PENYERBUKAN SILANG ALAMI BEBERAPA
GENOTIPE CABAI (Capsicum annuum L.) DAN PENENTUAN
METODE PEMULIAANNYA
ARYA WIDURA RITONGA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Studi Penyerbukan Silang Alami Beberapa Genotipe Cabai (Capsicum annuum L.) dan Penentuan Metode Pemuliaannya adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, 6 Mei 2013
RINGKASAN
ARYA WIDURA RITONGA. Studi Penyerbukan Silang Alami Beberapa Genotipe Cabai (Capsicum annuum L.) dan Penentuan Metode Pemuliaannya. Dibawah bimbingan MUHAMAD SYUKUR, RAHMI YUNIANTI dan SOBIR.
Cabai dikenal sebagai tanaman menyerbuk sendiri. Tanaman menyerbuk sendiri memiliki ciri-ciri persentase penyerbukan silang alaminya dibawah 4 - 5%, umumnya diarahkan menjadi varietas galur murni, memiliki nilai heterosis dan
inbreeding depression yang rendah. Akan tetapi, varietas cabai di Indonesia saat ini didominasi oleh varietas hibrida bukan varietas galur murni. Hal ini terjadi karena diduga terdapat genotipe-genotipe cabai yang berperilaku menyerupai tanaman menyerbuk silang, seperti memiliki persentase penyerbukan silang alami dan nilai heterosis yang tinggi. Informasi sistem penyerbukan silang yang belum konsiten tersebut menyebabkan studi penyerbukan silang alami pada tanaman cabai menjadi penting untuk dilakukan, sehingga dapat digunakan dalam penentuan metode dan arah pemuliaan tanaman cabai. Penelitian ini bertujuan untuk (1) memperoleh marka evaluasi penyerbukan silang alami yang efektif dan efisien pada tanaman cabai, (2) membandingkan tingkat penyerbukan silang alami beberapa genotipe cabai, (3) membandingkan nilai heterosis, inbreeding depression, dan respon seleksi dua populasi cabai.
Penelitian ini terdiri atas 3 percobaan, yaitu studi pola pewarisan sifat karakter kualitatif dan kuantitatif pada hipokotil dan kotiledon cabai, studi penyerbukan silang alami pada beberapa genotipe cabai dan pendugaan parameter genetik pada dua populasi cabai. Hasil percobaan pertama menunjukkan bahwa karakter warna hipokotil dan warna warna kotiledon merupakan karakter yang efektif dan efisien untuk mengevaluasi kejadian penyerbukan silang alami pada tanaman cabai. Hal ini karena kedua karakter tersebut sama-sama dikendalikan oleh 1 gen dan tidak dipengaruhi lingkungan serta dapat diamati dalam waktu singkat dan tidak membutuhkan biaya yang mahal.
Hasil percobaan kedua menunjukkan bahwa genotipe cabai IPB C2 memiliki tingkat penyerbukan silang alami yang lebih tinggi dibandingkan genotipe cabai IPB C120 dan IPB C5. Perbedaan ini sangat dipengaruhi oleh tingkat crossability dan tipe posisi stigma yang dimiliki oleh genotipe-genotipe tersebut. Semakin tinggi tingkat crossability suatu genotype cabai yang disertai dengan posisi stigma yang lebih tinggi dari pada anther maka semakin tinggi pula tingkat penyerbukan silang alaminya.
SUMMARY
ARYA WIDURA RITONGA. Natural Cross-pollination in Chili Peppers (Capsicum annuum L.) and Its Breeding Methods Determination. Supervised by
MUHAMAD SYUKUR, RAHMI YUNIANTI and SOBIR.
Chili was known as pollinated crops. Some characteristics of self-pollinated crops are having a natural cross-pollination percentage below 4-5%, generally directed to be pure line varieties, and have a low heterosis and inbreeding depression. However, the chili pepper varieties in Indonesia currently are dominated by hybrid varieties and not pure line varieties. That is happens because there were chili pepper genotypes that behaves like a cross-pollinated, as have the high heterosis and the high percentages of natural cross-pollination. Natural cross-pollination information was not consistent led to studies of natural cross-pollination in chili pepper is important to do, so it can be used to determining the methods and the direction in chili pepper program. The aims of this study were to obtain the markers of natural cross-pollination in chili pepper, to compared the level of natural cross-pollination in some genotypes of chili pepper and to compared the heterosis, inbreeding depression, and response selection two population of chili pepper.
This study was consisted of three experiments. First, estimation of several genetic parameters on qualitative and quantitative characters on hypocotyl and cotyledon of chili peppers (Capsicum annuum L.). Second, study of natural cross-pollination in some genotypes of chili pepper. Third, estimation of several genetic parameters in two population of chili peppers that have different level of natural cross-pollination. The results of the first experiment showed that the color of hypocotyl and cotyledon characters were effective and efficient markers to evaluate the incidence of natural cross-pollination in chili pepper. These were because both of characters controlled by single gene and not influenced by environment. In addition, its can be observed in short time and does not require an expensive cost.
The result of the second experiment showed that the IPB C2 genotype has higher natural cross-pollination level than the IPB C5 and IPB C120 genotypes. This was highly influenced by level of crossability and stigma position each of genotypes. The higher level of crossability with stigma position higher than anther causes the higher level of natural cross-pollination in chili pepper.
The third experiment result showed that the IPB C2 x IPB C5 population has a higher heterosis and inbreeding depression than IPB C120 x IPB C5 population. This was highly influenced by the parents were used at the formation of the population. The population was formed by the parents with the high level of natural cross pollination has a higher heterosis and inbreeding depression than the population was formed by the parents with low level of natural cross-pollination.
© Hak Cipta Milik IPB, Tahun 2011
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
PENYERBUKAN SILANG ALAMI BEBERAPA
GENOTIPE CABAI (Capsicum annuum L.) DAN PENENTUAN
METODE PEMULIAANNYA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2013
NIM : A253100041
Disetujui oleh Komisi Pembimbing
Prof. Dr. Ir. Sobir, MSi.
Ketua Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasmjana
Pemuliaan dan
Bioteknologi Tanaman
Dr. Ir. Trikoesoemarrtfigtyas, MSc.
Tanggal Ujian: Tanggal Lulus:
2 2 JUL
2013
Judul Tesis : Penyerbukan Silang Alami Beberapa Genotipe Cabai (Capsicum annuum L.) dan Penentuan Metode Pemuliannya.
Nama : Arya Widura Ritonga
NIM : A253100041
Disetujui oleh Komisi Pembimbing
Dr. Muhamad Syukur, SP., MSi Ketua
Prof. Dr. Ir. Sobir, MSi. Anggota
Diketahui oleh
Ketua Program Studi Pemuliaan dan
Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, MSc.
Dekan Sekolah Pascasarjana
Dr. Ir. Dahrul Syah, MScAgr.
Tanggal Ujian: 27 Mei 2013
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala rahmat dan karuni-Nya, sehingga karya ilmiah ini berhasil diselesaikan. Tesis yang berjudul Penyerbukan Silang Alami Beberapa Genotipe Cabai (Capsicum annuum
L.) dan Penentuan Metode Pemuliaannya ini merupakan tugas akhir penulis untuk memperoleh gelar Magister Sains dari Program Studi Pemuliaan dan Bioteknologi Tanaman, Sekolah Pascasarjana Institut Pertanian Bogor.
Terima kasih yang sebesar-besarnya penulis sampaikan kepada:
1. Allah SWT atas kemudahan, kekuatan, karunia, dan rahmat-Nya sehingga penulis dapat menyelesaikan tugas akhir ini
2. Dr. Muhamad Syukur, SP. MSi, Dr. Rahmi Yunianti, SP. MSi, dan Prof. Dr. Ir. Sobir, MS. selaku dosen pembimbing atas segala arahan, saran, masukan, kesabaran, dan motivasi yang telah diberikan selama ini,
3. Prof. Dr. Ir. Surjono H. Sutjahtjo dan Dr. Ir. Eny Widajati, MS selaku dosen penguji yang telah memberikan arahan dan masukan sehingga tesis ini menjadi lebih baik
4. Kedua orang tua (Bapak Kalpen Ritonga dan Ibu Sri Nuryanti) yang selalu berdoa dan memberikan dukungan tiada henti
5. Yusnita Sari yang selalu setia membantu dan menjadi pendengar keluh kesah penulis
6. Abdul Hakim, Mas Undang, Muhamad Alfarabi, Helfi Saputra, Tiara Yudilastari, Mba Vitria Rahardi, Mba Sri Wahyuni, Mba Siti Marwiyah, dan Mang Darwa atas segala bantuannya selama ini.
7. Rekan-rekan Program Studi Pemuliaan Tanaman, Sekolah Pascasarjana tahun 2010 atas dukungan dan kerjasamanya selama ini.
8. DIKTI atas beasiswa pendidikan yang telah diberikan melalui program I-MHERE B2c
9. Semua pihak yang telah membantu atas terselesaikannya penelitian ini
Mohon maaf atas segala kekurangan dan kesalahan dalam penulisan karya ilmiah ini. Semoga karya ilmiah ini bermanfaat bagi pengembangan ilmu pengetahuan, khususnya bidang pemuliaan tanaman dan pertanian pada umumnya.
Bogor, 6 Mei 2013
DAFTAR ISI xi
DAFTAR TABEL xii
DAFTAR GAMBAR xiii
DAFTAR LAMPIRAN xiii
DAFTAR LAMPIRAN PENDAHULUAN
Latar Belakang 1
Tujuan 1
Ruang Lingkup 2
TINJAUAN PUSTAKA 4
PEWARISAN SIFAT KARAKTER KUALITATIF DAN
KUANTITATIF PADA HIPOKOTIL DAN KOTILEDON CABAI (Capsicum annuum L.)
Pendahuluan 11
Bahan dan Metode 12
Hasil dan Pembahasan 15
Simpulan 23
STUDI PENYERBUKAN SILANG ALAMI DAN BUATAN BEBERAPA GENOTIPE CABAI (Capsicum annuum L.) DAN PENENTUAN METODE PEMULIAANNYA
Pendahuluan 24
Bahan dan Metode 25
Hasil dan Pembahasan 30
Simpulan 37
PENDUGAAN PARAMETER GENETIK DUA POPULASI CABAI (Capsicum annuum L.)PADA BEBERAPA GENERASI
Pendahuluan 39
Bahan dan Metode 40
Hasil dan Pembahasan 42
Simpulan 53
PEMBAHASAN UMUM 57
SIMPULAN UMUM 59
SARAN 59
DAFTAR PUSTAKA 58
LAMPIRAN 64
1. Nisbah fenotipe karakter kualitatif yang terkait adaptasi terhadap
cekaman yang dikendalikan oleh gen mayor pada populasi F2 9 2. Nilai X2hitung warna hipokotil dan warna kotiledon pada populasi
BCP1, BCP2 dan F2 16
3. Uji pengaruh tetua betina populasi F1 dan F1R pada beberapa karakter
cabai 18
4. Jumlah gen pengendali dan aksi gen pada beberapa karakter cabai 20 5. Uji kecocokan model genetik beberapa karakter cabai 20 6. Pendugaan komponen genetik beberapa karakter cabai 21 7. Komponen ragam dan heritabilitas beberapa karakter cabai 27 8. Warna mahkota, warna anther, warna putik, warna stigma, dan tipe
posisi stigma beberapa genotipe cabai 32
9. Rekapitulasi hasil analisis uji-z karakter umur berbunga, panjang tangkai hari, panjang anther, panjang putik, tinggi tanaman dan posisi
stigma beberapa genotipe cabai 34
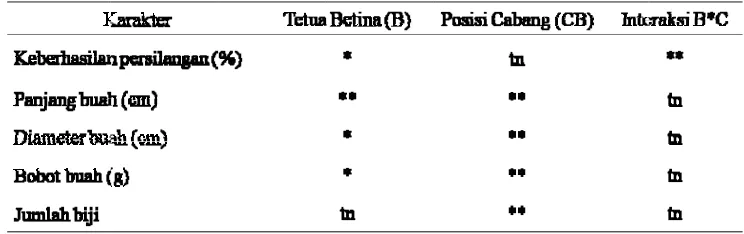
10.Rekapitulasi sidik ragam pengaruh genotipe, posisi cabang, dan interaksi genotipe x posisi cabang terhadap beberapa karakter cabai 35 11.Pengaruh genotipe terhadap jumlah biji, panjang, diameter, dan bobot
buah cabai hasil persilangan buatan 37
12.Pengaruh posisi cabang terhadap jumlah biji, panjang, diameter, dan
bobot buah hasil persilangan 37
13.Nilai heterosis, heterobeltiosis, dan inbreeding depression dua
populasi cabai 43
14.Ragam genetik beberapa karakter tanaman cabai pada dua populasi
cabai 49
15.Ragam genetik beberapa karakter tanaman cabai populasi cabai IPB
C2 x IPB C5 pada berbagai generasi 50
16.Heritabilitas arti luas beberapa karakter tanaman cabai populasi cabai
IPB C120 x IPB C5 pada berbagai generasi 51
17.Heritabilitas arti luas beberapa karakter tanaman cabai populasi cabai
1. Diagram alir penelitian penyerbukan silang alami beberapa genotipe cabai (Capsicum annuum L.) dan penentuan metode pemuliaannya 3 2. Warna hipokotil dan kotiledon genotipe cabai IPB C20, F1 IPB C20 x
IPB C2, dan IPB C2 15
3. Nilai tengah dan simpangan baku populasi P1, P2, F1, F2, BCP1 dan
BCP 2 17
4. Sebaran populasi F2 (IPB C2 x IPB C20) 18
5. Sebaran populasi F2 (IPB C2 x IPB C20) 19
6. Layout percobaan penyerbukan silang alami beberapa genotipe cabai
(Capsicum annuum L.) 26
7. Pola penyerbukan silang alami beberapa populasi cabai pada berbagai
minggu panen 30
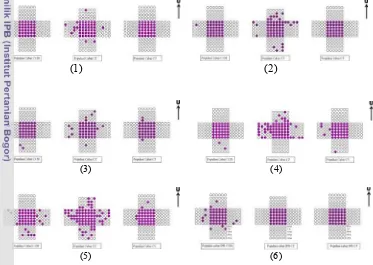
8. Pola penyerbukan silang alami beberapa genotipe cabai di lapang
setelah 6 minggu 31
9. Persentase tanaman yang mengalami penyerbukan silang alami 32 10.Tipe – tipe posisi stigma beberapa genotipe cabai 33 11.Persentase keberhasilan persilangan buatan pada beberapa genotipe
cabai 35
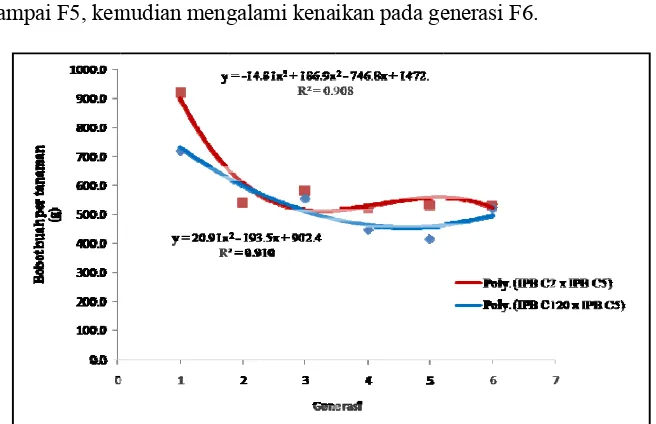
12.Pengaruh interaksi genotipe dan posisi cabang terhadap persentase keberhasilan persilangan buatan beberapa genotipe cabai 36 13.Bobot per tanaman dua populasi cabai pada enam generasi 44 14.Jumlah buah per tanaman dua populasi cabai pada enam generasi 45 15.Nilai tengah beberapa karakter buah dua populasi cabai pada enam
generasi 46 16.Nilai tengah beberapa karakter vegetatif dua populasi cabai enam
generasi 48
DAFTAR LAMPIRAN
1. Tabel nilai tengah beberapa karakter populasi cabai IPB C120 x IPB
C5 pada enam generasi 64
2. Tabel nilai tengah beberapa karakter populasi cabai IPB C2 x IPB C5
PENDAHULUAN
Latar Belakang
Cabai (Capsicum annuum L.) merupakan salah satu spesies tanaman yang memiliki nilai ekonomi penting di dunia (Bosland 1996). Tanaman ini juga merupakan salah satu tanaman sayuran penting di Indonesia. Hal ini dapat dilihat dari luas pertanaman cabai di Indonesia yang mencapai 237 ribu ha pada tahun 2009 (Kementerian Pertanian RI 2011) yang merupakan luasan terbesar pada komoditi sayuran. Namun, sering kali jumlah produksi cabai tidak mampu mencukupi kebutuhan permintaan pasar, sehingga menyebabkan tingginya harga cabai di pasaran. Dilaporkan bahwa harga cabai merah dan keriting mencapai lebih Rp 50.000 per kg pada bulan Desember 2010 (Kementerian Pertanian RI 2010). Salah satu penyebab rendahnya produksi cabai adalah penggunaan benih unggul cabai yang masih rendah sehingga menyebabkan pertanaman cabai lebih mudah teserang hama penyakit dan produktivitasnya tidak optimal.
Cabai dikenal sebagai tanaman menyerbuk sendiri. Tanaman menyerbuk sendiri adalah tanaman - tanaman yang persentase penyerbukan silang alaminya dibawah 4 - 5% (Sleper dan Phoelman 2006). Tanaman menyerbuk sendiri umumnya diarahkan untuk menjadi varietas galur murni. Hal ini karena nilai heterosis yang rendah dan tidak adanya inbreeding depression pada tanaman menyerbuk sendiri. Namun varietas – varietas cabai yang dihasilkan di Indonesia, saat ini didominasi oleh varietas hibrida bukan varietas galur murni. Sebanyak lebih dari 80% varietas cabai yang sudah dilepas di Indonesia merupakan varietas cabai hibrida (Direktorat Perbenihan dan Sarana Produksi 2011). Hal ini terjadi karena diduga terdapat genotipe-genotipe cabai yang perilakunya menyerupai tanaman menyerbuk silang, seperti memiliki persentase penyerbukan silang alami dan nilai heterosis yang tinggi. Campodonico (1983), Corella et al. (1986), dan Csillery (1986) melaporkan bahwa penyerbukan silang alami pada cabai dapat mencapai lebih dari 50%, sedangkan Sujiprihati et al. (2007) dan Mantri (2007) melaporkan bahwa nilai heterosis pada persilangan cabai dapat mencapai lebih dari 60%. Studi penyerbukan silang alami pada tanaman cabai penting untuk dilakukan. Informasi ini berguna dalam penentuan metode dan arah pemuliaan tanaman cabai.
Tujuan
Tujuan yang ingin dicapai dari penelitian ini adalah:
1. Memperoleh marka yang efektif dan efisien untuk mengevaluasi penyerbukan silang alami pada tanaman cabai.
2. Membandingkan tingkat penyerbukan silang alami beberapa genotipe cabai. 3. Membandingkan nilai heterosis, inbreeding depression, dan respon seleksi dua
Ruang Lingkup Penelitian
Penelitian ini terdiri atas 3 percobaan. Percobaan pertama, studi pola pewarisan sifat karakter kualitatif dan kuantitatif pada hipokotil dan kotiledon cabai yang bertujuan untuk mendapatkan karakter pada hipokotil atau kotiledon yang dapat dijadikan marka dalam mengevaluasi kejadian penyerbukan silang alami tanaman cabai. Kemudian dilanjutkan dengan percobaan kedua, studi penyerbukan silang alami pada beberapa genotipe cabai yang bertujuan untuk memperoleh informasi tentang pengaruh genotipe terhadap kejadian penyerbukan silang alami pada tanaman cabai, sehingga diketahui genotipe-genotipe cabai yang memiliki persentase penyerbukan silang alami rendah dan tinggi. Setelah itu, dibentuk dua populasi cabai yang terdiri dari populasi cabai dengan penyerbukan silang alami rendah dan tinggi untuk digunakan pada percobaan ketiga yang berjudul pendugaan parameter genetik pada dua populasi cabai selama enam generasi. Percobaan ini bertujuan untuk memperoleh informasi tentang heterosis,
G
Gambar 1 c
Diagram ali cabai (Capsi
ir penelitian
icum annuu
penyerbuka
um L.) dan
an silang ala penentuan
ami beberap metode pem
TINJAUAN PUSTAKA
Botani dan Morfologi Cabai
Cabai termasuk dalam tanaman kingdom Plantae, divisi Magnoliophyta, kelas Magnoliopsida, Ordo Solanales, famili Solanaceae, genus Capsicum, dan spesies Capsicum annuum L. Terdapat setidaknya dua puluh lima spesies liar dan lima spesies Capsicum yang telah dibudidayakan secara luas, yaitu Capsicum annuum (cabai besar), Capsicum frutescens (cabai kecil), Capsicum chinense, Capsicum baccatum, dan Capsicum pubescens (Bosland dan Votava 1999). Perbedaan diantara spesies cabai dapat diketahui pada ketidakmampuan terjadinya persilangan diantaranya dan perbedaan morfologi (warna, bentuk bunga dan biji) pada tanaman – tanaman yang mampu bersilang dengan keberhasilan yang rendah (Andrew 1995). Capsicum merupakan tanaman diploid yang sebagian besar memiliki jumlah kromosom 2n = 2x = 24, namun pada beberapa spesies liar memiliki jumlah kromosom 2n = 2x = 26 (Bosland dan Votava 1999).
Cabai pada umumnya merupakan tanaman semusim di wilayah temperate, namun dapat menjadi tanaman tahunan pada wilayah tropis dan dalam rumah kaca (OECD 2006). Cabai merupakan tanaman terna atau setengah perdu dengan tinggi tanaman 0.5 – 1.5 m. Tanaman cabai memiliki batang yang tegak dan berkayu pada bagian pangkalnya, sedangkan pada bagian atas terdapat banyak percabangan (Rubatzky dan Yamaguchi 1999).
Tanaman cabai memiliki perakaran yang kuat. Akar tanaman cabai merupakan akar tunggang yang tumbuh lurus ke bawah secara vertikal. Akar cabang akan tumbuh dari samping akar tunggang, kemudian akar rambut akan tumbuh pada sekitar akar cabang tersebut. Akar tanaman cabai bisa tumbuh mencapai kedalaman satu meter (Rubatzky dan Yamaguchi 1999).
Daun tanaman cabai merupakan daun tunggal dengan bentuk yang bervariasi, mulai dari lancip sampai dengan bulat telur dengan ujung daun yang lancip dan tepinya yang rata. Warna daun cabai bervariasi, dari mulai hijau, hijau tua, sampai hijau keunguan. Daun tumbuh pada tunas samping secara berurutan, sedangkan pada cabang utama secara spiral (Kusandriani 1996).
Bentuk dan warna buah cabai bervariasi. Falusi dan Morakinyo (1994) menjelaskan bahwa terdapat berbagai variasi bentuk buah pada Capsicum annuum
L. Bentuk buah cabai, mulai dari pendek, panjang, bulat, oval, sampai keriting. Warna buah cabai sangat bervariasi, yaitu: hijau, kuning, atau bahkan ungu ketika muda dan kemudian berubah menjadi merah, jingga, atau campuran bersamaan dengan meningkatnya umur buah (Rubatzky dan Yamaguchi 1999). Buah cabai merupakan buah tunggal yang tumbuh pada buku, namun juga terdapat buah yang lebih dari satu (fasiculate) tumbuh dalam satu buku (Kusandriani 1996). Arah tumbuh buah cabai juga bervariasi antar kultivar, ada yang ke bawah, menyamping, dan ke atas.
Pemuliaan Tanaman Cabai di Indonesia
Pemuliaan tanaman adalah suatu ilmu dan seni dalam merakit suatu tanaman untuk kepentingan manusia (Sleper dan Poehlman 2006). Program pemuliaan tanaman bertujuan untuk menghasilkan varietas unggul dengan sifat dan karakter tanaman yang lebih baik. Suatu program pemuliaan tanaman terdiri dari berbagai kegiatan yang saling berhubungan satu sama lainnya. Kegiatan – kegiatan tersebut terdiri dari pembentukan populasi genetik (hibridisasi, mutasi, variasi somaklonal, transformasi gen), seleksi (seleksi massa, seleksi pedigree, dan seleksi berulang), dan pengujian (uji daya hasil, uji BUSS, dan multilokasi). Namun demikian, metode pemuliaan suatu tanaman berbeda antara satu tanaman dengan tanaman lainnya. Sleper dan Poehlman (2006) membagi metode pemuliaan tanaman menjadi metode pemuliaan tanaman menyerbuk sendiri, pemuliaan tanaman menyerbuk silang, pemuliaan tanaman yang diperbanyak secara vegetatif, dan pemuliaan varietas hibrida.
Cabai dikenal sebagai tanaman menyerbuk sendiri di Indonesia. Hal ini karena tanaman cabai memiliki persentase penyerbukan sendiri yang tinggi. Tanaman menyerbuk sendiri umumnya adalah tanaman yang memiliki tingkat penyerbukan silang alami yang rendah, yaitu 4 – 5% (Sleper dan Poehlman 2006). Tingginya persentase penyerbukan sendiri pada tanaman cabai dapat terjadi karena cabai memiliki bunga hermaprodit yang self-compatible. Struktur bunga yang hermaprodit dan self – compatible sangat berperan dalam tingginya tingkat penyerbukan sendiri pada suatu tanaman (Damgaard et al. 1992). Oleh karena itu, metode pemuliaan tanaman cabai umumnya mengikuti metode pemuliaan tanaman pada tanaman menyerbuk sendiri, seperti pada tanaman padi dan kedelai.
Pemuliaan tanaman cabai diawali dengan pembentukan populasi genetik yang dilanjutkan dengan seleksi dan persilangan diantara tetua – tetua terpilih. Setelah itu, dilakukan seleksi selama beberapa generasi yang kemudian dilanjutkan dengan uji daya hasil, uji multilokasi, dan pelepasan varietas. Pemuliaan tanaman cabai, pada awalnya diarahkan untuk merakit varietas cabai berdaya hasil tinggi. Namun, pada saat ini pemuliaan tanaman cabai diarahkan untuk merakit varietas cabai yang selain berdaya hasil tinggi juga memiliki penampilan yang baik dan tahan terhadap serangan hama dan penyakit.
pada tanaman menyerbuk silang (Wehner 1997). Peningkatan hasil akibat heterosis pada tanaman padi hanya mencapai 23% (Lestari dan Nugraha, 2006), 20% (Sukirman 2005), dan 26% (Satoto et al. 2007). Sedangkan peningkatan hasil akibat heterosis pada tanaman jagung dapat mencapai lebih 100% (Suwarno 2000; Setiyono dan Subandi 1996) dan pada tanaman bunga matahari dapat mencapai 300% (Ahmad et al. 2005). Namun, varietas – varietas cabai yang dihasilkan di Indonesia saat ini didominasi oleh varietas hibrida bukan varietas galur murni. Sebanyak 80% varietas cabai yang dilepas di Indonesia merupakan cabai hibrida.
Salah satu penyebabnya diduga karena varietas – varietas tersebut dapat memiliki nilai heterosis yang tinggi. Nilai heterosis pada hasil persilangan dialel tanaman cabai dapat mencapai 63% dan nilai heterobeltiosisnya dapat mencapai 44 % (Sujiprihati et al. 2007), bahkan Mantri (2006) menyatakan bahwa nilai heterosis pada cabai dapat mencapai lebih dari 100%. Nilai heterosis yang tinggi pada cabai diduga karena tanaman cabai ternyata memiliki kemampuan untuk melakukan penyerbukan silang secara alami yang cukup tinggi.
Penyerbukan Silang pada Tanaman Cabai
Penyerbukan silang adalah peristiwa jatuhnya polen dari anther ke kepala putik tanaman yang berbeda. Penyerbukan silang dapat terjadi pada grup tanaman monoecious (jagung, kelapa sawit, melon, dan lain - lain) dan dioecious (salak, kurma, melinjo, vanili dan lain - lain). Penyerbukan silang juga dapat terjadi pada grup tanaman hermaprodit. Penyerbukan silang pada grup tanaman hermaprodit dapat terjadi karena adanya modifikasi bunga hermaprodit (kasmogami, dikogami, dan herkogami), self-incompability, dan male sterility.
Tingginya persentase penyerbukan sendiri alami pada tanaman cabai dapat terjadi karena tanaman cabai memiliki bunga hermaprodit yang self-compatible. Struktur bunga yang hermaprodit dan self – compatible sangat berperan dalam tingginya tingkat penyerbukan sendiri pada suatu tanaman (Damgaard et al.
1992). Tanaman menyerbuk sendiri umumnya adalah tanaman yang memiliki tingkat penyerbukan silang alami yang rendah, yaitu 4 – 5% (Sleper and Poehlman, 2006). Namun, beberapa penelitian melaporkan bahwa terdapat tingkat penyerbukan silang alami yang tinggi pada tanaman cabai. Tingkat penyerbukan silang alami pada tanaman cabai di Italia dan Spanyol dapat mencapai lebih dari 50% (Campodonico 1983; Corella et al. 1986; Csillery 1986).
Penyerbukan silang buatan pada cabai umumnya masih mudah dilakukan jika masih dalam satu spesies. Namun, jika penyerbukan silang dilakukan antar spesies cabai, maka umumnya persilangan akan mengalami hambatan. Hambatan yang sering terjadi diantaranya adalah sulit tejadi fertilisasi dan jika fertilisasi berhasil, maka tanaman tersebut akan steril. Emboden (1964) melaporkan bahwa persilangan buatan antara C. frutescens x C. baccatum dan C. Chinense x C. baccatum tidak dapat menghasilkan biji.
salah satu imbalan yang penting dari bunga bagi polinator (Robertson et al. 1999). Vektor penyerbukan silang alami yang utama pada tanaman cabai diduga adalah lebah. Raw (2000) melaporkan bahwa perilaku makan lebah berpotensi dalam mempengaruhi tingkat penyerbukan silang alami pada tanaman cabai. Cruz et al.
(2005) juga melaporkan bahwa lebah (Melipona subnitida) potensial dalam penyerbukan silang pada tanaman cabai di dalam green house. Penyerbukan silang alami pada tanaman cabai memiliki jangkauan radius yang luas. Kim et al. (2009) melaporkan bahwa penyerbukan silang alami pada tanaman cabai dapat mencapai jarak 18 m.
Antosianin
Antosianin merupakan salah satu pigmen yang berwarna merah muda sampai ungu. Antosianin merupakan bagian dari flavonoid yang berwarna ungu. Antosianin terletak di dalam vakuola sel (Lightbourn et al. 2008). Antosianin memiliki peranan yang sangat penting bagi tanaman. Antosianin berperan dalam melindungi tanaman dari radiasi sinar UV, sebagai zat anti-mikrobial, dan sebagai antioksidan (Harborne dan Williams 2000). Intensitas dan stabilitas pigmen antosianin dipengaruhi oleh berbagai faktor, yaitu struktur dan konsentrasi pigmen, PH, temperatur, intensitas cahaya, dan keberadaan pigmen lainnya (Laleh
et al. 2006). Peningkatan radiasi UV-B menyebabkan peningkatan intensitas antosianin pada daun tanaman cabai (Mahdavian et al. 2008). Antosianin pada tanaman kolesom dipengaruhi oleh pemupukan (Mualim et al. 2009)
Akumulasi antosianin pada tanaman cabai dapat dilihat pada bagian hipokotil, kotiledon, batang, bunga, dan buah yang belum matang (Wang dan Bosland 2006). Genotipe – genotipe cabai yang mengekspresikan antosianin memperlihatkan bahwa antosianin pada bagian buah cabai bersifat sementara yaitu hanya saat buah muda saja, sedangkan antosianin pada daun, batang, bunga, dan organ lainnya lebih stabil. Warna ungu akibat akumulasi antosianin pada batang, daun, bunga, dan buah muda cabai dikendalikan oleh gen dominan tidak lengkap anthocyanin (A) dan diperkuat oleh gen Modifier of A (MoA) (Lightbourn 2008). Warna ungu pada tanaman merupakan salah satu marka morfologi yang efektif dan efisien. Motten dan Antonovics (1992) telah menggunakan warna ungu pada bunga dan hipokotil untuk menentukan tingkat penyerbukan silang alami pada tanaman Datura stramoium. Warna ungu pada hipokotil dan kotiledon juga merupakan salah satu karakter yang di uji dalam uji BUSS tanaman cabai (UPOV 2006).
Komponen Ragam dan Heritabilitas
Berdasarkan komponen ragam genotipenya, nilai heritabilitas dapat dibedakan menjadi dua, yaitu heritabilitas arti luas (broad-sense heritability) dan heritabilitas arti sempit (narrow-sense heritability). Heritabilitas arti luas (h2bs)
merupakan perbandingan antara ragam genotipe total (Vg) yang merupakan
gabungan dari ragam aditif (Va), ragam dominan (Vd), dan ragam epistasis (Vi)
dengan ragam fenotipenya (Vp) (Sleper dan Poehlman 2006). Sedangkan
heritabilitas arti sempit adalah perbandingan hanya antara ragam aditif (Va)
dengan ragam fenotipenya (Vp). Berdasarkan hal tersebut, maka haritabilitas arti
sempit lebih sering diperhatikan dibandingkan heritabilitas arti luas. Hal ini dikarenakan hanya ragam aditif saja yang dapat diwariskan. Keberadaan ragam dominan dan ragam epistasis pada perhitungan heritabilitas arti luas dapat mengakibatkan bias ketika akan melakukan kegiatan pemuliaan tanaman.
Terdapat berbagai metode untuk menduga nilai heritabilitas. Nilai heritabilitas dapat diduga dengan perhitungan ragam turunan, regresi parent offspring, dan komponen ragam hasil analisis ragam. Metode yang digunakan disesuaikan dengan tujuan dan ketersediaan populasi yang dimiliki. Poespodarsono (1988) menyatakan bahwa nilai heritabilitas dinyatakan dalam bilangan desimal atau persentase, yang berkisar antara 0 – 1. Kriteria nilai heritabilitas arti luas, menurut Whirter (1979) dalam Mursito (2003) dapat dibagi menjadi tiga, yaitu heritabilitas rendah (h2bs < 20%), heritabilitas sedang (20% <
h2bs < 50%), dan heritabilitas tinggi (50% < h2bs). Nilai heritabilitas suatu sifat
akan bervariasi antar populasi. Karakter – karakter yang mudah dipengaruhi lingkungan umumnya memiliki nilai heritabilitas yang rendah, sedangkan karakter – karakter yang sulit dipengaruhi oleh lingkungan umumnya memiliki nilai heritabilitas yang tinggi (Sleper dan Poehlman 2006). Sujiprihati et al. (2003) menyatakan bahwa nilai heritabilitas sangat tergantung dari metode dan populasi yang digunakan.
Nilai heritabilitas yang rendah menunjukkan bahwa keragaman yang terjadi lebih dipengaruhi oleh faktor lingkungan dibandingkan faktor genetiknya, sedangkan nilai heritabilitas yang tinggi menunjukkan bahwa keragaman yang timbul lebih dipengaruhi oleh faktor genetik dibandingkan lingkungan (Sleper dan Poehlman 2006). Terhadap karakter – karakter yang memiliki nilai heritabilitas tinggi sudah dapat dilakukan kegiatan seleksi pada generasi awal, sedangkan terhadap karakter – karakter dengan nilai heritabilitas rendah sebaiknya dilakukan seleksi pada generasi lanjut agar gen-gen aditifnya sudah terfiksasi (Sleper dan Poehlman 2006).
Karakter Kualitatif dan Karakter Kuantitatif
kualitatif dapat langsung dilakukan secara visual baik dengan kontrol yang sudah distandarisasi maupun dengan skoring. Karakter kualitatif lebih cenderung mengikuti rasio mendel.
Tanaman pada generasi F2 akan mengalami segregasi sesuai dengan hukum mendel. Adanya aksi dan interaksi gen yang berbeda akan menghasilkan pola segregasi yang berbeda pula. Tipe aksi gen yang mengikuti sebaran mendel umumnya dibedakan menjadi dua, yaitu interaksi antar alel dalam lokus yang sama (intralokus) dan interaksi alel pada lokus yang berbeda (interlokus). Karakter kualitatif yang dikendalikan oleh dua alel dalam satu lokus akan menghasilkan segregasi 3 : 1 jika terdapat dominansi, namun jika tidak terdapat dominansi maka akan menghasilkan pola segregasi 1 : 2 : 1 pada F2. Pola segregasi akan lebih berbeda jika karakter kualitatif tersebut dikendalikan oleh interaksi interlokus. Tabel 1, memperlihatkan contoh nisbah fenotipe karakter kualitatif yang terkait adaptasi terhadap cekaman yang dikendalikan oleh gen mayor pada populasi F2 (Crowder1993).
Tabel 1 Nisbah fenotipe karakter kualitatif yang terkait adaptasi terhadap cekaman yang dikendalikan oleh gen mayor pada populasi F2 (Crowder1993) Jumlah gen dan tipe aksi gen berdasarkan nisbah Nisbah fenotipe
T AT AP P Satu (1) pasang gen:
A Dominan penuh 3 - - 1
B Resesif 1 - - 3
C Tidak ada dominansi 1 2 - 1
Dua (2) pasang gen:
A Tidak ada epistasis 9 3 3 1
B Epistasis resesif 9 3 - 4
C Epistasis dominan 12 - 3 1
D Epistasi dominan - resesif 13 - - 3
E Resesif ganda 9 - - 7
F Isoepistasis 15 - - 1
G Semi epistasis 9 6 - 1
H Interaksi kompleks 10 3 - 3
Keterangan: T = toleran AT = agak toleran AP = agak peka P = peka
Seleksi dan Respon Seleksi
Seleksi merupakan salah satu kegiatan penting dalam suatu program pemuliaan tanaman. Kegiatan seleksi bertujuan untuk meningkatkan frekuensi gen – gen yang diinginkan (Falconer dan Mackay 1996). Metode yang digunakan dalam kegiatan seleksi sebaiknya disesuaikan dengan tujuan dan jenis tanaman yang digunakan. Metode seleksi tanaman menyerbuk sendiri berbeda dengan metode seleksi pada tanaman menyerbuk silang. Pada tanaman menyerbuk sendiri dikenal seleksi untuk populasi heterozigot homozigot (seleksi massa) dan untuk populasi bersegregasi (seleksi pedigree, seleksi bulk, single seed descent, dan seleksi silang balik). Sedangkan pada tanaman menyerbuk silang, metode seleksi yang digunakan berupa seleksi massa, seleksi ear to raw, dan seleksi berulang (seleksi berulang fenotipe, seleksi berulang daya gabung umum, seleksi berulang untuk daya gabung khusus, dan seleksi berulang resiprokal) (Sleper dan Poehlman 2006).
Efektivitas kegiatan seleksi dapat dilihat dari nilai respon seleksinya. Respon seleksi adalah perubahan nilai rata – rata fenotipe dari generasi berikutnya sebagai akibat adanya kegiatan seleksi terhadap suatu populasi (Hamdan 2005). Nilai respon seleksi sangat tergantung pada nilai heritabilitas populasi tanaman contoh dan besar intensitas seleksi yang digunakan. Semakin tinggi nilai heritabilitas dan intensitas seleksi yang digunakan, menyebabkan nilai respon seleksi akan semakin tinggi. Nilai respon seleksi pada generasi awal umumnya lebih tinggi dibandingkan dengan nilai respon seleksi pada generasi lanjut. Hal ini terkait dengan tingkat keragaman genetik dan nilai heritabilitas yang masih tinggi pada generasi awal. Intensitas seleksi adalah jumlah indidu yang akan diseleksi dari suatu populasi. Pada populasi dengan tingkat keragaman yang tinggi sebaiknya digunakan intensitas seleksi yang tinggi pula. Namun, penggunan intensitas seleksi yang terlalu tinggi akan dapat menyebabkan penyempitan keragaman genetik pada turunannya yang nantinya dapat menyebabkan random genetic drift (Falconer dan Mackay 1996).
PEWARISAN SIFAT KARAKTER KUALITATIF DAN
KUANTITATIF PADA HIPOKOTIL DAN KOTILEDON
CABAI (Capsicum annuum L.)
Abstrak
Penggunaan marka yang efektif dan efisien sangat menentukan keberhasilan studi penyerbukan silang alami pada tanaman cabai. Marka yang efektif dan efisien dapat diperoleh jika didukung oleh pengetahuan yang lengkap tentang pewarisan sifat berbagai karakter. Tujuan dari penelitian ini adalah untuk mempelajari pola pewarisan sifat beberapa karakter kualitatif dan kuantitatif pada hipokotil dan kotiledon cabai sehingga dapat diperoleh marka yang efektif dan efisien untuk mengevaluasi kejadian penyerbukan silang alami tanaman cabai. Penelitian ini menggunakan 6 populasi cabai, yaitu P1 (IPB C20), P2 (IPB C2), F1, F1R, BCP1, BCP2, dan F2. Hasil penelitian menunjukkan bahwa warna hipokotil dan kotiledon dikendalikan oleh 1 gen, dimana gen pengendali warna ungu dominan terhadap gen pengendali warna hijau pada hipokotil dan gen pengendali warna hijau dominan terhadap gen pengendali warna ungu pada kotiledon. Model genetik dominan dengan interaksi aditif dan aditif-dominan sesuai untuk karakter panjang hipokotil. Model genetik aditif-aditif-dominan dengan interaksi aditif-dominan dan dominan-dominan sesuai untuk karakter diameter hipokotil. Model genetik aditif-dominan dengan interaksi aditif-aditif dan dominan-dominan sesuai untuk karakter panjang dan lebar kotiledon. Heritabilitas dalam arti luas pada karakter yang diamati berada pada kisaran tinggi, sedangkan heritabilitas arti sempit berada pada kisaran sedang-tinggi.
Kata kunci: warna ungu, model genetik, heritabilitas
PENDAHULUAN
Cabai dikenal sebagai tanaman menyerbuk sendiri. Tanaman menyerbuk sendiri adalah tanaman yang memiliki tingkat penyerbukan silang alami kurang dari 5%. Tanaman menyerbuk sendiri umumnya memiliki nilai heterosis yang rendah dan lebih sering diarahkan untuk menjadi varietas galur murni. Akan tetapi beberapa penelitian melaporkan bahwa tanaman cabai memiliki persentase penyerbukan silang alami yang lebih tinggi dari 50% (Campodonico 1983) dan memiliki nilai heterosis yang tinggi (Mantri 2006; Sujiprihati et al. 2007; Daryanto et al. 2010; Sitaresmi et al. 2010). Selain itu, lebih dari 80% varietas cabai yang ada di Indonesia adalah varietas hibrida bukan varietas galur murni (Direktorat Perbenihan dan Sarana Produksi 2011).
menggunakan warna buah, sedangkan Kim et al. (2009) menggunakan marka molekuler untuk mengevaluasi kejadian penyerbukan silang alami tanaman cabai. Penggunaan warna buah muda dan marka molekuler sebagai marka dalam evaluasi kejadian penyerbukan silang alami dinilai masih kurang efektif dan efisien karena membutuhkan waktu yang lama dan biaya yang cukup mahal. Oleh karenanya perlu dilakukan pencarian marka lain yang lebih efektif dan efesien.
Karakter – karakter pada hipokotil dan kotiledon sangat potensial untuk dijadikan sebagai marka morfologi yang efektif dan efisien. Pemilihan suatu karakter untuk dijadikan sebagai marka morfologi memerlukan informasi tentang pola pewarisan sifat dari karakter tersebut. Informasi tentang pola pewarisan sifat karakter kualitatif dan kuantitatif pada hipokotil dan kotiledon cabai masih belum banyak diketahui. Oleh karena itu, penelitian tentang pola pewarisan sifat karakter kualitatif dan kuantitatif pada hipokotil dan kotiledon cabai menjadi penting untuk dilakukan. Tujuan dari penelitian ini adalah untuk (1) untuk mempelajari pola pewarisan sifat beberapa karakter kualitatif dan kuantitatif pada hipokotil dan kotiledon cabai, serta (2) menentukan karakter yang efektif dan efisien untuk dijadikan marka evaluasi kejadian penyerbukan silang alami.
BAHAN DAN METODE
Waktu dan Tempat
Percobaan ini dilakukan pada bulan Mei – Oktober 2012. Pembentukan enam populasi cabai dilakukan di Citeureup, Bogor. Kegiatan perkecambahan dan pengamatan dilakukan di Labdik. Pemuliaan Tanaman, Departemen Agronomi dan Hortikultura, FAPERTA, IPB.
Bahan Tanam
Bahan tanaman yang digunakan pada percobaan ini merupakan cabai koleksi Laboratorium Pemuliaan Tanaman, Departemen Agronomi dan Hortikultura, FAPERTA – IPB. Bahan tanam yang digunakan terdiri atas tetua cabai ungu (IPB C20) dan tetua cabai hijau (IPB C2); turunan pertama (F1) dan turunan pertama resiprokal (F1R) masing-masing 24 bibit; backcross ke tetua betina (BCP1) sebanyak 62 bibit dan backcross ke tetua jantan sebanyak 91 bibit; dan populasi turunan kedua (F2) sebanyak 305 bibit.
Pelaksanaan Percobaan
P c p b k c p m c p p 2 3 4 5 6 K k y k g y
P2, dan F2 cara menyun persilangan benihnya. Perk kotak plastik cabai sebany plastik diba menggunaka cahaya mata pada saat pe
Terd percobaan in 1. Warna h 2. Warna k 3. Panjang kotiledon 4. Diamete kotiledon 5. Panjang ujung ko 6. Lebar k kotiledon
Anal Karakter ku karakter kua yaitu, uji n karakter, be genetik dan yang dilakuk 1. Penduga Pend kuadrat. (1979): Keterang
diperoleh d ngkup tanam
dan selfing
ecambahan k transparan yak 20-24 b asahi sampai an staples. K ahari. Tisu d engamatan. dapat bebera ni. Karakter-hipokotil, dia kotiledon, dia hipokotil (m n er hipokotil n mmbuka s
hipokotil (m otiledon sete kotiledon (m
n telah mem
lisis data d ualitatif diana antitatif men normalitas, p esaran nilai
nilai heritab kan pada per aan nisbah fe dugaan nisb Perhitungan
gan: X2 = X Oi2 = N Ei2 = N
dengan mela man tetua d g matang, b
dilakukan p n berukuran benih tiap ko
i jenuh terle Kotak plasti dibasahi kem
P
apa karakter -karakter ters amati pada sa amati pada s mm), diamat (mm), diam sempurna. mm), diamat lah kotiledon mm), diamat mbuka sempu A dilakukan te alisis berdas ngacu pada L
pendugaan derajat dom bilitas. Beriku rcobaan ini: enotipe bah fenotipe
n uji Chi-kua
X2 hitung Nilai hasil p Nilai yang d
akukan selfi dan turunan buah dipane
pada media t 20 cm x 10 otak. Sebelu ebih dahulu ik kemudian mbali setiap 2
Pengamatan
kualitatif d sebut adalah aat daun kot saat daun kot ti dari bagia
mati pada b
ti dari bagian n membuka ti pada bag urna
Analisis Data
erhadap ka sarkan analis Limbongan e
pengaruh te minansi, kom
ut adalah an
e dilakukan adrat (X2) m
engamatan diharapkan
ing, yang p pertama (F en dan diek
tisu basah ya 0 cm. Setiap um benih dis u kemudian n diletakan d
2 hari sekali
n
dan kuantita h:
tiledon sudah tiledon suda an atas akar
bagian hipo
n pangkal ko sempurna. gian kotiledo
a
arakter kual sis genetika
et al. (2008) etua, jumlah mponen rag nalisis data k
n dengan m mengacu pada
elaksanaann 1). Setelah kstraksi untu
ang diletaka p kotak ditan
semai, tisu p kotak ditut ditempat ya i. Kotak plas
atif yang dia
h membuka ah membuka sampai bag okotil terbes otiledon sam on terlebar litatif dan mendel. An ) dan Arif et
h gen-gen gam, kelayak kualitatif dan
menggunakan a Singh and
nya dengan buah hasil uk diambil
an di dalam nami benih pada kotak tup dengan ang terkena stik dibuka amati pada sempurna sempurna gian bawah sar setelah mpai bagian pada saat kuantitatif. nalisis data
t al. (2012) pengendali kan model n kuantitatif
2. Pendugaan pengaruh tetua betina
Pengaruh tetua betina dilakukan pada karakter kuantitatif dengan cara membandingkan F1 dan F1R dengan uji-t. Rumus uji-t mengacu pada Strickberger (1976):
Keterangan: YF1 = Nilai tengah populasi F1
YF1R = Nilai tengah populasi F1R
SYF1-YF1R = Simpangan baku selisih populasi F1-F1R
3. Uji normalitas pada populasi F2
Uji normalitas digunakan untuk mengetahui sebaran frekuensi populasi F2, sehingga dapat diketahui aksi gen dari masing-masing karakter yang diamati. Uji normalitas menggunakan metode kolmogorov-smirnov.
4. Pendugaan jumlah gen-gen pengendali karakter
Perhitungan pendugaan jumlah gen-gen pengendali karakter mengacu pada Lande (1981):
5. Pendugaan nilai derajat dominansi
Nilai derajat dominansi diduga dengan menghitung potensi rasio (hp). Perhitungan hp mengacu pada Peth and Frey (1966):
Keterrangan:
hp = Potensi rasio MP = Nilai rata-rata dua tetua XF1 = Nilai rata-rata F1 HP = Nilai rata-rata tetua tertinggi
6. Pendugaan kelayakan model genetik
Pendugaan kelayan model genetik dilakukan dengan melakukan uji skala gabungan. Uji skala gabungan mengacu pada Mather dan Jink (1982).
7. Pendugaan komponen ragam
Komponen ragam yang dihitung terdiri atas ragam fenotipe (VF2), ragam
fenotipe backcros (VBCP), ragam lingkungan (VE), ragam genotipe (VG), dan
ragam aditif (VA).
8. Pendugaan nilai heritabilitas
Pendugaan heritabilitas yang dihitung terdiri atas heritabilitas arti luas dan heritabilitas arti sempit. Perhitungan heritabilitas arti luas mengacu pada Allard (1960), sedangkan heritabilitas arti sempit mengacu pada Warner (1952):
Keterangan:
h2bs = Heritabilitas arti luas VF1 = Ragam populasi F1
h2ns = Heritabilitas arti sempit VF2 = Ragam populasi F2
VBCP1 = Ragam populasi silang balik ke P1 VP1 = Ragam Populasi P1
y b b w F y G n h h p p b p t w g p m a w t ( m p Terd yaitu ungu berwarna hij berwarna un warna hipok F1R IPB C2 yang berwar
Gambar 2 W
Hasi nisbah fenot hijau pada p hipokotil se pengendali w pada karakte bean dikend pengendali tanaman cab warna ungu Hal y genetik men pada popula menunjukka alel dan gen warna ungu tanaman lim
Anal (dihibrid), y mendel pada pada popula
H
dapat dua m dan hijau. G ijau, sedangk ngu. Tanam kotil yang be
2 x IPB C2 rna hijau (Ga
Warna hipok IPB C2, dan x IPB C2 : H kotiledon hij
l analisis gen tipe nisbah 3 populasi BCP ecara sederh
warna ungu er ini. Menu dalikan oleh warna ungu bai, sementa bersifat dom yang berbed ndel pada ka asi F2 dan 1 an bahwa kar n pengendal pada karak ma bean diken
lisis genetik yaitu warna a kedua kara asi F2 (Tabel
HASIL DA
Warna Hip
macam warna Genotipe cab kan genotip man F1 dan erbeda deng 20 memiliki ambar 2).
kotil dan kot n IPB C2. IP Hipokotil un ijau
netik mende 3 ungu : 1 h P1 (Tabel 2) hana dikend u bersifat do urut ICRISA
3 gen. Wang u bersifat do ara Jones et
minan pada b da dihasilkan arakter ini m 1 hijau : 1 u
rakter warna li warna hija kter ini. Men ndalikan ole k mendel jug a hipokotil
akter tersebu l 2). Padahal
AN PEMB
pokotil dan
a hipokotil d bai IPB C2 pe IPB C20 F1R genoti gan warna ko
hipokotil ya
tiledon geno PB C20: hipo ngu dan kotil
el pada karak hijau pada p
). Hal ini m dalikan oleh
ominan terh T (1975), w g dan Bossla ominan pada
al. (2003) m buah tomat. n pada karak menghasilkan
ungu pada p a kotiledon d
au bersifat d nurut ICRIS eh 1 lokus de ga dilakukan
dan warna ut tidak men l hasil analis
AHASAN
Kotiledon
dan kotiledo memiliki h memiliki h ipe IPB C2 otiledonnya. ang berwarn
otipe cabai IP okotil dan ko
ledon hijau,
kter warna h opulasi F2 d menunjukkan
1 lokus de adap gen pe warna hipoko and (2006) m a karakter b melaporkan
kter warna ko n nisbah fen
populasi BC dikendalikan
dominan ter SAT (1975), engan 2 alel. n terhadap d
kotiledon. nghasilkan n sis genetik m
N
on pada pen hipokotil dan ipokotil dan x IPB C20 Baik tanam na ungu dan
PB C20, F1 otiledon ungu
IPB C2 : hip
ipokotil me dan nisbah bahwa kara engan 2 ale engendali w otil pada tan melaporkan bunga dan b
bahwa gen
otiledon. Ha notipe 3 hija CP2 (Tabel n oleh 1 loku rhadap gen warna hipo dua karakter Hasil analis nisbah fenot mendel karak nelitian ini, n kotiledon n kotiledon 0 memiliki man F1 dan
n kotiledon
IPB C20 x u, IPB C20 pokotil dan
warna Hal in hipoko Tabel Keteran 9:3:3: dengan yang t tipe re yang s Peters bentuk
K
lebar k tengah Hal in tersebu dan P kotiled bahwa karakt diband kotiled hipokotil da ni memuncu
otil dengan w
2 Nilai X2h (IPB C2
ngan : HU = H Kotiled mendel
Jin et al. ( 1 pada popu n ketahanan terjadi antara epulsi. Hal i
sama-sama m on (1959) k buah pada
Keragaan K
Nilai tenga kotiledon po h F1 semua ni menunjukk
ut.
Nilai tenga 1 sedangkan don mendek a aksi gen ad ter panjang dingkan aksi don.
an warna ko lkan dugaan warna kotile
hitung warna x F1), BCP2
Hipokotil ung
don hijau, tn ra
l
1993) melap ulasi F2 men
terhadap pe a warna hipo ini ditandai memberikan melaporkan tanaman cab
Kotiledon da
ah panjang h opulasi P2
karakter ber kan bahwa t
ah F1 karakt n nilai tenga kati salah satu
ditif diduga hipokotil se i gen aditif p
otiledon men n bahwa terd
don.
a hipokotil d 2 (IPB C20 x
gu, HH = Hipo asio sesuai nisb
porkan bahw andakan ada enyakit karat okotil dan w
dengan gen n satu gen r n bahwa ter
bai. an Hipokoti hipokotil, di selalu lebih rada diantar tidak terdap
ter panjang h ah F1 karakte u tetua P1 at lebih berper edangkan ak
pada karakte
nghasilkan n dapat keterp
dan warna ko x F1) dan F2
okotil hijau, K bah mendel, **
wa segregasi anya keterpa t daun pada warna kotiled notipe tetua
esesif dan d rdapat linka
il pada Beb
iameter hipo besar diban ra nilai teng
at heterobel
hipokotil ber er diameter tau P2 (Gam ran dibandin ksi gen dom
er diameter
nisbah fenotip pautan antara
otiledon pad 2 (IPB C2 x
KU = Kotiledo * rasio tidak se
gen Rph7 d autan antara tanaman bar don cabai di cabai IPB C dominan kep age antara w
erapa Popu
okotil, panjan ndingkan po ah kedua ke ltiosis pada k
rada tepat d hipokotil, p mbar 3). Hal ngkan aksi ge minan didug
hipokotil, p
pe 3:1 (Tabe a karakter w
da populasi B IPB C20)
on ungu, dan esuai dengan n
dan Xa yang
warna kecam rley. Keterpa iduga merup C2 dan IPB pada turunan warna buah ulasi Cabai ng kotiledon opulasi P1. etua (Gamba karakter-kar
i tengah tetu panjang dan
G d F t p p ( p b W p p u
Gambar 3 N
Popu dibandingka F2 memiliki tertinggi pa populasi ters Hasi perbedaan y (Tabel 3). H pewarisan ka bahwa hany Weber (195 pewarisan p penelitian T ukuran kotil
(a)
(c)
Nilai tengah BCP 2 (a) hipokotil, da
ulai F2 pad an populasi l
i keragaman ada populas
sebut.
l uji penga yang nyata Hal ini menun
arakter-kara ya gen-gen d 59) melapor panjang hip Thayer (1934 edon tanama
h dan simpan panjang hi an (d) lebar k
da semua k lainnya (Gam
yang terting si F2 terjad
Pengar
aruh tetua antara F1 d njukkan bah akter tersebut dalam inti ya rkan bahwa pokotil dan 4) menunjuk an melon.
ngan baku p ipokotil, (b) kotiledon be
karakter me mbar 3). Hal ggi dibandin di karena a
ruh Tetua B
betina men dan F1R un hwa tidak ter
t pada tanam ang mengen
tidak terda n kotiledon kan bahwa te
(b) (d populasi P1, panjang k eberapa popu emiliki jang l ini menunj ngkan popula adanya segr
Betina
nunjukkan b ntuk semua rdapat penga man cabai. H ndalikan kara
apat pengaru tanaman erdapat peng
d)
P2, F1, F2, otiledon, (c ulasi cabai
gkauan yan jukkan bahw asi lainnya. K regasi maks
bahwa tida peubah yan aruh tetua be Hal ini mengi akter-karakte uh tetua bet tomat. Nam garuh tetua b
Tabel 3. Uji pengaruh tetua betina populasi F1 dan F1R pada beberapa karakter cabai
Karakter Nilai Tengah t-value p-value
F1 F1R
Panjang hipokotil 11.62 12.36 -0.97 0.35
diameter hipokotil 0.59 0.57 -1.89 0.08
Panjang kotiledon 8.82 9.13 -1.05 0.32
Lebar kotiledon 1.82 1.84 -0.35 0.73
Adanya pengaruh tetua betina pada pewarisan suatu karakter akan menyebabkan keturunan persilangan resiproknya memberikan hasil yang berbeda, dimana ciri tetua betina akan lebih dominan tampak pada keturunannya. Hal ini menyebabkan tidak dapat digabungkannya analisis pewarisan sifat F1 dan F1R karena akan menghasilkan segregasi F2 berbeda dan tidak sesuai dengan segregasi mendel. Arif et al. (2012) melaporkan bahwa juga tidak terdapat pengaruh tetua betina pada pewarisan karakter umur berbunga, umur berbungan, dan bobot per buah tanaman cabai.
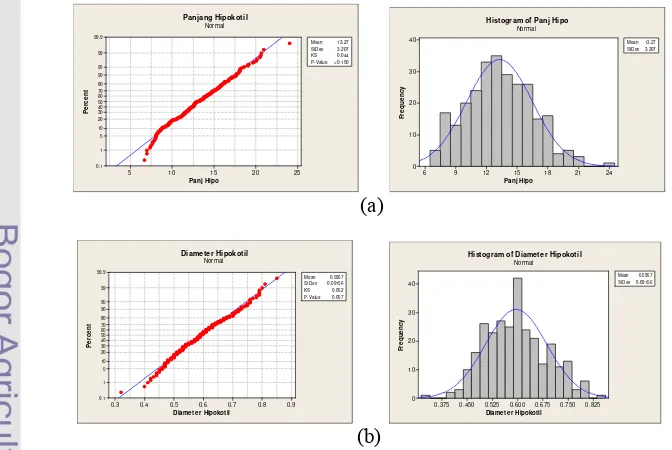
Uji Normalitas F2
Uji normalitas dilakukan untuk melihat sebaran frekuensi populasi F2. Semua karakter yang diamati memiliki sebaran frekuensi populasi F2 yang normal dan kontinyu (Gambar 4 dan 5). Sebaran yang kontinu dan normal pada F2 mengindikasikan bahwa karakter-karakter tersebut dikendalikan banyak gen (poligenik). 25 20 15 10 5 99.9 99 95 90 80 70 60 50 40 30 20 10 5 1 0.1 Panj Hipo P e rc e n t Mean 13.27 StDev 3.207 KS 0.044 P- Value> 0.150 Panjang Hipokotil Normal 24 21 18 15 12 9 6 40 30 20 10 0 Panj Hipo Fr e q u e n c y Mean 13.27 StDev3.207 Histogram of Panj Hipo
Normal (a) 0.9 0.8 0.7 0.6 0.5 0.4 0.3 99.9 99 95 90 80 70 60 50 40 30 20 10 5 1 0.1
Diamet er Hipokot il
P e rc e n t Mean 0.5957 StDev 0.09156 KS 0.052 P-Value 0.057 Diameter Hipokotil Normal 0.825 0.750 0.675 0.600 0.525 0.450 0.375 40 30 20 10 0
Diamet er Hipokot il
Fr e q u e n c y Mean 0.5957 StDev0.09156 Histogram of Diameter Hipokotil
Normal
(b)
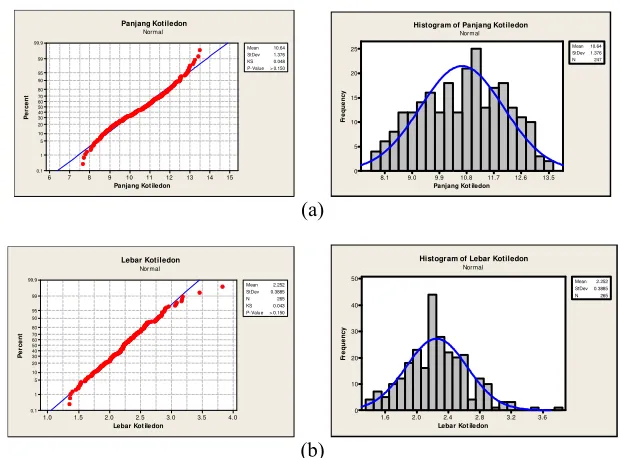
[image:32.612.108.444.435.660.2]15 14 13 12 11 10 9 8 7 6 99.9 99 95 90 80 70 60 50 40 30 20 10 5 1 0.1
Panjang Kot iledon
P e rc e n t Mean 10.64 StDev 1.376 KS 0.048 P-Value> 0.150 Panjang Kotiledon Normal 13.5 12.6 11.7 10.8 9.9 9.0 8.1 25 20 15 10 5 0
Panjang Kot iledon
Fr e q u e n c y Mean10.64 StDev1.376 N 247 Histogram of Panjang Kotiledon
Normal (a) 4.0 3.5 3.0 2.5 2.0 1.5 1.0 99.9 99 95 90 80 70 60 50 40 30 20 10 5 1 0.1
Lebar Kot iledon
P e rc e n t Mean 2.252 StDev 0.3885 N 265 KS 0.043 P- Value> 0.150 Lebar Kotiledon Normal 3.6 3.2 2.8 2.4 2.0 1.6 50 40 30 20 10 0
Lebar Kot iledon
Fr e q u e n c y Mean 2.252 StDev0.3885 N 265 Histogram of Lebar Kotiledon
Normal
[image:33.612.163.473.75.304.2](b)
Gambar 5 Sebaran populasi F2 (IPB C2 x IPB C20). (a) karakter panjang hipokotii dan (b) diameter hipokotil
Beberapa penelitian pada tanaman yang berbeda memberikan hasil yang berbeda. Weber (1959) melaporkan bahwa panjang hipokotil dan panjang kotiledon tanaman tomat dikendalikan 1 gen dengan 2 alel per lokus, sedangkan Walters (2001) melaporkan bahwa panjang hipokotil tanaman mentimun dikendalikan secara sederhana oleh 1 gen dengan 2 alel per lokus.
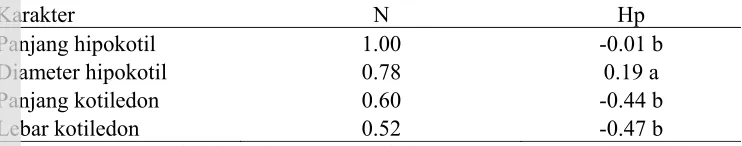
Aksi Gen dan Jumlah Pengendali Gen
Tabel 4 Jumlah gen pengendali dan aksi gen pada beberapa karakter cabai
Karakter N Hp
Panjang hipokotil 1.00 -0.01 b
Diameter hipokotil 0.78 0.19 a
Panjang kotiledon 0.60 -0.44 b
Lebar kotiledon 0.52 -0.47 b
Keterangan: N = Jumlah kelompok gen pengendali, hp = potensi rasio, a = aksi gen dominan positif tidak sempurna, b = aksi gen dominan negatif tidak sempurna
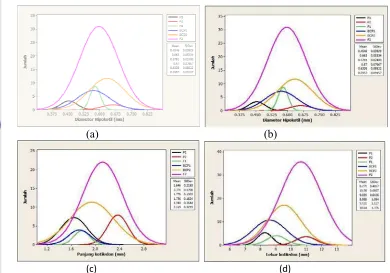
Karakter panjang hipokotil, diameter hipokotil, panjang kotiledon, dan lebar kotiledon memiliki jumlah gen pengendali yang sama. Karakter-karakter tersebut dikendalikan oleh 1 kelompok gen (Tabel 3). Satu kelompok gen dapat terdiri atas beberapa atau banyak gen. Karakter lain pada cabai yang dikendalikan oleh 1 kelompok adalah umur berbunga (Arif et al. 2012).
Model Genetik
Hasil uji kecocokan model genetik menunjukkan bahwa terdapat model genetik yang berbeda antar karakter yang berbeda pada tanaman cabai. Model genetik yang sesuai untuk karakter panjang hipokotil adalah model aditif-dominan dengan pengaruh interaksi aditif-aditif dan interaksi aditif-dominan dengan lima komponen m[d][h][i][j], sedangkan model genetik yang sesuai untuk karakter diameter hipokotil adalah model dominan dengan pengaruh interaksi aditif-dominan dan interaksi aditif-dominan-aditif-dominan dengan lima komponen m[d][h][j][l]. Model aditif-dominan dengan pengaruh interaksi aditif-aditif dan interaksi dominan-dominan dengan lima komponen m[d][h][i][l] paling sesuai untuk karakter panjang dan lebar kotiledon (Tabel 5).
Tabel 5 Uji kecocokan model genetik beberapa karakter cabai
Model genetik Panjang
hipokotil
Diameter hipokotil
Panjang kotiledon
Lebar kotiledon
m d 125.16** 26.483** 189.93 ** 136.50**
m d h 121.21** 26.413** 189.10** 123.37 **
m d h i 90.95** 11.759** 72.71** 17.65**
m d h j 20.10** 18.454** 175.65** 121.19**
m d h l 114.69** 5.828* 168.53** 67.27**
m d h i j 2.89 tn 6.735** 65.32 ** 15.07 **
m d h i l 73.30** 5.589* 3.62 tn 0.235 tn
m d h j l 12.47** 0.574 tn 155.76** 61.213**
Keterangan : ** model tidak sesuai pada taraf α = 1%, * model tidak sesuai pada taraf α = 5%,
tn model genetik sesuai pada taraf α = 5%
[image:34.612.106.478.472.591.2]g d ( p D c d H m d t k d d m d t k e m T K g p c i l k d genetik dom dibandingka (aditif-aditif pengaruh ak Dengan dem cabai adalah Kara dominan ber Hal ini me mengarah k dominan-dom tinggi. Nilai komponen g dibandingka dengan nila menunjukan diameter hip tanda antara ketahanan p epistasis dup mengendalik
Tabel 6 Pen
Keterangan: m a d α Kara genetik inter positif (Tab cenderung m interaksi dom lebih tinggi. komponen g dibandingka
minan menu an gen dom f) sama-sam
ksi gen epi mikian didug
h aditif komp akter diamet
rnilai positif enunjukkan ke tetua yan minan cend i komponen genetik aditi an gen aditi ai kompone n bahwa terd pokotil. Uta a nilai kompo
adi terhadap plikat pada k kan karakter
ndugaan kom
m = nilai tengah aditif x aditif, dominan x dom
α = 1%, tn = ti
akter panjan raksi aditif-a bel 6). Hal i mengarah ke minan-domi Nilai komp genetik aditi an gen aditi
unjukkan b minan. Nilai ma bertanda n
istasis kom ga aksi gen y plementer. ter hipokotil
f dan interak bahwa kom ng nilai rata-derung meng n genetik do if menunjuk if. Nilai ko en genetik dapat penga ami et al. ( onen genetik p penyakit b
karakter ters r diameter hi
mponen gene
h, d = pengaru j = pengaruh minan,* berbed
idak berbeda n
ng dan leba aditif bernila ini menunju e tetua yang inan cenderu onen genetik if menunjuk if. Nilai ko
bahwa gen i komponen
negatif. Hal mplementer p
yang mengen
l memiliki p ksi dominan mponen int -ratanya leb garah ke te ominan yan kkan bahwa omponen ge
interaksiny aruh aksi gen (2005) mela k dominan te blas ras 033, sebut. Denga ipokotil adal
etik beberapa
uh aditif, h = p h interaksi ad a nyata pada ta nyata pada taraf
ar kotiledon ai negatif dan ukkan bahwa nilai rata-ra ung mengara
k dominan y kkan bahwa omponen ge
aditif berk n genetik ad
ini menunj pada karakt ndalikan kara
parameter g n-dominan y teraksi aditi bih besar da etua yang n ng lebih bes gen domin netik domin ya (domina
n epistasis d aporkan bah
erhadap inte mengindika an demikian ah dominan
a karakter ca
pengaruh domi ditif x domina
araf α = 5%, ,*
f α = 5%
n sama-sama n interaksi d a komponen atanya lebih
ah ke tetua y yang lebih be
gen domin netik domin
kontribusi le ditif dan in jukkan bahw ter panjang akter panjan enetik intera yang negatif if-dominan an kompone nilai rata-rat sar dibandin nan lebih be
nan berlawa an-dominan) duplikat pad hwa adanya eraksinya pad asikan adany n diduga aks
duplikat.
abai
inan, i = penga an, l = pengar ** berbeda nya
a memiliki dominan-dom n interaksi a rendah dan yang nilai r esar dibandin nan lebih be
nan berlawa ebih besar nteraksinya wa terdapat hipokotil. ng hipokotil aksi aditif-f (Tabel 6).
cenderung n interaksi anya lebih ngkan nilai erkontribusi anan tanda ). Hal ini da karakter
perbedaan da karakter ya aksi gen si gen yang
aruh interaksi ruh interaksi ata pada taraf
dengan menun karakt panjan kisaran sedang hipoko sempit bahwa berbed aditif adanya Tabel kisaran sedang diband lebih skala diband panjan dalam propor dengan lebih
n nilai ko njukan bahw ter tersebut. ng dan lebar
Heritabilita n tinggi seda g (Tabel 7) otil Pinus ha
t yang jauh a proporsi ra da dengan h lebih besar a pengaruh i
7. Kompone
Heritabilita n tinggi seda g (Tabel 7 dingkan heri besar diban gabungan dingkan peng
Heritabilita ng dan lebar m arti sempi rsi ragam ad n hasil uji
besar diban
omponen ge wa terdapat Dengan dem hipokotil ad
as dalam art angkan herit ). Nilai her
alepensis M lebih renda agam domin hasil uji ska dibandingka interaksi adi
en ragam dan
as dalam art angkan herit ). Heritabil itabilitas art ndingkan rag yang menu garuh aditif as dalam ar r kotiledon it yang men ditif lebih be skala gabun ndingkan pe
enetik inter pengaruh a mikian didug dalah domina
Heritab
ti luas (h2bs)
tabilitas dala ritablitias ya Mill (Harfauc ah dibanding nan lebih be ala gabunga an pengaruh tif-dominan
n heritabilita
ti luas (h2bs)
tabilitas dala litas dalam ti luas menu gam aditif. H unjukkan b
(Tabel 6). rti luas (h2b
berada pada ndekati her esar dibandi ngan yang m engaruh dom
raksinya (d aksi gen ep ga aksi gen y an duplikat.
bilitas
) karakter pa am arti semp ang tinggi j che et al. 20 gkan heritab esar dibandi an yang me h dominan. P
pada karakt
as beberapa k
karakter dia am arti semp arti sempi unjukan bahw
Hal ini sesu ahwa peng
bs) dan arti
a kisaran tin ritabilitas ar ingkan ragam menunjukkan minan pada dominan-dom pistasis dupl yang mengen anjang hipok pit (h2ns) ber
juga terdapa 003). Heritab bilitas arti lu
ingkan ragam enunjukkan b
Perbedaan in ter ini (Tabel
karakter cab
ameter hipok pit (h2ns) ber
it yang jau wa proporsi uai dengan aruh domin
sempit (h2n
nggi (Tabel rti luas men m dominan. n bahwa pe karakter pa
minan). Hal likat pada k ndalikan kar
kotil berada rada pada kis at pada pan bilitas dalam uas menunju
m aditif. Ha bahwa peng ni diduga ka l 6)
bai
kotil berada rada pada kis uh lebih re i ragam dom
dengan has nan lebih b
ns) pada kar
7). Heritab nunjukan ba
kotiledon. Perbedaan ini diduga karena adanya pengaruh interaksi aditif-aditif pada karakter ini (Tabel 6). Beberapa penelitian lain pada cabai menunjukkan bahwa terdapat nilai heritabilitas yang tinggi karakter umur berbunga (Lestari et al. 2006) umur panen (Arif et al. 2012), ketahanan terhadap Phytophthora capsici
(Rahmi et al. 2011), bobot buah (Marame et al. 2008; Sharma et al. 2010; Syukur
et al. 2010), bobot buah per tanaman (Syukur et al. 2010), panjang dan diameter buah (Syukur et al. 2011).
SIMPULAN
Karakter warna hipokotil dan kotiledon dikendalikan oleh satu gen. Gen pengendali warna ungu bersifat dominan terhadap gen pengendali warna hijau pada hipokotil cabai, sedangkan gen pengendali warna hijau bersifat dominan terhadap gen pengendali warna ungu pada kotiledon cabai.
Pewarisan panjang hipokotil, diameter hipokotil, panjang dan lebar kotiledon tidak dipengaruhi oleh tetua betina. Model genetik aditif-dominan dengan interaksi aditif-aditif dan interaksi aditif-dominan sesuai untuk karakter panjang hipokotil. Model genetik aditif-dominan dengan interaksi aditif-dominan dan interaksi dominan-dominan sesuai untuk karakter diameter hipokotil. Model genetik aditif-dominan dengan interaksi aditif-aditif dan interaksi dominan-dominan sesuai untuk karakter panjang dan lebar kotiledon. Heritabilitas dalam arti luas pada karakter yang diamati berada pada kisaran tinggi, sedangkan heritabilitas arti sempit berada pada kisaran sedang-tinggi.
STUDI PENYERBUKAN SILANG ALAMI DAN BUATAN
PADA BEBERAPA GENOTIPE CABAI (Capsicum annuum L.)
Abstrak
Informasi sistem penyerbukan sangat penting bagi penentuan metode dan arah pemuliaan suatu tanaman. Namun, informasi sistem penyerbukan pada cabai masih belum konsisten sehingga menyebabkan studi penyerbukan silang alami dan buatan pada tanaman cabai penting untuk dilakukan. Percobaan ini bertujuan untuk memperoleh informasi tentang penyerbukan silang alami beberapa genotipe cabai. Percobaan ini dilakukan pada bulan September 2011 sampai Maret 2012 di Labdik Pemuliaan Tanaman dan Kebun Percobaan, Leuwikopo, Departemen Agronomi dan Hortikulura, FAPERTA-IPB. Percobaan ini terdiri dari dua kegiatan, yaitu (1) studi penyerbukan silang alami pada beberapa genotipe cabai dan (2) studi penyerbukan silang buatan pada beberapa genotipe cabai. Hasil pengamatan menunjukkan bahwa terdapat perbedaan tingkat penyerbukan silang alami antar genotipe cabai yang berbeda. Kemampuan menyerbuk silang yang tinggi dan posisi stigma yang lebih tinggi dibandingkan anther diduga berperan besar dalam peningkatan kejadian penyerbukan silang alami pada tanaman cabai. Hasil pengamatan juga menunjukkan bahwa tidak terdapat perbedaan tingkat penyerbukan silang alami antar radius tanaman yang berbeda. Pola penyerbukan silang alami yang terbentuk bersifat acak, menyebar, dan tidak teratur. Hal ini mengindikasikan bahwa penyerbukan silang alami pada tanaman cabai tidak dibantu oleh angin, melainkan dibantu oleh serangga lebah.
PENDAHULUAN
Tujuan yang ingin dicapai dari percobaan ini, yaitu 1) membandingkan tingkat penyerbukan silang alami beberapa genotipe cabai. 2) membandingkan morfologi bunga dan tingkat crossability beberapa genotipe cabai, dan 3) mengetahui pengaruh morfologi bunga dan tingkat crossability terhadap tingkat penyerbukan silang alami beberapa genotipe cabai.
BAHAN DAN METODE
Studi Penyerbukan Silang Buatan Beberapa Genotipe Cabai
Waktu dan Tempat
Percobaan ini dilakukan pada bulan September 2011 sampai Maret 2012. Kegiatan persemaian dan penanaman dilakukan di tempat yang terpisah. Kegiatan persemaian dilakukan di Laboratorium Pendidikan (Labdik) Pemuliaan Tanaman, sedangkan kegiatan penanaman dilakukan di Kebun Percobaan Leuwikopo, Departemen Agronomi dan Hortikultura, FAPERTA – IPB.
Bahan Tanaman
Genotipe-genotipe cabai yang digunakan pada percobaan ini terdiri atas genotipe cabai IPB C20, IPB C2, IPB C5, dan IPB C120. Genotipe cabai IPB C20 yang memiliki hipokotil berwarna ungu digunakan sebagai tetua donor polen, sedangkan genotipe cabai IPB C2, IPB C5, dan IPB C120 yang memiliki hipokotil berwarna hijau digunakan sebagai tetua resipien polen dan merupakan genotipe-genotipe cabai yang diuji tingkat penyerbukan silang alaminya.
Pelaksanaan Percobaan
Gambar 6 Layout percobaan penyerbukan silang alami beberapa genotipe cabai (Capsicum annuum L.). genotipe cabai IPB C20 (hipokotil, kotiledon, bunga, dan buah muda ungu, genotipe cabai IPB C120,
IPB C2, atau IPB C5 (hipokotil, kotiledon, bunga, dan buah muda daun)
Percobaan diawali dengan kegiatan penyemaian. Benih disemai sebanyak 2 benih per lubang tray. Penyiraman dilakukan setiap hari pada pagi dan sore hari. Pemupukan dilakukan satu minggu sekali setelah bibit berumur 2 minggu setelah semai (MSS). Pupuk yang digunakan adalah pupuk NPK Mutiara dengan konsentrasi 10 g L-1 dan Gandasil 2 g L-1. Penyemprotan pestisida dilakukan jika terlihat gejala serangan hama dan penyakit pada persemaian.
Kegiatan pengolahan lahan dan pembuatan bedengan dilakukan bersamaan pada saat melakukan kegiatan penyemaian. Setelah dilakukan pengolahan lahan dan pembuatan bedengan, maka dilakukan pemasangan mulsa. Penanaman dilakukan setelah bibit cabai berumur 35 hari setelah semai (HSS) atau minimal sudah memiliki empat helai daun dewasa. Lubang tanam dibuat dengan menggunakan tugal. Sebelum dilakukan penanam, terlebih dahulu dilakukan pemasangan ajir pada dekat lubang tanam. Penanaman (transplanting) dilakukan pada sore hari dengan jumlah tanaman satu tanaman per lubang tanam. Penyulaman bibit dilakukan 1-2 minggu setelah tanam.
Kegiatan pemeliharaan tanaman terdiri atas penyiraman, pemupukan, pemberian pestisida, pewiwilan tunas air, pembumbunan, dan penyiangan gulma. Penyiraman dilakukan pada pagi dan sore hari jika tidak terjadi hujan. Pemupukan dilakukan setiap satu minggu sekali setelah satu minggu setelah tanam (1 MST) dengan menggunakan pupuk NPK mutiara dengan konsentrasi 10 g L-1 sebanyak 250 ml per tanaman. Penyemprotan pestisida dilakukan satu minggu sekali dengan menggunakan fungisida Dithane M-45 atau Antracol 2 g L-1, insektisida Curacron 2 ml L-1 dan akarisida Kelthine 2 cc L-1. Pengendalian gulma dilakukan secara manual. Pewiwilan tunas air dan pembumbunan dilakukan agar tanaman dapat tumbuh optimal. Kegiatan pemanenan dilakukan pada saat cabai telah mencapai tingkat kematangan 75%. Pemanenan dilakukan setiap minggu selama 8 minggu.
Buah cabai hasil panen dari genotipe cabai IPB C2, IPB C5 dan IPB C120 dipisahkan berdasarkan tanamannya, lalu diambil bijinya untuk diekstraksi menjadi benih. Ekstraksi benih cabai dilakukan secara manual dengan menggunakan alat bantu pinset dan pisau cutter. Cabai dibelah dengan pisau
menggunakan pinset. Biji cabai yang sudah terpisah dari buahnya kemudian dikeringanginkan selama satu minggu.
Biji cabai yang sudah siap menjadi benih, selanjutnya disemai dan dievaluasi kejadian penyerbukan silang alaminya. Penyemaian dilakukan di dalam kotak plastik transparan dengan menggunakan media tisu basah. Kegiatan evaluasi dilakukan setelah kecambah memiliki kotiledon sempurna. Kejadian penyerbukan silang alami ditandai dengan munculnya hipokotil berwarna ungu pada benih yang dikecambahkan. Hal ini karena warna kecambah genotipe cabai IPB C2, IPB C5, dan IPB C120 yang sebenarnya adalah hijau, sehingga munculnya warna ungu pada hipokotil menandakan sudah tercemarnya genotipe-genotipe tersebut oleh polen cabai ungu (genotipe-genotipe cabai IPB C20). Warna ungu bersifat dominan terhadap warna hijau pada hipokotil cabai (hasil percobaan 1).
Pengamatan
Pengamatan yang dilakukan pada percobaan ini terdiri atas pengamatan karakter kualitatif dan karakter kuantitatif. Pengamatan-pengamatan tersebut terdiri atas:
1. Pengamatan posisi stigma, diamati pada minggu ke enam setelah tanam (MST) di lapang
2. Umur berbunga (hari setelah semai), dihitung dari awal persemaian sampai tanaman memiliki satu bunga yang telah mekar sempurna
3. Panjang tangkai anther (mm), diamati pada minggu ke enam setelah tanam (MST) di lapang
4. Panjang anther (mm), diamati setelah tanaman berumur 6 MST. 5. Panjang putik (mm), diamati setelah tanaman berumur 6 MST 6. Tinggi tanaman (cm), diamati setelah mulai panen minggu ke-2 7. Warna mahkota, diamati setelah tanaman berumur 6 MST 8. Warna anther, diamati setelah tanaman berumur 6 MST 9. Warna putik, diamati setelah tanaman berumur 6 MST 10.Warn stigma, diamati setelah tanaman berumur 6 MST
11.Warna hipokotil, diamati pada saat kotiledon telah