KEANEKARAGAMAN DAN TEMPAT BERSARANG LEBAH
TAK BERSENGAT (HYMENOPTERA: APIDAE) DI
SULAWESI TENGAH
NELKY SURIAWANTO
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis yang berjudul Keanekaragaman dan Tempat Bersarang Lebah Tak Bersengat (Hymenoptera: Apidae) di Sulawesi Tengah adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2016
RINGKASAN
NELKY SURIAWANTO. Keanekaragaman dan Tempat Bersarang Lebah Tak Bersengat (Hymenoptera: Apidae) di Sulawesi Tengah. Dibimbing oleh TRI ATMOWIDI dan SIH KAHONO.
Lebah tak bersengat termasuk dalam famili Apidae dan subfamili Apinae. Lebah ini merupakansalah satu kelompok serangga eusosial yang hidup bersama di dalam sarang. Dalam koloni lebah ini terdiri dari seekor lebah ratu, puluhan jantan, dan ribuan lebah pekerja yang dapat mengembangkan komunikasi kompleks dan mempertahankan sistem kasta. Lebah ini memiliki keanekaragaman yang bervariasi antar pulau dan tipe lingkungan, misalnya hutan dan permukiman. Pemukiman merupakan salah satu habitat lebah tak bersengat yang memiliki kekhasan lingkungan.
Salah satu tipe lingkungan sebagai habitat bersarang lebah tak bersengat adalah permukiman dan sarangnya dapat dijumpai di hampir semua bagian rumah seperti dinding rumah, rongga atap dan rongga pintu. Penelitian keanekaragaman lebah tak bersengat di lingkungan pedesaan di Sulawesi yang dikaitkan dengan tempat bersarangnya dan kepadatan sarangnya belum pernah dilaporkan. Penelitian ini bertujuan untuk mempelajari keanekaragaman, sebaran, tempat bersarang, dan kepadatan sarang lebah tak bersengat di Sulawesi Tengah.
Pulau Sulawesi yang terletak di sebelah timur garis Wallacea memiliki keunikan dalam proses terbentuknya daratan dibandingkan pulau-pulau lainnya, sehingga dikenal memiliki keunikan dan endemisitas flora dan fauna yang tinggi. Penelitian ini dilakukan di daerah permukiman desa Watumaeta, kecamatan Lore Utara, kabupaten Poso dengan ketinggian 1200 meter di atas permukaan laut (mdpl), desa Pakuli, kecamatan Gumbasa, kabupaten Sigi (130 mdpl), dan desa Purwosari, kecamtan Torue, kabupaten Parigi Moutong Sulawesi Tengah (10 mdpl). Pengamatan dilakukan pada bulan Juli 2015 – Januari 2016 menggunakan metode jelajah. Keanekaragaman lebah tak bersengat dihitung berdasarkan jumlah spesies dan sarang, sebaran sarang dipetakan menggunakan software ArcGis 10.1. Karakter pintu masuk sarang yang diamati adalah diameter, panjang, dan ketinggian dari permukaan tanah, warna pintu masuk sarang, dan teksturnya. Identifikasi spesimenlebah berdasarkan karakter morfologi dan morfometri pada lebah pekerja.
sapiens (5 sarang) seluruhnya ditemukan di rongga batu pondasi, dan T. laeviceps (1 sarang) yang ditemukan di dinding kayu.
Diameter pintu masuk terlebar adalah pada T. sapiens (2.18 cm), diikuti T. biroi (1.86 cm), dan T. fuscobalteata (1.24 cm). Pintu masuk terpanjang terjadi pada T. fuscobalteta (3.70 cm), diikuti T. biroi (2.23), dan T. sapiens (1.88 cm). Diameter dan panjang pintu masuk sarang T. laeviceps tidak teramati. Pintu masuk sarang dari permukaan tanah yang tertinggi adalah T. laeviceps (321 cm), diikuti oleh T. fuscobalteata (116.90 cm), T. biroi (56.64 cm), dan T. sapiens (1.88 cm). Karateristik pintu masuk sarang pada setiap spesies Tetragonula bervariasi. Pintu masuk sarang T. fuscobalteata berbentuk corong, berwarna coklat dan coklat terang dengan tekstur lembek. Pintu masuk sarang T. biroi berbentuk corong, berwarna coklat terang dan hitam dengan tekstur lembek, pada T. sapiens berbentuk corong, berwarna hitam, tekstur lembek, sedangkan pada T. laevieps berwarna coklat dan tekstur keras. Tempat bersarang dari lebah tak bersengat ditemukan di rongga dinding batako, rongga dinding kayu, rongga batu pondasi, rongga bambu, rongga besi, dan batang pohon.
Lebah pekerja T. fuscobalteata memiliki panjang tubuh 3.47 – 3.54 mm metasoma berwarna coklat, tergum pertama dan kedua berwarna pucat, sementara tergum keempat dan kelima gelap.
Lebah T. fuscobalteata dan T. sapiens ditemukan di dataran rendah dengan ketinggian 10 mdpl dan 130 mdpl. Lebah T. biroi ditemukan di dataran rendah dan tinggi (10 mdpl dan 1200 mdpl), dan T. laeviceps ditemukan di dataran tinggi (1200 mdpl). Empat spesies Tetragonula di Sulawesi hasil penelitian ini mempunyai morfologi yang mirip dengan spesimen spesies yang sama dari daerah lainnya. Morfometri dari empat spesies memiliki variasi ukuran tubuh dibandingkan dengan spesies yang sama yang telah dilaporkan di daerah lain.
SUMMARY
NELKY SURIAWANTO. Diversity and Nesting Sites of Stingless Bees (Hymenoptera: Apidae) in Central Sulawesi. Supervised by TRI ATMOWIDI and SIH KAHONO.
Stingless bees belonging to family Apidae and subfamily Apinae. Stingless bee is a group eusocial insect that live together in a nest. Nest of stingless bees consist of a queen, males, and workers that develop complex communication and maintain a division of caste. Diversity of stingless bees varied greatly between island and type of environments, such as forests and settlements. Settlement area is a habitat of stingless bees that has typical environment.
In the settlement areas, the nests of stingless bees can be found in almost parts of the house, such as the walls of the house, a roof cavity, and the cavity of door. The study of diversity of stingless bees in settlement areas in Sulawesi associated with nesting sites and nests density have not been reported. The aims of the study were to determine the diversity, distribution, nesting sites, and nests density of stingless bees in Central Sulawesi.
Sulawesi Island located in east of the Wallace line has unique process of formation of the land than the other islands. The island have high levels of endemicity of flora and fauna. This research was carried out in settlement areas in Watumaeta village, Lore Utara Sub-distric, Poso Regency located in 1200 meters above sea level (masl), Pakuli village, Gumbasa Sub-distric, Sigi Regency (130 masl), and Purwosari village, Torue Sub-distric, Parigi Moutong Regency (10 masl) in Central Sulawesi. Observations were carried out in July 2015 to January 2016 using cruising method. Diversity of stingless bees were analysed base on the number of species and colonies. Distribution of stingless bees nests ware mapped by ArcGIS software 10.1. Nest characters observed were diameter, length, and height from ground surface, entrance color and texture. Identification of specimen stingless bees based on morphological and morphometric characters.
A total of 170 nests of stingless bees belong to four species (Tetragonula fuscobalteata, T. biroi, T. sapiens, dan T. laeviceps) were found. In this study, T. biroi and T. sapiens are new record for Sulawesi islands. In Watumaeta village was found three nests belong to T. laeviceps (1 nest) and T. biroi (2 nests), in Pakuli village was found, 93 colonies belong to T. fuscobalteata (88 nests) and T. sapiens (5 nests), and in Purwosari village was found 74 colonies, belong to T. fuscobalteata (68 nests) and T. biroi (6 nests). The most number colonies found were T. fuscobalteata (155 nests) that found in wood walls (74 nests), stone cavity (40 nests), brick walls (30 nests), bamboos (6 nests), red brick walls (1 nest), iron cavity (4 nests), and trunk (1 nests), followed by T. biroi (7 nests) that found in the wood walls (3 nests), stone cavity (2 nests), red brick wall (1 nest), brick wall (1 nest) and trunk (1 nest), T. sapiens (5 nests) that found in stone cavity, and T. laeviceps (1 nest) that found in wood wall (1 nests).
occurred in T. laeviceps (321 cm), followed by T. fuscobalteata (116.90 cm), T. biroi (56.64 cm), and T. sapiens (1.88 cm). Characteristic of the nests entrance of each species Tetragonula varied. Nests entrance of T. fuscobalteata are funnel shape, brown and light brown, and soft texture. Nests entrance of T. biroi are funnel shape, light brown and dark, and soft texture. Nests entrance of T. sapiens are funnel shape, black, and soft texture. Nest entrance of T. laevieps are funnel shape, brown and hard texture. Nesting sites of stingless bees were found in cavity brick walls, wooden wall cavity, the cavity foundation stone, bamboo cavity, the iron cavity, and tree trunks.
The workers of T. fuscobalteata has body length 3.47 – 3.54 mm, with color of blackish brown. Species T. biroi has body length 4.00 – 4.17 mm, black color, abdomen brown pale. Species T. sapiens has body length of 3.69 - 3.80 mm, color black, metasoma brown, first and second tergum are dark brown, while tip of tergum black. Species T. laeviceps has body length 3.40 – 3.43 mm, body color black, metasoma brown, first and second tergum pale while the fourth and fifth tergum are dark.
Stingless bees, T. fuscobalteata and T. sapiens were found in the lowlands with a height of 10 (masl) and 130 (masl). Species T. biroi was found in lowland and highland (10 masl and 1200 masl), and T. laeviceps was found in the highland (1200 masl). Four species of Tetragonula found in Sulawesi have similar morphology with others specimens from other areas. Morphometry of the four species have variations in body size compared to the same species reported in other areas.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
KEANEKARAGAMAN DAN TEMPAT BERSARANG LEBAH
TAK BERSENGAT (HYMENOPTERA: APIDAE) DI
SULAWESI TENGAH
NELKY SURIAWANTO
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biosains Hewan
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Keanekaragaman dan Tempat Bersarang Lebah Tak Bersengat (Hymenoptera: Apidae) di Sulawesi Tengah
Nama : Nelky Suriawanto NIM : G352140111 Mayor : Biosains Hewan
Disetujui Oleh Komisi Pembimbing
Dr. Tri Atmowidi, M.Si Ketua
Dr. Sih Kahono Anggota
Diketahui
Ketua Program Studi Biosains Hewan
Dr. Ir. R.R. Dyah Perwitasari M.Sc
Dekan Sekolah Pascasarjana
Dr. Ir. Dahrul Syah, M.ScAgr.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karuniaNya. Sehingga karya ilmiah ini berhasil diselesaikan. Penelitian ini
berjudul “Keanekaragaman dan Tempat Bersarang Lebah Tak Bersengat (Hymenoptera: Apidae) di Sulawesi Tengah” yang dilaksanakan sejak bulan Juli 2015 sampai Januari 2016. Terima kasih penulis ucapkan kepada Dr Tri Atmowidi, M.Si dan Dr. Sih Kahono yang telah memberikan bimbingan dan arahan selama penelitian. Di samping itu terima kasih juga disampaikan kepada Dr. Clause Rasmussen dan Anne E. Dollin, Ph.D. yang telah membantu memberikan pustaka dan saran saat identifikasi, Lembaga Pengelolaan Dana Pendidikan (LPDP) yang telah membiayai penelitian ini dan Lembaga Ilmu Pengetahuan Indonesia (LIPI) yang telah membantu dalam proses identifikasi dan verifikasi spesimen. Penghargaan juga penulis sampaikan kepada Ibu Suhartini selaku laboran Fungsi dan Perilaku Hewan FMIPA IPB. Ungkapan terima kasih juga disampaikan kepada kedua orang tua atas doa, dukungan, dan perhatiannya yang tak terhingga, serta kepada keluarga, sahabat yang membantu di lapangan, dan teman-teman seperjuangan Biosains Hewan IPB 2014 dan teman-teman Asrama SULTENG atas segala doa dan dukungannya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juni 2016
DAFTAR ISI
Pengamatan Tempat Bersarang Lebah Tak Bersengat 7
Koleksi dan Mounting Spesimen Lebah 8
Identifikasi Spesimen Lebah 8
Pengamatan dan Karakter Pintu dan Tempat Bersarang 9
Analisis Data 9
HASIL DAN PEMBAHASAN 10
HASIL 10
Keanekaragaman dan Tempat Bersarang Lebah Tak bersengat 10
Karakteristik Pintu Masuk Sarang 13
Karakter Morfologi dan Morfometri Tetragonula 14
DAFTAR TABEL
1 Jumlah sarang Tetragonula yang ditemukan di lokasi penelitian 12 2 Karakteristik pintu masuk sarang pada setiap spesies 13 3 Karakter morfometri lebah pekerja T. fuscobalteata 16
4 Karakter morfometri lebah pekerja T. biroi 18
5 Karater morfometri lebah pekerja T. sapiens 20 6 Karater morfometri lebah pekerja T. laeviceps 22
DAFTAR GAMBAR
1 Bagan rumusan masalah 2
2 Sketsa struktur sarang lebah tak bersengat 5
3 Sketsa morfometri beberapa bagian karakter tubuh lebah tak bersengat 6
4 Peta lokasi penelitian 7
5 Koleksi dan mounting lebah tak bersengat 8
6 Pintu masuk sarang lebah tak bersengat 9
7 Morfologi lebah tak bersengat 10
8 Peta distribusi lebah tak bersengat di desa Watumaeta 11 9 Peta distribusi lebah tak bersengat di desa Pakuli 11 10 Peta distribusi lebah tak bersengat di desa Purwosari 12
11 Tempat bersarang 4 spesies lebah tak bersengat 13
12 Karateristik pintu masuk sarang lebah tak bersengat 14
13 Karakteristik morfologi T. fuscobalteata 15
14 Karakteristik morfologi T. biroi 17
15 Karakteristik morfologi T. sapiens 19
PENDAHULUAN
Latar Belakang
Lebah tak bersengat berperan penting dalam membantu proses penyerbukan tanaman (Inoue et al. 1985), penghasil madu, dan propolis (Lourino et al. 2006; Kumar et al. 2012). Walaupun lebah tak bersengat dapat menghasilkan bahan penting bagi manusia dan hidup bersama di sekitar perumahan, namun banyak yang tidak mengenalnya.
Indonesia memiliki banyak spesies lebah tak bersengat yang tersebar luas hampir di seluruh pulau. Keanekaragaman lebah tak bersengat bervariasi di ekosistem hutan dan permukiman (Boontop et al. 2008; Salim et al. 2012; Kelly et al. 2014; Syafrizal et al. 2014; Rahman et al. 2015). Di permukiman, sarang lebah tak bersengatdapat dijumpai di hampir semua bagian rumah, seperti dinding rumah, rongga atap, dan rongga pintu (Erniwati 2013).
Sulawesi merupakan pulau dengan tingkat endemisitas yang tinggi. Pulau Sulawesi terbentuk dari proses geologi dan ekologi yang panjang, membentuk bentang pulau yang unik dengan keanekaragaman flora dan fauna yang tinggi. Spesies dan subspesies lebah madu endemik yang ditemukan di Sulawesi, yaitu Apis nigrocincta dan A. dorsata binghami (Hadisoesilo 2001; Engel 2012). Spesies lebah tak bersengat yang endemik di Sulawesi adalah Geniotrigona incisa yang ditemukan di hutan Sulawesi pada ketinggian 900 meter di atas permukaan laut (mdpl) (Sakagami dan Inoue 1989). Dengan eksplorasi dan kegiatan identifikasi lebah tak bersengat yang lebih banyak masih dimungkinkan ditemukan jenis baru atau catatan baru di Sulawesi.
Sulawesi Tengah merupakan salah satu provinsi di pulau Sulawesi, yang memiliki banyak lahan perkebunan, pertanian, hutan sekunder, hutan primer, dan permukiman. Daerah ini terdapat tanaman tropis dari berbagai jenis yang dapat mendukung kehidupan lebah tak bersengat. Informasi dan publikasi tentang keberadaan lebah tak bersengat di Sulawesi Tengah masih sangat kurang, disebabkan oleh sedikitnya penelitian yang dilakukan. Masyarakat di Sulawesi Tengah pada umumnya kurang menyukai ternak lebah tak bersengat daripada ternak lebah Apis, karena lebah Apis pada umumnya dapat menghasilkan lebih banyak madu. Namun data terkini yang diperoleh dari beberapa daerah di Sulawesi dan Indonesia secara umum menunjukkan bahwa minat beternak lebah tak bersengat semakin meningkat karena lebih menguntungkan dibandingkan dengan ternak lebah Apis.
Perumusan Masalah
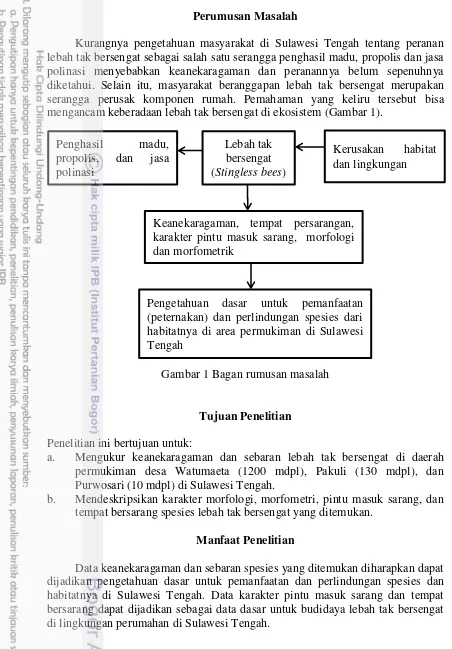
Kurangnya pengetahuan masyarakat di Sulawesi Tengah tentang peranan lebah tak bersengat sebagai salah satu serangga penghasil madu, propolis dan jasa polinasi menyebabkan keanekaragaman dan peranannya belum sepenuhnya diketahui. Selain itu, masyarakat beranggapan lebah tak bersengat merupakan serangga perusak komponen rumah. Pemahaman yang keliru tersebut bisa mengancam keberadaan lebah tak bersengat di ekosistem (Gambar 1).
Gambar 1 Bagan rumusan masalah
Tujuan Penelitian
Penelitian ini bertujuan untuk:
a. Mengukur keanekaragaman dan sebaran lebah tak bersengat di daerah permukiman desa Watumaeta (1200 mdpl), Pakuli (130 mdpl), dan Purwosari (10 mdpl) di Sulawesi Tengah.
b. Mendeskripsikan karakter morfologi, morfometri, pintu masuk sarang, dan tempat bersarang spesies lebah tak bersengat yang ditemukan.
Manfaat Penelitian
Data keanekaragaman dan sebaran spesies yang ditemukan diharapkan dapat dijadikan pengetahuan dasar untuk pemanfaatan dan perlindungan spesies dan habitatnya di Sulawesi Tengah. Data karakter pintu masuk sarang dan tempat bersarang dapat dijadikan sebagai data dasar untuk budidaya lebah tak bersengat di lingkungan perumahan di Sulawesi Tengah.
TINJAUAN PUSTAKA
Biologi Lebah Tak Bersengat
Lebah tak bersengat termasuk dalam famili Apidae dan subfamili Apinae. Lebah tak bersengat merupakan salah satu serangga eusosial yang hidup bersama di dalam sarang. Sarang lebah tak bersengat terdiri dari seekor lebah ratu, puluhan lebah jantan, dan ribuan lebah pekerja yang mengembangkan komunikasi kompleks dan mempertahankan sistem kasta (Michener 2007). Ratu dan pejantan adalah lebah produktif, sedangkan lebah pekerja adalah lebah betina steril. Lebah ratu berperan penting dalam mengatur sistem kerja di dalam sarang (Roopa et al. 2015), menghasilkan anakan baru, pejantan, dan pekerja. Lebah pekerja memiliki fungsi mengumpulkan bahan makanan berupa polen dan nektar, serta resin sebagai bahan untuk membangun sarang (Michener 2013).
Di Indonesia, lebah tak bersengat memiliki nama yang berbeda-beda, antara lain klanceng (Jawa), teuwel (Sunda), galo-galo (Minang) (Syafrizal et al. 2014), dan Tannese (Kaili-Sulawesi Tengah). Indonesia memiliki beberapa spesies lebah tak bersengat yaitu dari genus Tetragonula (T. iridipennis, T. laeviceps, T. fuscobalteata, T. minagkabau, T.canifrons, T. clypearis, T. drescheri, T. melina, T. reepeni, dan T. sapiens), Tetrigona (T. apicalis dan T. vidua), Geniotrigona (G. Secara umum pekerja dari lebah tak bersengat memiliki ukuran tubuh 2 - 8 mm (Erniwati 2013). Tubuh terdiri dari kepala, thorak, abdomen, sepasang sayap dengan 3 pasang tungkai dan warna tubuh bervariasi antar spesies.
Distribusi Lebah Tak Bersengat
ditemukan genus Austroplebeia, Platytrigona, dan Tetragonula. Kawasan Indonesia bagian tengah merupakan wilayah distribusi campuran antara Indo-Malayan dan Australian, terdistribusi genus Geniotrigona dan Tetragonula (Rasmussen dan Cameron 2007; Rasmussen 2008).
Di daerah tropis, lebah ini dilaporkan 8 spesies di India (Rasmussen 2013), 19 spesies di Siam, satu spesies di Burma termasuk Tenasserim, 26 spesies di Malaysia, 5 spesies di kepulauan Fillipina, satu spesies di Taiwan (Formosa), 32 spesies di Thailand (Schwarz 1939; Sakagami et al. 1985; Michener dan Boongird 2004; dan Klakasikorn et al. 2005), dan 12 spesies di Australia (Dollin et al. 1997; Dollin et al. 2015).
Peranan Lebah Tak Bersengat
Lebah tak bersengat merupakan salah satu lebah penghasil madu dan propolis (Michener 2007). Di beberapa negara, peternakan lebah tak bersengat untuk tujuan komersil telah lama dilakukan, yaitu untuk mendapatkan madu, propolis, dan membantu proses penyerbukan tanaman (Erniwati 2013). Tubuh lebah tak bersengat memiliki rambut-rambut yang bercabang yang dapat menempelkan serbuk sari pada saat lebah berkunjung di bunga. Selain rambut ditubuhnya, terdapat tempat penyimpanan khusus serbuk sari dibagian tungkai belakang yang disebut corbicula (pollen basket).
Di Indonesia, lebah tak bersengat telah dilaporkan oleh beberapa peneliti. Spesies T. laeviceps dapat meningkatkan dalam produksi biji kailan (Brassica oleraceae var. Alboglabra) sebesar 231% jumlah polong per tanaman, 48% jumlah biji per polong, 204% bobot biji per tanaman, 24% perkecambahan biji, dan 80.8% viabilitas polen (Wulandari et al. 2015). Selain sebagai penyerbuk tanaman pertanian yang memiliki nilai ekonomi tinggi, lebah tak bersengat berperan dalam penyerbukan tumbuhan di hutan primer dan hutan skunder sehingga membantu regenerasi hutan tersebut (Erniwati 2013). Kahono et al. (2012) melaporkan T. laeviceps dan T. melina mempunyai potensi tinggi sebagai penyerbuk kelapa sawit pada bagian permukaaan bunga. Di Australia, lebah tak bersengat merupakan lebah penyerbuk yang paling efisien pada tanaman Mangifera indica, karena banyaknya serbuk sari yang menempel di tubuhnya setelah mengunjugi bunga (Anderson et al. 1982).
Struktur Sarang Lebah Tak Bersengat
Gambar 2 Sketsa struktur sarang lebah tak bersengat: pintu masuk sarang (a), pollen cells (b), saluran masuk bagian dalam (c), brood cells (d), honey cells (e), dan lapisan-lapisa batument (f) (Michener 2013).
Lokasi Persarangan
Sarang lebah tak bersengat umumnya dapat ditemukan di hutan dan area permukiman (Erniwati 2013). Sarang lebah tak bersengat sebagian besar ditemukan di daerah terbuka yang memiliki suhu udara yang cukup tinggi dan memudahkan untuk mencapai sumber makanan (Nagamitsu dan Inoue, 1998; Eltz 2001; Michener 2007). Di hutan, sarang lebah tak bersengat dapat ditemukan antara lain di pohon Ficus religiosa, Diospyros rhodocalyx, Irvingia malayana, Dipterocarpus alatus, Homalium grandiflorum, Hevea brasiliensis, Tectona grandis, dan Eusideroxylon zwageri (Klakasikorn et al. 2005; Syafrizal et al. 2014). Di permukiman penduduk sarang lebah tak bersengat dapat ditemukan di rongga atap, rongga pintu, dan ronggajendela(Erniwati 2013).
Pada saat di dalam sarang lebah tak bersengat terdapat calon ratu baru, beberapa lebah pekerja akan pergi mencari tempat baru dengan membawa bahan dari sarang lama. Karateristik bentuk pintu masuk sarang selalu dibangun pertama kali. Setelah sarang baru jadi, ratu muda akan terbang menuju sarang baru dengan membawa sebagian lebah jantan dan pekerjanya. Untuk beberapa waktu (minggu atau bahkan bulan), pekerja terus terbang bolak-balik membawa bahan dari sarang lama ke sarang baru, sampai akhirnya kontak tersebut berhenti dan sarang baru menjadi independen (Michener 2013).
Morfometri
Morfometri adalah suatu metode untuk mengidentifikasi spesies dengan mendeskripsikan melalui pengukuran, penghitungan atau pemberian nilai (skor). Morfometri dapat diaplikasikan untuk mengetahui kekerabatan suatu spesies, diferensiasi dari berbagai spesies, dan variasi spesies (Bookstein dan Strauss 1982).
Dalam morfometri, deskripsi bisa berupa deskripsi kualitatif atau kuantitatif. Deskripsi kualitatif, misalnya bentuk tubuh lebih kecil dibandingkan dengan
(a) (b)
spesies lainnya. Deskriptif kualitatif dianggap belum memadai, sehingga diperlukan ekspresi kuantitatif dengan mengambil berbagai ukuran dari individu-individu dan dinyatakan dengan nilai statistik seperti rata-rata, kisaran, ragam dan korelasi (Bookstein dan Strauss 1982). Strauss dan Bond (1990) menyatakan studi morfometri secara kuantitatif dapat membedakan individu antar jenis kelamin atau spesiesnya, menggambarkan keragaman pola-pola morfometri antar populasi atau spesies dan dapat mengklarifikasi hubungan filogenetik.
Lebah tak bersengat merupakan serangga yang memiliki ukuran tubuh kecil (2 – 8 mm). Dalam proses identifikasi, morfologi tubuh dan warna sangat sulit untuk menentukan ke level spesies, meskipun dapat dilakukan pada level genus. Namun, untuk genus yang berukuran sangat kecil, seperti Tetragonula, diperlukan pengukuran karakter tubuh, seperti panjang tubuh, panjang sayap depan dari tegula, lebar maximum kepala, jarak antara percabangan M-Cu dan ujung dasar marginall cell, dan jarak antara dasar atas sampai ujung bawah hind tibia (Sakagami 1978; Sung et al. 2004) (Gambar 3).
BL
(a) (b)
METODE
Waktu dan Lokasi Penelitian
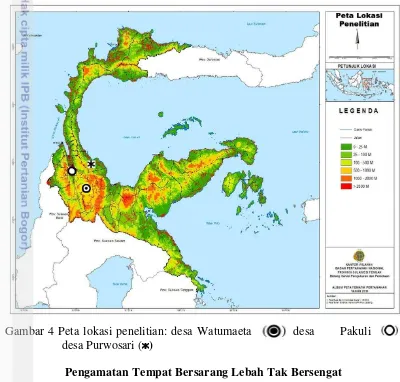
Penelitian dilaksanakan pada bulan Juli 2015 – Januari 2016. Pengambilan sampel dilakukan pada bulan Juli – September 2015 di tiga desa yaitu, desa Watumaeta, Kecamatan Lore Utara, Kabupaten Poso (1.200 mdpl), desa Pakuli Kecamatan Gumbasa, Kabupaten Sigi (130 mdpl), dan desa Purwosari, Kecamatan Torue, Kabupaten Parigi Moutong (10 mdpl) (Gambar 4). Luas lokasi permukiman di desa Watumaeta, Pakuli, dan Purwosari berturut-turut adalah 20.000 ha, 502 ha, dan 94 ha.
Gambar 4 Peta lokasi penelitian: desa Watumaeta desa Pakuli desa Purwosari ( )
Pengamatan Tempat Bersarang Lebah Tak Bersengat
Koleksi dan Mounting Spesimen Lebah
Lebah tak bersengat dikoleksi dari pintu masuk sarang (Gambar 5a) menggunakan jaring serangga dari setiap sarang yang ditemukan. Sepuluh individu lebah pekerja lebah tak bersengat (Gambar 5b) dari setiap sarang dimasukan ke dalam botol berisikan ethanol 90% (Gambar 5c), 7 individu untuk keperluan identifikasi dan 3 individu untuk keperluan morfometri.
Sebelum diidentifikasi, spesimen lebah dilakukan proses mounting (Gambar 5d) agar karakter-karakter yang penting pada tubuh lebah mudah diamati. Proses mounting dilakukan degan cara menusukkan jarum serangga di bagian toraks dan merapikan semua bagian tubuhnya. Selanjutnya, spesimen lebah dioven pada suhu 35oC selama satu minggu dan setelah itu spesimen dimasukan dalam freezer pada suhu -15oC selama satu minggu.
(a) (b)
(c) (d)
Gambar 5 Koleksi dan mounting lebah tak bersengat: koleksi lebah di pintu masuk sarang (a), lebah pekerja (b), spesimen dalam botol sampel dengan ethanol 90% (c), dan spesimen yang telah di mounting (d).
Identifikasi Spesimen Lebah
Spesimen lebah diidentifikasi berdasarkan karakter morfologi dan verifikasi menggunakan spesimen di Museum Zoologicum Bogoriense (MZB), Bidang Zoologi, Pusat Penelitian Biologi, Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong. Identifikasi spesimen berdasarkan karakter morfometri dilakukan di Laboratorium Biosistematika dan Ekologi Hewan, Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor (IPB) Dramaga.
Sakagami et al. (1990), dan Smith (2012). Karakter morfometrik diamati menggunakan Mikroskop stereo model Nikon C-LEDS yang dilengkapi dengan kamera, software Optilab viewer, dan Image Raster. Karakter morfometri yang diukur adalah panjang tubuh (BL), lebar kepala (LK), panjang dan lebar mata (PM dan LM), jarak maximum dan minimum interorbital (JMI dan LOD), jarak interocellar atas (JIA), jarak interocellar bawah (IOD), jarak ocellocular (JO), lebar gena (LG), panjang malar (PML), panjang dan lebar flagellomere IV (PF dan LF), panjang sayap depan dari tegula (WL1), panjang jarak antara venasi M-Cu (WL2), panjang tibia belakang (PTB), lebar tibia belakang (LTB), lebar dan panjang basitarsus (LBB dan PBB) (Sakagami 1978). Selain itu, pengukuran dilakukan untuk panjang kepala (PK), panjang clypeus (PC), jarak terpanjang dan terdekat interocular (JIB dan JIA), jarak interantennal (JI), antennocellar (JA), antennocular (JO) (Michener 2007), panjang dan lebar mandibula (PM dan LM), panjang dan lebar mesoscutum (PMS dan LMS) (Rasmussen 2013), panjang dan lebar sayap depan (PSD dan LSD), panjang dan lebar sayap belakang (PSB dan LSB), jumlah hamuli (JH), panjang femur (PJB), lebar dan panjang basitarsus (LBB dan PBB) (Klakasikorn et al. 2005) (Lampiran 1).
Pengamatan Karakteristik Pintu dan Tempat Bersarang
Karakteristik pintu masuk sarang dideskripsi secara kualitatif dan kuantitatif. Karakter kuantitatif yang diukur yaitu ketinggian dari permukaan tanah, panjang dan diameter pintu masuk sarang (Gambar 6). Karakter kualitatif pintu masuk sarang yang diamati mengikuti Kelly et al. (2014), yaitu warna (coklat, coklat terang, atau hitam), dan tekstur (lembek atau keras). Tempat bersarang lebah tak bersengat pada bagian rumah, juga diamati jumlah sarang dari setiap spesies yang ditemukan dimasing-masing lokasi juga dihitung.
Gambar 6 Pintu masuk sarang lebah tak bersengat: ketinggian dari permukaan tanah (T), panjang pintu masuk (P), dan diameter pintu masuk sarang (D).
Analisis Data
Keanekaragaman spesies lebah tak bersengat disetiap desa pengamatan ditampilkan dalam tabel. Sebaran sarang ditampilkan dalam peta distribusi spasial menggunakan menggunakan software ArcGis 10.1. Karakter pintu masuk sarang dan tempat bersarang dideskripsikan. Karakter morfologi dan morfometri dideskripsikan.
D P
HASIL
Keanekaragaman dan Tempat Bersarang Lebah Tak Bersengat
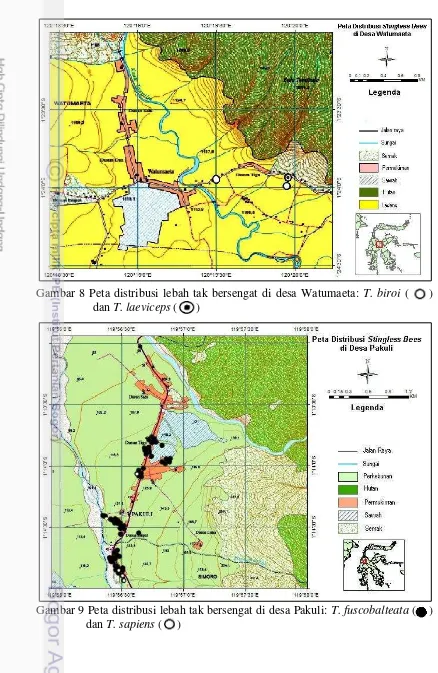
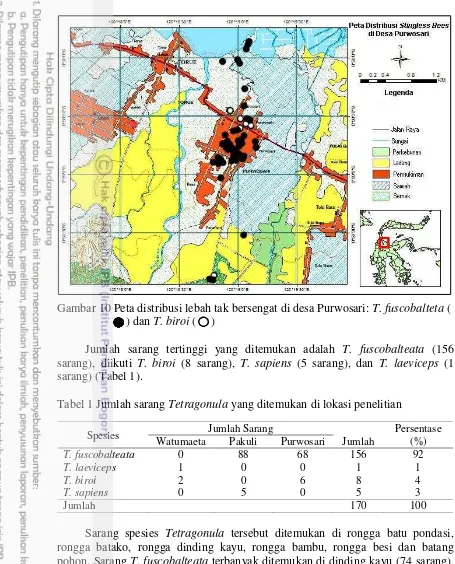
Total 170 sarang ditemukan dipenelitian ini yang terdiri dari 4 spesies, yaitu Tetragonula fuscobalteata (Cameron, 1908), T. biroi (Friese, 1898), T. sapiens (Cockerell, 1911), dan T. laeviceps (Smith, 1857) (Gambar 7). Di desa Watumaeta ditemukan 2 spesies, yaitu T. laeviceps (1 sarang) dan T. biroi (2 sarang). Di desa Pakuli ditemukan 2 spesies, yaitu T. fuscobalteata (88 sarang) dan T. sapiens (5 sarang), sedangkan di desa Purwosari ditemukan T. fuscobalteata (68 sarang) dan T. biroi (6 sarang) (Gambar 8, 9, dan 10). Titik kordinat tempat ditemukan sarang lebah tak bersengat terdapat di lampiran 2.
(a) (b)
(c) (d)
Gambar 8 Peta distribusi lebah tak bersengat di desa Watumaeta: T. biroi ( ) dan T. laeviceps ( )
Gambar 10 Peta distribusi lebah tak bersengat di desa Purwosari: T. fuscobalteta ( ) dan T. biroi ( )
Jumlah sarang tertinggi yang ditemukan adalah T. fuscobalteata (156 sarang), diikuti T. biroi (8 sarang), T. sapiens (5 sarang), dan T. laeviceps (1 sarang) (Tabel 1).
Tabel 1 Jumlah sarang Tetragonula yang ditemukan di lokasi penelitian
Sarang spesies Tetragonula tersebut ditemukan di rongga batu pondasi, rongga batako, rongga dinding kayu, rongga bambu, rongga besi dan batang pohon. Sarang T. fuscobalteata terbanyak ditemukan di dinding kayu (74 sarang), diikuti oleh rongga batu pondasi (40 sarang), dinding batako (31 sarang), bambu (6 sarang), rongga besi (4 sarang), dan batang pohon (1 sarang). Sarang T. biroi ditemukan di dinding kayu (3 sarang), rongga batu pondasi (2 sarang), dinding batako (2 sarang), dan batang pohon (1 sarang). Sarang T. sapiens ditemukan di rongga batu pondasi (5 sarang). Tempat bersarang T. laeviceps ditemukan di dinding kayu (1 sarang) (Gambar 11).
Spesies Jumlah Sarang Persentase
(%) Watumaeta Pakuli Purwosari Jumlah
T. fuscobalteata 0 88 68 156 92
T. laeviceps 1 0 0 1 1
T. biroi 2 0 6 8 4
T. sapiens 0 5 0 5 3
0 batako (DB), rongga batu pondasi (BP), rongga dinding kayu (DK), rongga bambu (RB), rongga besi (BS), dan batang pohon (BP).
Karakteristik Pintu Masuk Sarang dengan ketinggian dari permukaan tanah yaitu 321 cm (Tabel 2).
Tabel 2. Karakteristik pintu masuk sarang pada setiap spesies
Spesies
Keterangan : Rata-rata (minimum – maximum) (-) tidak diukur
bentuk tidak dapat diamati karena telah patah dan berwarna coklat dengan tekstur keras (Gambar 12i).
(a) (b) (c)
(d) (e) (f)
(g) (h) (i)
Gambar 12 Karakteristik pintu masuk sarang lebah tak bersengat: T. fuscobalteata (a-c), T. biroi (d-f), T. sapiens (g-h), dan T. laeviceps (i).
Karakter Morfologi dan Morfometri Tetragonula
Tetragonula fuscobalteata
Tubuh pekerja T. fuscobalteata berwarna coklat kehitaman, mesoscutum terdiri dari 6 rambut longitudinal dan masing-masing dipisahkan oleh 5 glabrous yang mencolok, clypeus pucat, anterior tungkai belakang terdapat rambut bagian pinggir berwarna abu-abu keperakan dan warna abdomen pucat serta tergit gelap kecoklatan (Gambar 13).
(a) (b)
(c) (d)
Gambar 13 Karakter morfologi T. fuscobalteata: lebah pekerja (a), mesoscutum terdiri dari 6 rambut longitudinal (b), clypeus pucat (c), dan abdomen terdiri dari tergit hitam kecoklatan (d).
Tetragonula biroi
Lebah pekerja T. biroi berwarna hitam, abdomen berwarna coklat kehitaman, clypeus dan tegula gelap, mesoscutum dan mesoscutellum terdapat rambut yang berdiri berwarna hitam, malar space tereduksi tetapi jelas memisahkan mandibula dengan mata majemuk (Gambar 14).
(a) (b)
(c) (d)
Gambar 14 Karakter morfologi T. biroi: lebah pekerja berwarna hitam (a), abdomen kecoklatan (b), mesoscutum dan mesoscutellum terdiri dari rambut hitam yang berdiri (c), dan malar space tereduksi (d).
Tetragonula sapiens
Tubuh pekerja T. sapiens berwarna hitam, metasoma coklat, tergum pertama dan kedua coklat gelap sedangkan tergum ujung hitam, mesoscutum terdiri dari glabrous hampir bisa dilihat, clypeus berwarna hitam, rambut bagian pinggir anterior tungkai belakang gelap sampai hitam kecoklatan sedangkan bagian posterior kuning kecoklatan (Gambar 15).
(a) (b)
(c) (d)
Gambar 15 Karakter morfologi T. sapiens: lebah pekerja berwarna hitam (a), abdomen coklat gelap (b), mesoscutum terdiri dari glabrous hampir bisa dilihat (c), clypeus coklat gelap (d).
Tetragonula laeviceps
Tubuh lebah pekerja T. laeviceps berwarna hitam, metasoma berwarna coklat, tergum pertama dan kedua berwarna pucat sementara tergum keempat dan kelima gelap, permukaan atas dari pedicel kehitaman, segmen antena (flagella) kehitaman, mesoscutum bagian atasterdapat rambut hitam dan tidak ada globarus, rambut anterior tugkai belakang kehitaman (Gambar 16).
(a) (b)
(c) (d)
Gambar 16 Karakter morfologi T. laeviceps: lebah pekerja berwarna hitam (a), abdomen hitam kecoklatan (b), mesoscutum bagian atas terdiri dari rambut hitam (c), dan segmenantena(flagella) kehitaman (d).
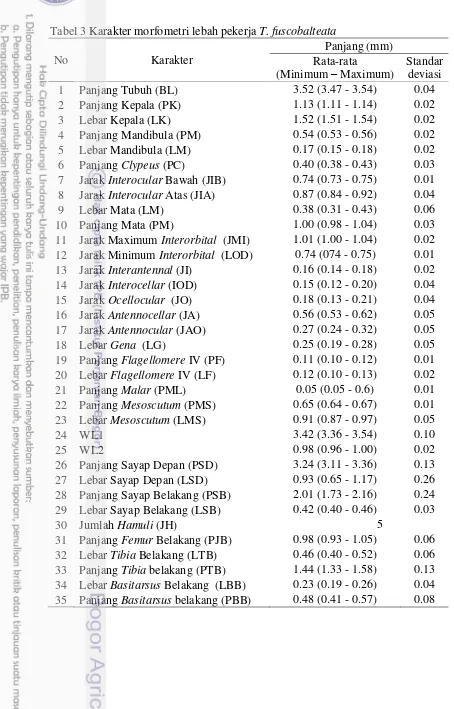
Morfometri lebah pekerja T. laeviceps adalah: panjang tubuh 3.40 - 3.43 mm, panjang sayap depan dari tegula (WL1)3.60 - 3.76 mm, panjang jarak antara venasi M-Cu (WL2)1.10 - 1.16 mm, panjang tungkai belakang 1.35 - 1.41 mm, panjang malar lebih pendek daripada lebar segmen antena IV (0.07 + 0.01 dan 0.13 + 0.00 mm), panjang sayap depan 3.54 - 3.62 mm, lebar sayap depan 1.04 - 1.31 mm, warna stigma sayap depan coklat tua, panjang hind basitarsus 0.57 - 0.61 mm, dan lebar hind basitarsus 0.24 - 0.31 mm (Tabel 6).
PEMBAHASAN
Keanekaragaman dan Tempat Bersarang
Empat spesies lebah tak bersengat yang ditemukan pada penelitian ini termasuk dalam genus Tetragonula, yaitu T. fuscobalteata, T. biroi, T. sapiens, dan T. laeviceps. Empat spesies ini di kumpulkan dari 3 lokasi yang memiliki ketinggian berbeda-beda, yaitu desa Purwosari (10 mdpl), Pakuli (130 mdpl), dan Watumaeta (1200 mdpl). Spesies T. fuscobalteata ditemukan di ketinggian 10 mdpl dan 130 mdpl. Lebah T. biroi ditemukan di ketinggian 10 mdpl dan 1200 mdpl. Lebah T. sapiens hanya ditemukan diketinggian 130 mdpl dan T. laeviceps di ketinggian 1200 mdpl. Salim et al. (2012) melaporkan T. fuscobalteata dapat ditemukan di ketinggian 100-600 mdpl di hutan lindung di Perak, Malaysia dan T. laeviceps 200 – 1128 mdpl. Lebah T. biroi dan T. sapiens belum banyak dilaporkan.
Dua spesies lebah tak bersengat yaitu T. fuscobalteata dan T. laeviceps yang ditemukan di penelitian ini sebelumnya telah dilaporkan di Sulawesi dan 2 spesies lainnya yaitu T. biroi dan T. sapien belum pernah dilaporkan. Dua spesies, yaitu Lebah T. fuscobalteata dan T. laeviceps memiliki distribusi yang luas. T. fuscobalteata dengan penyebaran di Asia Tenggara (Thailand, Kamboja, Semenanjung Malaysia, Borneo, Sumatra, Sulawesi, Pulau Palau, Pulau Caroline, Philippines-Palawan dan Pulau Oceanic) sampai kawasan Australia. Spesies T. laeviceps tersebar dari India, Sri Lanka, Asia Tenggara, dan Australia (Sakagami 1978; Sakagami et al. 1990; Rasmussen 2008; Smith 2012; Erniwati 2013). Lebah T. sapiens terdistribusi dari Filipina, Indonesia-Maluku, Papua Nugini dan Australia (Dollin et al. 1997; Rasmussen 2008; Smith 2012). Lebah T. biroi terdistribusi di Filipina–Pulau Oceanic dan Papua Nugini (Rasmussen 2008; Smith 2012). Selama ini belum dilaporkan spesies T. sapiens dan T. biroi ditemukan di Sulawesi, sehingga 2 spesies tersebut merupakan catatan baru (new record) dari Pulau Sulawesi.
Sarang empat spesies Tetragonula tersebut ditemukan di rongga dinding batako, rongga dinding kayu, rongga pondasi, rongga bambu, rongga besi, dan batang pohon yang merupakan area terbuka, dengan sinar matahari yang cukup. Michener (2007) dan Syafrizal (2014) melaporkan lokasi yang terbuka dengan suhu yang tinggi membuat lebah tidak membutuhkan energi yang banyak untuk memanaskan suhu tubuh untuk aktivitas mencari pakan. Starr dan Sakagami (1987) melaporkan di Filipina, sarang T. fuscobalteta dan T. sapiens ditemukan di batang bambu. Sakagami et al. (1983) melaporkan T. laeviceps bersarang pada hampir semua bagian rumah. Chinh et al. (2005) dan Kelly et al. (2014) melaporkan sarang T. laeviceps dapat ditemukan di batang pohon hidup atau mati dengan diameter 30 – 50 cm. Dalam penelitian ini T. biroi banyak dijumpai di lokasi permukiman, dan selama ini belum ada publikasi tempat bersarangnya.
Karakteristik Pintu Masuk Sarang
publikasi Starr dan Sakagami (1987) di Fillipina, bahwa T. fuscobalteata yang bersarang di bambu memiliki pintu masuk berbentuk corong. Selama ini belum ada publikasi tentang pintu masuk sarangn T. biroi. Pada T. laevicep,pintu masuk sarang tidak teramati dengan jelas karena telah patah. Kelly et al. (2014) melaporkan pintu masuk sarang T. laeviceps berbentuk corong. Roubik (2006) melaporkan bentuk, warna, dan kekakuan pintu masuk sarang lebah tak bersengat dipengaruhi faktor internal (umur sarang dan genetik lebah) dan eksternal (intensitas cahaya, resin tumbuhan, predator dan simbiosis).
Morfologi dan Morfometri Tetragonula
Karakter morfologi T. fuscobalteta yang dideskripsikan dalam penelitian ini sesuai dengan deskripsi Schwarz (1939), Sakagami (1978), Sakagami et al. (1990), dan Smith (2012). Namun, tubuh T. fuscobalteata hasil penelitian ini(3.47 - 3.54 mm) lebih panjang dari yang dideskripsikan oleh Sakagami (1978) dengan sampel dari Asia dan Sri Lanka (2.8 – 3.2 mm). Perbedaan ini juga pernah dilaporkan oleh Schwarz (1939), bahwa T. fuscobalteata dari Sulawesi dan kepulauan Filipina mempunyai tubuh lebih panjang dari spesimen-spesimen yang pernah dilaporkan dari Malaysia dan Borneo.
Morfologi lebah pekerja T. biroi mirip dengan T. iridipennis yaitu memiliki malar space yang tereduksi namun masih bisa dilihat jarak antara mata majemuk dan mandibula. Rambut-rambut hitam di mesoscutum dan mesoscutellum merupakan karakter khusus pada T. biroi, seperti yang telah dideskripsikan oleh Schwarz (1939), Sakagami (1978), dan Vijayakumar et al. (2014). Karakter morfometri T. biroi yang dilaporkan oleh Schwarz (1939) dan Smith (2012) adalah panjang sayap depan dari tegula yaitu 4.2 dan 4.4 mm, yang tidak berbeda jauh dengan spesimen dalam penelitian ini (4.09 - 4.20 mm).
Morfologi lebah pekerja T. sapiens yang dideskripsikan dalam penelitian ini sesuai dengan deskripsi Dollin et al. (1997). Panjang tubuh T. sapiens dipenelitian ini (3.69 - 3.80 mm) sesuai dengan yang dilaporkan oleh Dollin et al. (1997) (3.6 - 4.2 mm) yang merupakan spesimen dari Papua Nugini. Panjang sayap depan dari tegula T. sapiens dipenelitian ini (4.16 - 4.19 mm) tidak telalu berbeda dengan spesimen dari Papua Nugini (4.2–4.5 mm), Maluku (3.5–4.5 mm), dan Pulau Solomon 4.2–4.5 mm.
SIMPULAN
DAFTAR PUSTAKA
Anderson DL, Sedgley M, Short JRT, Allwood AJ. 1982. Insect pollination of mango in Northern Australia. Aust J AgricRes. 33(1):41-48.
Bookhout TA. 1996. Research And Management Techniques For Wildlife And Habitats. Kansas (US): Allen Press Inc.
Bookstein FL, Strauss RE. 1982. The truss: body form reconstruction in morphometrics. Systematic Zoology. 3: 113-115.
Boongird S. 2011. Aspects of culturing, reproductive behavior, and colony formation in the stingless bees Tetragonula fuscobalteata (Hymenoptera: Apidae: Meliponini). J Kans Entomol Soc. 84(3): 190–196.
Boontop Y, Malaipan S, Chareansom K, Wiwatwittaya D. 2008. Diversity of stingless bees (Apidae: Meliponini) in Thong Pha Phum district, Kanchanaburi province, Thailand. Kas J Nat Sc. 42: 444-456.
Chinh TX, Sommeijer MJ, Boot WJ, Michener CD. 2005. Nest and colony characteristics of three stingless bee species in Vietnam with the first description of the nest of Lisotrigona carpenter (Hymenoptera: Apidae: Meliponini). J Kans Entomol Soc. 78(4): 363–372.
Dollin AE, Dollin LJ, Rasmussen C. 2015. Australian and New Guinean stingless bees of the genus Austroplebeia moure (Hymenoptera: Apidae) a revision. Zootaxa. 4047 (1): 1–73.doi:10.11646/zootaxa.4047.1.1
Dollin AE, Dollin LJ, Sakagami SF. 1997. Australian stingless bees of the genus Trigona (Hymenoptera: Apidae). Invertebrate Taxonomy. 11: 861–896. Eltz T. 2001. Ecology of stingless bee (Apidae: Meliponini) in lowland
dipterocarp forest in Sabah, Malaysia, and an evaluation of logging impact on populations and communities. [dissertasi]. Munchen (GMN): Universitaet Wuerzburg.
Engel MS. 2012. The honey bees of Indonesia (Hymenoptera: Apidae). Treubia. 39:1-85.
Erniwati. 2013. Kajian biologi lebah tak bersengat (Apidae: Trigona) di Indonesia. MZI. 12(1): 29-34.
Free JB. 1982. Bees and Mankind. London (GB): George Allen & Unwin.
Gonzalez VH, Engel MS. 2012. A new species of Geotrigona Moure from the Caribbean coast of Colombia (Hymenoptera: Apidae). Zookeys. 172: 77-87.doi:10.3897/zookeys.172.2735.
Hadisoesilo S. 2001. Keanekaragaman spesies lebah madu asli Indonesia pusat penelitian dan pengembangan hutan dan konservasi alam, Bogor. Biodiversitas. 2: 123-128.
Inoue T, Salamah S, Abbas I, Yusuf E. 1985. Foraging behavior of individual workers and foraging dynamics of colonies of three sumatran stingless bees. Res Popul Ecol. 27: 373-392.
Kelly N, Farisya MSN, Kumara TK, Marcela P. 2014. Species diversity and external nest characteristics of stingless bees in meliponiculture. PerJ Trop Agric Sc. 37 (3): 293 – 298.
Klakasikorn A, Wongsiri S, Deowanish S, Duangphakdee O. 2005. New record of stingless bees (Meliponini: Trigona) in Thailand. Nat Hist J Chulalongk Univ. 5(1): 1-7.
Kumar MS, Singh AJAR, Alagumuthu G. 2012. Traditional beeskeeping of stingless bees (Trigona sp.) by kani tribes of Western Ghats, Tamil Nadu, India. Indian J Tradit Knowledge. 11(12): 342-345.
Lourino MC, Fonseca VLI, Roubik DW, Dollin A, Heard T, Aguilar IB, Venturieri GC, Eardley C, Neto PN. 2006. Global meliponiculture: challenges and opportunities. Apidologie. 37: 1-18.doi: 10.1051/apido:2006027.
Michener CD. 2007. The Beess of The World. Second Edition. Baltimore (US): The Johns Hopkins Univ. Pr.
Michener CD. 2013. Pot-Honey: A Legacy of Lebah tak bersengat. Vit P, Pedro SRM, Roubik DW. Editor. New York (US): Springer Pr. 3-17.doi: bersengat (Hymenoptera: Apidae, meliponinae) assosiated with floral shape and location in an Asian tropical rainforest. Entomological Science. 1: 189-194.
Rahman A, Das PK, Rajkumari P, Saikia J, Sharmah D. 2015. Stingless bees (Hymenoptera: Apidae: Meliponini) diversity and distribution in India. Inter J Sc Res. 4(1): 77-81.
Rasmussen C. 2008. Catalog of the Indo-Malayan/Australasian stingless bees (Hymenoptera: Apidae: Meliponini). Zootaxa. 1935:1–80.
Rasmussen C. 2013. Stingless bees (Hymenoptera: Apidae: Meliponini) of the Indian subcontinent: diversity, taxonomy and current status of knowledge. Zootaxa 3647(3): 401–428.doi: 10.11646/zootaxa.3647.3.1.
Rasmussen C, Cameron SA. 2007. A molecular phylogeny of the old world Lebah tak bersengat (Hymenoptera: Apidae: Meliponini) and the non-monophyly of the large genus Trigona. Systematic Entomology. 32:26-39.doi: 10.1111/j.1365-3113.2006.00362.x.
Rasmussen C, Michener CD. 2010. The identity and neotype of Trigona laeviceps Smith (Hymenoptera: Apidae). J Kans Entomol Soc. 83(2): 129–133.doi: 10.2317/JKES0907.08.1.
Roopa AN, Eswarrapa G, Sanganna M, Sajjanar, Gowda G. 2015. Study on nesting characteristics and biology of stingless bees (Trigona iridipennis Smith.). IOSR-JAVS. 8(10): 34-36.
Roubik DW. 2006. Stingless bee nesting biology. Apidologie. 37: 124-143.doi: 10.1051/apido:2006026.
Sakagami SF, Inoue T. 1989. Stingless bees of the genus Trigona (subgen: Geniotrigona) (Hymenoptera: Apidae) with description of T. (G.) incisa sp, nov. from Sulawesi. Jpn J Entomol. 57(3): 605-620.
Sakagami SF, Inoue T. 1987. Stingless bees of the genus Trigona (subgenus Trigonella) with notes on the reduction of spatha in male genitalia of the subgenus Tetragonula (Hymenoptera: Apidae). Kontyu. 55(4): 610-627. Sakagami SF, Inoue T, Yamane S, Salmah S. 1983. Nest architecture and colony
composition of the Sumatran stingless bees Trigona (Tetragonula) laeviceps. Kontyû. 51(1):100-111.
Sakagami SF, Inoue T, Salmah S. 1990. Stingless bees of Central Sumatra. Sakagami SF, Ohgushi R, Roubik DW. Editor. Sapporo (JP): Hokkaido Univ. Pr. 125-137.
Sakagami SF, Salmah S, Abbas I, Yusuf I. 1985. Foraging behavior of individual workers and foraging dynamics of colonies of three Sumatran stingless bees. Res Popul Ecol. 27: 373-392.
Salim HMW, Dzulkiply AD, Harrison RD, Fletcher C, Kassim AR, Potts MD. 2012. Stingless bee (Hymenoptera: Apidae: Meliponini) diversity in dipterocarp forest reserves in Peninsular Malaysia. Raffl Bull Zool. 60(1): 213–219
Schwarz HF. 1939. The Indo-Malayan species of Trigona. Bull AMNH. 76: 83– 141.
Smith DR. 2012. Key to workers of Indo-Malayan stingless bees. For use in the Stingless BeesWorkshop. 1(1): 1-42.
Sung IH, Yamane S, Ho KK, Wu WJ, Chen YW. 2004. Morphological caste and sex differences in the Taiwanese stingless bee Trigona ventralis hoozana (Hymenoptera: Apidae). Entomological Science. 7: 263-269.
Syafrizal, Tarigan D, Yusuf R. 2014. Biodiversity and habitat of Trigona at secondary tropical rain forest of Lempake education forest, Samarinda, Kalimantan Timur. JTP. 9(1): 34-38.
Vijayakumar K, Jeyaraaj R. 2014. Taxonomic notes on stingless bee Trigona (Tetragonula) iridipennis Smith (Hymenoptera: Apidae) from India. J Threat Taxa. 6(11):6480-6484.
Lampiran 1. Sketsa ukuran morfometri lebah tak bersengat
1 2
4
3
5
6
7 8
Lanjutan lampiran 1
9 10
11 12
13 14
15 16
Lanjutan lampiran 1
17 18
PF LF
19 20
21 22
23 24
Lanjutan lampiran 1
25 26
27 28
29 30
31 32 33
Lanjutan lampiran 1
34 35
Lampiran 2. Titik kordinat sarang yang ditemukan di setiap lokasi penelitian
Spesies Titik Kordinat Sarang
Desa Watumaeta
Riwayat Hidup
Peneliti dilahirkan di desa Purwosari, kecamatan Toure, kabupaten Parigi Moutong, Provinsi Sulawesi Tengah pada tanggal 15 November 1991 dari Pasangan Bapak Suwito, S.Pd dan Ibu Nurmini. Penulis merupakan anak pertama dari tiga bersaudara.
Penulis lulus dari SMA 1 Torue pada tahun 2009. Pendidikan sarjana ditempuh penulis di Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Tadulako (UNTAD) dan lulus pada tahun 2013. Pada tahun 2014 penulis melanjutkan studi strata 2 (S2) di Program Studi Biosains Hewan (BSH), Departemen Biologi pada program Pascasarjana Institut Pertanian Bogor (IPB). Penulis mendapatkan program Beasiswa Tesis dari Lembaga Pengelolan Dana Pendidikan (LPDP).
Publikasi ilmiah dengan judul “Diversity and Nesting Sites of Stigless Bees