KERAGAMAN GENETIK TIGA POPULASI KELAPA
SAWIT (Elaeis guinensis Jacq.) TIPE PISIFERA
BERDASARKAN MARKA RAPD
SKRIPSI
OLEH :
HERU PRAYOGI 111201104/ Budidaya Hutan
PROGRAM STUDI KEHUTANAN
FAKULTAS PERTANIAN
PERTUMBUHAN DAN KOMPOSISI RANTAI PANJANG
POLYISOPRENOID PADA MANGROVE
Avicennia marina (Forsk). DI BAWAH
CEKAMAN SALINITAS
SKRIPSI
Oleh :
HAMSYAH R HARAHAP 111201134
BUDIDAYA HUTAN
Skripsi sebagai salah satu syarat untuk memperoleh gelar sarjana di Fakultas Kehutanan
Universitas Sumatera Utara
FAKULTAS KEHUTANAN
UNIVERSITAS SUMATERA UTARA
HALAMAN PENGESAHAN
Judul Penelitian: Keragaman Genetik Tiga Populasi Kelapa Sawit
(Elaeis guinensis Jacq.) Tipe Pisifera Berdasarkan Marka RAPD
Nama : Heru Prayogi
NIM : 111201104
Program studi : Kehutanan
Disetujui Oleh
Komisi Pembimbing
Mohammad Basyuni, S.Hut.,M.Si.,Ph.D
Ketua Anggota
Dr. Ir. Lollie A.P. Putri, M.Si.
Mengetahui
i
ABSTRAK
HERU PRAYOGI. Keragaman Genetik Tiga Populasi Kelapa Sawit (Elaeis guinensis Jacq.) Tipe Pisifera Berdasarkan Marka RAPD Dibimbing oleh MOHAMMAD BASYUNI dan LOLLIE AGUSTINA P. PUTRI
Kelapa sawit sebagai tanaman penghasil minyak kelapa sawit merupakan salah satu primadona tanaman perkebunan yang menjadi sumber penghasil devisa non-migas bagi Indonesia. Varietas unggul kelapa sawit yang dihasilkan oleh berbagai lembaga riset adalah tenera yang merupakan hibrida dura x pisifera (DxP). Pisifera memiliki fungsi yang penting dalam produksi benih kelapa sawit. Oleh karena itu populasi pisifera sangat penting untuk dikelola dan dikembangkan. Metode RAPD merupakan metode untuk mengidentifikasi sejumlah besar polimorfisme DNA pada genom dengan cepat, efisien dan cocok untuk studi keanekaragaman genetik. Tujuan dari penelitian ini adalah untuk menganalisis keragaman genetik tipe sawit dari tiga populasi yakni Yangambi origin, Lame origin dan Lame silang lanjut di koleksi kebun Bangun Bandar PT. Socfin Indonesia menggunakan marka RAPD. Dari 18 sampel untuk setiap populasi Yangambi origin, Lame origin dan Lame silang lanjut sawit tipe pisifera dapat diketahui tingkat keragaman genetik yang sedang dengan nilai rata-rata 0,24 dengan nilai tertinggi 0,28 dan nilai terendah 0,21. Persentase pita polimorfik tertinggi terdapat pada primer 15 dan primer 19 Lame origin yang mencapai nilai 100%. Nilai PIC tertinggi terdapat pada primer 11 Yangambi origin dan primer 10 Lame sl sebesar 0,49, sedangkan untuk nilai PIC terendah terdapat pada primer 21 Lame origin sebesar 0,01.
ii
ABSTRACT
HERU PRAYOGI. The genetic diversity, palm oil (Elaeis guinensis Jacq.) of
pisifera using RAPD markers under academic supervision by MOHAMMAD
BASYUNI and LOLLIE AGUSTINA P. PUTRI.
Palm oil as a plant producing oil is one of the primadonna oil palm plantations that become a source of producer non-migas for Indonesia foreign exchange. Superior varieties of palm oil produced by various research institutions that is a hybrid is tenera dura x pisifera (DxP). Pisifera having an important function in the production of seed oil palm. Hence the population pisifera very important to be managed and developed . A method of RAPD is a method to identify a number of large polymorphism DNA in the genome quickly, efficient and suitable for the study of genetic diversity. The purpose of this research is to analyze genetic diversity type palm oil by three population namely Yangambi origin, Lame origin and Lame silang lanjut in collections garden bangun bandar PT Socfin Indonesia use RAPD markers. Sample of 18 for each oil palm population Yangambi origin, Lame origin and Lame silang lanjut pisifera type can be seen the level of genetic diversity is the average value 0.24 with the highest 0,28 and the lowest value 0.21. The percentage of ribbon polymorphic highest on the primary 15 and primary 19 Lame origin which reached 100 percent. The PIC highest on the primary 11 Yangambi origin and primary 10 lame sl of 0.49, while to the PIC the lowest on primary 21 Lame origin of 0.01.
iii
RIWAYAT HIDUP
Penulis lahir di desa Serapuh kabupaten Simalungun pada 17 November
1992 dari Misman dan Tukiyawati. Penulis merupakan anak keempat dari empat
bersaudara.
Penulis Lulusan SD Negeri 097320 Serapuh pada Tahun 2005, lulusan
SMP Negeri 2 Siantar pada tahun 2008, dan lulusan SMA Negeri 3 Pematang
Siantar pada tahun 2011. Juni 2011 penulis diterima di Program Studi Kehutanan,
Fakultas Pertanian Universitas Sumatera Utara melalui jalur SNMPTN tulis.
Selanjutnya penulis memilih peminatan Budidaya Hutan.
Penulis mengikuti Praktik Pengenalan Ekosistem Hutan (P2EH) 22-31
Agustus 2013 di hutan pendidikan USU Tahura, Tongkoh, Kabupaten Karo.
Penulis melaksanakan Praktik Kerja Lapang di Taman Nasional Gunung Halimun
iv
KATA PENGANTAR
Puji dan syukur penulis panjatkan kehadirat Allah SWT atas segala rahmat
dan karunia-Nya kepada penulis, sehingga penulis dapat menyelesaikan
penyusunan skripsi yang berjudul “Keragaman Genetik Kelapa Sawit Elaeis
guinensis (Jacq.) Tipe Pisifera dari Tiga Populasi Berdasarkan Marka RAPD.”.
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar sarjana di
Program Studi Kehutanan, Fakultas Pertanian, Universitas Sumatera Utara.
Pada kesempatan ini penulis mengucapkan terima kasih kepada kedua
orang tua penulis yang telah membesarkan dan mendidik penulis selama ini.
Penulis juga menyampaikan ucapan terima kasih kepada Mohammad Basyuni,
S.Hut, M.Si, Ph.D sebagai ketua komisi pembimbing skripsi dan Dr. Ir. Lollie
A.P. Putri, M.Si. sebagai anggota komisi pembimbing skripsi, yang telah banyak
membimbing dan memberikan arahan serta masukan kepada penulis sehingga
penulis dapat menyelesaikan penelitian ini.
Disamping itu penulis juga mengucapkan terima kasih kepada kepada
semua staf pengajar dan pegawai di Program Studi Kehutanan, serta semua
teman-teman mahasiswa yang telah membantu penulis dalam menyelesaikan
skripsi ini. Semoga skripsi ini dapat bermanfaat bagi pengembangan ilmu
pengetahuan.
Medan, Juni 2015
v
DAFTAR ISI
Halaman
ABSTRAK ... i
ABSTRACT ... ii
RIWAYAT HIDUP ... iii
KATA PENGANTAR ... iv
DAFTAR TABEL ... vii
DAFTAR GAMBAR ... viii
PENDAHULUAN Latar Belakang ... 1
Tujuan Penelitian ... 3
Manfaat Penelitian ... 3
TINJAUAN PUSTAKA Kelapa sawit secara umum ... 4
Keragaman genetik... 5
Isolasi DNA ... 5
Reaksi polymerase chain reaction (PCR) ... 6
Marka RAPD ... 7
BAHAN DAN METODE Waktu dan Tempat ... 9
Alat dan Bahan ... 9
Prosedur Penelitian... 10
Pengambilan sample ... 10
Isolasi DNA ... 10
Pengukuran konsentrasi DNA ... 11
Gel agarose 1 % ... 11
Running gel elektroforesis ... 12
Reaksi PCR primer dengan marka RAPD ... 12
Penanda RAPD ... 12
Penentuan Skoring Marka RAPD ... 13
Penentuan Ukuran Pasangan Basa ... 13
HASIL DAN PEMBAHASAN Analisa Profil Pita Hasil Isolasi DNA Tanaman Kelapa Sawit Pisifera ... 14
Analisa Profil Pita Hasil Isolasi DNA Tanaman Kelapa Sawit Pisifera ... 19
KESIMPULAN DAN SARAN Kesimpulan ... 35
vi DAFTAR PUSTAKA
vii
DAFTAR TABEL
No. Halaman
1. Nama Primer RAPD... 10
2. Program Mesin PCR ... 12
3. Konsentrasi DNA Yangambi Origin ... 18
4. Konsentrasi DNA Lame Origin ... 18
5. Konsentrasi DNA Lame Silang Lanjut ... 19
6. Keragaman Genetik Sawit Tipe Pisifera ... 20
viii
DAFTAR GAMBAR
No. Halaman
1. Uji Kualitas 18 DNA Tanaman Kelapa Sawit Yangambi Origin
Tipe Pisifera.. ... 14
2. Uji Kualitas 18 DNA Tanaman Kelapa Sawit Lame Origin Tipe
Pisifera.. ... 15
3. Uji Kualitas 18 DNA Tanaman Kelapa Sawit Lame Silang Lanjut
Tipe Pisifera.. ... 15
4. Amplifikasi 18 DNA Kelapa Sawit Yangambi Origin Tipe Pisifera
Menggunakan Primer 06.. ... 21
5. Amplifikasi 18 DNA Kelapa Sawit Lame Origin Tipe Pisifera
Menggunakan Primer 06.. ... 21
6. Amplifikasi 18 DNA Kelapa Sawit Lame Silang Lanjut Tipe
Pisifera Menggunakan Primer 06.. ... 22
7. Amplifikasi 18 DNA Kelapa Sawit Yangambi Origin Tipe Pisifera
Menggunakan Primer 10.. ... 23
8. Amplifikasi 18 DNA Kelapa Sawit Lame Origin Tipe Pisifera
Menggunakan Primer 10.. ... 23
9. Amplifikasi 18 DNA Kelapa Sawit Lame Silang Lanjut Tipe
Pisifera Menggunakan Primer 10.. ... 24
10.Amplifikasi 18 DNA Kelapa Sawit Yangambi Origin Tipe Pisifera
Menggunakan Primer 11.. ... 25
11.Amplifikasi 18 DNA Kelapa Sawit Lame Origin Tipe Pisifera
Menggunakan Primer 11.. ... 25
12.Amplifikasi 18 DNA Kelapa Sawit Lame Silang Lanjut Tipe
Pisifera Menggunakan Primer 11.. ... 26
13.Amplifikasi 18 DNA Kelapa Sawit Yangambi Origin Tipe Pisifera
Menggunakan Primer 15.. ... 27
14.Amplifikasi 18 DNA Kelapa Sawit Lame Origin Tipe Pisifera
ix
15.Amplifikasi 18 DNA Kelapa Sawit Lame Silang Lanjut Tipe
Pisifera Menggunakan Primer 15.. ... 28
16.Amplifikasi 18 DNA Kelapa Sawit Yangambi Origin Tipe Pisifera
Menggunakan Primer 19.. ... 29
17.Amplifikasi 18 DNA Kelapa Sawit Lame Origin Tipe Pisifera
Menggunakan Primer 19.. ... 29
18.Amplifikasi 18 DNA Kelapa Sawit Lame Silang Lanjut Tipe
Pisifera Menggunakan Primer 19.. ... 30
19.Amplifikasi 18 DNA Kelapa Sawit Yangambi Origin Tipe Pisifera
Menggunakan Primer 21.. ... 31
20.Amplifikasi 18 DNA Kelapa Sawit Lame Origin Tipe Pisifera
Menggunakan Primer 21.. ... 31
21.Amplifikasi 18 DNA Kelapa Sawit Lame Silang Lanjut Tipe
i
ABSTRAK
HERU PRAYOGI. Keragaman Genetik Tiga Populasi Kelapa Sawit (Elaeis guinensis Jacq.) Tipe Pisifera Berdasarkan Marka RAPD Dibimbing oleh MOHAMMAD BASYUNI dan LOLLIE AGUSTINA P. PUTRI
Kelapa sawit sebagai tanaman penghasil minyak kelapa sawit merupakan salah satu primadona tanaman perkebunan yang menjadi sumber penghasil devisa non-migas bagi Indonesia. Varietas unggul kelapa sawit yang dihasilkan oleh berbagai lembaga riset adalah tenera yang merupakan hibrida dura x pisifera (DxP). Pisifera memiliki fungsi yang penting dalam produksi benih kelapa sawit. Oleh karena itu populasi pisifera sangat penting untuk dikelola dan dikembangkan. Metode RAPD merupakan metode untuk mengidentifikasi sejumlah besar polimorfisme DNA pada genom dengan cepat, efisien dan cocok untuk studi keanekaragaman genetik. Tujuan dari penelitian ini adalah untuk menganalisis keragaman genetik tipe sawit dari tiga populasi yakni Yangambi origin, Lame origin dan Lame silang lanjut di koleksi kebun Bangun Bandar PT. Socfin Indonesia menggunakan marka RAPD. Dari 18 sampel untuk setiap populasi Yangambi origin, Lame origin dan Lame silang lanjut sawit tipe pisifera dapat diketahui tingkat keragaman genetik yang sedang dengan nilai rata-rata 0,24 dengan nilai tertinggi 0,28 dan nilai terendah 0,21. Persentase pita polimorfik tertinggi terdapat pada primer 15 dan primer 19 Lame origin yang mencapai nilai 100%. Nilai PIC tertinggi terdapat pada primer 11 Yangambi origin dan primer 10 Lame sl sebesar 0,49, sedangkan untuk nilai PIC terendah terdapat pada primer 21 Lame origin sebesar 0,01.
ii
ABSTRACT
HERU PRAYOGI. The genetic diversity, palm oil (Elaeis guinensis Jacq.) of
pisifera using RAPD markers under academic supervision by MOHAMMAD
BASYUNI and LOLLIE AGUSTINA P. PUTRI.
Palm oil as a plant producing oil is one of the primadonna oil palm plantations that become a source of producer non-migas for Indonesia foreign exchange. Superior varieties of palm oil produced by various research institutions that is a hybrid is tenera dura x pisifera (DxP). Pisifera having an important function in the production of seed oil palm. Hence the population pisifera very important to be managed and developed . A method of RAPD is a method to identify a number of large polymorphism DNA in the genome quickly, efficient and suitable for the study of genetic diversity. The purpose of this research is to analyze genetic diversity type palm oil by three population namely Yangambi origin, Lame origin and Lame silang lanjut in collections garden bangun bandar PT Socfin Indonesia use RAPD markers. Sample of 18 for each oil palm population Yangambi origin, Lame origin and Lame silang lanjut pisifera type can be seen the level of genetic diversity is the average value 0.24 with the highest 0,28 and the lowest value 0.21. The percentage of ribbon polymorphic highest on the primary 15 and primary 19 Lame origin which reached 100 percent. The PIC highest on the primary 11 Yangambi origin and primary 10 lame sl of 0.49, while to the PIC the lowest on primary 21 Lame origin of 0.01.
1
PENDAHULUAN
Latar belakang
Tanaman kelapa sawit disebut juga dengan Elaeis guinensis Jacq. Elaeis
berasal dari kata Elaion yang dalam bahasa Yunani berarti minyak. Guinensis
berasal dari kata Guinea yaitu Pantai Barat Afrika dan Jacq singkatan dari Jacquin
seorang Botanist dari Amerika. Varietas dari Elaeis guinensis Jacq. cukup banyak
dan biasanya diklasifikasikan atas tipe buah, bentuk luar, tebal cangkang, warna
buah dan lain-lain. Perluasan lahan kelapa sawit (E. guinensis Jacq.) di Indonesia
selalu meningkat setiap tahunnya, bahkan perusahaan perkebunan negara yaitu
PT. Perkebunan Nusantara berencana untuk mengembangkan sekitar 1,8 juta
hektar perkebunan kelapa sawit di kawasan perbatasan Indonesia dan Malaysia.
Dengan demikian diperlukan bibit dalam jumlah yang sangat banyak.
Perbanyakan secara generatif akan menghasilkan tanaman yang beragam karena
kelapa sawit merupakan tanaman yang menyerbuk silang. Dengan demikian harus
dilakukan perbanyakan secara vegetatif (Badan litbang pertanian, 2013).
Tingkat produksi yang dicapai pertama-tama ditentukan oleh potensi
genetik varietas, faktor lingkungan dan pengelolaan. Varietas unggul kelapa sawit
yang dihasilkan oleh berbagai lembaga riset adalah tenera yang merupakan
hibrida dura x pisifera (DxP), yaitu bunga jantan (pollen) dari jenis pisifera
dikawinkan pada bunga betina dari jenis dura. Jadi, benih yang membawah sifat
gabungan kedua jenis sawit tersebut adalah biji dari dura. Benih biji hibrida
dengan yang bukan hibrida tidak bisa dibedakan baik dalam hal ukuran, bentuk
dan warnanya sehingga membuka peluang pemalsuan. Begitu pula dengan
2
pemalsu benih juga menjual biji dari tenera yang dipanen dari kebun produksi.
Kekeliruan memilih benih baru disadari ketika tanaman sudah memasuki fase
berproduksi. Jika biji palsu berasal dari dura, maka seluruh tanaman adalah dura,
tetapi kalau berasal dari biji tenera, maka sebagian akan kembali ke induk dura,
sebagian kembali ke induk pisifera, dan sisanya seperti tenera sehingga kerugian
yang ditimbulkan sangat besar. Untuk meminimalisir kesalahan, usahakan
membeli benih kelapa sawit hanya dari produsen resmi yang bersertifikat melalui
prosedur resmi yang ditetapkan oleh produsen bersangkutan (Syakir, 2010).
Pisifera memiliki fungsi yang penting dan nilai ekonomis yang tinggi
dalam produksi benih kelapa sawit. Dalam industri perbenihan kelapa sawit,
konservasi yang baik pada serbuk sari pisifera akan dapat menunjang program
penyerbukan buatan yang berkelanjutan dalam produksi benih dan pemuliaan
yang terus berkembang. Oleh karena itu populasi pisifera merupakan salah satu
faktor penting untuk dikelola dan dikembangkan (Raganata, 2006).
Dewasa ini penelitian dengan menggunakan metode marka molekuler
berkembang sangat pesat, salah satu metode itu adalah dengan menggunakan
penanda Random amplified polymorpism DNA (RAPD). Metode RAPD
merupakan metode untuk mengidentifikasi sejumlah besar polimorfisme DNA
pada genom dengan cepat dan efisien. Tipe polimorfisme ini membuat RAPD
cocok untuk studi keanekaragaman genetik, hubungan kekerabatan, peta genetik,
sidik jari DNA. Sidik jari DNA banyak digunakan untuk kasus paternity dan
forensik. Metode RAPD menggunakan oligonukleotida pendek (biasanya 10 bp)
sebagai primer yang akan berikatan dengan bagian (sites) komplemennya
3
Oleh karena itu diperlukan studi keragaman genetik tipe pisifera
berdasarkan marka RAPD. Metode RAPD digunakan untuk mendeteksi
polimorfisme DNA yang digunakan sebagai genetik marker dan menentukan
hubungan kekerabatan pada bermacam-macam tanaman. Metode RAPD
merupakan penanda molekuler berbasis PCR. PCR adalah suatu teknik sintesis
dan amplifikasi DNA secara in vitro.
Tujuan
Tujuan dari penelitian ini adalah untuk menganalisis keragaman genetik
tipe sawit dari tiga populasi yakni Yangambi origin, Lame origin dan Lame silang
lanjut di koleksi kebun Bangun Bandar PT. Socfin Indonesia.
Manfaat
Manfaat dari penelitian ini diharapkan dapat memberikan informasi
mengenai keragaman genetik kelapa sawit tipe pisifera dari tiga populasi yang
4
TINJAUAN PUSTAKA
Kelapa sawit secara umum
Divisi : Embryophyta siphonagama
Kelas : Angiospermae
Ordo : Monocotyledonae
Famili : Arecaceae (Dahulu Palmae)
Sub-famili : Cocoideae
Genus : Elaeis
Spesies : E. guineensis Jacq.
Kelapa sawit sebagai tanaman penghasil minyak kelapa sawit (CPO- crude
palm oil) dan inti kelapa sawit (CPO) merupakan salah satu primadona tanaman
perkebunan yang menjadi sumber penghasil devisa non-migas bagi Indonesia.
Cerahnya prospek komoditi minyak kelapa sawit dalam perdagangan minyak
nabati dunia telah mendorong pemerintah Indonesia untuk memacu
pengembangan areal perkebunan kelapa sawit (Manurung, 2001).
Varietas unggul kelapa sawit yang dihasilkan oleh berbagai lembaga riset
adalah tenera. Tenera merupakan hibrida dura x pisifera (DxP), yaitu bunga jantan
(pollen) dari jenis pisifera dikawinkan pada bunga betina dari jenis dura (Syakir,
2010). Persilangan antara varietas dura dengan pisifera menghasilkan hibrida
tenera, yang memperlihatkan peningkatan hasil sebesar 30% dibandingkan
varietas dura (Fathurrahman, 2013).
Kelapa sawit tipe pisifera memiliki kelamin betina steril sehingga di dalam
program pemuliaan kelapa sawit tipe pisifera ini umumnya hanya digunakan
5
(Toruan, 1997). Pengamatan genotipik pada tingkat DNA tidak dipengaruhi oleh
umur tanaman atau faktor lingkungan sehingga sama pada setiap fase atau tahap
pertumbuhan dan perkembangan tanaman. Analisis pada tingkat DNA dapat
digunakan untuk deteksi sedini mungkin pada fase pembibitan atau bahkan saat
perbanyakan dalam kultur jaringan, khususnya tanaman perkebunan seperti
tanaman kelapa sawit. Dengan demikian program pemuliaan tanaman dalam
melakukan seleksi akan dipercepat, sehingga dapat memberi rekomendasi lebih
awal (Yuniastuti, 2005).
Keragaman Genetik
Guna mendukung upaya pemberdayaan potensi plasma nutfah pada
program seleksi maka mutlak diperlukan kelengkapan informasi yang berkaitan
dengan berbagai karakter morfologi maupun genetiknya. Marka molekuler dapat
memberi gambaran yang akurat tentang perbedaan genetik individu, baik pada
tingkat spesies maupun dengan kerabat jauhnya (Zulhermana, 2009).
Keragaman genetik merupakan faktor yang sangat berpengaruh dalam
menyusun strategi pemuliaan pohon. Karakter genetik suatu jenis pohon baik
yang terdapat dalam satu tempat tumbuh maupun yang berbeda provenansinya,
hal ini disebabkan karena perbedaan genetik. Kondisi ini menunjukkan sifat dan
kekhasan suatu tegakan. Sehingga tegakan atau provenansi yang memiliki
karakter genetik yang baik dapat menjadi sumber yang tepat untuk kegiatan
pemuliaan pohon (Langga dkk, 2012).
Isolasi DNA
Isolasi DNA merupakan langkah awal yang harus dikerjakan dalam
6
total DNA dari jaringan adalah dengan memecah dan mengekstraksi jaringan
tersebut sehingga akan terbentuk ekstrak sel yang terdiri atas sel-sel jaringan,
DNA, dan RNA. Kemudian ekstrak sel dipurifikasi sehingga dihasilkan pelet sel
yang mengandung DNA total. Prinsip-prinsip isolasi DNA plasmid hampir sama
dengan isolasi total DNA dari jaringan (Faatih, 2009).
Ada tiga langkah utama dalam ekstraksi DNA, yaitu perusakan dinding sel
(lisis), pemisahan DNA dari bahan padat seperti selulosa dan protein, serta
pemurnian DNA. Melalui proses tersebut DNA dipisahkan dari komponen seluler
lain seperti protein, RNA (Riboksi nukleat acid) dan lemak. Banyak metode yang
digunakan untuk mengisolasi DNA, tergantung spesimen yang akan dideteksi.
Metode tersebut pada dasarnya memiliki prinsip yang sama, namun ada beberapa
hal tertentu yang biasanya digunakan modifikasi untuk dapat menghancurkan
inhibitor yang ada di dalam masing-masing sumber spesimen (Langga dkk, 2012).
Analisis molekuler tumbuhan tergantung pada jumlah dan kemurnian sampel
DNA serta kondisi optimum reaksi. Oleh karena itu diperlukan metode yang tepat
dalam ekstraksi DNA dan reaksi molekuler seperti PCR (Pharmawati, 2009).
Reaksi Polymerase Chain Reaction (PCR)
PCR adalah suatu metode enzimatis untuk amplifikasi DNA dengan cara
in vitro. Pada proses PCR diperlukan beberapa komponen utama, yaitu DNA
cetakan, Oligonukleotida primer, Deoksiribonukelotida trifosfat (dNTP), Enzim
DNA Polimerase dan komponen pendukung lain adalah senyawa buffer. PCR
menggunakan alat termosiklus, sebuah mesin yang memiliki kemampuan untuk
memanaskan sekaligus mendinginkan tabung reaksi dan mengatur temperatur
7
terulang dalam 30-40 siklus dan berlangsung dengan cepat yaitu denaturasi,
annealing, dan pemanjangan untai DNA. Produk PCR dapat diidentifikasi melalui
ukurannya dengan menggunakan elektroforesis gel agarosa (Yusuf, 2010).
Teknik PCR pertama kali dikembangkan oleh Karry Mullis pada tahun
1985. Teknik PCR dapat digunakan untuk mengamplifikasi segmen DNA dalam
jumlah jutaan kali hanya dalam beberapa jam. Dengan ditemukannya teknik PCR
di samping juga teknik-teknik lain seperti sekuensing DNA, telah merevolusi
bidang sains dan teknologi khususnya di bidang diagnosa penyakit genetik,
kedokteran forensik dan evolusi molekular (Handoyo dan Ari, 2000). Amplikasi
PCR dari DNA menggunakan genomik primer acak penanda DNA polimer dan
keterkaitan genetik antara kultivar dan varietas (Sathish and Mohankumar, 2006).
Teknologi PCR terus disederhanakan dan dikembangkan, sehingga biaya
relatif rendah, kecepatan tinggi, membutuhkan contoh uji sangat sedikit, metode
ekstraksi dan amplifikasi yang sederhana sehingga membuat penanda berdasarkan
PCR dapat diaplikasikan pada semua spesies (Zulfahmi, 2013). Williams, et al.
(1990), telah mengembangkan metode RAPD (Random Amplified Polymorphic
DNA) yang hanya menggunakan random primer dalam PCR untuk menghasilkan
marker polimorfik yang tepat dan dapat digunakan untuk menentukan variasi dan
kekerabatan genetik pada tumbuhan (Pratiwi, 2012).
Marka RAPD
Analisis keragaman genetik tanaman dapat diketahui dengan identifikasi
secara molekuler dan identifikasi morfologi. Salah satu teknik molekuler yang
dapat digunakan adalah metode RAPD (Random Amplified Polymorphic DNA)
8
Teknik RAPD mendeteksi polimorfisme ruas nukleotida pada DNA
dengan menggunakan sebuah primer tunggal yang memiliki rangkaian nukleotida
acak. Pada reaksi PCR-RAPD ini, sebuah primer menempel pada DNA genomik
pada dua tempat berbeda dari DNA komplementer. Jika tempat penempelan
primer ini berada pada daerah yang dapat diamplifikasi, maka hasil DNA tertentu
dapat dihasilkan melalui amplifikasi siklus termal (Pharmawati, 2009).
Teknik RAPD dipilih untuk analisis genetik dengan berbagai alasan,
antara lain tidak membutuhkan latar belakang tentang genom yang akan
dianalisis, menggunakan bahan-bahan yang relatif murah dan amplifikasinya tidak
bergantung pada bahan radioaktif. Meskipun demikian hasilnya kadang kurang
konsisten. Untuk mengatasi hal tersebut, beberapa parameter yang mempengaruhi
kekhususan PCR perlu dioptimasi dengan tepat, misalnya pada suhu penempelan
(annealing), jumlah DNA polimerase dan konsentrasi primer. Selain itu juga
diperlukan standar pengerjaan yang sangat baik (Setiyo, 2001).
Penelitian menggunakan marka RAPD untuk kelapa sawit ini bukan yang
pertama kali dilakukan. Beberapa penelitian yang menggunakan marka RAPD
untuk kelapa sawit antara lain dilakukan oleh Zakaria (2005) Hetharie (2010)
9
BAHAN DAN METODE
Waktu dan Tempat
Peneltian ini dilakukan pada bulan Agustus 2014 sampai dengan bulan
Desember 2014. Penelitian ini dilaksanakan di laboratorium DNA, Pusat Seleksi
Bangun Bandar PT. SOCFINDO, Desa Martebing, Kecamatan Dolok Masihul,
Kabupaten Serdang Bedagai, Sumatera Utara.
Alat
Alat yang digunakan dalam penelitin ini adalah mortar, alumunium foil,
pastle, mikropipet, rak tube, waterbath, stirer, sentrifuge, kulkas, freezeer,
komputer, nanospec, timbangan elektrik, erlenmeyer, microwave, cetakan
agarose, bak elektroforesis, PCR, UV Transilluminator, Geldoc dan spin.
Bahan
Bahan yang digunakan dalam penelitian ini adalah daun kelapa sawit
muda dari tiga origin pisifera yaitu: Yangambi origin, Lame origin, Lame silang
lanjut masing-masing 18 daun untuk tiap origin, air, vinyl-polypyrolidone,
nitrogen cair, buffer ekstraksi castillo, β-merkaptoethanol,tube 2 ml, tube 1,5 ml,
tube 0,2 ml, chloroform : isoamil alkohol, isopropanol, buffer TE, CH3COONa 3
M pH 5.2, ethanol absolute, ethanol 70%, RNA-se, larutan TBE 0,5x, gel agarose,
gelred, es, nuclease free water, loading buffer dan primer RAPD.
Primer RAPD yang digunakan dalam penelitian ini adalah sebanyak lima
primer, sequence dan suhu annealinng dari masing-masing primer dapat dilihat
10
Tabel 1. Nama primer RAPD
Primer Sequence annealing (oC)
P6 TACCACCCCG 35
P10 GGACCCAACC 35
P11 GTCGCCGTCA 35
P15 TTGGCACGGG 35
P19 AGCGCCATTG 35
P21 GGGGTGACGA 35
Prosedur penelitian
A. Pengambilan sampel
Semua alat disiapkan seperti pisau dan plastik sampel, lalu diambil sampel
daun muda di lapangan dari masing-masing populasi. Daun dibersihkan dengan
air bersih kemudian dibuang tulang daunnya dan daun dipotong kecil-kecil.
Dimasukkan potongan daun ke alumunium foil lalu simpan ke dalam kulkas.
B. Isolasi DNA
Daun kelapa sawit muda yang sudah dicuci dan bersih ditambahkan ke
dalam mortar dingin sebanyak 0,1 gr. Kemudian ditambahkan ± 0,1 gr
Vinyl-Polypyrolidone dan nitrogen cair, gerus hingga halus. Serbuk halus yang
dihasilkan kemudian dimasukkan ke dalam tube 1,5 ml yang berisi 1 ml buffer
Castillo yang telah dipanaskan dan diberi 2 µl β-merkaptoethanol 1%. Campuran
tersebut kemudian dikocok menggunakan stirer, lalu dipanaskan selama 30 menit
pada suhu 65oC di waterbath. Larutan dibiarkan dingin pada suhu kamar lalu,
ditambahkan chloroform : isoamil alkohol 1 Volume. Larutan tersebut
disentrifuge selama 10 menit dengan kecepatan 11.000 rpm.
Supernatan diambil dan dipindahkan ke dalam tube 2 ml yang baru.
Kemudian ditambahkan 1 volume Chloroform : Isoamil alkohol, Tube tersebut
11
kecepatan 11.000 rpm, cairan bagian atas didalam tube kemudian dipipet dan
dimasukkan kedalam tube baru kemudian ditambahkan isopropanol dingin
sebanyak 1 volume. Dikocok perlahan hingga homogen kemudian simpan dalam
kulkas 1 - 4 jam, kemudian disentrifuge selama 10 menit dengan kecepatan
11.000 rpm. Cairan dan pellet DNA dibuang dan dikeringkan dengan cara
membalikkan tabung.
Pellet DNA dilarutkan dalam 0,1 ml buffer TE, kemudian ditambahkan
CH3COONa 3 M pH 5.2 sebanyak 1/10 volume dan ethanol absolute sebanyak 2,5
volume. Larutan tersebut dikocok hingga homogen dan disimpan dalam freezeer
selama 30 menit atau semalam. Kemudian larutan tersebut disentrifuge selama 10
menit dengan kecepatan 12.000 rpm, Cairan tersebut kemudian dibuang dan pellet
DNA diambil kemudian dicuci dengan ethanol 70% lalu dikeringkan. Pellet DNA
yang sudah kering dilarutkan dalam 50 µl buffer TE atau nuclease free water,
Ditambahkan RNA-se 25 µl, kemudian simpan dalam freezer. Kualitas DNA
kemudian diuji dengan elektroforesis gel agarose 2% dan konsentrasinya diukur
dengan nanospec pada panjang gelombang 230, 260 dan 280 nm.
C. Pengukuran konsentrasi DNA
Sebanyak 1 µl larutan diukur menggunakan nanospec, kemudian sebagai
blanko digunakan nuclease free water. Absorban diukur pada panjang gelombang
230, 260 dan 280 nm. Kemurnian DNA ditentukan dari nilai perbandingan
A260/A280.
D. Gel agarose 1%
Agarose ditimbang sebanyak 0,2 gr lalu dimasukkan ke dalam erlenmeyer
12
microwave. Angkat dan biarkan sampai suhu turun, siapkan cetakan gel
elektroforesis. Tambahkan gelred 1% atau 2 µ, kocok hingga homogen, lalu tuang
larutan agarose ke cetakan, tunggu hingga padat ± 30 menit.
E. Running gel elektroforesis
Bak elektroforesis disiapkan dan diisi dengan TBE 0,5x sebanyak 300 ml.
Kemudian cetakan gel yang sudah kering dimasukkan ke dalam bak, cetakan gel
dipastikan terendam dalam buffer. kemudian loading buffer disiapkan dalam
parafilm dan dicampurkan dengan sampel. Dimasukkan sampel ke dalam
sumur-sumur elektroforesis lalu dijalankan alat elektroforesis dengan
menghubungkannya dengan power supply.
F. Reaksi PCR primer dengan RAPD.
Semua reagen PCR disiapkan di atas es dan dibuat campuran primer
RAPD yang dimasukkan dalam tube 0,2 ml. Kemudian dijalankan reaksi pada
mesin PCR dengan program :
Tabel 2. Program mesin PCR (Sathis dan Mohankumar 2006).
Step Suhu (oC) Waktu Jumlah siklus
Pre Denaturasi 95 4 menit 1
Denaturasi 95 45 detik
Annealing 35 45 detik 40
Ekstensi 72 1 menit
Ekstensi Akhir 72 7 menit 1
Cooling 4 Unlimited
G. Penanda RAPD
Gel agarose disiapkan dengan konsentrasi 2%, kemudian dicetak dan
dimasukkan ke dalam bak elektroforesis, loading buffer kemudian disiapkan
dalam parafilm. Loading buffer dan sampel kemudian dimasukkan ke dalam
sumur-sumur elektroforesis. Alat elektroforesis dijalankan dengan
13
kemudian dikeluarkan dari bak elektroforesis kemudian hasil running
elektroforesis difoto menggunakan Geldoc.
Penentuan Skoring Marka RAPD
Untuk menentukan keragaman genetik, produk PCR – RAPD diskoring
berdasarkan muncul tidaknya pita DNA. Pita yang muncul pada gel diasumsikan
sebagai alel RAPD. Keragaman alel RAPD ditentukan dari perbedaan migrasi alel
pada gel dari masing- masing individu sampel. Berdasarkan ada atau tidaknya
pita, profil pita diterjemahkan kedalam data biner. Pita yang muncul diberi kode 1
(ada) dan 0 (tidak ada).
Menurut Roldan-Ruiz (2000) nilai PIC (Polymorphic Information
Content) beberapa marker dihitung dengan menggunakan rumus :
PICi = 2fi (1-fi)
Dengan:
PICi = Polymorphic Information Content pada marker
fi = frekuensi pita primer yang muncul
1-fi = frekuensi pita primer yang tidak muncul
Penentuan Ukuran Pasangan Basa
Ukuran fragmen basa (pasangan basa = bp) produk PCR ditentukan
dengan log jarak menggunakan program regresi linier. Fragmen DNA standar
(DNA ladder) digunakan sebagai absis (x) dan log jarak migrasi sebagai ordinat
(y). Dari persamaan ini ditentukan ukuran pasangan basa dari fragmen produk
PCR berdasarkan log jarak dari fragmen tersebut. Untuk selanjutnya perhitungan
dan analisis keragaman genetik penelitian ini menggunakan software GenAlEx 6.5
14
HASIL DAN PEMBAHASAN
Analisis Profil Pita Hasil Isolasi DNA Tanaman Kelapa Sawit Tipe Pisifera
Hasil dari proses isolasi DNA tanaman kelapa sawit yang dilakukan
kemudian diuji kualitasnya menggunakan gel agarose. Harahap (2014)
menyatakan uji kualitatif terhadap sampel DNA dilakukan dengan elektroforesis
gel agarose 0,8%. Uji ini dilakukan untuk mengetahui kualitas DNA yang
diperoleh. Dari uji kualitatif DNA tanaman kelapa sawit diperoleh hasil yang
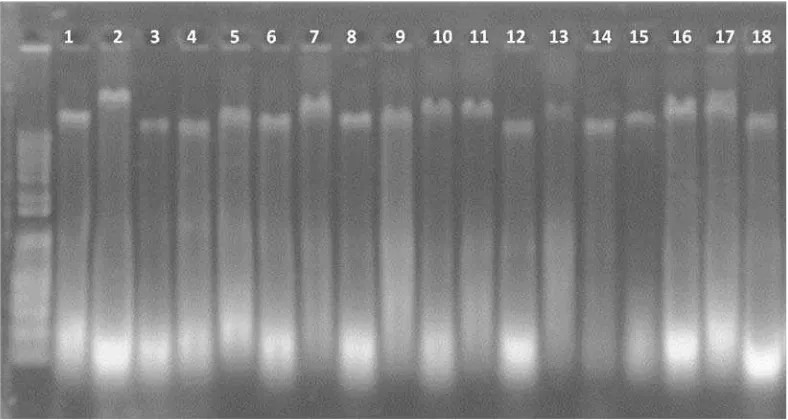
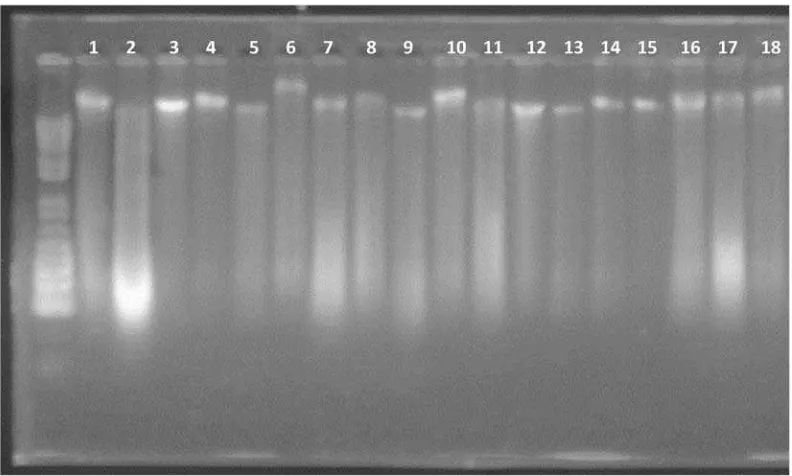
dapat dilihat pada Gambar 1-3.
15
Gambar 2. Uji kualitas 18 DNA tanaman kelapa sawit Lame origin tipe pisifera
Gambar 3. Uji kualitas 18 DNA tanaman kelapa sawit Lame silang lanjut tipe
pisifera
Uji kualitas DNA tanaman kelapa sawit Yangambi origin tipe pisifera
(Gambar 1) menunjukkan 18 sampel DNA memiliki pita yang relatif terang pada
16
pisifera (Gambar 2) menunjukan pita yang terang untuk sampel nomor 1, 2, 3, 4,
5, 14, 16, 17 dan sisanya menunjukkan pita yang tipis dan kurang terang. Uji
kualitas DNA kelapa sawit Lame silang lanjut tipe pisifera (Gambar 3)
menunjukkan pita yang terang untuk sampel nomor 1, 3, 4, 10, 12, 16 dan sisanya
menunjukkan pita yang kurang terang. Intensitas pita DNA hasil amplifikasi pada
setiap primer sangat dipengaruhi oleh kemurnian dan konsentrasi cetakan DNA
(Poerba, 2008).
Dari gambar 1-3 dapat dilihat bahwa pita dari DNA memiliki karakter
yang beragam, Prana dan Hartati (2003) menyatakan walaupun karakter yang
dihasilkan melalui penggunaan primer RAPD bisa sangat banyak, namun dalam
menentukan jenis primer (sekuen primer) dan kondisi PCR yang sesuai (dapat
annealing) untuk menghasilkan produk amplifikasi yang maksimum perlu
dilakukan penelitian tersendiri. Tobing (2014) juga menyatakan pita yang muncul
memiliki ukuran basa dan intensitas yang bervariasi. Perbedaan intensitas pita
DNA dipengaruhi oleh sebaran situs penempelan primer pada genom, kemurnian
DNA dan konsentrasi DNA dalam reaksi. Kriteria primer yang dapat digunakan
untuk analisis RAPD adalah primer yang dapat menghasilkan pita-pita polimorfik,
pita-pita yang dihasilkan jelas dan mudah dibaca (Hartati, 2007). DNA yang
diisolasi dari tanaman seringkali terkontaminasi oleh polisakarida dan metabolit
sekunder.
Teknik yang digunakan dalam percobaan kali ini adalah kombinasi
penambahan antioksidan polivinilpolipirolidon (PVPP) dan mercaptoethanol pada
buffer ekstraksinya (Syafaruddin, 2011). Pada teknik RAPD tingkat kemurnian
17
kemurnian DNA. Keuntungan lain metode RAPD adalah kuantitas DNA yang
dibutuhkan hanya sedikit (Latief, 2014), dengan kuantitas DNA yang sedikit ini,
segmen DNA nantinya dapat diamplifikasi sampai jutaan kali dalam beberapa
jam, hal ini dijelaskan oleh Handoyo dan Ari (2000) yang menyatakan bahwa
Teknik PCR dapat digunakan untuk mengamplifikasi segmen DNA dalam jumlah
jutaan kali hanya dalam waktu beberapa jam.
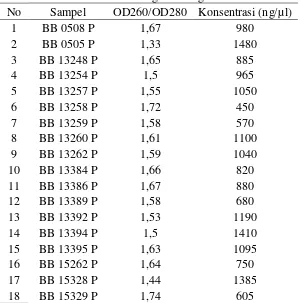
Nilai kemurnian dari sampel Yangambi origin (Tabel 3) menunjukkan
nilai terendah 1,33 dan nilai tertinggi 1,74 dengan rata-rata 1,58, sementara nilai
konsentrasi tertinggi adalah 1480 ng/µl dan terendah 450 ng/µl dengan rata-rata
963,05 ng/µl. Nilai kemurnian dari sampel Lame origin (Tabel 4) menunjukkan
nilai terendah 1,55 dan nilai tertinggi 1,77 dengan rata-rata 1,66, sementara nilai
konsentrasi tertinggi adalah 1330 ng/µl dan terendah 310 ng/µl dengan rata-rata
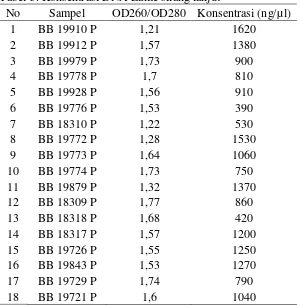
619,16 ng/µl. Nilai kemurnian dari sampel Lame silang lanjut (Tabel 5)
menunjukkan nilai terendah 1,21 dan nilai tertinggi 1,77 dengan rata-rata 1,55,
sementara nilai konsentrasi tertinggi adalah 1620 ng/µl dan terendah 390 ng/µl
dengan rata-rata 1004.44 ng/µl.
Nilai kemurnian yang didapat untuk analisis molekuler dipengaruhi oleh
metode yang yang digunakan, hal ini berkaitan dengan Pharmawati (2009) yang
menyatakan analisis molekuler tumbuhan tergantung pada jumlah dan kemurnian
sampel DNA serta kondisi optimum reaksi. Oleh karena itu diperlukan metode
yang tepat dalam ekstraksi DNA dan reaksi molekuler seperti PCR. PCR adalah
18
Tabel 3. Konsentrasi DNA Yangambi origin
No Sampel OD260/OD280 Konsentrasi (ng/µl)
1 BB 0508 P 1,67 980
2 BB 0505 P 1,33 1480
3 BB 13248 P 1,65 885
4 BB 13254 P 1,5 965
5 BB 13257 P 1,55 1050
6 BB 13258 P 1,72 450
7 BB 13259 P 1,58 570
8 BB 13260 P 1,61 1100
9 BB 13262 P 1,59 1040
10 BB 13384 P 1,66 820
11 BB 13386 P 1,67 880
12 BB 13389 P 1,58 680
13 BB 13392 P 1,53 1190
14 BB 13394 P 1,5 1410
15 BB 13395 P 1,63 1095
16 BB 15262 P 1,64 750
17 BB 15328 P 1,44 1385
18 BB 15329 P 1,74 605
Tabel 4. Konsentrasi DNA Lame origin
No Sampel OD260/OD280 Konsentrasi (ng/µl)
1 BB 8914 P 1,72 355
2 BB 8957 P 1,75 350
3 BB 8977 P 1,77 565
4 BB 8978 P 1,72 505
5 BB 8979 P 1,69 815
6 BB 8980 P 1,71 520
7 BB 8981 P 1,63 660
8 BB 8985 P 1,6 335
9 BB 8988 P 1,73 715
10 BB 8992 P 1,75 310
11 BB 10450 P 1,69 375
12 BB 13203 P 1,58 1125
13 BB 13211 P 1,55 640
14 BB 13212 P 1,61 370
15 BB 14139 P 1,54 1330
16 BB 14141 P 1,73 930
17 BB 17313 P 1,72 815
[image:32.595.114.413.429.729.2]19
Tabel 5. Konsentrasi DNA Lame silang lanjut
No Sampel OD260/OD280 Konsentrasi (ng/µl)
1 BB 19910 P 1,21 1620
2 BB 19912 P 1,57 1380
3 BB 19979 P 1,73 900
4 BB 19778 P 1,7 810
5 BB 19928 P 1,56 910
6 BB 19776 P 1,53 390
7 BB 18310 P 1,22 530
8 BB 19772 P 1,28 1530
9 BB 19773 P 1,64 1060
10 BB 19774 P 1,73 750
11 BB 19879 P 1,32 1370
12 BB 18309 P 1,77 860
13 BB 18318 P 1,68 420
14 BB 18317 P 1,57 1200
15 BB 19726 P 1,55 1250
16 BB 19843 P 1,53 1270
17 BB 19729 P 1,74 790
18 BB 19721 P 1,6 1040
Analisis Profil Pita Hasil Amplifikasi PCR Tanaman Kelapa Sawit Tipe Pisifera
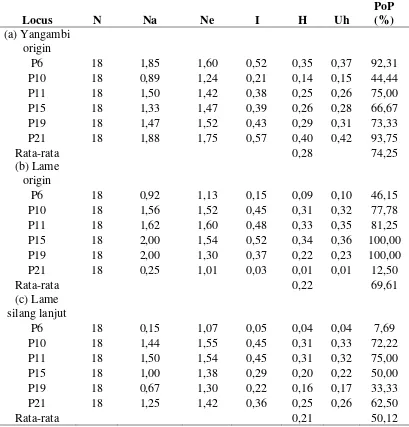
Pada Tabel 6 diketahui bahwa tingkat keragaman tertinggi terdapat
pada populasi Yangambi origin dengan rata-rata sebesar 0,28, sedangkan
keragaman terendah terdapat pada populasi Lame silang lanjut dengan rata-rata
0,21 dengan rata-rata total 0,24. Menurut Olivia (2012) dominant marker seperti
RAPD hanya dapat memproduksi dua alel pada masing-masing lokus. Maka dari
itu nilai He maksimum adalah 0,5. Dari hasil penelitian yang telah dilakukan
didapatkan nilai keragaman rata-rata adalah 0,24 yang termasuk kedalam kategori
sedang. Nilai keragaman ini nantinya akan digunakan untuk menyusun strategi
pemuliaan sesuai pernyataan dari Langga (2012) yang menyatakan bahwa
20
strategi pemuliaan pohon. Karakter genetik suatu jenis pohon baik yang terdapat
dalam satu tempat tumbuh maupun yang berbeda provenansinya, hal ini
[image:34.595.115.524.183.610.2]disebabkan karena perbedaan genetik.
Tabel 6. Keragaman genetik sawit tipe pisifera
Locus N Na Ne I H Uh
PoP (%) (a) Yangambi
origin
P6 18 1,85 1,60 0,52 0,35 0,37 92,31
P10 18 0,89 1,24 0,21 0,14 0,15 44,44
P11 18 1,50 1,42 0,38 0,25 0,26 75,00
P15 18 1,33 1,47 0,39 0,26 0,28 66,67
P19 18 1,47 1,52 0,43 0,29 0,31 73,33
P21 18 1,88 1,75 0,57 0,40 0,42 93,75
Rata-rata 0,28 74,25
(b) Lame origin
P6 18 0,92 1,13 0,15 0,09 0,10 46,15
P10 18 1,56 1,52 0,45 0,31 0,32 77,78
P11 18 1,62 1,60 0,48 0,33 0,35 81,25
P15 18 2,00 1,54 0,52 0,34 0,36 100,00
P19 18 2,00 1,30 0,37 0,22 0,23 100,00
P21 18 0,25 1,01 0,03 0,01 0,01 12,50
Rata-rata 0,22 69,61
(c) Lame silang lanjut
P6 18 0,15 1,07 0,05 0,04 0,04 7,69
P10 18 1,44 1,55 0,45 0,31 0,33 72,22
P11 18 1,50 1,54 0,45 0,31 0,32 75,00
P15 18 1,00 1,38 0,29 0,20 0,22 50,00
P19 18 0,67 1,30 0,22 0,16 0,17 33,33
P21 18 1,25 1,42 0,36 0,25 0,26 62,50
Rata-rata 0,21 50,12
(ket. Na= jumlah alel yang berbeda, Ne= jumlah alel efektif, I= index Shannon, h= keragaman dan uh= keragaman tidak bias, PoP= persentase polimorfik)
Rata-rata nilai persentase polimorfik tertinggi terdapat pada populasi
Yangambi origin sebesar 74,25%, hal ini menunjukkan bahwa tingkat persentase
21
Hartati (2007) yang menyatakan bahwa jumlah pita DNA polimorfik dalam
analisis keragaman genetik menentukan tingkat keragaman dalam suatu populasi,
sehingga banyaknya pita DNA yang polimorfis akan menggambarkan keadaaan
suatu genom dan memperkecil bias yang disebabkan oleh tidak terwakilinya
bagian-bagian genom. Penanda molekuler yang digunakan dapat menunjukkan
[image:35.595.118.512.251.429.2]perubahan yang terjadi pada material genetik.
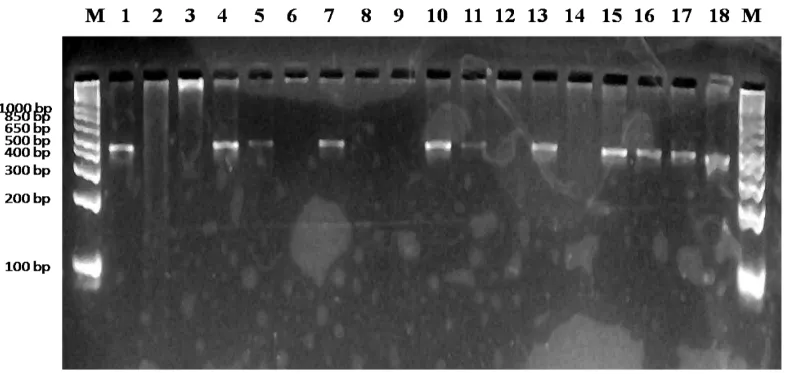
Gambar 4. Amplifikasi 18 DNA kelapa sawit Yangambi origin tipe pisifera menggunakan primer 06.
Gambar 5. Amplifikasi 18 DNA kelapa sawit Lame origin tipe pisifera
[image:35.595.119.509.487.669.2]22
Gambar 6. Amplifikasi 18 DNA kelapa sawit Lame silang lanjut tipe pisifera
menggunakan primer 06.
Pada hasil elektroforesis DNA kelapa sawit tipe pisifera menggunakan
primer 06 menunjukkan pola pita yang berbeda antara Yangambi origin (Gambar
4), Lame origin (Gambar 5) dan Lame silang lanjut (Gambar 6) dengan total pita
yang dihasilkan sebanyak 13 dengan ukuran pita berkisar antara 300 bp - 3000 bp.
Persentase pita polimorfik sebesar 92,31% untuk Yangambi origin, 46,15% untuk
Lame origin dan 7,69% untuk Lame silang lanjut. Nilai PIC primer ini (Tabel 6)
sebesar 0,41 untuk Yangambi origin, 0,10 untuk Lame origin dan 0,08 untuk
23
Gambar 7. Amplifikasi 18 DNA kelapa sawit Yangambi origin tipe pisifera menggunakan primer 10.
Gambar 8. Amplifikasi 18 DNA kelapa sawit Lame origin tipe pisifera
[image:37.595.118.505.358.578.2]24
Gambar 9. Amplifikasi 18 DNA kelapa sawit Lame silang lanjut tipe pisifera
menggunakan primer 10.
Pada hasil elektroforesis DNA kelapa sawit tipe pisifera menggunakan
primer 10 menunjukkan pola pita yang berbeda antara Yangambi origin (Gambar
7), Lame origin Gambar 8) dan Lame silang lanjut (Gambar 9) dengan total pita
yang dihasilkan sebanyak 18 dengan ukuran pita berkisar antara 220 bp - 4000 bp.
Persentase pita polimorfik sebesar 44,44% untuk Yangambi origin, 77,78% untuk
Lame origin dan 72,22% untuk Lame silang lanjut. Nilai PIC primer ini (Tabel 6)
sebesar 0,38 untuk Yangambi origin, 0,34 untuk Lame origin dan 0,49 untuk
25
Gambar 10. Amplifikasi 18 DNA kelapa sawit Yangambi origin tipe pisifera menggunakan primer 11.
Gambar 11. Amplifikasi 18 DNA kelapa sawit Lame origin tipe pisifera
[image:39.595.117.504.348.562.2]26
Gambar 12. Amplifikasi 18 DNA kelapa sawit Lame silang lanjut tipe pisifera
menggunakan primer 11.
Pada hasil elektroforesis DNA kelapa sawit tipe pisifera menggunakan
primer 11 menunjukkan pola pita yang berbeda antara Yangambi origin (Gambar
10), Lame origin (Gambar 11) dan Lame silang lanjut (Gambar 12) dengan total
pita yang dihasilkan sebanyak 16 dengan ukuran pita berkisar antara 170 bp -
3500 bp. Persentase pita polimorfik sebesar 75% untuk Yangambi origin, 81,25%
untuk Lame origin dan 75% untuk Lame silang lanjut. Nilai PIC primer ini (Tabel
6) sebesar 0,49 untuk Yangambi origin, 0,39 untuk Lame origin dan 0,48 untuk
27
Gambar 13. Amplifikasi 18 DNA kelapa sawit Yangambi origin tipe pisifera menggunakan primer 15.
Gambar 14. Amplifikasi 18 DNA kelapa sawit Lame origin tipe pisifera
[image:41.595.117.509.348.564.2]28
Gambar 15. Amplifikasi 18 DNA kelapa sawit Lame silang lanjut tipe pisifera
menggunakan primer 15.
Pada hasil elektroforesis DNA kelapa sawit tipe pisifera menggunakan
primer 15 menunjukkan pola pita yang berbeda antara Yangambi origin (Gambar
13), Lame origin (Gambar 14) dan Lame silang lanjut (Gambar 15) dengan total
pita yang dihasilkan sebanyak 12 dengan ukuran pita berkisar antara 200 bp -
3000 bp. Persentase pita polimorfik sebesar 66,67% untuk Yangambi origin,
100% untuk Lame origin dan 50% untuk Lame silang lanjut. Nilai PIC primer ini
(Tabel 6) sebesar 0,44 untuk Yangambi origin, 0,36 untuk Lame origin dan 0,34
29
Gambar 16. Amplifikasi 18 DNA kelapa sawit Yangambi origin tipe pisifera menggunakan primer 19.
Gambar 17. Amplifikasi 18 DNA kelapa sawit Lame origin tipe pisifera
[image:43.595.117.511.341.550.2]30
Gambar 18. Amplifikasi 18 DNA kelapa sawit Lame silang lanjut tipe pisifera
menggunakan primer 19.
Pada hasil elektroforesis DNA kelapa sawit tipe pisifera menggunakan
primer 19 menunjukkan pola pita yang berbeda antara Yangambi origin (Gambar
16), Lame origin (Gambar 17) dan Lame silang lanjut (Gambar 18) dengan total
pita yang dihasilkan sebanyak 15 dengan ukuran pita berkisar antara 150 bp -
4000 bp. Persentase pita polimorfik sebesar 73,33% untuk Yangambi origin,
100% untuk Lame origin dan 33,33% untuk Lame silang lanjut. Nilai PIC primer
ini (Tabel 6) sebesar 0,37 untuk Yangambi origin, 0,23 untuk Lame origin dan
31
Gambar 19. Amplifikasi 18 DNA kelapa sawit Yangambi origin tipe pisifera menggunakan primer 21.
Gambar 20. Amplifikasi 18 DNA kelapa sawit Lame origin tipe pisifera
[image:45.595.117.507.357.575.2]32
Gambar 21. Amplifikasi 18 DNA kelapa sawit Lame silang lanjut tipe pisifera
menggunakan primer 21.
Pada hasil elektroforesis DNA kelapa sawit tipe pisifera menggunakan
primer 21 menunjukkan pola pita yang berbeda antara Yangambi origin (Gambar
19), Lame origin (Gambar 20) dan Lame silang lanjut (Gambar 21) dengan total
pita yang dihasilkan sebanyak 16 dengan ukuran pita berkisar antara 200 bp -
4000 bp. Persentase pita polimorfik sebesar 93,75% untuk Yangambi origin,
12,50% untuk Lame origin dan 62,50% untuk Lame silang lanjut. Nilai PIC
primer ini (Tabel 6) sebesar 0,47 untuk Yangambi origin, 0,01 untuk Lame origin
dan 0,39 untuk Lame silang lanjut.
Ukuran pita tertinggi (Tabel 7) terdapat pada primer 10, 19 dan 21
sebesar 4000 bp untuk masing-masing populasi sedangkan untuk ukuran pita
terkecil terdapat pada primer 19 sebesar 150 bp untuk masing-masing populasi.
Frekuensi pola pita tertinggi terdapat pada primer 11 Yangambi origin sebesar
33
Tabel 7. Lokus, ukuran pita, frekuensi pola pita, persentase pita polimorfik dan
polymorphic information content (PIC) DNA sawit
Ukuran Frekuensi Persentase Pita
No Lokus Pita (bp) Pola Pita Polimorfik (%) PIC
(a) Yangambi origin
1 P6 300-3000 0,30 92,31 0,41
2 P10 220-4000 0,26 44,44 0,38
3 P11 170-3500 0,53 75,00 0,49
4 P15 200-3000 0,33 66,67 0,44
5 P19 150-4000 0,25 73,33 0,37
6 P21 200-4000 0,39 93,75 0,47
(b) Lame origin
7 P6 300-3000 0,05 46,15 0,10
8 P10 220-4000 0,22 77,78 0,34
9 P11 170-3500 0,27 81,25 0,39
10 P15 200-3000 0,24 100,00 0,36
11 P19 150-4000 0,13 100,00 0,23
12 P21 200-4000 0,01 12,50 0,01
(c) Lame silang lanjut
13 P6 300-3000 0,05 7,69 0,08
14 P10 220-4000 0,44 72,22 0,49
15 P11 170-3500 0,41 75,00 0,48
16 P15 200-3000 0,22 50,00 0,34
17 P19 150-4000 0,14 33,33 0,24
18 P21 200-4000 0,27 62,50 0,39
Total 4,53 1163,92 6,01
Rata-rata 0,25 64,66 0,33
Persentase pita polimorfik tertinggi (Tabel 7) terdapat pada primer 15
dan primer 19 Lame origin yang mencapai 100%, sedangkan persentase terendah
terdapat pada primer 06 Lame silang lanjut dengan nilai 7,69%. Nilai PIC
tertinggi terdapat pada primer 11 Yangambi origin dan primer 10 Lame silang
lanjut sebesar 0,49, sedangkan untuk nilai PIC terendah terdapat pada primer 21
Lame origin sebesar 0,01. Tasma (2013) menyatakan nilai PIC dan jumlah alel per
lokus sangat bergantung dari keragaman aksesi plasma nutfah yang diuji.
34
menggunakan teknik RAPD sangat ditentukan oleh urutan basa primer yang
digunakan serta kualitasnya atau kandungan primer dalam setiap reaksi.
Persentase nilai polimorfik yang didapatakan pada penelitian ini
menunjukkan bahwa metode RAPD cocok untuk studi tentang keragaman genetik
sesuai pernyataan Anggereini (2008) yang menyatakan bahwa Metode RAPD
merupakan metode untuk mengidentifikasi sejumlah besar polimorfisme DNA
pada genom dengan cepat dan efisien. Tipe polimorfisme ini membuat RAPD
cocok untuk studi keanekaragaman genetik, hubungan kekerabatan, peta genetik,
sidik jari DNA.
Prana dan Hartati (2003) menyatakan untuk tujuan analisis keragaman,
hal penting lainnya adalah pemilihan primer yang dapat menampilkan
polimorfisme pita-pita DNA diantara individu yang diuji. Kualitas pita DNA yang
tajam juga penting untuk memudahkan interpretasi dan keakuratan data. Beberapa
faktor yang dapat mempengaruhi kualitas pita DNA produk amplifikasi PCR
dalam analisis RAPD adalah konsentrasi MgCl2, konsentrasi DNA, konsentrasi
enzim polimerase, primer dan suhu siklus PCR terutama suhu annealing. Langga
(2012) menyatakan bahwa keragaman genetik dapat diamati dengan pengamatan
karakter genetik, sifat yang diamati adalah DNA yang sulit dipengaruhi
lingkungan. Yusuf (2010) menyatakan bahwa variasi genetik dilihat dari
35
KESIMPULAN DAN SARAN
Kesimpulan
Dari 18 sampel untuk setiap populasi Yangambi origin, Lame origin dan
Lame silang lanjut sawit tipe pisifera dapat diketahui tingkat keragaman genetik
yang sedang dengan nilai rata-rata 0,24 dengan nilai tertinggi 0,28 dan nilai
terendah 0,21. Persentase pita polimorfik tertinggi terdapat pada primer 15 dan
primer 19 Lame origin yang mencapai nilai 100%. Nilai PIC tertinggi terdapat
pada primer 11 Yangambi origin dan primer 10 Lame sl sebesar 0,49, sedangkan
untuk nilai PIC terendah terdapat pada primer 21 Lame origin sebesar 0,01.
Saran
Perlu dilakukan penelitian lebih lanjut untuk menambah keragaman
genetik tanaman kelapa sawit tipe pisifera dengan menggunakan primer yang
DAFTAR PUSTAKA
Aggereini, E. 2008. Random amplified polymorphic DNA (RAPD), suatu metode analisis DNA dalam menjelaskan berbagai fenomena biologi. Biospecies, 1 (2): 73 - 76.
Arumsari, S. 2013. Analisis diversitas genetik aksesi kelapa sawit Kamerun berdasarkan marka SSR. Jurnal Littri, 19 (4): 194 - 202.
Badan litbang pertanian. 2013. Inovasi Kultur Jaringan Kelapa Sawit. Sinartani Edisi 23-29 Januari 2013 No.3491 Tahun XLIII.
Dwiatmini, K., N.A. Mattjik, H. Aswidinoor, N.I.T. Matius. 2003. Analisis
pengelompokan dan hubungan kekerabatan spesies Anggrek Phalaenopsis
berdasarkan kunci determinasi fenotifik dan marka molekuler RAPD. J. Hort., 13(1): 16-27.
Faatih, M. 2009. Isolasi dan digesti DNA kromosom. Jurnal Penelitian Sains & Teknologi, 10 (1): 61 - 67.
Fathurrahman. 2013. Perbandingan komposisi asam lemak kelapa sawit
(Elaeis guineensis Jacq.) hasil transformasi genetik. Jurnal Agroteknologi, 3 (2):11 - 20.
Gusmiaty. 2012. Seleksi primer untuk analisis keragaman genetik jenis bitti
(Vitex coffassus). Jurnal Perennial, 2012 8 (1): 25-29.
Handoyo, D dan A. Rudiretna. 2000. Prinsip umum dan pelaksanaan
polymerase chain reaction (PCR), Unitas, 9 (1):17 - 29.
Hartati, D. 2007. Pendugaan keragaman genetik di dalam dan antar provenan pulai (Alstonia scholaris(L.) R. Br. ) menggunakan marka RAPD. Jurnal pemuliaan tanaman hutan, 1(2): 1-9.
Harahap, A. S. 2014. Keragaman genetik aren asal Sulawesi tenggara berdasarkan
marka Random Amplified Polymorphic DNA. Tesis. USU. Tidak
dipublikasikan.
Hetharie, H. 2010. deteksi perubahan genetik pada kelapa sawit
(Elaeis guinnensis jacq.) abnormal dengan teknik RAPD. Jurnal Budidaya Pertanian, 6: 45-50.
Langga, I. F., M. Restu, T. Kuswinanti. 2012. Optimalisasi suhu dan lama
inkubasi dalam ekstraksi DNA tanaman bitti (Vitex cofassus Reinw)
Latief, W. dan Amien, S. 2014. Studi awal pemanfaatan marka molekuler RAPD untuk penentuan kebenaran tiga kultivar nilam. Bionatura, 16: 109 - 113.
Manurung, E. G. Togu. 2001. Analisis valuasi ekonomi investasi perkebunan kelapa sawit di Indonesia. Environmental Policy and Institutional Strengthening IQC OUT-PCE-I-806-96-00002-00.
Olivia, R.D. dan Siregar, U.J. 2012. Keragaman genetik populasi sengon (Paraserianthes falcataria (L) Nielsen) pada hutan rakyat di Jawa berdasarkan penanda RAPD. Jurnal silvikultur tropika, 3(2): 130 – 136.
Peakall R. and Smouse P.E. (2012) GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research – an
update. Bioinformatics, 28, 2537-2539.
Pharmawati, M. 2009. Optimalisasi ekstraksi DNA dan PCR-RAPD pada
Grevillea spp. (Proteaceae). Jurnal Biologi, 13(1): 12 - 16.
Poerba, Y.S dan Martanti, D. 2008. Keragaman genetik berdasarkan marka
Random Amplified Polymorphic DNA pada Amorphophallus muelleri
Blume di Jawa. Jurnal biodiversitas, 9 (4): 245-249.
Prana, T.K. dan Hartati, N.S. 2003. Identifikasi Sidik Jari DNA Talas
(Colocasia esculenta L. Schott) Indonesia dengan Teknik RAPD
(Random Amplified Polymorphic DNA) Skrining Primer dan Optimalisasi Kondisi PCR. Jurnal Natur Indonesia, 5(2): 107-112.
Pratiwi, P. 2012. Analisis Variasi Genetik Beberapa Populasi Globba leucantha
Miq. di Sumatera Barat dengan Random Amplified Polymorphic DNA (RAPD). Program Pascasarjana Universitas Andalas.
Raganata, A.P. 2006. Kajian pengelolaan serbuk sari kelapa sawit
(Elaeis guinnensis jacq.) Pisifera. Thesis. IPB. Tidak dipublikasikan.
Roldan-Ruiz, I., Dendauw, J., Van, B. E. and Depicker, A. 2000. AFLP markers
reveal high polymorphic rates in ryegrasses (Lollium spp.) Mol. Breed, 6: 125-134.
Sathish, D. K and C. Mohankumar. 2007. RAPD markers for identifying oil palm
(Elaeis guineensis Jacq.) parental varieties (dura & pisifera) and the hybrid tenera. Indian Journal of Biotechnology, 6: 354 - 358.
Setiyo, I.E. (2001) Pemetaan dan keragaman genetik RAPD pada kelapa sawit Sungai Pancur (RISPA). Thesis. IPB. Tidak dipublikasikan.
Susantidiana. 2009. Identifikasi beberapa aksesi jarak pagar ( Jatropha curcas L.)
Syafaruddin. 2011. Optimasi teknik isolasi dan purifikasi DNA yang efisien
dan efektif pada kemiri sunan (Reutalis trisperma (Blanco) Airy Shaw).
Jurnal Littri, 17(1): 11 - 17.
Syakir, M. 2010. Budidaya kelapa Sawit. Aska media. Bogor.
Tasma, M. 2013. Analisis diversitas genetik aksesi kalepa sawit Kamerun berdasarkan marka SSR. Jurnal Littri, 19(4): 194 - 202.
Tobing, D. H. Y. L. 2014. Analisis RAPD (Random Amplified Polymorphic
DNA) populasi manggis (Garnicia mangostana L.) di Sumatera utara.
Tesis. USU. Tidak dipublikasikan.
Toruan M. 1997. Identification of oil palm (Elaeis guineensis Jacq.) Dura, Pisifera
and Tenera by RAPD markers. Proc. IBC., 97: 237 – 248.
Yuniastuti, E. 2005. Analisis AFLP pada abnormalitas klon-klon kelapa sawit (Elaeis Guineensis Jacq.) hasil kultur jaringan yang berbuah normal dan abnormal. Agrosains, (1): 1 - 12.
Yusuf, Z. K. 2010. Polymerase chain reaction (PCR). Saintek, 5 (6).
Zakaria, L. 2005. Random amplified polymorphic DNA (RAPD) and random
amplified microsatellite (RAMS) of ganoderma from infected oil palm and coconut stumps in Malaysia. Asia Pacific Journal of Molecular Biology and Biotechnology, 13 (1): 23-34.
Zulfahmi. 2013. Penanda DNA untuk analisis genetik tanaman. Jurnal Agroteknologi, 3 (2): 41-52.
LAMPIRAN
CTAB buffer 10%
- CTAB : 10 gr
- NaCl : 4,1 gr
Larutkan dalam akuades 100ml (Autoclave sebelum digunakan).
Buffer TRIS HCl 1 M pH 8,0
- Tris-Base : 12,11 gr
- HCl : 4,2 ml
- Akuades : 80 ml
Atur pH larutan hingga 8,0, tepatkan volume hingga 100 ml (Autoclave
sebelum digunakan).
Larutan EDTA 0,5 M pH 8,0
- EDTA : 18,61 gr
- Akuades : 80 ml
Kocok perlahan diatas magnetik stirer atur pHhingga mencapai 8,0 dengan
NaOH, tepatkan volume dengan akuades hingga 100 ml (Autoclave sebelum
digunakan).
NaCl 5 M
29,22 gr NaCl dilarutkan dengan akuades hingga 100 ml (Autoclave
sebelum digunakan).
Natrium Asetat 3 M ph 5,2
- CH3COONa : 24,609 gr
- Akuades : 80 ml
Atur pH lrutan hingga 5,2 tepatkan volume dengan akuades hingga 100 ml
Buffer ekstraksi Castillo
- CTAB 10% : 10 ml
- EDTA 0,5 M pH 8.0 : 2 ml
- Tris - HCl 1 M pH 8.0 : 5 ml
- NaCl 5 M : 12,6 ml
- Aquades steril : 20,4 ml
Setelah pelarutan, larutan kemudian di inkubasi dalam waterbath 65oC
selama minimum 30 menit.
Buffer TE ( Tris - HCl : EDTA)
- Tris - HCl 1 M pH 8.0 : 1 ml
- EDTA 0,5 M pH 8.0 : 0,2 ml
Larutkan dengan aquades hingga menjadi 100 ml.
Buffer TBE 50x
- Tris base : 27 gr
- EDTA 0,5 M pH 8.0 : 10 ml
- Boric acid : 13,75 gr