primer PCR
spesifik. ... 85
24. Deteksi PCR gen AV1 pada 46 tanaman tembakau transgenik putatif
generasi T0 menggunakan primer spesifik ... 98
25. Bioasai tanaman tembakau transgenik menggunakan vektor
26. Analisis hibridisasi Southern Blot pada sampel tanaman tembakau trasngenik putatif generasi T0 yang positif PCR dan 2 tanaman yang
negatif PCR (no.10 & 20) dengan pelacak gen AV1 ... 102
27. Deteksi keberadaan Begomovirus dengan teknik PCR menggunakan primer universal pada tanaman tembakau transgenik generasi T0
setelah bioasai ... 103
28. Skrining tanaman F1-TYLCV (F1 FLA456/Intan dan FLA456/CL6046) dengan TYLCV menggunakan vektor kutu kebul
di rumah kaca ... 121
29. Beberapa gejala tanaman F1-CMV setelah inokulasi dengan CMV .. 122
30. Amplifikasi gen CP pada tanaman generasi F1 Intan/R8-110-11; Varietas Intan; Air; Galur transgenik R8-110-11 menggunakan
teknik PCR ………... 123
31. Amplifikasi gen CP-CMV pada tanaman generasi F1 CL6046/R8-110-11; Varietas CL6046; Air; Galur transgenik R8-110-11
menggunakan teknik PCR ……….. 123
32. Amplifikasi gen CP-CMV pada tanaman generasi F1
IC-Intan/R8-110-11//FLA456/Intan (39); Varietas Intan; Air; Galur transgenik
R8-110-11(+) menggunakan teknik PCR ………... 126
33. Amplifikasi gen CP-CMV pada tanaman generasi F1
IC-CL6046/R8-110-11//FLA456/CL6046 (38); Varietas CL6046; Air; Galur
DAFTAR LAMPIRAN
Halaman
1. Amplifikasi PCR dengan primer universal untuk mendeteksi
Begomovirus pada tanaman tomat dari beberapa daerah ... 153
2. Komposisi Media Dasar Murashige and Skoog ... 154
I. PENDAHULUAN
Latar Belakang
Tomat (Lycopersicum esculentum Mill.) adalah salah satu komoditas
sayuran penting secara ekonomi yang dibudidayakan hampir di seluruh dunia
termasuk Indonesia. Komoditas ini mempunyai banyak fungsi di antaranya adalah
bahwa buah tomat dapat berfungsi sebagai sayuran, buah meja, minuman, bahkan
sebagai bahan kosmetik dan obat-obatan (Duriat 1996). Di bidang kesehatan,
tomat merupakan salah satu komoditas yang mendapat perhatian besar karena
selain kandungan vitamin dan mineral, tomat juga mengandung senyawa
antioksidan yang bermanfaat untuk pencegahan penyakit kronis termasuk di
antaranya adalah penyakit jantung koroner dan beberapa penyakit kanker
(Weisburger 1998). Buah tomat kaya akan senyawa-senyawa karotenoid termasuk
likopin (lycopene). Tomat juga termasuk dalam lima besar tanaman sayuran
penting di Indonesia selain kubis, bawang putih, kacang kapri dan cabai. Produksi
tomat pada tahun 2006 mencapai hampir 640.385 ton dengan produktifitas 11,74
ton/ha dan luas panen 54.527 ha (Deptan 2007).
Infeksi virus daun (kuning) menggulung yang menyebabkan penyakit
“keriting daun” pada tomat [Tomato (Yellow) Leaf Curl Virus, TYLCV/ToLCV]
dari salah satu anggota genus Begomovirus (Famili Geminiviridae), adalah salah
satu kendala biotik yang serius pada produksi tomat di seluruh dunia.
Gejala-gejala tanaman yang terinfeksi virus ini diantaranya adalah penghambatan
pertumbuhan, daun menguning dan menggulung (keriting) serta tanaman menjadi
kerdil. Virus ini dapat menginfeksi tanaman tomat baik pada tanaman muda atau
tua yang ditanam di lapang terbuka atau di rumah kaca, dan menyebabkan
kehilangan produksi yang dapat mencapai 100% apabila menginfeksi tanaman
sewaktu masih muda. Virus ini telah ditemukan di beberapa negara tropik,
subtropik dan mediterania seperti negara-negara di Timur Tengah, Eropa Barat
Daya, Afrika, Asia Tenggara dan kepulauan Karibia (Green & Kalloo 1994;
Czosnek & Laterrot 1997; Jones 2003), bahkan juga ditemukan di daerah dengan
iklim temperate (Moriones & NavasCastillo 2000) yang kejadian penyakitnya
berkisar antara 20 - 100% dan dapat menyebabkan kehilangan hasil sampai 100%
virus ini dapat menginfeksi hampir 90-100% tanaman tomat dan mengakibatkan
pengurangan hasil antara 50-100% (AVRDC Centerpoint newsletter – spring 2003
issue). Sudiono et al. (2001) melaporkan bahwa serangan virus daun kuning
menggulung pada tanaman tomat di daerah Bogor dan sekitarnya mencapai
50-70%.
Usaha pengendalian penyakit keriting yang disebabkan infeksi TYLCV
sampai saat ini masih sulit untuk dilakukan karena tidak ada pestisida yang dapat
diaplikasikan secara langsung untuk mengontrol virus tersebut. Pengendalian
biasanya dilakukan secara tidak langsung antara lain dengan mengurangi sumber
inokulum dengan cara mencabut atau menghilangkan tanaman-tanaman yang
telah menunjukkan gejala serangan virus, mengendalikan perkembangan serangga
vektor, melakukan pergiliran tanaman, dan pemberantasan gulma yang dapat
menjadi inang pembawa virus. Akan tetapi cara-cara pengendalian ini terkadang
kurang efektif karena proses penularan virus ini dapat terjadi dengan cepat
mengingat penularan virus ini dilakukan oleh serangga vektor. Penggunaan
varietas tahan merupakan pilihan yang tepat untuk mengendalikan virus karena
metode ini relatif lebih aman dan murah bila dibandingkan dengan metode
pengendalian yang lain.
Terdapat dua pendekatan utama untuk pengembangan ketahanan genetik
terhadap virus yang tergantung pada sumber gen yang digunakan (Dasgupta et al.
2003). Gen ketahanan dapat berasal dari virus itu sendiri atau berasal dari sumber
yang lain. Pendekatan pertama didasarkan pada konsep ketahanan yang berasal
dari patogen (pathogen-derived resistance, PDR). Pendekatan PDR memanfaatan
elemen genetik yang berupa gen utuh atau bagian gen dari genom virus kemudian
diklon dan diintroduksikan ke tanaman, yang selanjutnya akan mempengaruhi
satu atau beberapa tahap penting dalam siklus hidup virus. Pemanfaatan gen
selubung protein (coat protein gene) (Vidya et al. 2000) merupakan salah satu
contoh dari pendekatan PDR ini. Pendekatan yang kedua adalah ketahanan yang
berasal bukan dari patogen (non pathogen-derived resistance), yang didasarkan
pada pemanfaatan gen-gen ketahanan dari tanaman inang dan gen-gen lain yang
bertanggungjawab untuk adaptasi dan respon tanaman inang terhadap serangan
tersebut. Penggunaan pendekatan non-PDR, diantaranya dilakukan oleh Hanson et
al. (2000). Meskipun tidak sepopuler pendekatan PDR, pendekatan non PDR
memberikan harapan dan peluang yang besar untuk mengembangkan ketahanan
yang bersifat durabel (dapat bertahan lama dan berkelanjutan) ketika
dikombinasikan dengan pendekatan PDR.
Galur-galur tomat hasil pemuliaan secara konvensional yang mempunyai
ketahanan terhadap TYLCV (Begomovirus) telah dikembangkan oleh The Asian
Vegetables Research and Development Center (AVRDC), Taiwan dan telah diuji
serta terbukti efektif terhadap beberapa strain TYLCV Asia termasuk diantaranya
Taiwan, India Selatan dan Thailand (AVRDC Centerpoint newsletter – spring
2003 issue). Galur-galur tomat yang tahan CMV juga telah dikembangkan oleh
AVRDC melalui pendekatan rekayasa genetik menggunakan gen protein selubung
(coat protein gene). Sampai sekarang ini, galur transgenik tahan CMV tersebut
telah dievaluasi di lapang dan menunjukkan tingkat ketahanan yang memadai
untuk mengendalikan infeksi virus. Melalui proyek kerjasama ABSP II yang
didanai oleh USAID, persilangan antara tomat varietas Indonesia (Intan dan
CL6046) dengan varietas tomat yang tahan TYLCV (FLA 456 dan FLA 478) atau
varietas tomat transgenik tahan CMV (R7-110-11) telah dilakukan di AVRDC
dan menghasilkan tanaman tomat generasi F1 dari masing-masing persilangan
(tanaman F1-TYLCV dan F1-CMV). Tanaman tomat generasi F1-TYLCV dan
F1-CMV tersebut kemudian didonasikan ke Indonesia (BB BIOGEN) sebagai
materi untuk pengembangan tomat tahan multi-virus.
Pendekatan konvensional untuk pengembangan varietas tahan virus
memiliki beberapa keterbatasan. Di antaranya adalah sumber gen ketahanan
belum ditemukan pada koleksi plasmanutfah tomat di Indonesia. Selain itu,
kultivar tahan yang dihasilkan melalui pemuliaan konvensional dengan
memanfaatkan gen-gen ketahanan dari kerabat liar akan cepat terpatahkan.
Kondisi tersebut disebabkan perubahan dari virus yang cepat akibat adanya
rekombinasi dan adanya variasi genetik yang tinggi dari virus. Kultivar tahan
yang dihasilkan mungkin hanya spesifik untuk strain atau isolat tertentu. Oleh
karena itu, diperlukan pendekatan lain seperti pemanfaatan teknik rekayasa
ketahanan yang tinggi.
Pemanfaatan teknik rekayasa genetik memberikan wahana baru bagi para
pemulia tanaman untuk memperoleh gen baru yang lebih luas (Greenberg & Glick
1993). Di samping itu, munculnya teknik rekayasa genetik dapat mengatasi
masalah inkompatibilitas dan linkage drag karena introgresi gen-gen penting
dilakukan dengan mengintroduksikan gen secara langsung ke dalam genom
tanaman. Sifat ketahanan tanaman terhadap beberapa cekaman biotik seperti
misalnya gulma, virus, serangga dan mikroorganisme telah dapat diperbaiki
dengan pendekatan ini. Demikian pula terhadap cekaman abiotik dan modifikasi
kualitas dan kuantitas produk tanaman (Bennet 1993). Teknologi rekayasa genetik
dapat digunakan sebagai mitra dan pelengkap teknik pemuliaan tanaman
konvensional yang telah digunakan dengan sukses selama bertahun-tahun
(Riazudin 1994). Suatu gen yang tidak terdapat pada suatu spesies tanaman
tertentu dimungkinkan untuk dapat diperoleh dari organisme lain, seperti bakteri,
virus, binatang dan tanaman lain dan dipindahkan ke tanaman (Herman 1996).
Sebagai contoh gen penyandi protein selubung virus (virus coat protein gene),
diisolasi dari virus untuk memperoleh resistensi non-konvensional terhadap virus.
Gen ini digabungkan dengan suatu sekuen pengendali (promoter dan terminator)
dan ditransformasikan ke dalam tanaman. Bila gen tersebut terekspresi ke dalam
tanaman akan terjadi akumulasi protein pembungkus virus. Mekanisme resistensi
ini berperan pada tingkat awal proses replikasi virus, dengan menghalangi proses
replikasi secara tidak terkendali dari partikel virus (Aswidinnoor 1995).
Mekanisme lain yang juga berperan di dalam ketahanan terhadap virus yang
dikembangkan melalui pendekatan rekayasa genetik adalah mekanisme
pembungkaman gen paska transkripsi (post transcriptional gene silencing)
(Dasgupta et al. 2003). Berdasarkan informasi ini, perlu dilakukan pemanfaatan
gen-gen Begomovirus untuk pengembangan varietas tahan.
Di dalam pengembangan tanaman tomat tahan virus, adanya informasi
tentang keragaman genetik virus akan dapat bermanfaat dalam hal pemilihan
lokasi untuk pengujian (uji multi-lokasi). Selain itu, informasi mengenai suatu
strain virus yang dominan menginfeksi tanaman tomat perlu diketahui sehingga
negara India dan Taiwan, telah berhasil diidentifikasi secara molekuler. Urutan
DNA genom dari isolat-isolat tersebut telah dapat dibandingkan sehingga dapat
diketahui tingkat kesamaannya (Zeidan et al. 1998). Kemajuan di bidang biologi
molekuler telah menghadirkan beberapa teknik yang dapat digunakan untuk
mendeteksi dan mengidentifikasi virus. Salah satu teknik molekuler yang banyak
diaplikasikan adalah Polymerase Chain Reaction (PCR) karena teknik ini sangat
sensitif dan spesifik untuk mendeteksi dan mengidentifikasi patogen-patogen
tanaman. Selain itu, PCR dapat digunakan untuk mengetahui komposisi populasi
patogen dan diversitas genetik virus (Rojas et al. 1993). Spesifisitas PCR
didasarkan pada penggunaan primer-primer oligonukleotida yang komplementer
dengan daerah yang mengapit sekuen DNA yang diamplifikasi. Deteksi virus
dengan metode serologi mempunyai beberapa kelemahan diantaranya adalah
rendahnya titer dari antigen sehingga virus sulit untuk dideteksi, adanya reaksi
silang antibodi dengan antigen heterolog dan adanya pengaruh pengaturan
produksi antibodi oleh lingkungan dan tahap perkembangan dari tanaman.
Sedangkan metode deteksi dengan PCR mempunyai keuntungan antara lain
metode ini hanya membutuhkan sampel DNA yang sedikit yang dapat diperoleh
dari jaringan tanaman yang segar, disimpan di lemari es atau bahkan jaringan
yang telah kering. Selain itu deteksinya tidak dipengaruhi oleh tahap
perkembangan tanaman dan faktor lingkungan. Teknik ini juga relatif lebih mudah
untuk dilakukan dan memungkinkan untuk analisis sekuen (sequencing)
berdasarkan fragmen produk PCR yang terbentuk.
Di Indonesia, keragaman genetik Begomovirus pada tingkat molekuler
(urutan basa DNA) belum banyak dilaporkan. Usaha identifikasi melalui teknik
hibridisasi asam nukleat dan polymerase chain reaction (PCR) telah dirintis oleh
beberapa peneliti (Hidayat et al. 1999; Aidawati et al. 2005). Namun demikian,
informasi yang lebih mendetail mengenai urutan sekuen DNA dari Begomovirus
yang mungkin berkaitan dengan sekuen-sekuen fungsional atau yang dapat
menunjukkan adanya keragaman genetik di antara Begomovirus belum pernah
Tujuan Penelitian
1. Memperoleh informasi tentang adanya infeksi Begomovirus pada
pertanaman tomat di beberapa daerah sentra produksi melalui deteksi
menggunakan teknik Polymerase Chain Reaction (PCR) dan menentukan
identitas serta keragaman genetiknya berdasarkan teknik PCR-RFLP dan
analisis sekuen nukleotida dan asam amino gen AV1.
2. Memperoleh klon gen AV1 Begomovirus pada vektor kloning dan
mendapatkan konstruksi gen tersebut pada vektor ekspresi untuk
digunakan dalam kegiatan transformasi genetik tanaman.
3. Mendapatkan tanaman-tanaman tembakau transgenik (sebagai tanaman
model) yang membawa gen AV1 untuk mempelajari keefektifan gen
tersebut dalam hubungannya dengan ketahanan terhadap Begomovirus.
4. Mendapatkan galur-galur tanaman tomat yang tahan terhadap
Begomovirus (TYLCV) yang dikombinasikan dengan ketahanan terhadap
CMV melalui pendekatan konvensional
Strategi dan Alur Penelitian
Untuk dapat mencapai tujuan-tujuan tersebut di atas maka strategi
penelitian yang dilakukan meliputi beberapa pendekatan, yaitu diantaranya adalah
melakukan survei dan mengumpulkan tanaman tomat sakit atau bagiannya yang
menunjukkan gejala-gejala spesifik terinfeksi oleh Begomovirus dari beberapa
sentra produksi di Indonesia. Asam nukleat total dari jaringan tanaman tomat sakit
diisolasi dan digunakan sebagai cetakan untuk amplifikasi DNA genom
Begomovirus dengan teknik Polymerase Chain Reaction (PCR) dengan
menggunakan primer universal (top primer) untuk Begomovirus (Percobaan 1). Hal ini diharapkan akan diperoleh informasi tentang adanya kejadian penyakit
akibat infeksi Begomovirus pada sentra-sentra produksi di beberapa daerah dan
juga didapatkan isolat-isolat Begomovirus yang dapat digunakan untuk materi
percobaan selanjutnya.
Isolat-isolat Begomovirus yang menginfeksi tomat yang telah dikoleksi
genetiknya untuk menentukan identitas dan hubungan kekerabatan antar isolat
Begomovirus tersebut. Studi diversitas genetik dilakukan dengan melihat adanya
perbedaan situs enzim restriksi dari masing-masing isolat Begomovirus
berdasarkan teknik PCR-RFLP (Percobaan 2). Untuk mempelajari secara lebih detail adanya diversitas genetik di antara isolat Begomovirus, dilakukan analisis
sekuen nukleotida dan asam amino dari gen AV1 yang merupakan gen yang
mempunyai sekuen yang konservatif (conserved sequences) (Percobaan 3). Di samping informasi tentang keragaman genetik dari isolat-isolat Begomovirus yang
menginfeksi tomat, dari penelitian 3 juga diharapkan dapat diperoleh identitas
genetik Begomovirus Indonesia dengan Begomovirus yang ada di database DNA
(GenBank).
Upaya untuk mengendalikan penyakit yang berasosiasi dengan
Begomovirus dapat ditempuh dengan menggunakan varietas-varietas tomat yang
tahan, maka pada penelitian disertasi ini dilakukan dua pendekatan yang berbeda
untuk merakit tanaman tahan terhadap Begomovirus. Pertama, pendekatan
non-konvensional melalui teknik rekayasa genetik dengan menggunakan gen yang
berasal dari Begomovirus itu sendiri, yang sering disebut dengan
pathogen-derived resistance (PDR). Untuk pendekatan ini dilakukan konstruksi gen AV1
Begomovirus (menyandikan protein selubung) pada vektor ekspresi dan konstruk
gen AV1 diintroduksikan ke tanaman tembakau menggunakan vektor bakteri A.
tumefaciens (Percobaan 4). Transformasi genetik tanaman tembakau (tanaman model) dengan gen AV1 dimaksudkan untuk mempelajari fungsi dan efektifitas
gen AV1 sebelum diintroduksikan ke tanaman target. Tanaman-tanaman tembakau
transgenik putatif yang dihasilkan pada penelitian 4 digunakan sebagai materi
untuk analisis deteksi keberadaaan gen AV1 pada genom menggunakan teknik
PCR dan Southern Blot serta untuk evaluasi keefektifan gen AV1 terhadap
Begomovirus (Percobaan 5). Kedua, pendekatan konvensional dilakukan dengan memanfaatkan gen ketahanan terhadap TYCLV yang ada pada galur-galur dari
AVRDC melalui persilangan dengan tomat-tomat Indonesia untuk mendapatkan
tanaman tomat Indonesia yang tahan terhadap TYLCV (Begomovirus)
Gambar 1 Diagram alur strategi penelitian dan keterkaitan antar percobaan dari seluruh kegiatan penelitian
Analisis keragaman genetik
Begomovirus berdasarkan teknik PCR-RFLP (Percobaan 2)
TOMAT TAHAN BEGOMOVIRUS
Koleksi tanaman tomat terinfeksi Begomovirus
Deteksi Begomovirus yang menginfeksi tomat (Percobaan 1)
Amplifikasi PCR DNA Begomovirus dengan primer
universal (top primer)
Analisis sekuen produk PCR berdasarkan primer gen AV1 Pemotongan produk PCR dengan
enzim restriksi
Gen AV1 dari isolat Begomovirus terpilih
Konstruksi, introduksi gen AV1 pada tanaman model tembakau dan analisis molekuler serta bioasai (Percobaan 4 & 5)
Galur tomat rentan X CMV®
Galur F1-TYLCV® Galur F1-CMV®
Persilangan untuk mendapatkan galur TYLCV® dikombinasikan dg CMV®
(Percobaan 6)
Galur F1-TYLCV® (+ CMV®)
Analisis keragaman genetik
Begomovirus berdasarkan sekuen gen AV1 (Percobaan 3)
Amplifikasi PCR DNA Begomovirus dengan primer
spesifik gen AV1
[image:31.612.101.515.78.679.2]II. TINJAUAN PUSTAKA
Famili Geminiviridae
Geminivirus merupakan salah satu kelompok virus tanaman terbesar dan
penting yang meliputi virus-virus yang menginfeksi sejumlah spesies tanaman
baik monokotil atau dikotil. Geminivirus ini secara struktural mempunyai
morfologi berupa partikel virion isometrik kembar yang selalu berpasangan
(twinned-geminate) yang berukuran sekitar 18-30 nm dan secara genetik
mempunyai sebuah DNA genom yang terdiri dari satu atau dua molekul DNA
berutas tunggal (ssDNA) yang berbentuk sirkuler (Gutierrez 2000).
Taksonomi dari famili Geminiviridae terdiri dari empat genus yaitu
Mastrevirus, Curtovirus, Topocuvirus dan Begomovirus (van Regenmortel et al.
1999) yang dibedakan berdasarkan organisasi genetik, tanaman inang dan vektor
yang menginfeksi (Gambar 2). Organisasi genetik dari masing-masing genus dari
famili Geminiviridae berbeda satu sama lain (Gambar 3).
(Ribeiro 2006)
Mastrevirus mempunyai sebuah genom monopartit, terdiri dari sebuah
DNA utas tunggal berbentuk sirkuler (circular ssDNA) dengan ukuran sekitar 2,6
– 2,8 kb. Kelompok virus ini biasanya menginfeksi tanaman monokotil dan
ditularkan oleh kutu daun (leafhoppers, Hemiptera dari famili Cicadellidae)
dengan cara persisten, sirkulatif dan non-propagatif. Genom dari genus ini
mengkodekan empat protein: dua pada utas v-sense (movement protein, MP dan
capsid protein, CP) dan dua pada utas c-sense (RepA dan Rep). Genus ini banyak
ditemukan di Afrika dan termasuk dalam genus ini adalah Maize streak virus
(MSV) dan Wheat dwarf virus (Agrios 1997; van Regenmortel et al. 1999;
Gutierrez 2000).
Curtovirus mempunyai sebuah genom monopartit dan ditularkan oleh kutu
daun (leafhopper) dengan cara persisten, sirkulatif dan non-propagatif. Virus ini
menginfeksi tanaman dikotil. Protein selubungnya lebih mirip dengan protein
selubung dari genus Mastrevirus, akan tetapi ssDNA tunggalnya diorganisasi
lebih mirip dengan DNA A bipartit dari genus Begomovirus. Di samping
menyandikan movement protein (MP) dan coat protein (CP), genom dari genus ini
juga menyandikan protein (V2) pada utas v-sense-nya sedangkan empat protein
dikodekan pada utas c-sense. Protein-protein tersebut adalah Rep, Rep yang
homolog pada genus mastrevirus, protein C2, REn (Replication enhancer protein)
dan protein C4. Virus yang termasuk dalam genus ini adalah beet curly top virus
(BCTV). Genus ini kebanyakan ditemukan di India, Amerika dan negara-negara
Mediterania (van Regenmortel et al. 1999).
Genus Topocuvirus sebenarnya hampir mirip dengan Curtovirus dan
hanya dibedakan dalam famili vektor yang menularkan. Virus dari genus ini
ditularkan oleh treehopper (Hemiptera: Micrutalis malleifera) dan bukan kutu
daun dan menginfeksi tanaman dikotil. Hasil penelitian menunjukkan bahwa
genus ini merupakan hasil rekombinasi dengan virus lain dari genus yang berbeda
(Briddon et al. 1996). Virus dari genus ini pertama kali ditemukan di Florida
(Stoner & Hogan 1950). Genom dari virus genus ini adalah berukuran sekitar
2861 nukleotida dan mengkodekan 6 protein yang mirip dengan Curtovirus
(Briddon et al. 1996). Hanya satu virus yang termasuk dalam genus ini yaitu
Gambar 3 Organisasi genom masing-masing genus dari famili Geminiviridae dan serangga vektor utamanya. MSV=Maize streak virus, BCTV=Beet
curly top virus, TPCTV=Tomato pseudo-curly top virus,
Genus Begomovirus meliputi virus-virus yang menginfeksi tanaman
dikotil. Genus ini terdiri dari virus-virus dengan genom bipartit yang mempunyai
gen-gen yang terletak pada dua molekul DNA utas tunggal sirkuler yang berbeda
(DNA A dan DNA B dengan ukuran masing-masig 2,6-2,8 kb)) atau monopartit
dengan semua gen-nya terletak pada satu DNA utas tunggal sirkuler (2,8 kb).
Begomovirus ini ditularkan oleh serangga kutu kebul (whiteflies) dari genus
Bemisia dengan sifat penularan persisten, sirkulatif dan non-propagatif.
Komponen DNA A dan DNA B mengandung gen-gen yang menyandikan protein
pada utas sense virus (v-sense) dan utas sense komplementer (c-sense).
Komponen DNA A mengandung satu gen (AV1) pada v-sense dan 3 gen (AC1,
AC2, dan AC3) pada c-sense. Pada komponen DNA B mempunyai satu gen
(BV1) pada v-sense dan satu gen (BC1) pada c-sense. Produk protein dari gen
BV1 ditempatkan pada inti sel dan berfungsi mengikat DNA, sehingga genom
virus yang baru dibentuk dapat dipindahkan ke sitoplasma. Produk protein BC1
ditempatkan pada dinding sel dan membran seluler, dan berfungsi untuk
meningkatkan kerja eksklusif dari plasmodesmata dalam pergerakan virus dari sel
ke sel. Kedua movement protein ini berhubungan dalam penentuan kisaran inang
virus, namum hanya gen BC1 yang berperan dalam menentukan keparahan gejala
dan patogenisitas pada Begomovirus. Contoh virus yang termasuk kelompok ini
adalah Bean golden mosaic virus (BGMV) dan Tomato yellow leaf curl virus
(TYLCV) (van Regenmortel et al. 1999).
Karakteristik molekuler dari Begomovirus
Genom dari Begomovirus dapat berupa monopartite (Mediterania,
Amerika Tengah dan Utara, serta sebagian negara di Asia) atau bipartit (Thailand)
(Fauquet et al. 2003; Fauquet & Stanley 2003). Genom bipartit Begomovirus
terdiri dari 2 komponen ssDNA (DNA A dan DNA B) dengan ukuran hampir
sama. Urutan nukleotida DNA A dan DNA B adalah cukup berbeda, kecuali
untuk “common region” pendek berukuran sekitar 200 nukleotida yang sangat
mirip. Daerah tersebut meliputi sebuah struktur stem-loop yang mengandung
dari keempat genus geminivirus dan meliputi sekuen origin untuk replikasi rolling
circle (Horrison & Robinson 2002; Zhou et al. 2003). Genom Begomovirus
mengkodekan 6 open reading frame (ORF) yang saling tumpang tindih secara
parsial (V1,V2, C1, C2, C3, dan C4) dan transkripsi gen-gen dari Begomovirus
terjadi dalam 2 arah pada kedua komponen transkripsi dari genom yang
dipisahkan oleh daerah intergenik (Rybicki et al. 2000).
Protein-protein yang disandikan oleh genus Begomovirus adalah:
- Protein selubung (capsid protein, CP), ORF V1; yang digunakan untuk
menyelubungi genom dan juga sangat penting untuk penyebaran virus
(Briddon et al. 1989). CP dan pre-CP (V2) juga penting untuk pergerakan
lokal atau sistemik yaitu untuk pergerakan keluar masuk genom virus dari
inti sel inang (Gafni & Epel 2002). Komponen AV1 juga berperan dalam
melindungi ssDNA virus dan penularan oleh serangga vektor. Protein ini
juga penting untuk perpindahan virus ketika masuk ke dalam sistem
pencernaan serangga kutu kebul untuk melindungi partikel virus dari
degradasi (Morin et al. 2000).
- Protein yang berhubungan dengan replikasi (replication-associated
protein, Rep), ORF C1; merupakan protein yang hanya terlibat dalam
proses replikasi virus (Desbiez et al. 1995).
- Protein untuk aktivasi transkripsi (transcriptional activator protein), ORF
C2; protein yang terlibat dalam pengaktifan transkripsi dari promoter
protein selubung. Protein ini ditemukan terlokalisasi pada inti dan
berperan dalam patogenisitas virus (van Wezel et al. 2001).
- Protein untuk meningkatkan replikasi (replication enhancer protein), ORF
C3; protein ini berinteraksi dengan protein C1 dan meningkatkan
akumulasi DNA virus (HanleyBowdoin et al. 2000)
- Protein C4 merupakan protein yang penting untuk penentu gejala dan
terlibat dalam inisiasi pembelahan sel (Krake et al. 1998). Protein C4
mungkin berinteraksi dengan ORF RepC1 dan mematahkan mekanisme
pertahanan tanaman (van Wezel et al. 2002).
- Produk protein yang disandikan oleh pre-CP (V2/MP) dan ORF C4 diduga
DNA A dari spesies bipartit mempunyai susunan yang hampir sama
dengan genom dari Begomovirus monopartit. Untuk Begomovirus bipartit Dunia
Baru, komponen DNA A tidak mempunyai gen AV2. Komponen DNA B
menyandikan BV1 dan BC1, protein-protein yang penting untuk pergerakan virus
dari sel ke sel dan untuk infeksi sistemik (Sanderfoot et al. 1996), dan dapat
mempengaruhi kisaran inang (Ingham et al. 1995). Meskipun tidak secara
langsung terlibat dalam interaksi dengan vektor kutu kebul, sekuen DNA B
mempengaruhi efisiensi akuisisi virus oleh serangga dengan menentukan lokasi
Begomovirus pada jaringan tanaman (Liu et al. 1997).
Infeksi Begomovirus ini telah terjadi pada beberapa tanaman penting
seperti kacang-kacangan, mentimun, tomat, cabai dan ubikayu pada daerah tropis
dan sub-tropis serta beberapa rumput (Roye et al. 1997; Ambrozevicius et al.
2002). Di beberapa negara di Timur Tengah, Eropa Barat Daya, Afrika Tropis,
Asia Timur dan Tenggara dan Australia, Begomovirus yang menyerang tanaman
tomat adalah Tomato yellow leaf curl virus (TYLCV) atau Tomato leaf curl virus
(ToLCV) (Zeidan et al. 1998). Sedikitnya 17 Begomovirus telah dilaporkan
menginfeksi tomat di daerah Amerika dan Karibia, seperti misalnya Texas pepper
virus, TYLCV, ToMoV, TGMV, Tomato yellow mosaic virus dan lain-lain.
Begomovirus ditularkan oleh serangga vektor kutu kebul (Bemisia tabaci,
ordo Hemiptera, famili Aleyrodidae) dengan cara persisten sirkulatif (Idris et al.
2001; Brown & Czosnek 2002). Periode makan akuisisi dan inokulasi
minimumnya telah banyak dilaporkan untuk banyak Begomovirus dan pada
umumnya masing-masing adalah 10-60 menit dan 10-30 menit (Idris & Brown
1998; Brown & Czosnek 2002). Periode laten virus ini di dalam vektornya lebih
dari 20 jam. Virus dapat bertahan di dalam vektor selama lebih dari 20 hari namun
tidak sepanjang masa hidup kutu kebul. Virus tersebut dapat dibawa oleh serangga
pada tahapan larva atau dewasa namun tidak diturunkan ke keturunannya.
Keragaman genetik dari Begomovirus
Begomovirus saat ini telah mendapat perhatian yang cukup serius.
Beberapa alasan yang mendasari hal ini adalah antara lain bahwa Begomovirus
(Harrison & Robinson 1999; Morales & Anderson 2001); pemanfaatannya sebagai
vektor dan induser pembungkaman gen (Atkinson et al. 1998; Kjemtrup et al.
1998); dan kontribusinya sebagai model untuk mempelajari mekanisme
pergerakan makromolekul secara intraseluler dan interseluler (Rojas et al. 1998;
Gutierrez 1999; Lazarowitz 1999).
Di samping itu, perhatian yang serius terhadap kelompok virus ini
dikarenakan oleh munculnya strain-starin Begomovirus baru melalui rekombinasi
dan pseudo-rekombinasi di antara strain dan/atau spesies pada berbagai tanaman,
peran dari komponen DNA-β seperti satelit virus dan penemuan adanya integrasi
sekuen Begomovirus ke dalam genom tanaman seperti pada spesies Nicotiana
(Navas-Castillo et al. 2000; Saunders et al. 2000; Harper et al. 2002; Ribeiro et al.
2002). Penemuan-penemuan ini mengindikasikan bahwa rekombinasi telah
berkontribusi terhadap keragaman genetik dari Begomovirus dan terhadap
munculnya varian-varian dan spesies virus baru. Adanya infeksi yang bersamaan
(mixed infection) dari dua atau lebih Begomovirus pada satu tanaman juga
merupakan aspek yang penting dalam memunculkan keragaman genetik dari
Begomovirus. Hal ini disebabkan karena infeksi yang bersamaan memberikan
pre-kondisi untuk terjadi rekombinasi yang dapat memunculkan strain virus baru yang
lebih ganas atau spesies Begomovirus yang baru (Sanz et al. 2000; Ribeiro et al.
2003).
Beberapa peneliti telah mempelajari adanya keragaman genetik dari
Begomovirus, diantaranya adalah keragaman genetik Begomovirus yang
menginfeksi kedelai, kacang-kacangan dan rumput-rumputan (Rodriguez-Pardina
et al. 2006), keragaman genetik pada infeksi campuran dari Begomovirus yang
menginfeksi tomat, cabai dan ketimun (Ala-Poikela et al. 2005; Ambrozevicious
et al. 2002), dan pada ubikayu (Bull et al. 2006).
Teknik deteksi dan identifikasi Begomovirus
Beberapa teknik telah digunakan untuk mendeteksi keberadaan dan
akumulasinya pada jaringan tanaman Begomovirus. Metode serologi atau
immunoasai seperti enzyme-linked immunosorbent assay (ELISA) telah menjadi
menggunakan antiserum yang disiapkan untuk mendeteksi virus tertentu.
Antiserum dengan bantuan bufer alkalin digunakan pada plate plastik mikrotiter
untuk menguji sap tanaman yang terinfeksi virus. Di antara jenis teknik ELISA
adalah double antibody sandwich-ELISA (DAS-ELISA) dan triple antibody
sandwich-ELISA (TAS-ELISA). Nono-Womdim & Atibalentja (1993)
menggunakan DAS-ELISA untuk mengidentifikasi PVMV pada cabai (Capsicum
annuum). TAS-ELISA menggunakan monoklonal antibodi untuk mendeteksi
virus, seperti Begomovirus pada tomat (Credi et al. 1989; Pico et al. 1999).
Teknik ELISA ini relatif lebih murah khususnya apabila antiserum dapat
diproduksi secara lokal dan juga cukup memadai untuk diagnosa virus. Namun
demikian, metode deteksi serologi mempunyai beberapa kelemahan diantaranya
adalah rendahnya titer dari antigen, adanya reaksi silang antibodi dengan antigen
heterolog dan adanya pengaruh pengaturan produksi antibodi oleh lingkungan dan
tahap perkembangan(Harrison 1991; Pico et al. 1999).
Teknik PCR adalah sebuah teknik molekuler yang sangat sensitif dan
spesifik untuk deteksi dan identifikasi patogen tanaman (Rojas et al. 1993), dan
teknik tersebut dapat digunakan untuk mempelajari dengan akurat komposisi
populasi patogen dan keragaman genetik virus (Gilbertson et al. 1991; Robertson
et al. 1991). Kespesifikan dari teknik PCR didasarkan pada penggunaan
primer-primer oligonukleotida yang komplementer dengan daerah yang diapit pada
sekuen DNA yang diamplifikasi. Karena PCR mengamplifikasi asam nukleotida,
teknik ini sangat bermanfaat untuk mengatasi kesulitan-kesulitan yang dihadapi
oleh metode deteksi serologi, seperti rendahnya jumlah antigen, reaksi silang dari
antibodi dengan antigen-entigen heterolog dan regulasi produksi antigen yang
dipengaruhi oleh tahap perkembangan atau lingkungan. Selain itu, dengan teknik
PCR, jumlah sampel DNA yang sedikit dari sampel tanaman yang segar, atau
disimpan dilemari es serta kering dapat digunakan untuk analisis PCR.
Metode PCR telah digunakan untuk mendeteksi dan menentukan
variabilitas genetik virus tanaman, termasuk diantaranya luteovirus (Robertson et
al. 1991), potyvirus (Langeveld et al. 1991), geminivirus yang ditularkan hama
wereng yang menginfeksi tanaman monokotil (Rybicki & Hughes 1990) dan
1992; Hidayat et al. 1999; Aidawati et al. 2005).
Rampersad & Umaharan (2003) bahkan telah mengembangkan suatu
teknik untuk mendeteksi Begomovirus menggunakan PCR. Ada tiga teknik PCR
yang digunakan yaitu PCR standar, PCR penempelan langsung (direct-binding
PCR) dan immunocapture PCR. Dari hasil penelitiannya menunjukkan bahwa
teknik immunocapture PCR yaitu teknik PCR yang menggunakan interaksi
antibodi-antigen untuk mengikat virus kemudian digunakan sebagai cetakan untuk
analisis PCR merupakan teknik yang paling efektif untuk mendeteksi
Begomovirus.
Teknik deteksi secara molekuler yang lain adalah teknik hibridisasi asam
nukleat. Teknik juga merupakan teknik yang sensitif untuk mendeteksi dan
mengidentifikasi Begomovirus pada tanaman yang terinfeksi (Pico et al. 1999;
Rodriguez et al. 2003). Hibridisasi asam nukleat menggunakan sebuah membran
nilon untuk memblot sap virus dan sebuah pelacak (probe). Teknik ini dapat
digunakan untuk mendeteksi virus dalam jumlah sampel yang cukup banyak
dalam waktu yang sama, namun biasanya teknik ini mempunyai banyak tahapan
yang harus dilakukan dan hanya dapat dilakukan di laboratorium yang
mempunyai fasilitas untuk itu.
Pemuliaan konvensional untuk ketahanan terhadap Begomovirus Kebanyakan kultivar-kultivar tomat komersial rentan terhadap infeksi
Begomovirus (TYLCV). Hal ini yang mendorong para pemulia untuk
mengembangkan tanaman tahan dengan memanfaatkan sumber gen ketahanan
dari spesies liar (Pilowsky & Cohen 2000). Sampai saat ini hanya satu gen
ketahanan mayor terhadap TYCLV yang telah diidentifikasi yaitu gen ty-1 (Zamir
et al. 1994) pada kromosom 6 dari Lycopersicon chilense. Dua gen ketahanan
yang lain telah dipetakan pada kromosom 3 dan 7 (Zamir et al. 1994) dari spesies
yang sama. Gen ketahahan terhadap TYLCV yang lain berasal dari L.
pimpinellifolium telah dipetakan menggunakan marka berbasis PCR RAPD pada
kromosom 6 namun berbeda lokus dengan ty-1 (Chague et al. 1997). Selain itu,
gen ketahanan terhadap ToLCV Taiwan dipetakan pada kromosom 8 dan 11 dari
tanaman tahan terhadap TYCLV menggunakan spesies liar yang berbeda seperti
L. peruvianum (Lapidot et al. 1997; Vidavsky & Czosnek 1998), L. chilense
(Scott et al. 1996), L. pimpinellifolium (Vidavsky et al. 1998), dan L. hirsutum
(Vidavsky & Czosnek 1998; Hanson et al. 2000).
Kultivar tomat komersial yang tahan TYLCV hasil pemuliaan
konvensional adalah TY20 yang membawa gen ketahanan dari L. peruvianum,
yang menunjukkan penundaan perkembangan gejala dan akumulasi virus
(Pilowsky & Cohen 1990; Rom et al. 1993). Pada kebanyakan kasus, sumber
ketahanan TYLCV dikendalikan oleh banyak gen (Pico et al. 1996; Pico et al.
1999). Setelah hampir 20 tahun, program pemuliaan secara konvensional hanya
menghasilkan sedikit kultivar tomat komersial yang ada di pasaran.
Rekayasa genetik untuk ketahanan terhadap Begomovirus
Rekayasa genetik untuk mendapatkan sifat ketahanan terhadap virus
biasanya menggunakan pendekatan konsep ketahanan yang berasal dari patogen
(pathogen-derived resistance, PDR) yang dikembangkan oleh Sanford & Johnson
(1985). Konsep strategi PDR ini didasarkan pada transformasi tanaman inang
yang rentan dengan gen yang berasal dari patogen itu sendiri. Ekspresi produk gen
tertentu dari patogen pada tanaman dapat mengganggu infeksi dari virus-virus
yang menginfeksi (Sanford & Johnson 1985). Keberhasilan pemanfaatan PDR
telah dilaporkan pada Begomovirus. Meskipun demikian, pada umumnya, sifat
ketahanan dengan level yang tinggi terhadap virus-virus DNA lebih sulit untuk
direkayasa.
Ketahanan terhadap TYLCV telah dihasilkan dengan menggunakan 5
strategi, yaitu i) ketahanan berdasarkan protein selubung (CP) (Kunik et al. 1994;
Bendahmane et al. 1997; Sinisterra et al. 1999), ii) ketahanan berdasarkan protein
movement (MP) (Malyshenko et al. 1993; Hou et al. 2000), iii) defektif
interferring DNA virus (Stanley et al. 1990), iv) gen-gen dengan orientasi
antisense (Day et al. 1991; Bendahmane & Gronenborn 1997) dan gen replikase
(rep, C1, AC1) yang terpotong (truncated) (Noris et al. 1996) dan hasil mutasi
(Yang et al. 2004). Pendekatan berdasarkan CP dan MP melibatkan ekspresi dari
Sedangkan strategi yang lain, meskipun berbeda dalam konstruk gen yang
digunakan, semua bertujuan untuk menghalangi replikasi dari virus dengan
meng-nonaktifkan gen Rep. Yang et al. (2004) telah berhasil merekayasa tomat tahan
TYLCV menggunakan konstruk transgen Rep dan C4 yang menunjukkan tidak
adanya DNA virus dan gejala yang dapat diamati pada tanaman tomat
transforman.
Dari hasil penelitian, dari pendekatan PDR yang dilakukan (terutama
untuk non-begomovirus), ketahanan tanaman transgenik yang diperoleh
disebabkan oleh ekspresi sekuen transgen virus pada tahap transkripsi dan bukan
pada tahap translasi (Chellappan et al. 2004; Vanitharani et al. 2004). Mekanisme
yang mendasari kasus ini adalah adanya pembungkaman RNA (RNA silencing)
atau interferensi RNA (RNA interference, RNAi), sebuah mekanisme
penghancuran sekuen spesifik pada tanaman yang menggambarkan mekanisme
pertahanan antivirus secara alami (Voinnet 2001; Vanitharani et al. 2003;
Chellappan et al. 2004). Karena Begomovirus mempunyai genom DNA maka
prospek penggunaan pendekatan berdasarkan RNAi masih terbatas.
Pembungkaman RNA berdasarkan transgen pada gen Rep dan C4 belum begitu
berhasil. Studi ini menunjukkan bahwa jika virus mencapai level threshold dari
ekspresi replikasi pada sel-sel yang terinfeksi awal, maka penyebaran virus tidak
dapat lagi dihalangi (Noris et al. 2004).
Sekuen Begomovirus yang tidak menyandikan protein (IR) juga telah
diteliti untuk menghasilkan ketahanan terhadap virus. Pooggin & Hohn (2003)
menjelaskan bahwa ekspresi baik sense dan antisense sekuen promoter dari Vigna
mungo yellow mosaic virus (VMYMV) pada IR menghasilkan ketahanan pada
III. DETEKSI BEGOMOVIRUS YANG MENGINFEKSI
TOMAT MENGGUNAKAN TEKNIK POLYMERASE CHAIN
REACTION (PCR)
Abstrak
Infeksi Begomovirus telah dilaporkan dan ditemukan pada beberapa tanaman sayuran penting seperti tomat dan cabai. Namun demikian, informasi mengenai deteksi Begomovirus yang menginfeksi tanaman tomat pada beberapa area produksi tomat menggunakan teknik PCR belum banyak dilaporkan. Tujuan penelitian adalah untuk mendeteksi Begomovirus yang menginfeksi tanaman tomat pada beberapa daerah area produksi tomat di Jawa Timur, Jawa Tengah, Daerah Istimewa Jogjakarta dan Jawa Barat menggunakan teknik polymerase c