EKTOMIKORIZA PADA
Shorea
spp. DI KAWASAN HUTAN
HAURBENTES JASINGA BOGOR
FAJAR ISLAM SITANGGANG
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Ektomikoriza pada Shorea spp. di Kawasan Hutan Haurbentes Jasinga Bogor adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
FAJAR ISLAM SITANGGANG. Ektomikoriza pada Shorea spp. di Kawasan Hutan Haurbentes Jasinga Bogor. Dibimbing oleh NAMPIAH SUKARNO dan SRI LISTIYOWATI.
Shorea spp. merupakan anggota famili Dipterocarpaceae. Shorea spp.
merupakan tanaman hutan hujan tropis yang bernilai ekonomi penting karena kualitas produk kayu dan non kayunya yang tinggi. Cendawan ektomikoriza digunakan sebagai pupuk hayati untuk Shorea spp. Penelitian ini bertujuan untuk mempelajari morfotipe akar ektomikoriza Shorea spp. dan mengisolasi serta mengidentifikasi cendawan ektomikorizanya. Sebanyak enam puluh sampel akar diambil dari tegakan Shorea saminis (plot 1), Shorea palembanica (plot 2), dan
Shorea stenoptera (plot 3). Hasil penelitian menunjukkan bahwa semua tanaman
berasosiasi dengan cendawan ektomikoriza dengan persentase kolonisasi sekitar 60%. Kelimpahan akar ektomikoriza paling banyak ditemukan pada plot 2, kemudian diikuti plot 3, dan paling sedikit pada plot 1. Data analisis akar menunjukkan bahwa terdapat 5 macam morfotipe utama, yakni irregular pinnate cokelat, irregular pinnate hitam, irregular pinnate perak, monopodial pinnate cokelat, dan monopodial pinnate hitam. Irregular pinnate cokelat ialah morfotipe dominan dengan persentase sebesar 60.9%. Sebanyak 15 isolat yang memiliki morfologi koloni berbeda berhasil diisolasi dari 3 macam morfotipe ektomikoriza, yaitu irregular pinnate cokelat, irregular pinnate hitam, dan monopodial pinnate cokelat. Seluruh isolat merupakan miselia sterilia. Beberapa cendawan memiliki karakteristik miselium khusus dengan membentuk anastomosis hifa dan menghasilkan percabangan miselium yang intensif.
ABSTRACT
FAJAR ISLAM SITANGGANG. Ectomycorrhiza of Shorea spp. in Haurbentes Research Forest Jasinga Bogor. Supervised by NAMPIAH SUKARNO and SRI LISTIYOWATI.
Shorea spp. are member of Dipterocarpaceae family. They are tropical
rainforest plant which have high economic value due to high quality of their timber and non-timber products. Ectomycorrhizal fungi had been used as biofertilizer for Shorea spp. This research aimed to study the ectomycorrhizal root morphotypes, isolation and identification of ectomycorrhizal fungi associated with
Shorea spp. Sixty ectomycorrhyzal root tip samples were taken from Shorea
saminis (plot 1), Shorea palembanica (plot 2), and Shorea stenoptera (plot 3). The results showed that all plants associated with ectomycorrhiza, with percentage of roots colonization around 60%. The highest abundance of associated root was found at plot 2, followed by plot 3, and the lowest was found at plot 1. Data of root morphotype analysis indicated that there were 5 major of root tip morphotypes. There were brown irregular pinnate, black irregular pinnate, silver irregular pinnate, brown monopodial pinnate, and black monopodial pinnate. Brown irregular pinnate was the dominant root tips morphotypes with the frequency was 60.9%. A total of 15 different isolates were isolated from 3 ectomycorrhizal root tips morphotypes, e.g. brown irregular pinnate, black irregular pinnate, and brown monopodial pinnate. All of the isolates were mycelia sterilia. Some of the fungi had special mycelial characteristics by producing hyphal anastomosis and produce intensive branching mycelium.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
pada
Departemen Biologi
EKTOMIKORIZA PADA
Shorea
spp. DI KAWASAN HUTAN
HAURBENTES JASINGA BOGOR
FAJAR ISLAM SITANGGANG
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM INSTITUT PERTANIAN BOGOR
Judul Skripsi : Ektomikoriza pada Shorea spp. di Kawasan Hutan Haurbentes Jasinga Bogor
Nama : Fajar Islam Sitanggang NIM : G34090019
Disetujui oleh
Dr Ir Nampiah Sukarno Pembimbing I
Dr Sri Listiyowati, MSi Pembimbing II
Diketahui oleh
Dr Ir Iman Rusmana, MSi Ketua Departemen
PRAKATA
Puji syukur penulis panjatkan kehadirat Allah SWT atas limpahan rahmat dan karunia-Nya sehingga dapat menyelesaikan karya ilmiah yang berjudul Ektomikoriza pada Shorea spp. di Kawasan Hutan Haurbentes. Jasinga Bogor. Kegiatan Penelitian ini berlangsung dari bulan Maret-September 2013 di Laboratorium Mikologi Departemen Biologi IPB.
Terima kasih penulis ucapkan kepada Dr. Ir. Nampiah Sukarno selaku pembimbing I dan pemberi dana penelitian, Dr. Sri Listiyowati, MSi selaku pembimbing II atas bimbingan dan pengarahan yang telah diberikan, dan Hadi Sunarso, MSi selaku penguji atas saran dan masukannya. Terima kasih kepada Buya, Ibu, serta seluruh keluarga atas segala dukungan baik semangat, materi, serta doa selama penulis menempuh pendidikan hingga karya ilmiah ini terselesaikan. Ungkapan terima kasih juga ditujukan kepada Ibu Emi, Bapak Kusnadi, Bapak Adi, Kak Erwin, Ivan Permana Putra, Sepriyadi Rihi, Nicho Nurdebyandaru, Muhammad Ginanjar dan teman-teman Biologi 46 untuk bantuan dan kerja samanya.
Semoga karya ilmiah ini dapat bermanfaat untuk perkembangan ilmu pengetahuan serta dapat menambah wawasan dan pengetahuan bagi kita semua.
DAFTAR ISI
DAFTAR ISI ix
DAFTAR TABEL x
DAFTAR GAMBAR x
DAFTAR LAMPIRAN x
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
Ruang Lingkup Penelitian 2
METODE 2
Bahan dan Alat 2
Pengukuran Kondisi Lingkungan 3
Koleksi Ektomikoriza dari Kawasan Hutan Haurbentes 3
Pembersihan Sampel Akar 3
Identifikasi Morfotipe Ektomikoriza dan Analisis Data 3
Isolasi Cendawan Ektomikoriza 3
Pemurnian Isolat Cendawan 4
Analisis Morfologi 4
HASIL DAN PEMBAHASAN 4
Hasil 4
Kondisi Lingkungan Kawasan Hutan Haurbentes Jasinga 4
Analisis Morfotipe Ektomikoriza 5
Isolasi Ektomikoriza, Pemurnian, dan Analisis Morfologi Isolat Cendawan 6
Pembahasan 9
SIMPULAN DAN SARAN 12
Simpulan 12
Saran 12
DAFTAR PUSTAKA 13
DAFTAR TABEL
1 Jenis pohon dan kondisi lingkungan pengambilan sampel
ektomikoriza dari Shorea spp. di Kawasan Hutan Haurbentes ... 4
2 Persentase kolonisasi ektomikoriza Shorea spp. di Kawasan Hutan Haurbentes ... 6
3 Kelimpahan relatif morfotipe ektomikoriza Shorea spp. di Kawasan Hutan Haurbentes (Lampiran 1-5) ... 7
4 Analisis morfologi mikroskopis dan makroskopis isolat cendawan ektomikoriza Shorea spp. di Kawasan Hutan Haurbentes ... 8
DAFTAR GAMBAR
1 Morfotipe ektomikoriza yang berasal dari Kawasan Hutan Haurbentes Jasinga: (a) Irregular pinnate cokelat (b) Irregular pinnate hitam (c) Irregular pinnate perak (d) Monopodial pinnate cokelat (e) Monopodial pinnate hitam ... 52 Ciri mikroskopis isolat cendawan ektomikoriza hasil isolasi: (A) P14118a (B) P13211 (C) P14118a (D) P14118a (E dan F) P23157. Anastomosis (a), struktur hifa gelap (b), sekat hifa (c), klamidospora (d), dan percabangan hifa intensif (e dan f) ... 9
DAFTAR LAMPIRAN
1 Jumlah akar irregular pinnate cokelat per plot 152 Jumlah akar irregular pinnate hitam per plot 16
3 Jumlah akar irregular pinnate perak per plot 17
4 Jumlah akar monopidial pinnate cokelat per plot 18
5 Jumlah akar monopodial pinnate hitam per plot 19
PENDAHULUAN
Latar Belakang
Genus Shorea merupakan anggota dari famili Dipterocarpaceae. Anggota famili Dipterocarpaceae berjumlah kurang lebih 512 jenis yang termasuk ke dalam 16 genus dan 3 subfamili, yakni Dipterocarpaceae, Pakaraimoideae dan Monotoideae. Jenis Dipterocarpaceae umumnya berupa pohon menjulang (emergent trees) yang pertumbuhannya lambat. Kebanyakan jenis tersebut berupa pohon-pohon besar dengan tajuk yang sangat dominan (Al Rasyid et al. 1991).
Sebagian besar dari jenis Dipterocarpaceae terdapat pada daerah beriklim basah, memiliki kelembapan tinggi, memiliki ketinggian tempat 0 – 800 m dpl, serta curah hujan di atas 2.000 mm/th dengan rentang musim kemarau yang pendek. Jenis yang mampu tumbuh di ketinggian tempat di atas 800 m dpl sangat sedikit, di antaranya adalah anggota dari genus Shorea, Dipterocarpus dan Vatica. (Al Rasyid et al. 1991).
Sebagian besar Dipterocarpaceae menyukai hidup di tanah yang kering, masam, dan liat, meskipun sebagian kecil di antaranya dapat tumbuh di tanah dengan kondisi berkapur, berpasir, dan gambut. Pada kondisi tanah asam, Dipterocarpaceae seringkali berasosiasi dengan cendawan membentuk ektomikoriza sebagai cara bertahan hidup di lingkungan cekaman asam (Fajri 2008)
Ektomikoriza merupakan simbiosis mutualisme antara cendawan ektomikoriza dengan akar tumbuhan (Brundrett 2004). Simbiosis ektomikoriza dicirikan dengan terbentuk struktur tubuh buah jamur di luar akar, jalinan hifa yang menyelubungi akar tanaman (mantel), hifa eksternal untuk mengeksplorasi tanah yang berfungsi sebagai perpanjangan akar, dan jaringan hartig pada epidermis maupun korteks akar (Olsson et al. 2000). Pembentukan struktur ini menyebabkan bentuk akar menjadi khas, yakni pendek, menebal, dan memiliki warna yang berbeda karena diselubungi hifa atau mantel cendawan (Brundrett et al. 1996).
Ektomikoriza berperan penting dalam memacu pertumbuhan inang, di antaranya karena hifa eksternal berfungsi meningkatkan penyerapan unsur hara dan air untuk tanaman sehingga meningkatkan ketahanan terhadap kekeringan dan defisiensi hara. Selain itu, jaringan hifa mantel meningkatkan ketahanan tanaman terhadap serangan patogen (Smith dan Read 1997). Cendawan ektomikoriza memperoleh unsur karbon hasil fotosintesis tumbuhan inang dari proses pertukaran nutrisi antara cendawan dan tumbuhan inang di jaringan hartig (Smith dan Read 1997).
Ektomikoriza pada Shorea spp. ditemukan di hutan-hutan tropis di banyak negara. Amornpitak et al (2006) melaporkan keberadaan asiosiasi Theleporaceae sp. pada Shorea farinosa, Tomentella sp. pada Shorea roxburghii dan Shorea
guiso, Scleroderma bovista pada S. roxburghii, dan Lepiota sp. pada Shorea
obtusa di hutan Thailand. Sedangkan Nuhamara (1987) melaporkan keberadaan cendawan ektomikoriza pada Shorea javanica yaitu Amanita hemibapha, Cantharellus cibarius, Russula sp., Lactarius spp., dan Scleroderma sp.
sehingga struktur ini dapat digunakan untuk mengidentifikasi morfotipenya (Agerer 1996).
Ektomikoriza dapat digunakan sebagai pupuk hayati. Penelitian Martiningsih (2000) menunjukkan perlakuan ektomikoriza Pisolitus sp. terhadap
Shorea sp. memiliki perbedaan positif yang signifikan terhadap parameter
pertumbuhan, seperti tinggi tajuk, diameter batang, berat basah, dan nisbah pucuk akar. Namun, penggunaan ektomikoriza sebagai pupuk hayati untuk tanaman
Shorea di Indonesia belum maksimal. Hal ini karena biologi ektomikoriza
termasuk keragaman jenis cendawan ektomikoriza, dan keragaman morfotipe pada tanaman Dipeterocarpaceae belum dipelajari dengan baik, termasuk di Kawasan Hutan Haurbentes. Oleh karena itu biologi ektomikoriza Dipterocarpaceae di Kawasan Hutan Haurbentes perlu dipelajari.
Perumusan Masalah
Berdasarkan latar belakang penelitian, rumusan masalah yang akan digunakan pada penelitian ini yaitu masih sedikit penelitian yang melaporkan tentang data biologi ektomikoriza termasuk keragaman jenis cendawan ektomikoriza, dan keragaman morfotipe ektomikoriza pada tanaman Dipeterocarpaceae terutama di Kawasan Hutan Haurbentes Jasinga Bogor.
Tujuan Penelitian
Penelitian ini bertujuan untuk mempelajari morfotipe ektomikoriza pada Dipterocarpaceae khususnya Shorea, mengisolasi dan mengkarakterisasi cendawan ektomikoriza melalui analisis morfologi.
Manfaat Penelitian
Penelitian ini diharapkan mampu memberikan data mengenai keragaman cendawan ektomikoriza dan keragaman morfotipe ektomikoriza pada tanaman Dipeterocarpaceae. Data tersebut dapat digunakan sebagai bahan untuk penelitian lanjutan mengenai ektomikoriza pada Dipterocarpaceae di Kawasan Hutan Haurbentes.
Ruang Lingkup Penelitian
Ruang lingkup penelitian ini mencakup analisis morfotipe ektomikoriza, persentase kolonisasi masing-masing morfotipe pada setiap plot dan kelimpahan relatif morfotipe pada setiap plot. Adapun cakupan yang lain yaitu mengisolasi cendawan ektomikoriza dan mengkarakterisasinya secara makroskopis maupun mikroskopis.
METODE
Bahan dan Alat
Bahan yang digunakan adalah sampel akar Shorea yang bersimbiosis dan yang tidak bersimbiosis dengan cendawan ektomikoriza yang diperoleh dari Kawasan Hutan Haurbentes, dan medium Modified Merlin Nokrans (MMN). Alat–alat yang digunakan ialah autoklaf, inkubator, Laminar Air Flow Cabinet, mikroskop stereo, mikroskop cahaya, tabung 1.5 mL, dan peralatan laboratorium lain yang umum digunakan.
Pengukuran Kondisi Lingkungan
Parameter yang diukur pada lokasi pengambilan sampel meliputi suhu, kelembapan, dan intensitas cahaya.
Koleksi Ektomikoriza dari Kawasan Hutan Haurbentes Jasinga
Pengambilan sampel dilakukan dengan membuat plot berukuran 20 m x 20 m sebanyak 3 ulangan. Sebanyak 5 pohon Dipterocarpaceae dipilih secara acak dari setiap plot, kemudian pada masing-masing pohon ditentukan 4 titik pengambilan akar. Pengambilan akar dilakukan dengan metode perunutan akar. Sampel akar diambil dengan menggali akar utama sampai mendapat akar yang memiliki struktur ektomikoriza. Sampel akar yang diambil kemudian dimasukkan ke dalam wadah plastik dan diberi keterangan tiap plot pengambilan dan ulangannya sebagai identitas sampel.
Pembersihan Sampel Akar
Sampel akar diletakkan pada saringan plastik dan dicuci menggunakan air mengalir. Akar dipisahkan dari tanah dan dibersihkan dengan menggunakan kuas. Akar yang sudah dicuci dibagi ke dalam dua bagian, yaitu untuk analisis morfotipe dan isolasi cendawan.
Identifikasi Morfotipe Ektomikoriza dan Analisis Data
Analisis data morfotipe dihitung dengan metode grid line berukuran 1x1 cm, yaitu akar dipotong dengan ukuran 1 cm kemudian disebar secara acak ke dalam cawan grid line. Selanjutnya potongan akar yang sudah tersebar secara acak tersebut diamati dengan mikroskop stereo dan mikroskop cahaya, kemudian difoto dan dicatat. Sampel akar yang diketahui memiliki struktur ektomikoriza kemudian sebagian disimpan dalam formaldehid: asam asetat: alkohol= 1: 0.5: 5 (FAA). Masing-masing morfotipe kemudian dihitung. Persentase kolonisasi (K) dan kelimpahan relatif (KR) dihitung dengan menggunakan rumus:
K =
x 100%
KR =
x 100%
Isolasi Cendawan Ektomikoriza
Isolasi cendawan ektomikoriza merujuk pada metode Brundrett et al. (1996). Akar tumbuhan Dipterocarpaceae yang terkolonisasi cendawan ektomikoriza dipotong-potong dengan ukuran sekitar 0.5 cm sebanyak sepuluh potong untuk tiap morfotipe pada masing-masing sampel. Potongan akar kemudian disterilisasi permukaannya dengan diawali perendaman dalam air steril selama 15 menit sebanyak tiga kali. Akar kemudian direndam dalam etanol 75% selama 10 menit, direndam kembali dalam air steril selama 10 menit, direndam dalam larutan natrium hipoklorit (NaOCl) 1% selama 30 detik. Selanjutnya akar direndam kembali dengan air steril selama 5 menit dan dikeringkan dengan kertas tisu steril. Potongan akar kemudian diletakkan pada cawan berisi medium MMN steril yang mengandung antibiotik chloramphenicol (500 mg/liter), selanjutnya cawan diinkubasi pada suhu ruang hingga tumbuh hifa cendawan.
Pemurnian Isolat Cendawan
Hifa cendawan yang tumbuh dari akar Dipterocarpaceae, dimurnikan pada medium MMN yang mengandung antibiotik chloramphenicol hingga diperoleh kultur murni cendawan.
Analisis Morfologi
Pengamatan morfologi terhadap cendawan dilakukan menggunakan metode Riddle (1950), yaitu menumbuhkan cendawan pada potongan MMN yang diletakkan pada bagian tengah kaca objek steril. Kaca objek yang telah diberi kaca penutup dimasukkan ke dalam cawan petri steril yang mengandung kertas saring steril yang lembap dan diinkubasi selama 15-30 hari. Kaca penutup yang ditumbuhi cendawan dipindahkan ke kaca objek steril yang telah ditetesi larutan shears. Morfologi cendawan diamati dengan mikroskop cahaya pada perbesaran 400 dan 1000 kali.
HASIL DAN PEMBAHASAN
Hasil
Kondisi Lingkungan Kawasan Hutan Haurbentes Jasinga
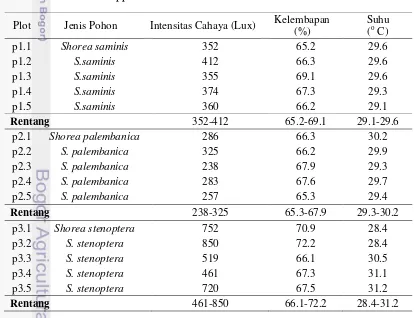
Suhu rata-rata seluruh plot pada saat pengambilan sampel adalah 29.7oC dengan suhu terendah adalah 28.4oC dan suhu tertinggi 31.2oC (Tabel 1).
Tabel 1 Jenis pohon dan kondisi lingkungan pengambilan sampel ektomikoriza dari Shorea spp. di Kawasan Hutan Haurbentes
Plot Jenis Pohon Intensitas Cahaya (Lux) Kelembapan
(%)
Rentang 352-412 65.2-69.1 29.1-29.6
p2.1 Shorea palembanica 286 66.3 30.2
p2.2 S. palembanica 325 66.2 29.9
p2.3 S. palembanica 238 67.9 29.3
p2.4 S. palembanica 283 67.6 29.7
p2.5 S. palembanica 257 65.3 29.4
Rentang 238-325 65.3-67.9 29.3-30.2
p3.1 Shorea stenoptera 752 70.9 28.4
p3.2 S. stenoptera 850 72.2 28.4
p3.3 S. stenoptera 519 66.1 30.5
p3.4 S. stenoptera 461 67.3 31.1
p3.5 S. stenoptera 720 67.5 31.2
Rentang 461-850 66.1-72.2 28.4-31.2
Pemilihan pohon dilakukan secara acak sesuai pemilihan tempat plot. Sampel akar diambil dari 3 jenis Shorea, yakni S. saminis, S. palembanica, dan S.
stenoptera Burck. Masing-masing plot memiliki faktor kondisi lingkungan yang
berbeda. Plot 3 memiliki intensitas paparan cahaya yang paling tinggi, sedangkan plot 2 memiliki intensitas paparan cahaya paling rendah. Kelembapan tertinggi terdapat pada plot 3, sedangkan kelembapan terendah pada plot 1.
Analisis Morfotipe Ektomikoriza
Morfologi ektomikoriza berdasarkan Colour Atlas of Ectomycorrhizae (Agerer 1996), menunjukkan bahwa potongan akar yang berasal dari tiga plot pengamatan tergolong ke dalam 5 tipe percabangan morfotipe utama, yaitu
irregular pinnate hitam, irregular pinnate cokelat, irregular pinnate perak,
monopodial pinnate cokelat, dan monopodial pinnate hitam (Gambar 1).
Gambar 1 Morfotipe ektomikoriza yang berasal dari Kawasan Hutan Haurbentes Jasinga: (a) Irregular pinnate cokelat (b) Irregular pinnate hitam, (c)
Irregular pinnate perak, (d) Monopodial pinnate cokelat, (e)
Monopodial pinnate hitam.
Persentase kolonisasi cendawan ektomikoriza pada seluruh sampel ditemukan kurang dari 60%, sehingga akar yang tidak terkolonisasi memiliki porsi yang lebih kecil pada perakaran. Masing-masing plot memiliki persentase kolonisasi yang berbeda. Persentase kolonisasi tertinggi ditemukan pada plot 3 (59,5%), diikuti plot 2 (53,7%), dan plot 1 (49,4%).
Persentase kolonisasi pada masing-masing plot menunjukkan komposisi morfotipe yang berbeda. Persentase kolonisasi morfotipe pada plot 1 dan 2 memiliki pola urutan yang serupa, yakni irregular pinnate cokelat sebagai morfotipe dengan persentase kolonisasi tertinggi (37,5% dan 36,6%), diikuti dengan irregular pinnate hitam, monopodial pinnate cokelat, dan irregular
pinnate perak. Morfotipe monopodial pinnate hitam tidak ditemukan pada plot 1
dan 2, hanya ditemukan pada plot 3 (Tabel 2)
5
a b
e d
Tabel 2 Persentase kolonisasi ektomikoriza Shorea spp. di Kawasan Hutan Haurbentes
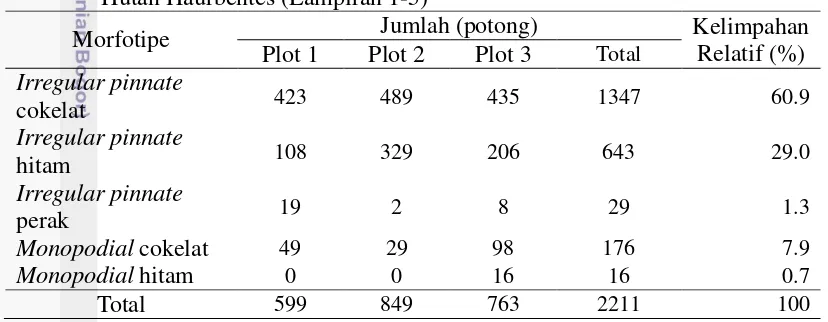
Sebanyak 2211 potong akar ektomikoriza berhasil diamati dari 60 sampel yang berasal dari tiga plot pengamatan. Jumlah akar ektomikoriza paling banyak terdapat pada plot pengamatan 2 dan jumlah ektomikoriza yang paling sedikit terdapat pada plot 1 (Tabel 3). Morfotipe Irregular pinnate cokelat memiliki kelimpahan relatif tertinggi yaitu sebesar 60.9% (1347 potong akar). Sedangkan morfotipe dengan kelimpahan relatif terendah yaitu monopodial hitam sebesar 0.7% (16 potong akar) (Tabel 3). Kelimpahan relatif morfotipe pada masing-masing plot memiliki pola yang serupa dengan persentase kolonisasinya.
Tabel 3 Kelimpahan relatif morfotipe ektomikoriza Shorea spp. di Kawasan Hutan Haurbentes (Lampiran 1-5)
Morfotipe Jumlah (potong) Kelimpahan
Relatif (%)
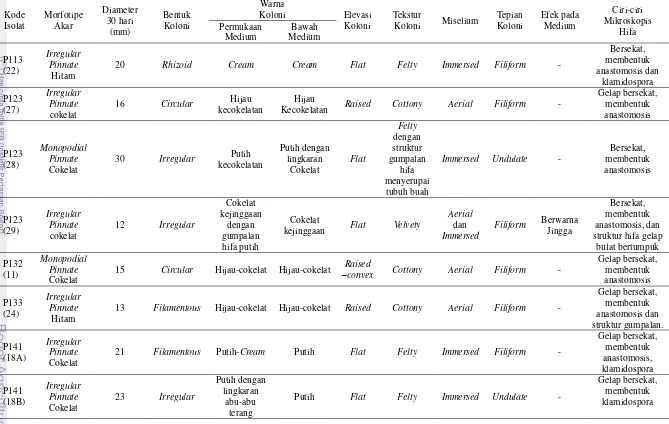
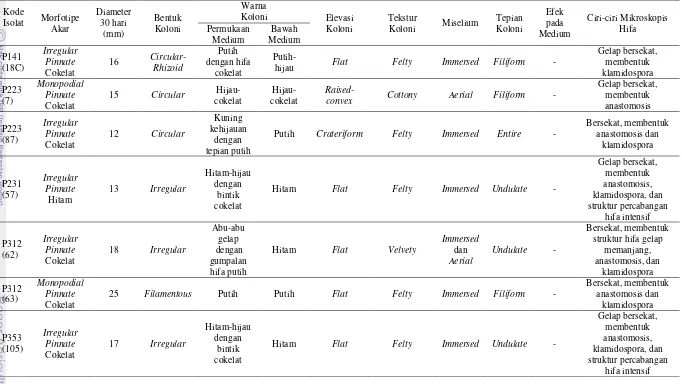
Isolasi Ektomikoriza, Pemurnian, dan Analisis Morfologi Isolat Cendawan Tingkat pertumbuhan hifa cendawan ektomikoriza dari akar yang terkolonisasi pada medium sintetis sangat rendah (3.5%), begitu pula tingkat keberhasilan pemurniannya, sehingga hanya diperoleh 15 isolat cendawan (Lampiran 7). Hasil pengamatan menunjukkan, cendawan yang diperoleh memiliki ciri-ciri makroskopis yang berbeda, namun keseluruhan isolat merupakan cendawan yang tidak bersporulasi (mycelia sterilia) (Tabel 4). Karakterisasi koloni cendawan mengacu pada Lampiran 6.
Pengamatan mikroskopis menunjukkan masing-masing isolat memiliki ciri yang berbeda, namun sebagian besar memiliki hifa gelap, septat (bersekat), membentuk klamidospora dan membentuk jembatan antar hifa (anastomosis). Beberapa di antara isolat cendawan memiliki struktur percabangan hifa intensif (Tabel 4, Gambar 2, Lampiran 7).
Morfotipe Persentase Kolonisasi Per plot (%)
1 2 3
Irregular pinnate cokelat 37.5 36.6 29.3
Kode
Kecokelatan Raised Cottony Aerial Filiform -
Gelap bersekat,
kejinggaan Flat Velvety
Aerial
15 Circular Hijau-cokelat Hijau-cokelat –Raised
convex Cottony Aerial Filiform -
13 Filamentous Hijau-cokelat Hijau-cokelat Raised Cottony Aerial Filiform -
Gelap bersekat,
Tabel 4 Analisis morfologi mikroskopis dan makroskopis isolat cendawan ektomikoriza Shorea spp. Kawasan Hutan Haurbentes
Kode
Isolat Morfotipe Akar
15 Circular
Hijau-cokelat struktur hifa gelap
memanjang,
Tabel 4 Lanjutan Analisis morfologi mikroskopis dan makroskopis isolat cendawan ektomikoriza Shorea spp. Kawasan Hutan Haurbentes
Pembahasan
Kawasan Hutan Haurbentes Jasinga merupakan hutan hujan tropis dataran rendah (250 m dpl). Kawasan ini memiliki curah hujan rata-rata sebesar 4276 mm per tahun. Jenis tanah yang mendominasi ialah podsolik merah kuning, regosol, dan acid brown forest soil. Sebanyak 13 jenis Shorea, 5 jenis Hopea, 2 jenis Dipterocarpus, dan 1 jenis Anisoptera telah ditanam di Haurbentes sampai dengan tahun 1997, seluruhnya telah beradaptasi yang ditandai dengan telah terjadinya regenerasi secara alami (Balitbanghut 2007). Pohon-pohon S. stenoptera di Haurbentes tumbuh dari benih yang berasal dari Pontianak, Kalimantan Barat sekitar 40 tahun yang lalu (Suzuki dan Gadrinab 1989).
Hasil karakterisasi morfologi ektomikoriza berdasarkan Colour Atlas of Ectomycorrhizae (Agerer 1996), menunjukkan bahwa ke-2211 potong akar tergolong ke dalam 5 tipe ramifikasi morfotipe utama, yaitu irregular pinnate hitam, irregular pinnate cokelat, irregular pinnate perak, monopodial pinnate cokelat, dan monopodial pinnate hitam dengan persentase yang berbeda pada tiap plot sampel (Gambar 1).
Persentase kolonisasi per plot menunjukkan bahwa kolonisasi oleh cendawan ektomikoriza pada akar relatif cukup besar, yakni mendekati 60%. Hal ini terjadi karena kondisi lingkungan Haurbentes cocok untuk pertumbuhan 9
optimal ektomikoriza. Faktor yang mempengaruhi perkembangan cendawan ektomikoriza pada rizosfer di antaranya faktor suhu dan kondisi tanah. Cendawan ektomikoriza berkembang dengan baik dalam kondisi tanah yang masam dan lembap. Faktor suhu juga berperan penting dalam perkembangan cendawan ektomikoriza. Cendawan ektomikoriza memiliki suhu optimum yang berbeda tiap jenisnya, namun 80% Pisolithus sp. mampu berkembang pada suhu 34 oC (Hadi 1999).
Isolat cendawan yang berhasil dimurnikan hanya berasal dari potongan akar ektomikoriza dengan morfotipe irreguler pinnate cokelat, irregular pinnate hitam
dan monopodial pinnate cokelat. Masing-masing potongan akar bermorfotipe
sama menghasilkan isolat cendawan dengan karakteristik morfologi yang berbeda, baik secara makroskopis maupun mikroskopis.
Perbedaan karakteristik isolat cendawan yang tumbuh dari potongan akar bermorfotipe utama yang sama dapat terjadi karena perbedaan ciri-ciri dalam taraf khusus yakni bentuk ujung akar, bentuk khas permukaan mantel, dan bentuk
rhizomorf atau hifa yang berada di sekitar akar ektomikoriza. Struktur yang
berbeda pada akar ektomikoriza dapat saja menunjukkan bahwa cendawan yang bersimbiosis dengan akar tersebut berbeda (Agerer1996). Kondisi lingkungan abiotik seperti suhu, intensitas cahaya dan kelembapan mempengaruhi ramifikasi, persentase ramifikasi akar, dan struktur khusus ektomikoriza (Santoso 1997).
Identifikasi morfotipe akar ektomikoriza merupakan metode estimasi langsung mengenai kekayaan dan keragaman spesies cendawan ektomikoriza yang bersimbiosis tanpa bergantung pada tubuh buah yang ditemukan. Tubuh buah cendawan ektomikoriza biasanya ditemukan dalam jumlah yang sangat sedikit sehingga tidak efektif dalam penelitian mengenai kekayaan dan diversitas cendawan ektomikoriza (Moser et al. 2005).
Becerra et al. (2005) melaporkan beberapa genus cendawan berdasarkan morfotipenya pada tanah asam. Morfotipe irregular pinnate yang ditemukan merupakan hasil simbiosis akar Alnus acuminata dengan cendawan Tomentella sp., Russula sp., dan Alnirhiza sp. Morfotipe monopodial pinnate merupakan simbiosis dengan cendawan Naucoria sp. dan Lactarius sp. Selanjutnya morfotipe yang menyerupai irregular pinnate-monopodial pinnate ditemukan pada simbiosis cendawan Gyrodon sp. dan Tomentella sp. dengan akar Quercus garryana. Tomentella sp. memiliki morfotipe monopodial pinnate berwarna hitam dengan ujung putih (Moser et al. 2009).
Informasi mengenai simbiosis ektomikoriza pada akar Shorea spp. di Kawasan Hutan Haurbentes dilaporkan oleh Sukarno et al. (2013). Berdasarkan analisis morfotipe dan molekuler diperoleh 50 morfotipe ektomikoriza yang terdiri dari 43 cluster cendawan. Beberapa morfotipe tersebut di antaranya unramified hitam dikolonisasi oleh Pleurotus ostreatus, dan Russula sp. Morfotipe monopodial pinnate coklat, irregular pinnate cokelat, dan irregular pinnate hitam dikolonisasi oleh Russula sp.
(2006) yang menyebutkan bahwa ektomikoriza yang umum ditemukan pada hutan-hutan tua merupakan jenis dari genus Russula, Amanita, dan Cortinarius. Namun demikian, dugaan perbandingan tersebut diperlukan analisis lanjutan berupa analisis molekuler terhadap akar-akar yang terkolonisasi tersebut.
Isolat cendawan yang berhasil dimurnikan sebagian besar berasal dari plot 1, yakni 9 isolat. Isolat cendawan yang berhasil dimurnikan yang berasal dari plot 2 dan plot 3 masing-masing 3 isolat cendawan. Hal ini mungkin terjadi akibat lama penyimpanan yang berbeda sebelum diisolasi. Pengerjaan sampel dilakukan secara berurut, sehingga identifikasi morfotipe akar dan isolasi akar pada plot 1 dilakukan dalam kondisi akar yang lebih segar daripada plot 2 dan plot 3.
Lama waktu penyimpanan sampel akar sebelum diisolasi memungkinkan cendawan ektomikoriza menjadi rusak atau cendawan tidak mampu tumbuh secara optimal (Smith dan Read 1997). Selain itu, cendawan non ektomikoriza yang tumbuh di sekitar ektorizosfer diduga memiliki kemampuan saprofitik lebih tinggi sehingga dapat bersaing dengan cendawan ektomikoriza ketika ditumbuhkan pada medium MMN, walaupun telah dilakukan sterilisasi permukaan (Marx dan Kenny 1982). Hal ini diduga menjadi penyebab tingkat keberhasilan isolasi yang rendah.
Sebagian besar ciri mikroskopis isolat cendawan yang berhasil diisolasi ialah hifa bersekat, tidak memiliki sambungan apit, membentuk jembatan antar hifa (anastomosis) dan membentuk klamidospora. Beberapa di antara isolat cendawan memiliki hifa gelap berwarna cokelat kehitaman. Nurhayat (2012) berhasil mengisolasi dan mengidentifikasi cendawan ektomikoriza pada pohon pinus secara morfologi dan molekuler yakni Cenococcum. Karakteristik morfologi baik secara mikroskopis maupun makroskopis cendawan Cenococcum memiliki kesamaan dengan beberapa isolat cendawan yang berhasil diisolasi pada penelitian ini, yakni isolat dengan kode P132(11) dan P223(7), meskipun morfotipe akarnya berbeda. Karakteristik Cenococcum yakni miselium berwarna hitam, memiliki diameter pertumbuhan miselium yang relatif lambat, hifa berwarna cokelat kehitaman, hifa septat, tidak memiliki sambungan apit (clamp connection), dan terdapat fusi antar hifa atau anastomosis. Berdasarkan kesamaan morfologi ini, terdapat kemungkinan bahwa isolat cendawan tersebut memiliki genus yang sama yakni Cenococcum, namun masih membutuhkan identifikasi molekuler untuk memastikannya.
Cendawan ektomikoriza termasuk cendawan yang sulit ditumbuhkan diluar habitat aslinya. Banyak cendawan ektomikoriza memiliki pertumbuhan yang sangat rendah pada medium buatan. Medium MMN merupakan medium khusus yang digunakan untuk mengisolasi cendawan yang sulit ditumbuhkan di luar habitat alamiahnya seperti cendawan ektomikoriza (Brundrett et. al. 1996). Medium MMN merupakan medium kaya nutrisi yang menunjang pertumbuhan cendawan ektomikoriza.
SIMPULAN DAN SARAN
Simpulan
Sebanyak 2211 potong akar ektomikoriza berhasil diamati dari 60 sampel yang berasal dari tiga plot pengamatan. Jumlah akar ektomikoriza paling banyak terdapat pada plot pengamatan 2 dan jumlah ektomikoriza yang paling sedikit terdapat pada plot 1. Hasil karakterisasi morfologi ektomikoriza menunjukkan bahwa terdapat 5 ramifikasi morfotipe utama ektomikoriza pada Shorea spp. di Kawasan Hutan Haurbentes yaitu Irregular pinnate cokelat, Irregular pinnate hitam, Irregular pinnate perak, monopodial pinnate cokelat, dan monopodial
pinnate hitam. Morfotipe irregular pinnate cokelat merupakan tipe yang
mendominasi sampel akar ektomikoriza pada setiap plot dengan persentase sebesar 60.9 % Sebanyak 15 isolat berbeda diisolasi dari morfotipe ektomikoriza. Seluruh isolat tersebut tidak bersporulasi. Beberapa cendawan memiliki karakteristik miselium khusus dengan memproduksi anastomosis dan menghasilkan percabangan miselium intensif.
Saran
DAFTAR PUSTAKA
Agerer R. 1996. Colour Atlas of Ectomycorrhizae. Schwabish Gmund (DE): Einhorn-Verlag.
Al Rasyid H, Marfuah H, Wijayakusuma, Hendarsyah D. 1991. Vademikum
Dipterocarpaceae. Jakarta (ID): Badan Penelitian dan Pengembangan
Departemen Kehutanan.
Amornpitak TW, Vichitsoonthonkul T, Tanticharoen M, Cheevadhanarak S, Ratchadawong S. 2006. Diversity of Ectomycorrhyzal Fungi of
Dipterocarpaceae in Thailand. J. Bio. Sci 6 (6): 1059-1064.
Ashton PS. 1982. Dipterocarpaceae. In: Van Steenis, C.G.G.J (ed.) Flora Malesiana (9): 237-552.
[Balitbanghut] Badan Penelitan dan Pengembangan Kehutanan. 2007. Kawasan Hutan dengan Tujuan Khusus Haurbentes. [internet]. Bogor: [diunduh 2013 Des 7]. Tersedia pada: http://www.dephut.go.id/index.php/news/details/951. Becerra A et al. 2005. Ectomycorrhizal colonization of Alnus acumiata Kunth in northwestern Argentina in relation to season and soil parameters. Ann. For. Sci 65: 325-332.
Brundrett M. 2004. Diversity and Classification of Mycorrhizal Associations.
Biol.Rev 79:473–495.
Brundrett M, Bougher N, Dell B, Grove T, Malajczuk N. 1996. Working With Mycorrhizae in Forestry and Agriculture. Canberra (AU): Monograph ACIAR.
Fajri M. 2008. Pengenalan Umum Dipterocarpaceae, Kelompok Jenis Bernilai Ekonomi Tinggi. Inf. Tek. Dipter 2(1): 9-21.
Hadi S. 1999. Status ektomikoriza pada tanaman hutan di Indonesia. Prosiding Seminar Nasional Mikoriza I; [Bogor 15-16 Nov 1999] Bogor (ID): Asosiasi Mikoriza Indonesia.
Martiningsih SH. 2000. Pengaruh Bio-Stimulant dan Inokulasi Cendawa Ektomikoriza Pisolithus tinctorius Coker and Couch Terhadap Pertumbuhan Semai Shorea leprosula Miq [skripsi]. Bogor (ID). Institut Pertanian Bogor. Marx DH, Kenney DS. 1982. Production of ectomycorrhizal fungus inoculum.
Schenk NC, editor. Minnesota (US): The American Phytopathological. Moser AM, Petersen CA, D’Allura JA, Southworth D. 2005 Comparison of
ectomycorrhyzas of Quercus garryana (Fagaceae) on serpentine and non-serpentine soils in southwestern Oregon. Am. Jour. Bot 92 (2): 224-230.
Moser AM, Frank JL, D’Allura JA, Southworth D. 2009. Ectomycorrhizal
communities of Quercus garryana are similar on serpentine and nonserpentine soils. Plant Soil 305: 185-194.
Nara K. 2006. Ectomycorrhizal network and seedling establishment during early primary succesion. New Phyt 169: 169-78.
Nuhamara ST. 1987. Mycorrhizae in Agroforestry: A Case Study. Biotropia 1(1): 53-57.
Nurhayat OD. 2012. Ektomikoriza Pinus merkusii di Bagian Kesatuan Pemangku Hutan (BKPH) Lembang [skripsi]. Bogor (ID): Institut Pertanian Bogor. Olsson PA, Munzenberger B, Mahmood S, Erland S. 2000. Molecular and
the ectomycorrhizal fungi Suillus bovinus and Gomphidius roseus. J. Mycol 16 (1): 8-9.
Riddle RW. 1950. Permanent stained mycological preparation obtained by slide culture. Mycol Res 42:265-270.
Santoso E. 1997. Hubungan Perkembangan Ektomikoriza dengan Populasi Jasad Renik dalam Rizosfer dan Pengaruhnya Terhadap Pertumbuhan Eucalyptus
pellita dan Eucalyptus urophylla [tesis]. Bogor (ID): Program Pascasarjana
Institut Pertanian Bogor.
Smith SE, Read DJ. 1997. Mychorrhizal symbiosis. Second edition. London (GB): Academic Press.
Sukarno N, Listiyowati S, Nurcahyo OD, Retnowati A, Nara K 2013. Diversity of
Shorea ectomycorrhyza. Proceeding of International Conference on
Mycorrhiza; [Sendai 16 Nov 2013] Sendai,(JP): Tohoku University.
LAMPIRAN
Lampiran 1 Jumlah Akar irregular pinnate cokelat per plot PLOT 1 JUMLAH
AKAR PLOT 2
JUMLAH
AKAR PLOT 3
JUMLAH AKAR
P111 15 P211 6 P311 10
P112 0 P212 12 P312 0
P113 32 P213 38 P313 16
P114 34 P214 10 P314 32
Subtotal 81 Subtotal 66 Subtotal 58
P121 21 P221 9 P321 12
P122 20 P222 0 P322 33
P123 34 P223 23 P323 35
P124 28 P224 11 P324 34
Subtotal 103 Subtotal 43 Subtotal 114
P131 12 P231 28 P331 27
P132 23 P232 0 P332 22
P133 40 P233 42 P333 17
P134 17 P234 9 P334 15
Subtotal 92 Subtotal 79 Subtotal 81
P141 25 P241 29 P341 21
P142 26 P242 36 P342 27
P143 9 P243 26 P343 48
P144 24 P244 17 P344 0
Subtotal 84 Subtotal 108 Subtotal 96
P151 12 P251 39 P351 16
P152 25 P252 20 P352 24
P153 22 P253 63 P353 34
P154 4 P254 71 P354 12
Subtotal 63 Subtotal 193 Subtotal 86
Total 423 Total 489 Total 435
Lampiran 2 Jumlah akar irregular pinnate hitam per plot PLOT 1 JUMLAH
AKAR PLOT 2
JUMLAH
AKAR PLOT 3
JUMLAH AKAR
P111 15 P211 0 P311 0
P112 13 P212 5 P312 28
P113 18 P213 20 P313 0
P114 9 P214 0 P314 12
Subtotal 55 Subtotal 25 Subtotal 40
P121 0 P221 0 P321 7
P122 0 P222 21 P322 17
P123 0 P223 10 P323 31
P124 0 P224 0 P324 20
Subtotal 0 Subtotal 31 Subtotal 75
P131 0 P231 29 P331 0
P132 13 P232 38 P332 0
P133 16 P233 17 P333 13
P134 0 P234 0 P334 21
Subtotal 29 Subtotal 84 Subtotal 34
P141 0 P241 49 P341 13
P142 10 P242 15 P342 0
P143 0 P243 51 P343 0
P144 0 P244 31 P344 29
Subtotal 10 Subtotal 146 Subtotal 42
P151 0 P251 5 P351 0
P152 9 P252 5 P352 15
P153 5 P253 29 P353 0
P154 0 P254 4 P354 0
Subtotal 14 Subtotal 43 Subtotal 15
Total 108 Total 329 Total 206
Lampiran 3 Jumlah akar irregular pinnate perak per plot PLOT 1 JUMLAH
AKAR PLOT 2
JUMLAH
AKAR PLOT 3
JUMLAH AKAR
P111 0 P211 0 P311 0
P112 0 P212 2 P312 0
P113 12 P213 0 P313 0
P114 0 P214 0 P314 0
Subtotal 12 Subtotal 2 Subtotal 0
P121 0 P221 0 P321 0
P122 0 P222 0 P322 8
P123 0 P223 0 P323 0
P124 0 P224 0 P324 0
Subtotal 0 Subtotal 0 Subtotal 8
P131 0 P231 0 P331 0
P132 0 P232 0 P332 0
P133 7 P233 0 P333 0
P134 0 P234 0 P334 0
Subtotal 7 Subtotal 0 Subtotal 0
P141 0 P241 0 P341 0
P142 0 P242 0 P342 0
P143 0 P243 0 P343 0
P144 0 P244 0 P344 0
Subtotal 0 Subtotal 0 Subtotal 0
P151 0 P251 0 P351 0
P152 0 P252 0 P352 0
P153 0 P253 0 P353 0
P154 0 P254 0 P354 0
Subtotal 0 Subtotal 0 Subtotal 0
Total 19 Total 2 Total 8
Lampiran 4 Jumlah akar monopodial pinnate cokelat PLOT 1 JUMLAH
AKAR PLOT 2
JUMLAH
AKAR PLOT 3
JUMLAH AKAR
P111 0 P211 0 P311 0
P112 0 P212 0 P312 9
P113 0 P213 0 P313 0
P114 0 P214 0 P314 0
Subtotal 0 Subtotal 0 Subtotal 9
P121 10 P221 0 P321 6
P122 0 P222 3 P322 0
P123 16 P223 7 P323 8
P124 0 P224 0 P324 0
Subtotal 26 Subtotal 10 Subtotal 14
P131 0 P231 0 P331 4
P132 9 P232 10 P332 4
P133 0 P233 0 P333 0
P134 4 P234 0 P334 12
Subtotal 13 Subtotal 10 Subtotal 20
P141 0 P241 0 P341 24
P142 0 P242 8 P342 0
P143 0 P243 0 P343 15
P144 10 P244 0 P344 10
Subtotal 10 Subtotal 8 Subtotal 49
P151 0 P251 0 P351 0
P152 0 P252 1 P352 6
P153 0 P253 0 P353 0
P154 0 P254 0 P354 0
Subtotal 0 Subtotal 1 Subtotal 6
Total 49 Total 29 Total 98
Lampiran 5 Jumlah akar monopodial pinnate hitam PLOT 1 JUMLAH
AKAR PLOT 2
JUMLAH
AKAR PLOT 3
JUMLAH AKAR
P111 0 P211 0 P311 0
P112 0 P212 0 P312 0
P113 0 P213 0 P313 0
P114 0 P214 0 P314 0
Subtotal 0 Subtotal 0 Subtotal 0
P121 0 P221 0 P321 0
P122 0 P222 0 P322 0
P123 0 P223 0 P323 6
P124 0 P224 0 P324 9
Subtotal 0 Subtotal 0 Subtotal 15
P131 0 P231 0 P331 1
P132 0 P232 0 P332 0
P133 0 P233 0 P333 0
P134 0 P234 0 P334 0
Subtotal 0 Subtotal 0 Subtotal 16
P141 0 P241 0 P341 0
P142 0 P242 0 P342 0
P143 0 P243 0 P343 0
P144 0 P244 0 P344 0
Subtotal 0 Subtotal 0 Subtotal 0
P151 0 P251 0 P351 0
P152 0 P252 0 P352 0
P153 0 P253 0 P353 0
P154 0 P254 0 P354 0
Subtotal 0 Subtotal 0 Subtotal 0
Total 0 Total 0 Total 16
Lampiran 7 Mikroskopis dan makroskopis isolat hasil isolasi Kode
Isolat Morfotipe Mikroskopis Makroskopis
P113 (22)
Irregular pinnate cokelat
P123 (27)
Irregular Pinnate cokelat
P123 (28)
Monopodial Pinnate Cokelat
P123 (29)
Irregular Pinnate cokelat
P132 (11)
Monopodial pinnate Cokelat
Lampiran 7 Lanjutan Mikroskopis dan makroskopis isolat hasil isolasi Kode
Isolat Morfotipe Mikroskopis Makroskopis
P134 (24)
Irregular Pinnate hitam
P141 (18A)
Irregular Pinnate Cokelat
P141 (18B)
Irregular Pinnate Cokelat
P141 (18C)
Irregular Pinnate Cokelat
P223 (7)
Lampiran 7 Lanjutan Mikroskopis dan makroskopis isolat hasil isolasi Kode
Isolat Morfotipe Mikroskopis Makroskopis
P223 (87)
Irregular Pinnate Cokelat
P231 (57)
Irregular Pinnate Hitam
P312 (62)
Irregular Pinnate Cokelat
P312 (63)
Irregular Pinnate Cokelat
P353 (105)
Irregular Pinnate Cokelat
RIWAYAT HIDUP
Penulis dilahirkan di Deli Serdang pada tanggal 16 April 1991 dari ayah Dr Ir. H Masri Sitanggang dan ibu Dra.Khairita Thamrin. Penulis merupakan anak ke 4 dari 10 bersaudara. Penulis menyelesaikan pendidikannya di SDN 101764 Tembung 2003, SMPN 1 P.S. Tuan pada tahun 2006, SMAN 11 Medan pada tahun 2009. Tahun yang sama penulis lulus seleksi masuk Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Masuk IPB (USMI) di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam.
Selama mengikuti perkuliahan, penulis pernah menjadi asisten praktikum mata kuliah Biologi Cendawan pada tahun ajaran 2013/2014. Penulis aktif di beberapa organisasi, di antaranya sebagai Kepala Departemen Pendidikan Bina Desa FMIPA pada tahun 20010-2011 dan Ketua Umum Himpunan Mahasiswa Biologi pada tahun 2011-2012.
Tahun 2010, penulis melakukan Studi Lapang di Hutan Pendidikan Gunung Walat Sukabumi Jawa Barat dengan judul laporan “Ekologi Goa Kapur Hutan Pendidikan Gunung Walat”. Tahun 2012, penulis melakukan Praktik Kerja Lapang di PT. Sinar Pusaka Lestari dari bulan Juli sampai bulan Agustus dengan
judul laporan “Pemantauan Kualitas Air Kolam Tambak Udang Vaname
(Litopenaeus vannamei) di PT. Sinar Pusaka Lestari ”.
Sebagai salah satu syarat kelulusan studi di Fakultas Matematika dan Ilmu
Pengetahuan Alam, penulis melaksanakan penelitian yang berjudul “Ektomikoriza
pada Shorea spp. di Kawasan Hutan Haurbentes Jasinga Bogor” di bawah bimbingan Ibu Dr Ir Nampiah Sukarno dan Ibu Dr Sri Listiyowati, MSi.