AKTIVITAS INHIBISI ENZIM PENGUBAH ANGIOTENSIN
PADA HIDROLISAT TEMPE KORO PEDANG (
Canavalia
ensiformis
L.) SECARA
IN VITRO
IRMAN FADLY
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Aktivitas Inhibisi Enzim Pengubah Angiotensin pada Hidrolisat Tempe Koro Pedang (Canavalia ensiformis L.) secara In Vitro adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Penelitian ini merupakan bagian dari kegiatan penelitian Teknologi Pengolahan Kacang-kacangan sebagai Sumber Protein untuk Substitusi Impor Kedelai dengan penanggung jawab Dr Ir Endang Yuli Purwani Msi, dkk. Penelitian ini didanai oleh DIPA BB Pascapanen 2013 dengan nomor DOK-INT-RE-3.2/032/2013. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Februari 2014
ABSTRAK
IRMAN FADLY. Aktivitas Inhibisi Enzim Pengubah Angiotensin pada Hidrolisat Tempe Koro Pedang (Canavalia ensiformis L.) secara In Vitro. Dibimbing oleh MARIA BINTANG dan ENDANG YULI PURWANI.
Tempe merupakan salah satu olahan pangan yang memiliki aktivitas inhibisi ACE (Angiotensin Converting Enzyme). Penelitian ini bertujuan mengukur aktivitas inhibisi ACE pada hidrolisat tempe koro pedang secara in vitro. Analisis proksimat menggunakan metode AOAC (2005) dan dihidrolisis dengan gastrointestinal simulation (GIS) digestions. Derajat hidrolisis mengacu pada modifikasi metode Benjakul & Morrissesey. Pengukuran ACE inhibitor, berdasarkan laju pembentukan asam hippurat dari hippuryl-L-histidyl-L-leusine (HHL). Analisis proksimat koro pedang menunjukkan kadar lemak yang lebih rendah dan karbohidrat yang lebih tinggi dari kedelai. Derajat hidrolisis dari tempe koro pedang menunjukkan hasil baik yakni sebesar 10.62% untuk tempe mentah dan 16.24% untuk tempe kukus. Hal ini sebanding dengan konsentrasi protein yang diperoleh antara tempe mentah dan tempe kukus. Hasil tersebut berpengaruh terhadap inhibisi ACE. Nilai IC50
menunjukkan tempe mentah lebih efektif menghambat aktivitas ACE sebesar 0.26 mg/g sampel (bk), sedangkan tempe kukus diperoleh nilai IC50 sebesar 1.11 mg/g
sampel (bk). Hasil IC50 kacang koro ini lebih kecil dibandingkan dengan kedelai dan
kaptropil (kontrol positif). Secara umum, tempe koro pedang mampu menghambat aktivitas ACE dalam menurunkan tekanan darah secara in vitro
Kata kunci: ACE, inhibisi, HHL, in vitro, koro pedang, tempe
ABSTRACT
IRMAN FADLY. Inhibition of angiotensin-converting enzyme activity in the hydrolyzate “Tempe” from Jack Bean (Canavalia ensiformis L.) in In Vitro. Supervised by MARIA BINTANG and ENDANG YULI PURWANI.
Tempe is one of the processed food that has ACE (Angiotensin Converting Enzyme) inhibitory activity. This study aims to measure the activity of ACE inhibition in jack bean in vitro. Proximate analysis used AOAC method (2005) and hydrolyzed by gastrointestinal simulation (GIS) digestions, and then tested the degree of hydrolysis by Benjakul & Morrissesey modification method. Measurement of ACE inhibitors, based on the rate of formation of hippuric acid from substrate hippuryl-L-histidyl-L-leusine (HHL). Proximate analysis showed fat value of jack bean is lower than soybean and carbohydrate value is higher than soybean. Degree of hydrolysis (DH) of tempe from jack bean showed good results which amounted to 10.62% for crude tempe and 16.24% for steamed tempe. Of the IC 50 values obtained, crude tempe 50% more effective at inhibiting the activity of ACE by 0.26 mg/g sample. However, for the steamed tempeh also obtain poor results, because of the results obtained IC50 1:11 mg/g sample.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biokimia
AKTIVITAS INHIBISI ENZIM PENGUBAH ANGIOTENSIN
PADA HIDROLISAT TEMPE KORO PEDANG (
Canavalia
ensiformis
L.) SECARA
IN VITRO
IRMAN FADLY
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul Skripsi : Aktivitas Inhibisi Enzim Pengubah Angiotensin pada Hidrolisat Tempe Koro Pedang (Canavalia ensiformis L.) secara In Vitro Nama : Irman Fadly
NIM : G84090011
Disetujui oleh
Prof Dr drh Maria Bintang, MS Pembimbing I
Dr Endang Yuli Purwani, MSi Pembimbing II
Diketahui oleh
Dr Ir I Made Artika, MAppSc Ketua Departemen Biokimia
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan April 2013 ini ialah diversifikasi pangan, dengan judul Aktivitas Inhibisi Enzim Pengubah Angiotensin pada Hidrolisat Tempe Koro Pedang (Canavalia ensiformis L.) secara In Vitro
Terima kasih penulis ucapkan kepada BB Pasca Panen yang telah mendanai penelitian ini serta Ibu Maria Bintang dan Ibu Endang Yuli Purwani selaku pembimbing yang telah banyak memberi masukan terhadap penelitian ini. Di samping itu, penulis juga megucapkan terima kasih kepada pak Tri, Ibu Ika, Ibu Mely, Pak Yudi, Wiwik, beserta staf dan teknisi lain BB Pasca Panen yang telah membantu dalam pengumpulan data. Ungkapan terima kasih juga penulis sampaikan kepada Bapak Mansyur Sarip dan Ibu Iriani serta seluruh keluarga, atas segala dukungan, doa, dan kasih sayangnya kepada penulis sehingga karya ilmiah ini dapat diselesaikan.
Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
PENDAHULUAN 1
METODE 2
Waktu dan Tempat Penelitian 2
Bahan 2
Alat 3
Prosedur Penelitian 3
HASIL DAN PEMBAHASAN 7
Hasil 7
Pembahasan 11
SIMPULAN DAN SARAN 16
Simpulan 16
Saran 17
DAFTAR PUSTAKA 17
LAMPIRAN 20
DAFTAR TABEL
1 Hasil pengukuran aktivitas inhibisi ACE 10
2 Nilai IC 50 dari masing-masing sampel hidrolisat 11
DAFTAR GAMBAR
1 Visualisasi kacang koro pedang dan tempe koro pedang 7 2 Kadar proksimat sampel kedelai dan koro pedang 8 3 Perbandingan hasil pengukuran derajat hidrolisis 9 4 Kadar protein bobot kering sampel hidrolisat kedelai 9
DAFTAR LAMPIRAN
1 Diagram alir penelitian 20
2 Hasil pengukuran kadar air kedelai dan koro pedang 21 3 Hasil pengukuran kadar abu kedelai dan koro pedang 22 4 Hasil pengukuran kadar protein kedelai dan koro pedang 23 5 Hasil pengukuran kadar lemak kedelai dan koro pedang 24 6 Hasil pengukuran kadar karbohidrat (by difference) kedelai dan koro
pedang 25
7 Hasil pengukuran derajat hidrolisis 26
8 Hasil pengukuran konsentrasi standar BSA 27
PENDAHULUAN
Indonesia terkenal sebagai negara yang memiliki sumber daya hayati yang melimpah, salah satunya tanaman kacang-kacangan. Kacang-kacangan merupakan bahan pangan yang memiliki kandungan protein, mineral, vitamin, dan serat pangan yang tinggi, serta telah lama dikenal sebagai sumber protein pelengkap. Semua kandungan tersebut sangat dibutuhkan oleh tubuh, seperti protein. Protein memiliki manfaat yang penting bagi tubuh, yakni: pembangun struktur, biokatalis, pengatur pH, sebagai salah satu sumber energi, dan sebagai pembawa sifat turunan. Kacang kedelai merupakan salah satu kacang-kacangan yang memiliki protein yang tinggi. Kadar protein kedelai bisa mencapai 35% berdasarkan bobot keringnya, bahkan pada varietas unggul kadar proteinnya mencapai 40% - 43%. Selama ini kedelai digunakan sebagai bahan baku utama untuk pembuatan tempe. Namun, beberapa tahun terakhir produksi kedelai Indonesia semakin merosot, sedangkan permintaannya kian naik dari tahun ke tahun (BPS 2012).
Departemen Pertanian Indonesia pada tahun 2005, melaporkan tingkat konsumsi kedelai nasional per tahun 2004 adalah 4.186.157 ton, sedangkan kedelai yang tersedia adalah 1.311.196 ton, sehingga menyebabkan impor sebesar 2.874.961 ton. Sampai dengan tahun 2011 produksi kedelai nasional masih mengalami penurunan sebesar 55.74 ribu ton (±6.15%) dibandingkan produksi tahun 2010. Sementara perkiraan produksi di tahun 2012 hanya sebesar 779.74 ribu ton biji kering. Konsekuensi dari penurunan produksi adalah terjadinya defisit kedelai yang terus bertambah, karena konsumsi nasional cenderung meningkat dari tahun ke tahun. Penurunan ini disebabkan berkurangnya luas panen kedelai sebanyak 19 ribu hektar (3.05%) serta terjadi penurunan produktivitas sebesar 0.38 % (Sudaryanto dan Swastika 2007). Data lain menyebutkan penurunan luas panen mencapai 55.56 ribu hektar, yang diperkirakan terjadi di Jawa (BPS 2012). Untuk mengatasi kekurangan tersebut perlu dicari alternatif pemanfaatan kacang-kacangan selain kedelai. Disamping untuk menutup kekurangan produksi kedelai, juga meningkatkan kualitas tempe (Edinur 2008).
Tempe adalah sumber protein yang penting dalam menu makanan Indonesia yang merupakan bahan makanan lauk pauk nabati atau sebagai sumber protein nabati. Di Indonesia sendiri sekitar 24% masyarakatnya mengkonsumsi tempe setiap harinya. Selain mudah ditemui dan harganya murah, tempe memiliki kandungan gizi yang baik untuk tubuh. Selain memiliki kandungan gizi dan kandungan protein yang tinggi, tempe memiliki manfaat lain dari segi kesehatan. Sebagai sumber nutrisi, tempe berperan sebagai sumber protein dan mineral, sedangkan sebagai obat dan penunjang kesehatan, tempe berperan sebagai antidiare, antibakteri dan antihipertensi. Wang dan Hesseltine (1981) menyatakan bahwa Rhizopus oligosporus bahkan dapat mencegah akumulasi aflatoksin yang ada pada kedelai dengan melakukan hidrolisis.
2
yang berfungsi sebagai antihipertensi (Suarsana et al. 2008). Penelitian yang dilakukan oleh Astawan et al. (1997), hidrolisat tempe kacang kedelai terbukti memiliki aktifitas sebagai antihipertensi baik secara in vitro maupun secara in vivo. Secara in vitro, hidrolisat tersebut mampu menghambat aktivitas Angiotensin Converting Enzim (ACE) atau enzim pengubah angiotensin. Selain itu juga dibuktikan melalui hewan coba dengan tikus hipertensi, bahwa tikus yang diberi hidrolisat tempe terjadi penurunan tekanan darah, baik sistolik maupun diastolik (Astawan et al. 1997). Hal ini berkaitan dengan peptida yang dimiliki tempe sebagai hasil dari hidrolisis enzim protease yang mampu menghambat kerja enzim pengubah angiotensin (Angiotensin Converting Enzyme (ACE)) yang dapat meningkatkan tekanan darah.
Salah satu jenis kacang-kacangan yang dapat dijadikan alternatif pengganti kedelai untuk bahan baku tempe adalah kacang koro pedang. Protein yang terdapat pada kacang koro pedang lebih besar dibanding dengan kacang-kacangan lain, yaitu sekitar 27.4 g. Menurut Windrati et al. (2010), PRF (protein rich flour) koro pedang memiliki kandungan protein 37.61%, lemak 4.49%, dan pati 36.70% (amilosa 31.12% dan amilopektin 68.88%). Asam amino terbesar dalam PRF koro pedang yakni asam glutamate (5.47%) (Windrati et al. 2010). Penelitian lainnya yang dilakukan oleh Suciati (2012) mengenai pembuatan tempe dari kacang koro pedang, menyatakan bahwa tempe dari kacang koro pedang memiliki tampilan dan rasa yang tidak berbeda dengan tempe kedelai. Kandungan gizi yang terkandung didalamnya pun tidak menunjukkan hasil yang terlalu berbeda (Suciati 2012). Sampai saat ini belum ada yang melaporkan kemampuan tempe koro pedang sebagai ACE inhibitor seperti tempe kedelai. Oleh sebab itu, dilakukan penelitian untuk mengukur kemampuannya sebagai inhibitor terhadap aktivitas enzim pengubah angiotensin (ACE).
Penelitian ini dilakukan untuk mengnalisis aktivitas inhibisi enzim pengubah angiotensin dari hidrolisat tempe yang menggunakan bahan baku kacang koro pedang. Peptida yang terdapat pada hidrolisat tempe koro pedang memiliki aktivitas penghambatan terhadap enzim ACE seperti halnya pada tempe kedelai. Kegiatan penelitian ini dilakukan sebagai langkah awal pemanfaatan kacang koro pedang sebagai pangan fungsional serta diversifikasi kacang kedelai khususnya untuk bahan baku tempe.
METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan selama 5 bulan (April-September 2013). Tempat pelaksanaannya di laboratorium kimia, BB Pasca Panen; Laboratorium Balai Tanah; dan Laboratorium Biologi Molekuler, BB Biogen.
Bahan
-3 amilase (A 6380), pepsin (P 6887), tripsin (T 0303), α-kemotripsin (C 4129), ACE (A 6778), dan Hippuril-His-Leu (HHL) (H 1635). Bahan kimia lain yang digunakan adalah NaOH, HCl, natrium sulfit, coomassie brilliant blue G-250, 2.4.6-trinitrobenzenesulphonic acid (TNBS), etil asetat, asam fosfat, protein BSA, buffer glisin, buffer fosfat, NaCl, dan buffer borrat. Bahan lain yang digunakan, yakni: ragi tempe, air destilata, dan aqua bidest (ddH2O) .
Alat
Peralatan yang digunakan adalah proofer untuk fermentasi tempe, waterbath, sentrifuse Beckman GA-J20, sentrifuse Tomy High Speed Refrigerator Micro TX-160, dan Spetrofotometer BIO-RAD SmartSpec Plus. Alat-alat lain yang digunakan yakni alat gelas, pipet mikro, pipet tetes, pipet volumetrik, pH meter, dan stirer.
Prosedur Penelitian
Secara keseluruhan alur penelitian dalam pengukuran aktivitas inhibisi enzim pengubah angiotensin (ACE) pada hidrolisat tempe koro pedang (Canavalia ensiformis L.) secara in vitro adalah pembuatan tempe, analisis proksimat (kadar air, kadar abu, kadar protein, kadar lemak, dan kadar karbohidrat), hidrolisis protein, pengukuran kadar protein hidrolisat, pengukuran derajat hidrolisis, dan pengukuran aktivitas enzim (Lampiran 1). Sampel yang digunakan pada penelitian ini adalah kacang kedelai mentah, tempe kedelai mentah, tempe kedelai kukus, kacang koro pedang mentah, tempe koro pedang mentah, dan tempe koro pedang kukus. Kedelai dan kacang mentah digunakan sebagai pembanding untuk melihat aktivitas inhibisi. Tahapan metode penelitian secara lebih lanjut akan dibahas pada pemaparan dibawah ini :
Pembuatan Tempe Koro Pedang
Proses pembuatan tempe koro pedang dilakukan dengan modifikasi dari metode Suciati (2012) yang dikembangkan oleh BB Pasca Panen. Sebanyak 300 g kacang koro direndam selama 48 jam pada wadah plastik dengan air bersih. Koro selanjutnya dikeringkan dengan oven pengering bersuhu 70 ˚C selama 24 jam. Koro pedang kering dikupas dengan mesin pengupas kacang dan digiling dengan disc mill. Setelah ukurannya kecil, kacang koro dicuci kembali dan direbus selama 20 menit dilanjutkan dengan perendaman kembali dengan air bersih selama 50 jam hingga pH-nya menjadi ±4.5. Setelah pH mencapai 4.5 kacang dicuci, dikukus selama 15 menit dan ditiriskan. Setelah kacang dalam suhu ruang, selanjutnya diberi ragi dengan perbandingan ragi 0.1% dari total berat bahan. Kacang yang telah diberi ragi, dibungkus dengan plastik yang telah dilubangi lalu dirapatkan. Kemudian dilakukan fermentasi dengan menyimpan bahan di dalam proofer (alat untuk menginkubasi tempe) pada suhu 30 ˚C selama 48 jam. Setelah proses fermentasi, tempe siap untuk dilakukan uji selanjutnya.
Pembuatan Tempe Kedelai
4
kemudian direndam selama 60 menit. Selanjutnya kacang direbus selama 50 menit dilanjutkan dengan perendaman kembali dengan air bersih selama 24-36 jam hingga pH-nya menjadi ±4.5. Setelah pH mencapai 4.5 kacang dicuci, dikukus selama 20 menit dan ditiriskan. Setelah kacang dalam suhu ruang, selanjutnya diberi ragi dengan perbandingan ragi 0.1% dari total berat bahan. Kacang yang telah diberi ragi, dibungkus dengan plastik yang telah dilubangi lalu dirapatkan. Kemudian dilakukan fermentasi dengan menyimpan bahan di dalam proofer (alat untuk menginkubasi tempe) pada suhu 30 ˚C selama 48 jam. Setelah proses fermentasi tempe siap untuk dilakukan uji selanjutnya.
Analisis Proksimat (AOAC 2005)
Analisis proksimat dilakukan melalui beberapa tahapan, meliputi preparasi sampel, penentuan kadar air, penentuan kadar abu, penentuan kadar lemak, penentuan kadar protein, dan penentuan kadar karbohidrat. Sampel terdiri dari 3 perlakuan yaitu kacang koro pedang mentah, tempe koro pedang mentah, dan tempe koro pedang kukus. Sebagai pembanding dari kacang koro pedang digunakan kedelai dengan perlakuan yang sama, yaitu kacang kedelai, tempe kedelai mentah, dan tempe kedelai kukus. Sampel dihaluskan hingga menjadi bubuk.
Pengukuran kadar air pada penelitian ini menggunakan metode sebagai berikut: Cawan keramik yang akan digunakan untuk mengukur bobot sampel, dioven terlebih dahulu selama 30 menit pada suhu 100-105oC, kemudian didinginkan dalam desikator untuk menghilangkan uap air dan ditimbang. Sampel ditimbang sebanyak 2 gram dalam cawan yang sudah dikeringkan kemudian dioven pada suhu 100-105oC selama 6 jam lalu didinginkan dalam desikator selama 30 menit dan ditimbang. Tahap ini diulangi hingga dicapai bobot yang konstan. Kadar air dihitung dengan rumus:
adarair bb Cx 00
adar air bk 00 kadar air ( bb)kadar air ( bb) x 00
Keterangan :
A = bobot cawan kosong, B = bobot cawan + sampel , C = bobot cawan akhir
Pengukuran kadar abu pada penelitian ini menggunakan metode oven. Cawan porselin yang akan digunakan untuk mengukur bobot sampel, dikeringkan menggunakan oven selama 30 menit pada suhu 100-105 oC. Cawan yang sudah dikeringkan tersebut kemudian didinginkan dalam desikator untuk menghilangkan uap air dan ditimbang. Sampel ditimbang sebanyak 2 gram dalam cawan yang sudah dikeringkan kemudian dibakar di atas nyala pembakar sampai tidak berasap dan dilanjutkan dengan pengabuan di dalam tanur pada suhu 550-600 oC sampai pengabuan sempurna. Sampel beserta cawan porselin di masukkan ke dalam desikator, kemudian bobotnya ditimbang. Tahap pembakaran dalam tanur diulangi sampai didapat bobot yang konstan. Kadar abu dihitung dengan rumus:
adar abu bb C x 00
adar abu bk 00kadar abu ( bb)kadar air ( bb)x 00
Keterangan :
5 Analisis kadar protein dilakukan dengan metode Kjeldahl. Sampel yang akan diuji ditimbang sebanyak 0.1-0.5 gram dimasukkan ke dalam labu kjeldahl 100 ml, ditambahkan dengan ¼ buah tablet kjedahl, kemudian didekstruksi (pemanasan dalam keadaan mendidih) sampai larutan menjadi hijau jernih dan SO2 hilang. Larutan dibiarkan dingin dan dipindahkan ke labu 50 ml dan diencerkan dengan akuades sampai tanda tera, dimasukkan ke dalam alat destilasi, ditambahkan dengan 5-10 ml NaOH 30-33% dan dilakukan destilasi. Destilat ditampung dalam larutan 10 ml asam borat 3% dan beberapa tetes indikator (larutan bromcresol green 0.1% dan larutan metil merah 0.1% dalam alkohol 95% secara terpisah dan dicampurkan antara 10 ml bromcresol green dengan 2 metil merah) kemudian dititrasi dengan larutan HCl 0.02 N sampai larutan berubah warnanya menjadi merah muda. Kadar protein dihitung dengan menggunakan persamaan sebagai berikut:
adar protein bb ol. Cl ol. blanko . Cl 4.007 00 obot sampel (mg)
Keterangan :
FK = Faktor koreksi, 5.75 untuk kedelai dan 6.25 untuk koro pedang
Pengukuran kadar lemak menggunakan metode soxhlet. Labu lemak yang akan digunakan dioven selama 30 menit pada suhu 100-105oC, kemudian didinginkan dalam desikator untuk menghilangkan uap air dan ditimbang. Sampel yang akan diukur kadar ditimbang sebanyak 2 gram lalu dibungkus dengan kertas saring, ditutup dengan kapas bebas lemak dan dimasukkan ke dalam alat ekstraksi sokhlet yang telah dihubungkan dengan labu lemak yang telah dioven dan diketahui bobotnya. Pelarut heksan dituangkan sampai sampel terendam dan dilakukan refluks atau ekstraksi lemakselama 5-6 jam atau sampai pelarut lemak yang turun ke labu lemak berwarna jernih. Pelarut lemak yang telah digunakan, disuling dan ditampung setelah itu ekstrak lemak yang ada dalam labu lemak dikeringkan dalam oven bersuhu 100-105oC selama 1 jam, lalu labu lemak didinginkan dalam desikator dan ditimbang. Tahap pengeringan labu lemak diulangi sampai diperoleh bobot yang konstan. Kadar lemak dihitung dengan menggunakan persamaan berikut:
adar lemak bb C 00
Keterangan:
A = bobot labu lemak kosong , B = bobot sampel, C = bobot akhir labu lemak
Kadar karbohidrat dihitung sebagai sisa dari kadar air, abu, lemak dan protein. Pada analisis ini diasumsikan bahwa karbohidrat merupakan bobot sampel selain air, abu, lemak dan protein. perhitungan kadar karbohidrat dengan metode by difference menggunakan persamaan sebagai berikut:
kadarkarbohidrat 00 (kadarair kadarabu kadarprotein kadarlemak)
Hidrolisis Protein (Metode Rui et al. 2012)
6
suhu 37°C selama 3 menit. Hidrolisis dilanjutkan secara berturut-turut oleh
pepsin, tripsin dan α-kemotripsin dengan E/S: 1/250 (b/b, berdasarkan kandungan
protein). Hidrolisis berlangsung selama 10 menit dan selanjutnya enzim diinaktivasi dengan pemanasan dalam air mendidih selama 10 menit. Campuran disentrifugasi (12.000 g, 4°C selama 20 menit). Supernatan disimpan untuk uji lebih lanjut.
Derajat Hidrolisis (DH) (Modifikasi Benjakul & Morrissesey 1997)
Derajat hidrolisis (DH, %) diukur berdasarkan perbandingan total leusin dengan jumlah kadar protein total. Metode ini diacu dari modifikasi Benjakul and Morrissesey (1997). Hidrolisat sebanyak 125 µl ditambahkan dengan 2.0 ml bufer fosfat 0.2 M pH 8.2 dan 1 ml larutan TNBS (trinitrobenzenesulfonic acid) 0.01%.
Larutan dicampur seluruhnya dan diinkubasi pada water bath pada suhu 50 ˚C
selama 30 menit pada kondisi gelap. Untuk menghentikan reaksi, 2 ml natrium sulfit 0.1 M ditambahkan. Campuran didinginkan selama 15 menit pada suhu ruang. Absorbansi diukur pada panjang gelombang 420 nm dan kurva standar yang digunakan yakni kurva standar L-leusin.
Analisis Protein (Bradford 1976)
Analisis konsentrasi protein hidrolisat dilakukan menggunakan metode Bradford (1976). Larutan Bradford dibuat dengan cara sebagai berikut: sebanyak 100 mg coomassie brilliant blue (CBB) G-250 dilarutkan dalam 50 ml etanol 95%. Setelah itu 100 ml asam fosfat 85% ditambahkan. Terakhir larutan diencerkan hingga 1 liter. Larutan disaring dengan kertas saring.
Larutan standar dibuat menggunakan protein BSA (bovine serum albumin). Sebanyak 100 mg BSA ditimbang dan ditambahkan 25 ml akuades. Larutan dikocok perlahan, setelah larut, diencerkan sampai 50 ml. Konsentrasi akhir larutan stok untuk standar ini adalah 2 mg/ml. Sederetan larutan standar disiapkan menggunakan stok.
Langkah selanjutnya adalah memipet masing-masing larutan dalam tiap tabung sebanyak 0.1 ml dan dimasukkan ke dalam tabung reaksi lain yang bersih. Sebanyak 5 ml pereaksi Bradford ditambahkan ke dalam masing-masing tabung reaksi. Blanko dibuat dengan cara mencampurkan 0.1 ml aquades dan direaksikan dengan 5 ml pereaksi Bradford. Setelah sekitar 20 menit, masing-masing
campuran reaksi diukur absorbansinya pada 595 nm.
Penentuan Daya Inhibisi Terhadap Aktivitas ACE secara In Vitro
7
HASIL DAN PEMBAHASAN
Hasil
Tempe Koro Pedang
Hasil pembuatan tempe koro pedang dapat dilihat pada Gambar 1. Kacang koro pedang memiliki ukuran yamg jauh lebih besar daripada kacang kedelai, sehingga diperlukan pencacahan sebelum dijadikan tempe (Gambar 1). Tempe koro pedang memiliki tampilan yang menyerupai tempe kedelai. Miselium dari kapang menutupi permukaannya secara merata dengan warna putih. Tempe koro pedang memiliki aroma tempe normal. Tempe ini memiliki tekstur yang hampir mirip dengan tempe kedelai, padat dan kompak. Tekstur padat dan biji koro kompak karena adanya miselia yang mengikat sehingga jika dipotong tempe tidak terburai atau pecah.
Gambar 1 Visualisasi kacang koro pedang dan tempe koro pedang
Kacang koro mentah yang belum dikupas (a), kacang koro pedang yang telah direndam, dikeringkan, dan dicacah (b), dan tempe koro pedang dengan waktu fermentasi 48 jam (c)
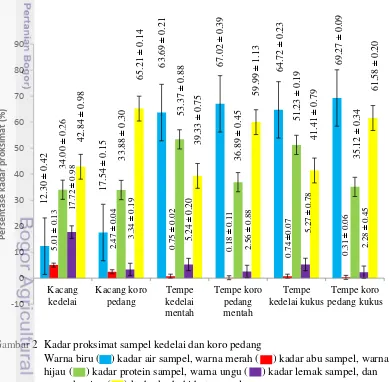
Kadar Proksimat dari Sampel Kedelai dan Koro Pedang
Analisis proksimat merupakan analisis yang meliputi uji kadar air, abu, protein, lemak, dan karbohidrat. Analisis ini dilakukan untuk memperoleh nilai kandungan gizi dalam sampel kedelai dan koro pedang yang dianalisis, meliputi: kacang mentah, tempe mentah, dan tempe kukus. Kedelai digunakan sebagai pembanding dari kacang koro pedang. Hasil pengukuran kadar air ini menunjukkan perbedaan antara kedelai dan koro pedang, yakni pengolahan kacang menjadi tempe meningkatkan kadar air, baik tempe dari kacang kedelai maupun kacang koro pedang (Gambar 2). Gambar 1 juga memperlihatkan bahwa kandungan air kacang koro pedang lebih tinggi dari kacang kedelai. Sehingga, saat diolah menjadi tempe pun memiliki kadar air yang jauh lebih tinggi dari tempe kedelai. Dari hasil tersebut juga terlihat perubahan yang tetap dari kacang mentah maupun olahannya.
Hasil pengukuran kadar abu menunjukkan bahwa kandungan senyawa anorganik pada kedelai lebih besar dibandingkan dengan koro pedang. Hasil ini juga memperllihatkan bahwa pengolahan bahan dasar kacang menjadi tempe, menurunkan kadar abu (Gambar 2). Hasil kadar abu tertinggi diperoleh dari kacang kedelai sebesar 5.01%, sedangkan kacang koro hanya memiliki kadar abu sebesar 2.47 % (Gambar 2).
8
Hasil pengukuran kadar protein, kedelai memiliki kadar protein yang lebih unggul dibandingkan dengan koro pedang. Saat bahan baku mentah berupa kacang, kadar protein kedelai mencapai 34% dan kacang koro pedang 33.85 %. Setelah diolah menjadi tempe, kedelai memiliki kadar protein total yang jauh lebih tinggi dibandingkan koro pedang. Tempe kedelai memiliki kadar protein sebesar 53.37% dan tempe koro pedang 36.89% (Gambar 2). Dari hasil ini, kedelai memiliki kandungan protein yang lebih unggul dibandingkan koro pedang.
Hasil analisis kadar lemak yang dilakukan pada sampel menunjukkan hasil yang jauh berbeda antara kedelai dan koro pedang (Gambar 2). Kadar lemak pada biji kacang kedelai sebesar 17.72% jauh lebih tinggi dibandingkan dengan biji kacang koro yang hanya meperoleh 3.34%. Saat diolah menjadi tempe, kadar lemak pada kedelai mengalami penurunan yang cukup drastis menjadi 5.24%, sedangkan koro pedang tidak mengalami banyak penurunan. Analisis proksimat yang terakhir adalah kadar karbohidrat. Analisis yang dilakukan berdasarkan metode by difference, artinya jumlah karbohidrat yang terdapat pada sampel diperoleh dari perbedaan antara jumlah kandungan air, protein, lemak dan abu dengan rumus karbohidrat yaitu 100%-(protein+lemak+abu+air). Hasilnya diperoleh jumlah karbohidrat koro pedang lebih tinggi dibandingkan dengan kedelai (Gambar 2).
Gambar 2 Kadar proksimat sampel kedelai dan koro pedang
9 Derajat Hidrolisis
Hasil dari pengukuran derajat hidrolisis, sampel dari masing-masing kacang mentah (kedelai dan koro pedang) memiliki derajat hidrolisis yang tinggi, yakni 18.86% untuk kedelai dan 16.19% untuk koro pedang (Gambar 3). Setelah kacang tersebut diolah menjadi tempe, derajat hidrolisis mengalami penurunan. Secara garis besar, tempe mentah memiliki derajat hidrolisis yang lebih rendah dibandingkan dengan tempe kukus. Akan tetapi, tempe kacang koro pedang memiliki persentase derajat hidrolisis yang lebih besar dibandingkan dengan kacang kedelai, baik tempe mentah maupun tempe kukus. Perbandingan hasil derajat hidrolisis ini dapat dilihat pada Gambar 3. Data derajat hidrolisis secara lengkap disajikan pada Lampiran 7.
Gambar 3 Perbandingan hasil pengukuran derajat hidrolisis
Warna biru ( ) merupakan sampel kedelai dan warna merah ( ) merupakan sampel koro pedang
Kadar Protein Terlarut dari Sampel Hidrolisat
Penentuan kadar protein Bradford untuk mendapatkan konsentrasi protein terlarut pada sampel hidrolisat. Hasil yang diperoleh, kacang kedelai memiliki konsentrasi protein terlarut sebesar 165.30 mg/g sampel berat kering (bk). Setelah diolah menjadi tempe, terjadi penurunan konsentrasi protein terlarut yakni 16.46 mg/g sampel bk untuk tempe kedelai mentah dan 26.22 mg/g sampel bk untuk tempe kedelai kukus (Gambar 4). Kacang koro pedang hanya memperoleh konsentrasi protein terlarut sebesar 37.52 mg/g sampel bk. Tempe koro pedang memperoleh konsentrasi sebesar 0.67 mg/g sampel bk untuk tempe mentah dan 1.84 mg/g sampel bk (Gambar 4).
Gambar 4 Kadar protein bobot kering sampel hidrolisat kedelai
10
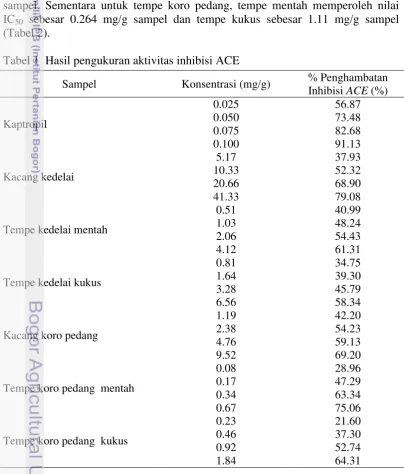
Aktivitas Inhibisi ACE
Hasil pengukuran aktivitas ACE Inhibitor tertera pada Tabel 1. Data tersebut menunjukkan tempe koro pedang mentah memiliki range penghambatan ACE dari 28.96%-75.06%. Sedangkan tempe koro pedang kukus mengalami penurunan aktivitas penghambatan (Tabel 1). Kacang kedelai dan tempe kedalai masih unggul dari hasil pengukuran tersebut. Aktivitas penghambatan tertinggi pada kedelai ditemukan pada perlakuan sampel kacang kedelai dengan konsentrasi 41.33 mg/g sampel, dengan aktivitas penghambatan sebesar 79.08%. Sementara, untuk koro pedang, diperoleh aktivitas penghambatan tertinggi pada tempe koro mentah dengan konsentrasi 0.67 mg/g sampel, dengan aktivitas penghambatan sebesar 75.06%.
Nilai IC50 yang diperoleh, kacang kedelai sebesar 10.36 mg/g sampel, tempe kedelai mentah 1.69 mg/g sampel, dan tempe kedelai kukus 17.83 mg/g sampel. Untuk koro pedang, kacang mentah memperoleh nilai IC50 sebesar 2.3 mg/g sampel. Sementara untuk tempe koro pedang, tempe mentah memperoleh nilai IC50 sebesar 0.264 mg/g sampel dan tempe kukus sebesar 1.11 mg/g sampel (Tabel 2).
Tabel 1 Hasil pengukuran aktivitas inhibisi ACE
11 Tabel 2 Nilai IC 50 dari masing-masing sampel hidrolisat
Sampel Nilai IC 50
Kaptropil 0.0044 mg/g
Kacang kedelai 10.36 mg/g sampel kering
Tempe kedelai mentah 1.69 mg/g sampel kering Tempe kedelai kukus 17.83 mg/g sampel kering Kacang koro pedang 2.30 mg/g sampel kering Tempe koro pedang mentah 0.26 mg/g sampel kering Tempe koro pedang kukus 1.11 mg/g sampel kering
Pembahasan
Tempe Koro Pedang
Bahan dasar yang digunakan adalah kacang koro pedang (Canavalia ensiformis L.) yang berasal dari petani di daerah Temanggung. Biji kacang koro pedang kemudian dilakukan perendaman selama 48 jam. Tujuan dari perendaman ini untuk menghilangkan kandungan HCN pada kacang koro pedang. HCN pada kacang koro pedang berupa glikosianida sianogenik, yang akan terurai menjadi asam sianida saat teroksidasi (Suciati 2012). Di dalam proses pencernaan, senyawa ini mudah terserap dan masuk ke dalam darah sehingga sangat berbahaya apabila terdapat dalam konsentrasi yang tinggi. Menurut Mahendradatta (2007), konsentrasi HCN yang berbahaya bagi tubuh berkisar antara 0.5-3.5 mg/kg berat badan.
Pengolahan tempe koro pedang ini, berdasarkan pengembangan metode dari BB Pascapanen. Pertama kacang koro pedang dikeringkan selama 24 jam untuk mempermudah pengupasan kulitnya. Setelah itu, kacang digiling dan dicuci kembali, kemudian dilakukan proses perebusan selama 20 menit. Proses perebusan awal ini agar kacang tidak terlalu keras. Selanjutnya kacang direndam kembali kurang lebih 2 hari untuk menurunkan pH sekitar 4.2 - 4.8. pH tersebut merupakan pH optimum agar kapang pada tempe dapat tumbuh. Terakhir adalah proses peragian, konsentrasi ragi yang digunakan 0.1% atau 0.15% dari berat kacang yang diolah. Proses peragian ini harus merata, karena apabila tidak merata miselium kapang yang tumbuh tidak menutupi semua bagian tempe. Proses tersebut terjadi perubahan baik oleh proses fisik maupun proses enzimatik karena adanya aktivitas mikroorganisme. Keterlibatan mikroorganisme pada proses pembuatan tempe terutama terjadi pada proses perendaman oleh bakteri-bakteri pembentuk asam dan proses fermentasi oleh kapang khususnya Rhizopus oligosporus (Ariani 2001).
Akibat perubahan-perubahan tersebut tempe menjadi lebih enak, lebih bergizi, dan lebih mudah dicerna. Salah satu faktor penting dalam perubahan tersebut adalah terbebasnya senyawa-senyawa isoflavon dalam bentuk bebas (aglikon) dan adanya 6,7,4’-trihidroksi-isoflavon, yang terdapat pada tempe tetapi tidak terdapat pada kacang. Senyawa tersebut ternyata berpotensi tinggi sebagai antioksidan (Gyorgy et al. 1964), antihemolitik, penurun tekanan darah, dan anti kanker (Zilleken 1986) dibandingkan dengan jenis isoflavon yang lainnya.
12
kuat dan merata, sehingga bila diiris tempe tersebut tidak hancur. Tempe yang kurang baik dicirikan dengan kondisi tempe yang terlalu basah, tidak kompak, perrmukaan tempe bercak-bercak hitam, tempe berbau amoniak dan alkohol, tempe pecah-pecah, pertumbuhan kapang tidak merata, dan tempe mengandung racun. Faktor-faktor yang sangat mempengaruhi keberhasilan pembuatan tempe adalah oksigen, suhu, jenis laru tempe, dan nilai pH (derajat keasaman) (Syarif et al.1999).
Persentase Proksimat Sampel Kedelai dan Koro Pedang
Analisis proksimat meliputi analisis kadar air, kadar abu, kadar protein, kadar lemak, dan kadar karbohidrat. Analisis ini dilakukan untuk memperoleh nilai kandungan gizi pada kacang mentah serta olahannya (tempe) yang dihasilkan.
Air dalam bahan pangan akan mempengaruhi penampakan, tekstur, serta cita rasa makanan. Air juga akan mempengaruhi daya tahan bahan pangan terhadap serangan mikroba (Fardiaz 1989). Semakin tinggi kadar air suatu bahan pangan maka semakin rendah mutu bahan pangan tersebut. Penentuan kadar air bertujuan sebagai data pendukung untuk menghitung kadar protein terhidrolisis basis kering. Tingginya kadar air pada produk pangan dapat menyebabkan kerusakan baik itu secara mikrobiologis, kimiawi, maupun enzimatis (Fardiaz 1989). Berdasarkan Gambar 1, kadar air tempe sangat meningkat dibandingkan dengan kacang mentah. Hal ini disebabkan saat proses pembuatan tempe terjadi hidrasi terutama pada saat perendaman dan perebusan, sehingga berat kacang dapat meningkat karena air akan mudah berdifusi ke dalam dinding sel. Proses menunjukkan total mineral yang terkandung dalam bahan tersebut (Faridah et al. 2008). Kadar abu kacang kedelai jauh lebih tinggi dibandingkan dengan kacang koro pedang. Hal ini berarti, kacang kedelai memiliki kandungan mineral yang lebih banyak dibandingkan dengan koro pedang. Setelah kacang diolah menjadi tempe, banyak mineral yang hilang sehingga kadar abunya menjadi menurun. Penurunan ini dapat disebabkan proses yang terjadi pada pembuatan tempe, karena pada proses pembuatan tempe terjadi pengupasan kulit kacang. Sebagian besar mineral pada kacang terkandung dikulitnya. Kadar abu yang diperoleh dari tempe kedelai dan tempe koro pedang sesuai dengan syarat mutu tempe berdasarkan SNI (BSN 2009) yang tidak melebihi 1.5% (b/b).
13 bentuk yang lebih sederhana, seperti asam amino bebas, sehingga lebih banyak lagi asam amino yang terukur (Syarif et al. 1999). Nitrogen terlarut juga meningkat selama proses fermentasi. Peningkatan nitrogen terlarut akan turut meningkatkan kadar protein kasar. Selain itu, Abu et al. (1999) juga menyatakan bahwa selama tidak ada sumber nitrogen dari luar yang disediakan, peningkatan protein kemungkinan diakibatkan oleh konversi beberapa protein tanaman atau komponen nitrogen lain.
Hasil pengukuran kadar lemak, kacang kedelai memiliki kandungan lemak tertinggi. Apabila dibandingkan dengan kacang koro pedang, kacang kedelai memiliki kandungan lemak yang jauh lebih tinggi yakni 17.72% : 3.34%. Setelah mengalami pengolahan, masing-masing kacang mengalami penurunan kandungan lemak. Tempe kedelai hanya memperoleh kandungan lemak 5.24% dan tempe koro pedang memperoleh 2.56%. Kasmidjo (1996) menyatakan bahwa proses fermentasi pada pembuatan tempe dapat menurunkan kadar lemak.
Karbohidrat mempunyai peranan penting dalam menentukan karakteristik bahan makanan, misalnya rasa, warna, tekstur, dan lain-lain. Sedangkan dalam tubuh, karbohidrat berguna untuk mencegah timbulnya ketosis, pemecahan protein tubuh yang berlebihan, kehilangan mineral, dan berguna untuk membantu metabolisme lemak dan protein. Kacang mentah memiliki kandungan karbohidrat yang lebih banyak dibandingkan dengan tempe. Data tersebut menunjukkan bahwa proses fermentasi dapat menurunkan kadar karbohidrat kacang komak. Selama perendaman kacang komak selama dua malam pada pembuatan tempe kacang komak menyebabkan penurunan kadar karbohidrat karena karbohidrat tersebut digunakan oleh bakteri asam laktat, terutama karbohidrat golongan oligosakarida, seperti stakiosa, verbakosa, dan rafinosa. Hermana et al. (2001) menyatakan bahwa penurunan kadar karbohidrat juga terjadi selama proses fermentasi yang disebabkan oleh pemecahan gula-gula kompleks seperti pati, stakiosa, dan rafinosa menjadi gula-gula yang mudah dicerna (digestible sugars) oleh kapang.
Persentase Derajat Hidrolisis dari Sampel Hidrolisat
Hidrolisat protein merupakan protein yang mengalami proses hidrolisis atau degradasi secara kimia maupun secara enzimatik. Derajat hidrolisis (DH) menyatakan kemampuan suatu enzim untuk menghidrolisis atau menguraikan protein (Lieske and Konkrad 1994). Derajat hidrolisis substrat bergantung pada keadaan substrat dan kondisi lingkungan, disamping pada spesifikasi enzim. Derajat hidrolisis merupakan salah satu parameter dasar yang perlu dikendalikan karena sifat dari hidrolisat protein berhubungan erat dengan parameter tersebut (Nielsen 1997). Pengendalian ini diperlukan karena daya hidrolitik suatu enzim dapat bervariasi berdasarkan sumber dan substrat yang digunakan. Melalui teknik hidrolisis, protein dari suatu bahan dapat diubah menjadi senyawa asam amino L, nukleotida, dan berbagai ragam peptida.
14
menguntungkan karena memungkinkan untuk memproduksi hidrolisat fragmen peptida yang berbeda. Menurut Kunts (2000), hidrolisat protein mempunyai range aplikasi yang sangat luas terkait dengan sifat fungsional atau sifat nutrisinya.
Berdasarkan hasil dari Gambar 3, terlihat bahwa kacang mentah memiliki derajat hidrolisis (DH) yang tinggi dibandingkan dengan produk tempe. Kacang kedelai memiliki derajat hidrolisis (DH) sebesar 18.86% dan kacang koro pedang sebesar 16.19% (Gambar 3). Pengolahan kacang menjadi tempe, ternyata dapat menurunkan derajat hidrolisis (DH). Hal ini terlihat pada Gambar 3, tempe kedelai maupun tempe koro pedang mengalami penurunan persentase derajat hidrolisis. Tempe kukus memiliki persentase derajat hidrolisis lebih tinggi daripada tempe mentah (Gambar 3). Nilai DH yang tinggi mengidentifikasikan bahwa protein tersebut mudah untuk dihidrolisis. Hal ini dipengaruhi oleh masih banyaknya ikatan peptida pada protein tersebut. Besarnya derajat hidrolisis terkait erat dengan jumlah produk hidrolisat yang dihasilkan, atau dengan kata lain besarnya derajat hidrolisis memiliki kecenderungan yang sama dengan jumlah protein terlarut atau gugus amino bebas (Hernandez et al. 2011). Derajat hidrolisis ini sebanding dengan konsentrasi terlarutnya. Semakin besar kadar protein terlarutnya maka akan semakin besar pula derajat hidrolisis dari sampel tersebut (Hernandez et al. 2011).
Protein pada tempe sudah didegradasi oleh enzim protease yang dihasilkan kapang tempe saat proses fermentasi. Fermentasi tempe, memecah sebagian protein menjadi peptida-peptida oleh enzim protease yang dihasilkan oleh kapang. Pemecahan peptida tersebut dipengaruhi oleh konsentrasi enzim dan lamanya waktu fermentasi. Hubungannya dengan hidrolisis protein ini, protein pada kacang mentah resisten terhadap daya cerna enzim pencernaan (tripsin dan α-kemotripsin). Hal inilah yang diduga menyebabkan tempe memiliki derajat hidrolisis yang lebih rendah dari kacang mentah. Dugaan sementara, proses fermentasi ini dapat meningkatkan resistensi, sehingga derajat hidrolisis yang dihasilkan dari tempe lebih kecil daripada kacang mentah. Rui et al. (2012) menyatakan kacang-kacangan memiliki protein dominan berupa phaseolin yang resisten terhadap
pencernaan enzim tripsin dan α-kemotripsin. Hal tersebut dipengaruhi oleh
struktur yang kompak serta tingginya konformasi lembar-β (Rui et al. 2012). Peningkatan derajat hidrolisis setelah pengukusan tempe disebabkan proses pemanasan saat tempe dikukus mampu mendegradasi protein. Saat tempe mengalami pengukusan, protein terdenaturasi yang menyebabkan protein menjadi terdegradasi sehingga derajat hidrolisisnya meningkat.
Konsentrasi Protein Terlarut dari Sampel Hidrolisat
Konsentrasi protein terlarut hidrolisat menggunakan metode Bradford. Hasil yang diperoleh tertera pada Gambar 4. Hidrolisat kacang mentah memiliki konsentrasi protein terlarut yang jauh lebih besar dari pada tempe. Hal ini dikarenakan, kacang mentah belum mengalami proses pemanasan yang dapat merusak protein di dalamnya. Lain halnya dengan tempe, saat proses pembuatannya, kacang berkali-kali mengalami proses pemanasan seperti perebusan dan pengukusan. Akan tetapi, saat tempe dikukus, konsentrasi protein terlarutnya meningkat dari tempe mentah (Gambar 4).
15 terputus menjadi peptida-peptida sederhana sehingga kelarutan protein semakin meningkat. Hal inilah yang terjadi pada kacang mentah. Protein pada kacang mentah terputus menjadi peptida pendek yang menyebabkan kelarutannya meningkat. Semakin lama reaksi hidrolisis, reaksi enzim dengan substrat juga semakin lama, sehingga tingkat hidrolisis semakin tinggi dan dihasilkan molekul-molekul protein yang pendek dan kelarutannya meningkat.
Konsentrasi protein terlarut ini sebanding dengan derajat hidrolisis. Saat kacang diolah menjadi tempe, derajat hidrolisisnya menurun begitu pula dengan konsentrasi protein terlarutnya. Dari hasil ini dapat diketahui, peptida yang dihasilkan dari fermentasi tempe sebagian besar protein tak terlarut. Hal ini terbukti dari total protein meningkat setelah proses fermentasi (setelah menjadi tempe) yang ditunjukkan dari hasil proksimat kadar protein (Gambar 2). Hal inilah yang menyebabkan nilai derajat hidrolisis dan konsentrasi terlarut tempe menurun.
Aktivitas Inhibisi ACE
Angiotensin Converting Enzyme (ACE) merupakan glikoprotein peptidildipeptida hidrolase yang termasuk ke dalam kelas enzim hidrolase. enzim ini membutuhkan senyawa zink dan klorida untuk mendapatkan bentuk aktifnya. Jenis peptida ini pada dasarnya mengatalisis reaksi angiotensin I menjadi angiotensin II, dengan memecah dipeptida histidil-leusina dari ujung˗C angiotensin I menghasilkan angiotensin II dan asam hipurat. Ada dua bentuk dari enzim ini pada manusia yaitu ubiquitous somatic ACE dan the sperm-specific germinal ACE. ACE sudah lama dikenal sebagai bagian kunci pada sistem renin angiotensin yang penting pada pengaturan tekanan darah. Pada saat terjadi penurunan tekanan darah di arteriol ginjal (glomerulus), maka sel ginjal membentuk renin. Renin dirilis dalam plasma dan membentuk kesetimbangan berupa angiotensinogen yang diubah menjadi angiotensin I. Setelah terbentuk angiotensin I, oleh ACE kemudian diubah menjadi Angiotensin II serta berikatan dengan reseptor. Hal tersebut mengakibatkan terjadinya tiga peristiwa yaitu vasokonstriksi kuat, stimulasi sekresi hormon aldosteron, dan terjadi retensi garam dan air (peningkatan absorbsi natrium oleh ginjal dan volume intravaskular). Hasil akhirnya adalah tekanan darah meningkat (Bicket 2002).
ACEmerupakan enzim multifungsi yang juga dapat mengkatalisis degradasi bradikinin (vasodilator yang dapat melebarkan pembuluh darah) dan enkefalin. Oleh karena itu, penghambatan aktivitas ACE dapat mengurangi peranan angiotensin II tetapi dapat meningkatkan kadar bradikinin dan enkefalin, sehingga akan menurunkan tekanan darah. Hasil dari pengukuran aktivitas inhibisi ACE, diperoleh daya hambat yang cukup tinggi antara kaptropil, kedelai dan koro pedang. Nilai aktivitas inhibisi kedelai masih lebih besar dibandingkan dengan koro pedang (Tabel 1). Kaptropil digunakan sebagai kontrol positif terhadap penghambatan aktivitas ACE, sedangkan kedelai digunakan sebagai pembanding karena kedelai digunakan sebagai bahan baku tempe yang saat ini ada di pasaran.
16
aktivitas penghambatan yang tinggi. Aktivitas tertinggi dari kontrol positif diperoleh sebesar 91.13% dengan konsentrasi sampel sebesar 0.100 mg/g. Kaptropil adalah obat antihipertensi yang menghambat pembentukan angiotensin I menjadi angiotensin II dengan mengikat sisi aktif dari enzim ACE. Kaptropil memiliki afinitas yang tinggi terhadap ACE dan berkompetisi secara kompetitif dengan substrat untuk mencegah terbentuknya angiotensin II (Hansen 1995).
IC50 digunakan untuk mengetahui kemampuan hidrolisat sampel untuk menghambat 50 % aktivitas enzim ACE. Berdasarkan nilai IC50 yang diperoleh, kacang koro pedang lebih efektif menghambat aktivitas ACE karena memiliki nilai IC50 yang lebih kecil daripada kacang kedelai. Nilai IC50 untuk tempe, tempe koro pedang memiliki nilai yang jauh lebih kecil dibandingkan dengan tempe kedelai, baik dengan perlakuan tempe mentah maupun tempe kukus. Kaptropil sebagai kontrol positif memiliki nilai IC50 yang paling rendah sebesar 0.0044 mg/g. Hasil tersebut masih berbeda jauh apabila dibandingkan dengan sampel. Hal ini disebabkan, bahwa kaptropil yang digunakan sebagai obat antihipertensi memang sudah teruji secara klinis.
Berdasarkan hasil yang diperoleh, peptida yang terfragmen saat proses fermentasi maupun hidrolisis protein, mampu bertindak sebagai ACE inhibitor. Peptida tersebut berasal dari protein kompleks koro pedang yang diurai oleh enzim protease. Menurut Astawan et al. (1997) ada berbagai faktor yang dapat mempengaruhi aktivitas penghambatan ACE. Faktor-faktor tersebut adalah enzim yang digunakan, waktu hidrolisis, dan waktu fermentasi. Penelitian ini menggunakan gabungan dari ketiga faktor itu, berdasarkan acuan dari Rui et al. (2012). Astawan et al. (1997) menyatakan, penggunaan enzim pepsin menghasilkan hidrolisat dengan aktivitas ACE inhibitor yang baik.
Waktu hidrolisis yang digunakan, disamakan untuk setiap perlakuan. Akan tetapi, untuk enzim amilase digunakan waktu yang sangat cepat. Hal ini dikarenakan enzim amilase mampu mendegradasi karbohidrat dengan cepat, sehingga tidak dibutuhkan waktu yang lama untuk mendegradasi karbohidrat yang ada pada tempe. Faktor yang ketiga adalah waktu fermentasi. Untuk koro pedang, digunakan waktu fermentasi selama 48 jam. Lama fermentasi antara 36-72 jam menyebabkan aktivitas enzim protease tidak hanya untuk menghidrolisis protein menjadi asam amino tetapi juga pembentukan amonia (Astawan et al. 1997).
SIMPULAN DAN SARAN
Simpulan
17 Saran
Perlu dilakukan penelitian lanjutan untuk menjadikan koro pedang ini sebagai bahan pangan fungsional, berdasarkan potensi yang dimiliki sebagai inhibitor ACE. Untuk itu perlu dilakukan uji in vivo. Identifikasi asam amino yang menjadi ACE inhibitor juga diperlukan untuk mengetahui secara spesifik jenis peptidanya.
DAFTAR PUSTAKA
[AOAC] Association of Official Analytical Chemist. 2005. Official Method of Analysis of The Association Analytical of Chemist. The Association of Official Analysis Chemist, Inc., Arlington.
[BPS] Balai Pusat Statistik. 2006. Angka Tetap Tahun 2005 dan Angka Ramalan II Tahun 2006 Produksi Tanaman Pangan. Badan Pusat Statistik, Jakarta. [BPS] Balai Pusat Statistik. 2012. Produksi Padi, Jagung, dan Kedelai (Angka
Tetap 2011 dan Angka Ramalan I 2012). Berita Resmi Statistik : No. 43/07/Th. XV.
[BSN] Badan Standarisasi Nasional Indonesia. 2009. SNI 3144-2009 tentang Tempe Kedelai. Jakarta: Badan Standarisasi Nasional Indonesia.
[BSN] Badan Standarisasi Nasional. 1998. Standar Nasional Indonesia untuk Tempe Kedelai. SNI 01-3144-1998. Jakarta: Badan Standarisasi Nasional. [Deptan] Departemen Pertanian. 2005. Prospek dan Arah Pengembangan
Agribisnis Kedelai. [internet] [diacu 2013 Oktober 16]. Tersedia dari: http://www.litbang.deptan.go.id
Abu OA, OO Tewe, DM Losel, and AA Onifade. 1999. Changes in lipids, fatty acids, and protein composition in sweet potato (Ipomomea batatas) after solid state fungal fermentation. Bioresource Technology,72: 189-192.
Ariani SRD. 2001. Identifikasi Senyawa Faktor-2 (Suatu Senyawa Isoflavon) dari Tempe Selama Proses Fermentasi Hari ke-0,1,2,3,4, dan 5. Paedagogia, 4(1): 2-8.
Astawan M dan Mita W. 2003. Teknologi Pengolahan Pangan Nabati Tepat Guna. Jakarta (ID): Akademika Pressindo.
Astawan M, S Yasni, Suliantari, dan Olih. 1997. Mempelajari Potensi Hidrolisat Tempe Campuran Kedelai-Beras sebagai Penurun Tekanan Darah pada Tikus Percobaan. Bul. Teknol. dan Industri Pangan, VIII (3): 45-51.
Benjakul S and Morrissey M. 1997. Protein hydrolisate from pasific whiting solid waste. J Agric Food Chem 45:3423-3430.
18
Bradford MM. 1976. A rapid and sensitive method for quantitation of microgram quantities of protein untilizing the principles of protein dye-binding. Anal Biochem 72: 234-254.
Cushman DW and Cheung HW. 1971. Spectrophotometric assay and properties of the angiotensin converting enzyme of the rabbit lung. Biochem Pharmacol 20:1637-1648.
Edinur. 2008. Standardisasi Tempe di Indonesia. Prosiding Simposium Nasional: Perkembangan Terkini tentang Tempe: Teknologi, Standardisasi dan Potensinya dalam Perbaikan Gizi serta Kesehata. Forum Tempe Indonesia, Yayasan Tempe Indonesia, dan PERGIZI Pangan Indonesia.
Fardiaz D. 1989. Hidrokoloid. Laboratorium Kimia dan Biokimia Pangan. Bogor (ID): Pusat Antar Universitas Pangan dan Gizi, Institut Pertanian Bogor. Gyorgy P, K Murata, and H Ikehata. 1964. Antioxidants isolated from fermented
soybeans tempeh. Nature. 203: 872-875.
Hermana, M Mahmud, and D Karyadi. 2001. Composition and Nutritional Value
of Tempeμ It’s Role in The Improvement of The Nutritional Value of Food.
Agranoff, J. (ed). The Complete Handbook of Tempe. Jakarta: Indonesian Tempe Foundation.
Hernandez-Ledesma B, del Mar Contreras M, Recio I. 2011. Antihypertensive peptides: Production, bioavailability and incorporation into foods. Adv Colloid Interface Sci 165: 23-35.
Hansen K et al. 1995. In vitro screening of traditional medicines for anti-hypertensive effect based on inhibiton of the angiotensin converting enzyme (ACE). J Ethnopharmacol 48:43-51.
Kasmidjo RB. 1996. Tempe : Mikrobiologi dan Kimia Pengolahan serta Pemanfaatannya. Yogyakarta: PAU Pangan dan Gizi UGM.
Kunts A. 2000. Enzymatic Modification of Soy Proteins to Improve Their Functional Properties. Magazine of Industrial Protein, 8(3): 9–11.
Lieske B and G Konkrad. 1994. Protein Hydrolysis The Key to Daging Flavoring System. Food Reviews International, 10(3): 287-312.
Mahendradatta M. 2007. Pangan Aman dan Sehat, Prasyarat Kebutuhan Mutlak Sehari-hari. Makassar: Lembaga Penerbitan Universitas Hasanuddin.
Nielsen PM. 1997. Food Proteins and Their Applications. New York: Marcel Dekker, Inc.
Rui et al. 2012. Electrophoretic profiles and angiotensin I-converting enzyme inhibitory activities of nine varieties of Phaseolus vulgaris protein hydrolysates. J Nutr Food Sci, 2(8) :1-6.
Suciati A. 2012. Pengaruh Lama Perendaman dan Fermentasi terhadap Kandungan HCN pada Tempe Kacang Koro. [Skripsi]. Makassar: Jurusan Teknologi Pertanian, Fakultas Pertanian, Universitas Hasanuddin.
19 Kedelai: teknik produksi dan pengembangan. Bogor: Puslitbang Tanaman Pangan.
Suarsana IN, BP Priosoeryanto, M Bintang, dan T Wresdiyati. 2008. Efek Hipoglikemik Ekstrak Tempe pada Tikus Diabetes yang Diinduksi dengan Aloksan. Prosiding Simposium Nasional: Perkembangan Terkini tentang Tempe: Teknologi, Standardisasi dan Potensinya dalam Perbaikan Gizi serta Kesehatan. Forum Tempe Indonesia, Yayasan Tempe Indonesia, dan PERGIZI Pangan Indonesia.
Van den Hil PJR, Nout MJR. 2011. Anti-diarrhoeal aspects of fermented soya beans, soybean and health, Prof. Hany El-Shemy (ed.), InTech, doi: 10.5772/17997 [Internet]. [diunduh 2013 Mar 6]. Tersedia pada: http://www.intechopen.com/books/soybean-and-health/anti-diarrhoeal-aspects-of-fermented-soya-beans.
Wang HL dan CW Hesseltine. 1981. Use of microbial cultures in legumes and cereal products. Food Technol. 1:79.
Windrati WS, Ahmad N, and Puspa DA. 2010. Sifat Nutrisional Protein Rich Flour (PRF) Koro Pedang (Canavalia ensiformis L.). Agrotek, 4(1): 18-26. Zilleken F. 1986. First draft meeting on biotechnology. Jakarta: BPP Teknologi,
Lampiran 1 Diagram alir penelitian
Pengolahan kacang menjadi tempe
Tempe dikukus selama 10 menit
Kacang Kedelai Kacang Koro
Pedang
Tempe
Tempe Kukus
Hidrolisis Protein
Analisis Proksimat
Kadar Lemak Kadar Protein Kadar Abu
Kadar Air
Pengukuran Derajat Hidrolisis
Pengukuran konsentrasi protein sampel
21 Lampiran 2 Hasil pengukuran kadar air kedelai dan koro pedang
Sampel Bobot Cawan Kosong Bobot Sampel
Bobot Cawan +Sampel Kering
U 1 U 2 U 3 U 1 U 2 U 3 U 1 U 2 U 3
kacang kedelai 23.029 23.444 18.357 2.001 2.001 2.001 24.789 25.202 20.102
tempe kedelai mentah 23.453 21.208 24.301 5.005 5.004 5.005 25.259 23.030 26.126
tempe kedelai kukus 23.039 23.157 21.918 5.003 5.007 5.007 24.818 24.915 23.680
kacang koro 21.012 22.981 23.132 2.000 2.001 2.000 22.663 24.621 24.783
kacang koro kering 21.557 22.955 22.832 2.005 2.004 2.004 23.352 24.718 24.617
tempe koro mentah 23.318 23.031 23.494 5.069 5.067 5.070 24.972 24.723 25.163
tempe koro kukus 22.856 25.593 24.288 5.005 5.005 5.007 24.399 27.130 25.823
Sampel
Kadar Air (% bb)
Rataan SD
Ulangan 1 Ulangan 2 Ulangan 3
kacang kedelai 12.0014 12.1270 12.7849 12.3044 0.4208
tempe kedelai mentah 63.9270 63.5951 63.5374 63.6865 0.2102
tempe kedelai kukus 64.4520 64.8933 64.8099 64.7184 0.2344
kacang koro 17.4674 17.7253 17.4374 17.5433 0.1583
tempe koro mentah 67.3736 66.6075 67.0809 67.0206 0.3866
tempe koro kukus 69.1802 69.2972 69.3445 69.2739 0.0846
Perhitungan:
adarair bb C x 00
Keterangan :
Lampiran 3 Hasil pengukuran kadar abu kedelai dan koro pedang
Sampel Bobot Cawan Kosong Bobot Sampel
Bobot Cawan +Sampel Kering
U 1 U 2 U 3 U 1 U 2 U 3 U 1 U 2 U 3
kacang kedelai 23.029 23.444 18.357 2.001 2.001 2.001 23.122 23.543 18.452
tempe kedelai mentah 23.453 21.208 24.301 5.005 5.004 5.005 23.492 21.245 24.338
tempe kedelai kukus 23.040 23.157 21.918 5.003 5.007 5.007 23.077 23.190 21.959
kacang koro 21.012 22.981 23.132 2.000 2.001 2.000 21.061 23.029 23.179
kacang koro kering 21.557 22.955 22.833 2.005 2.004 2.004 21.610 23.004 22.885
tempe koro mentah 23.318 23.031 23.494 5.069 5.067 5.070 23.324 23.039 23.510
tempe koro kukus 22.856 25.593 24.288 5.005 5.005 5.007 22.869 25.612 24.303
Sampel
Kadar Abu (% bk)
Rataan SD
Ulangan 1 Ulangan 2 Ulangan 3
kacang kedelai 4.8972 5.1566 4.9848 5.0129 0.1320
tempe kedelai mentah 0.7792 0.7530 0.7285 0.7536 0.0254
tempe kedelai kukus 0.7572 0.6634 0.8094 0.7433 0.0740
kacang koro 2.5217 2.4524 2.4484 2.4741 0.0412
tempe koro mentah 0.1086 0.1403 0.3047 0.1845 0.1052
tempe koro kukus 0.2524 0.3730 0.3106 0.3120 0.0603
Perhitungan:
adar abu bb C x 00
adar abu bk 00kadar abu ( bb)kadar air ( bb)x 00
Keterangan :
Lampiran 4 Hasil pengukuran kadar protein kedelai dan koro pedang
Sampel Bobot sampel Volume HCl %N
U 1 U 2 U 3 U 1 U 2 U 3 U 1 U 2 U 3
kacang kedelai 0.502 0.500 0.503 18.350 18.420 18.500 5.162 5.197 5.195
tempe kedelai mentah 0.504 0.504 0.504 11.850 11.950 12.300 3.318 3.345 3.448
tempe kedelai kukus 0.504 0.504 0.504 11.300 11.220 11.150 3.164 3.141 3.126
kacang koro 0.502 0.500 0.503 15.800 15.750 16.100 4.445 4.444 4.521
kacang koro kering 0.506 0.505 0.506 14.750 14.850 14.800 4.118 4.150 4.129
tempe koro mentah 0.502 0.501 0.502 6.900 6.900 6.950 1.942 1.943 1.954
tempe koro kukus 0.502 0.503 0.503 6.100 6.200 6.150 1.714 1.741 1.725
Sampel
Kadar Protein (% bk)
Rataan SD
Ulangan 1 Ulangan 2 Ulangan 3
kacang kedelai 33.7300 34.0087 34.2498 33.9962 0.2601
tempe kedelai mentah 52.8940 52.8332 54.3807 53.3693 0.8765
tempe kedelai kukus 51.1840 51.4400 51.0789 51.2343 0.1857
kacang koro 33.6589 33.7584 34.2242 33.8805 0.3018
tempe koro mentah 37.1982 36.3665 37.0976 36.8874 0.4539
tempe koro kukus 34.7645 35.4408 35.1673 35.1242 0.3402
Perhitungan:
adar protein bb ol. Cl ol. blanko . Cl 4.007 00 obot sampel (mg)
Keterangan :
FK = Faktor koreksi,
Lampiran 5 Hasil pengukuran kadar lemak kedelai dan koro pedang
Sampel Bobot Cawan Kosong Bobot Sampel
Bobot Cawan +Sampel Kering
U 1 U 2 U 3 U 1 U 2 U 3 U 1 U 2 U 3
kacang kedelai 32.610 32.659 25.987 2.001 2.002 2.002 32.922 32.988 26.279
tempe kedelai mentah 31.770 31.778 32.865 2.001 2.005 2.002 31.810 31.815 32.902
tempe kedelai kukus 32.510 32.758 32.865 2.001 2.002 2.001 32.542 32.801 32.901
kacang koro 32.610 32.659 25.987 2.001 2.002 2.002 32.668 32.714 26.039
kacang koro kering 32.510 32.758 32.865 2.002 2.002 2.003 32.568 32.804 32.912
tempe koro mentah 32.339 30.138 32.758 2.003 2.004 2.001 32.363 30.152 32.771
tempe koro kukus 32.610 32.339 30.138 2.005 2.004 2.005 32.627 32.352 30.151
Sampel
Kadar Lemak (% bk)
Rataan SD
Ulangan 1 Ulangan 2 Ulangan 3
kacang kedelai 17.7357 18.6939 16.7423 17.7240 0.9758
tempe kedelai mentah 5.4723 5.1376 5.1239 5.2446 0.1973
tempe kedelai kukus 4.5549 6.1032 5.1404 5.2662 0.7818
kacang koro 3.5302 3.3327 3.1523 3.3384 0.1890
tempe koro mentah 3.5812 2.0622 2.0495 2.5643 0.8807
tempe koro kukus 2.8003 1.9829 2.0658 2.2830 0.4499
Perhitungan:
adar lemak bb C 00
Keterangan:
A = bobot labu lemak kosong, B = bobot sampel,
Lampiran 6 Hasil pengukuran kadar karbohidrat (by difference) kedelai dan koro pedang
Sampel
Kadar Karbohidrat (% bb)
Rataan SD
Ulangan 1 Ulangan 2 Ulangan 3
kacang kedelai 43.2289 41.7168 43.5637 42.8365 0.9840
tempe kedelai mentah 39.4904 39.9762 38.5119 39.3262 0.7459
tempe kedelai kukus 42.1469 40.5796 41.4991 41.4085 0.7876
kacang koro 59.8307 59.9995 59.7289 59.8530 0.1367
tempe koro mentah 65.1006 65.1452 65.3914 65.2124 0.1566
tempe koro kukus 58.8881 61.1517 59.9302 59.9900 1.1330
Perhitungan:
Lampiran 7 Hasil pengukuran derajat hidrolisis
Tempe Kedelai Mentah 0.583 0.560 0.535 1367.75 1310.25 1247.75
Tempe Kedelai Kukus 0.757 0.775 0.767 1802.75 1847.75 1827.75
Kacang Koro 0.726 0.720 0.718 1725.25 1710.25 1705.25
Tempe Koro Mentah 0.512 0.524 0.541 1190.25 1220.25 1262.75
Tempe Koro Kukus 0.720 0.721 0.727 1710.25 1712.75 1727.75
Berdasarkan persamaan yang didapat dari kurva standar yaitu:
y = 0.0004x + 0.0359. maka [Leusin] didapat dengan memasukan absorbansi sampel (A)
Sampel Derajat Hidrolisis (%) Derajat Hidrolisis
(%) Ulangan 1 Ulangan 2 Ulangan 3
Kacang Kedelai 18.9006 18.8766 18.8048 18.8607 ±0.0451
Tempe Kedelai Mentah 8.2009 7.8561 7.4814 7.8461 ±0.3311
Tempe Kedelai Kukus 11.2606 11.5417 11.4167 11.4063± 0.1296
Kacang Koro Pedang 16.2952 16.1535 16.1063 16.1850 ±0.0983
Tempe Koro Pedang Mentah 10.3275 10.5879 10.9566 10.6240 ±0.3161
Tempe Koro Pedang Kukus 15.5831 15.6059 15.7426 15.6439 ±0.0862
27
Lampiran 8 Hasil pengukuran konsentrasi standar BSA
Konsentrasi BSA (mg/ml) Absorbansi
0.00 0.000
0.20 0.215
0.40 0.340
0.60 0.485
0.80 0.590
1.00 0.690
1.20 0.810
Kurva standar BSA
0.000 0.100 0.200 0.300 0.400 0.500 0.600 0.700 0.800 0.900
0.00 0.20 0.40 0.60 0.80 1.00 1.20 1.40
Abs
o
rba
ns
i
Konsentrasi BSA (mg/ml) y = 0,6482x + 0,0582
29
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 26 Juni 1991 sebagai anak kedua dari dua bersaudara dari Bapak Mansyur Sarip dan Ibu Iriani.
Penulis menyelesaikan sekolah dasar di SDN UKS 17 Pagi Jakartapada tahun 2003 dan melanjutkan sekolah menengah pertama di SMPN 7 Jakarta sejak tahun 2003-2006. Sekolah menengah atas penulis selesaikan pada tahun 2009 di SMAN 31 Jakarta. Pada tahun yang sama penulis diterima di Institut Pertanian Bogor pada Program Studi Biokimia.
Pengalaman organisasi penulis yakni menjadi ketua seksi pembinaan demokrasi. hak asasi manusia. dan pendidikan politik di SMPN 7 Jakarta massa jabatan 2005-2006. Penulis juga pernah menjadi divisi HUMAS OSIS di SMAN 31 Jakarta massa jabatan 2006-2007. Selama kuliah penulis menjadi anggota divisi Communication and Information Center (CIC) perhimpunan mahasiswa CREBS tahun 2010-2011. Pengalaman kepanitian yang pernah penulis ikuti. diantaranya: Kepanitian Seminar Kesehatan 2011 sebagai Humas. Kepanitian Masa Perkenalan Departemen Biokimia tahun 2011 sebagai Humas. Kepanitian Biokimia Fair (BIK FAIR) tahun 2011 sebagai ketua pelaksana. Kepanitiaan Pesta Sains 2011 sebagai Lead Officer (LO). Kepanitian Seminar Kesehatan 2012 sebagai anggota tim acara. Kepanitiaan Masa Perkenalan Departemen Biokimia 2012. Kepanitiaan Pesta Sains 2012 sebagai anggota tim acara. dan lain sebagainya.