FILOGENI KERBAU RAWA
(Bubulus bubalis)
BERDASARKAN GENOM MITOKONDRIA
SYLVIA NOVITA PALUPI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAIIUAN ALAM
INSTITUT PERTANIAN
BOGOR
ABSTRAK
SYLVIA NOVITA PALUPI. Filogeni Kerbau Rawa (Bubalus bubalis) berdasarkan Genom Mitokondria. Dibimbing oleh ACHMAD FARAJALLAH dan DYAH PERWITASARI.
Kerbau yang ada saat ini merupakan hasil domestikasi kerbau liar Asia, Btrbalus arnee.
Kerbau basil domestikasi dibedakan menjadi dua jenis, yaitu kerbau rawa dati kerbau sungai. Keduanya berbeda baik secara fisik, kariotipe dan pola penyebar'mya. Penelitian ini dilakukan untuk mengetahui posisi filogeni B. bubalis dalam Bovidae dan mempelajari pola penyebarannya berdasarkan genom mitokondria. Sampel yang digunakan adalah kerbau rawa lokal yang berasal dari Sumatera Utara, Banten, Jawa Tengah dan NTB. Amplifikasi dilakukan secara in vilro dengan teknik Polymerase Chain Reaction (PCR). Produk amplifikasi sebesar 1145 pb dirunutkan dan dianalisis filogeninya. Empat dari delapan sampel berhasil diamplifikasi yaitu kerbau asal Sumatera Utara, Banten dan NTB. Semua sampel kerbau rawa lokal mengelompok dalam subfamili Bovinae dali famili Bovidae .Pola penyebaran kerbau mengikuti pola ekspansi pertanian manusia dari daratan Cina, tempat nenek moyang kerbau, menuju ke semanjung Malaya (Sumatera Utara) dan ke arah Jawa kemudian tnenuju ke Indonesia Timur. Hal ini merupakan penjelasan bahwa sampel asal Sumatera Utara adalah haplotipe yang berbeda tlari sanlpel asal Banten dan NTB berdasarkan aualisis ~ne~lggunakan dua basa pertama pada tiap kodon C:yt b.
ABSTRACT
SYLVIA NOVITA PALUPI. Phylogeny of Swamp Buffalo (Btrbnhrs bubnlis) Based on Mitochondrial Genome. Supervised by ACHMAD FARAJALLAH and D Y N I PERWITASARI.
FILOGENI KERBAU
RAWA
(Bubalus bubalis)
BERDASARKAN GENOM MITOKOmEUA
SYLVIA NOVITA PALUPI
Skripsi
sebagai salah satu syarat
untukme~nperoleh
g e l a
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAEIUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul Skripsi : Filogeni Kerbau Rawa (Bubal~is bubalis) berdasarkan Genom Mitokondria
1
Nama : Sylvia Novita PalupiNIM : G34050562
1
~
Dr. Ir. Achmad Farajallah, M.Si NIP. 19650427 199002 1002
Menyetujui:
Dr. Ir. R. R Dyah Perwitasari, M.Sc NIP 1
a
660403 199003 2 001$.
Dekau Fakultas Mate~natika dan Ilmu Pengetahuan Alan Institut Pertanian BogorPRAKATA
Puji dan syukur penulis panjatkan kepada Allah swt. atas segala karunia dan berkat-Nya sehingga karya ilmiah ini berhasil diselesaikan.
Penulis mengucapkan terima kasih kepada Bapak Dr. Ir. Achnad Farajallah M.Si dan Ibu Dr. Ir. R. R. Dyab Penvitasari, M.Sc. Di sampinbg itu, penghnrgaan penulis sampaikan kepada staf pengajar Zoologi, kepada Pak Chairul, Bu Bibah, Mbak Tini, Mbak Ani, I'ak Adi, Kak Wildan, Kak Erik, Kak Ogi, Kak Ruth, Kak Uce, Ichay, Jazy, Dewi, Ika, Prista serta teman-teman Biologi khususnya Bio 42 atas persahabatan yang indah selama ini. Ucapan terima kasih tak terhingga penulis berikan untuk Mama, Papa dan Mas Panji atas segala doa, kasih saynng, dan dukungannya kepada penulis.
Semoga kalya ilmiah ini dapat bermanfaat bagi pengembangan ilmu pengetahuan.
RIWAYAT HIDUP
Penulis dilahirkan di Jember pada tanggal 28 November 1986 dari ayahanda Supamo, SE dan ibunda Subaida. Penulis merupakan putri tunggal.
Tahun 2005 penulis lulus dari SMAN 1 Jember dan pada tal~un yang sanla lulus Undangan Seleksi Masuk IPB (USMI). Penulis memasuki Program Shldi Biologi, Falmltas Mateinatika dan Ilmu Pengetahuan Alam (MIPA). Selama mengikuti perkuliahan, penulis pemah menjadi asisten praktikum mata kuliah Biologi Dasar pada tahun ajaran 2006/2007 dan 200812009,
Stmktur Hewan pada tahun ajaran 200812009 dau pengantar Genetika Molekuler pada tahun ajaran
DAFTAR IS1
DAFTAR
TABE
viii ...DAFRAT GAMBAR VIII
...
DAETAR LAMPIRAN
...
VIIIPENDAHULUAN
...
...
Latar Belakang
.
.
1Tujuan
...
1 BAHAN DAN METODEHASIL
Alnplifikasi dan Visualisasi Fraglnel~ DNA 2
Pelunutall Produk PCR
...
3Analisis Filoge~ 3
PEMBAFIASAN
5 5
G
SIMPULAN
...
7S A R A N
...
.
.
.
.
... 7DAPTAR TABEL
. .
1 Panjang mas hasil pensejajaran (nt)
...
3...
...
2 Hasil pensejajaan nukleotida DNA yang berheda
.
.
.
33 Nilai jarak genetik (bawah diagonal) dan standar eror (atas diagonal) antar B. bttbalis
...
dari beberapa lokasi berdasarkan metode Kimttra-2-paranieter 5DAFTAR GAMBAR
Halaman
1 Hasil an~plifikasi luas D-loop dalanl PAGE 6% dengan pewarnaan sensitif pcrak ... 2
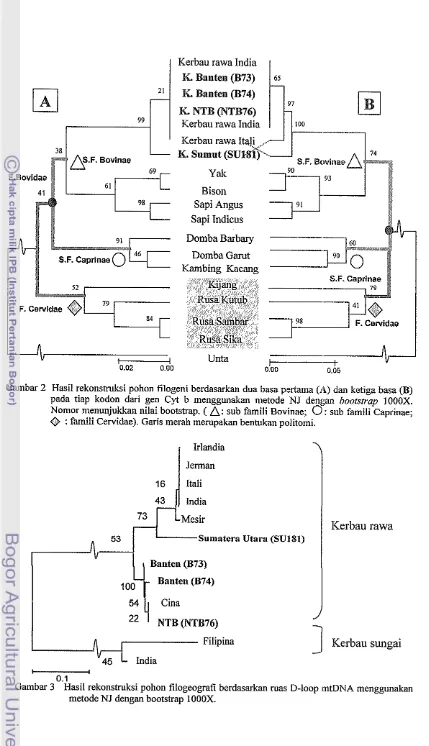
2 Hasil I-ekonstruksi pohon filogeni herdasarkan dua basa pertaina (A) dan ketiga basa (B) pada tiap kodon dari gen Cyt b tnenggunakan n~etode NJ dengan bor~lstrrrp 1000X. 4
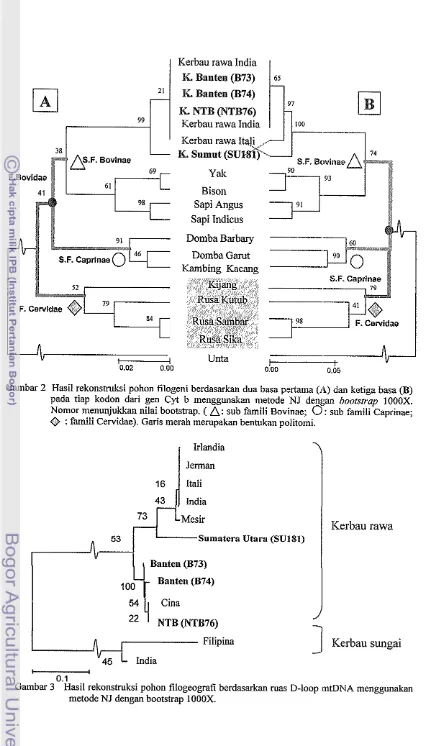
3 Hasil rekonstruksi pollon filogeografi berdasarkan ruas D-loop mtDN.4 menggunakan metode NJ dengan bootstrap 1000X ...
.
.
.
...
4DAFTAR LAMPIRAN
f l a l a r n a ~ ~
1 Spesies pelnbanding dalaln analisis filogeni yang diperoleh da1-i GuleB017k ... 10
2 EIasil penjajaran DNA ruas Cyt b dan D-loop genonl nlitokondria kerbau rawa
(B. bubalis)
...
.
.
.
...
11PENDAHULUAN
L a t a r Belakang
Kerbau (Bubalus bubalis) me~upakan anggota Famili Bovidae, Subfaniili Bovinae, genus Bubalus (Williamson & Payne 1993). Bubalus memiliki morfologi berbeda dari Bovinae lailmya yaitu Bos, Bison, Pseudoryx, dan Syncerus. Tanduknya cenderung bersudut bahkan membentuk sabit. Selain itu, telinganya berukuran kecil dan tulang tengkoraknya panjang dan senipit (Willianlson & Payne 1993).
Ulrbalus b ~ ~ b u l i s dikenal sebagai kerbau Indian yang diduga hasil dotnestikasi kerbau liar Asia, Bubalis anlee (Lau el a1 1998). Kerbau hasil domestikasi bisa dibedakan menjildi kehau rawa (swanzp bufiff dan kcrbau sungai (rh~er b~rffnlo). Kerbau rawa ~ilenliliki kulit benvarna hitam abu-abu dengan tanda putih abu-abu pada lipatan bagiati bawah leliernya dan tanduk yang tunlbuh ~nelengkung ke belakang seperti sabit. Tubuh kerbau sungai berukuran lebih besar dibandingkan kerbau rawa, kulit dominan berwarna hitam dengan tanda putih di bagian dahi dan ujung ekor, dan tanduk berukuran relatif pendek, melengkung ke depan, ke bawah atau ke belakang. Selain itu, jumlah krotnosom kerbau rawa 2n=4S sedangkan kerbau sungai 2n=50 (FIilmi 1991). Pola penyebaran keduanyapun berbeda. Kerbau rawa tersebar di daratan India, Timur Tengah dan Eropa bagian barat, sedangkan kerbau sungai tersebar di Cina, Bangladesh, negara- negara di Asia Tenggara dan Barat daya India. Filogeni adalah sejarah mengenai garis evolusi suatu kelo~npok organisme. Analisis filogeni pada satu spesies yang nienyebar nlengikuti pola geografi dikenal dengan istilah filogeografi. Salah satu penanda molekular yang banyak digunakan dalam analisis filogeui maupun filogeografi adalah genoni mitokondria (mtDNA) (Coccone 1999). Perbedaan penggunaan mtDNA baik untuk analisis filogeni maupun filogeografi adalah pada karakter laju mutasi autar bagian-bagian mtDNA. Bagian mtDNA yang paling stabil dan banyak digunakan untuk menganalisis filogeni pada tingkat interspesies adalah tRNA dan rRNA. Bagian genom yang menyandikan protein yang paling sering digunakan untuk menganalisis filogeni anggota taksa dibawah ordo atau famili adalah Cyt b. Bagian D-loop dari genom adalah yang paling cepat laju mutasinya sehingga sangat populer digunakan untuk menganalisis
keragaman intraspesies atau filogeografi (Farajallah A 8 Juni 2009, komunikasi pribadi). Ruas genonl mitokondria yang banyak digunakan sebagai penanda molekuler untuk menganalisis filogeni pada B~rbalus yakni Cyt b (Lau et ul. 1993, Hassanin & Douzety 1999) dan D-loop (Lau et al. 1993).
Parma et nl. (2004) menjabarkan posisi filogenetik Bubalus dalam kelima matnalia lainnya, yaitu sapi (Bos inclicus), kambing, domba, rusa (Mcuitiacus nzwztjak) dan Ilama (Lanta pacos) berdasarkan 13 gen penyandi protein mtDNA. Bubalus berkerabat dekat dengau sapi sebagai ang, aota run~inausia besar. ICeduanyn niengelo~i~pok dengan kambing dan domba sebagai anggota dari mminansia kecil. Elubungan kekerabatan Bubalus jauh dengan rusa dati Ilama.
Tujuan
Penelitan ini be]-tujuan mempelajari posisi filogeni B. b~fbalis dalanl Fnniili Bovidae dari berbagai daerah di Indonesia dan menipelajari pola penyebal-annya berdasarkan genom mitokondria.
BAHAN DAN METODE
Bahan
Sampel darah kerbau rawa yaug digunakan dalam penelitian ini adalah sampel darah yang diawetkan dalam alkohol 70% koleksi Bagian Fungsi Hnyati dan Perilaku Hewan Departemen Biolog~. Sebailyak delapan sampel dipilih dari keenlpat lokasi yang berbeda, yaitu Surnatera Utara (SU181 dan SU182), Nusa Tenggara Barat (NTB)
(NTB76 dan NTB77), Banten (B73 dan I374) dan Jawa Tengah (JT38 dan JT39). Sa~npel asal Suniatera Utara dipilih berdasarkan infonnasi yang diberikan ole11 Ta~winangsih (2009) sebagai haplotipe yang berbeda berdasarkan metode PCR-RFLP.
Waktu dan Ternpat P e ~ ~ c l i t i a ~ l
Penelitian i ~ i i dilaksanakan pada bulan Feb~uari sampai dengan Mei 2009 bertempat di Laboratoriu~u Bagian Fungsi Hayati dan Perilaku Hewan Departemen Biologi, Institut Pertanian Bogor, Bogor.
Metode
Ekstraksi dan Isolasi DNA
yang sudah bersih dari alkohol kemudian disuspensikan dalam bufer STE (NaCI lM, Tris-HCI lOmM, EDTA 0,) mM, pH 8,O) sampai volume 350 p1. Sel-sel darah dilisis dengan SDS 1% dan proteinase K 0,125 mglml, kemudian dimkubasi pada suhu 55'C selama 1 jam sambil dikocok pelan. Metode ekshaksi DNA selanjutnya mengikuti petunjuk Genolnic DNA mini kit (Geneaid) for blood.
Amplifikasi dan Visualisasi Fragmen DNA Amplifikasi genom mitokondria menggunakan pasangan primer koleksi Dr.Ir. Achmad Farajallah, M. Si, yaitu AF22 (5'- GCGTACGCAATCTTACGAT CA-3') yang menempel pada basa ke-15357 dan AF23 (5'-
ATGCAGTTAAGTCCAGCTAC-3') yang
menempel pada basa ke-143. Pasangan primer ini mengapit mas D-loop mulai dari bagian akhir gen Cyt b sampai ke bagian akhir D- loop dengan panjang 1145 pb. Knmposisi reaksi PCR dengan volume 25 p1 terdiri atas sampel DNA 2 PI, Taq Polymerase (RBC) 0.75 unit beserta bufernya, MgC12 0.2 mM,
dNTPs mix 0.2 mM dan masing-masing primer 1
nM.
Reaksi PCR dilakukan menggunakan mesin Thern~ocycler TaKaRa MP4 dengan kondisi sebagai berikut: tahap denaturasi awal pada suhu 94 OC selama lima menit;
tahap kedua yang terdiri dari 30 siklus yang masing-masing siklus terdiri dari denaturasi pada suhu 94 OC selama satu menit,
penempelan primer pada suhu 59 OC selama dua menit dan pemanjangan pada suhu 72 OC
selama dua menit; tahap terakhir yaitu pemanjangan akhir pada suhu 72 OC selama sepuluh menit. Kualitas produk PCR diuji menggunakan PAGE 6% dalam bufer 1X TBE (10 mM Tris-HCI, 1 M asam borat, 0.1 mM EDTA) yang dijalankan pada kondisi 180 mV selama 40 menit. Selanjutnya DNA diwarnai dengan pewamaan sensitif p e r k (Tegelshom 1986).
Perunutan Prnduk PCR
Produk PCR bempa pita tunggal berukuran sesuai desain primer dimurnikan dengan QIAQuick PCR Purification Kit
(QIAGEN). Produk PCR yang sudah diiumikan kemudian dijadikan sampel dalam reaksi PCR untuk perunutan nukleotida dengan metode Dye Dideoxi Terminator
menggunakan primer AF 22 (PT. CHAROEN POKPHAND INDONESIA). Perunutan nukleotida menggunakan mesin ABI Prism 3700-Avant Genetic Analyzer.
Anaiisis Filogeni
Runutan nukleotida yang dipcroleh diedit secara manual lnenggunaban program Bioedit versi 7.0.9.0 berdasarkan pada kromatogram hasil perunutan. Runutan nukleotida yang telah diedit disejajarkau dengan urutan nukleotida beberapa spesies lain yang bomolog dari GeneBank (Lampiran 1). Proses pensejajaran dilakukan menggunakan program Clustal W1.8 dalam MEGA 4.0 yang kemudian diedit secara manual. Perhitungan komposisi nukleotida, tipe subtitusi, jarak genetik dan konstruksi pohon filogeni dilakukan menggunaka~i program MEGA 4.0 dengan metode Ki~ntrra-2-parameter.
Rekonstruksi pohon filogeni berdasarkan mas Cyt b, sedangkan rekonstmksi pohon filogeogmfi bet'dasarltan mas D-loop. Rekonshuksi keduanya menggunakan metode
Neighboor Joining (NJ) dengan bootshap lOOOX (Nei & Kumar 2000).
HASIL
Amplifikasi &an Visualisasi Fragmen DNA Dari kedelapdn sampel yang digunakan, sebanyak empat san~pel yang berhasil diamplifikasi, yaitu sampel kerbau rawa asal Sumatera Utara (SUI&l), NTB (NTB76), dan Banten (B73 dan B74). Amplifikasi menggunakan pasangan primer AF22 dan AF23 menghasilkan pita tunggal bem6mn sekitar 1145 pb (Gambar 1). Sampel asal Sumatera Utara (SU182), NTB (NTB77) dan Jawa Tengah (JT38 dan JT39) tidak berhasil diamplifikasi sehingga sampel ini tidak dilakukan perunutan nukleotida.
M I 2 3 4
Perunutan Produk PCR
Perunutan DNA dilakukan dari arah primer
AF
22. Panjang DNA hasil perunutan setelah diedit bervariasi yakni 572 nt untuk sampel SUl 81, 563 nt untuk sampel NTB76, 525 nt untuk sampel B73 dan 527 nt unhlk sampel B74.Analisis Filogeni
Proses pensejajaran menghasilkan runutan sepanjang 516-518 nt dengan rata-rata komposisi nukleotida A=35.6%; T=23.9%; G=13.5%; dan C=27%. Presentase A+T (59.5%) lebih besar daripada C+G (40.5%). Berdasarkan hasil pensejajaran dengan data dari GeneBnnk (Lampiran 1) dapat diketahui bahwa hasil perunutatl keempat san~pel tersebut n~eliputi enlpat domain, yaitu bagian akhir Cyt b (203 nt), tRNA Tlx (70 nt), tRNA Pro (65 nt) dan bagian awal D-loop (178-180 111) (Tabel 1). Panjang gel1 Cyt b dan tRNA stabil antar satnpel sedangkan paiijang D-loop beragam karena terdapat insersi pada basa ke- 159 sa~npel asal NTB (NTB76) atau delesi pada basa ke-67 sampel Sumatera Utara (SU18 1) (lainpiran 2).
Ruas yang digunakan untuk analisis filogeui
B. bllbnlis adalah gen Cyt b. Hasil peusejajaran dengan data dari GnleBank
menunjukkan 172 nt yang identik dan 31 nt
yang berbeda. Basa yang berbeda disebabkan ole11 substitusi basa, baik transisi maupun transversi (Tabel 2). Contoh transisi pada luas Cyt b menyebabkan pe~ubahan asatn amino Met menjadi Val pada basa ke-174 dan Ile menjadi Val pada basa ke-177, sedangkan transisi pada basa ke-44 tidak merubah asam amino dimungkinkan karena terjadi pada basa ketiga dari kodon.
Topologi pohon filogeni hasil NJ dua basa pertama pada tiap kodon menunjukkan Hasil rekonstritksi berdasarkan dua basa pertama dari tiap kodon Cyt b tnetigelolnpokkan Subfamili Capriliae dan Subfamili bovinae dalam salu percabangan melnbentuk kelounpok Fanlili Bovidae. Fa~nili Cervidae berada di luar percabangan liienlbentuk sister grorrp dengan Falnili Bovidae (Ganlbar 2A). Lain halnya dengan hasil rekonstruksi herdasarkan tiga basa dal-i tiap kodon yang ~uembcntuk politotiii antara Subfatuili Bovinae, Subfamili Caprinae dan Fan~ili Cervidae (Gamnbar 28).
Secara filogeografi, sample kerbau rawa Sulilatera Utara te~pisah detigan ketiga sampel kerbau rawa lainnya dan n~engelonlpok dengan kerbau dari India, Jern~an, Irlandia, Itali dan Mesir ((ianlbar 3). Sa~npel kerbau rawa asal Banten dan NTB ~nengelompok dengan sampel kerbau dari Cina.
Tabel 1 Panjang mas hasil pensejajaran (nt)
Satnpel keibvu Cyt-b tRNA Thr IRNA Pro D-loop Pnnjang scluruh nl:s
Surnateta Utv~a (SUISl) 203 70 65 178 516
NTB (NTB76) 203 70 65 I SO 518
Bnnte~l (873) 203 70 65 179 517
Bullle11 (874) 203 70 65 179 517
Tabel 2 Hasil pensejajaran nukleotida DNA yang berbeda
Cyt-b D-loop
I I 1 1 1 1 I I I I 1
Sampel kerbau 4 7 7 1 1 1 2 4 4 5 6 6 6
4 4 7 3 6 4 8 3 6 9 9 1 4 8
Sumaten Utan (SU18 1) C G G
ctfb
T : ~ G G C G T CNTB (NTB76) T A A T;C,~A c~$,:A A T A
.r
TBallten (B73) T A A T ' G A C , C A A T A T T
Banten (B74) T A A
TMA
CiktA A , ~ A C TKerbau rawa India
K. NTB (NTB76)
Kerbau rawa IndiaDomba Gantt
Karnbing Kacang
0.02 0.00 0.00 0.05
Gambar 2 Hasil rekonstruksi pohon filogeni berdasarkan dua basa pertama (A) dan ketiga basa (B) pada tiap kodon dari gen Cyt b menggunakan metode NJ dengan bootstrap 1000X. Nomor menunjukkan nilai bootstrap. (
A:
sub famili Bovinae;0
: sub famili Caprinae;'@ : famili Cewidae). Garis merah merupakan benhlkan politomi.
\
Jerman
Mesir
Sumatera Utara (SUISI)
Banten (B73)
Banten (B74)
Kerbau
rawa
/J
Filipina
Kerbau sungai
-
0.1 [image:12.595.82.511.48.794.2]I
Tabel 3 Nilai jarak genetik (bawah diagonal) dan kesamaan genetik (atas diagonal) antar B. bubalis dari beberapa lokasi berdasarkan metode Ki1nu1-u-2-para1neier
I 2 3 4 5 6 7 8 9 10 I 1 12
I 0.907 0.902 0.082 0.168 0.941 0.994 0.989 1.000 0.909 0.909 0.909
2 0.093 0.994 0.012 0.111 0.919 0.976 0.907 0.975 1.000 1.000 1.000
3 0.098 0.006 0.063 0.151 0.908 0.909 0.903 0.902 0.994 0.994 0.994
4 0.918 0.988 0.937 0.888 -0.084 0.082 0.063 0.082 0.040 0.040 0.040
5 0.832 0.889 0.849 0.112 0.080 0.168 0.151 0.168 0.132 0.132 0.132
6 0.059 0.081 0.092 1.084 0.920 0.947 0.941 0.941 0.814 0.814 0.814
7 0.006 0.087 0.091 0.918 0.832 0.053 0.994 0.994 0.915 0.915 0.915
8 0.011 0.093 0.097 0.937 0.849 0.059 0.006 0.989 0.909 0.909 0.909
9 0,000 0.093 0.098 0.918 0.832 0.059 0.006 0.011 0.909 0.909 0.909
10 0.091 0.000 0.006 0.960 0.868 0.086 0.085 0.091 0.091 1.000 1.000
11 0.091 0.000 0.006 0.960 0.868 0.086 0.085 0.091 0.091 0.0011 1.000
12 0.091 0.000 0.006 0.960 0.868 0.086 0.085 0.091 0.091 0.000 0,000
Keterangan : (1) Cina, (2) Irlandia, (3) Mesir, (4) Filipina, (5) Itidia, (6) Sumatera Utara, (7) Banten (73), (8) Banten (74), (9) NTB, (10) India, (1 1) Italia, (12) Jemian.
G=13.5%; dan C=27%. Presentase A t T (59.5%) lebili besar daripada C+G (40.5%). Berdasarkan hasil pensejajaran deilgan data dari GeneBunk (Lampiran 1) dapnt diketahui bahwa hasil pemnutan keempat sanipel tersebut nieliputi empat domain, yaitu bagian akhir Cyt b (203 nt), tRNA Thr (70 tit), tRNA Pro (65 nt) dan bagian awal D-loop (178-180 nt) (Tabel 1). Panjang gen Cyt b dan tRNA stabil antar sampel sedangkan paiijang D-loop beragani karena terdapat insersi pada basa ke- 159 satlipel asal NTB (NTB76) atau delesi pada basa ke-67 sa~npel Sumatera Utara (SU181) (lainpiran 2).
Ruas yang digunakan untuk analisis filogeni
B. b~tbalis adalah gen Cyt b. Hasil pensejajaran dengan data dari GeneBunk
nienunjukkan 172 nt yang identik dan 31 nt yang berbeda. Basa yang berbeda disebabkan oleh substitusi basa, baik tra~isisi niaupun transversi (Tabel 2). Contoh transisi pada mas Cyt b inenyebabkan pe~ubalian asam ainino Met menjadi Val pada basa ke-174 dan Ile menjadi Val pada basa ke-177, sedangkan transisi pada basa ke-44 tidak inerubah asam amino dimungkinkan karena terjadi pada basa ketiga dari kodon.
Topologi pohon filogeni hasil NJ dua basa pertama pada tiap kodon menunjukkan Hasil rekonstruksi berdasarkan dua basa pertama dari tiap kodon Cyt b mengelompokkan Subfamili Caprinae dan Subfamili bovinae dalam satu percabangan membentuk kelompok Famili Bovidae. Fa~nili Cervidae berada di luar percabaigan membentuk sister group dengan Famili Bovidae (Gambar 2A). Lain halnya dengan hasil rekonstruksi berdasarkan tiga basa dari
tiap kodon yang ~nenibentuk politoiiii autara SubCamili Bovinae, Subfamili Caprinae dan Faulili Cervidae (Gamb;~r 2B).
Secara filogeografi, sampel kerbau rawa Surnatera Utara terpisah dengan ketiga sampel kerbau rawa Iainnya dan rnengelompok dengan kerbau dari India, Jcrman, Irlandia, Itali dan Mesir (Gambar 3). Sanipel kerbau rawa asal Ballten dan NTB ~nengelompok dengan sa~npel kerbau dari Cina.
PEMBAIIASAN
An~plifikasi dan Visualisnsi Fragmen DNA Sampel kerbau rawa asal Sumatera Utara, NTB, dan Ballten berhasil dianiplifikasi menggunakati pasangan primer AF22 dan AF23. Pita tunggal yang dihasilkan sesuai dengall hasil a~nplifikasi desaiu primer bemkuran sekitar 1145 pb (Gambar I).
Keetnpat sanipel lai~mya tidal< berhasil dianiplifikasi (38, 39, 77 dan 182) kemunglcinan karcna subu penempelan primer ( 5 9 ' ~ ) yang tidak sesuai sekalipun sampel yang digu~iakan merupakan spesies yang sama.
Perunutan Produk P C R
dimnutkan seperti pada karakterisasi genom mitokondria Platypus (Gemmell et a1 1994).
Analisis Filogeni
Rnas mtDNA harus perunutan mayoritas tersusun atas basa A+T. Banyaknya basa A t T pada D-loop erat kaitannya dengan karakter D-loop yang memiliki banyak situs awal replikasi untuk utas berat (Heavy slrand) mtDNA dan promotor bagi utas berat tnaupun utas ringan (Light strand) sebagai titik awal transkripsi (Hoelzel AR et (11. 1994).
Panjang mas Cyt b stabil pada keempat sample dan memiliki tingkat variabilitas yang rendah. Hal ini terlihat dari sedikitnya situs yang mengala~ni subtitusi (Tabel 2).
Transversi lianya dite~nukan pada luas D-loop dengan rasio transisi terhadap trausversi (tsltv) sebesar 4.5. Hal ini menunjukkan baliwa subtitusi tranversi jarang terjadi dibandingkan subtitusi twlsisi. Hasil ini sejalan dengan yang dikemukakan oleh Hixson & Brown (1986) bahwa pada 1ntDN.4 transisi lebih sering terjadi dibandingkan transversi.
Hasil rekonstruksi pohon filogeni herdasarkan urutan nukleotida Cyt b dilaknkan dengan membandinglcan dua basa pertanla dan ketiga basa pada tiap kodon. Rekonstmksi pollon filogeni menggunakan dua basa pertama pada tiap kodon lebih tepat menjelaskan kedudnkan Famili Cervidae yang merupakan sistei. group dari Famili Bovidae (Gambar 2A) sesuai dengan klasifikasi Williamson & Payne (1993) dan Kuznetsova el a[. (2004). Rekonstruksi pohon filogeni menggunakan ketiga basa pada tiap kodon menghasilkan politotni antara Subfamili Bovinae, Subfa~nili Capriiiae dan Famili Cervidae (Gambar 20). Politotiii mengindikasikan adanya liubungan filogenetik yang tidak pasti (politon~i lunak). Perbedaan hasil analisis antara dua basa pertama dan ketiga basa dari tiap kodon dimungkinkan karena posisi basa ketiga pada tiap kodon Cyt b yang tidak stabil. Basa ketiga pada tiap kodon tidak stabil karena sering mengalami transisi (Tanaka et a/. 1994; Matthee & Davis 2001). Kejadian transisi pada basa ketiga dari tiap kodon tidak berpengamh terhadap penyandian asam amino Transisi pada basa ketiga dari tiap kodon Cyt b (Tabel 2) merupakan mutasi bisn yang tidak berpengaruh terhadap penyandian asam amino. Basa pada Cyt b yang dibandingkan merupakan sebagian dari mas Cyt b yang seharusnya berukuran
2
1100 nt sehingga asumsi bahwa transisi pada basa ketigamelupakan jenis mutasi bisu belum kuat dibuktikan.
Kerbau rawa asal Sumatera Utara lebih berkerabat dekat dengan kerbau dari Irlandia, Jerman, Itali, Mesir dan India dengan rata-rata jarak genetik sebesar 0.0865 atau tingkat kesamaan gennya sebesar 91.35%. Kerbau rawa asal NTB dan Banten memiliki kedekatan dengan kerbau dari Cina dengan tingkat kesamaan gen sebesar 100% dan 91.5% (Tabel 3). Kerbau rawa asal Sumatera Utara mernang merup;lkan haplotipe yang berbeda dari kerbau rawa asal Baiiten dan NTB, namun ketiganya tetap mengelotnpok dala~n Subfamili Bovinae dengan nilai bootstrap sebesar 99%. Hal ini ~nemunculkan dugaan bahwa kerbau rawa asal Sumatera Utara berasal dari nenek ~noyang yalig salna dengan sampel asal Batilen daii NTB.
Pola persebaran kerbau yang luas diduga tnengiknti jalur niigr;isi inanusia karena kerbau merupakan lieu,an ~uminansia besar yang mernbutuhki~n media untuk migrasinya. Pola migrasi ~iianusia yang dijadikan acuali yakni pada masa menetap dan bercocok tanam yang mempakan penanda beralihnya kebudayaan berburu dan mengumpulkan makanan dari zaman batu lama (Paleolitikum) ke zaman batu baru li\ieolitikum). Daerah pertanian awal yang meliputi tiga area, yakni Asia barat daya, Cina dan Mesoamerica (Bellwood 2001) diduga tnerupakan awal tempat domestikasi hewnn, tern~asuk kerbau.
Cockrill (1981) berasumsi bahwa tempat domestikasi kerbau berawal di pusat peradaban India yakni daerah Yangtze, Eupluates and Tigris sekitar 5000 tahun lalu. Berbeda dengan asumsi Chen & Li (1989) yang menyebutkan baliwa Cilia merupakan tempat awal donlestikasi kerbau sejak 6000 tal~nn yang lalu. Kedua lcontadibsi iiii menguatkan dugaan ICu~uar et nl (2007) bahwa kedua tipe kerbau merupakan hasil dotnestikasi dari nenek nloyang yang sama dan domestikasi kednanya dilaknkan secara terpisah.
Sumatera Utara berbeda dengan sampel kerbau rawa asal Banten dan NTB. Meskipun berbeda dalam ha1 proses domestikasi lebih lanjut, kerbau rawa asal Sumatera Utara, Banten, dan NTB tetap berasal dari nenek moyang yang sama di Cina. Jawa (Banten) dan Indonesia Timur (NTB).
Penyebaran hewan domestikasi memiliki kecendemngan menghimdari hariel. geografi. Kerbau dari Cina tidak menyebar ke arah utara Asia karena adanya Gurun Gobi dan tidak adanya ekspat~si dari J a m ke Sumatera karena di Sumatera terdapat pegunu~lga~l Bukit Barisan.
Kerbau berkenlbang di daerah dengan agroekosiste~n yang bemariasi seperti padang penggemhalaa~l dengan iklim kering di NTB, NTT di daerah persawahan irigasi ~naupull non irigasi (Banten); di daerah pegunungan (Sumatera Utara) dan dataran rendah yang berawa-rawa seperti di daerah Kalimantan Selatan. Hal ini menunjukka~~ bahwa daya adaptasi kerbau pada berbagai koi~disi agroekosistem sangat besar. Sebagai akibat pengamh lingkungan telah terjadi semacam evolusi sehingga timbul semacam sub gmp pada kerbau di Indonesia, seperti (1) timbulnya kerbau-kerbau yang berbadan besar dan yang berbadan kecil, (2) perbedaan terhadap daya tahan terhadap panas, dan (3) kegenlaran hidup di dalam air (Fahimuddin 1975).
Hubuugan filogeni berdasarkan gen Cyt b rnenempatkan Bubalus bubalis sebagai anggota Subfamili Bovinae, Fa~nili Bovidae. Analisis filogeni n~enggut~akan d ~ ~ a basa pertama pada tiap kodot~ Cyt b lehi11 stabil dibandingkan ketiga basa pada tiap kodon Cyt b. Kerbau rawa asal Sunlatera Utara lnempakan haplotipe yang berbeda dari kerbau rawa asal Banten dan NTB dalam proses domestikasi lebih lanjut, namun tetap berasal dari satu nenek moyang kerbau yang sama yakni dari Cina. Pola persebaran kerbau mengikuti pola ekspansi pertanian manusia.
SARAN
Ruas Cyt b dan D-loop yang digunakan dalam menganalisis filogeni hendaknya bemkuran lebih panjang mendekati ukuran keselumhan. Basa ketiga dari tiap kodon sebaiknya tidak diikutsertakan dalam analisis filogeni.
DAFTAR I'USTAKA
I
Bellwood P. 2001. Early agriculturalist population diasporas? farming,
- . -
languages, and genes. Annu. Rev. ~ n & i - o i o l 3 0 : 181-207.
Chen YS, Li XH. 1989. New evidence of the oiigin and doinestication of the Chinese swalno buffalo (Bubal~rs bubalis). Buffalo / 1: 51-5.
Coccone A, Anlato G, Gratry OC, Bel~ler J, Powell JR. 1999. A nlolecular phylogeny of four endangered madagascar tortoises based on mtDNA sequences. Mol P/ij,/ Ei,oI 12: 1-9. Cockrill WR. 1981 The water buffalo: a
review. British Vet J 137: 8-16.
Fahimuddin M. 1975. Dor71estic Water B t ~ ~ ~ l o . New Delhi: Oxrord & IBH. Gray RD, Jordan EM. 2000. Languages trees
support the express train sequence of Austronesia~~ expansion. Nnf~rre 405: 1052-1055.
Gemmell NJ el ul. 1994. Cloning and characterization of the Platypus mitochondrial genome. J Mol Evol 39: 200-205.
I-Iilmi M. 1991. Karyotypes of water buffalo crosses (swamp x river). Di dalam: Tulloh NH, editor. Buffr,lo and Goal in Asir~: Genetic Dii~ersity arid its Applicalion. Proceedings of The Atrstruliar~ Centre for Internationul Agriccrltural Resem.ch; Kuala Lumpur, 10-14 Feb 1991. Canberra: Goa~ma. 1991. h l n ~ 53-56.
Hivsoi~ JE, Brown WM. 1986. A comparison of the snlall ribosomal RNA genes f i o n ~ the mitochondrial DNA of the great apes and humans: sequences, structure, evolution, and phytogenetic imnplications. MolBiolE1~ol3:I-15.
Hoelzel AR, Lopez JV, Dover GA, Brien JO. 1994. Rapid evolution of a heteroplasmic repetitive sequences in the mitochondrial DNA control region of carnivores. J Mol Eva1 39: 191-199.
Kumar S, Nagarajan M, Sandhu JS, Ku~nar N. Behl V, Nishanth G. 2007. Mitochon DNA analyses of Indian water bu support a distinct genetic origin of river and swamp buffalo. Anirn Gene1 38: 227- 232.
Kuznetsova MV, Khoiodova MV, Danilkin
Lau CH, Drinkwater RD, Yusoff K, Tan SG, Hetzel DJS, Barker JSF. 1998. Genetic diversity of Asian water buffalo (Bubalus bubalis): mitochondrial DNA D-loop and cytochrome b sequence variation. ilnin~ Genet 29(4): 253-264.
Matthee CA, Davis SK. 2001. Molecular insights into the evolution of the family Bovidae: a nuclear DNA perspective. Mol
BiolEvol18(7):1220-1230.
Nei M, Kumar S. 2000. Molecldur Evolzltiorl artd Phylogenetics. New York: Oxford Univ Pr.
Parnla P, Era-Pujada M, Feligini M, Greppi G, Enne G. 2004. Water buffalo (Bubalrrs bubalis): complete nucleotide mitochondrial genonie sequence. DNA sequence 15: 369-373.
Tanaka T, Matsui M, Takenaka 0. 1994. Estimation of phylogenetic relationships among japanese brown frogs from mitochondrial cytoclvon~e b gene (Anlphibia: Anura). Zoo1 Sci 1 l(5): 753- 757.
Taswinangsih W. 2009. Analisis keragaman genetic kerbau lokal (BL~~NILIS b~ibalis) berdasarkan haplotipe DNA mitokondria
[skripsi]. Bogor: Fakultas Peternakan, Institut Pertanian Bogor.
Tegelstrom H. 1986. Mitochondria1 DNA in natural populations: an improved routine for the screening of genetic variation based on sensilive silver staining. Electrophoresis 7:226-229.
Lampiran 1. Spesies pembanding dalam analisis filogeni yang diperoleh dari GerzeBank
No Nama spesies Nama lokal Lokasi No. Akses
1 Bttbalus bubalis kerbau rawa Cina AY702618
2 B~tbalus btrbalis kerbau sungai (Carabao) Filipina FJ873683
3 Bzrbalus bubalis kerbau rawa Ittdia AF547270
4 Bubaltrs bubalis kerbau rawa India-2 FM179473
5 Bubaltts bubalis Kerbau rawa Itali AY488491
6 Bubalus bubalis kerbau rawa Irlandia DQ995713
7 Bubalcrs bubalis Kerbau rawa Jernian AF197199
8 Bubalus bztbalis kerbau rawa Mesir EU268909
Bos grunniens Bisorz bison Bos faurus Bos inn'ictrs Anznlofragus lwvia Ovis aries
Capra hircits Mzrntincrts crinifr.ons
Yak Bison Sapi angus Sapi indicus Dotnba barbary Dotnba gamt Kanibing kacang Kij ang
Cina USA USA Jennan Itali Jerman Itali Cina
17 Rangifer furandus Rusa kutub Jepang A8245426
18 Cervus mic color swinhoei Rusa sambar Taiwan EF035448
19 Cervus rzipporr fniovanrs Rusa sika Taiwan EF058308
Lampiran 2 Hasil penjajaran DNA mas Cyt b dan D-loop genom mitokondria kerbau rawa (Bubalus bubalis). Keterangan: nolnor posisi nukleotida dibaca secara vertil~al di tiga baris pertama
Ruas Cyt b
1 1111111112 2222222223 3333333334 4444444445 1234567890 1234567890 1234567890 1234567890 1234567890 Sumut-(SU181) GAAGTATGAT GTTCCGGCCA TTCAGCCAAT GCCTATTCTG AATCCTAGTA Banten (8731
-
. . .
T.... . . Banten-(B74). . .
T.... . .
NTB - (NTB76) T...
5555555556 6666666667 7777777778 8888888889 9999999990 1234567890 1234567890 1234567890 1234567890 1234567890 Sumut_(SU181) TAGTGCTART ACCAACGGCC AGCGTAGTCG APJATPATCT C T T W T G A
. . .
Banten-(B73) A..A..
. . .
. . .
Banten-(B74) A..A...
. . .
. . .
NTB (NTB76) - A..A.
. . .
123 Sumut-(SU181) AGA Banten-(B73)
. . .
Banten-(B74). . .
NTB-(NTB76) . . .456 Smut-(SU181) CAA Banten-(B73)
...
Banten (B74)...
NTB-
( N T B ~ ~ )...
tRNA Thr
-
. . .
Banten-(B73)
. . .
Banten-(B74)
tRNA Pro
7890123456 78901 Sumut (SU181) TTTAAATACT CCCTG
! l l - : - - l l ? , ! 2 i i 2 > , . ' . . 2 :::!.:.>>:. ; 1:.:.%.;.14::.2
. . L , 4 ; b ' d i ! -. : i j ; 5 ( : 7 6 ? ? 12j.l:~5-8,C 1 : : 4 : ( 7 5 i . 1:.?35616;1
Sunur
-
( S U l i 1 r:r,C:.C'!Al l'fi. .V.'hrNl C?:C .>CNU%GC;LU. A'.;!.!;Zl-T'l '. !:;!.4?3TC&AABanten-(B73) .. T
. . .
G... . .
Banten_(B74)
. .
T. . .
G..... . .
NTB-(NTB76) . . T....
. . .
G..... . .
~ ~ ~ ~ ~ ~ ~
1 2 ? L ! , C ' b S l 123.1-1C1190 1 2 3 < 5 6 - 8 3 0 I T ? : i i r - F ' L : : ' t , j G - B ? C
Ecx-r - 1'; 16 1. 'I '!'C.ACTIL:JA (: I"I'CCTi- :<LA CI'T?-:,CII 3 I C: A: l'i'. ;I /.:. :' C'I :IG u u - - I q ' .,-%C
. . .
. . .
Banten__(B73) A...
Banten-(B74i
. . .
A,... . .
NTB-(NTB76). . .
A,... . .
-
. . .
Banten (B73)
. . .
A...C.. A..A.-
FILOGENI KERBAU RAWA
(Bubulus bubalis)
BERDASARKAN GENOM MITOKONDRIA
SYLVIA NOVITA PALUPI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAIIUAN ALAM
INSTITUT PERTANIAN
BOGOR
PENDAHULUAN
L a t a r Belakang
Kerbau (Bubalus bubalis) me~upakan anggota Famili Bovidae, Subfaniili Bovinae, genus Bubalus (Williamson & Payne 1993). Bubalus memiliki morfologi berbeda dari Bovinae lailmya yaitu Bos, Bison, Pseudoryx, dan Syncerus. Tanduknya cenderung bersudut bahkan membentuk sabit. Selain itu, telinganya berukuran kecil dan tulang tengkoraknya panjang dan senipit (Willianlson & Payne 1993).
Ulrbalus b ~ ~ b u l i s dikenal sebagai kerbau Indian yang diduga hasil dotnestikasi kerbau liar Asia, Bubalis anlee (Lau el a1 1998). Kerbau hasil domestikasi bisa dibedakan menjildi kehau rawa (swanzp bufiff dan kcrbau sungai (rh~er b~rffnlo). Kerbau rawa ~ilenliliki kulit benvarna hitam abu-abu dengan tanda putih abu-abu pada lipatan bagiati bawah leliernya dan tanduk yang tunlbuh ~nelengkung ke belakang seperti sabit. Tubuh kerbau sungai berukuran lebih besar dibandingkan kerbau rawa, kulit dominan berwarna hitam dengan tanda putih di bagian dahi dan ujung ekor, dan tanduk berukuran relatif pendek, melengkung ke depan, ke bawah atau ke belakang. Selain itu, jumlah krotnosom kerbau rawa 2n=4S sedangkan kerbau sungai 2n=50 (FIilmi 1991). Pola penyebaran keduanyapun berbeda. Kerbau rawa tersebar di daratan India, Timur Tengah dan Eropa bagian barat, sedangkan kerbau sungai tersebar di Cina, Bangladesh, negara- negara di Asia Tenggara dan Barat daya India. Filogeni adalah sejarah mengenai garis evolusi suatu kelo~npok organisme. Analisis filogeni pada satu spesies yang nienyebar nlengikuti pola geografi dikenal dengan istilah filogeografi. Salah satu penanda molekular yang banyak digunakan dalam analisis filogeui maupun filogeografi adalah genoni mitokondria (mtDNA) (Coccone 1999). Perbedaan penggunaan mtDNA baik untuk analisis filogeni maupun filogeografi adalah pada karakter laju mutasi autar bagian-bagian mtDNA. Bagian mtDNA yang paling stabil dan banyak digunakan untuk menganalisis filogeni pada tingkat interspesies adalah tRNA dan rRNA. Bagian genom yang menyandikan protein yang paling sering digunakan untuk menganalisis filogeni anggota taksa dibawah ordo atau famili adalah Cyt b. Bagian D-loop dari genom adalah yang paling cepat laju mutasinya sehingga sangat populer digunakan untuk menganalisis
keragaman intraspesies atau filogeografi (Farajallah A 8 Juni 2009, komunikasi pribadi). Ruas genonl mitokondria yang banyak digunakan sebagai penanda molekuler untuk menganalisis filogeni pada B~rbalus yakni Cyt b (Lau et ul. 1993, Hassanin & Douzety 1999) dan D-loop (Lau et al. 1993).
Parma et nl. (2004) menjabarkan posisi filogenetik Bubalus dalam kelima matnalia lainnya, yaitu sapi (Bos inclicus), kambing, domba, rusa (Mcuitiacus nzwztjak) dan Ilama (Lanta pacos) berdasarkan 13 gen penyandi protein mtDNA. Bubalus berkerabat dekat dengau sapi sebagai ang, aota run~inausia besar. ICeduanyn niengelo~i~pok dengan kambing dan domba sebagai anggota dari mminansia kecil. Elubungan kekerabatan Bubalus jauh dengan rusa dati Ilama.
Tujuan
Penelitan ini be]-tujuan mempelajari posisi filogeni B. b~fbalis dalanl Fnniili Bovidae dari berbagai daerah di Indonesia dan menipelajari pola penyebal-annya berdasarkan genom mitokondria.
BAHAN DAN METODE
Bahan
Sampel darah kerbau rawa yaug digunakan dalam penelitian ini adalah sampel darah yang diawetkan dalam alkohol 70% koleksi Bagian Fungsi Hnyati dan Perilaku Hewan Departemen Biolog~. Sebailyak delapan sampel dipilih dari keenlpat lokasi yang berbeda, yaitu Surnatera Utara (SU181 dan SU182), Nusa Tenggara Barat (NTB)
(NTB76 dan NTB77), Banten (B73 dan I374) dan Jawa Tengah (JT38 dan JT39). Sa~npel asal Suniatera Utara dipilih berdasarkan infonnasi yang diberikan ole11 Ta~winangsih (2009) sebagai haplotipe yang berbeda berdasarkan metode PCR-RFLP.
Waktu dan Ternpat P e ~ ~ c l i t i a ~ l
Penelitian i ~ i i dilaksanakan pada bulan Feb~uari sampai dengan Mei 2009 bertempat di Laboratoriu~u Bagian Fungsi Hayati dan Perilaku Hewan Departemen Biologi, Institut Pertanian Bogor, Bogor.
Metode
Ekstraksi dan Isolasi DNA
PENDAHULUAN
L a t a r Belakang
Kerbau (Bubalus bubalis) me~upakan anggota Famili Bovidae, Subfaniili Bovinae, genus Bubalus (Williamson & Payne 1993). Bubalus memiliki morfologi berbeda dari Bovinae lailmya yaitu Bos, Bison, Pseudoryx, dan Syncerus. Tanduknya cenderung bersudut bahkan membentuk sabit. Selain itu, telinganya berukuran kecil dan tulang tengkoraknya panjang dan senipit (Willianlson & Payne 1993).
Ulrbalus b ~ ~ b u l i s dikenal sebagai kerbau Indian yang diduga hasil dotnestikasi kerbau liar Asia, Bubalis anlee (Lau el a1 1998). Kerbau hasil domestikasi bisa dibedakan menjildi kehau rawa (swanzp bufiff dan kcrbau sungai (rh~er b~rffnlo). Kerbau rawa ~ilenliliki kulit benvarna hitam abu-abu dengan tanda putih abu-abu pada lipatan bagiati bawah leliernya dan tanduk yang tunlbuh ~nelengkung ke belakang seperti sabit. Tubuh kerbau sungai berukuran lebih besar dibandingkan kerbau rawa, kulit dominan berwarna hitam dengan tanda putih di bagian dahi dan ujung ekor, dan tanduk berukuran relatif pendek, melengkung ke depan, ke bawah atau ke belakang. Selain itu, jumlah krotnosom kerbau rawa 2n=4S sedangkan kerbau sungai 2n=50 (FIilmi 1991). Pola penyebaran keduanyapun berbeda. Kerbau rawa tersebar di daratan India, Timur Tengah dan Eropa bagian barat, sedangkan kerbau sungai tersebar di Cina, Bangladesh, negara- negara di Asia Tenggara dan Barat daya India. Filogeni adalah sejarah mengenai garis evolusi suatu kelo~npok organisme. Analisis filogeni pada satu spesies yang nienyebar nlengikuti pola geografi dikenal dengan istilah filogeografi. Salah satu penanda molekular yang banyak digunakan dalam analisis filogeui maupun filogeografi adalah genoni mitokondria (mtDNA) (Coccone 1999). Perbedaan penggunaan mtDNA baik untuk analisis filogeni maupun filogeografi adalah pada karakter laju mutasi autar bagian-bagian mtDNA. Bagian mtDNA yang paling stabil dan banyak digunakan untuk menganalisis filogeni pada tingkat interspesies adalah tRNA dan rRNA. Bagian genom yang menyandikan protein yang paling sering digunakan untuk menganalisis filogeni anggota taksa dibawah ordo atau famili adalah Cyt b. Bagian D-loop dari genom adalah yang paling cepat laju mutasinya sehingga sangat populer digunakan untuk menganalisis
keragaman intraspesies atau filogeografi (Farajallah A 8 Juni 2009, komunikasi pribadi). Ruas genonl mitokondria yang banyak digunakan sebagai penanda molekuler untuk menganalisis filogeni pada B~rbalus yakni Cyt b (Lau et ul. 1993, Hassanin & Douzety 1999) dan D-loop (Lau et al. 1993).
Parma et nl. (2004) menjabarkan posisi filogenetik Bubalus dalam kelima matnalia lainnya, yaitu sapi (Bos inclicus), kambing, domba, rusa (Mcuitiacus nzwztjak) dan Ilama (Lanta pacos) berdasarkan 13 gen penyandi protein mtDNA. Bubalus berkerabat dekat dengau sapi sebagai ang, aota run~inausia besar. ICeduanyn niengelo~i~pok dengan kambing dan domba sebagai anggota dari mminansia kecil. Elubungan kekerabatan Bubalus jauh dengan rusa dati Ilama.
Tujuan
Penelitan ini be]-tujuan mempelajari posisi filogeni B. b~fbalis dalanl Fnniili Bovidae dari berbagai daerah di Indonesia dan menipelajari pola penyebal-annya berdasarkan genom mitokondria.
BAHAN DAN METODE
Bahan
Sampel darah kerbau rawa yaug digunakan dalam penelitian ini adalah sampel darah yang diawetkan dalam alkohol 70% koleksi Bagian Fungsi Hnyati dan Perilaku Hewan Departemen Biolog~. Sebailyak delapan sampel dipilih dari keenlpat lokasi yang berbeda, yaitu Surnatera Utara (SU181 dan SU182), Nusa Tenggara Barat (NTB)
(NTB76 dan NTB77), Banten (B73 dan I374) dan Jawa Tengah (JT38 dan JT39). Sa~npel asal Suniatera Utara dipilih berdasarkan infonnasi yang diberikan ole11 Ta~winangsih (2009) sebagai haplotipe yang berbeda berdasarkan metode PCR-RFLP.
Waktu dan Ternpat P e ~ ~ c l i t i a ~ l
Penelitian i ~ i i dilaksanakan pada bulan Feb~uari sampai dengan Mei 2009 bertempat di Laboratoriu~u Bagian Fungsi Hayati dan Perilaku Hewan Departemen Biologi, Institut Pertanian Bogor, Bogor.
Metode
Ekstraksi dan Isolasi DNA
yang sudah bersih dari alkohol kemudian disuspensikan dalam bufer STE (NaCI lM, Tris-HCI lOmM, EDTA 0,) mM, pH 8,O) sampai volume 350 p1. Sel-sel darah dilisis dengan SDS 1% dan proteinase K 0,125 mglml, kemudian dimkubasi pada suhu 55'C selama 1 jam sambil dikocok pelan. Metode ekshaksi DNA selanjutnya mengikuti petunjuk Genolnic DNA mini kit (Geneaid) for blood.
Amplifikasi dan Visualisasi Fragmen DNA Amplifikasi genom mitokondria menggunakan pasangan primer koleksi Dr.Ir. Achmad Farajallah, M. Si, yaitu AF22 (5'- GCGTACGCAATCTTACGAT CA-3') yang menempel pada basa ke-15357 dan AF23 (5'-
ATGCAGTTAAGTCCAGCTAC-3') yang
menempel pada basa ke-143. Pasangan primer ini mengapit mas D-loop mulai dari bagian akhir gen Cyt b sampai ke bagian akhir D- loop dengan panjang 1145 pb. Knmposisi reaksi PCR dengan volume 25 p1 terdiri atas sampel DNA 2 PI, Taq Polymerase (RBC) 0.75 unit beserta bufernya, MgC12 0.2 mM,
dNTPs mix 0.2 mM dan masing-masing primer 1
nM.
Reaksi PCR dilakukan menggunakan mesin Thern~ocycler TaKaRa MP4 dengan kondisi sebagai berikut: tahap denaturasi awal pada suhu 94 OC selama lima menit;
tahap kedua yang terdiri dari 30 siklus yang masing-masing siklus terdiri dari denaturasi pada suhu 94 OC selama satu menit,
penempelan primer pada suhu 59 OC selama dua menit dan pemanjangan pada suhu 72 OC
selama dua menit; tahap terakhir yaitu pemanjangan akhir pada suhu 72 OC selama sepuluh menit. Kualitas produk PCR diuji menggunakan PAGE 6% dalam bufer 1X TBE (10 mM Tris-HCI, 1 M asam borat, 0.1 mM EDTA) yang dijalankan pada kondisi 180 mV selama 40 menit. Selanjutnya DNA diwarnai dengan pewamaan sensitif p e r k (Tegelshom 1986).
Perunutan Prnduk PCR
Produk PCR bempa pita tunggal berukuran sesuai desain primer dimurnikan dengan QIAQuick PCR Purification Kit
(QIAGEN). Produk PCR yang sudah diiumikan kemudian dijadikan sampel dalam reaksi PCR untuk perunutan nukleotida dengan metode Dye Dideoxi Terminator
menggunakan primer AF 22 (PT. CHAROEN POKPHAND INDONESIA). Perunutan nukleotida menggunakan mesin ABI Prism 3700-Avant Genetic Analyzer.
Anaiisis Filogeni
Runutan nukleotida yang dipcroleh diedit secara manual lnenggunaban program Bioedit versi 7.0.9.0 berdasarkan pada kromatogram hasil perunutan. Runutan nukleotida yang telah diedit disejajarkau dengan urutan nukleotida beberapa spesies lain yang bomolog dari GeneBank (Lampiran 1). Proses pensejajaran dilakukan menggunakan program Clustal W1.8 dalam MEGA 4.0 yang kemudian diedit secara manual. Perhitungan komposisi nukleotida, tipe subtitusi, jarak genetik dan konstruksi pohon filogeni dilakukan menggunaka~i program MEGA 4.0 dengan metode Ki~ntrra-2-parameter.
Rekonstruksi pohon filogeni berdasarkan mas Cyt b, sedangkan rekonstmksi pohon filogeogmfi bet'dasarltan mas D-loop. Rekonshuksi keduanya menggunakan metode
Neighboor Joining (NJ) dengan bootshap lOOOX (Nei & Kumar 2000).
HASIL
Amplifikasi &an Visualisasi Fragmen DNA Dari kedelapdn sampel yang digunakan, sebanyak empat san~pel yang berhasil diamplifikasi, yaitu sampel kerbau rawa asal Sumatera Utara (SUI&l), NTB (NTB76), dan Banten (B73 dan B74). Amplifikasi menggunakan pasangan primer AF22 dan AF23 menghasilkan pita tunggal bem6mn sekitar 1145 pb (Gambar 1). Sampel asal Sumatera Utara (SU182), NTB (NTB77) dan Jawa Tengah (JT38 dan JT39) tidak berhasil diamplifikasi sehingga sampel ini tidak dilakukan perunutan nukleotida.
M I 2 3 4
yang sudah bersih dari alkohol kemudian disuspensikan dalam bufer STE (NaCI lM, Tris-HCI lOmM, EDTA 0,) mM, pH 8,O) sampai volume 350 p1. Sel-sel darah dilisis dengan SDS 1% dan proteinase K 0,125 mglml, kemudian dimkubasi pada suhu 55'C selama 1 jam sambil dikocok pelan. Metode ekshaksi DNA selanjutnya mengikuti petunjuk Genolnic DNA mini kit (Geneaid) for blood.
Amplifikasi dan Visualisasi Fragmen DNA Amplifikasi genom mitokondria menggunakan pasangan primer koleksi Dr.Ir. Achmad Farajallah, M. Si, yaitu AF22 (5'- GCGTACGCAATCTTACGAT CA-3') yang menempel pada basa ke-15357 dan AF23 (5'-
ATGCAGTTAAGTCCAGCTAC-3') yang
menempel pada basa ke-143. Pasangan primer ini mengapit mas D-loop mulai dari bagian akhir gen Cyt b sampai ke bagian akhir D- loop dengan panjang 1145 pb. Knmposisi reaksi PCR dengan volume 25 p1 terdiri atas sampel DNA 2 PI, Taq Polymerase (RBC) 0.75 unit beserta bufernya, MgC12 0.2 mM,
dNTPs mix 0.2 mM dan masing-masing primer 1
nM.
Reaksi PCR dilakukan menggunakan mesin Thern~ocycler TaKaRa MP4 dengan kondisi sebagai berikut: tahap denaturasi awal pada suhu 94 OC selama lima menit;
tahap kedua yang terdiri dari 30 siklus yang masing-masing siklus terdiri dari denaturasi pada suhu 94 OC selama satu menit,
penempelan primer pada suhu 59 OC selama dua menit dan pemanjangan pada suhu 72 OC
selama dua menit; tahap terakhir yaitu pemanjangan akhir pada suhu 72 OC selama sepuluh menit. Kualitas produk PCR diuji menggunakan PAGE 6% dalam bufer 1X TBE (10 mM Tris-HCI, 1 M asam borat, 0.1 mM EDTA) yang dijalankan pada kondisi 180 mV selama 40 menit. Selanjutnya DNA diwarnai dengan pewamaan sensitif p e r k (Tegelshom 1986).
Perunutan Prnduk PCR
Produk PCR bempa pita tunggal berukuran sesuai desain primer dimurnikan dengan QIAQuick PCR Purification Kit
(QIAGEN). Produk PCR yang sudah diiumikan kemudian dijadikan sampel dalam reaksi PCR untuk perunutan nukleotida dengan metode Dye Dideoxi Terminator
menggunakan primer AF 22 (PT. CHAROEN POKPHAND INDONESIA). Perunutan nukleotida menggunakan mesin ABI Prism 3700-Avant Genetic Analyzer.
Anaiisis Filogeni
Runutan nukleotida yang dipcroleh diedit secara manual lnenggunaban program Bioedit versi 7.0.9.0 berdasarkan pada kromatogram hasil perunutan. Runutan nukleotida yang telah diedit disejajarkau dengan urutan nukleotida beberapa spesies lain yang bomolog dari GeneBank (Lampiran 1). Proses pensejajaran dilakukan menggunakan program Clustal W1.8 dalam MEGA 4.0 yang kemudian diedit secara manual. Perhitungan komposisi nukleotida, tipe subtitusi, jarak genetik dan konstruksi pohon filogeni dilakukan menggunaka~i program MEGA 4.0 dengan metode Ki~ntrra-2-parameter.
Rekonstruksi pohon filogeni berdasarkan mas Cyt b, sedangkan rekonstmksi pohon filogeogmfi bet'dasarltan mas D-loop. Rekonshuksi keduanya menggunakan metode
Neighboor Joining (NJ) dengan bootshap lOOOX (Nei & Kumar 2000).
HASIL
Amplifikasi &an Visualisasi Fragmen DNA Dari kedelapdn sampel yang digunakan, sebanyak empat san~pel yang berhasil diamplifikasi, yaitu sampel kerbau rawa asal Sumatera Utara (SUI&l), NTB (NTB76), dan Banten (B73 dan B74). Amplifikasi menggunakan pasangan primer AF22 dan AF23 menghasilkan pita tunggal bem6mn sekitar 1145 pb (Gambar 1). Sampel asal Sumatera Utara (SU182), NTB (NTB77) dan Jawa Tengah (JT38 dan JT39) tidak berhasil diamplifikasi sehingga sampel ini tidak dilakukan perunutan nukleotida.
M I 2 3 4
Perunutan Produk PCR
Perunutan DNA dilakukan dari arah primer
AF
22. Panjang DNA hasil perunutan setelah diedit bervariasi yakni 572 nt untuk sampel SUl 81, 563 nt untuk sampel NTB76, 525 nt untuk sampel B73 dan 527 nt unhlk sampel B74.Analisis Filogeni
Proses pensejajaran menghasilkan runutan sepanjang 516-518 nt dengan rata-rata komposisi nukleotida A=35.6%; T=23.9%; G=13.5%; dan C=27%. Presentase A+T (59.5%) lebih besar daripada C+G (40.5%). Berdasarkan hasil pensejajaran dengan data dari GeneBnnk (Lampiran 1) dapat diketahui bahwa hasil perunutatl keempat san~pel tersebut n~eliputi enlpat domain, yaitu bagian akhir Cyt b (203 nt), tRNA Tlx (70 nt), tRNA Pro (65 nt) dan bagian awal D-loop (178-180 111) (Tabel 1). Panjang gel1 Cyt b dan tRNA stabil antar satnpel sedangkan paiijang D-loop beragam karena terdapat insersi pada basa ke- 159 sa~npel asal NTB (NTB76) atau delesi pada basa ke-67 sampel Sumatera Utara (SU18 1) (lainpiran 2).
Ruas yang digunakan untuk analisis filogeui
B. bllbnlis adalah gen Cyt b. Hasil peusejajaran dengan data dari GnleBank
menunjukkan 172 nt yang identik dan 31 nt
yang berbeda. Basa yang berbeda disebabkan ole11 substitusi basa, baik transisi maupun transversi (Tabel 2). Contoh transisi pada luas Cyt b menyebabkan pe~ubahan asatn amino Met menjadi Val pada basa ke-174 dan Ile menjadi Val pada basa ke-177, sedangkan transisi pada basa ke-44 tidak merubah asam amino dimungkinkan karena terjadi pada basa ketiga dari kodon.
Topologi pohon filogeni hasil NJ dua basa pertama pada tiap kodon menunjukkan Hasil rekonstritksi berdasarkan dua basa pertama dari tiap kodon Cyt b tnetigelolnpokkan Subfamili Capriliae dan Subfamili bovinae dalam salu percabangan melnbentuk kelounpok Fanlili Bovidae. Fa~nili Cervidae berada di luar percabangan liienlbentuk sister grorrp dengan Falnili Bovidae (Ganlbar 2A). Lain halnya dengan hasil rekonstruksi herdasarkan tiga basa dal-i tiap kodon yang ~uembcntuk politotiii antara Subfatuili Bovinae, Subfamili Caprinae dan Fan~ili Cervidae (Gamnbar 28).
Secara filogeografi, sample kerbau rawa Sulilatera Utara te~pisah detigan ketiga sampel kerbau rawa lainnya dan n~engelonlpok dengan kerbau dari India, Jern~an, Irlandia, Itali dan Mesir ((ianlbar 3). Sa~npel kerbau rawa asal Banten dan NTB ~nengelompok dengan sampel kerbau dari Cina.
Tabel 1 Panjang mas hasil pensejajaran (nt)
Satnpel keibvu Cyt-b tRNA Thr IRNA Pro D-loop Pnnjang scluruh nl:s
Surnateta Utv~a (SUISl) 203 70 65 178 516
NTB (NTB76) 203 70 65 I SO 518
Bnnte~l (873) 203 70 65 179 517
Bullle11 (874) 203 70 65 179 517
Tabel 2 Hasil pensejajaran nukleotida DNA yang berbeda
Cyt-b D-loop
I I 1 1 1 1 I I I I 1
Sampel kerbau 4 7 7 1 1 1 2 4 4 5 6 6 6
4 4 7 3 6 4 8 3 6 9 9 1 4 8
Sumaten Utan (SU18 1) C G G
ctfb
T : ~ G G C G T CNTB (NTB76) T A A T;C,~A c~$,:A A T A
.r
TBallten (B73) T A A T ' G A C , C A A T A T T
Banten (B74) T A A
TMA
CiktA A , ~ A C TKerbau rawa India
K. NTB (NTB76)
Kerbau rawa IndiaDomba Gantt
Karnbing Kacang
0.02 0.00 0.00 0.05
Gambar 2 Hasil rekonstruksi pohon filogeni berdasarkan dua basa pertama (A) dan ketiga basa (B) pada tiap kodon dari gen Cyt b menggunakan metode NJ dengan bootstrap 1000X. Nomor menunjukkan nilai bootstrap. (
A:
sub famili Bovinae;0
: sub famili Caprinae;'@ : famili Cewidae). Garis merah merupakan benhlkan politomi.
\
Jerman
Mesir
Sumatera Utara (SUISI)
Banten (B73)
Banten (B74)
Kerbau
rawa
/J
Filipina
Kerbau sungai
-
0.1 [image:29.595.82.511.48.794.2]I
Tabel 3 Nilai jarak genetik (bawah diagonal) dan kesamaan genetik (atas diagonal) antar B. bubalis dari beberapa lokasi berdasarkan metode Ki1nu1-u-2-para1neier
I 2 3 4 5 6 7 8 9 10 I 1 12
I 0.907 0.902 0.082 0.168 0.941 0.994 0.989 1.000 0.909 0.909 0.909
2 0.093 0.994 0.012 0.111 0.919 0.976 0.907 0.975 1.000 1.000 1.000
3 0.098 0.006 0.063 0.151 0.908 0.909 0.903 0.902 0.994 0.994 0.994
4 0.918 0.988 0.937 0.888 -0.084 0.082 0.063 0.082 0.040 0.040 0.040
5 0.832 0.889 0.849 0.112 0.080 0.168 0.151 0.168 0.132 0.132 0.132
6 0.059 0.081 0.092 1.084 0.920 0.947 0.941 0.941 0.814 0.814 0.814
7 0.006 0.087 0.091 0.918 0.832 0.053 0.994 0.994 0.915 0.915 0.915
8 0.011 0.093 0.097 0.937 0.849 0.059 0.006 0.989 0.909 0.909 0.909
9 0,000 0.093 0.098 0.918 0.832 0.059 0.006 0.011 0.909 0.909 0.909
10 0.091 0.000 0.006 0.960 0.868 0.086 0.085 0.091 0.091 1.000 1.000
11 0.091 0.000 0.006 0.960 0.868 0.086 0.085 0.091 0.091 0.0011 1.000
12 0.091 0.000 0.006 0.960 0.868 0.086 0.085 0.091 0.091 0.000 0,000
Keterangan : (1) Cina, (2) Irlandia, (3) Mesir, (4) Filipina, (5) Itidia, (6) Sumatera Utara, (7) Banten (73), (8) Banten (74), (9) NTB, (10) India, (1 1) Italia, (12) Jemian.
G=13.5%; dan C=27%. Presentase A t T (59.5%) lebili besar daripada C+G (40.5%). Berdasarkan hasil pensejajaran deilgan data dari GeneBunk (Lampiran 1) dapnt diketahui bahwa hasil pemnutan keempat sanipel tersebut nieliputi empat domain, yaitu bagian akhir Cyt b (203 nt), tRNA Thr (70 tit), tRNA Pro (65 nt) dan bagian awal D-loop (178-180 nt) (Tabel 1). Panjang gen Cyt b dan tRNA stabil antar sampel sedangkan paiijang D-loop beragani karena terdapat insersi pada basa ke- 159 satlipel asal NTB (NTB76) atau delesi pada basa ke-67 sa~npel Sumatera Utara (SU181) (lainpiran 2).
Ruas yang digunakan untuk analisis filogeni
B. b~tbalis adalah gen Cyt b. Hasil pensejajaran dengan data dari GeneBunk
nienunjukkan 172 nt yang identik dan 31 nt yang berbeda. Basa yang berbeda disebabkan oleh substitusi basa, baik tra~isisi niaupun transversi (Tabel 2). Contoh transisi pada mas Cyt b inenyebabkan pe~ubalian asam ainino Met menjadi Val pada basa ke-174 dan Ile menjadi Val pada basa ke-177, sedangkan transisi pada basa ke-44 tidak inerubah asam amino dimungkinkan karena terjadi pada basa ketiga dari kodon.
Topologi pohon filogeni hasil NJ dua basa pertama pada tiap kodon menunjukkan Hasil rekonstruksi berdasarkan dua basa pertama dari tiap kodon Cyt b mengelompokkan Subfamili Caprinae dan Subfamili bovinae dalam satu percabangan membentuk kelompok Famili Bovidae. Fa~nili Cervidae berada di luar percabaigan membentuk sister group dengan Famili Bovidae (Gambar 2A). Lain halnya dengan hasil rekonstruksi berdasarkan tiga basa dari
tiap kodon yang ~nenibentuk politoiiii autara SubCamili Bovinae, Subfamili Caprinae dan Faulili Cervidae (Gamb;~r 2B).
Secara filogeografi, sampel kerbau rawa Surnatera Utara terpisah dengan ketiga sampel kerbau rawa Iainnya dan rnengelompok dengan kerbau dari India, Jcrman, Irlandia, Itali dan Mesir (Gambar 3). Sanipel kerbau rawa asal Ballten dan NTB ~nengelompok dengan sa~npel kerbau dari Cina.
PEMBAIIASAN
An~plifikasi dan Visualisnsi Fragmen DNA Sampel kerbau rawa asal Sumatera Utara, NTB, dan Ballten berhasil dianiplifikasi menggunakati pasangan primer AF22 dan AF23. Pita tunggal yang dihasilkan sesuai dengall hasil a~nplifikasi desaiu primer bemkuran sekitar 1145 pb (Gambar I).
Keetnpat sanipel lai~mya tidal< berhasil dianiplifikasi (38, 39, 77 dan 182) kemunglcinan karcna subu penempelan primer ( 5 9 ' ~ ) yang tidak sesuai sekalipun sampel yang digu~iakan merupakan spesies yang sama.
Perunutan Produk P C R
Tabel 3 Nilai jarak genetik (bawah diagonal) dan kesamaan genetik (atas diagonal) antar B. bubalis dari beberapa lokasi berdasarkan metode Ki1nu1-u-2-para1neier
I 2 3 4 5 6 7 8 9 10 I 1 12
I 0.907 0.902 0.082 0.168 0.941 0.994 0.989 1.000 0.909 0.909 0.909
2 0.093 0.994 0.012 0.111 0.919 0.976 0.907 0.975 1.000 1.000 1.000
3 0.098 0.006 0.063 0.151 0.908 0.909 0.903 0.902 0.994 0.994 0.994
4 0.918 0.988 0.937 0.888 -0.084 0.082 0.063 0.082 0.040 0.040 0.040
5 0.832 0.889 0.849 0.112 0.080 0.168 0.151 0.168 0.132 0.132 0.132
6 0.059 0.081 0.092 1.084 0.920 0.947 0.941 0.941 0.814 0.814 0.814
7 0.006 0.087 0.091 0.918 0.832 0.053 0.994 0.994 0.915 0.915 0.915
8 0.011 0.093 0.097 0.937 0.849 0.059 0.006 0.989 0.909 0.909 0.909
9 0,000 0.093 0.098 0.918 0.832 0.059 0.006 0.011 0.909 0.909 0.909
10 0.091 0.000 0.006 0.960 0.868 0.086 0.085 0.091 0.091 1.000 1.000
11 0.091 0.000 0.006 0.960 0.868 0.086 0.085 0.091 0.091 0.0011 1.000
12 0.091 0.000 0.006 0.960 0.868 0.086 0.085 0.091 0.091 0.000 0,000
Keterangan : (1) Cina, (2) Irlandia, (3) Mesir, (4) Filipina, (5) Itidia, (6) Sumatera Utara, (7) Banten (73), (8) Banten (74), (9) NTB, (10) India, (1 1) Italia, (12) Jemian.
G=13.5%; dan C=27%. Presentase A t T (59.5%) lebili besar daripada C+G (40.5%). Berdasarkan hasil pensejajaran deilgan data dari GeneBunk (Lampiran 1) dapnt diketahui bahwa hasil pemnutan keempat sanipel tersebut nieliputi empat domain, yaitu bagian akhir Cyt b (203 nt), tRNA Thr (70 tit), tRNA Pro (65 nt) dan bagian awal D-loop (178-180 nt) (Tabel 1). Panjang gen Cyt b dan tRNA stabil antar sampel sedangkan paiijang D-loop beragani karena terdapat insersi pada basa ke- 159 satlipel asal NTB (NTB76) atau delesi pada basa ke-67 sa~npel Sumatera Utara (SU181) (lainpiran 2).
Ruas yang digunakan untuk analisis filogeni
B. b~tbalis adalah gen Cyt b. Hasil pensejajaran dengan data dari GeneBunk
nienunjukkan 172 nt yang identik dan 31 nt yang berbeda. Basa yang berbeda disebabkan oleh substitusi basa, baik tra~isisi niaupun transversi (Tabel 2). Contoh transisi pada mas Cyt b inenyebabkan pe~ubalian asam ainino Met menjadi Val pada basa ke-174 dan Ile menjadi Val pada basa ke-177, sedangkan transisi pada basa ke-44 tidak inerubah asam amino dimungkinkan karena terjadi pada basa ketiga dari kodon.
Topologi pohon filogeni hasil NJ dua basa pertama pada tiap kodon menunjukkan Hasil rekonstruksi berdasarkan dua basa pertama dari tiap kodon Cyt b mengelompokkan Subfamili Caprinae dan Subfamili bovinae dalam satu percabangan membentuk kelompok Famili Bovidae. Fa~nili Cervidae berada di luar percabaigan membentuk sister group dengan Famili Bovidae (Gambar 2A). Lain halnya dengan hasil rekonstruksi berdasarkan tiga basa dari
tiap kodon yang ~nenibentuk politoiiii autara SubCamili Bovinae, Subfamili Caprinae dan Faulili Cervidae (Gamb;~r 2B).
Secara filogeografi, sampel kerbau rawa Surnatera Utara terpisah dengan ketiga sampel kerbau rawa Iainnya dan rnengelompok dengan kerbau dari India, Jcrman, Irlandia, Itali dan Mesir (Gambar 3). Sanipel kerbau rawa asal Ballten dan NTB ~nengelompok dengan sa~npel kerbau dari Cina.
PEMBAIIASAN
An~plifikasi dan Visualisnsi Fragmen DNA Sampel kerbau rawa asal Sumatera Utara, NTB, dan Ballten berhasil dianiplifikasi menggunakati pasangan primer AF22 dan AF23. Pita tunggal yang dihasilkan sesuai dengall hasil a~nplifikasi desaiu primer bemkuran sekitar 1145 pb (Gambar I).
Keetnpat sanipel lai~mya tidal< berhasil dianiplifikasi (38, 39, 77 dan 182) kemunglcinan karcna subu penempelan primer ( 5 9 ' ~ ) yang tidak sesuai sekalipun sampel yang digu~iakan merupakan spesies yang sama.
Perunutan Produk P C R
dimnutkan seperti pada karakterisasi genom mitokondria Platypus (Gemmell et a1 1994).
Analisis Filogeni
Rnas mtDNA harus perunutan mayoritas tersusun atas basa A+T. Banyaknya basa A t T pada D-loop erat kaitannya dengan karakter D-loop yang memiliki banyak situs awal replikasi untuk utas berat (Heavy slrand) mtDNA dan promotor bagi utas berat tnaupun utas ringan (Light strand) sebagai titik awal transkripsi (Hoelzel AR et (11. 1994).
Panjang mas Cyt b stabil pada keempat sample dan memiliki tingkat variabilitas yang rendah. Hal ini terlihat dari sedikitnya situs yang mengala~ni subtitusi (Tabel 2).
Transversi lianya dite~nukan pada luas D-loop dengan rasio transisi terhadap trausversi (tsltv) sebesar 4.5. Hal ini menunjukkan baliwa subtitusi tranversi jarang terjadi dibandingkan subtitusi twlsisi. Hasil ini sejalan dengan yang dikemukakan oleh Hixson & Brown (1986) bahwa pada 1ntDN.4 transisi lebih sering terjadi dibandingkan transversi.
Hasil rekonstruksi pohon filogeni herdasarkan urutan nukleotida Cyt b dilaknkan dengan membandinglcan dua basa pertanla dan ketiga basa pada tiap kodon. Rekonstmksi pollon filogeni menggunakan dua basa pertama pada tiap kodon lebih tepat menjelaskan kedudnkan Famili Cervidae yang merupakan sistei. group dari Famili Bovidae (Gambar 2A) sesuai dengan klasifikasi Williamson & Payne (1993) dan Kuznetsova el a[. (2004). Rekonstruksi pohon filogeni menggunakan ketiga basa pada tiap kodon menghasilkan politotni antara Subfamili Bovinae, Subfa~nili Capriiiae dan Famili Cervidae (Gambar 20). Politotiii mengindikasikan adanya liubungan filogenetik yang tidak pasti (politon~i lunak). Perbedaan hasil analisis antara dua basa pertama dan ketiga basa dari tiap kodon dimungkinkan karena posisi basa ketiga pada tiap kodon Cyt b yang tidak stabil. Basa ketiga pada tiap kodon tidak stabil karena sering mengalami transisi (Tanaka et a/. 1994; Matthee & Davis 2001). Kejadian transisi pada basa ketiga dari tiap kodon tidak berpengamh terhadap penyandian asam amino Transisi pada basa ketiga dari tiap kodon Cyt b (Tabel 2) merupakan mutasi bisn yang tidak berpengaruh terhadap penyandian asam amino. Basa pada Cyt b yang dibandingkan merupakan sebagian dari mas Cyt b yang seharusnya berukuran
2
1100 nt sehingga asumsi bahwa transisi pada basa ketigamelupakan jenis mutasi bisu belum kuat dibuktikan.
Kerbau rawa asal Sumatera Utara lebih berkerabat dekat dengan kerbau dari Irlandia, Jerman, Itali, Mesir dan India dengan rata-rata jarak genetik sebesar 0.0865 atau tingkat kesamaan gennya sebesar 91.35%. Kerbau rawa asal NTB dan Banten memiliki kedekatan dengan kerbau dari Cina dengan tingkat kesamaan gen sebesar 100% dan 91.5% (Tabel 3). Kerbau rawa asal Sumatera Utara mernang merup;lkan haplotipe yang berbeda dari kerbau rawa asal Baiiten dan NTB, namun ketiganya tetap mengelotnpok dala~n Subfamili Bovinae dengan nilai bootstrap sebesar 99%. Hal ini ~nemunculkan dugaan bahwa kerbau rawa asal Sumatera Utara berasal dari nenek ~noyang yalig salna dengan sampel asal Batilen daii NTB.
Pola persebaran kerbau yang luas diduga tnengiknti jalur niigr;isi inanusia karena kerbau merupakan lieu,an ~uminansia besar yang mernbutuhki~n media untuk migrasinya. Pola migrasi ~iianusia yang dijadikan acuali yakni pada masa menetap dan bercocok tanam yang mempakan penanda beralihnya kebudayaan berburu dan mengumpulkan makanan dari zaman batu lama (Paleolitikum) ke zaman batu baru li\ieolitikum). Daerah pertanian awal yang meliputi tiga area, yakni Asia barat daya, Cina dan Mesoamerica (Bellwood 2001) diduga tnerupakan awal tempat domestikasi hewnn, tern~asuk kerbau.
Cockrill (1981) berasumsi bahwa tempat domestikasi kerbau berawal di pusat peradaban India yakni daerah Yangtze, Eupluates and Tigris sekitar 5000 tahun lalu. Berbeda dengan asumsi Chen & Li (1989) yang menyebutkan baliwa Cilia merupakan tempat awal donlestikasi kerbau sejak 6000 tal~nn yang lalu. Kedua lcontadibsi iiii menguatkan dugaan ICu~uar et nl (2007) bahwa kedua tipe kerbau merupakan hasil dotnestikasi dari nenek nloyang yang sama dan domestikasi kednanya dilaknkan secara terpisah.
Sumatera Utara berbeda dengan sampel kerbau rawa asal Banten dan NTB. Meskipun berbeda dalam ha1 proses domestikasi lebih lanjut, kerbau rawa asal Sumatera Utara, Banten, dan NTB tetap berasal dari nenek moyang yang sama di Cina. Jawa (Banten) dan Indonesia Timur (NTB).
Penyebaran hewan domestikasi memiliki kecendemngan menghimdari hariel. geografi. Kerbau dari Cina tidak menyebar ke arah utara Asia karena adanya Gurun Gobi dan tidak adanya ekspat~si dari J a m ke Sumatera karena di Sumatera terdapat pegunu~lga~l Bukit Barisan.
Kerbau berkenlbang di daerah dengan agroekosiste~n yang bemariasi seperti padang penggemhalaa~l dengan iklim kering di NTB, NTT di daerah persawahan irigasi ~naupull non irigasi (Banten); di daerah pegunungan (Sumatera Utara) dan dataran rendah yang berawa-rawa seperti di daerah Kalimantan Selatan. Hal ini menunjukka~~ bahwa daya adaptasi kerbau pada berbagai koi~disi agroekosistem sangat besar. Sebagai akibat pengamh lingkungan telah terjadi semacam evolusi sehingga timbul semacam sub gmp pada kerbau di Indonesia, seperti (1) timbulnya kerbau-kerbau yang berbadan besar dan yang berbadan kecil, (2) perbedaan terhadap daya tahan terhadap panas, dan (3) kegenlaran hidup di dalam air (Fahimuddin 1975).
Hubuugan filogeni berdasarkan gen Cyt b rnenempatkan Bubalus bubalis sebagai anggota Subfamili Bovinae, Fa~nili Bovidae. Analisis filogeni n~enggut~akan d ~ ~ a basa pertama pada tiap kodot~ Cyt b lehi11 stabil dibandingkan ketiga basa pada tiap kodon Cyt b. Kerbau rawa asal Sunlatera Utara lnempakan haplotipe yang berbeda dari kerbau rawa asal Banten dan NTB dalam proses domestikasi lebih lanjut, namun tetap berasal dari satu nenek moyang kerbau yang sama yakni dari Cina. Pola persebaran kerbau mengikuti pola ekspansi pertanian manusia.
SARAN
Ruas Cyt b dan D-loop yang digunakan dalam menganalisis filogeni hendaknya bemkuran lebih panjang mendekati ukuran keselumhan. Basa ketiga dari tiap kodon sebaiknya tidak diikutsertakan dalam analisis filogeni.
DAFTAR I'USTAKA
I
Bellwood P. 2001. Early agriculturalist population diasporas? farming,
- . -
languages, and genes. Annu. Rev. ~ n & i - o i o l 3 0 : 181-207.
Chen YS, Li XH. 1989. New evidence of the oiigin and d