PENGENDALIAN FOLIKULOGENESIS OVARIUM MENCIT
(Mus musculus) DENGAN PEMBERIAN EKSTRAK BIJI
KAPAS (Gossypium hirsutum)
SISKA ADELYA RAMADHANI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Pengendalian Folikulogenesis Ovarium Mencit (Mus musculus) dengan Pemberian Ekstrak Biji Kapas (Gossypium hirsutum) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Desember 2016
Siska Adelya Ramadhani
RINGKASAN
SISKA ADELYA RAMADHANI. Pengendalian Folikulogenesis Ovarium Mencit (Mus musculus) dengan Pemberian Ekstrak Biji Kapas (Gossypium
hirsutum). Dibimbing oleh IMAN SUPRIATNA, NI WAYAN KURNIANI
KARJA, dan ADI WINARTO.
Gosipol adalah zat yang terkandung dalam ekstrak biji kapas dan diduga mempunyai kemampuan antifertilitas sehingga dapat digunakan sebagai bahan herbal kontrasepsi. Penelitian ini bertujuan mengkaji gambaran folikulogenesis serta perubahan morfologi ovarium dan siklus estrus mencit setelah pemberian ekstrak biji kapas. Sebanyak 60 ekor mencit betina DDY induk berumur 14-15 minggu dengan bobot badan 30-35 g dibagi menjadi empat kelompok dan diberi ekstrak biji kapas dengan konsentrasi 0; 1.5; 2.1; dan 2.7 g kg-1 BB (15 ekor per grup) selama 5, 10, 15, 24, dan 34 hari (3 ekor per periode grup) pada fase proestrus. Status estrus mencit dievaluasi berdasarkan gambaran sel dari ulas vagina. Pada setiap akhir periode perlakuan mencit di euthanasia untuk mengamati histomorfologi perkembangan folikel dan morfologi ovarium.
Hasil penelitian menunjukkan bahwa pemberian ekstrak biji kapas memperpanjang durasi siklus estrus. Panjang, lebar, dan bobot ovarium relatif sama antar grup (P > 0.05). Jumlah folikel tersier menurun seiring peningkatan lama waktu pemberian (P < 0.05). Jumlah folikel berkembang menurun setelah pemberian ekstrak biji kapas. Pemulihan jumlah folikel paling baik adalah dosis 1.5 g kg-1 BB pada mencit yang diberikan perlakuan ekstrak biji kapas. Data tersebut mengindikasikan bahwa efek pemberian ekstrak biji kapas selama 24 hari bersifat reversible.
SUMMARY
SISKA ADELYA RAMADHANI. Ovarian Folliculogenesis Control in Mice (Mus musculus) Treated by the Cottonseed Extract (Gossypium hirsutum). Supervised by IMAN SUPRIATNA, NI WAYAN KURNIANI KARJA, and ADI WINARTO.
Gossypol is a substance in cottonseed extract that has been thought commonly used as a herbal contraceptive due to its’ antifertility ability. The aim of the study were to assess the histomorphology of folliculogenesis, ovarian morphological exchange, and estrous cycle in cottonseed extract administrated mice. Sixty female mice DDY in 14-15 weeks old and 30-35 g body weight were randomly divided into four groups and then orally administrated with cottonseed extract in concentration of 0; 1.5; 2.1; and 2.7 g kg-1 BW (15 per group) for 5, 10, 15, 24, and 34 days (three per group period) at proestrus stage. The estrous status of mice was assessed during treatment period based on the description of the vaginal smear cells. At the end of the treatment period, all of each group was euthanized to observe of its morphological and histomorphological follicular development.
The results showed that administration of cottonseed extract prolonged the estrous cycle duration. The length, width, and weight of ovarium were similar among the group (P > 0.05). Whereas the number of tertiery follicle decreased as long as increasing of the period of the administration (P < 0.05). The number of developing follicles as long as increasing in cottonseed extract administration mice lower than control. The best follicle development recovery rate was found in 1.5 g kg-1 BW cottonseed administrated mice. These data indicated that effects of cottonseed extract administration for 24 days were reversible.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biologi Reproduksi
PENGENDALIAN FOLIKULOGENESIS OVARIUM MENCIT
(Mus musculus) DENGAN PEMBERIAN EKSTRAK BIJI
KAPAS (Gossypium hirsutum)
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Judul Tesis : Pengendalian Folikulogenesis Ovarium Mencit (Mus musculus) dengan Pemberian Ekstrak Biji Kapas (Gossypiumhirsutum) Nama : Siska Adelya Ramadhani
NIM : B352140051
Disetujui oleh Komisi Pembimbing
Prof Dr Drh Iman Supriatna Ketua
Drh Ni Wayan K Karja, MP., PhD Anggota
Drh Adi Winarto, PhD., PAVet Anggota
Diketahui oleh
Ketua Program Studi Biologi Reproduksi
Prof Dr Drh Mohamad Agus Setiadi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MSc. Agr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Penelitian telah dilaksanakan sejak bulan Januari sampai Juni 2016 dengan judul Pengendalian Folikulogenesis Ovarium Mencit (Mus musculus) dengan Pemberian Ekstrak Biji Kapas (Gossypiumhirsutum).
Terima kasih penulis ucapkan kepada Bapak Prof Dr Drh Iman Supriatna, Ibu Drh Ni Wayan Kurniani Karja, MP., PhD dan Bapak Drh Adi Winarto, PhD., PAVet selaku pembimbing atas semua arahan selama penulis melakukan persiapan, penelitian sampai dengan terlaksanannya ujian thesis ini, serta Bapak Prof Dr Drh Mohamad Agus Setiadi yang telah banyak memberi saran. Penulis juga mengucapkan kepada Institut Pertanian Bogor khususnya Sekolah Pascasarjana yang telah mengizinkan penulis untuk menempuh pendidikan magister di Institut Pertanian Bogor. Ucapan terima kasih juga penulis haturkan kepada ketua dan sekretaris program studi Biologi Reproduksi atas segala arahan yang diberikan selama penulis menjalani perkuliahan di Biologi Reproduksi, Ketua Departemen Klinik Reproduksi dan Patologi, ketua Divisi Kebidanan atas segala kebijakan yang diberikan selama penulis menjalani kuliah.
Ungkapan terima kasih penulis sampaikan kepada Ayahanda Mulyadi dan ibunda Delmawati atas segala doa dan kasih sayangnya yang tiada henti serta adik Alif Adelyadi dan Febriella Fauziah serta abang Asra Abdiatas dukungan selama penulis menyelesaikan studi. Ucapan terima kasih juga penulis haturkan kepada sahabat Nofri Zayani selaku teman seperjuangan yang mendampingi penulis hingga selesainya studi, teman diskusi di Laboratorium In Vitro Alvien Nuraini, Magfira, Yulida Nofa, dan Musthamin Balumbi serta teman-teman Biologi Reproduksi lainnya atas semua dukungan dan keceriaan yang diberikan selama penulis melaksanakan studi di IPB.
Semoga karya ilmiah ini bermanfaat.
Bogor, Desember 2016
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
1 PENDAHULUAN 1
Latar Belakang 1
Kerangka Pemikiran 2
Tujuan Penelitian 3
Manfaat Penelitian 3
2 TINJAUAN PUSTAKA 3
Ovarium Mencit (Mus musculus) 3
Biji Kapas (Gossypium hirsutum) 8
Mekanisme Kerja Gosipol pada Biji Kapas terhadap Ovarium 9
3 METODE 11
Waktu dan Tempat 11
Alat dan Bahan 11
Rancangan Penelitian 11
Prosedur Penelitian 12
Analisis Data 15
4 HASIL DAN PEMBAHASAN 15
5 SIMPULAN 25
DAFTAR PUSTAKA 26
LAMPIRAN 31
DAFTAR TABEL
1 Gambaran mikroskopis hasil vaginal smear pada masing-masing
fase siklus estrus 7
2 Morfologi vagina pada masing-masing fase dari siklus estrus 7
3 Klasifikasi folikel pada ovarium mencit 14
4 Jumlah folikel berkembang pada mencit setelah pemberian
ekstrak biji kapas 21
5 Jadwal pelaksanaan penelitian 32
DAFTAR GAMBAR
1 Biji kapas 8
2 Morfometri ovarium setelah pemberian ekstrak biji kapas 16 3 Struktur fungsional ovarium setelah pemberian ekstrak biji kapas 17 4 Morfologi ovarium setelah pemberian ekstrak biji kapas 18 5 Siklus estrus mencit setelah pemberian ekstrak biji kapas selama
24 hari 18
6 Ovarium mencit 19
7 Perkembangan folikel ovarium 20
8 Histologi ovarium setelah pemberian ekstrak biji kapas 21 9 Rata-rata jumlah folikel pada potongan histologi setelah
pemberian ekstrak biji kapas 23
10 Rata-rata jumlah folikel atresia setelah pemberian ekstrak biji kapas 24
DAFTAR LAMPIRAN
1 Jadwal pelaksanaan penelitian 32
2 Persetujuan perlakuan etik hewan 33
1
PENDAHULUAN
Latar Belakang
Fertilitas merupakan kemampuan organ reproduksi untuk bekerja optimal dalam menjalankan fungsi fisiologi reproduksi untuk menghasilkan keturunan. Pada betina, fertilitas berkaitan erat dengan folikulogenesis yaitu proses pertumbuhan dan perkembangan folikel yang terjadi di dalam ovarium (Leung dan Adashi 2004). Beberapa faktor dapat memengaruhi folikulogenesis seperti integrasi sinyal ekstraovarian dan faktor intrafolikuler (Webb et al. 2003), endokrinologi (Paris et al. 2009), perkembangan gonad, serta kondisi nutrisi (Chavatte-Palmer et al. 2014). Interaksi faktor-faktor tersebut menentukan folikel akan terus dikembangkan atau dialihkan ke jalur atresia. Oleh karena itu, pengendalian folikulogenesis dapat dijadikan sebagai salah satu cara untuk mencegah terjadinya fertilisasi dan dapat digunakan sebagai salah satu metode kontrasepsi.
Kontrasepsi memiliki peran penting dalam program keluarga berencana (KB), namun perlu diketahui cara pemilihan kontrasepsi yang cocok dan tepat. Kontrasepsi yang paling banyak dipakai adalah kontrasepsi hormonal jenis suntikan KB yang mengandung 2 komponen bahan aktif yaitu estrogen dan progesteron. Progesteron dalam kontrasepsi berfungsi untuk mengentalkan lendir serviks dan mengurangi kemampuan rahim untuk menerima sel yang telah dibuahi. Namun hormon ini juga mempermudah perubahan karbohidrat menjadi lemak, sehingga sering kali efek sampingnya adalah penumpukan lemak yang menyebabkan berat badan bertambah dan menurunnya keinginan seksual (Kusumaningrum 2009).
Standar pemilihan bahan kontrasepsi yang baik yaitu tidak memiliki efek samping yang buruk terhadap pengguna dan bersifat reversible. Persyaratan ini banyak terdapat pada bahan alam terutama tumbuhan. Penggunaan bahan kontrasepsi dari tumbuhan pada manusia dapat dikembalikan tingkat fertilitasnya jika telah dihentikan. Banyak zat tumbuhan telah dipercaya mempunyai aktivitas sebagai antifertilitas sehingga sering digunakan sebagai bahan kontrasepsi. Salah satu tumbuhan yang mengandung zat antifertilitas adalah tanaman kapas karena memiliki kandungan zat aktif berupa gosipol (Singla dan Garg 2013; Li et al. 1989; Lin et al. 1985). Banyak penelitian telah dilakukan untuk mengkaji pengaruh pemberian ekstrak biji kapas untuk mengetahui pengaruhnya terhadap reproduksi pada hewan model.
Pemberian gosipol secara oral sebanyak 30 mg kg-1 BB setiap hari selama lima minggu mampu menurunkan fertilitas (Singla dan Garg 2013) tikus jantan tanpa menurunkan libido (Hadley et al. 1981). Pemberian gosipol pada hewan jantan juga tidak memengaruhi laju pertumbuhan tikus, bobot testis, dan kelenjar aksesori seksual (White et al. 1988). Sedangkan pada betina, pemberian 25 mg kg -1
2
gosipol dihentikan (Gadelha et al. 2014a). Hal ini menyebabkan ekstrak biji kapas memiliki potensi besar jika dikembangkan sebagai kontrasepsi ketika dilakukan penghentian pemakaian maka kondisi fisiologis tubuh pengguna berangsur kembali normal (Rusmiati 2010).
Penelitian pada sapi dilaporkan bahwa gosipol yang dicampur dalam pakan mengakibatkan folikel besar (> 5 mm) pada ovarium relatif sedikit dibandingkan dengan yang diberi makan kedelai (Gadelha et al. 2014a). Hal ini dapat terjadi karena degenerasi yang luas pada folikel akibat perubahan struktur fungsional membran akibat gosipol berikatan dengan membran sel dan masuk ke dalam sel (Cuellar dan Ramirez 1993). Degenerasi biasa diikuti oleh kehilangan oosit, sel granulosa, dan reseptor hormon (Hafez dan Hafez 2000). Pada sel-sel tersebut terjadi piknosis, kromatolisis inti dan deskuamasi sel granulosa (Bansode 1994). Selain itu gosipol berpengaruh pada steroidogenesis ovarium serta ekspansi oosit kumulus sapi dan pematangannya. Gosipol juga menunjukkan penurunan Immunoglobulin G (IgG) (Gadelha et al. 2014a). Konsentrasi IgG diperlukan dalam cairan folikel untuk ovulasi. Peningkatan konsentrasi IgG merupakan akibat dari perluasan dan peningkatan cairan dalam folikel (Hafez dan Hafez 2000). Hambatan steroidogenesis akibat gosipol terjadi pada sel luteal ovarium yang diamati pada sapi betina (Yurekli et al. 2009). Gosipol menghambat sintesis progesteron dengan menekan aktivitas enzim steroidogenik yaitu beberapa enzim dehidrogenase (Bansode 1994). Penelitian Li et al. (1989) memberikan bukti bahwa gosipol dosis tinggi yang diberikan pada hewan bunting tidak memberi efek teratogenik dan retardasi pertumbuhan.
Banyak penelitian yang dilakukan untuk mengetahui pengaruh pemberian ekstrak biji kapas pada hewan coba, akan tetapi belum ditemukan laporan tentang pengaruh pemberian ekstrak biji kapas terhadap gambaran folikulogenesis. Oleh karena itu diperlukan kajian pengendalian folikulogenesis ovarium dan hubungannya dengan siklus estrus mencit setelah pemberian ekstrak biji kapas.
Kerangka Pemikiran
3 sehingga follicle stimulating hormone (FSH) dan luteinizing hormone (LH) yang dikeluarkan menjadi berkurang yang mengakibatkan proliferasi sel-sel di ovarium berkurang. Bahan yang bisa memasuki sel, menurunkan produksi energi, menekan sekresi sel, antiproliferatif, dan memengaruhi hormonal maka bahan ini berpotensi untuk digunakan dalam mengendalikan folikulogenesis. Pada dosis yang tepat sangat mungkin bisa dimanfaatkan sebagai bahan antifertilitas. Pendekatan secara
in vivo merupakan suatu pilihan dalam pembuktian bahwa pengontrolan folikulogenesis dalam dosis yang tepat dapat berperan sebagai antifertilitas.
Tujuan Penelitian
Penelitian ini bertujuan mengkaji pengendalian folikulogenesis ovarium dan status siklus estrus mencit dengan pemberian ekstrak biji kapas.
Manfaat Penelitian
Penelitian ini bermanfaat untuk.
1. Memberikan informasi ilmiah mengenai efek ekstrak biji kapas terhadap pengendalian folikulogenesis pada ovarium dan siklus estrus mencit.
2. Mengatasi permasalahan penggunaan kontrasepsi melalui pengembangan penggunaan herbal sebagai bahan kontrasepsi.
2
TINJAUAN PUSTAKA
Ovarium Mencit (Mus musculus)
4
Folikulogenesis
Semua folikel di korteks ovarium berada dalam fase folikel primordial sebelum mencapai masa pubertas (Lewis 2007). Folikel primordial ini berisi oosit imatur berukuran kecil (diameter20m) yang dikelilingi satu lapis sel granulosa bertipe pipih selapis. Mencit merupakan hewan yang memulai proses meiosis profase 1 pada hari ke 13.5 setelah konsepsi (Chavatte-Palmer et al. 2014). Oosit berada pada istirahat meiosis profase 1 dan tidak menyelesaikan pembelahan meiosis sebelum mencapai masa pubertas (Aerts dan Bols 2010; Irianto 2014). Jumlah folikel primordial dalam ovarium berhubungan dengan masa reproduksi betina (Patricia 2014). Durasi perkembangan folikel mencit dari primordial hingga folikel ovulasi rata-rata 20 hari (Aerts dan Bols 2010). Menurut Lewis (2007) folikulogenesis bertujuan untuk perekrutan folikel yang diovulasikan.
Folikulogenesis secara garis besar terbagi dalam dua fase yaitu fase preantral dan fase antral. Fase preantral atau gonadotropin-independent phase ditandai dengan pertumbuhan dan diferensiasi oosit yang dipengaruhi oleh faktor-faktor pertumbuhan melalui sistem autokrin atau parakrin (Lewis 2007). Folikel pada ovarium dikategorikan berdasarkan penampilan morfologi dari granulosa terkait lapisan sel menjadi primordial, primer, sekunder, folikel antral dan atresia (Myers
et al. 2004). Folikel primordial cenderung ditemukan menuju wilayah subkapsular dari korteks ovarium (Scudamore 2014). Fase antral atau gonadotropin-dependent phase ditandai dengan peningkatan pesat dari ukuran folikel itu sendiri. Fase antral diatur oleh follicle stimulating hormone (FSH) dan luteinizing hormone (LH) serta faktor-faktor pertumbuhan lainnya. Faktor-faktor pertumbuhan ini akan merangsang proliferasi sel dan memengaruhi aktivitas gonadotropin (Gannon 2013). Akinola et al. (2006) mengatakan bahwa folikulogenesis dipengaruhi oleh gonadotropin dan estrogen. Menurut Findlay et al. (1996) pengadaan kapasitas sel-sel folikel terhadap respon gonadotropin merupakan kunci folikulogenesis. Folikulogenesis dimulai dengan diambilnya folikel primordial ke dalam suatu kumpulan folikel-folikel yang sedang tumbuh berkembang dan dapat diakhiri dengan ovulasi maupun atresia.
Tahap-tahap Folikulogenesis sebagai berikut.
Folikel Primer
5
Follicle stimulating hormone (FSH) memengaruhi peningkatan jumlah sel-sel granulosa (Lewis 2007).
Folikel Sekunder
Perubahan utama yang terjadi selama perkembangan folikel sekunder adalah peningkatan jumlah sel granulosa dan pembentukan sel teka. Oosit terus tumbuh dan memengaruhi perkembangan folikuler (Eppig 2001). Pada tahap ini juga terjadi transisi sel granulosa dari epitel selapis kuboid menjadi epitel berlapis kolumnar. Perkembangan folikel diikuti oleh sel-sel teka folikuli yang tersusun menjadi satu lapisan dalam sel sekretorik membentuk teka interna dan satu lapisan luar jaringan ikat yang mengandung sel-sel mirip fibroblas membentuk teka eksterna (Patricia 2014). Sel teka memproduksi androgen yang digunakan sebagai substrat untuk menghasilkan estrogen (Lewis 2007; Peluso et al. 1984). Perkembangan sel teka menyebabkan kapiler-kapiler masuk ke dalam lapisan teka interna sehingga folikel mendapat suplai darah sendiri, sedangkan lapisan sel granulosa tetap avaskular (Patricia 2014). Sel-sel granulosa mengaktifkan dan meningkatkan jumlah reseptor FSH, estrogen, dan androgen (Lewis 2007).
Folikel Tersier
Proses yang terjadi pada folikel tersier yaitu perkembangan antrum yang tersebar menjadi satu kesatuan dalam folikel. Antrum mengandung cairan yang disebut cairan folikuler atau liquor folliculi yang berasal dari eksudat plasma (Aerts dan Bols 2010). Cairan folikuler merupakan hasil sekresi dari oosit dan sel granulosa yang mengandung steroid, elektrolit, serta proteoglikan. Follicle stimulating hormone merangsang sel-sel granulosa berdiferensiasi membentuk membran periantral, kumulus oophorus, dan lapisan korona radiata. Sel granulosa menghasilkan aktivin dan meningkatkan ekspresi p450 aromatase akibat stimulasi FSH. Enzim P450 aromatase berperan dalam mengonversi androgen menjadi estrogen. Aktivin akan meningkatkan ekspresi gen reseptor FSH di sel granulosa dan mempercepat folikulogenesis. Sel granulosa juga menghasilkan inhibin yang berperan dalam umpan balik negatif untuk menghambat hipofisa mensekresi FSH. Follicle stimulating hormone berperan penting dalam proses mitosis sel granulosa pada fase ini. Kekurangan FSH pada fase ini akan menghambat pertumbuhan dan perkembangan folikel sehingga folikel menjadi atresia. Pada bagian dalam sel teka interna terbentuk sel-sel teka interstisial. Follicle stimulating hormone dan estrogen menstimulus sel-sel teka interstisial agar meningkatkan jumlah reseptor LH dan meningkatkan aktivitas enzim-enzim steroidogenesis untuk sintesis androstenedion dan testosteron. Androstenedion mengalami aromatisasi menjadi estrogen dengan bantuan enzim p450. Estrogen akan meningkatkan jumlah reseptor FSH pada sel granulosa sehingga sel tersebut berproliferasi (Patricia 2014).
6
LH yang cukup di sel-sel granulosa untuk terjadinya luteinisasi (Leung dan Adashi 2004). Oosit dalam folikel memproduksi faktor ekspansi kumulus (Vanderhyden et al. 1990). Pada sapi terdapat korelasi antara jumlah folikel antral dengan ukuran ovarium dan berasosiasi dengan fertilitas (Evans et al. 2012). Jumlah folikel antral besar meningkat selama proestrus dan menurun sebagai akibat dari ovulasi pada masa estrus (Scudamore 2014).
Folikel de Graaf
Folikel antral yang matang disebut folikel de Graaf (Aerts dan Bols 2010). Oosit dipisahkan dari ruang antral tunggal oleh lapisan sel granulosa sekitarnya (kumulus granulosa). Kumulus ini akan dipertahankan bersama oosit ketika ovulasi (Scudamore 2014). Pada tahap ini terjadi seleksi folikel dominan yang akan berovulasi (Senger 2005). Penurunan kadar FSH menyebabkan folikel-folikel antral yang lebih kecil mengalami atresia, sedangkan folikel-folikel dominan terus mengalami pertumbuhan. Penurunan FSH juga menyebabkan lonjakan LH (Leung dan Adashi 2004). Setelah terjadi lonjakan LH maka folikel disebut folikel preovulatory (Eppig 2001). Lonjakan LH meningkatkan sintesis progesteron dalam sel-sel granulosa (Senger 2005). Progesteron merangsang enzim-enzim
collagenase untuk menghancurkan kolagen di dinding folikel sehingga mudah ruptur. Prostaglandin akan menyebabkan otot polos ovarium berkontraksi untuk membantu pelepasan ovum (Leung dan Adashi 2004). Pada saat yang sama sel-sel granulosa meningkatkan sekresi cairan folikel sehingga meningkatkan volume cairan dalam folikel. Dengan demikian, peningkatan volume ini mengakibatkan enzim-enzim collagenase semakin mendekati dinding folikel dan membentuk stigma (Senger 2005). Stigma merupakan tempat bakal terjadi ovulasi. Setelah terjadi ovulasi, sel-sel granulosa, jaringan ikat, dan pembuluh darah kecil di ovarium mengalami proliferasi. Sel-sel granulosa membesar dan mengandung lutein dengan banyak kapiler serta jaringan ikat sehingga disebut corpus luteum.
Corpus luteum akan dipertahankan mulai dari fertilisasi sampai terbentuknya plasenta. Sel-sel corpus luteum mengalami atrofi jika tidak terjadi fertilisasi, sehingga terbentuk corpus albicans (Patricia 2014).
Tampilan histologi ovarium akan memperlihatkan folikel-folikel yang terdapat pada organ ovarium. Folikel normal merupakan folikel yang memiliki oosit utuh. Folikel ini dibagi menjadi folikel primer, folikel sekunder, folikel tersier, dan folikel de Graaf. Folikel atretik merupakan folikel yang ditandai dengan degenerasi oosit, lapisan sel granulosa tidak teratur, zona pelusida melipat sebagian atau seluruhnya yang berpisah dari korona radiata, dan sel granulosa oosit. Folikel menjadi atretik dapat disebabkan oleh terjadinya apoptosis baik di dalam maupun di luar sel oosit dan sel granulosa (Aguirre-Samudio et al. 2006). Jumlah folikel dipengaruhi oleh jenis hewan, umur, siklus estrus, morfologi ovarium, status reproduksi, faktor genetik, dan faktor lingkungan seperti asupan nutrisi (Lin et al. 1992).
7
smear. Gambaran hasil mikroskopis vaginal smear pada masing-masing fase siklus estrus terlihat pada Tabel 1.
Tabel 1 Gambaran mikroskopis hasil vaginal smear pada masing-masing fase
Sel epitel berinti Sel-sel kecil dengan inti bulat
Proestrus dan estrus terjadi pada fase folikuler, sedangkan metestrus dan diestrus terjadi pada fase luteal ovarium. Mencit mengalami proestrus sekitar 12 jam, estrus 12 jam, metestrus 21 jam, dan diestrus 57 jam. Fase dari siklus estrus mencit juga dapat ditentukan berdasarkan pengamatan morfologi vagina. Gambaran hasil pengamatan morfologi vagina berbagai fase siklus estrus terlihat pada Tabel 2.
Tabel 2 Morfologi vagina pada masing-masing fase dari siklus estrus
Fase Morfologi vagina mencit
Diestrus Vagina terbuka kecil, jaringan berwarna kebiru-biruan hingga ungu, dan sangat lembab.
Proestrus Vagina bercelah, jaringan berwarna kemerah-merahan hingga pink, lembab, serta banyak lipatan longitudinal atau goresan tampak pada bibir dorsal dan ventral.
Estrus Vagina terlihat sama seperti fase proestrus, namun jaringan terlihat lebih pink cerah dan sedikit lembab serta goresan lebih jelas. Metestrus Jaringan vagina pucat, kering, bibir dorsal tidak edema seperti fase
estrus hingga surut, runtuhan seluler keputih-putihan pada dinding dalam atau sebagian yang mengisi vagina.
8
Biji Kapas (Gossypium hirsutum)
Kapas (Gossypium hirsutum) merupakan tanaman arboreus dari Familia Malvaceae. Tanaman kapas mengandung senyawa fenolik yang dihasilkan oleh kelenjar pigmen kapas batang, daun, biji, dan tunas bunga (Gossypium hirsutum
L.) yang disebut gosipol. Konsentrasi gosipol paling banyak terdapat pada biji kapas. Gosipol berbentuk bintik-bintik hitam yang memiliki berat molekul 518.55 Dalton, berpigmen kuning, dan kristal (Gadelha et al. 2014a). Gosipol bersifat polar tetapi tidak larut dalam air (Sutikno 2000) dan heksana (Gadelha et al.
2014a). Gosipol larut dalam aseton, asetonitril, etanol, metanol (Sutikno 2000), metil etil keton (butanone), dan sebagian larut dalam minyak sayur mentah (Gadelha et al. 2014a). Rumus molekulnya adalah C30H30O8 (Sutikno 2000) dan struktur formula kimianya adalah 2.2bis (8-formil-1,6,7-trihydroxy-5-isopropil-3methylnaphthalene) (Gadelha et al. 2014a). Menurut Matham (2011) kandungan gosipol mencapai 50% pada biji kapas. Sedangkan Gadelha et al. (2014a) mengatakan bahwa biji kapas mengandung 14 gram gosipol kg-1. Singla dan Garg (2013) menyatakan minyak biji kapas mentah mengandung 0.01% gosipol. Bentuk biji kapas dapat dilihat pada Gambar 1.
Gambar 1 Biji Kapas
9 diukur pada serum darah (Gadelha et al. 2014a). Gosipol tidak hanya berfungsi sebagai zat antifertilitas melainkan juga berpotensi untuk terapi. Pada tahun 1970 Cina menggunakan biji kapas untuk mengobati fibroids rahim, endometriosis, dan rahim pendarahan pada wanita (Gadelha et al. 2014a).
Mekanisme Kerja Gosipol pada Biji Kapas terhadap Ovarium
Gosipol yang berikatan dengan membran sel menyebabkan peroksidasi lipid pada membran sehingga dapat masuk ke dalam sel. Gosipol dapat bereaksi dengan fosfolipid dan mengganggu enzim-enzim pada membran sehingga menyebabkan perubahan struktur fungsionalnya (Brocas et al. 1997). Hal ini sesuai dengan penelitian Lin et al. (1992) yang menemukan pada kultur sel luteal ovarium sapi bahwa membran selnya memiliki kapasitas mengikat paling tinggi terhadap gosipol. Sedangkan Xue (1981) dalam Lin et al. (1992) dengan menggunakan fraksi subseluler testis menyatakan mitokondria memiliki tingkat penggabungan tertinggi di antara semua organel. Pada konsentrasi tinggi, gosipol merusak enzim pada proses pembentukan energi dari metabolisme oksidatif yaitu pada proses transportasi rantai elektron dan fosforilasi oksidatif (Gadelha et al. 2014a). Hal ini mengakibatkan penurunan produksi adenosine triphosphate (ATP) yang juga telah diteliti Yuan dan Shi (2000). Gosipol tidak hanya menghambat enzim pada membran mitokondria, tetapi juga menghambat enzim yang berhubungan dengan sitoplasma dan membran organel lainnya (Lin et al. 1992). Pada oosit tikus yang terpapar gosipol menunjukkan peningkatan jumlah lisosom di ooplasma dan berkurangnya perkembangan retikulum endoplasma halus. Pada retikulum endoplasma tersebut tidak terdapat sisterna yang membentang serta terjadi penekanan aktivitas sekresi dari sel-sel granulosa (Pan et al. 1987).
Ikatan gosipol dengan lipoprotein membran mengganggu jalur biokimia dan fungsi seluler. Gosipol juga menunjukkan efek antiproliferatif dengan menghambat low density lipoprotein (LDL) pada mitokondria (Gilbert 1995). Gosipol menghambat produksi progesteron dalam sel luteal sapi yang dikultur secara in vitro, terutama dengan menghambat aktivitas plasma membran terkait adenilat siklase dan membran mitokondria terkait sitokrom P450 rantai samping enzim cleavage. Gosipol menekan hormon adrenokortikotropik (ACTH) yang menginduksi produksi kortikosteron pada kultur sel adrenokortical tikus dengan menghambat pembentukan siklik AMP (cAMP) (Lin et al. 1992).
Gosipol merupakan agen antifertilitas yang menghambat steroidogenesis pada kedua jenis kelamin (Lin et al. 1992). Gosipol memiliki efek langsung pada folikel ovarium yaitu menghambat sintesis hormon ovarium (Akinola et al. 2006). Gosipol berkonsentrasi tinggi dapat meningkatkan O2 dengan menghambat aktivitas superokside dismutase (SOD). Karena itu gosipol akan memodulasi steroidogenesis pada ovarium sehingga terjadi pengurangan produksi hormon steroid pada sel granulosa (Basini et al. 2008). Berkurangnya produksi estradiol juga karena gosipol menghambat aktivitas aromatase yang diteliti dalam sel granulosa babi (Akira 1994).
10
antrum. Selain itu juga menginduksi sensitivitas sel granulosa terhadap LH dengan meningkatkan jumlah reseptor LH. Kenaikan reseptor LH mempersiapkan luteinisasi sel-sel granulosa dalam menanggapi LH ovulatory surge. Luteinizing hormone juga merangsang aktivitas adenilat siklase dan meningkatkan cAMP. Siklik AMP merangsang aktivitas protein kinase yang menyebabkan peningkatan ketersediaan kolesterol untuk belahan rantai samping dan meningkatkan produksi testosteron. Gangguan gosipol dapat menyebabkan berkurangnya kemampuan reaksi sel granulosa dan sel teka terhadap sinyal gonadotropin. Hafez dan Hafez (2000) mengemukakan bahwa kurangnya reaksi granulosa dan teka terhadap gonadotropin menyebabkan berhentinya pertumbuhan folikel dan menginisiasi folikel atresia. Viabilitas folikel juga bergantung pada rasio testosteron dan estrogen pada cairan folikel.
Penambahan gosipol pada media kultur dapat menghambat pematangan oosit sapi (Brocas et al. 1997). Gosipol mampu mengambat ekspansi kumulus, menghambat pembentukan cAMP, menghambat maturasi nukleus oosit, dan mengambat produksi progesteron. Pemberian gosipol menyebabkan proses meiosis dari oosit ditahan pada tahap diploten Prophase I. Selain menghambat progesteron, gosipol juga menghambat kortikosteron pada sel luteal (Lin et al.
1994). Pada sel luteal sapi gosipol menekan kegiatan adenilat siklase dan hidroksisteroid dehidrogenase pada jalur steroidogenesis ovarium. Hal ini mengakibatkan penekanan adrenokortikotropik hormon yang diinduksi oleh sekresi kortikosteron (Gilbert et al. 1995). Gosipol bertindak pada produksi siklik ATP, reseptor LH yaitu G-protein, dan unit katalitik atau konversi ATP menjadi AMP siklik. Gosipol menghambat aktivitas adenilat siklase langsung dengan interaksi di domain ikatan nukleotida dari enzim sub unit katalitik yang diamati pada sel Leydig (Pearce et al. 1986).
11
3
METODE
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Januari sampai Juni 2016. Penelitian ini dilaksanakan di Laboratorium Farmasi Fakultas Kedokteran Hewan (FKH) Institut Pertanian Bogor (IPB) untuk ekstraksi biji kapas, Laboratorium Teknologi Pangan Pusat Antar Universitas (PAU) IPB untuk evaporasi ekstrak biji kapas, rumah sakit hewan pendidikan (RSHP) IPB sebagai tempat pemeliharaan hewan dan penelitian, Laboratorium Histologi FKH IPB untuk pembuatan dan pengamatan preparat histologi serta Laboratorium In Vitro Fertilisasi FKH IPB untuk pembuatan dan pengamatan preparat ulas vagina.
Alat dan Bahan
Alat yang digunakan pada penelitian ini adalah neraca Ohause, oven, gelas beker, gelas ukur, batang pengaduk, Erlenmeyer, rotary vacum evaporator, petri dish, neraca analitik, sonde (jarum gavage), perangkat alat bedah, mikroskop stereo, jangka sorong, lup, botol sampel, syringe, mikrotom, embedding console,
cryo console, inkubator, dan mikroskop binokuler. Bahan yang digunakan yaitu biji kapas, etanol, aquadest, Giemsa, sodium chloride (NaCl) 0.9%, alkohol bertingkat (70%, 80%, 90%, 95%, absolut), xylol, formalin, asam asetat, asam pikrat, blok parafin, larutan haematoxylin, larutan eosin, dan larutan carboxy methyl cellulose (CMC) 0.2%.
Rancangan Penelitian
12
Prosedur Penelitian
Persiapan Hewan Coba
Hewan yang digunakan adalah 60 ekor mencit induk (Mus musculus L. DDY) umur 14 sampai 15 minggu dengan rata-rata berat badan 30 sampai 35 g. Mencit diperoleh dari Laboratorium Non Ruminansia dan Satwa Harapan Fakultas Peternakan IPB yang diadaptasikan selama dua minggu. Pada minggu kedua masa adaptasi mencit diberi obat pra perlakuan berupa dosis tunggal anthelmintik (Combantrin®) 1.4 mg kg-1 BB. Mencit strain ini adalah mencit yang memiliki reproduksi yang baik yang ditandai dengan memiliki anak 8 sampai 15 ekor per kelahiran. Selain itu mencit ini juga memiliki sensitivitas yang tinggi terhadap perlakuan secara oral. Mencit dipelihara secara kelompok (lima ekor mencit perkandang). Mencit dipelihara dalam kandang bentuk segi empat ukuran 38 cm(p) x 27 cm(l) x 13 cm(t) yang ditutupi dengan kawat. Bedding pada kandang berupa serutan kayu diberi setebal 5 cm yang diganti dua kali seminggu. Selama pemeliharaan pakan diberikan 5 g perhari berupa pellet mencit produksi Indo Feed® yang dibagi menjadi dua kali pemberian yaitu pagi dan sore hari. Air minum diberikan secara ad libitum melalui botol minum mencit. Pakan yang diberikan adalah pakan yang sesuai kebutuhan mencit dengan kandungan nutrisi: air 12%, protein K. 23%, lemak 4%, serat kasar 5%, abu 8%, protein D.D 17%, M.E Kcal-1 2750%, kalsium 1%, dan phospor 0.8%. Siklus pencahayaan diatur terang (06.00 - 18.00) dan gelap (18.00 - 6.00) WIB. Penggunaan hewan coba dalam penelitian ini sudah mendapatkan persetujuan dari Komisi Etik Hewan IPB nomor 15 – 2016 IPB.
Pembuatan Ekstrak Biji Kapas dan Pemberian Perlakuan
Bahan yang digunakan adalah biji kapas yang diperoleh dari Balai Penelitian Tanaman Pemanis dan Serat (Balittas), Malang, Provinsi Jawa Timur. Biji kapas dikoleksi dari tanaman kapas umur 5 bulan. Biji kapas yang digunakan berbentuk bulat utuh dan berwarna coklat kehitaman. Biji kapas kering yang masih utuh ditimbang kemudian digerus selanjutnya disaring untuk mendapatkan serbuk simplisia.
Ekstraksi dilakukan dengan metode maserasi (perendaman). Metode maserasi dimulai dengan perendaman serbuk simplisia dalam pelarut polar etanol 80% (1:10 b/v) selama 3 hari. Penggantian pelarut dan penampungan dilakukan setiap hari sampai larutan ekstrak nampak bening. Filtrasi dilakukan dengan menggunakan kapas sehingga diperoleh larutan ekstrak. Filtrat dari penyaringan dipekatkan menggunakan rotary vacuum evaporator pada suhu 68 oC hingga diperoleh ekstrak berbentuk serbuk (Chandrashekar et al. 2013). Kemudian dilakukan penimbangan dengan neraca analitik. Ekstrak disimpan dalam desikator.
13 pelarut carboxy methyl cellulose (CMC) 0.2% sebanyak 0.5 ml. Bubuk CMC ditimbang 2 g kemudian dilarutkan dengan sodium chloride (NaCl) 0.9% hingga volumenya mencapai 1 liter dan diaduk hingga homogen dengan batang pengaduk.
60 ekor mencit dibagi menjadi empat kelompok (15 ekor mencit setiap perlakuan) dan diberi ekstrak biji kapas dengan dosis 0; 1.5; 2.1 dan 2.7 g kg-1 BB selama 5 hari (D0-5; D1.5-5; 5 dan D2.7-5), 10 hari (D0-10; D1.5-10; D2.1-10 dan D2.7-D2.1-10), 15 hari (D0-15; D1.5-15; D2.1-15 dan D2.7-15), dan 24 hari (D0-24; D1.5-24; D2.1-24 dan D2.7-24). Ekstrak biji kapas diberikan pada fase proestrus dan didefinisikan sebagai hari pertama (H1). Pada setiap akhir periode pemberian ekstrak biji kapas, mencit dieuthanasia masing-masing 3 ekor dari tiap perlakuan untuk mengamati morfologi ovarium dan folikulogensis berdasarkan gambaran histomorfolgi perkembangan folikel. Sedangkan untuk sampel
reversible diambil 10 hari setelah pemberian ekstrak biji kapas selama 24 hari dihentikan.
Kajian siklus estrus mencit berdasarkan gambaran sel ulas vagina dengan pewarnaan Giemsa
Pengamatan terhadap siklus estrus dilakukan mulai dari hari pertama (H1) sampai hari pengambilan ovarium yaitu setelah hari ke-5, 10, 15, 24 dan 34. Ulas vagina dilakukan dengan bantuan cotton bud and NaCl fisiologis yang diulas 3-5 kali selanjutnya diulas pada gelas obyektif, kemudian diwarnai dengan giemsa, prerapat diamati di bawah mikroskop (mikrosokop binokuler) dengan perbesaran obyektif 10 dan 40 kali. Pada ulas vagina ditentukan fase yang sedang dialami hewan coba melalui tipe sel, proporsi masing-masing sel, dan keberadaan leukosit.
Kajian pemberian ekstrak biji kapas terhadap morfologi ovarium mencit berdasarkan morfometri ovarium
Setiap 5 hari mencit dieuthanasia sebanyak 3 ekor pada setiap perlakuan dengan anesthesia ketamine dan xylazine metode extinguish secara intracardial,
kemudian dilakukan pembedahan dimulai dari bagian inferior ke superior pada bagian abdominal. Ovarium dipisahkan dari lemak yang menempel. Kemudian melakukan pengamatan dengan mengukur morfometri ovarium mencit (Mus musculus L. DDY). Parameter yang diamati adalah panjang ovarium, lebar ovarium, bobot, dan struktur fungsional ovarium dari masing-masing mencit. Ovarium kemudian disimpan dalam NaCl untuk dibuatkan preparat histologinya.
Kajian pemberian ekstrak biji kapas terhadap gambaran folikulogensis dengan histomorfologi ovarium
14
sampel, setelah itu ovarium dipindahkan ke dalam larutan alkohol 70% selama 24 jam pada botol sampel, kemudian ovarium yang telah difiksasi di triming dan dimasukkan ke dalam tissue cassette. Selanjutnya dilakukan tahapan dehidrasi yaitu ovarium dalam tissue cassette direndam dalam larutan alkohol bertingkat (80%, 90%, dan 95%) masing-masing selama 24 jam, kemudian dipindahkan ke dalam alkohol absolut dengan tiga kali tingkatan dengan waktu masing-masing 60 menit. Tahapan clearing I dimulai dengan merendam ovarium ke dalam xylol
dengan tiga kali tingkatan dengan waktu masing-masing 60 menit. Tahapan infiltrasi yaitu ovarium direndam dalam parafin cair dengan tiga kali tingkatan dengan waktu masing-masing 60 menit pada suhu inkubator 65 oC. Tahapan
embedding yaitu ovarium ditanam dalam cetakan, kemudian parafin dibiarkan mengeras selama 24 jam. Tahapan pemotongan yaitu spesimen dipotong dengan mikrotom setebal 4-5 mikron, lalu tempelkan pada gelas objek, direnggangkan di atas alat pemanas untuk menghilangkan gelembung udara selama 1-2 jam pada suhu 45 oC. Tahapan deparafinisasi yaitu preparat direndam berturut-turut (xylol I, II, dan III). Tahapan rehidrasi yaitu preparat direndam berturut-turut dengan alkohol absolut I, II, III, 95%, 90%, 85%, 80%, dan 70%) masing-masing 3 menit, kemudian direndam dengan air keran selama 10 menit dan aquadest 5 menit. Tahapan pewarnaan yaitu preparat direndam dalam larutan haematoxylin selama 1 menit, kemudian direndam dengan air keran selama 10 menit dan aquadest 10 menit, selanjutnya direndam dalam larutan eosin selama 2 menit dan cuci dengan air keran. Tahapan dehidrasi yaitu preparat direndam berturut-turut di dalam alkohol 70%, 80%, 85%, 90%, 95% I, 95% II, absolut I, II, III masing-masing 1 menit. Tahapan clearing yaitu preparat direndam dalam xylol I, II, III masing-masing selama 1 menit, dan ditutup dengan cover glass, dikeringkan selama 24 jam. Preparat diberi label sesuai dengan perlakuan sehingga didapatkan preparat permanen histologis ovarium yang dapat diamati di bawah mikroskop.
Folikulogenesis diamati dengan melihat jumlah folikel primer, sekunder, dan tersier (Myers et al. 2004) sesuai dengan klasifikasi folikel pada ovarium mencit menurut Peters (1969) seperti disajikan pada Tabel 3. Folikel merupakan struktur selular bundar yang ditemukan di ovarium sebagai tempat berkembangnya ovum dan kemudian melepaskannya pada saat ovulasi. Penghitungan jumlah folikel yang terdapat pada ovarium merupakan pendekatan untuk studi folikulogenesis (Myers et al. 2004).
Tabel 3 Klasifikasi folikel pada ovarium mencit menurut Peters (1969) Klasifikasi
< 20 Oosit mulai dikelilingi selapis sel epitel pipih. Folikel primer > 20 Oosit dikelilingi selapis sel epitel kuboid (sel
granulosa) dan zona pelusida. Folikel
sekunder
70 Oosit dikelilingi beberapa lapis sel granulosa, zona pelusida, sudah mulai ada antrum yang tersebar pada lapisan granulosa.
15 Analisis Data
Penelitian ini menggunakan faktorial rancangan acak lengkap (RAL) dengan dua faktor perlakuan yaitu lama waktu pemberian dan dosis. Data berupa morfometri ovarium, jumlah struktur fungsional, jumlah folikel berkembang, persentase jumlah folikel primer, sekunder, dan tersier dianalisis dengan analisis varian (ANOVA). Perbedaan hasil ANOVA dilanjutkan dengan Duncan multiple range test (DMRT) pada taraf 5%. Data diolah menggunakan program SPSS Versi 16.
4
HASIL DAN PEMBAHASAN
16
Gambar 2 Morfometri ovarium setelah pemberian ekstrak biji kapas. a. Panjang. b. Lebar. c. Bobot. kontrol, 1.5 g kg-1 BB, 2.1 g kg-1 BB, 2.7 g kg-1 BB. Huruf kecil yang berbeda pada setiap histogram di antara kelompok perlakuan di atas menunjukkan perbedaan nyata (P < 0.05).
Data pada Gambar 2 menunjukkan rataan panjang, lebar, dan bobot ovarium setelah pemberian ekstrak biji kapas relatif sama antara kelompok perlakuan dengan kontrol. Pada Gambar 2 ini juga terlihat bahwa jika pemberian ekstrak biji kapas melebihi 24 hari diperkirakan dapat menyebabkan penurunan bobot ovarium. Bobot ovarium dipengaruhi oleh panjang, lebar, ketebalan serta struktur penyusun ovarium. Folikel ovarium yang telah mengovulasikan oosit akan mengalami diferensiasi sel granulosa menjadi luteal. Sel granulosa memiliki ukuran yang lebih kecil dibandingkan dengan luteal sehingga menyebabkan peningkatan ukuran dan bobot ovarium. Penelitian pada domba yang diberikan biji kapas tidak menunjukkan penurunan bobot ovarium (Camara et al. 2015). Hal ini dipengaruhi oleh jenis sistem pencernaan mencit yang lebih sensitif terhadap asupan makanan. Hewan monogastrik memiliki kepekaan lebih tinggi terhadap toksisitas gosipol yang dikandung oleh biji kapas daripada hewan ruminansia (Randel et al. 1992). Pada hewan ruminansia gosipol tidak diikat selama fermentasi ruminal (Randel et al. 1996).
17
Gambar 3 Struktur fungsional ovarium setelah pemberian ekstrak biji kapas. a. Jumlah folikel tersier. b. Jumlah CL. kontrol, 1.5 g kg-1 BB, 2.1 g kg-1 BB, 2.7 g kg-1 BB. Huruf kecil yang berbeda pada setiap histogram di antara kelompok perlakuan di atas menunjukkan perbedaan nyata (P < 0.05).
Pengamatan struktur fungsional dapat dilihat pada Gambar 3. Berdasarkan gambar tersebut teramati bahwa terjadi penurunan jumlah folikel tersier setelah pemberian ekstrak biji kapas selama 24 hari pada semua dosis perlakuan. Pengaruh pemberian ekstrak biji kapas sama di antara dosis 1.5; 2.1; dan 2.7 g kg -1
BB. Penghentian pemberian ekstrak selama 10 hari menyebabkan peningkatan kembali jumlah folikel tersier. Rataan jumlah CL ovarium setelah pemberian ekstrak biji kapas juga relatif sama di antara perlakuan. Penghentian pemberian ekstrak selama 10 hari juga menyebabkan peningkatan jumlah CL.
Jumlah CL terlihat banyak karena diduga CL yang terbentuk dari folikel unovulasi. Folikel unovulasi dicirikan dengan adanya folikel dominan yang akan mencapai ovulasi, namun mengalami regresi dan menjadi atretik. Folikel unovulasi diikuti oleh pembentukan folikel lutein. Dinding permukaan CL anovulasi secara morfologi lebih tebal (berselaput) dibandingkan CL ovulasi (Noakes 1986). Prostaglandin F2α memainkan peranan dasar dalam ruptur folikel, hambatan sintesis prostaglandin akan mencegah ovulasi, dan aksinya terjadi pada lapisan albuginea dan epitel. Ketika sintesis prostaglandin dihambat, oosit tetap berada pada sisi dalam folikel luteinisasi atau mungkin diovulasikan di sisi dalam ovarium. Prostaglandin F2α terlibat dalam ruptur folikel, memperbaiki lapisan folikel (PGE2α), dan mengakhiri pembentukan corpus luteum (Siregar 2010).
18
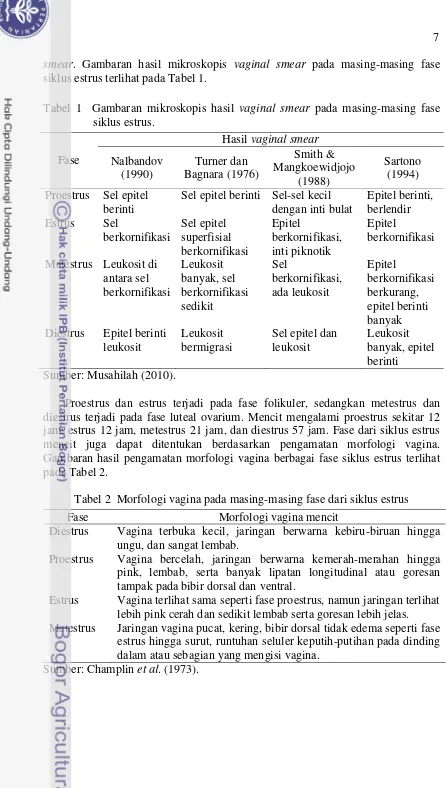
dengan pendapat Gu et al. (1991) bahwa gosipol menginduksi perubahan morfologi dalam organel seluler. Corpus luteum (CL) merupakan tempat utama produksi progesteron selama fase luteal pada betina siklik dan bunting yang normal. Progesteron penting untuk mengatur siklus estrus, implantasi embrio dan pemeliharaan kebuntingan (Lin et al. 1992).
Gambar 4 Morfologi ovarium mencit setelah pemberian ekstrak biji kapas. a kontrol, b perlakuan. Corpus luteum.
Corpus luteum yang berwarna merah muda disebabkan oleh banyak
pembuluh darah yang menunjukkan jaringan sedang aktif. Corpus luteum yang berwarna putih dikarenakan terjadi degenerasi. Selain itu, perubahan di dalam ovarium dipengaruhi oleh respon terhadap perubahan hormonal yang diamati dari perubahan siklus estrus. Perubahan siklus estrus setelah pemberian ekstrak biji kapas selama 24 hari dapat dilihat pada Gambar 5.
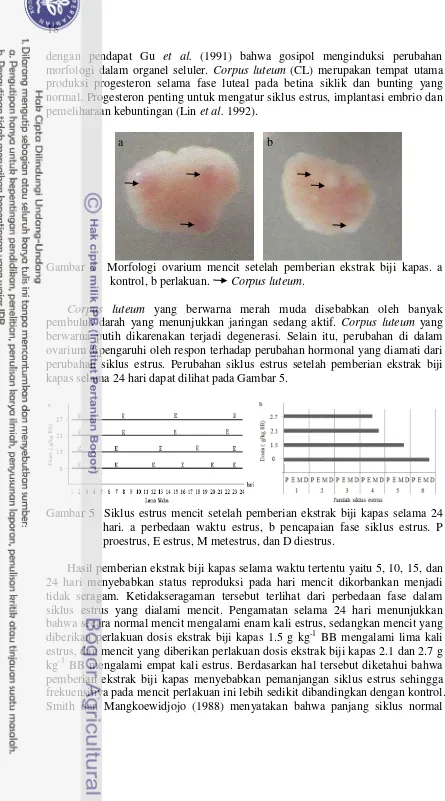
Gambar 5 Siklus estrus mencit setelah pemberian ekstrak biji kapas selama 24 hari. a perbedaan waktu estrus, b pencapaian fase siklus estrus. P proestrus, E estrus, M metestrus, dan D diestrus.
Hasil pemberian ekstrak biji kapas selama waktu tertentu yaitu 5, 10, 15, dan 24 hari menyebabkan status reproduksi pada hari mencit dikorbankan menjadi tidak seragam. Ketidakseragaman tersebut terlihat dari perbedaan fase dalam siklus estrus yang dialami mencit. Pengamatan selama 24 hari menunjukkan bahwa secara normal mencit mengalami enam kali estrus, sedangkan mencit yang diberikan perlakuan dosis ekstrak biji kapas 1.5 g kg-1 BB mengalami lima kali estrus, dan mencit yang diberikan perlakuan dosis ekstrak biji kapas 2.1 dan 2.7 g kg-1 BB mengalami empat kali estrus. Berdasarkan hal tersebut diketahui bahwa pemberian ekstrak biji kapas menyebabkan pemanjangan siklus estrus sehingga frekuensinya pada mencit perlakuan ini lebih sedikit dibandingkan dengan kontrol. Smith dan Mangkoewidjojo (1988) menyatakan bahwa panjang siklus normal
19 pada mencit adalah empat sampai lima hari. Pemberian gosipol yang terkandung dalam ekstrak biji kapas menyebabkan pemanjangan siklus estrus dengan meningkatkan durasi fase metestrus dan diestrus (Gadelha et al. 2014a). Metestrus dan diestrus seiring dengan fase luteal yang terjadi di ovarium. Fase luteal terjadi selama CL aktif. Fase luteal diperpanjang jika tidak terjadi luteolisis sehingga hewan tidak bisa mencapai siklus berikutnya (Senger 2005). Keberhasilan luteolisis sangat bergantung pada kondisi endometrium yang menghasilkan agen luteolitik berupa PGF2α (Senger 2005). Sedangkan penurunan sekresi hormon luteotropik pituitari tidak memengaruhi luteolisis CL (Hafez dan Hafez 2000). Gosipol mengganggu endometrium secara langsung yaitu aktivitas sekresi, proliferasi, dan pengecilan endometrium (Peng-di et al. 1984). Hal ini diduga menyebabkan sekresi PGF2α tidak mencukupi untuk melisiskan CL sehingga terjadi perpanjangan siklus estrus yaitu fase metestrus dan diestrus. Pemanjangan siklus estrus akan berkaitan dengan kondisi folikulogenesis pada ovarium.
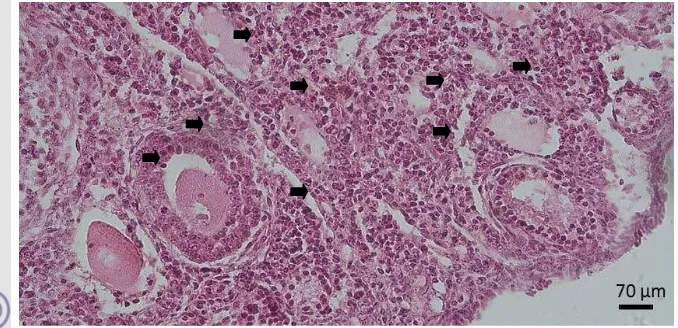
Hasil pengamatan mikroskopis berdasarkan histologi dari ovarium mencit dalam penelitian ini didapatkan bahwa pada ovarium terdapat folikel yang beragam. Keragaman folikel dapat dilihat dari hasil histologi ovarium pada Gambar 6.
Gambar 6 Ovarium mencit control (HE). a corpus luteum. b folikel sekunder. c folikel tersier. Korteks ovarium.
20
laporan Scudamore (2014) yang mengelompokkan folikel ovarium berdasarkan penampilan morfologi lapisan sel granulosa seperti yang disajikan pada Gambar 7. Pertumbuhan dan perkembangan folikel mencakup pertumbuhan oosit, proliferasi sel granulosa, pembentukan antrum, aktivasi reseptor follicle stimulating hormone (FSH) dan luteinizing hormone (LH), pembentukan hormon estrogen hingga oosit diovulasikan (Leung dan Adashi 2004). Secara histomorfologi terlihat bahwa pada saat pertumbuhan terjadi pertambahan ukuran folikel dari folikel primordial, primer, dan sekunder. Folikulogenesis dimulai dari perekrutan folikel primordial dalam jumlah banyak yang dijumpai pada bagian korteks ovarium. Perekrutan tersebut diatur oleh tekanan jaringan dalam stroma pada korteks ovarium. Kepadatan stroma menurun secara bertahap dari korteks hingga bagian medula (Woodruff dan Shea 2011).
Gambar 7 Perkembangan folikel ovarium (HE). a folikel primordial. b folikel primer. c dan d folikel sekunder. e folikel tersier. f corpus luteum
(CL).
Folikulogenesis dimulai saat folikel primordial menuju arah medula yang lebih lunak. Hal tersebut memicu selesainya istirahat meiosis oosit dalam folikel primordial (Silber 2015) dan berkembang menjadi folikel primer kemudian folikel sekunder. Pada folikel sekunder mulai terjadi pembentukan antral kecil dan tersebar, selanjutnya antral menyatu sehingga terbentuk folikel tersier. Pada masa pembentukan antrum terjadi dinamika folikel antral yang terdiri dari perekrutan, seleksi, dominan dan atresi. Selama perkembangan jumlah folikel terus berkurang hingga mencapai folikel preovulasi (Senger 2005).
21
Gambar 8 Histologi ovarium setelah pemberian ekstrak biji kapas (HE). Folikel degenerasi.
Hasil pengamatan pada Gambar 8 menunjukkan bahwa folikulogenesis mencit dipengaruhi oleh pemberian ekstrak biji kapas. Folikel mengalami degenerasi dengan perubahan yang bervariasi pada semua tahapan perkembangan folikel. Variasi bentuk kerusakan terlihat pada oosit dan sel granulosa yang mengelilinginya. Sel-sel mengalami perubahan berupa kariolisis (sel-sel yang kehilangan inti), nukleus yang bergranulasi, zona pelusida yang hilang, pembengkakan sel granulosa, inti sel granulosa terlihat mengecil, batas dan bentuk oosit yang tidak jelas, sel-sel granulosa yang pipih, dan susunan sel granulosa yang tidak kompak. Keseluruhan degenerasi tersebut mengarah pada kematian sel dengan berbagai tingkat proses kematian. Kerusakan folikel primer hingga tersier disebabkan oleh akumulasi ekstrak biji kapas yang mengakibatkan gangguan pada permeabilitas membran (Brocas et al. 1997). Hal ini berdampak pada terganggunya transpor nutrien yang diperlukan sel-sel selama folikulogenesis (Basini et al. 2008), hambatan proliferasi sel (Moon 2003), dan stimulasi produksi peptida proangiogenik sehingga terjadi peningkatan mekanisme apoptosis pada sel granulosa (Basini et al. 2008). Banyaknya jumlah folikel yang mengalami degenerasi maka dilakukan evaluasi terhadap ratio nilai hitungan kuantitatif pada histomorfologi ovarium. Hasil evaluasi histomorfologi tersebut disajikan pada Tabel 4.
Tabel 4 Jumlah folikel berkembang pada mencit setelah pemberian ekstrak biji kapas
Lama
pemberian Kontrol 1.5 g kg -1
BB 2.1 g kg-1 BB 2.7 g kg-1 BB 5 hari 24 ± 4.5de 33 ± 10.4bcde 42 ± 10.0abc 23 ± 3.6de 10 hari 19 ± 8.5e 36 ± 4.5bcde 31 ± 4.9bcde 23 ± 5.0de 15 hari 33 ± 7.4bcde 30 ± 12.7bcde 37 ± 12.5 bcd 19 ± 12.4e 24 hari 34 ± 12.3 bcde 25 ± 10.4de 27 ± 3.5cde 25 ± 10.1de 34 hari 44 ± 7.6ab 53 ± 10.8a 29 ± 6.2bcde 36 ± 6.0bcde Keterangan: huruf kecil yang berbeda di antara kelompok perlakuan di atas menunjukkan
22
Tabel 4 menyajikan jumlah total folikel berkembang dalam ovarium setelah pemberian ekstrak biji kapas. Jumlah folikel berkembang setelah pemberian ekstrak biji kapas selama 5, 10, dan 15 hari menunjukkan bahwa mencit yang diberi ekstrak biji kapas dengan dosis 2.7 g kg-1 BB memiliki jumlah folikel lebih rendah (P < 0.05) dibandingkan dengan dosis 1.5 dan 2.1 g kg-1 BB. Efektivitas ekstrak biji kapas dengan dosis 2.7 g kg-1 BB sama pada pada pemberian 5, 10, 15, dan 24 hari. Berdasarkan hasil tersebut diketahui bahwa pemberian ektrak biji kapas paling efektif selama 5 hari dengan dosis 2.7 g kg-1 BB. Sedangkan, pemberian ekstrak biji kapas selama 24 hari menunjukkan jumlah folikel berkembang rendah (P < 0.05) pada semua dosis perlakuan dibandingkan dengan kelompok pemberian ekstrak biji kapas selama 5, 10, dan 15 hari. Perbedaan jumlah folikel berkembang dapat disebabkan lama waktu pemberian dan dosis yang diberikan (Camara et al. 2015; Sirard et al. 2006).
Penurunan jumlah folikel setelah pemberian ekstrak biji kapas disebabkan oleh banyaknya folikel yang mengalami degenerasi sehingga menjadi atresia (Gambar 8). Gadelha et al. (2014a) juga melaporkan bahwa pemberian gosipol secara subkutan dengan dosis 25 mg kg-1 hari-1 juga menyebabkan banyak folikel yang mengalami atresia. Atresia pada folikel besar akan menyebabkan kerusakan pada lapisan kumulus yang akan mengakibatkan gangguan perkembangan embrio (Sirard et al. 2006). Selain itu folikel atresia akan menghasilkan oosit yang sulit menyelesaikan metafase II (Nasim et al. 1995). Gosipol merusak kompetensi meiosis yang diperoleh pada saat perkembangan folikel. Hal ini berdasarkan pernyataan Cecconi (2002) bahwa kemampuan meneruskan meiosis diperoleh bersamaan dengan pembentukan antrum. Pemberian ekstrak biji kapas walaupun memengaruhi ovarium tetapi tidak menyebabkan perubahan berat badan yang mengindikasikan tanda-tanda toksisitas. Hasil ini sesuai dengan penelitian pada domba yang diteliti oleh Camara et al. (2015).
Penghentian pemberian ekstrak biji kapas selama 10 hari menunjukkan jumlah folikel berkembang mengalami peningkatan (P < 0.05) yang dibandingkan dengan jumlah folikel berkembang setelah pemberian selama 24 hari. Pada dosis kontrol (0 g kg-1 BB) dan 1.5 g kg-1 BB menunjukkan jumlah yang lebih tinggi (P < 0.05) dibandingkan dengan dosis 2.1 dan 2.7 g kg-1 BB. Berdasarkan hal tersebut diketahui bahwa terjadi pemulihan ovarium yang dilihat dari jumlah folikel berkembang setelah penghentian pemberian ekstrak terjadi hingga dosis 2.7 g kg-1 BB. Efek pemulihan tersebut paling baik terlihat setelah penghentian pemberian ekstrak biji kapas pada dosis paling rendah yaitu 1.5 g kg-1 BB.
23
Gambar 9 Rata-rata jumlah folikel pada potongan histologi setelah pemberian ekstrak biji kapas. a folikel primer, b sekunder, dan c tersier.
kontrol, 1.5 g kg-1 BB, 2.1 g kg-1 BB, 2.7 g kg-1 BB. Huruf kecil yang berbeda pada setiap histogram di antara kelompok perlakuan di atas menunjukkan perbedaan nyata (P < 0.05)
Data pada Gambar 9 menunjukkan masing-masing populasi folikel primer (a), sekunder (b) dan tersier (c). Persentase populasi folikel primer setelah pemberian ekstrak biji kapas selama 15 dan 24 hari lebih rendah (P < 0.05) dibandingkan dengan 5 dan 10 hari. Jumlah folikel primer setelah pemberian ekstrak biji kapas selama 15 hari dengan dosis 2.7 g kg-1 BB memiliki persentase jumlah folikel yang sama dengan 24 hari pada dosis 2.1 dan 2.7 g kg-1 BB. Penghentian pemberian ekstrak selama 10 hari menunjukkan terjadi peningkatan jumlah folikel primer pada semua dosis perlakuan.
Persentase populasi folikel sekunder dan tersier belum bisa diduga karena jumlahnya yang fluktuatif (P < 0.05) antara kelompok kontrol dan perlakuan. Hasil pengamatan tersebut berbeda dengan penelitian pada jumlah folikel besar pada sapi yang menerima 5 g gosipol per ekor setiap hari yang mengalami penurunan kuantitas (Randel et al. 1996). Keberadaan folikel tersier tidak jelas berubah dari jumlahnya akibat pemberian ekstrak biji kapas namun pada perluasan perkembangan kumulus yang menjadi lapisan oosit pada folikel besar mengalami hambatan karena pemberian gosipol (Lin et al. 1994b).
Data pada Gambar 9 memperlihatkan folikel primer ditemukan dalam jumlah lebih sedikit dibandingkan folikel sekunder setelah pemberian ekstrak biji kapas. Hal tersebut mengakibatkan jumlah folikel yang direkrut dan diseleksi untuk menjadi folikel dominan berkurang. Penurunan tersebut berdampak pada jumlah ovulasi selanjutnya. Folikel sekunder setelah pemberian ekstrak biji kapas masih banyak ditemukan, namun pada proses perkembangan menjadi folikel tersier tidak
24
banyak folikel yang mampu terus berkembang. Folikel yang telah berkembang menjadi folikel sekunder kemungkinan telah atresi (Camara et al. 2015).
Akumulasi gosipol memengaruhi jumlah folikel yang sedang aktif berkembang (Gadelha et al. 2014b). Folikel primordial dan primer membutuhkan banyak nutrisi untuk melanjutkan perkembangan sehingga apapun zat yang diberikan akan memengaruhi perkembangan folikel tersebut. Gosipol bersifat sitotoksik (Randel et al. 1992) dan berkemampuan meningkatkan pembentukan
reactive oxygen species (ROS) sehingga menginduksi stres oksidatif (Kovaci 2003; Fornes et al. 1993) serta terjadi apoptosis pada folikel (Moon et al. 2011). Folikel pada masa awal perkembangan hanya dipengaruhi oleh nutrisi dan bebas dari pengaruh gonadotropin. Hal tersebut mengakibatkan perekrutan folikel primer menjadi lebih sedikit dibandingkan folikel sekunder dan tersier yang perkembangannya dipengaruhi oleh stimulasi gonadotropin (Baerwald 2009). Banyaknya jumlah folikel yang mengalami degenerasi maka dilakukan evaluasi terhadap ratio nilai hitungan kuantitatif folikel atresia pada histomorfologi ovarium. Hasil evaluasi tersebut disajikan pada Gambar 10.
Gambar 10 Rata-rata jumlah folikel atresia setelah pemberian ekstrak biji kapas. kontrol, 1.5 g kg-1 BB, 2.1 g kg-1 BB, 2.7 g kg-1 BB. Huruf kecil yang berbeda pada setiap histogram di antara kelompok perlakuan di atas menunjukkan perbedaan nyata (P < 0.05)
Pada Gambar 10 menunjukkan jumlah folikel atresia setelah pemberian ekstrak biji kapas lebih tinggi dibandingkan dengan kontrol. Jumlah folikel atresia tersebut meningkat setelah 10 hari (P < 0.05) perlakuan pada dosis 1.5, 2.1, dan 2.7 g kg-1 BB. Penghentian pemberian ekstrak biji kapas selama 10 hari menunjukkan terjadi penurunan jumlah folikel atresia pada semua dosis perlakuan.
Jumlah folikel ovarium memengaruhi jumlah oosit dan kompetensi perkembangan oosit (Mossa et al. 2007). Pemberian ekstrak biji kapas berkaitan dengan lingkungan folikel yang dapat memengaruhi fertilitas betina (Sirard et al.
2006). Beberapa studi menyatakan kerusakan akibat gosipol juga disebabkan oleh hambatan sekresi adrenocorticotropic hormone (ACTH), hambatan pembentukan
cyclic adenosine monophosphate (cAMP) pada sel-sel luteal (Lin et al. 1994b), gangguan komunikasi interseluler (Hervé et al. 1996), gangguan metabolisme
0
5 hari 10 hari 15 hari 24 hari reversible
25 energi seluler, gangguan transpor ion melalui membran, dan penurunan steroidogenesis yaitu estrogen serta progesteron (Gadelha et al. 2014b).
5
SIMPULAN
26
DAFTAR PUSTAKA
Aerts JMJ, Bols PEJ. 2010. Ovarian follicular dynamics: a review with emphasis on the bovine species. Part I: Folliculogenesis and pre-antral follicle development. Reprod Dom Anim. 45:171-179.
Aguirre-Samudio AJ, de la Fuente-Juárez G, Orozco A, Márquez-Orozco MC. 2006. Histological changes of the ovary of adult mice exposed prenatally to flunitrazepam. Int J Morphol. 24(4):651-658.
Akinola OB, Oderinde OO, Adejumo AT, Bayode ED. 2006. Effect of cottonseed oil on oestrous cycle and ovulation in albino rats of wistar strain. Niger Postgrad Med. 13(3):203-205.
Akira. 1994. Gossypol inhibits aromatase activity in cultured porcine granulosa cells. Theriogenology. 41:1489-l497.
Baerwald AR. 2009. Human antral folliculogenesis: what we have learned from the bovine and equine models. Anim Reprod. 6:20-29.
Bai J. 2002. Inhibition of T-type Ca2+ currents in mouse spermatogenic cells by gossypol, an antifertility compound. Eur J Pharmacol. 440:1-6.
Bansode FW. 1994. Genotoxic effects of gossypol acetate on the ovary of
Rhinopoma kinneari wroughton (Microchiroptera: Mammalia).
Contraception. 49:601-607.
Basini G, Bussolati S, Baioni L, Grasseli F. 2008. Gossypol, a polyphenolic aldehyde from cotton plant, interferes with swine granulosa cell function.
Dom Anim Endocrinol. 37:30-36.
Braw-Tal R. 2002. The initiation of follicle growth: the oocyte or the somatic cells? Mol Cell Endocrinol. 187:11–18.
Brocas C, Rivera MR, Paula-Lopes FF, McDowell LR, Calhoun MC, Staples CR, Wilkinson NS, Boning AJ, Chenoweth PJ, Hansen PJ. 1997. Deleterious actions of gossypol on bovine spermatozoa, oocytes, and embryos. Biol Reprod. 57:901-907.
Camara ACL, Gadelha ICN, Borges PAC, de Paiva SA, Melo MM, Soto-Blanco B. 2015. Toxicity of gossypol from cottonseed cake to sheep ovarian follicles. PloS ONE. 10:1-11.
Cecconi S. 2002. Growth and differentiation of small ovarian follicles in mammals: problems and future perspective. Reprod Dev. 48:431-445. Champlin AK, Dorr DL, Gates AH. 1973. Determining the stage of the estrous
cycle in the mouse by the appearance of the vagina. Biol Reprod. 8:491-494. Chandrashekar R, Kumar AK, Reddy YR, Chaitanya PJ, Bhavani NL,
Pochampalli J. 2013. Isolation of gossypol and analysis of phytochemicals in seed extract of Bt and non Bt varieties of cotton. Pharm Phytochem. 2(1): 180-186.
Chavatte-Palmer P, Dupont C, Debus N, Camous S. 2014. Nutritional programming and the reproductive function of the offspring. Anim Product Sci. 54:1166–1176.
Cuellar A, Ramirez J. 1993. Further studies on the mechanism of action of gossypol on mitochondrial membrane. Int Biochem. 25(8):1149-1155. Eppig JJ. 2001. Oocyte control of ovarian follicular development and function in
27 Evans ACO, Mossa F, Walsh SW, Scheetz D, Jimenez-Krassel F, Ireland JLH, Smith GW, Ireland JJ. 2012. Effects of maternal environment during gestation on ovarian folliculogenesis and consequences for fertility in bovine offspring. Reprod Dom Anim. 47(4):31–37.
Findlay JK, Drummond AE, Fry RC. 1996. Intragonadal regulation of follicular development and ovulation. Anim Reprod Sci. 42:321-331.
Fornes MW, Barbieri AM, Burgos MH. 1993. Sperm motility loss induced by gossypol: relation with OH scavengers, motile stimulators dan malondialdehyde production. Biochem Biophys Res Commun. 195:1289-1293.
Gadelha ICN, Fonsesa NBS, Oloris SCS, Melo MM, Blanco SB. 2014a. Gossypol toxicity from cottonseed products. Sci World. 14:1-9.
Gadelha ICN, de Macedo MF, Oloris SCS, Melo MM, Soto-Blanco B. 2014b. Gossypol promotes degeneration of ovarian follicle in rats. Sci World. 14:1-7.
Gannon AM. 2013. Cigarette smoke exposure and its impact on ovarian follicles [disertation]. McMaster University.
Gilbert NE, O’Reilly JE, Chang CJG, Lin YC, Brueggemeier RW. 1995. Antiproliferative activity of gossypol and gossypolone on human breast cancer cells. Life Sci. 57(1):61-67.
Gu Y, Li PK, Lin YC, Rikihisa Y, Brueggemeier RW. 1991. Gossypolone suppresses progesterone synthesis in bovine luteal cells. Steroid Biochem Mol Biol. 38:709-715.
Hadley MA, lin YC, Dym M. 1981. Effect of gossypol on the reproductive system of male rats. Androl. 2:190-199.
Hafez ESE, Hafez B. 2000. Reproduction in Farm Animal: Folliculogenesis, Egg Maturation, and Ovulation. seventh edition. Philadelphia (US): Lippincot Williams and Walkins.
Hassan ME, Smith GW, Ott RA, Faulkner DB, Firkins LD, Ehrharte EJ, Schaeffer DJ. 2004. Reversibility of the reproductive toxicity of gossypol in peripubertal bulls. Theriogenology. 61:1171–1179.
Hervé J-C, Pluciennik F, Bastide B, Cronier L, Verrecchia F, Malassiné A, et al. 1996. Contraceptive gossypolblocks cell-to-cell communication in human and rat cells. Eur J Pharmacol. 313:243–255.
Irianto K. 2014. Biologi Reproduksi. Bandung (ID): Alfabeta Bandung.
Kovaci P. 2003. Mechanism of drug and toxic action of gossypol: focus on reactive oxygen species and electron transfer. Curr Medicin Chem. 10(24):2711-2718.
Kusumaningrum R. 2009. Faktor-faktor yang mempengaruhi pemilihan jenis kontrasepsi yang digunakan pada pasangan usia subur [laporan akhir penelitian karya tulis ilmiah]. Semarang (ID): UNDIP.
Lagerlof RK, Tone JN. 1985. The effect of gossypol acetic acid on female reproduction. Drug Chem Toxicol. 8(6):469-482.
Leung PCK, Adashi EY. 2004. The Ovary: Dynamics of Primate Follicular Growth: A Physiological Perspective. second edition. San Diego (US): Elsevier Academic Pr.