KERAGAMAN DAN STRUKTUR GENETIK
POPULASI JATI SULAWESI TENGGARA
BERDASARKAN MARKA MIKROSATELIT
DIRVAMENA BOER
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi ”Keragaman dan Struktur Genetik Populasi Jati Sulawesi Tenggara Berdasarkan Marka Mikrosatelit” adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam
bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang
berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari
penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di
bagian akhir disertasi ini.
Bogor, Juli 2007
ABSTRACT
DIRVAMENA BOER. Diversity and Genetic Structure of Southeast Sulawesi Teak Populations Based on Microsatellite Markers. Supervisors: SUDARSONO, SATRIYAS ILYAS, ASEP SETIAWAN, and EDI GUHARDJA.
The aim of this research was to study diversity and genetic population structure, dynamical aspect of gene flow through pollens and seeds dispersal and also mating system. This study was carried out at three locations of teak stand populations which indicated human disturbance level using ten markers microsatellite (AGT10, AC44, AC01, AG14, ATC02, AC28, AAG10, dan CPIMS). A total of 46 alleles were detected, with six the highest number allele at AAG10 and AG16 loci. The mean value of polymorphic information content (PIC) of the 10 loci ranged from 0.442 to 0.580.
Study of genetic similarity to compare between mature tree and seedling progeny showed the average genetic similarity was calculated based on Dice coefficient for total population of mature and juvenile tree were 51.91% and 54.55%, respectively. The mature tree of Dolok, Warangga and Sampolawa have 60%, 55%, and 73% genetic similarity, respectively. While seedling progeny of Dolok, Warangga, and Sampolawa have 56%, 61%, and 74% genetic similarity, respectively.
Study of genetic structure showed the heterozigosity Ha and He were high
(for Dolok population were 0.630 and 0.645, respectively). Genetic differentiation FST was 0.112 (11.2% of total genetic variation among population)
and showed less deviation from Hardy-Weinberg expectation (Wright’s inbreeding coefficient FIS = 0.009). However, genetic differentiation using AMOVA showed 14% of total variation among population, the remaining 86% occurred within populations. Cluster analysis calculated by Nei’s Distance showed Dolok and Warangga population joint in the same cluster.
Parentage analysis successfully detected candidate male parent as much as 30% for Sampolawa, 81% for Dolok, and 87% for Warangga progenies. Analysis parentage for juvenile tree in the field successfully detected 76% pairs of candidate male and female parents. The gene flow through pollen dispersal showed that pollens spread out to all directions by insect vector. Source of pollen for the teak pollination trees mainly came from the teak around the mother tree (30%). The distance average pollen source could pollinate 30.23-39.43 m and furthermost more than 80 m. Whereas, the genetic migration through seed dispersal showed that seedling from their expected parents occurred by the distance average of 34.27 m and the furthermost 68.73 m.
The mating system parameters showed the singlelocus outcrossing rate (ts)
varied among loci and populations, but multilocus outcrossing rates (tm) were
equal to one for Sampolawa and Warangga populations and so it is with biparental inbreeding (tm-ts) was different from zero for Sampolawa and Warangga
populations. Biparental inbreeding occured for Dolok population and parental inbreeding for Sampolawa population.
ABSTRAK
DIRVAMENA BOER. Keragaman dan Struktur Genetik Populasi Jati Sulawesi Tenggara Berdasarkan Marka Mikrosatelit. Komisi Pembimbing: SUDARSONO, SATRIYAS ILYAS, ASEP SETIAWAN, dan EDI GUHARDJA.
Penelitian ini bertujuan untuk mempelajari kemiripan genetik individu di dalam populasi tanaman jati, keragaman dan struktur populasi genetik serta aspek dinamiknya akibatnya adanya perpindahan informasi genetik via serbuk sari dan migrasi biji, serta sistem perkawinan pada tanaman jati. Dalam penelitian ini diambil tiga lokasi populasi tegakan jati yang mempunyai level kerusakan akibat adanya aktifitas manusia kemudian dipelajari menggunakan 10 marka mikrosatelit (AG04, AG16, AGT10, AC44, AC01, AG14, ATC02, AC28, AAG10, dan CPIMS) menghasilkan total 43 alel dengan rata-rata banyaknya alel per lokus 4.6 dengan kisaran alel mulai dari dua sampai tujuh alel. Tingkat polimorfisme tertinggi pada lokus AG16 sebesar 0.767, rata-rata untuk semua lokus adalah 0.522.
Hasil studi kemiripan genetik terhadap tanaman dewasa dibandingkan dengan benih yang diunduh dari beberapa pohon menunjukkan rata-rata kemiripan genetika yang dihitung menggunakan koefisien Dice pada populasi total tanaman dewasa dan tanaman semai asal Sulawesi Tenggara mempunyai kemiripan genetik berturut turut 51.91% dan 54.55%. Untuk tanaman dewasa kelompok Dolok, Warangga dan Sampolawa mempunyai kemiripan genetika berturut-turut 60%, 55% dan 73%. Tanaman semai kelompok Dolok, Warangga dan Sampolawa mempunyai kemiripan genetika berturut-turut 56%, 61%, dan 74%.
Analisis struktur genetik memperlihatkan bahwa nilai heterozigositas Ha
dan He mempunyai nilai yang tinggi (tertinggi untuk Dolok adalah 0.630 dan
0.645) dengan nilai He selalu lebih besar daripada Ha. Nilai diferensiasi genetik
FST adalah 0.112 atau 11.2% dari total keragaman genetik di antara populasi dan
memperlihatkan sedikit penyimpangan dari keseimbangan Hardy-Weinberg harapan (Wright’s inbreeding coefficient FIS=0.009). Akan tetapi diferensiasi genetik yang dihitung dengan AMOVA memperlihatkan 14% terjadi keragaman di antara populasi dan sisanya sekitar 86% terjadi dalam populasi. Analisis cluster yang dihitung menggunakan jarak genetik Nei menunjukkan bahwa populasi Dolok dan Warangga berada pada satu cluster.
Analisis sistem perkawinan berupa parameter sistem perkawinan diduga di bawah model perkawinan percampuran menggunakan software MLTR menunjukkan derajat penyerbukan silang lokus tunggal (ts) bervariasi di antara
lokus dan populasi, tapi derajat penyerbukan silang multilokus (tm) secara statistik
sama dengan satu untuk populasi Sampolawa dan Warangga demikian pula dengan koefisien biparental inbreeding (tm-ts) sama dengan nol untuk populasi
Sampolawa dan Warangga. Terjadi biparental inbreeding pada populasi Dolok dan parental inbreeding (f) pada Sampolawa. Hal ini menunjukkan bahwa walaupun derajat penyerbukan silang besar namun pada lokasi Dolok dan Sampolawa terjadi proses silang dalam.
© Hak cipta milik IPB, tahun 2007
Hak cipta dilindungi
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
KERAGAMAN DAN STRUKTUR GENETIK
POPULASI JATI SULAWESI TENGGARA
BERDASARKAN MARKA MIKROSATELIT
DIRVAMENA BOER
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Departemen Agronomi dan Hortikultura
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup : Dr. Ir. Hajrial Aswidinnoor, M.Sc.
Penguji pada Ujian Terbuka : 1. Dr. Ir. Ulfah J. Siregar, M.Agr.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala
karunia-Nya sehingga disertasi yang berjudul ”Keragaman dan Struktur Genetik Populasi
Jati Sulawesi Tenggara Berdasarkan Marka Mikrosatelit” dapat diselesaikan
dengan baik.
Disertasi ini disusun berdasarkan empat topik penelitian yaitu: (1) Analisis
kemiripan genetik (genetic similarity) tanaman jati Sulawesi Tenggara menggunakan marka mikrosatelit, (2) Analisis struktur genetik (genetic structure) populasi jati Sulawesi Tenggara menggunakan marka mikrosatelit, (3) Analisis
aliran gen via serbuk sari (gene flow) dan biji (migration) pada tanaman jati
Sulawesi Tenggara menggunakan marka mikrosatelit, dan (4) Analisis sistem
perkawinan (mating system) tanaman jati Sulawesi Tenggara menggunakan marka mikrosatelit.
Penulis menyampaikan ucapan terimakasih dan penghargaan yang
setinggi-tingginya kepada Prof. Dr. Ir. Sudarsono, M.Sc. selaku ketua komisi
pembimbing, Dr. Ir. Satriyas Ilyas, MS., Dr. Ir. Asep Setiawan, MS. dan Prof. Dr.
Ir. Edi Guhardja, M.Sc. yang masing-masing sebagai anggota komisi, atas semua
arahan, bimbingan dan motivasi yang diberikan kepada penulis mulai dari
perencanaan, pelaksanaan, sampai penyelesaian penulisan penelitian disertasi ini.
Penulis juga mengucapkan terimakasih kepada Dr. Ir. Hajrial Aswidinnoor, M.Sc.
(selaku penguji luar komisi pada ujian tertutup), Dr. Ir. Ulfah J. Siregar, M.Agr.,
dan Dr. Ir. Sugiono Moeljopawiro, M.Sc. (selaku penguji luar komisi pada ujian
terbuka), atas masukan yang diberikan demi kesempurnaan disertasi ini.
Ucapan terimakasih juga penulis sampaikan kepada Dekan dan Rektor
Universitas Haluoleo atas izin dan kesempatan yang diberikan kepada penulis
dalam mengikuti pendidikan program doktor di Sekolah Pascasarjana IPB.
Ucapan terimakasih yang sama ditujukan kepada Rektor IPB, Dekan Sekolah
Pascasarjana IPB dan staf, Ketua Program Studi Agronomi dan staf atas segala
motivasi dan pelayanan akademik hingga segala persyaratan program doktor di
IPB dapat dipenuhi.
Begitu juga ucapan terimakasih yang tak terhingga kepada Ketua
PSPT-IPB, ketua laboratorium BIORIN, PAU-IPB. atas kesempatan untuk melakukan
kegiatan penelitian dan pemakaian peralatan laboratorium. Serta seluruh staf dan
rekan-rekan teknisi atas segala kesempatan, bantuan dan pelayanan kepada
penulis berupa bahan dan peralatan yang diperlukan dalam penelitian ini.
Terimakasih penulis sampaikan pula kepada Yth: Prof. Dr. Ir. Didy
Sopandie, M.Agr. yang telah memimpin sidang dalam Ujian Terbuka dan Dr. Ir.
Aris Munandar MS yang telah memimpin sidang Ujian Tertutup sehingga dapat
berjalan lancar dan penuh hikmat.
Terimakasih juga penulis sampaikan kepada pengelola Beasiswa Program
Pascasarjana (BPPS) Direktorat Jenderal Pendidikan Tinggi Departemen
Pendidikan Nasional atas dukungan finansial yang diberikan berupa bantuan
beasiswa pendidikan program doktor selama tiga tahun, juga kepada pemberi dana
penelitian yaitu European Commission, Inco-Dev ICA4-CT-2001-10094 yang
diketuai oleh Prof. Dr. Ir. Sudarsono, M.Sc.
Terimakasih juga disampaikan kepada semua rekan-rekan mahasiswa
seperjuangan di Laboratorium Molekular Biologi Tanaman (PMB) IPB atas
kebersamaan dan kesempatan saling diskusi selama penelitian berlangsung. Juga
kepada semua rekan-rekan sesama mahasiswa Pascarjana IPB.
Tak lupa disampaikan ucapan terimakasih kepada ayahanda Drs.
Boerhanudin Jacoub. Dt.T (alm) dan ibunda Bayana Biran tersayang atas segala
ketulusan doa restu dan dorongan moril, juga kepada seluruh kakak dan adik atas
semangat dan doanya. Kepada istri tercinta dan anak-anak tersayang, atas segala
pendampingan, doa dan dorongan semangat untuk meraih sukses. Semoga Allah
SWT menjadikan suatu keberkahan dan manfaat atas segala keberhasilan yang
penulis capai saat ini.
Bogor, Agustus 2007
RIWAYAT HIDUP
Penulis dilahirkan di Bandung pada tanggal 17 Desember 1962 sebagai
anak keempat dari enam bersaudara dari keluarga Boerhanuddin Jacoub dan
Bayana Biran.
Pendidikan sarjana ditempuh di Program Studi Biologi, FMIPA, Institut
Pertanian Bogor, lulus pada tahun 1989 dengan gelar Ir. Pada tahun 1995 penulis
melanjutkan pendidikan S2 di Universitas Goettingen dan menamatkannya pada
tahun 1997 dengan gelar M.Sc.Agr. Kesempatan untuk melanjutkan ke program
doktor pada program studi Agronomi, pada perguruan tinggi IPB diperoleh pada
tahun 2000. Beasiswa pendidikan pascasarjana diperoleh dari Departemen
Pendidikan Nasional Republik Indonesia.
Penulis bekerja sebagai staf pengajar di Jurusan Budidaya Pertanian,
Fakultas Pertanian, Universitas Haluoleo di Kendari sejak tahun.1990 sampai
xii
DAFTAR ISI
Halaman
DAFTAR TABEL...xv
DAFTAR GAMBAR ... xvii
DAFTAR LAMPIRAN... xviii
DAFTAR SINGKATAN ...xix
GLOSARI ...xx
1. PENDAHULUAN ...1
Latar Belakang ... 1
Tujuan Penelitian ... 3
2. TINJAUAN PUSTAKA ...4
Tanaman Jati (Tectona grandis Linn.f.)... 4
Klasifikasi Jati ... 4
Daerah Penyebaran Jati ... 4
Karakter Vegetatif, Generatif, dan Pembungaan Jati... 5
Karakter Vegetatif ... 5
Karakter Generatif ... 6
Karakter Biologi Pembungaan ... 6
Marka Genetik... 7
DNA Mikrosatelit... 8
Daerah DNA Mikrosatelit ... 8
Keungulan DNA Mikrosatelit ... 9
Sistem Reproduksi Seksual Tanaman ... 10
Sistem Seksual... 10
Sistem Aliran Informasi Genetik (Gene Flow) ... 11
Sistem Perkawinan (Mating System) ... 11
3. METODE UMUM PENELITIAN ...14
Ruang Lingkup Penelitian... 14
Tempat dan Waktu ... 14
Bahan dan Metode Penelitian ... 14
Bahan Tanaman ... 14
Prosedur Molekular dengan Marka Mikrosatelit... 16
4. ANALISIS KEMIRIPAN DAN KERAGAMAN GENETIK TANAM-AN JATI SULAWESI TENGGARA MENGGUNAKTANAM-AN MARKA MIKROSATELIT (Genetic similarity and diversity analysis of teak from originated Southeast Sulawesi by using microsatellite markers)...20
Abstract ... 20
Abstrak ... 20
xiii
Bahan dan Metode ... 22
Material Tanaman... 22
Isolasi serta Penentuan Kualitas dan Kuantitas DNA ... 22
Amplifikasi PCR dan Gel Electroforesis... 23
Analisis Data ... 23
Hasil ... 24
Profil Pita Mikrosatelit ... 24
Kemiripan Genetika di dalam Populasi... 27
Pembahasan... 32
Kesimpulan dan Saran... 34
Daftar Pustaka ... 34
5. ANALISIS STRUKTUR GENETIK POPULASI JATI ASAL SULA-WESI TENGGARA MENGGUNAKAN MARKA MIKROSATELIT (Genetic structure analysis of Southeast Sulawesi teak populations based on microsatellite markers)...36
Abstract ... 36
Abstrak ... 36
Pendahuluan ... 37
Bahan dan Metode ... 38
Tempat dan Waktu ... 38
Bahan Tanaman ... 38
Analisis Data ... 38
Hasil ... 45
Pembahasan... 50
Kesimpulan dan Saran... 52
Daftar Pustaka ... 52
6. ANALISIS ALIRAN INFORMASI GENETIK VIA SERBUK SARI DAN PENYEBARAN BENIH TANAMAN JATI ASAL SULAWESI TENGGARA MENGGUNAKAN MARKA MIKROSATELIT (Gene flow via pollen and seed dispersal analysis of teak from Southeast Sulawesi by using microsatellite markers) ...54
Abstract ... 54
Abstrak ... 54
Pendahuluan ... 55
Bahan dan Metode ... 56
Material Populasi dan Ekstrasi DNA ... 56
Analisis Marka Mikrosatelit ... 57
Analisis Data ... 57
Hasil ... 58
Pembahasan... 63
Kesimpulan dan Saran... 64
xiv 7. ANALISIS SITEM PERKAWINAN TANAMAN JATI SULAWESI
TENGGARA MENGGUNAKAN MARKA MIKROSATELIT (Mating system analysis of teak from Southeast Sulawesi by using microsatellite
markers) ...66
Abstract ... 66
Abstract ... 66
Pendahuluan ... 67
Bahan dan Metode ... 68
Material Tanaman dan Isolasi DNA... 68
Analisis Penanda Mikrosatelit... 68
Analisis Data ... 69
Hasil ... 70
Pembahasan... 73
Kesimpulan dan Saran... 75
Daftar Pustaka ... 76
8. PEMBAHASAN UMUM...77
9. KESIMPULAN DAN SARAN ...81
DAFTAR PUSTAKA ...83
xv
DAFTAR TABEL
Halaman 3.1. Koordinat posisi geografis populasi jati dari Sulawesi Tenggara ...15
3.2. Nama dan sekuen primer mikrosatelit berasal dari project jati TEAKDIV ICA4-2000-20053...19
4.1. Pohon induk dan tanaman semai jati yang dikoleksi dipilih secara acak pada tiga lokasi populasi jati asal Sulawesi Tenggara dianalisis menggunakan 10 lokus mikrosatelit ...22
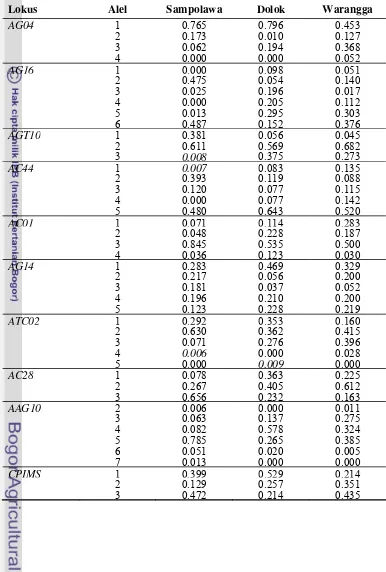
4.2. Jumlah alel dan polymorphic information content (angka yang di dalam tanda kurung) berdasarkan 10 lokus mikrosatelit pada populasi tanaman jati asal Sulawesi Tenggara ...25
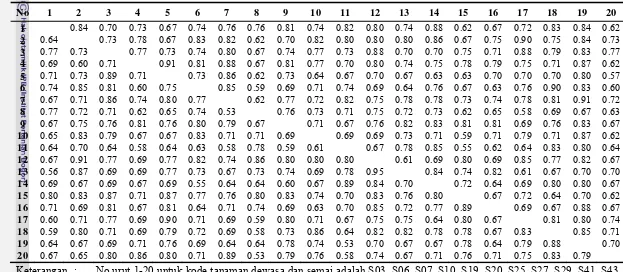
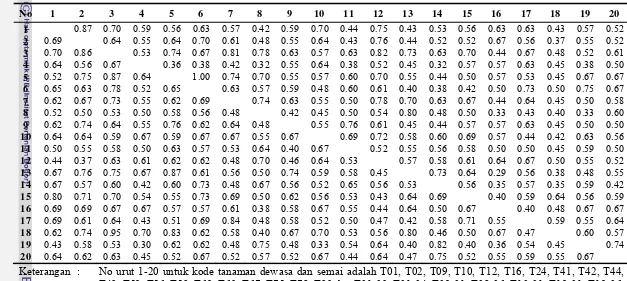
4.3. Matrik kemiripan genetik di antara 20 tanaman dewasa (di bawah diagonal) dan 20 tanaman semai (di atas diagonal) populasi jati asal Sampolawa ...28
4.4. Matrik kemiripan genetik 20 tanaman dewasa (di bawah diagonal) dan 20 tanaman semai (di atas diagonal) populasi jati asal Dolok...29
4.5. Matrik kemiripan genetik 20 tanaman dewasa (di bawah diagonal) dan 20 tanaman semai (di atas diagonal) populasi jati asal Warangga ...30
5.1. Jumlah alel dan tingkat polimorfisme 10 lokus mikrosatelit pada tanaman jati asal Sulawesi Tenggara...46
5.2. Frekuensi alel 10 lokus mikrosatelit pada tiga populasi jati asal Sulawesi
Tenggara ...47
5.3. Keragaman genetik jati berdasarkan nilai heterosigositas dan nilai FIS...48
5.4. Nilai F-statistik populasi jati asal Sulawesi Tenggara...48
5.5. AMOVA populasi jati asal Sulawesi Tenggara berdasarkan 10 lokus mikrosatelit ...49
6.1. Koleksi progeni famili half-sib jati dari 13-19 pohon induk benih serta semua tanaman jati yang diindentifikasi berpotensi sebagai sumber serbuk sari pada tiga lokasi yang memiliki level kerusakan akibat aktivitas manusia dianalisis menggunakan 10 penanda mikrosatelit ...57
xvi 7.2. Derajat penyerbukan silang berdasarkan lokus tunggal (ts) dan nilai
frekuensi serbuk sari (pollen) dan ovule dari alel yang sering muncul untuk tiga populasi jati asal Sulawesi Tenggara, Pop = populasi; A = banyaknya alel; f = koefisien parental inbreeding; SE = Standart Error; S = Sampolawa; T = Dolok; W = Warangga...72
7.3. Parameter sistem perkawinan dari tiga populasi jati Sulawesi Tenggara. Parameter yang diuji meliputi derajat outcrossing multilokus (tm), derajat
outcrossing rata-rata lokus tunggal (ts), biparental inbreeding (tm-ts),
koefisien parental inbreeding (f), korelasi t dugaan (rt), korelasi p
xvii
DAFTAR GAMBAR
Halaman 3.1. Peta lokasi tempat pengambilan sampel populasi jati di Kabupaten Muna
(Dolok dan Warangga) dan Kabupaten Buton (Sampolawa) ...15
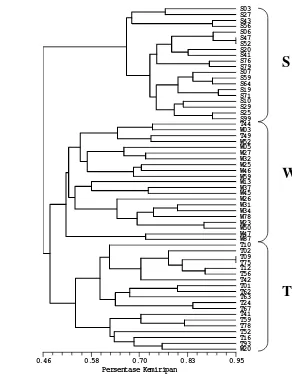
4.1. Dendrogram kemiripan genetika jati tanaman dewasa hasil analisis kluster dengan metode pengelompokan UPGMA berdasarkan 10 primer mikrosatelit hasil amplifikasi...26
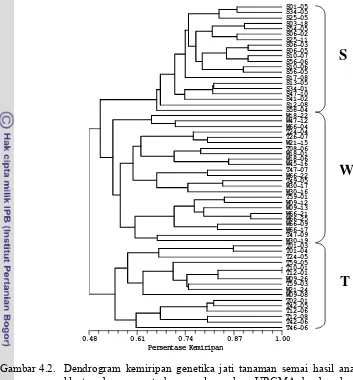
4.2. Dendrogram kemiripan genetika jati tanaman semai hasil analisis kluster dengan metode pengelompokan UPGMA berdasarkan 10 primer mikrosatelit hasil amplifikasi...27
4.3. Analisis komponen utama dari data kemiripan jati tanaman dewasa asal Sulawesi Tenggara berdasarkan 10 primer mikrosatelit hasil amplifikasi, yang dipetakan ke dalam bentuk tiga sumbu komponen utama yang pertama ...31
4.4. Analisis komponen utama dari data kemiripan jati tanaman semai asal Sulawesi Tenggara berdasarkan 10 primer mikrosatelit hasil amplifikasi, yang dipetakan ke dalam bentuk tiga sumbu komponen utama yang pertama ...31
5.1. Contoh profil pola pita lokus AC01 pada tanaman jati...45
5.2. Dendrogram jarak genetik antar populasi jati berdasarkan jarak genetik Nei ...50
6.1. Contoh profil pola pita lokus AG16 pada tanaman jati ...59
6.2. Perpindahan informasi genetik (gene flow) via serbuk sari ...59
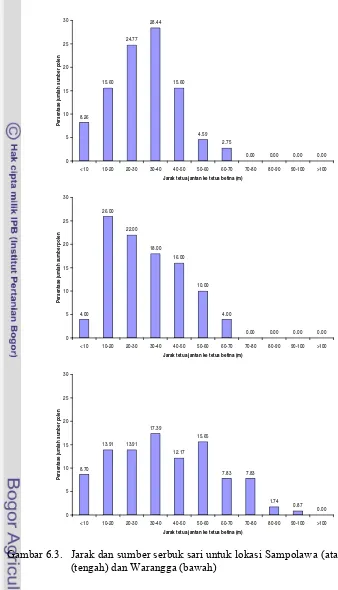
6.3. Jarak dan sumber serbuk sari untuk lokasi Sampolawa (atas), Dolok (tengah) dan Warangga (bawah)...60
6.4. Perpindahan informasi genetik melalui benih ...61
6.5. Analisis tetua (parentage analysis) via benih untuk populasi Dolok...62
xviii
DAFTAR LAMPIRAN
Halaman
1. Peta posisi relatif tanaman jati di lokasi Dolok ...88
2. Peta posisi relatif tanaman jati di lokasi Warangga...89
3. Peta posisi relatif tanaman jati di lokasi Wadila...90
4. Analisis tetua jantan terhadap famili half-sib turunan dari pohon induk benih jati asal populasi Sampolawa...91
5. Analisis tetua jantan terhadap famili half-sib turunan dari pohon induk benih jati asal populasi Dolok ...92
6. Analisis tetua jantan terhadap famili half-sib turunan dari pohon induk benih jati asal populasi Warangga ...94
7. Analisis tetua jantan terhadap tanaman juvenil (J) dari lapang untuk tanaman jati asal populasi Dolok...97
8. Prosedur isolasi DNA ...98
9. Prosedur elektroforesis polyacrylamide...108
10. Daftar bahan kimia yang dibutuhkan...115
xix
DAFTAR SINGKATAN
ADP : adenosine diphosphate
AFLP : Amplified Fragment Length Polymorphism AMOVA : analysis molecular of variance
ATGC : Adenine, Thymine, Guanine, dan Cytosine
bp : base pair
BT : Bujur Timur
cm : sentimeter
cpDNA : chloroplast DNA
cpSSR : chloroplast Simple Sequence Repeat
CTAB : hexadecyltrimethyl-ammonium bromide db : derajad bebas
dCTP : deoxy Cytidine Tri Phosphate dGTP : deoxy Guanidine Tri Phosphate DNA : deoxyribonucleid acid dNTP : deoxy Nucleotide Tri Phosphate EDTA : Ethylene Diamine Tetracetic Acid
F : Forward
HCl : asam chloride KCl : kalium chloride LS : Lintang Selatan LU : Lintang Utara
m : meter
MgCl2 : magnesium chloride
ml : mililiter
mM : milimolar
ng : nanogram
NH4 : methane
ns : non significant
NTSYS : numerical taxonomy system
o
C : derajat Celsius (centigrade) OD : optic dissociation P : probability
PCR : polymerase chain reaction pH : derajat kemasaman Pmol : piko mol
PVPP : polyvinylpolypyrrolidone.
R : reverse
RAPD : Random Amplified Polymorphic DNA RFLP : Restriction Fragment Length Polymorphism SAS : Statistical analysis system
SSR : Simple sequence repeats TBE : Trizma Boric EDTA
Tm : Temperature melting
xx
GLOSARI
Alel: Variasi gen pada satu lokus
Anelling: Temperatur untuk proses menempelnya primer pada utas cetakan Anemophily: Pergerakan gamet jantan (serbuk sari) dengan vektor berupa angin Apomiksis: Terjadinya biji tanpa penggabungan gamet jantan dan betina
Autochonous: Suatu populasi yang belum dipindahkan oleh manusia dalam hal ini jika tumbuh di habitat alami atau aslinya
Autogami: Tanaman menyerbuk sendiri
Biparental inbreeding, (tm - ts): Inbreeding yang terjadi karena perkawinan
kerabat dekat
Bootstrapping: Merupakan cara non-parametrik untuk mendapatkan nilai standar errror atau ragam dengan cara membuat set data ulangan dengan teknik sampling dari set data original dengan teknik pemulihan.
Cleistogami: Serbuk sari berasal dari bunga yang sama, fertilisasi terjadi saat bunga mekar
Differensiasi genetik: Perbedaan genetik antar populasi
Dioecy: Tanaman yang hanya memiliki bunga betina saja atau bunga jantan saja. Eksotik: Suatu populasi yang dipindahkan secara sengaja dan tumbuh di luar
penyebaran alaminya
Elektroforesis gel: Elektroforesis yang dikerjakan pada matriks gel sehingga molekul dengan muatan listrik yang sama dapat dipisahkan berdasarkan ukurannya.
Elektroforesis: Pemisahan molekul berdasarkan muatan listriknya
Etidium bromid: senyawa bersifat fluorescen yang dapat menyisip di antara pasangan basa dalam molekul DNA untai ganda, yang dipakai untuk deteksi DNA
Frekuensi alel: Proporsi tipe alel dari suatu lokus di dalam suatu populasi Frekuensi genotipe: Proporsi tipe genotipe di dalam suatu populasi
Galat lingkungan: Faktor kesalahan yang diperhitungkan akibat kemungkinan kesalahan waktu pemilihan tanaman, akibat dari faktor lingkungan yang dianggap tidak homogen.
Geitonogami: Serbuk sari berasal dari bunga berbeda pada tanaman yang sama Gen: Suatu segmen DNA yang mengkode molekul RNA dan atau molekul
polipeptida
Gene flow, aliran gen: Proses transportasi informasi genetik melalui transportasi serbuk sari (penyebaran gamet jantan) dan transportasi melalui benih.
Gene pool: All of the alleles present and carried by the population
Genetika: Suatu cabang ilmu dalam biologi yang mempelajari hal-hal yang berhubungan dengan gen.
Genom: Seperangkat lengkap gen dalam suatu organisme Half-sib: Keturunan dari garis ibu
Klon: Tanaman hasil perbanyakan secara vegetatif dimana tanaman tersebut merupakan duplikat yang susunan genetiknya sama.
Kromosom: Struktur pembawa sifat keturunan dan berada dalam nukleus.
xxi Mikrosatelit, simple sequence repeats, SSR: Merupakan unit pengulangan 1-6
pasangan basa. Primer SSR dibentuk berdasarkan kepada conserved flanking region (daerah pengapit konservatif) lokus SSR, yang mana bisa dihasilkan amplifikasi PCR pada lokus SSR tersebut. Hasil produk PCR bisa dielektroforesis yang dapat dibedakan menurut jumlah unit pengulangan dalam alel-alel SSR yang muncul.
Mixed mating model, model perkawinan campuran: Zigot yang terbentuk dapat berasal dari selfing atau outcrossing yang terjadi secara acak
Multilocus outcrossing rates, derajat penyerbukan silang multilokus, tm:
Derajat penyerbukan silang, yang dihitung berdasarkan pola variasi pada banyak lokus gen
Outcrossing rate, derajat outcrossing, t: Proporsi relatif dari ovul yang dibuahi oleh serbuk sari asing
Outcrossing, penyerbukan silang: Bila ovul dibuahi oleh serbuk sari dari tanaman lain
Parentage analysis, analisis tetua: Analisis untuk mendapatkan pasangan tetua dari setiap individu keturunan berdasarkan data genetik mereka.
Parental inbreeding, f: Inbreeding akibat dari selfing
Pemuliaan Tanaman: Suatu metode secara sistematis merakit keragaman genetik menjadi bentuk yang bermanfaat bagi kehidupan manusia.
Penguji (tester): Suatu tanaman atau barisan tanaman yang digunakan untuk mengevaluasi atau mengenal genotipe tanaman lain.
Plasmanutfah: Koleksi tanaman/gen yang merupakan material (bahan) keturunan.
Progeni: Keturunan
Protandri: Bunga yang benang sarinya lebih dulu matang daripada putik, bilamana putiknya masak, maka benangsarinya telah layu dengan semua tepungsari telah mati sehingga hampir selalu bunga tersebut mengalami penyerbukan silang.
Provenan: Merupakan contoh sejumlah pohon dari populasi yang telah beradaptasi secara evolusi di suatu habitat tertentu
Seleksi: Usaha untuk mendapatkan tanaman yang mempunyai sifat genetik yang baik, yaitu dengan cara memilihnya di antara tanaman lain dengan mengenali ciri-cirinya.
Selfing rate, derajat selfing, s: Proporsi relatif dari ovul yang dibuahi oleh serbuk sari sendiri atau kerabat dekat
Selfing, penyerbukan sendiri: Bila ovul dibuahi oleh serbuk sari sendiri atau kerabat dekat
Singlelocus outcrossing rate, derajat penyerbukan silang lokus tunggal, ts:
Derajat penyerbukan silang, yang dihitung berdasarkan pola variasi pada satu lokus gen
Tekanan lingkungan: Faktor pembatas dari lingkungan, yang mengakibatkan produksi tanaman menurun, misalnya pH, salinitas dan sebagainya.
Xenogami: Pembuahan terjadi bila serbuk sari dari satu tanaman sampai ke kepala putik yang receptive (siap dibuahi) dari tanaman lain dari jenis yang sama.
Latar Belakang
Jati (Tectona grandis Linn.f) merupakan salah satu jenis tumbuhan tropis yang sangat penting dengan reputasi internasional untuk sifat-sifat teknis dan
dekoratifnya. Sifat dekoratif kayu jati ditunjukkan oleh garis lingkar tumbuh yang
unik dan bernilai artistik tinggi, sehingga disukai konsumen. Selain itu kayu jati
termasuk kayu kelas awet I (mampu bertahan hingga 500 tahun) dan kelas kuat II
(Departemen Kehutanan Republik Indonesia, 1992), dan dari segi pengerjaannya,
kayu jati mudah dikerjakan serta punya berat jenis 0.78 sehingga jati termasuk
kelas kayu ringan dan terapung di atas air. Berdasarkan keunggulannya itu, kayu
jati dikelompokkan ke dalam jenis kayu mewah (fancy wood). Oleh karenanya di pasaran kayu jati termasuk kayu yang mahal harganya.
Kayu jati merupakan sumber penghasil devisa negara, sebagai gambaran
pada tahun 1981 volume kayu sebesar 18.000 m3 mempunyai nilai eksport sebesar
$ 10.943.000 dan pada tahun 1982 volume kayu sebesar 20.000 m3 mempunyai
nilai eksport sebesar $ 12.449.000 (Anonim, 1983). Pada tahun 1998, harga kayu
gelondongan di pasar luar negeri berkisar 9 sampai 15 juta rupiah per m3
(Sumarna, 2001).
Kebutuhan dalam negeri terhadap kayu jati sampai saat ini masih belum
terpenuhi. Dari kebutuhan sebesar 2.5 juta m3 per tahun, baru dapat dipenuhi oleh
Perum Perhutani sebesar 0.75 juta m3 per tahun, sehingga masih ada kekurangan
sekitar 1.75 juta m3. Walaupun permintaan dalam negeri masih belum terpenuhi
semua, namun kayu jati Indonesia juga ikut mengisi pasar dunia. Beberapa
negara yang mengimpor kayu dari Indonesia seperti Amerika, Taiwan, Hongkong,
Korea, Uni Emirat Arab, dan Italia.
Pemanfaatan sumberdaya jati alam terus semakin meningkat diikuti
adanya usaha ekploitasi terhadap hutan jati. Apabila hal tersebut tidak diikuti oleh
adanya upaya penanaman kembali dikhawatirkan akan mengancam punahnya
keragaman genetik plasmanutfah tanaman jati yang semenjak diintroduksikan ke
Sementara penanaman jati memerlukan investasi yang tinggi serta membutuhkan
waktu yang lama yaitu 60 tahun dan produksi optimumnya pada umur 80 tahun.
Sejalan dengan alternatif pemecahan masalah mengenai informasi potensi
keragaman genetik jati yang tersebar di seluruh kawasan Indonesia maka
diperlukan penelitian yang nantinya dapat digunakan oleh para pemulia jati untuk
meningkatkan potensi produksi jati sekaligus mengkonservasi hutan jati yang
keberadaanya terancam punah. Untuk konservasi dan program pemuliaan jati
diperlukan informasi keragaman genetik, struktur populasi genetik, serta aspek
dinamik gene flow dan sistem perkawinannya.
Sekarang ini terdapat beberapa metode molekular yaitu berdasarkan
polimorfisme DNA dalam memperoleh informasi keragaman genetik, struktur
populasi genetik serta aspek dinamik populasi genetik akibat adanya migrasi gen
(gene flow) yang disebabkan oleh perpindahan serbuk sari dan biji serta model sistem perkawinan. Metode molekular tersebut dikenal sebagai penanda (atau
marka) genetik pada tingkat DNA seperti RAPD, RFLP, AFLP dan mikrosatelit.
Keuntungan marka DNA adalah kemampuannya dalam menyediakan penanda
polimorfisme pola pita DNA dalam jumlah banyak, akurasi yang tinggi dan tidak
dipengaruhi lingkungan.
Marka mikrosatelit dibuat berdasarkan jumlah sekuen DNA sederhana
yang berulang-ulang sehingga sering disebut juga dengan simple sequence repeat
(SSR), merupakan salah satu penanda DNA yang menggunakan prinsip kerja reaksi polimerisasi berantai dengan menggunakan mesin PCR (Polymerase Chain
Reaction), yang dapat mengamplifikasi sekuen DNA tertentu secara in vitro. Sekarang ini mikrosatelit menjadi salah satu marka yang paling banyak digunakan
secara luas untuk pemetaan genetik, analisis keragaman, dan studi evolusi
(Temnykh et al., 2000).
Penggunaan penanda mikrosatelit mempunyai beberapa keunggulan
dibanding marka lainnya seperti (1) bersifat kodominan, (2) polimorfisme tinggi,
(3) lokus tersebar merata dalam genom dan dalam jumlah sangat banyak, dan (4)
dideteksi berbasis PCR sehingga diperlukan DNA dalam jumlah sedikit.
Marka mikrosatelit telah digunakan untuk menganalisis keragaman genetik
apel dan pear (Yamamoto, 2001), mempelajari struktur populasi genetik pada
kakao dan padi (Goran, 2000 dan Gao, 2002), serta untuk mempelajari sistem
perkawinan (mating system) dan gene flow pada pohon tropika (Collevatti, 2001).
Tujuan Penelitian
Penelitian molekular genetik dengan menggunakan penanda mikrosatelit
ini, bertujuan mempelajari (1) kemiripan genetik (genetic similarity) individu di dalam populasi tanaman jati, (2) keragaman dan struktur genetik (genetic
structure) populasi jati dikaitkan dengan aspek dinamika dari sistem genetik, (3) sistem aliran informasi genetik (gene flow) melalui serbuk sari dan biji serta (4)
Tanaman Jati (Tectona grandis Linn.f.) Klasifikasi Jati
Jati (Tectona grandis Linn. f.) adalah salah satu anggota famili Verbenaceae, Ordo Tubiflorae. Ada empat spesies yang tergolong dalam genus
Tectona yaitu Tectona grandis Linn. f, Tectona hamiltoniana Wall, Tectona philippinensis Benth and Hook. f. dan Tectona abludens (Hedegart, 1976).
Tectona grandis mempunyai beberapa nama seperti jati (Indonesia), teak (Inggris), lyiu (Burma), sagun (India), maisak (Thailand), teck (Perancis), teca
(Brasilia), birma, sian atau java teak (Jerman) (Samingan, 1991).
Daerah Penyebaran Jati
Tanaman jati merupakan tanaman asli daerah-daerah Asia Selatan dan
Tenggara, yang secara alami terdapat di India, Myanmar, Thailand dan bagian
barat Laos meliputi kisaran 9o LU di India sampai 25o LU di Myanmar dan antara
73o sampai 104o BT (Kaosa-ard, 1986).
Di Cina tanaman jati ditemukan di lembah sungai Jieyang bagian barat
Yunnan dalam jumlah yang sedikit kisaran 26o LU dan 98o BT (Kaosa-ard, 1995).
Pada abad 19 jati juga mulai ditanam di daerah tropis benua Amerika seperti
Trinidad dan Nicaragua. Akhir-akhir ini jati juga ditanam di Nigeria dan beberapa
negara Afrika tropik lainnya (Cordes, 1992).
Di Indonesia jati ditemukan terutama di Jawa, Kangean, Bali dan Muna.
Selain itu ditemukan pula di Buton, Maluku (Wetar), Sumbawa dan Lampung
berada pada posisi 7° LS dan 106° sampai 123° BT (Sastrosumarto dan Suhaendi,
1985). Menurut Wepf (1954) jati di Indonesia tumbuh secara alami di Jawa dan
Muna. Hutan jati di Jawa kebanyakan terdapat di Jawa Tengah dan Jawa Timur
dan terutama pada tanah kapur napal yang berasal dari zaman tersier di
tanah-tanah rendah 0 - 500 meter di atas permukaan laut. Persyaratan tumbuh lainnya
untuk jati adalah membutuhkan iklim musim yang nyata, yaitu dengan musim
kemarau yang periodik. Tanah yang beraerasi baik sangat dibutuhkan oleh jenis
tanaman ini, sedangkan ketinggian tempat tumbuh pada umumnya di bawah 700
(Departement Kehutanan Republik Indonesia, 1992). Indonesia memiliki luas
areal pertanaman jati yang relatif tinggi. Sampai tahun 1975, tercatat ada sekitar
774.000 hektar tanaman jati yang sebagian besar berada di Jawa, Sulawesi
Selatan, Sulawesi Tenggara, NTB, Maluku, dan Lampung (Sumarna, 2001).
Spesies jati di Indonesia diyakini merupakan introduksi dari India pada abad ke 14
(Kaosa-ard, 1986). Walaupun demikian jati yang ada di Jawa ternyata memiliki
variasi genetik yang berbeda dengan jati di India yang dianggap sebagai pusat
diversitas jati. Para ahli menyatakan bahwa spesies tersebut telah beradaptasi
dengan kondisi edafis dan iklim setempat sehingga membentuk vegetasi yang
berbeda.
Karakter Vegetatif, Generatif, dan Pembungaan Jati Karakter Vegetatif
Jati adalah salah satu jenis pohon berdaun lebar. Pada kondisi tempat
tumbuh yang sesuai, tinggi total dapat mencapai 30 - 40 m pada umur masak
tebang 70 - 80 tahun, sedangkan pada kondisi yang kurang baik pertumbuhannya
agak terhambat. Di daerah subur dengan kondisi lingkungan yang mendukung,
tinggi bebas cabang dapat mencapai 15 - 20 m atau lebih dan diameter dapat
mencapai 150 cm atau lebih (Departemen Kegutanan Republik Indonesia, 1992;
Keiding, 1985).
Daun jati berukuran relatif besar, panjangnya berkisar antara 25 - 50 cm
dan lebarnya 15 - 35 cm, berbentuk bulat telur dengan permukaan luar kasar.
Warna daun hijau sampai hijau tua dan kedudukan pada satu tangkai saling
bersilangan (Keiding, 1985).
Bentuk tajuk tak beraturan, menyerupai kubah, agak lebar dan termasuk
jenis menggugurkan daun (Departemen Kehutanan Republik Indonesia, 1992).
Batang umumnya bulat dan lurus, batang yang besar pada umumnya berbanir dan
berlekuk-lekuk; warna kulit agak kelabu muda, agak tipis dan beralur memanjang
agak dalam (Departemen Kehutanan Repubublik Indonesia, 1992).
Bagian vegetatif lain dari pohon jati yang perlu diketahui adalah sistem
perakarannya. Pada umumnya salah satu ciri dari perakaran jati adalah tidak
tanaman yang tahan terhadap kekurangan air untuk selang waktu 0 - 10 hari dan
jika lebih dari itu tanaman akan tumbuh merana dan mati.
Karakter Generatif
Bunga jati berukuran kecil (diameter 6 – 8 mm), berwarna keputihan
dengan tangkai yang pendek dan termasuk jenis biseksual atau hermaprodit, yaitu
dalam satu bunga terdapat putik dan benang sari. Jati juga disebut tanaman
berumah satu (monoecious), karena bunga jantan dan bunga betina ada pada pohon yang sama (Samingan, 1991). Tipe bunga jati adalah bunga majemuk tak
terbatas, yaitu dikenal dengan bentuk malai atau tandan majemuk. Ukuran malai
biasanya besar, terdiri atas ratusan kuncup bunga yang keluar secara serentak.
Kuncup ini akan bermekaran dalam waktu yang cukup singkat yaitu dalam selang
2 - 4 minggu (Keiding, 1985).
Menurut Kaosa-ard (1995) dan Cordes (1992) bunga jati merupakan bunga
sempurna yang terdiri atas bagian kelopak bunga berwarna coklat terang,
berbentuk lonceng, dan tidak terpisah pada bagian bawahnya, yang berfungsi
untuk melindungi kuncup bunga selama tahap perkembangannya. Mahkota bunga
berwarna keputih-putihan, berbentuk tabung sama panjang dengan kelopaknya,
yang berfungsi merangsang penyerbukan. Mahkota bunga terdiri atas enam buah
petal pada bagian tabung mahkota yang mengandung sedikit nectar pada bagian
basal. Di sebelah mahkota ada enam stamen dan masing-masing terdiri atas
sebuah filament dan sebuah antera bercaping dua (two-lobed) berwarna kuning. Pada bagian tengah ada sebuah pistil yang terdiri atas sebuah ovari, sebuah stilus
bercabang dua dan berbulu halus. Pada ujung masing-masing stilus terdapat
sebuah stigma.
Karakter Biologi Pembungaan
Pembungaan pada tanaman jati terjadi tidak seragam, tergantung pada
tempat tumbuh, iklim, sistem silvikultur serta faktor genetik (Keiding, 1985).
Pada daerah dengan iklim yang memungkinkan tanaman jati pada saat fase
juvenilnya tumbuh cepat, pembungaan terjadi setelah tanaman berumur 2 tahun,
(tempat tumbuh alaminya), pembungaan terjadi setelah tanaman berumur 6 - 8
tahun, malah kadang-kadang lebih 1ambat lagi (Keiding, 1985).
Pembungaan mulai terjadi pada bulan Juni sampai dengan Oktober dan
berbuah antara bulan Mei sampai dengan Desember tahun berikutnya, terutama
Juni sampai September (Departement Kehutanan Republik Indonesia, 1992).
Penyerbukan secara alami pada bunga jati terutama dengan bantuan serangga,
walaupun dengan bantuan angin dapat terjadi (Bryndum dan Hedegart, 1969). Di
Thailand, Heriodes biparvula dan Ceratina hieroglyphica yang menjadi penyerbuk utama (Hedegart, 1976). Meskipun demikian berbagai macam lalat
dan kupu-kupu juga terlibat (Bryndum dan Hedegart, 1969).
Persentase pembuahan pada jati dalam penyerbukan alami umumnya
rendah, karena tidak cukupnya serangga penyerbuk (Hedegard, 1973) dan
proporsi self incompatibility tinggi yaitu dapat mencapai 96 - 100% (Hedegart, 1976). Pembuahan terjadi 24 jam setelah penyerbukan, dan zigot mulai
berkembang pada 3 - 5 hari setelah penyerbukan. Buah mencapai ukuran penuh
sekitar 50 hari setelah terjadi penyerbukan terkendali, tetapi mencapai tingkat
kematangan penuh setelah berumur 120 – 200 hari setelah penyerbukan. Tanda
buah sudah masak adalah jika pohon ranting digoyangkan maka buah akan jatuh,
atau jatuh sendiri ke tanah (Hedegart, 1976).
Marka Genetik
Terdapat tiga jenis marka genetik yang dapat digunakan untuk analisis
genom dan analisis populasi seperti mendeteksi keragaman genetik suatu individu
di dalam atau antar populasi yaitu marka morfologi, marka biokimia dan marka
molekular. Marka morfologi adalah penanda yang dapat diamati secara langsung
seperti jumlah anakan, karakteristik batang, daun, bunga, buah, biji, dan lain
sebagainya. Keuntungan dari penanda jenis ini adalah pengamatannya mudah,
namun demikian penanda ini memiliki kelemahan karena dapat dipengaruhi oleh
tahap perkembangan tanaman dan lingkungan serta jumlahnya sangat terbatas,
sehingga kadangkala tidak dapat dibedakan antar genotipe yang diamati karena
secara morfologi kelihatannya sama, tetapi sebenarnya berbeda akibat adanya
sangat terbatas dan mempunyai tingkat polimorfisme yang relatif rendah, serta
ekspresinya sedikit dipengaruhi oleh lingkungan.
Marka molekuler seringkali juga dikenal sebagai sidik jari DNA karena
mengacu pada pita polimorfisme berupa fragmen DNA. Keunggulan utama
penanda molekular adalah (1) keakuratan yang tinggi dan tidak dipengaruhi oleh
lingkungan yang mempengaruhi ekspresi gen tersebut, (2) dapat diuji pada semua
tingkat perkembangan tanaman, (3) pada pengujian ketahanan hama dan penyakit
tidak tergantung pada organisme pengganggu tersebut, dan (4) seleksi pada
tingkat genotipe ini dapat mempercepat proses seleksi dan hemat pada pengujian
selanjutnya di lapang.
Analisis genetik berdasarkan penanda molekular dapat dilakukan dengan
hibridisasi fragmen DNA dengan penanda DNA pada teknik non-PCR seperti RFLP (Restriction Fragment Length Polymorphism) (Serret et al., 1997), dengan amplifikasi fragmen DNA dalam mesin PCR (Polymerase Chain Reaction) seperti
RAPD (Random Amlified Polymorphic DNA) (William et al., 1990), AFLP (Amplified Fragmen Length Polymorphisms), SSR (Simple Sequence Repeat) dan lain-lain.
DNA Mikrosatelit
DNA mikrosatelit merupakan rangkaian pola nukleotida antara dua sampai
enam pasang basa yang berulang secara berurutan. DNA mikrosatelit biasa
digunakan sebagai penanda genetik untuk menguji kemurnian galur, studi
filogenetik, lokus pengendali sifat kuantitatif dan forensik.
DNA mikrosatelit diamplifikasi menggunakan teknik PCR dengan beberapa pasang primer mikrosatelit. DNA produk PCR dideteksi menggunakan teknik elektroforesis gel poliakrilamida (PAGE) yang dilanjutkan dengan
pewarnaan perak.
Daerah DNA Mikrosatelit
DNA genom terdiri atas DNA ruas khas (specific sequence) dan DNA ruas berulang (repetitive sequence). DNA ruas berulang dalam genom eukariot dapat mencapai > 90% DNA total yang ada dalam genom tanaman (Weising et al.,
pula proporsi DNA ruas berulangnya. Dari beberapa hasil penelitian diketahui
proporsi DNA ruas berulang dalam genom tanaman jagung mencapai 60% (Gupta
et al., 1984), gandum dan kerabat liarnya mencapai 70% (Flavel, 1980), kedelai mencapai 60% (Walbot dan Goldberg, 1979), dan pada padi mencapai 50%
(McCouch et al., 1988).
DNA ruas berulang dapat berupa DNA yang ruasnya berulang secara
moderat atau berselang-seling (intersperse), dan DNA yang ruasnya berulang secara berurutan (tandem). Perulangan ruas DNA secara tandem diklasifikasikan berdasarkan panjang dan jumlah ruas berulang di dalam genom, dan dapat berupa:
(1) DNA satelit yang biasanya jarang ditemukan dalam lokus genom, karena
perulangannya sangat tinggi (biasanya antara 1000 sampai 100000 kopi) dan
bentuknya sangat panjang, sering berada pada bagian heterokromatin dengan
panjang fragmen berulang antara 2 sampai beberapa ribu pasang basa tetapi
umumnya ditentukan antara 100-300 pasang basa, (2) minisatelit yang biasanya
banyak terdapat dalam genom, bentuknya lebih pendek (10 - 60 bp) dan
menunjukkan derajat perulangan yang lebih rendah, (3) fragmen sederhana atau
biasa disebut mikrosatelit (disebut juga fragmen berulang sederhana atau
perulangan tanden sederhana), ruas perulangannya pendek dan derajat
perulangannya lebih sedikit, terdistribusi lebih banyak pada lokus genom, dan (4)
midisatelit untuk kategori DNA yang tipenya merupakan kombinasi dari satelit
(perulangan yang panjang pada lokus genom tunggal) dan minisatelit (fragmen
berulang secara tandem kira-kira 40 bp) (Weising et al., 1995).
Pola ulangan DNA mikrosatelit terdiri atas pola di-, tri-, atau
tetranukleotide berulang. Pola ini ditemukan dalam semua organisme, baik
prokariot maupun eukariot. Ulangan dinukleotide yang paling sering ditemukan
pada tanaman adalah AA/TT dan AT/TA, sedangkan pada hewan GT/AC
(Hoelzel, 1998).
Keungulan DNA Mikrosatelit
Hoelzel (1998) menyebutkan bahwa DNA mikrosatelit merupakan salah
satu penciri genetik yang ideal untuk analisa genom karena jumlahnya cukup
banyak di dalam genom. DNA mikrosatelit memiliki tingkat ulangan 5-100 tiap
sehingga polimorfismenya lokus tersebut dapat diketahui berdasarkan jumlah
ulangannya yang berbeda. DNA mikrosatelit memiliki jumlah lokus yang banyak
(104 - 105) dengan tingkat variabilitas yang tinggi dalam hal jumlah salinan
berulangnya (Tautz, 1993). Tingkat polimorfismenya tinggi serta mudah untuk
dianalisis, misalnya dengan teknik PCR (Ellegreen et al., 1993). DNA
mikrosatelit lebih sedikit dalam penggunaan enzim dan runutan inti lokus dapat
digunakan sebagai probe dibandingkan penciri genetik lainnya sehingga akan
mengurangi biaya (Winaya, 2000). Alel-alel DNA mikrosatelit berekspresi dalam
bentuk kodominan. Sifat tersebut memungkinkan untuk membedakan antara
individu-individu homozigot dari heterozigotnya (Wright dan Benzen, 1994).
Sistem Reproduksi Seksual Tanaman
Proses reproduksi seksual pada tanaman merupakan proses penggabungan
gamet jantan dan betina yang akan menghasilkan zigot. Pada proses reproduksi
seksual ini akan terbentuk berbagai macam kombinasi gen sehingga menghasilkan
turunan dengan berbagai genotipe. Dengan demikian reproduksi seksual akan
menghasilkan sejumlah besar turunan yang berbeda secara genetik. Fase
reproduksi selama perkembangan populasi merupakan hal yang menarik dari sisi
genetik karena fase ini menyebabkan terjadinya perubahan yang dinamis dari
struktur genetik dalam populasi tanaman (Finkeldey, 2005). Aspek dinamik
struktur populasi genetik sangat penting dipelajari terutama dalam program
breeding dan untuk koservasi tanaman tersebut (Yeh, 2000).
Sistem reproduksi seksual terdiri atas beberapa sub-sistem yang berbeda
yaitu sistem seksual, aliran gen (gene flow), dan sistem perkawinan (mating
system).
Sistem Seksual
Sistem seksual pada tanaman sangat beragam, yaitu kemampuan tanaman
dalam suatu populasi untuk dapat saling berkawin, tergantung dari kemampuan
tanaman dalam menghasilkan gamet jantan (serbuk sari atau polen) atau gamet
betina (sel telur atau ovul) atau keduanya. Secara umum sistem seksual ini
betina terletak pada tanaman yang berbeda, (ii) hermaprodit dimana
individu-individu dari suatu spesies mempunyai gamet jantan dan betina dalam bunga yang
sama, dan (iii) monoecious dimana individu memiliki bunga jantan dan betina terpisah dalam satu tanaman (Boshier, 2000)
Tanaman dioecious tidak mungkin kawin sendiri (self pollination)
sehingga derajat inbreeding dalam populasi rendah. Mayoritas spesies tanaman dalam hutan tropikal adalah hermaprodit, sedangkan di hutan temperet spesiesnya monoecious. Pada hutan tropical sistem polinasi bunga hermaprodit dominan terjadi disebabkan oleh bantuan hewan terutama serangga (Gailing et al., 2003)
Sistem Aliran Informasi Genetik (Gene Flow)
Gene flow adalah perpindahan informasi genetik melalui serbuk sari dan biji. Jadi gene flow bertanggung jawab terhadap distribusi informasi genetik dalam populasi (intrapopulasi gene flow) dan antar populasi (interpopulation gene
flow). Efisiensi gene flow melalui serbuk sari dan biji merupakan hal yang sangat penting berkenaan dengan ukuran populasi efektif dalam bereproduksi. Ukuran
populasi sangat penting dalam menggambarkan pola penyebaran keragaman
genetik dan perbedaan genetik di antara populasi (Hamrick, 2000 dan Gailing et al., 2003).
Banyaknya tanaman, jarak serta arah gene flow melalui serbuk sari dalam tegakan menentukan apakah perkawinan yang terjadi antar tanaman yang berbeda
(outcrossing) atau dalam tanaman itu sendiri (selfing). Oleh karena itu, gene flow bila dikaitkan dengan tipe perkawinan dan segala aspek-aspeknya merupakan hal
yang sangat menarik dalam menganalisis sistem perkawinan (mating system). Dalam sistem pembungaan tanaman ada dua tipe transpor serbuk sari yaitu
melalui angin (anemophily) dan hewan (zoophily). Namun polinasi pada hutan
tropis terutama disebabkan oleh serangga (entomophily) (Hamrick, 2000 dan Gailing et al., 2003).
Sistem Perkawinan (Mating System)
Tipe mating system sangat menentukan struktur genetik dalam suatu populasi. Tujuan dari analisis mating system adalah untuk menentukan asal gamet
populasi sangat tergantung pada faktor lingkungan seperti densitas populasi, tata
letak tanaman, fenologi, dan vektor polinasi (Collevatti, 2001).
Pendugaan laju selfing dan outcrossing sangat dipengaruhi oleh kondisi lapang. Untuk menghindari selfing dikembangkan berbagai mekanisme seperti sistem inkompatibilitas, perbedaan waktu pembungaan pada bunga jantan dan
betina, terpisahnya bunga jantan dan betina. Selfing merupakan proses inbreeding dalam penurunan heterozigositas. Hal tersebut dapat menyebabkan penurunan
viabilitas dalam turunannya berkaitan dengan peningkatan kerusakan alel-alel
dalam genotipe-genotipe homosigositas. Khususnya untuk spesies yang langka
dengan densitas rendah, selfing dapat mempercepat kepunahan. Jadi ukuran laju
selfing dapat digunakan untuk mengembangkan prioritas dan strategi program konservasi (Gailing et al., 2003).
Proporsi ovul yang terbuahi secara selfing disebut laju selfing (s) dan frekuensi ovul yang dibuahi secara outcrossing disebut laju outcrossing (t). Dimana s + t = 1. Nilai laju outcrossing berkisar dari t = 0 (100% selfing) sampai
t = 1 (100% outcrossing). Nilai-nilai yang signifikan kurang dari t=1 mengindikasikan terjadi inbreeding yang disebabkan oleh selfing atau perkawinan antar saudara. Struktur tata letak tanaman, komposisi tanaman dan densitas
populasi, serta adanya variasi pembungaan dan mekanisme inkompatibilitas dapat
menghasilkan variasi dalam laju outcrossing pada tegakan dan individu tanaman
(Gailing et al., 2003).
Mating system dapat dipelajari bila terdapat tipe alel atau genotip yang unik dalam populasi, dan tujuan ini dapat dicapai dengan tersedianya marka
molekular dengan polimorfik yang tinggi seperti mikrosatelit atau paling sedikit
terdapat satu alel unik pada tanaman dalam suatu populasi (Boshier, 2000).
Koefisien inbreeding atau indek fiksasi adalah ukuran deviasi antara pengamatan struktur genotipe dan struktur genotipik sesungguhnya di bawah
asumsi random mating, dan asumsi lainnya seperti tidak ada seleksi, mutasi,
migrasi, dan lain sebagainya. Koefisien inbreeding untuk setiap single lokus
dihitung dari perbedaan struktur genotipik berupa heterozigot yang diamat
terhadap heterozigot harapan pada kondisi keseimbangan Hardy-Weinberg, yang
Ruang Lingkup Penelitian
Disertasi ini disusun berdasarkan empat topik penelitian yaitu: (1) Analisis
kemiripan genetik (genetic similarity) tanaman jati Sulawesi Tenggara
menggunakan marka mikrosatelit, (2) Analisis struktur genetik (genetic structure) populasi jati Sulawesi Tenggara menggunakan marka mikrosatelit, (3) Analisis
aliran gen via serbuk sari (gene flow) dan biji (migration) pada tanaman jati Sulawesi Tenggara menggunakan marka mikrosatelit, dan (4) Analisis sistem
perkawinan (mating system) tanaman jati Sulawesi Tenggara menggunakan marka
mikrosatelit.
Tempat dan Waktu
Penelitian ini dilaksanakan di laboratorium Biologi Molekuler Tanaman
(PMB), Fakultas Pertanian, Institut Pertanian Bogor. Sampel daun dan benih dari
tanaman jati diperoleh terutama dari hutan jati di kabupaten Muna dan Buton,
Sulawesi Tenggara. Lama penelitian ini adalah 3 tahun, dimulai Mei 2003.
Bahan dan Metode Penelitian Bahan Tanaman
Populasi tegakan atau hutan jati yang dipilih adalah yang mempunyai level
kerusakan atau gangguan terhadap tegakan jati tersebut akibat adanya aktivitas
manusia. Untuk itu telah dipilih tiga lokasi populasi di Sulawesi Tenggara
(Gambar 3.1 dan Tabel 3.1) dengan berbagai level gangguan yaitu dua populasi
dari Kabupaten Muna dan satu populasi dari kabupaten Buton, sebagai berikut:
Lokasi Warangga terletak dekat kota Raha, Kabupaten Muna. Lokasi
tanaman jati di Warangga ini relatif terjaga dari penebangan liar. Tanaman jati di
lokasi ini ditanam sekitar tahun 1938. Lokasi ini sekarang dipertahankan dan
mulai dipagari untuk dijadikan kebun bibit.
Lokasi Dolok terletak di Tampo, Kabupaten Muna. Tanaman jati di lokasi
ini diperkirakan berumur 40 tahun. Lokasi ini relatif tidak terganggu oleh
Lokasi Wadila terletak di Gunung Sejuk, Sampolawa, kabupaten Buton.
Lokasi ini awalnya merupakan lokasi pertanaman jati yang pernah ditebang habis
pada tahun 1958, kemudian dibiarkan sampai sekarang, dan pada lokasi ini
kemudian tumbuh secara alami tanaman jati dan sekarang dikenal oleh
masyarakat sebagai jati alam.
Tabel 3.1. Koordinat posisi geografis populasi jati dari Sulawesi Tenggara
Populasi Lintang Bujur Lokasi
Dolok 4.63 LS 122.70 BT Pulau Muna
Warangga 4.84 LS 122.65 BT Pulau Muna Sampolawa 5.53 LS 122.68 BT Pulau Buton
Pengambilan sampel tanaman berupa daun muda serta buah jati ditujukan
untuk mempelajari keragaman genetik, struktur genetik serta aspek dinamika
akibat adanya perpindahan informasi genetik berupa aliran gen (gene flow) melalui serbuk sari dan biji, serta untuk mempelajari sistem perkawinan (mating
system) pada tanaman jati.
Material jaringan daun yang diambil untuk diekstrasi DNA nya adalah
daun yang masih muda dan masih akan berkembang, kira-kira berukuran 8 - 12
cm dan diambil sebanyak 5 - 7 helai. Daun-daun tersebut kemudian diletakan di
antara dua kertas yang dapat menyerap air kemudian dikeringkan pada ruangan
ber AC (air-conditioning room) atau dikering anginkan. Material daun dapat juga
disimpan dalam kantong ziplock yang berisi silika gel. Jaringan tersebut juga
dapat diambil langsung dilapang menggunakan tabung microtube 1.5-2.0 ml yang
mengandung buffer ekstraksi, namun dengan cara ini daun tersebut harus segera
di gerus untuk diekstrasi atau dapat disimpan dalam waktu cukup lama sebelum
diekstrak pada suhu -20oC.
Sedangkan benih diambil dengan cara memanen benih sebanyak 100-150
biji dari 15-20 pohon yang dianggap sebagai induk potensial, untuk
masing-masing populasi. Benih-benih tersebut dikecambahkan untuk kemudian diisolasi
DNA nya.
Prosedur Molekular dengan Marka Mikrosatelit Isolasi DNA
Isolasi DNA daun jati dilakukan menggunakan metode CTAB. Sebanyak
1 gram daun jati muda ditambahkan nitrogen cair kemudian digerus dengan
mortal, serbuk halusnya kemudian dimasukkan ke dalam tabung eppendorf .
Untuk sampel sebanyak 0.3 g yang telah dimasukkan ke dalam tabung eppendorf tersebut kemudian ditambahkan 700 µL buffer CTAB (100 mM Tris-HCL pH 8.0,
1.4 M NaCl, 20 mM EDTA, 2% (b/v) CTAB), yang sebelumnya dipanaskan pada
suhu 65oC dan 0.2% β-mercaptoethanol yang ditambahkan pada saat melakukan
ekstraksi) dan digoyang-goyang supaya tercampur sempurna selama 30 detik.
Sampel dalam buffer diinkubasi dalam penangas air pada suhu 65oC selama 30
menit dan sesekali dibolak-balik secara perlahan supaya buffer tercampur sempurna dengan sampel. Kemudian campuran tersebut dibiarkan pada suhu
ruang selama beberapa menit untuk menurunkan suhu.
Untuk memisahkan larutan DNA dengan kotoran lainnya ditambahkan
kloroform/isoamilalkohol (24:1) sebanyak 700 µL dan digoyang-goyang sampai
menit pada suhu ruang. Larutan bagian atas dipipet dan dimasukan ke dalam
tabung yang baru kemudian ditambahkan 750 µL isopropanol dingin dan
digoyang-goyang secara perlahan. Penambahan isopropanol dingin akan
menyebabkan terbentuknya benang-benang DNA yang halus berwarna putih.
Pengendapan DNA dilakukan dengan sentifugasi pada kecepatan 15000
rpm selama 15 menit pada suhu ruang. Larutan bagian atas dibuang dan pellet
dicuci dengan 200 µL ethanol 70% dengan cara mengoyang-goyang dan
disentrifugasi 10000 rpm selama 10 menit kemudian ethanol 70% dibuang dengan
cara dipipet kemudian pellet dikeringkan dengan cara membalikan tabung di atas
kertas tisue dan divacum selama 10 menit sampai kering. Endapan DNA
dilarutkan dengan 50 µL aquabidest dengan cara digoyang-goyang secara perlahan dan diinkubasi selama 30 menit atau satu malam pada suhu 37oC,
sebelum digunakan disimpan pada -20oC , untuk analisa selanjutnya.
Penetapan Kualitas dan Kuantitas serta Visualisasi DNA.
Ukuran dan integritas DNA ditentukan berdasarkan elektroforesis gel
agarose 1%. Pembuatan gel agarose 1% (b/v) dilakukan dengan cara melarutkan
1.0 g tepung agarose ke dalam 100 ml larutan buffer 1 x TAE (50 x TAE untuk 1
L mengandung 242 g Tris-base, 57.1 mL asam asetat glasial dan 100 mL 0.5 M
EDTA-Na2 pH 8.0) kemudian dipanaskan dalam microwave selama 2 menit
sampai agarose benar-benar larut. Larutan agarose diinkubasi selama 30 menit
untuk menurunkan suhunya hingga mencapai 65oC. Kemudian dituangkan ke
dalam cetakan gel yang sudah dipasang sisir dan dibiarkan sampai mengeras
kira-kira 1 jam kemudian sisir dicabut dari gel secara perlahan. Gel yang telah
mengeras dimasukan ke dalam bak elektroforesis dengan posisi sisir pada
elektroda negatif dan ke dalam bak dimasukkan larutan buffer 1 x TAE sampai
seluruh terendam.
Pengujian dilakukan dengan menggunakan 10 µL sampel DNA dilarutkan
dengan 5 µL aquabides dan 5 µL loading buffer FDEU (90% delonized
formamide, 0.1 M EDTA, 10% xylene cyanol, 10% bromophenol blue dan 8%
(w/v) urea) dan dicampur merata. Elektroforesis DNA dilakukan dengan
memasukan sebanyak 20 µL campuran DNA, aquabides dan loading buffer ke
model 1000/500 (BIORAD) pada tegangan konstan 75 volt, setelah pewarna
pewarna mencapai jarak 1 cm dari pinggir bawah gel, power suplai listrik
dimatikan (kira-kira selama 60 menit). Gel hasil elektroforesis direndam dalam
larutan buffer 1 x TAE yang diberi 0.5 µg/mL etidium bromida sambil
digoyang-goyangkan selama 20 menit kemudian dibilas dengan aquades selama 10 menit.
Hasil elektroforesis dilihat dengan menggunakan transiluminator UV model
T2202 (Sigma) untuk melihat pendaran DNA yang diberikan etidium bromida dan
hasilnya difoto dengan menggunakan film Polaroid 667.
Amplifikasi DNA.
DNA diamplifikasi dengan menggunakan primer spesifik yang telah
dikembangkan untuk jati dari proyect ICA4-2000-20053 (Tabel 3.2). Reaksi
amplifikasi dilakukan dengan menggunakan 25 µL yang merupakan campuran
larutan yang terdiri atas AmpliTaq DNA polimerase dan Stoffel fragment, dNTPs
(masing-masing dATP, dCTP, dGTP, dan dTTP 0.4 mM), 2.5 µg bovine serum
albumin (BSA) dan buffer (3 mM MgCl2, 30 mM KCl dan 10 mM Tris, pH 8.3),
25 pmol primer, 2 µL DNA cetakan, dan 18 µL dH2O supaya mencapai volume
akhir 25 µL dimasukkan ke dalam tabung eppendorf volume 500 µL dispin secara pelahan lahan supaya semua larutan tercampur sempurna. Minyak mineral
ditambahkan keatas campuran larutan PCR dan DNA cetakan sebanyak 20 µL
Tabel 3.2. Nama dan sekuen primer mikrosatelit berasal dari project jati TEAKDIV ICA4-2000-20053
No Primer Sekuen
1 AG04 for: 5’-AGAGGAGGTGCAGAGAGCAG-3’ rev: 5’-TAGCATTTGCTGCAAGCTGT-3’ 2 AG16 for: 5’-ATGCAAAAACGGAGTCTTGG-3’
rev: 5’-GGCAGAGCTATCTGAAGATCC-3’ 3 AGT10 for: 5’-TGCAGATAAAATGCTTGTGGA-3’
rev: 5’-CGCGAGAAATAGACCAGTGC-3’ 4 AC44 for: 5’-ACGCGGGTGTTAGGAAAATG-3’
rev: 5’-CCCATCAAACTGAGACAACCA-3’ 5 AC01 for: 5’-CATGTTGTATCATGAATGTG-3’
rev: 5’-CCTAGAAGAGAACCCCATGC-3’ 6 AG14 for: 5’-TCCACGACTCATGCAGGCTA-3’
rev: 5’-CCAACCAACCCTTTCAAATCC-3’ 7 ATC02 for: 5’-TCAAAGCTTGGCTACCACCA-3’
rev: 5’-GCCGAATTGGGACGACTTTA-3’ 8 AC28 for: 5’-ACGGCTATCAGACCAGCAGA-3’ rev: 5’-ATGCATGGCATGTTCTACCC-3’ 9 AAG10 for: 5’-GTGCACCAAGTCCGAGCAAT-3’
rev: 5’-CGAGAACCCGAACCTAACCA-3’ 10 CPIMS for: 5’-TTTCCCGTTATGTAGAGAATTGA-3’
rev: 5’-CCCAAATTGTGAACGATGAA-3’
Tabung berisi campuran larutan PCR dan DNA cetakan dimasukkan ke
dalam mesin PCR. Reaksi amplifikasi DNA dilakukan dengan menggunakan mesin PCR (Gene Amp PCR system 2400 Perkin-Elmer) dan diprogramkan untuk
PCR awal pada suhu 95oC selama 5 menit satu siklus. Denaturasi DNA cetakan dari utas ganda menjadi utas tunggal pada suhu 95oC selama 1 menit, penempelan
primer ke DNA cetakan pada suhu 36oC selama 1 menit dan pemanjangan pada
72oC selama 2 menit sebanyak 45 siklus, dan pemanjangan akhir pada suhu 72oC
selama 5 menit satu siklus, dan pendinginan pada suhu 4oC selama tidak terhingga
4. ANALISIS KEMIRIPAN DAN KERAGAMAN GENETIK
TANAMAN JATI SULAWESI TENGGARA MENGGUNAKAN
MARKA MIKROSATELIT
(Genetic similarity and diversity analysis of teak from originated
Southeast Sulawesi by using microsatellite markers)
Abstract
The objective of this research was to analyze genetic similarity and diversity of three populations of teak from Southeast Sulawesi using ten microsatellite loci. The result of the research showed that the average genetic similarity, which calculated based on Dice coefficient, for total population of mature and juvenile tree were 51.91% and 54.55%, respectively. Mature tree of Dolok, Warangga and Sampolawa had genetic similarity 60%, 55%, and 73%, respectively. Juvenile tree of Dolok, Warangga, and Sampolawa had genetic similarity 56%, 61%, and 74%, respectively. The juvenile tree harvested from 13-19 mature trees showed high genetic diversity similar with mature tree. Principal component analysis showed by the first two principal components explained 38% and 40% of total diversity of mature and juvenile trees. Population was separated into tree groups according locations. But condition of harvest of seeds of this research done before illegal logging on large scale, so that source of pollen very abundances. At present teak in Southeast Sulawesi are endangered, constraint of reboisation is due to not availability of seed. Bulk harvest from a lot of tree and a lot of location represent a strategy to prevent inbreeding and genetic drift.
Keywords: Tectona grandis, genetic similarity, mikrosatelit
Abstrak
Penelitian bertujuan untuk mempelajari kemiripan dan keragaman genetika tiga populasi tanaman jati asal Sulawesi Tenggara. Hasil penelitian menunjukkan rata-rata kemiripan genetika yang dihitung menggunakan koefisien Dice (Nei dan Li, 1979) pada populasi total tanaman dewasa dan semai asal Sulawesi Tenggara berturut turut 51.91% dan 54.55%. Untuk tanaman dewasa kelompok Dolok, Warangga dan Sampolawa mempunyai kemiripan genetika berturut-turut 60%, 55% dan 73%. Sedangkan tanaman semai kelompok Dolok, Warangga dan Sampolawa mempunyai kemiripan genetika berturut-turut 56%, 61%, dan 74%. Tanaman semai yang diunduh dari 13-19 pohon induk benih memperlihatkan keragaman yang tinggi serupa dengan tanaman induknya (dewasa). Namun kondisi pengunduhan benih pada penelitian ini terjadi sebelum penebangan hutan secara besar-besaran, sehingga sumber serbuk sari sangat berlimpah. Saat ini jati di Sulawesi Tenggara sudah demikian parah, kendala reboisasi hutan tidak tersedianya benih. Pengunduhan yang berasal dari banyak tanaman dan berasal dari banyak lokasi merupakan strategi yang dapat mengurangi terjadinya inbreeding dan penghanyutan genetik.
Pendahuluan
Kawasan hutan jati di Kabupaten Muna, Sulawesi Tenggara kini dalam
kondisi rusak parah, akibat penebangan liar yang tidak terkendali. Sekitar 29.000
hektar lebih dari luas total areal semula yang mencapai 30.000 hektar, sudah
berubah menjadi kawasan gundul. Hingga di awal tahun 2004 luas areal jati yang
tersisa tinggal sekitar 1.000 hektar lebih, itu pun luasannya tidak terkonsentrasi
pada satu kawasan, melainkan tersebar di beberapa titik lokasi.
Untuk mengembalikan kerusakan hutan jati yang sudah cukup serius itu,
pemerintah pusat melalui dana reboisasi hutan, berusaha menanam kembali
tanaman jati melalui proyek tersebut yang sepenuhnya diserahkan kepada petani.
Namun demikian yang menjadi kendala sekarang adalah tidak tersedianya sumber
benih tanaman lokal sedangkan benih yang berasal dari tempat lain belum tentu
sesuai dan harus diuji terlebih dahulu.
Pembangunan hutan dengan cara mengunduhan benih yang berasal hanya
dari beberapa tanaman saja dari tanaman yang tersisa akan menimbulkan masalah
seperti inbreeding dan penghanyutan genetik, padahal untuk jangka panjang keragaman genetik total yang tinggi perlu dipertahankan. Dengan demikian
diperlukan penelitian yang dapat memberi gambaran atau bayangan tentang
kosekuensi genetik sendainya benih hanya diunduh dari beberapa tanaman saja
atau dari beberapa area saja, sehingga dapat dilakukan langkah-langkah yang tepat
untuk menghindari hilangnya keragaman genetik.
Studi tentang kemiripan genetik di antara individu tanaman dan di antara
populasi dapat digunakan untuk mengambarkan keragaman genetik yang ada dan
dapat digunakan dalam memilih individu tanaman dari kelompok populasi.
Selama ini pemilihan individu tanaman berdasarkan fenotipe (tanaman plus),
namun karakter fenotipe sangat dipengaruhi oleh lingkungan, dengan demikian
informasi tambahan secara genetik sangat diperlukan guna mendapatkan tanaman
plus tersebut.
Dalam program pemuliaan dan konservasi keberhasilannya sangat
digunakan sebagai sumber material genetik yang memiliki keragaman genetik
total yang tinggi.
Keragaman dan kemiripan genetika dapat dipelajari melalui analisis
langsung terhadap sifat morfologi, biokimia atau melalui penanda DNA seperi
mikrosatelit. Penanda mikrosatelit merupakan salah satu penanda genetik yang
bersifat unggul untuk digunakan karena terdistribusi secara melimpah dan merata
dalam genom, variabilitasnya sangat tinggi (banyak alel dalam lokus), bersifatnya
kodominan, bersifat polimorfik yang tinggi dan berbasis PCR
Adapun tujuan penelitian ini adalah untuk (1) mempelajari kemiripan
genetik individu-individu tanaman di dalam populasi, (2) mempelajari keragaman
genetik jati di dalam populasi dan antar populasi, dan (3) membandingkan
keragaman genetik populasi jati dewasa serta progeni dari bulk benih jati
Bahan dan Metode Material Tanaman
Material tanaman jati berupa daun dan benih diperoleh dari dua kabupaten
di Sulawesi Tenggara yaitu Kabupaten Muna (Dolok dan Warangga) dan
Kabupaten Buton (Sampolawa), dikoleksi masing-masing 20 sampel seperti
tertera pada Tabel 4.1 sebagai berikut:
Tabel 4.1. Pohon induk dan tanaman semai jati yang dikoleksi dipilih secara acak pada tiga lokasi populasi jati asal Sulawesi Tenggara dianalisis menggunakan 10 lokus mikrosatelit
Asal Populasi Kode Jumlah Tanaman
Pohon T 20 Dolok, Muna
Semai T 20
Pohon W 20 Warangga, Muna
Semai W 20 Pohon S 20 Sampolawa, Buton
Semai S 20
Total 6 populasi 120 tanaman
Isolasi serta Penentuan Kualitas dan Kuantitas DNA
Untuk mendapatkan DNA total genom dilakukan isolasi DNA dari daun
dengan menambahkan nitrogen cair. Isolasi DNA dilakukan menggunakan
procedur CTAB yang telah dikembangkan dari project ICA4-2000-20053 (Bab 3
dan Lampiran 8).
Kualitas dan kuantitas DNA yang diperoleh dicek terlebih dahulu dengan
merunning dalam gel agarose 0.8% dengan menggunakan alat elektroforesis.
Spektrofotometer digunakan untuk menentukan kemurnian dan konsentrasi DNA.
Amplifikasi PCR dan Gel Electroforesis
DNA tanaman hasil isolasi kemudian diamplifikasi menggunakan mesin
PCR menggunakan 10 primer mikrosatelit. Program