SEBARAN DAN KARAKTERISTIK PERSARANGAN Apis dorsata
binghami Cockerell (HYMENOPTERA: APIDAE)
DI HUTAN MAROS, SULAWESI SELATAN
MUHAMMAD TEGUH NAGIR
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis yang berjudul Sebaran dan Karakteristik Persarangan Apis dorsata binghami Cockerell (Hymenoptera: Apidae) di Hutan Maros, Sulawesi Selatan adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2016
Muhammad Teguh Nagir
4
RINGKASAN
MUHAMMAD TEGUH NAGIR. Sebaran dan Karakteristik Persarangan Apis dorsata binghami Cockerell (Hymenoptera: Apidae) di Hutan Maros, Sulawesi Selatan. Dibimbing oleh TRI ATMOWIDI dan SIH KAHONO.
Lebah madu hutan, Apis dorsata Fabricius merupakan penghasil utama madu di Indonesia yang mendukung sektor ekonomi nasional dan berkontribusi pada proses regenerasi hutanmelalui jasa polinasi. Lebah A. dorsata dikenal juga sebagai lebah madu hutan yang memiliki ukuran tubuh dan sarang paling besar, sarangnya berbentuk sisiran tunggal, bersarang di tempat terbuka, dan biasanya menggantung pada dahan pohon besar. Lebah Apis dorsata binghami merupakan subspesies A. dorsata yang hanya terdapat di Pulau Sulawesi dan pulau-pulau sekitarnya. Penelitian ini bertujuan untuk mengukur dan memetakan daerah sebaran sarang dan pohon persarangan, jumlah koloni, karakter pohon persarangan, dan perilaku bersarang A. d. binghami, serta kondisi biotik dan fisik dari hutan di Kabupaten Maros, Sulawesi Selatan.
Pengamatan dilakukan pada bulan Juli-November 2015 di kawasan hutan desa Laiya dan Cenrana Baru, Kecamatan Cenrana, Kabupaten Maros, Sulawesi Selatan. Waktu pengamatan dimulai pukul 07.00-17.00 WITA. Persebaran sarang diamati menggunakan metode jelajah dengan mengikuti jalur yang dilalui pencari madu di hutan dan mencatat sarang aktif dan bekas sarang yang ditemukan. Metode wawancara digunakan untuk mengumpulkan informasi tentang keberadaan sarang di lokasi tersebut. Pohon persarangan difoto bagian yang pentingnya (daun, bunga, dan buah) dan dibuat herbarium untuk keperluan identifikasi. Pengamatan karakteristik sarang meliputi tinggi sarang dari permukaan tanah, diameter pohon, karakter permukaan cabang, kemiringan dan arah percabangan, dan posisi terlindung atau tidaknya sarang. Kondisi sekitar sarang yang diamati meliputi tipe habitat, jarak pohon persarangan dari sumber air, dan pengukuran parameter fisik lingkungan meliputi kelembaban relatif, suhu, kecepatan angin, dan intensitas cahaya serta data rata-rata curah hujan, kelembaban, dan kecepatan angin dari Badan Meteorologi, Klimatologi, dan Geofisika (BMKG), Makassar, Sulawesi Selatan.
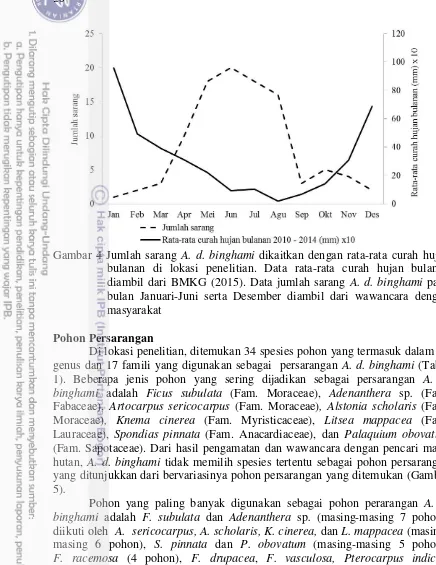
Jumlah sarang A. d. binghami yang ditemukan berjumlah 102 sarang terdiri 17 sarang aktif dan 85 bekas sarang (75 sarang di desa Laiya dan 27 sarang di desa Cenrana Baru). Sarang A. d. binghami ditemukan berkelompok dan juga menyebar pada kawasan hutan. Semua sarang ditemukan pada daerah dengan ketinggian 520-750 m dpl. Hasil pengamatan dan wawancara dengan masyarakat menunjukkan bahwa koloni A. d. binghami banyak ditemukan pada bulan April-Agustus dan menurun pada bulan Desember-Maret. Hal ini berkaitan dengan perilaku migrasi lebah A. d. binghami.
Tiga puluh empat jenis, dalam 27 genus, dan 17 famili pohon digunakan sebagai tempat persarangan A. d. binghami. Pohon yang paling sering digunakan sebagai tempat bersarang A. d. binghami yaitu Ficus subulata (Moraceae),
Adenanthera sp. (Fabaceae), Spondias pinnata (Anacardiaceae), Artocarpus sericoarpus (Moraceae), Alstonia scholaris (Moraceae), Knema cinerea
(Myristicaceae), Litsea mappacea (Lauraceae), dan Palaquium obovatum
ketinggian 11-20 m, dan 51 sarang pada ketinggian >21 m di atas permukaan tanah dengan rata-rata ketinggian dari permukaan tanah adalah 19.2 m. Pohon persarangan yang ditemukan memiliki diameter 0.1-2.52 m dengan rata-rata 0.7 m. Lima sarang dan 42 bekas sarang ditemukan pada cabang dengan sudut kemiringan 10-30°, 11 sarang dan 38 bekas sarang ditemukan pada cabang dengan sudut kemiringan 31-60°, 1 sarang dan 5 bekas sarang ditemukan pada cabang dengan dengan sudut kemiringan 61-90° dan rata-rata sudut kemiringan percabangan adalah 38°. Sembilan puluh empat persen A. d. binghami bersarang pada percabangan dengan sudut kemiringan 10-60°. Sebanyak 10 sarang (1 aktif, 9 bekas) ditemukan pada percabangan yang menghadap ke barat, 37 sarang (6 aktif, 31 bekas) menghadap ke selatan, 35 sarang (6 aktif, 29 bekas) menghadap ke timur, dan 20 sarang (4 aktif, 16 bekas) menghadap ke utara. Sembilan puluh enam persen sarang A. d. binghami ditemukan pada percabangan dengan kulit yang tidak mudah mengelupas (98 sarang) dan 4% (4 sarang) ditemukan pada pohon dengan kulit yang mudah mengelupas (Pterocarpus indicus dan
Wendlandia glabrata). Sembilan puluh persen A. d. binghami bersarang pada percabangan yang terlindungi oleh tanaman liana, daun, atau keduanya.
Sembilan puluh dua sarang (16 aktif, 76 bekas) ditemukan di hutan primer dan 10 sarang (1 aktif, 9 bekas) ditemukan di dekat perkebunan. Rata-rata jarak sarang dari sumber air yaitu 118.4 m. Rata-rata kelembaban udara pada sekitar sarang sebesar 42%, suhu sebesar 30°C, kecepatan angin sebesar 5 km/jam, dan intensitas cahaya sebesar 790 lux. Rata-rata curah hujan, kelembaban, dan kecepatan angin bulanan di lokasi penelitian 5 tahun terakhir masing-masing 310.63 mm/tahun, 81.26%, dan 6.2 km/jam.
6
SUMMARY
MUHAMMAD TEGUH NAGIR. The Distribution and Characteristic of Nesting-Site of Apis dorsata binghami Cockerell (Hymenoptera: Apidae) at Maros Forest, South Sulawesi. Supervised by TRI ATMOWIDI and SIH KAHONO.
The giant honey bee Apis dorsata Fabricius is a major producer of honey in Indonesia that support national economic sectors and contributes to the regeneration of forests and various crops through pollination services. A. dorsata
is known as the giant honey bee with large body size (>15 mm). Their nest has a large size, reaching to 1 m2, single comb, build in the open, and usually hung on the branches of a large tree. Apis dorsata binghami is subspecies of A. dorsata, that is only found in the Sulawesi and surrounding islands as endemic bee. This research aimed to measure the distribution of nest and nesting trees, characteristics of nesting trees, nesting behavior, biotic and physical condition of forest in Maros, South Sulawesi.
Observation of giant honey bees was carried out from July to November 2015 in forest at Laiya and Cenrana Baru village, Cenrana district, Maros, South Sulawesi. Survey method is used to observe giant honey bee colony, starting from 7:00 am to 5:00 pm in the forest to find the A. d. binghami nest by following the path that has been known by the wild honey hunter. The active and abandoned combs found were then recorded. Interview method also was used to find information about the existence of the nests in that location. Trees used for nesting bees were recorded, i.e., local name, photograph, and some parts of the plant (leaves, flowers, and fruits) were taken for identification. The characteristics of nest observed were height from ground level, tree diameter, characteristic, slope and direction of branching, as well as the protected and unprotected nest of honey bee. The conditions around the nest observed were the type of habitat, nest distance from the water source, and environmental parameters, i.e., relative humidity and temperature, wind speed, and light intensity. Monthly average data of rainfall, humidity, and wind speed were found from Meteorology, Climatology and Geophysics office, Makassar, South Sulawesi.
We found 102 nests of A. d. binghami in Laiya and Cerana Baru village consist of 17 active nests and 85 abandoned combs (75 nests in Laiya and 27 nests in Cenrana baru). Nests of A. d. binghami are clump in an area and spread in the forest areas. The nests were found in altitude interval 520-750 m above sea level. Based on observation and interviews with surrounding community showed that colonies of A. d. binghami are higher in April to August and lower in December to March. This is due to the migratory behavior of honey bee A. d. binghami.
Thirty four species belonging to 27 genera and 17 families of trees used as nesting site of A. d. binghami. The common trees species used as nesting site of A. d. binghami are Ficus subulata (Fam. Moraceae), Adenanthera sp. (Fam. Fabaceae), Spondias pinnata (Fam. Anacardiaceae), Artocarpus sericocarpus
(Fam. Moraceae), Alstonia scholaris (Fam. Moraceae), Knema cinerea (Fam. Myristicaceae), Litsea mappacea (Fam. Lauraceae), and Palaquium obovatum
found in tree with diameter 0.01-2.52 m, and the average diameter of trees was 0.7 m. The average branches elevation of A. d. binghami nest was in 38°. Ninety four percent of nests were found in branch with elevation 10-60°. Branches direction of nesting trees also varied. We found 10 nests (1 active, 9 abandoned combs) in branch direction to west, 37 nests (6 active, 31 abandoned combs) in branch direction to south, 35 nests (6 active, 29 abandoned combs) in branch direction to east, and 20 nests (4 active, 16 abandoned combs) in branch direction to north. Ninety nine percent of A. d. binghami nests were found in branch with hard to peel (98 nests) and 4% (4 nests) in branch with easy to peel (Pterocarpus indicus and Wendlandia glabrata). Ninety percent of nests were found in protected areas with canopy and protected by liana plants, leaves, or both.
Ninety two nests (16 active, 76 abandoned combs) were found at primary forests and 10 (1 active, 9 abandoned combs) nests at a location near the plantation. The average distance of nesting tree from water sources was 118.4 m. Around the nest, the average of air humidity was 42 %, temperature was 30°C, wind speed was 4 km/h, and light intensity was 790 lux. Monthly average data of rainfall, humidity, and wind speed at study site last five years were 310.63 mm/year, 81.26%, and 6.2 km/hour.
8
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
SEBARAN DAN KARAKTERISTIK PERSARANGAN Apis dorsata
binghami Cockerell (HYMENOPTERA: APIDAE)
DI HUTAN MAROS, SULAWESI SELATAN
MUHAMMAD TEGUH NAGIR
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biosains Hewan
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
10
Judul Tesis : Sebaran dan Karakteristik Persarangan Apis dorsata binghami
Cockerell (Hymenoptera: Apidae) di Hutan Maros, Sulawesi Selatan
Nama : Muhammad Teguh Nagir NIM : G352140091
Mayor : Biosains Hewan
Disetujui Oleh Komisi Pembimbing
Dr. Tri Atmowidi, M.Si Ketua
Dr. Sih Kahono Anggota
Diketahui
Ketua Program Studi Biosains Hewan
Dr. Ir. R.R. Dyah Perwitasari M.Sc
Dekan Sekolah Pascasarjana
Dr. Ir. Dahrul Syah, M.Sc.Agr
12
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karuniaNya sehingga karya ilmiah ini berhasil diselesaikan. Penelitian ini berjudul
“Sebaran dan Karakteristik Persarangan Apis dorsata binghami Cockerell (Hymenoptera: Apidae) di Hutan Maros, Sulawesi Selatan.” yang dilaksanakan sejak bulan Juli sampai November 2015. Terima kasih penulis ucapkan kepada Bapak Dr. Tri Atmowidi, M.Si dan Bapak Dr. Sih Kahono yang telah memberikan bimbingan dan arahan selama penelitian. Terima kasih kepada pemerintah Kabupaten Maros, Sulawesi Selatan yang telah memberikan izin untuk melakukan penelitian di kawasan hutan Maros. Terima kasih kepada Direktorat Jenderal Pendidikan Tinggi Kementerian Pendidikan dan Kebudayaan atas beasiswa Fresh Graduate dengan nomor surat 1476.2/E4.4/2014, SK Prodi 027/KL3/PP/2007, tanggal 22 Maret 2007. Terima kasih kepada Badan Meteorologi, Klimatologi dan Geofisika (BMKG) Makassar yang membantu dalam penelitian ini, kepada staf lapangan Muh. Ikhsan Ismail, Bahtiar Anas, dan masyarakat setempat di desa Laiya dan Cenrana Baru yang mendampingi selama di lapangan. Penghargaan yang setinggi-tingginya kepada Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor, dan Departemen Biologi,
Fakultas Matematika dan Ilmu Pengetahuan Alam, Fakultas Kehutanan, Universitas Hasanuddin, serta Laboratorium Herbarium, Bidang Botani, Lembaga Ilmu Pengetahuan Indonesia (LIPI) yang membantu dalam identifikasi spesimen. Penghargaan juga penulis sampaikan kepada Ibu Suhartini selaku laboran Fungsi dan Perilaku Hewan FMIPA IPB. Ungkapan terima kasih juga disampaikan kepada kedua orang tua atas doa, dukungan, dan perhatiannya yang tak terhingga, serta kepada keluarga, para sahabat, dan teman-teman seperjuangan Biosains Hewan IPB pascasarjana angkatan 2014 atas segala doa dan dukungannya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juni 2016
Muhammad Teguh Nagir
DAFTAR ISI
DAFTAR TABEL ii
DAFTAR GAMBAR ii
PENDAHULUAN 1
Latar Belakang 1
Rumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
TINJAUAN PUSTAKA 4
Lebah Madu Indonesia 4
Morfologi A. dorsata 4
Persarangan Lebah A. dorsata 5 Perilaku Migrasi A. dorsata 5
BAHAN DAN METODE 7
Lokasi dan Waktu Penelitian 7 Pengamatan Sebaran Sarang dan Identifikasi Pohon Persarangan 7 Pengukuran Karakteristik Sarang dan Kondisi Lingkungan 7
Analisis Data 7
HASIL DAN PEMBAHASAN 8
Jumlah dan Sebaran Sarang 8
Pohon Persarangan 10
Karakteristik Sarang 14
Kondisi Lingkungan Sekitar sarang 17
Pembahasan 18
SIMPULAN 21
Simpulan 21
DAFTAR PUSTAKA 22
14
DAFTAR TABEL
1 Spesies pohon persarangan A. d. binghami dan lokasi ditemukan 11 2 Parameter lingkungan di sekitar sarang A. d. binghami 18
DAFTAR GAMBAR
1 Bagan rumusan masalah 2 2 Lebah dan sarang A. d. binghami 8 3 Distribusi sarang A. d. binghami di desa Laiya dan Cenrana Baru 9 4 Jumlah sarang A. d. binghami dikaitkan dengan rata-rata curah hujan
bulanan 10
5 Pohon persarangan A. d. binghami 13 6 Jumlah dan ketinggian sarang A. d. binghami dari permukaan tanah 14 7 Lokasi sarang A. d. binghami pada pohon persarangan 14 8 Ukuran pohon persarangan A. d. binghami 15 9 Persentase sudut percabangan sarang dan kecenderungan
arah percabangan sarang A. d. binghami 16 10 Karakteristik lokasi sarang A. d. binghami 16 11 Karakteristik kondisi sarang A. d. binghami 17 12 Jarak pohon persarangan A. d. binghami dari sumber air 18
DAFTAR LAMPIRAN
PENDAHULUAN
Latar Belakang
Lebah madu hutan, Apis dorsata Fabricius merupakan penghasil utama madu di Indonesia yang mendukung sektor ekonomi nasional dan berkontribusi pada proses regenerasi hutan melalui jasa polinasi (Starr et al. 1987; Appanah 1993; Momose et al. 1998; Itioka et al. 2001). Hutan sangat bergantung kepada hewan penyerbuk, seperti lebah madu untuk penyerbukan pohon-pohon yang terpisah secara spasial (Bawa 1990; Corlett 2004; Faheem et al. 2011; Jasmi et al. 2014). Kelangsungan hidup lebah madu hutan didukung oleh ketersediaan sumber pakan dan tempat persarangan. Lebah A. dorsata selain memanfaatkan tumbuhan hutan, juga memanfaatkan tumbuhan liar dan tanaman pertanian yang ada di sekitar hutan sebagai sumber pakannya (Bawa 1990; Kahono et al. 1999; Rianti et al. 2010; Depra et al. 2014). Secara umum, penyerbukan oleh lebah madu dapat meningkatkan produksi tanaman labu-labuan 200-300%, jagung 100-150%, apel 35-60%, jeruk 300-400%, dan mentimun sebanyak 60% sehingga keberadaannya sangat penting bagi ekosistem (Sarwono 2001).
Potensi A. dorsata sebagai penghasil madu menyebabkan perburuan oleh masyarakat semakin meningkat. Perhatian yang tinggi hanya terfokus pada bidang produksi dan ekonomi, menyebabkan kurangnya perhatian terhadap peran penting
A. dorsata dalam membantu penyerbukan tanaman hutan. Kegiatan yang hanya berfokus pada pemanenan madu dapat mengancam kelangsungan hidup koloni A. dorsata. Hal ini akan berdampak pada berkurangnya jasa polinasi bagi tanaman hutan.
Lebah A. dorsata sering melakukan migrasi hingga 200 km dari sarang lama. Kondisi lingkungan yang menurun termasuk berkurangnya ketersediaan makanan dan adanya parasit larva dan pupa juga dapat menyebabkan koloni lebah bermigrasi ke lokasi lain (Koeniger dan Koeniger 1980; Paar et al. 2004; Woyke
et al. 2004; Rattanawannee dan Chanchao 2011; Makinson et al. 2014).
Lebah A. dorsata binghami Cockerell merupakan subspesies dari
A. dorsata yang hanya terdapat di Sulawesi dan pulau-pulau sekitarnya dan sampai saat ini belum berhasil dibudidayakan (Hadisoesilo 2001: Lo et al. 2010). Penelitian tentang jenis-jenis pohon yang digunakan untuk tempat bersarang lebah
A. d. dorsata, perilaku bersarang, kondisi biotik dan fisik di sekitar pohon persarangan telah dilakukan di beberapa kawasan di Jawa yang sangat penting untuk memetakan kondisinya untuk pemanfaatan dan konservasinya (Kahono et al. 1999). Penelitian serupa belum dilakukan pada lebah A. d. binghami yang berada di pulau Sulawesi dan sekitarnya.
Kawasan hutan di Kabupaten Maros merupakan tempat migrasi dari lebah
A. d. binghami, sehingga masyarakat di sekiar hutan memanfaatkannya sebagai sumber madu alam. Namun pengetahuan tentang kondisi koloni lebah dan habitat pendukungnya belum diketahui. Kawasan hutan ini merupakan salah satu ekosistem penting yang menjadi bagian dari gugusan hutan tropis dataran rendah di Sulawesi sebagai tempat migrasi lebah A. d. binghami.
2
60-82%, curah hujan tahunan 347 mm/bulan dengan rata-rata hari hujan sekitar 16 hari, temperatur udara 29oC, dan kecepatan angin 2-3 knot/jam. Musim hujan terjadi pada periode bulan Oktober-Maret dan musim kemarau pada bulan April-September (BMKG 2015).
Dalam penelitian ini dipetakan daerah sebaran sarang dan pohon persarangan, jumlah koloni, karakter pohon persarangan, perilaku bersarang A. d. binghami, dan kondisi biotik dan fisik dari hutan di Kabupaten Maros, Sulawesi Selatan. Hasil penelitian ini diharapkan dapat digunakan sebagai data dalam strategi pengelolaan dan perlindungan dari koloni dan habitat lebah A. d. binghami di Kabupaten Maros, Sulawesi Selatan.
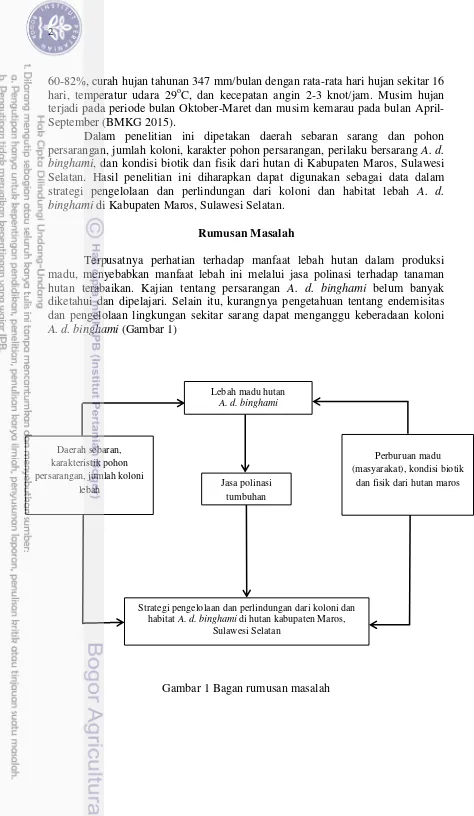
Rumusan Masalah
Terpusatnya perhatian terhadap manfaat lebah hutan dalam produksi madu, menyebabkan manfaat lebah ini melalui jasa polinasi terhadap tanaman hutan terabaikan. Kajian tentang persarangan A. d. binghami belum banyak diketahui dan dipelajari. Selain itu, kurangnya pengetahuan tentang endemisitas dan pengelolaan lingkungan sekitar sarang dapat menganggu keberadaan koloni
A. d. binghami (Gambar 1)
Gambar 1 Bagan rumusan masalah
Lebah madu hutan A. d. binghami
Jasa polinasi tumbuhan Daerah sebaran,
karakteristik pohon persarangan, jumlah koloni
lebah
Perburuan madu (masyarakat), kondisi biotik
dan fisik dari hutan maros
Strategi pengelolaan dan perlindungan dari koloni dan habitat A. d. binghami di hutan kabupaten Maros,
Tujuan Penelitian
Penelitian ini bertujuan untuk:
1. Memetakan daerah sebaran sarang dan pohon persarangan A. d. binghami di Desa Laiya dan Cenrana Baru, Kabupaten Maros, Sulawesi Selatan.
2. Mengukur jumlah koloni lebah A. d. binghami, karakter pohon persarangan, perilaku bersarang, dan kondisi biotik dan fisik dari hutan di Kabupaten Maros, Sulawesi Selatan.
Manfaat Penelitian
1. Informasi sebaran sarang dan pohon persarangan A. d. binghami dapat dijadikan gambaran keberadaan lebah madu hutan di kawasan hutan Maros. 2. Memberikan informasi pentingnya keberadaan A. d. binghami sebagai lebah
endemik dan agen penting penyerbuk tanaman hutan untuk menjaga kelangsungan hidup ekosistem hutan di Kabupaten Maros, Sulawesi Selatan. 3. Data dasar untuk strategi pengelolaan dan perlindungan koloni dan habitat dari
4
TINJAUAN PUSTAKA
Lebah Madu Indonesia
Lebah madu yang merupakan spesies asli Indonesia terdapat 5 jenis yaitu
Apis dorsata, A. andreniformis, A. cerana, A. koschevnikovi, dan A. nigrocincta
(Engel 2012). Lebah A. florea yang tersebar di Indonesia, sampai saat ini belum diketahui secara pasti area penyebarannya. Spesimen A. florea yang ada dikoleksi dari museum di Jakarta dan Surabaya (Hadisoesilo 2001). Selain itu, lebah A. nuluensis sampai saat ini baru dilaporkan di Gunung Emas, Sabah, Borneo pada ketinggian di atas 1700 m dpl (Tingek et al. 1996).
Lebah A. dorsata merupakan lebah madu dengan ukuran paling besar dan tersebar luas di Indonesia. A. dorsata dapat ditemukan hampir di seluruh kepulauan di Indonesia, kecuali Maluku dan Papua. Selain ukurannya yang besar, spesies ini juga terkenal dengan nama lebah hutan yang sangat agresif dibandingkan dengan spesies lebah madu lainnya. A. andreniformis merupakan
spesies asli dan yang paling umum dari subgenus Micrapis di Indonesia, dengan penyebaran di wilayah Kepulauan Sunda Besar dan sedikit ditemukan di Selat Makassar, Sumbawa, dan Flores. A. cerana tersebar hampir di semua kepulauan di Indonesia sampai ke Timor, kecuali Maluku dan Papua. A. cerana yang ada di Ambon dan Papua bukanlah lebah asli pulau itu melainkan didatangkan dari luar daerah tersebut. A. koschevnikovi tersebar di Semenanjung Malaya, Kalimantan, Sumatera, dan Jawa. A. nigrocincta merupakan lebah endemik Sulawesi (Ruttner 1988; Otis 1996; Kahono et al. 1999; Oldroyd et al 2000; Hadisoesilo 2001;
Hepburn dan Radloff 2011; Engel 2012).
Sarang A. dorsata berbentuk sisiran tunggal terdapat di tempat terbuka, dengan ukuran besar, luas bisa mencapai 1 m2. Pada pohon kempas (Kompassia excelsa), sarang A. dorsata dapat ditemukan pada ketinggian lebih dari 10 m di atas permukaan tanah. Sarang A. Andreniformis pada umumnya di tempat terbuka, menggantung di ranting atau dahan semak-semak atau pohon yang kecil serta terlindung dedaunan. Ketinggian sarang dari atas tanah hanya berkisar 5 m dan hanya terdiri dari satu sisir (Hepburn dan Radloff 2011). Berbeda dengan sarang A. dorsata dan A. Andreniformis, sarang dari A. cerana. A. koschevnikovi, A. nigrocincta terdiri dari beberapa sisiran dan pada umumnya terdapat di tempat yang tertutup, seperti di lubang pepohonan, celah bangunan rumah, atau tempat tertutup lainnya (Hadisoesilo 2001; Hepburn dan Radloff 2011).
Morfologi A. dorsata
sel calon ratu (Tan 2007; Beaurepaire et al. 2014). Lebah A. dorsata mempunyai panjang sayap depan mencapai 14 mm, panjang tungkai mencapai 11.5 mm dan panjang probosis mencapai 6.5 mm (Hadisoesilo 2001). Berbeda dengan lebah sosial lainnya, A. dorsata mampu melakukan pencarian pakan mulai pagi hingga malam hari karena mata tunggalnya (ocelli) berkembang baik (Dyer 1985; dan Crozier 2007; Hepburn dan Radloff 2011). A. d. brevilugula merupakan sub spesies dari A. dorsata yang area persebarannya di Filipina dan pulau-pulau sekitarnya (Sakagami et al. 1980). Perbedaan A. d. dorsata dan A. d. binghami
dapat diketahui dari warna abdomen lebah pekerjanya. Warna andomen dari A. d. dorsata agak kecoklatan dengan strip oranye, sedangkan A. d. binghami hitam dengan strip putih yang jelas (Hadisoesilo 2001).
Persarangan Lebah A. dorsata
Sarang lebah A. dorsata biasanya menggantung pada dahan pohon besar dan bergerombol pada satu pohon dengan jumlah dapat mencapai puluhan sampai ratusan sarang (Kahono et al. 1999; Hadisoesilo 2001; Hepburn dan Radloff 2011;
Mead 2013). Secara umum, A. dorsata cenderung menyukai habitus pohon yang tinggi dengan percabangan relatif terbuka dan tajuk tidak terlalu padat sebagai tempat bersarang (Starr et al. 1987; Hadisoesilo dan Kuntadi 2007; Roy et al.
2011), walaupun pernah ditemukan hanya ada satu koloni dalam satu pohon. Berbeda dengan A. d. dorsata, sarang A. d. binghami hanya ditemukan 2-3 sarang per pohon dan dibangun pada tempat yang tidak terlalu terbuka, namun tetap dapat menerima cahaya matahari (Hadisoesilo dan Kuntadi 2007). Sahebzadeh et al. (2013) melaporkan bahwa di hutan Terengganu, Malaysia, sarang A. dorsata
yang berkelompok pada satu pohon yang sama adalah unit populasi yang berbeda (P>0.05) berdasarkan perbandingan genotip ratu. Pemilihan lokasi bersarang sangat penting untuk serangga sosial, seperti A. dorsata karena berhubungan dengan keberlangsungan hidup koloni dari resiko predator, ancaman cuaca yang buruk, serta kegagalan dalam produksi (Franks et al. 2002; Neupane et al. 2013).
Perilaku Migrasi A. dorsata
Lebah A. dorsata memiliki perilaku migrasi berulang, yaitu pergi dan kembali di tempat persarangan sebelumnya (Neumann et al. 2000). Koloni lebah hutan memanfaatkan bunga di sekitarnya dalam periode waktu tertentu. Pada periode waktu lainnya, lebah ini meninggalkan tempat tersebut untuk tinggal di pohon persarangan lainnya yang memiliki musim bunga yang berbeda. Koloni
6
Tiga sifat migrasi A.dorsata dapat terjadi, yaitu migrasi koloni yang diakibatkan karena musim pembungaan, abscond yangmerupakan migrasi karena kondisi lingkungan yang memburuk, sehingga memaksa koloni untuk segera berpindah tempat secara total, dan swarming (pecah koloni) yaitu migrasi yang terjadi karena koloni sudah penuh, sehingga ratu sulit menempatkan telur (Bertoni 2013; Makinson 2013).
Perubahan lingkungan, seperti kerusakan hutan, kekurangan air dan sumber pakan, perburuan madu, dan kerusakan sarang merupakan penyebab lebah
A. dorsata bermigrasi (Oldroyd dan Nanork 2009). Populasi A. dorsata di Hutan Terengganu, Malaysia terancam akibat kurangnya usaha restorasi hutan dan perburuan lebah madu (Sahebzadeh et al. 2013). Ratu lebah akan berhenti bertelur satu minggu sebelum melakukan migrasi, sementara lebah pekerja menghabiskan madu dan memasukkannya ke dalam honey crop (Weihmann et al. 2014). Perilaku migrasi karena swarming dilaporkan di India, yaitu terjadi penambahan jumlah koloni A. dorsata pada bangunan di Bangalore dan tujuh koloni diantaranya dalam keadaan swarming. Musim pembungaan pada awal Desember di Nepal juga menyebabkan banyaknya koloni yang bermigrasi ke lokasi tersebut, tanpa ada koloni yang pergi ke tempat lain (Woyke et al. 2012). Lebah A. dorsata
BAHAN DAN METODE
Lokasi dan Waktu Penelitian
Pengamatan persarangan A. d. binghami dilakukan di kawasan hutan desa Laiya dan Cenrana Baru, Kecamatan Cenrana, Kabupaten Maros, Sulawesi Selatan pada bulan Juli sampai November 2015. Pengamatan harian dimulai pukul 07.00-17.00 WITA.
Pengamatan Sebaran Sarang dan Identifikasi Pohon Persarangan
Pengamatan sebaran sarang A. d. binghami menggunakan metode jelajah (Bookhout 1996) yaitu menjelajahi hutan untuk mencari sarang dan pohon persarangannya dengan mengikuti jalur yang telah diketahui oleh para pencari madu hutan. Metode wawancara juga digunakan untuk mengumpulkan informasi tentang keberadaan sarang pada lokasi tersebut (Tongco 2007; Thomas et al.
2009). Daftar pertanyaan untuk wawancara terdapat dalam Lampiran 1. Nama lokal pohon persarangan dicatat dan difoto bagian penting pohonnya (daun, bunga, buah, serta bagian penting lainnya) dan dibuat herbarium untuk keperluan identifikasi. Jumlah koloni A. d. binghami yang masih aktif, bekas sarang, dan posisi sarang pada dahan pohon diamati dan dicatat (Neupane et al. 2013). Posisi kordinat pohon persarangan lebah diukur menggunakan GPS Garmin map 62sc. Identifikasi koleksi spesimen pohon persarangan dilakukan dengan membandingkan dengan spesimen herbarium koleksi Fakultas Kehutanan, Universitas Hasanuddin, Makassar. Spesimen herbarium lainnya diidentifikasi di Herbarium Bogoriense, Bidang Botani, Pusat Penelitian Biologi-LIPI, Bogor, Jawa Barat.
Pengukuran Karakteristik Sarang dan Kondisi Lingkungan
Karakteristik sarang yang diukur adalah tinggi sarang dari permukaan tanah menggunakan hagameter, diameter pohon persarangan menggunakan DBH meter (diameter at breast high), karakter permukaan cabang, kemiringan dan arah percabangan, dan terlindung atau tidaknya sarang. Kondisi lingkungan sekitar sarang dan jarak pohon persarangan dari sumber air juga diamati. Pengukuran parameter fisik lingkungan meliputi kelembaban relatif dan suhu menggunakan thermohygrometer, kecepatan angin menggunakan anemometer, dan intensitas cahaya menggunakan lux meter. Data curah hujan, kelembaban udara, dan kecepatan angin didapatkan dari Badan Meteorologi, Klimatologi, dan Geofisika (BMKG), Makassar, Sulawesi Selatan.
Analisis Data
8
HASIL DAN PEMBAHASAN
Hasil
Jumlah dan Sebaran Sarang
Jumlah sarang A. d. binghami (Gambar 2a) yang ditemukan di hutan desa Laiya dan Cenrana Baru, Kecamatan Cenrana, Kabupaten Maros sebanyak 102 sarang yang terdiri 17 sarang aktif dan 85 bekas sarang (75 sarang di Laiya dan 27 sarang di Cenrana Baru) (Gambar 2b dan 2c). Sarang aktif yang ditemukan merupakan sarang yang sudah ditandai oleh pemiliknya dan siap untuk dipanen. Bekas sarang yang ditemukan merupakan sarang yang sudah dipanen oleh pemiliknya.
Gambar 2 Lebah dan sarang A. d. binghami: Lebah pekerja (a), sarang aktif (b), dan bekas sarang (c) (dalam lingkaran).
(a)
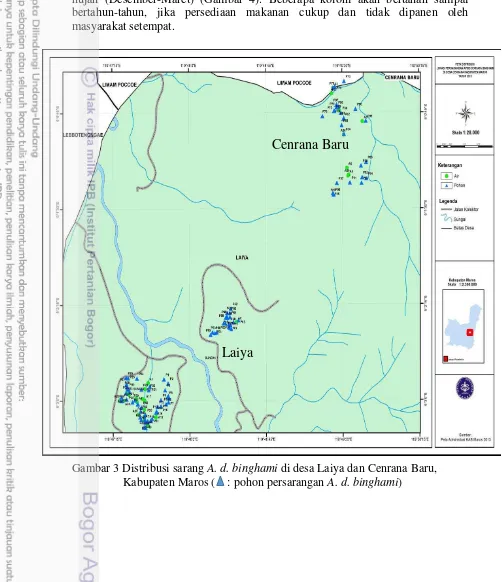
Sarang A. d. binghami ditemukan berkelompok dan juga menyebar pada kawasan hutan (Gambar 3). Semua sarang ditemukan pada daerah dengan ketinggian 520-750 m dpl. Hasil pengamatan dan wawancara dengan masyarakat menunjukkan bahwa koloni A. d. binghami banyak ditemukan pada bulan April-Agustus dengan puncak kedatangannya pada bulan Juni dan menurun pada musim hujan (Desember-Maret) (Gambar 4). Beberapa koloni akan bertahan sampai bertahun-tahun, jika persediaan makanan cukup dan tidak dipanen oleh masyarakat setempat.
Gambar 3 Distribusi sarang A. d. binghami di desa Laiya dan Cenrana Baru, Kabupaten Maros ( : pohon persarangan A. d. binghami)
Laiya
10
Gambar 4 Jumlah sarang A. d. binghami dikaitkan dengan rata-rata curah hujan bulanan di lokasi penelitian. Data rata-rata curah hujan bulanan diambil dari BMKG (2015). Data jumlah sarang A. d. binghami pada bulan Januari-Juni serta Desember diambil dari wawancara dengan masyarakat
Pohon Persarangan
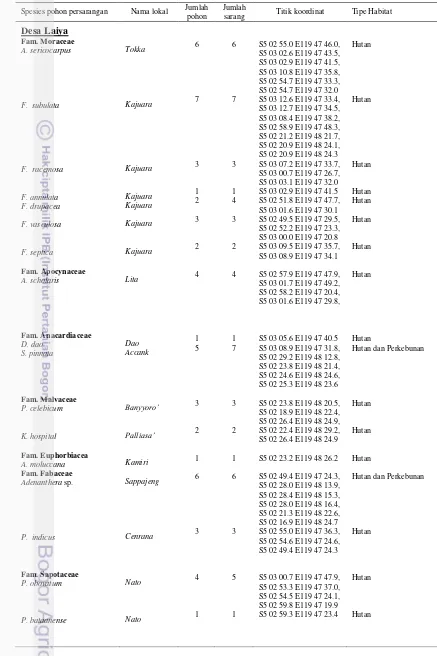
Di lokasi penelitian, ditemukan 34 spesies pohon yang termasuk dalam 27 genus dan 17 famili yang digunakan sebagai persarangan A. d. binghami (Tabel 1). Beberapa jenis pohon yang sering dijadikan sebagai persarangan A. d. binghami adalah Ficus subulata (Fam. Moraceae), Adenanthera sp. (Fam. Fabaceae), Artocarpus sericocarpus (Fam. Moraceae), Alstonia scholaris (Fam. Moraceae), Knema cinerea (Fam. Myristicaceae), Litsea mappacea (Fam. Lauraceae), Spondias pinnata (Fam. Anacardiaceae), dan Palaquium obovatum
(Fam. Sapotaceae). Dari hasil pengamatan dan wawancara dengan pencari madu hutan, A. d. binghami tidak memilih spesies tertentu sebagai pohon persarangan yang ditunjukkan dari bervariasinya pohon persarangan yang ditemukan (Gambar 5).
Pohon yang paling banyak digunakan sebagai pohon perarangan A. d. binghami adalah F. subulata dan Adenanthera sp. (masing-masing 7 pohon), diikuti oleh A. sericocarpus, A. scholaris, K. cinerea, dan L. mappacea
(masing-masing 6 pohon), S. pinnata dan P. obovatum (masing-masing 5 pohon),
F. racemosa (4 pohon), F. drupacea, F. vasculosa, Pterocarpus indicus, P. bataanense, Pterospermum celebicum, Pinus merkusii (masing-masing 3
pohon), dan F. septica, Toona sureni, Barringtonia sp., Syzygium sp., Acer laurinum, Klenhovia hospita, Mangifera indica, Lithocarpus celebicus (masing-masing 2 pohon).
Tumbuhan yang paling sedikit dijadikan sebagai persarangan yaitu
Baccaurea sp., F. annulata, Dracontamelon dao, Barringtonia acutangula,
Aleurites moluccana, Buchanania arborescens, Arthrophyllum diversifolium,
Macaranga sp., Melochia umbellata, Persea sp., dan Wendlandia glabrata
Tabel 1 Spesies pohon persarangan A. d. binghami dan lokasi ditemukan
Spesies pohon persarangan Nama lokal Jumlah pohon
Jumlah
Gambar 5 Pohon persarangan A. d. binghami: F. subulata (a), A. scholaris (b),
P. merkusii (c), F. racemosa (d), A. moluccana (e), F. drupacea (f),
L. mappacea (g), dan L. celebicus (h).
(a)
(c) (d)
(b)
(e)
(g) (h)
14
Karakteristik Sarang
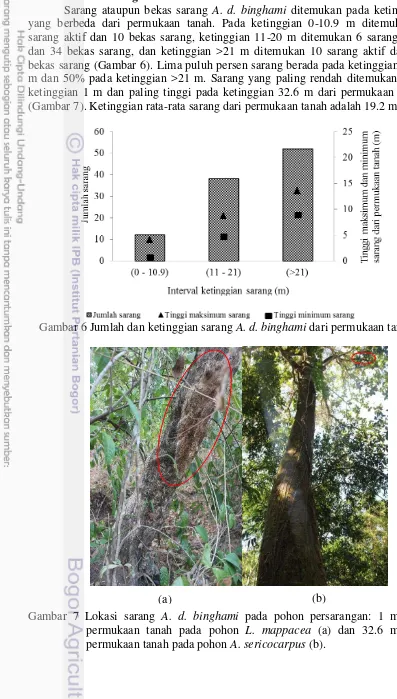
Sarang ataupun bekas sarang A. d. binghami ditemukan pada ketinggian yang berbeda dari permukaan tanah. Pada ketinggian 0-10.9 m ditemukan 1 sarang aktif dan 10 bekas sarang, ketinggian 11-20 m ditemukan 6 sarang aktif dan 34 bekas sarang, dan ketinggian >21 m ditemukan 10 sarang aktif dan 41 bekas sarang (Gambar 6). Lima puluh persen sarang berada pada ketinggian 0-20 m dan 50% pada ketinggian >21 m. Sarang yang paling rendah ditemukan pada ketinggian 1 m dan paling tinggi pada ketinggian 32.6 m dari permukaan tanah (Gambar 7). Ketinggian rata-rata sarang dari permukaan tanah adalah 19.2 m.
Gambar 6 Jumlah dan ketinggian sarang A. d. binghami dari permukaan tanah
Gambar 7 Lokasi sarang A. d. binghami pada pohon persarangan: 1 m dari permukaan tanah pada pohon L. mappacea (a) dan 32.6 m dari permukaan tanah pada pohon A. sericocarpus (b).
Pohon persarangan yang ditemukan memiliki diameter yang bervariasi. Diameter terkecil (0.1 m) pada pohon S. pinnata dan diameter terbesar (2.52 m) pada pohon A. scholaris (Gambar 8) dengan rata-rata diameter pohon 0.7 m.
Gambar 8 Ukuran pohon persarangan A. d. binghami: pohon terkecil (diameter 0.1 m) pada pohon S. pinnata (tanda panah) (a) dan pohon terbesar (diameter 2.52 m) pada pohon A. scholaris (tanda panah) (b).
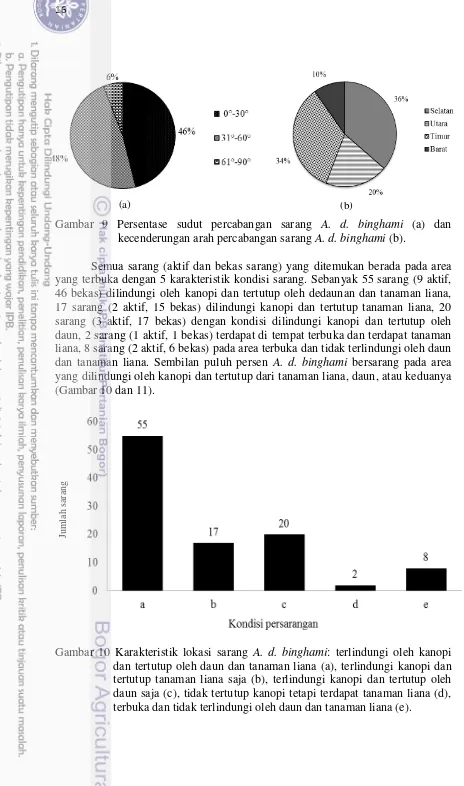
Sarang A. d. binghami ditemukan pada cabang pohon dengan sudut yang bervariasi. Lima sarang dan 42 bekas sarang ditemukan pada sudut kemiringan 10-30°. Sebanyak 11 sarang dan 38 bekas sarang ditemukan pada cabang dengan sudut kemiringan 31-60°. Sebanyak 1 sarang dan 5 bekas sarang ditemukan pada cabang dengan sudut kemiringan 61-90°. Sembilan puluh empat persen A. d. binghami bersarang pada percabangan dengan sudut kemiringan 10-60° (Gambar 9). Arah percabangan pohon persarangan juga bervariasi. Sepuluh sarang (1 aktif, 9 bekas) ditemukan pada percabangan yang menghadap ke barat, 37 sarang (6 aktif, 31 bekas) ditemukan pada percabangan yang menghadap ke selatan, 35 sarang (6 aktif, 29 bekas) ditemukan pada percabangan yang menghadap ke timur, 20 sarang (4 aktif, 16 bekas) yang menghadap ke utara. Sarang A. d. binghami
ditemukan pada percabangan dengan kulit yang tidak mudah mengelupas (96%) dan 4% ditemukan pada pohon dengan kulit yang mudah mengelupas (P. indicus
dan W. glabrata).
16
Ju
m
la
h
sa
ra
n
g
Gambar 9 Persentase sudut percabangan sarang A. d. binghami (a) dan kecenderungan arah percabangan sarang A. d. binghami (b).
Semua sarang (aktif dan bekas sarang) yang ditemukan berada pada area yang terbuka dengan 5 karakteristik kondisi sarang. Sebanyak 55 sarang (9 aktif, 46 bekas) dilindungi oleh kanopi dan tertutup oleh dedaunan dan tanaman liana, 17 sarang (2 aktif, 15 bekas) dilindungi kanopi dan tertutup tanaman liana, 20 sarang (3 aktif, 17 bekas) dengan kondisi dilindungi kanopi dan tertutup oleh daun, 2 sarang (1 aktif, 1 bekas) terdapat di tempat terbuka dan terdapat tanaman liana, 8 sarang (2 aktif, 6 bekas) pada area terbuka dan tidak terlindungi oleh daun dan tanaman liana. Sembilan puluh persen A. d. binghami bersarang pada area yang dilindungi oleh kanopi dan tertutup dari tanaman liana, daun, atau keduanya (Gambar 10 dan 11).
Gambar 10 Karakteristik lokasi sarang A. d. binghami: terlindungi oleh kanopi dan tertutup oleh daun dan tanaman liana (a), terlindungi kanopi dan tertutup tanaman liana saja (b), terlindungi kanopi dan tertutup oleh daun saja (c), tidak tertutup kanopi tetapi terdapat tanaman liana (d), terbuka dan tidak terlindungi oleh daun dan tanaman liana (e).
Gambar 11 Karakteristik kondisi persarangan A. d. binghami: terlindungi oleh kanopi dan tertutup oleh daun dan tanaman liana (a), terlindungi kanopi dan tertutup tanaman liana saja (b), terlindungi kanopi dan tertutup oleh daun saja (c), tidak tertutup kanopi tetapi terdapat tanaman liana (d), terbuka dan tidak terlindungi oleh daun dan tanaman liana (e).
Kondisi Lingkungan Sekitar Sarang
Sarang A. d. binghami yang ditemukan sebanyak 102 sarang (17 aktif, 85 bekas) ditemukan di hutan primer dan di dekat perkebunan masyarakat. Sembilan puluh dua sarang (16 aktif, 76 bekas) ditemukan di hutan primer dan 10 sarang (1 aktif, 9 bekas) ditemukan di dekat perkebunan masyarakat. Jarak pohon persarangan dari sumber air juga bervariasi (1.5-470 m). Koloni A. d. binghami
cenderung bersarang di dekat sumber air (Gambar 12). Rata-rata jarak pohon persarangan dari sumber air yaitu 118.4 m. Rata-rata kelembaban udara di sekitar sarang adalah 42%, suhu berkisar 30°C, kecepatan angin sebesar 5 km/jam, dan intensitas cahaya sebesar 790 lux (Tabel 2). Rata-rata curah hujan, kelembaban, dan kecepatan angin dari tahun 2010-2014 di lokasi penelitian masing-masing 310.63 mm/tahun, 81.26%, dan 6.2 km/jam.
(a) (b) (c)
18
Gambar 12 Jarak pohon persarangan A. d. binghami dari sumber air Tabel 2 Parameter lingkungan di sekitar sarang A. d. binghami
Pembahasan
Berdasarkan pengamatan dan wawancara dengan masyarakat sekitar lokasi penelitian, bahwa koloni A. d. binghami tidak ditemukan sepanjang tahun. Hal ini berkaitan dengan perilaku migrasi A. d. binghami. Koloni A. d. binghami banyak ditemukan pada bulan April-Agustus dengan puncak kedatangannya di bulan Juni dan sedikit ditemukan pada bulan Desember-Maret. Hal ini berkaitan dengan tingginya curah hujan pada bulan Desember-Maret (BMKG 2015). Lebah
A. dorsata melakukan migrasi paling sedikit dua kali dalam setahun pada dua tempat secara bergantian dan mampu melakukan migrasi hingga 200 km dari sarang lama. Migrasi tersebut dapat dipicu oleh kerusakan lingkungan, berkurangnya ketersediaan makanan, dan parasit pada larva dan pupa (Koeniger dan Koeniger 1980; Momose et al. 1998; Paar et al. 2004; Woyke et al. 2004; Rattanawannee dan Chanchao 2011; Makinson et al. 2014). Dalam pengamatan, satu sarang A. d. binghami selama dua tahun tetap bersarang di pohon tanpa melakukan migrasi. Hal ini karena pemilik sarang tidak memanen madu dari sarang selama kurun waktu tersebut dan diduga tersedianya pakan dan air yang cukup. Oldroyd dan Nanork (2009) menyatakan bahwa kondisi hutan yang rusak, kekurangan air dan pakan, serta perburuan lebah dengan merusak sarang, merupakan penyebab lebah bermigrasi ke lokasi yang lebih baik.
Parameter Lingkungan Nilai Minimum
Nilai Maksimum
Rata-rata
Suhu (°C) 24 34.4 30.08 ± 1.85
Kelembaban (%) 29 75 42.19 ± 8.45
Intensitas Cahaya x10 30 512 78.76 ± 63.5
Pohon persarangan A. d. binghami sangat bervariasi mulai dari pohon yang besar dan tinggi sampai pada pohon yang rendah. Kecenderungannya, lebah A. d. binghami tidak memilih jenis pohon, ketinggian dari tanah, dan arah percabangan dalam membuat sarang. Starr et al. (1987) melaporkan bahwa pada A. d. dorsata
di Kalimantan bersarang pada 15 jenis pohon dan cenderung tidak memilih pohon dalam membuat sarang. Hadisoesilo dan Kuntadi (2007) juga melaporkan di Riau,
A. d. dorsata bersarang pada beberapa pohon, seperti beringin (Ficus sp.), kempas (Koompassia excelsa), lumbuai (Metroxylon sp.), siluang (Polythias hipolenca),
jangkang (Dellenia exinia), dan mahang (Macaranga sp.). Di Sulawesi, A. d. binghami ditemukan bersarang pada pohon durian (Durio zibethinus), mangga (Mangifera sp.), jambu (Syzygium sp.), dan kapuk randu (Ceiba pentandra).
Beberapa penelitian juga melaporkan A. d. dorsata cenderung menyukai pohon kempas (Koompassia excelsa) sebagai tempat bersarang (Starr et al. 1987; Hadisoesilo 2001). Di Kebun Raya Bogor, Jawa Barat A. d. dorsata cenderung memilih F. albipila sebagai pohon persarangan (72%) (Kahono et al. 1999). Koloni A. d. binghami dalam membangun sarang tidak memilih jenis, namun beberapa karakteristik pohon dipilih sebagai pohon persarangan (Hadisoesilo dan Kuntadi 2007).
Sepuluh persen sarang A. d. binghami ditemukan pada ketingian <10 m dan 90% pada ketinggian >10 m dari permukaan tanah. Kahono et al. (1999) juga melaporkan sarang A. d. dorsata terletak pada ketinggian <10 m dari permukaan tanah selama jangka waktu dua tahun. Weihmann et al. (2014) juga melaporkan bahwa A. d. dorsata memilih pohon yang tinggi untuk membuat sarangnya. Beberapa penelitian juga melaporkan A. d. dorsata cenderung menyukai tempat yang tinggi (>10 m) untuk membuat sarang (Starr et al. 1987; Kahono et al. 1999; Hadisoesilo 2001). Pemilihan tempat bersarang A. d. binghami yang tinggi dari permukaan tanah bertujuan untuk menghindari ancaman vertebrata (Starr et al. 1987). Sarang A. d. binghami yang rendah mengindikasikan kurang atau tidak adanya predator yang dapat membahayakan koloni lebah.
Lebah A. d. binghami cenderung bersarang pada percabangan dengan sudut kemiringan cabang 10-60° (94%) dan pada cabang pohon yang kuat dan kulit cabangnya tidak mudah mengelupas (96%). Hal ini didukung publikasi Kahono et al. (1999) bahwa koloni A. d. dorsata yang ditemukan di Kebun Raya Bogor, Jawa Barat cenderung bersarang pada cabang dengan sudut kemiringan 50°. Koloni A. d. dorsata tidak membuat sarang pada bangunan tua, cabang yang rapuh, ataupun pohon yang sudah mati (Neupane et al. 2013).
Lebah A. d. binghami bersarang dengan menggantung di cabang pohon yang terbuka dan cenderung menyukai pohon dengan kondisi yang terlindungi oleh tanaman liana, daun, atau terlindungi oleh keduanya. Hasil penelitian ini berbeda dengan penelitian Starr et al. (1987) yang melaporkan bahwa A. d. dorsata cenderung membuat sarang pada lokasi yang terbuka dan bebas dari tanaman liana. Weihmann et al. (2014) juga melaporkan, A. d. dorsata cenderung membuat sarang pada lokasi lebih terbuka dan tidak terlindungi. Lebah A. d. dorsata pada umumnya membangun sarang secara berkelompok dalam satu pohon pada cabang yang bersih dan bebas dari tanaman epifit (Roy et al. 2011).
20
predator seperti, beberapa jenis burung yang sering menyerang sarang A. d. binghami. Oldroyd dan Nanork (2009) melaporkan beberapa jenis burung predator lebah madu, antara lain Indicator xanthonotus, I. archipelagicus, Pernis ptilorhyncus, dan P. celebensis.
Lokasi persarangan A. d. binghami yang diamati terdapat di hutan primer dan perkebunan dengan kondisi vegetasi yang rapat dan jenis pohon yang bervariasi. Kondisi ini mendukung keberlangsungan hidup koloni A. d. binghami
dengan ketersediaan sumber makanan. Lebah menunjukkan flower constancy
terhadap sumber pakan utama yang terdekat dari sarangnya (Sadeh et al. 2007; Bernardino dan Gaglianone 2008; Rianti et al. 2010). Keberadaan bunga sebagai sumber polen dan nektar merupakan hal yang sangat penting untuk kelangsungan hidup koloni lebah. Penelitian lainnya melaporkan bahwa Apis sp. merupakan lebah penyerbuk yang sangat potensial pada berbagai jenis tanaman. Pada tanaman pertanian seperti caisin (Brassica rapa), A. dorsata berperan penting sebagai lebah penyerbuk (Atmowidi 2008).
SIMPULAN
Di dua desa yang diamati yaitu desa Laiya dan Cenrana Baru ditemukan 102 sarang A. d. binghami yang terdiri dari 17 sarang aktif dan 85 bekas sarang. Tiga puluh empat spesies pohon yang digunakan sebagai persarangan A. d. binghami yang termasuk dalam 27 genus dan 17 famili. Pohon yang sering
dijadikan sebagai persarangan adalah F. subulata, Adenanthera sp.,
A. sericocarpus, A. scholaris, K. cinerea, L. mappacea, S. pinnata, dan P. obovatum. Lima puluh persen sarang A. d. binghami ditemukan pada ketinggian 0-20 m dan 50% pada ketinggian >21 m di atas permukaan tanah. Koloni A. d. binghami cenderung membuat sarang pada cabang yang berkayu dan kulitnya tidak mudah mengelupas dengan sudut kemiringan 10-60°, serta terlindungi kanopi dan tertutupi oleh tanaman liana, daun, atau keduanya.
22
DAFTAR PUSTAKA
Appanah S. 1993. Mass flowering of dipterocarp forests in the aseasonal tropics.
J Biosci. 18(4): 457–474.
Atmowidi T. 2008. Keanekaragaman dan Perilaku Kunjungan Serangga Penyerbuk serta Pengaruhnya dalam Pembentukan Biji Tanaman Caisin (Brassica rapa L: Brassicaceae) [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Bawa KS. 1990. Plant-pollinator interactions in tropical rain forest. Annu Rev Ecol Syst 21:399-422
Bernardino AS, Gaglianone MC. 2008. Nest distribution and nesting habits of
Xylocopa ordinaria Smith (Hymenoptera, Apidae) in a resting area in the northern rio de Janeiro State, Brazil. Revis Brasil Entomol. 52(3):434-440. Bertoni R. 2013. Perbandingan Ukuran-Ukuran Bagian Tubuh Lebah Pekerja Apis
dorsata (Lebah Hutan) pada Empat Lokasi. skripsi. Bogor (ID). Institut Pertanian Bogor.
Beaurepaire AL, Kraus BF, Koeniger G, Koeniger N, Lim H, Moritz RFA. 2014. Extensive population admixture on drone congregation areas of the giant honey bee, Apis dorsata (Fabricius, 1793). Ecol and Evol 4(24):4669-4677.doi:10.1002/ece3.1284.
[BMKG] Badan Meteorologi Klimatologi, dan Geofisika. 2015. Data Iklim Bulanan Stasiun Klimatologi Maros. Makassar (ID): BMKG.
Bookhout TA. 1996. Research and Management Techniques for Wildlife and Habitats. Kansas (US): Allen Press Inc.
Corlett RT. 2004. Flower visitors and pollination in the oriental (Indomalayan) region. Biol Rev. 79, 497–532.
Depra MS, Delaqua GCG, Freitas L, Gaglianone MC. 2014. Pollination deficit in open-field tomato crops (Solanum lycopersicon L., Solanaceae). J Poll Ecol. 12(1):1-8.
Dyer FC. 1985. Nocturnal orientation by the asian honey bee, Apis dorsata.
Animal Behavior. 33:769–774.
Engel MS. 2012. The honey bees of Indonesia (Hymenoptera: Apidae). Treubia.
39:41-49.
Faheem M, Aslam M, Razaq M. 2004. Pollination ecology with special reference to insects a review. J Res Sci. 4:395-409.
Fransk NR, Pratt SC, Mallon EB, Britton NF, Sumpter DJT. 2002. Information flow, opinion polling and collective intelligence in house-hunting social insects. PTRSL. 357: 1567-1583.
Gerling D, Velthuis HHW, Hefetz A. 1989. Bionomics of the large carpenter bee
Xylocopa pubescens and its implications for the evolution of sociality.
Annu Rev Entomol. 2:123–128.
Gottlieb D, Keasar T, Shimida A, Motro U. 2005. Possible foraging benefits of bimodal daily activity in ProXylocopa olivieri (Lepeletier) (Hymenoptera: Anthophoridae). Environ Entomol. 34:417-424.
Hadisoesilo S. 2001. Keanekaragaman spesies lebah madu asli Indonesia. Pusat Penelitian dan Pengembangan Hutan dan Konservasi Alam, Bogor.
Hadisoesilo S, Kuntadi. 2007. Kearifan Tradisional dalam Budidaya Lebah Hutan (Apis dorsata). Departemen Kehutanan. Badan Penelitian dan Pengembangan Kehutanan. Pusat Penelitian dan Pengembangan Hutan dan Konservasi Alam. Bogor. ISBN: 978-979-3145-38-9.
Heinrich B. 1979. Bumblebee Economics. London (GB). Harvard Univ Pr.
Hepburn R, Radloff SE. 2011. Honeybees of Asia. New York (US). Springer-Verlag Berlin Heidelberg.
Itioka T, Inoue T, Kaliang H, Kato M, Nagamitsu T, Momose K, Sakai S, Yumoto T, Mohamad SU, Hamid AA, dan Yamane S. 2001. Six-year population fluctuation of giant honeybee Apis dorsata (Hymenoptera: Apidae) in a tropical lowland dipterocarp forest in Sarawak. Annals Entomol Soc America. 94:545–549.
Jasmi, Salmah S, Dahelmi S. 2014. Nesting sites of Apis cerana Fabr. (Hymenoptera: Apidae) in two different altitudes of polyculture plantations in West Sumatera. Hayati J Biosci. 21(3):135-143.
Kahono S, Nakamura K, Amir M. 1999. Seasonal migration and colony behavior of the tropical honeybee Apis dorsata F. (Hymenoptera: Apidae). Treubia. 31(3)283-297.
Koeniger N, Koeniger G. 1980. Observations and experiments on migration and dance communication of Apis dorsata in Sri Lanka. J Apicul Research. 19(1):21–34.
Lo N, Gloag RS, Anderson DL, Oldroyd BP. 2010. A molecular phylogeny of the genus Apis suggests that the giant honey bee of the Philippines, A. breviligula Maa, and the plains honey bee of Southern India, A. indica
Fabricius, are valid species. Systematic Entomology. 35:226-233.
Luca PAD, Marin MV. 2013. What‟s the „Buzz‟ about? the ecology and
evolutionary significance of buzz pollination [current opinion]. Plant Biology 16:429-435.
Makinson JC. 2013. Collective Decision-Making in Honey Bee During Nest-Site Selection. [thesis]. Australia. School of Biological Science.
Makinson JC, Schaerf TM, Rattanawannee A, Oldroyd BP, Beekman M. 2014. Consensus building in giant honey bee, Apis dorsata, swarms on the move.
Animal behavior. 93:191-199.
Mead D. 2013. A guide to some bees and wasps of Indonesia. Sulang Language Data and Working Papers: Topics in Lexicography, no. 11. Sulawesi Language Alliance. http://sulang.org/.
Momose K, Yumoto T, Nagamitsu T, Kato M, Nagamasu H, Sakai S, Harrison RD, Itioka T, Hamid AA, Inoue T. 1998. Pollination biology in a lowland dipterocarp forest in Sarawak, Malaysia. I. characteristics of the plant-pollinator community in a lowland dipterocarp forest. American J Bot. 85(10):1477–1501. ISSN 1537-2197.
24
Neupane KR, Woyke J, Poudel SM. 2013. Nesting site-preference and behavior of giant honey bee Apis dorsata. Paper presented on Apimondia, 29 September-4 October 2013 , Kyiv, Ukraine and published on abstract and working procedure Apimondia, 2013.
Apis dorsata populations: the significance of migration and colony aggregation. J Heredity. 95:119-126.
Purwatiningsih B, Leksono AS, Yanuwiadi B. 2012. Kajian komposisi serangga polinator pada tumbuhan penutup tanah di Poncokusumo Malang. Berk Penel Hayati.17:165-172.
Raffiudin R, Crozier R. H. 2007. Phylogenetic analysis of honeybee behavioral evolution. MPE. 43:543-552.
Rattanawannee A, Chanchao C. 2011. Bee diversity in Thailand and the applications of bee products, changing diversity in changing environment, PhD. Oscar Grillo (Ed.), ISBN: 978-953-307-796-3, InTech, Available from: http://www.intechopen.com/books/changing-diversity-in-changing-known world largest honeybee (Hymenoptera:Apidae). Insect Matsum. 19:47-77.
Sarwono B. 2001. Lebah Madu. Jakarta: Agromedia Pustaka.
Sadeh A, Shmida A, Keasar T. 2007. The carpenter bee Xylocopapubescens as an agricultural pollinator in greenhouse. Apidologie. 38:508-517.
Sahebzadeh N, Nurazura A, Lau WH, Mardan M, Ali MA, Tan SG. 2013. Genetic structure of malaysian Apis dorsata in Marang District (Terengganu). J Agricul Research. 52(5):202-209.
Seeley TD. 1985. Honeybee Ecology A Study of Adaptation in Social Life. Princeton University Press. Princeton, New Jersey, United Kingdom. Starr CK, Schmidt PJ, Schmidt JO. 1987. Nest site-preferences of the giant honey
Tan NQ. 2007. Biology of Apis dorsata in Vietnam. Apidologie. 38:221–229. Tingek S, Koeniger G, Koeniger N. 1996. Description of a new cavity dwelling
species of Apis (Apis nuluensis) from Sabah, Borneo with note on its occurrence and reproductive biology (Hymenoptera, Apoidea, Apini).
Senckenbergiana Biologica 76: 115-119.
Thomas SG, Varghese A, Roy P, Bradbear N, Potts SG, Davidar P. 2009. Characteristics of trees used as nest sites by Apis dorsata (Hymenoptera, Apidae) in the Nilgiri Biosphere Reserve, India. J Trop Ecol. 25:559-562 Tongco MDC. 2007. Purposive sampling as a tool for Informant selection.
Ethnobot Resear Appl. 5:147-158.
Weihmann F, Waddoup D, Hotzl T, Katzberger G. 2014. Intraspesific aggression in giant honey bees (Apis dorsata). Insects. 5:689-704.doi:10.3390/insects 5030689.
Woyke J, Wilde J, Reddy CC. 2004. Open air nesting honey bees Apis dorsata
and Apis laboriosa differ from the cavity-nesting Apis mellifera and Apis cerana in brood hygiene behaviour. J Invert Phatol. 86:1-6.
Woyke J, Wilde J, Wilde M. 2012. Swarming and migration of Apis dorsata and
Apis laboriosa honey bees In India, Nephal and Bhutan. J Apicul Science.
26
Lampiran 1 Daftar pertanyaan wawancara
Maros, 2015
Nama koresponden : Umur : Jenis kelamin : Pekerjaan : Desa/Kelurahan :
Daftar pertanyaan wawancara :
1. Pohon apa saja yang biasa digunakan sebagai pohon persarangan lebah madu hutan? Mulai bulan apa saja lebah madu hutan datang untuk bersarang?
2. Apakah ada waktu tertentu lebah madu hutan banyak/sedikit ditemukan? 3. Apa yang mempengaruhi keberadaan sarang di hutan Maros?
4. Berapa banyak sarang yang biasa ditemukan dalam satu pohon?
5. Apakah sarang yang ditemukan semakin banyak atau sebaliknya dari tahun ke tahun? Kira kira apa penyebabnya?
6. Bagaimana metode penandaan kepemilikan sarang di hutan, serta cara pengambilan madu dari sarang sampai pemasarannya ?
7. Apakah bapak menjadikan pekerjaan mencari madu sebagai pekerjaan utama?
8. Bagaimana kondisi hutan sekarang? Menurut bapak, apakah mempengaruhi keberadaan lebah madu hutan (jumlah sarang makin sedikit atau ukuran sarang makin kecil atau produksi madu semakin sedikit?
SEBARAN DAN KARAKTERISTIK PERSARANGAN LEBAH MADU HUTAN Apis dorsata binghami Cockerell
28