KARAKTERISASI PROTEASE DARI ISOLAT BAKTERI ASAL TUMBUHAN RAWA DARI INDRALAYA
Teks penuh
Gambar

Dokumen terkait
Hasil pengujian aktivitas enzim protease dari sembilan isolat bakteri, ditemukan tiga isolat bakteri yang menghasilkan enzim protease yaitu nomor 2,6, dan 7.
Tujuan penelitian ini adalah memproduksi enzim alginat liase dari isolat bakteri Bacillus megaterium asal Indonesia dan mengetahui optimasi suhu, pH, dan
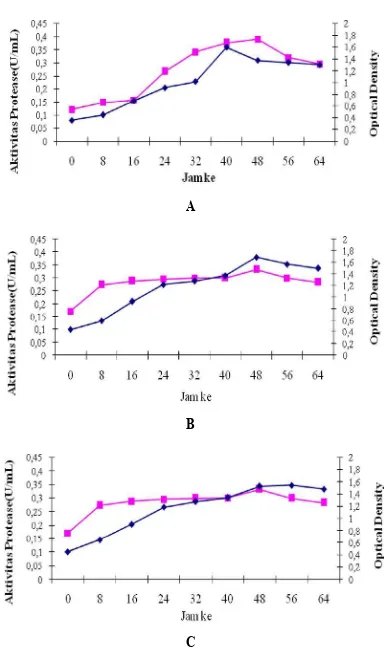
Dari hasil percobaan diketahui bahwa enzim protease dihasilkan pada waktu produksi optimum 18 jam dengan aktivitas protease sebesar 0,09 U/mL.. Suhu optimum enzim ini yaitu pada
isolat BB200109, dan mengkarakterisasi aktivitas kitin- ase pada berbagai kondisi pH, suhu, stabilitas waktu inkubasi, ion logam, kinetika enzim, dan menentukan bobot
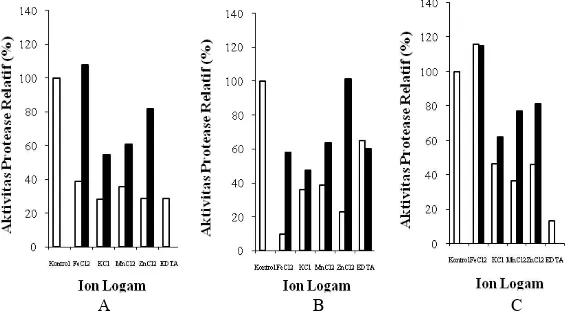
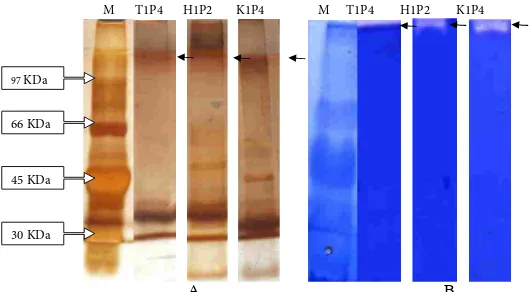
Gambar 4 Pengaruh ion logam pada protease dari bakteri tumbuhan rawa.. Hasil analisis zimogram menunjukkan protease asal isolat T1P4 memiliki berat molekul 151 kD, asal
Indeks uji proteolitik bakteri tumbuhan rawa Isolat Bakteri Indeks Proteolitik AR1TS1 1,31 AR2TS1 1,28 AR3TS1 1,30 AR1TS2 1,30 AR2TS2 1,08 Berdasarkan Tabel 2 menunjukkan
Hasil penelitian menunjukkan bahwa enzim amilase isolat bakteri R2M larva kumbang sagu yang direaksikan dengan substrat amilum optimum pada suhu 70°C, pH 4, dan konsentrasi substrat
Produksi Optimum Enzim Amilase dari Isolat Bakteri Terpilih Kurva pertumbuhan sel dan aktivitas amilase optimum diukur menggunakan isolat PKL2 karena memiliki nilai indeks amilolitik
