IMUNOGLOBULIN Y ANTITETANUS
DIISOLASI DARI TELUR AYAM SEBAGAI
PENGGANTI ANTITETANUS
SERUM KUDA
I NYOMAN SUARTHA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Karakteristik Imunoglo bulin Y Antitetanus Diisolasi Dari Telur Ayam Sebagai Pengganti Antite tanus Serum Kuda adalah karya saya sendiri dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutif dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka dibagian akhir disertasi ini.
I NYOMAN SUARTHA. Karakteristik Imunoglobulin Y Antitetanus Diisolasi dari Telur Ayam sebagai Pengganti Antitetanus Serum Kuda. Dibawah bimbingan I WAYAN TEGUH WIBAWAN, RETNO DAMAYANTI SOEJOEDONO, dan BIBIANA W. LAY.
Produksi antibodi poliklonal pada ayam lebih menguntungkan dibandingkan dengan produksi pada mamalia. Pemeliharaan ayam lebih sederhana dan murah, ekstraksi dan purifikasi imunoglobulin Y (IgY) dari telur lebih sederhana, cepat dan biayanya murah. Penggunaan ayam juga dapat mengurangi pemakaian hewan mamalia dan menjadikan kondisi hewan coba lebih baik. Selain itu IgY di dalam telur memberikan prospek yang sangat berarti, untuk mengatasi masalah produksi antitetanus serum pada kuda. Telur dikoleksi dari ayam betina jenis Isa Brown yang telah diimunisasi dengan toksoid tetanus dosis bertingkat. Toksoid tetanus dicampur dengan Freund adjuvant complete dan Freund adjuvant incomplete. Ekstraksi IgY antitetanus dari kuning telur dilakukan dengan metode PEG–Kloroform selanjutnya dipurifikasi dengan fast protein liquid chromatography. Identifikasi IgY antitetanus ditentukan secara fotometris dengan metode Bradford (λ = 595 nM), uji Agar Gel Presipitation. Berat molekul IgY antitetanus dideteksi dengan Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis. IgY antitetanus diber ikan perlakuan pH, panas , dan enzim. Aktivitas biologis IgY antitetanus setelah perlakuan diamati dengan metode
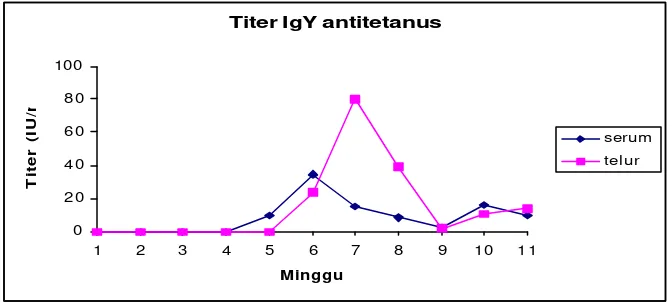
enzyme linked imunosorbent assay. Uji Potensi IgY anti tetanus ditentukan dengan metode Spearman-Karber. Titer IgY antitetanus mencapai punca k pada telur minggu ke tujuh dari awal imunisasi. Rataa n titer tertinggi pada telur 80.16 ± 33.55 IU/ml dan terendah 1.69 ± 0.63 IU/ml. Konsentrasi protein (IgY) setelah ekstraksi diperoleh sebesar 0.652 ± 0.041 mg/ml. Sedangkan setelah purifikasi konsentrasi protein sebesar 1.644 ± 0.424 mg/ml. Aktivitas biologis IgY anti tetanus menurun sangat nyata (p<0.01) pada pH 2 dan pH 3, suhu 72.5 oC, dan hilang pada perlakuan suhu 90 oC. Larutan sukrosa konsentrasi 25% mampu mempertahankan aktivitas IgY antitetanus sampai 50 % pada suhu 75 oC selama 5 menit, sedangkan larutan glukosa konsentrasi 50% mampu meningkatkan aktivitas IgY antitetanus pada suhu 75 oC dan suhu 80 oC selama 5 menit. Aktivitas IgY antitetanus menurun setelah perlakuan enzim pepsin, tripsin dan protease. Berdasarkan perhitungan Spearman-Karber diperoleh nilai potensi IgY antitetanus sebesar 35 IU/ml. Pemberian IgY antitetanus 0.2 dan 0.4 IU/ekor mampu melindungi mencit dari dosis letal toksin tetanus . Pada pemberian dosis IgY lebih tinggi (0.8 IU/ekor dan 1 IU/ekor) mampu melindungi mencit dari peningkatan dua kali dosis letal toksin. Pemberian IgY antitetanus secara oral pada hewan dewasa tidak efektif untuk tujuan imunoterapi pasif dalam usaha pencegahan penyakit secara sistemik . Kesimpulan dari penelitian ini yaitu ayam mampu memproduksi IgY antitetanus pada kuning telur dan dapat digunakan sebagai sumber antitetanus serum untuk menggantikan produksi antitetanus serum di kuda.
I NYOMAN SUARTHA. The Characteristic of Antitetanus Immunoglobulin Y Isolated from Eggs to substitute Horse Antitetanus Serum. Under advisory of I WAYAN TEGUH WIBAWAN, RETNO DAMAYANTI SOEJOEDONO, and BIBIANA W. LAY.
Laying hens are highly profitable producers of polyclonal antibodies in comparison with mammals as antibodies can be purified from egg yolks. The cost for producing IgY is lower than for mammalian antibodies since chicken housing is cheap and isolation process is economical, high yielding, uncomplicated and fast. The purpose of the study was to explore the opportunity of using antitetanus IgY from egg yolks to substitute the production of antitetanus serum from horses. The eggs were collected from adult Isa brown hens which have been immunized by tetanus toxoid. The immunization was applied intra venously with an initial dose of 15 Lf. The immunization was repeated three times with gradual dose of 100, 200, and 300 Lf with an interval of one week intra muscularly. The first immunization was tetanus toxoid mixed with Freund adjuvant complete and subsequently mixed with Freund adjuvant incomplete. Antitetanus IgY was extracted from egg yolks by means of PEG–Chloroform and purifie d using fast protein liquid chromatography. The purity of antitetanus IgY was determined by Bradford method (λ = 595 nm) and Agar Gel Precipitation test. The molecular weight of purified antitetanus IgY was determined with Sodium Dodecyl Sulphate Polyacrylamide Gel Electrophoresis. The antitetanus IgY were treated by pH, heat, and enzyme. The biological activities of treated antitetanus IgY was determined by enzyme linked immunosorbent assay. Neutralization potency test of antitetanus IgY was determined by Spearman-Karber method. The antitetanus IgY highest titer of egg yolks was 80.16 ± 33.55 IU/ml reached at seven weeks after starting immunization schedule and the lowest was 1.69 ± 0.63 IU/ml. Protein concentration (IgY) after extraction and purification were 0.652 ± 0.041 mg/ml and 1.644 ± 0.424 mg/ml respectively . Biological activity of antitetanus IgY decreased significantly (p<0.01) at of pH 2, pH 3, and at 72.5 oC, and lost its activity at 90 oC after 20 minute s. Addition of 25% sucrose solution was able to maintain antitetanus IgY activity until 50 % at 75 oC for 5 minutes, while addition of 50% glucose solution increase antitetanus IgY activity at 75 oC and 80 oC for 5 minutes. The effect of pepsin, trypsin, and protease enzyme s decreased the activities of antitetanus IgY. Spearman-Karber value of potency of antitetanus IgY was 35 IU/ml. The injection of 0.2 and 0.4 IU antitetanus IgY per mice respectively, protect all mice from the letal dose tetanus toxin. The doses of 0.8 IU and 1 IU anti tetanus IgY per mice protected twice dose of letal tetanus toxin. Antitetanus IgY given orally on adult animal was ineffective as passive immunotherapy. This research concluded that the hens were capable of produc ing antitetanus in egg yolks and can be used as a resource of antitetanus serum to substitute horse antitetanus serum.
Hak cipta milik I nstitut Pertanian Bogor , tahun 2006 Hak cipta dilindungiDilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apa pun, baik cetak, fotokopi,
IMUNOGLOBULIN Y ANTITETANUS DIISOLASI DARI
TELUR AYAM SEBAGAI PENGGANTI ANTITETANUS
SERUM KUDA
I NYOMAN SUARTHA
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Sains Veteiner
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
ANTITETANUS DIISOLASI DARI TELUR AYAM SEBAGAI PENGGANTI ANTITETANUS SERUM KUDA
Nama Mahasiswa : I NYOMAN SUARTHA Nomor pokok : B161030021
Program Studi : SAINS VETERINER
Disetujui Komisi Pembimbing
Dr. drh. I Wayan Teguh Wibawan, MS. Ketua
Dr. Retno Damayanti Soejoedono, MS. Prof. Dr. Bibiana W. Lay, M.Sc. Anggota Anggota
Diketahui
Ketua Program Studi Sains Veteriner Dekan Sekolah Pascasarjana
drh. Bambang P. Priosoeryanto, MS, PhD. Dr.Ir.Khairil Anwar Notodiputro,MS.
Puji syukur penulis panjatkan kehadapan Ida Sang Hyang Widi Wasa, Tuhan Yang Maha Esa atas segala rahmatNya, sehingga penulisa n disertasi ini dapat diselesaikan. Karya ilmiah ini berjudul : Karakteristik Imunoglobulin Y Antitetanus Diisolasi Dari Telur Ayam Sebagai Pengganti Antitetanus Serum Kuda.
Ucapan terimakasih dan penghargaan penulis sampaikan kepada Yang terhormat: Dr. drh. I Wayan Teguh Wibawan MS selaku ketua komisi pembimbing, Dr. Retno Damayanti Soejoedono, MS, dan Prof. Dr. Bibiana W. Lay, MSc. Sebagai anggota komisi pembimbing yang telah memberikan semangat, bimbingan, saran, petunjuk dan tuntunan yang tulus selama mengikuti pendidikan, persiapan dan perencanaan penelitian, pencarian dana penelitian sampai pada penyusunan disertasi ini.
Terima kasih penulis sampaikan kepada DIRJEN DIKTI atas beasiswa BPPS yang telah diberikan, Direktur P3M DIKTI atas sebagian dana penelitian yang diberikan melalui proyek Hibah bersaing XII/tahun ke-1 dan ke-2, Dr. drh Risa Tiuria, MS atas masukannya saat ujian tertutup. Prof. Dr. Fachriyan H. Pasaribu, dan Dr. drh. A.E.T.H. Wahyuni, MSi atas masukannya saat sidang terbuka untuk memperluas kasanah tulisan ini. Ibu drh Lia Siti Halimah, MSi dan staf di laboratorium uji hewan PT Bio Farma Bandung atas segala bantuan yang telah diberikan, Dr Ita Djuwitha, MPhil dan staf di laboratorium Embriologi FKH IPB atas bantuannya. Ucapan terima kasih juga penulis sampaikan kepada drh Okti Nadia putri, drh Putu Ika Mayangsari, drh Candramaya Siska Damayanti, Dra Elisabeth Maria, MSi, saudara Imam Bayu serta tema n–teman Punhawacana Bali atas bantuannya.
Kepada ayahanda I Wayan Budiasa, BA, Ibunda Ni Ketut Sangi, AMAPd, Ayah mertua I Nyoman Padet dan Ibunda Ni Wayan Rambu (Alm) , Kakak, Adik serta seluruh keluarga, penulis mengucapkan terima kasih dan penghargaan atas segala doa, bimbingan dan dorongan semangat yang telah diberikan selama ini. Terima kasih, penghargaan dan hormat yang tulus penulis sampaikan kepada istri tercinta drh Ni Made Sri Widiyani, yang penuh rasa kasih dan pengertian, sabar, selalu mendoakan, memberikan dorongan, semangat, dan banyak berkorban selama penulis mengikuti pendidikan sampai selesainya disertasi ini. Terima kasih pula untuk anak-anak tercinta dan tersayang Ni Luh Vigyan Witharni, Ni Made Praba Viswandari, dan I Nyoman Gede Prajnam Magha Vadantha yang senantiasa memberikan semangat dan dorongan dalam kehidupan penulis.
Penulis dengan rendah hati mohon maaf atas segala kekurangan dalam disertasi ini dan semoga karya ini bermanfaat.
Bogor, September 2006
Penulis dilahirkan di Badung Bali pada tanggal 1 Maret 1968, sebagai anak kedua dari empat saudara dari pasangan ayah I Wayan Budiasa BA, dan Ibu Ni Ketut Sangi, AMAPd. Pendidikan sarjana dan profesi dokter hewan di tempuh di Fakultas Kedokteran Hewan Universitas Udayana, lulus tahun 1993. Pada tahun 1997 penulis mendapat kesempatan mengikuti pendidikan program Magister di Program Studi Sains Veteriner Program Pascasarjana Institut Pertanian Bogor dan menamatkannya pada tahun 1999. Kesempatan untuk melanjutkan ke program doktor pada program studi dan pada perguruan tinggi yang sama di peroleh pada tahun 2003. Beasiswa pendidikan pascasarjana diperoleh dari Direktorat Jenderal Pendidikan Tinggi, Departemen Pendidikan Nasional Republik Indonesia melalui Biaya Pendidikan Program Pascasarjana (BPPS).
Latar Belakang
Kejadian penyakit tetanus dilaporkan masih tetap tinggi, setiap tahun dilaporkan terjadi 350 000 sampai 400 000 kasus tetanus di seluruh dunia. Kejadian kasus lebih tinggi di negara berkembang terutama negara yang program imunisasinya tidak teratur dan tingkat kesadaran masyarakat tentang kesehatan rendah (Bruggemann et al. 2003; Kiefer 2004). Pada negara industri dilaporkan kasus tetanus terjadi secara sporadik. Penyakit tetanus dilaporkan bersifat endemis pada 90 negara berkembang (Bruggemann et al. 2003).
Strategi pencegahan penyakit tetanus dengan meningkatkan kebersihan dan imunisasi. Imunisasi dapat dilakukan secara aktif dengan toksoid tetanus dan pasif dengan antitetanus serum. Pada daerah yang beresiko tinggi, imunisasi aktif minimal dilakukan sebanyak tiga kali (Vandelaer et al. 2003). Antibodi yang terbentuk akan berikatan dan menginaktivasi toksin sebelum toksin itu menyerang otot dan saraf. Clostridium tetani dapat dibunuh dengan antibiotik penicilin, tetapi antibiotik tidak mampu menetralisasi toksin (Kiefer 2004). Pencegahan dengan antitetanus serum dan pengobatan dengan antibiotik merupakan pendekatan yang masih relevan untuk mengurangi penyakit tetanus (Covarelli dan Marconi 1980; Bleck 1991; Forrat et al. 1998; Bruggemann et al. 2003).
Pencegahan tetanus difokuskan pada imunisasi awal. Imunisasi diulang setiap 10 tahun atau pada saat berumur 40 sampai 50 tahun untuk menghindari terinfeksi tetanus saat umur tua (Schatz et al. 1998; Cavuslu et al. 2003). Maral et al. (2001) melaporkan kejadian tetanus menyerang lebih dari satu juta orang di seluruh dunia, dengan 80% kejadian terjadi pada masa neonatal.
Kekhawatiran akan kejadian tetanus sampai saat ini pada manusia masih mendapat perhatian yang serius, khususnya aplikasi antitetanus ser um (ATS) tetap dilakukan pada saat terjadi perlukaan. Hal yang sama juga terjadi pada kesehatan hewan, terutama pada kawasan kebun binatang atau kawasan wisata dengan obyek binatang. Penggunaan kandang yang sempit, daya dukung kawasan yang terbatas akan mempermudah kejadian luka, seperti halnya pada daerah kawasan wisata monkey forest Ubud Bali yang dihuni oleh monyet ekor panjang, kejadian luka berkisar sampai 10%. Tipe luka yang banyak diderita adalah kulit robek, luka tusuk, dan kulit robek sampai otot rusak. Kondisi luka ini akan mempermudah kejadian tetanus terutama luka yang terjadi di daerah badan yang sulit dibersihkan. Kesembuhan luka yang diderita oleh monyet itu berkisar antara satu minggu sampai 3 minggu (Suartha et al. 2002). Waktu itu merupakan periode masa inkubasi dari kuman tetanus (Lewis 1998). Hal yang sama juga sangat rentan dialami oleh praktisi kesehatan hewan di lapangan, terutama yang bertugas di wilayah terpencil. Seorang praktisi kesehatan hewan dilaporkan tewas akibat tetanus sete lah terinjak sapi, begitu juga halnya di daerah bekas bencana (Soeharsono 2005).
Produksi ATS saat ini umumnya dilakukan pada kuda, yakni dengan menyuntikkan toksoid tetanus pada kuda yang terpilih. Masalah sering muncul karena penggunaan serum spesifik sering menyebabkan reaksi silang seperti reaksi anafilaktik atau serum sickness. Hal itu telah mendorong penggunaan imunoglobulin antitetanus dari sumber lain seperti serum manusia, tetapi kesulitan menyedia kan donor (Forrat et al. 1998). Penggunaan telur (Imunoglobulin Y) diharapkan dapat mengurangi resiko itu dan digunakan sebagai sumber antitetanus .
menggunakan imunoglobulin dari mamalia seperti kelinci, tikus, quinea pig, dan hewan mamalia besar seperti kuda, kambing, domba, dan sapi (Svendsen et al.
1995).
Beberapa peneliti melaporkan keuntungan penggunaan telur sebagai pabrik bahan biologi adalah : kandungan IgY tinggi dalam telur, mudah diproduksi dalam jumlah besar, tidak menimbulkan efek samping karena tidak bereaksi silang dengan faktor rheumatoid dan komponen jaringan mamalia, tidak bereaksi dengan reseptor Fc yang dimiliki oleh mikrob sehingga bereaksi lebih spesifik terhadap antigen yang dikehendaki, memiliki aktivitas dan daya netralisasi lebih tinggi dibandingkan IgG mamalia, biaya produksi secara masal lebih murah dibandingkan menggunakan mamalia, dan dari segi Animal welfare
lebih dapat diterima dibandingkan dengan menggunakan mamalia (Davis dan Reeves 2002).
Pemanfaatan IgY untuk pengobatan dan pencegahan penyakit masih sedikit dan terbatas pada skala laboratorium. Perkembangan dan penelitian ke arah pemanfaatan telur unggas baru berkembang dalam satu setengah dekade terakhir. Antibodi spesifik di dalam kuning telur dapat ditimbulkan dari berbagai macam patogen. Kermani-Arab e t al. (2001) melaporkan IgY spesifik terhadap penyakit Marek yang diaplikasi secara pasif mampu menahan infeksi virus Marek. Efek yang sama diamati terhadap berbagai penyakit misalnya influenza (Bogoyavlensky et al. 1999), EPEC K11 (Rawendra 2005) , Salmonella enteridis
dan typhimurium (Lee et al. 2002; Babu et al. 2003) , Helicobacter pylori (Shin
et al. 2002; Shin et al. 2004).
IgY juga digunakan untuk deteksi antigen pe rmukaan penderita hepatitis B (Makvandi dan Fiuzi 2002), caries gigi (Hamada et al.1991), pembentukan
plaque gigi karena Streptokokus mutans (Hatta et al. 1997), diare pada pedet (Erhard et al. 1997), deteksi kanker (Yang et al. 1997; Fortgens et al. 1997; Sasse et al. 1998; Sriram et al. 1999), Virus horsesickness (Plessis et al. 1999) , Bovine coronavirus (Ikemori et al. 1997) , dan deteksi insulin (Song et al. 1985).
Teknologi IgY sangat baik digunakan untuk produksi antibodi dari antigen
al.1997; Losonczy et al.1999; Coillie et al. 2004; Ester 2004), dan diagnostik yang lain (Katz et al.1985). Pemberian secara oral sebagai food suplemen untuk pencegahan kuman (Shimizu et al. 1988; Yokohama et al. 1998; Carlander et al.
2000; Hedlund dan Hau 2001, Sunwoo et al. 2002). IgY dapat di absor psi dan ditransfer secara efisien sebagai antibodi kolustrum (Yokohama et al. 1993).
Adanya IgY dalam telur memberikan prospek untuk pemberian kekebalan pasif pada kasus penya kit (Polson et al. 1980). Prinsip pengebalan adalah pasif, artinya transfer kekebalan terhadap beberapa penyakit dilakukan dengan mengkonsumsi telur yang mengandung zat kebal dan dipreparasi secara khusus. IgY unggas mengenal lebih banyak epitop protein mamalia dibandingkan dengan imunoglobulin kelinci, sehingga cocok untuk percobaan imunologi protein mamalia (Schade et al.1996). IgY unggas dapat diproduksi apabila antigen dalam jumlah sedikit atau memerlukan pengawetan tinggi seperti hormon. Dalam pemerik saan imunologi, IgY memberikan hasil lebih akurat karena dapat mengurangi ikatan dengan antigen non spesifik, yang menyebabkan hasil negatif palsu atau positif palsu (Schade et al. 1996; Warr dan Higgins 1995).
Laporan penelitian di atas memberikan inspirasi untuk mempelajari karakteristik dan efikasi atau daya netralisasi IgY spesifik terhadap toksin tetanus. Sampai saat ini keperluan akan antitetanus serum (ATS) untuk pencegahan infeksi tetanus, khususnya pada kasus perlukaan masih sangat dibutuhkan.
Pe rumusan masalah
Hal ini menyebabkan penderitaan kronis pada kuda. Endapan amiloid sering dijumpai pada organ limpa, limfoglandula dan organ limfoid lainnya. Produksi ATS pada kuda sangat mahal.
Adanya masalah-masalah di atas maka terpikirkan untuk memproduksi ATS pada telur ayam. Hal ini sangat mungkin dilakukan karena antibodi dalam darah induk ayam dapat ditransfer ke dalam telur dalam jumlah yang cukup banyak. Schade et al. (1996) melaporkan imunisasi pada ayam menghasilkan konsentrasi antibodi spesifik yang sama antara serum da n kuning telur. Konsentrasi IgY pada kuning telur konstan sampai oosit matang (maturasi), dengan kandungan 10 sampai 20 mg/ml (Carlander 2002). Biaya produksi imunoglobulin pada telur unggas sangat murah (Warr dan Higgins 1995; Makvandi da n Fiuzi 2002)
Penggunaan ayam untuk produksi antibodi menghilangkan dan mengurangi penggunaan mamalia sebagai hewan laboratorium. Menghilangkan yang di maksud adalah menghilangkan langkah yang menyakitkan saat koleksi darah, yang digantikan dengan ekstraksi antibodi dari kuning telur. Pengurangan yang di maksud adalah mengurangi jumlah hewan yang digunakan, sebab ayam menghasilkan antibodi yang lebih efisien dibandingkan hewan mamalia seperti kelinci maupun mamalia besar lainnya (Karlsson et al. 2004). Berkenaan dengan
animal welfare dan efisiensi biaya, penggunaan antibodi dalam telur lebih bisa diterima dibandingkan dengan penggunakan hewan percobaan mamalia (Svendsen
et al. 1995).
Tujuan Penelitian
Tujuan penelitian ini adalah mengetahui sifat karakteristik IgY spesifik terhadap toksin tetanus yang diisolasi dari telur, untuk menggantikan produksi antitetanus serum pada kuda.
Manfaat Penelitian
produksinya lebih murah, aman, mudah, dan kualitas yang lebih baik daripada ATS konvensional yang diproduksi dari kuda.
Hipotesis
Berdasarkan atas latar belakang dan permasalahan yang diajukan di atas, maka disusun suatu hipotesis sebagai berikut :
1 Ayam mampu memproduksi IgY spesifik terhadap toksin tetanus yang tersimpan pada telur,
2 IgY yang terbentuk mempunyai daya tahan terhadap pengaruh suhu, pH, dan enzim pencernaan,
viii Halaman
DAFTAR TABEL ... x
DAFTAR GAMBAR ... xi
DAFTAR LAMPIRAN ... xiii
PENDAHULUAN ... 1
Latar Belakang ... 1
Perumusan masalah ... 4
Tujuan Penelitian ... 5
Manfaat Penelitian ... 5
Hipotesis ... 6
TINJAUAN PUSTAKA ... 7
Sistem Imun ... 7
Imunoglobulin ... 9
Imunoglobulin Y ... 10
Keuntungan Penggunaan Imunoglobulin Y ... 14
Penggunaan Teknologi IgY ... 17
Stabilitas IgY ... 18
Tetanus ... 19
Toksin Tetanus ... 22
Mekanisme Kerja Toksin Tetanus ... 25
Imunitas Terhadap Toksin Tetanus ... 34
Pencegahan dengan Antitoksin ... 35
MATERI DAN METODE ... 36
Lokasi dan Waktu Penelitian ... 36
Materi ... 36
Hewan Percobaan ... 36
Toksoid Tetanus ... 36
Bahan ... 37
Peralatan ... 37
Metode ... 38
Produksi IgY pada Telur Ayam ... 38
Ekstraksi IgY dari Kuning Telur ... 38
Purifikasi IgY ... 39
Identifikasi Kemurnian IgY ... 39
ix
Sodium Dodecyl Sulphate Polyacrylamide Gel
Electrophoresis ... 41
Teknik ELISA ... 42
Aktivitas Biologis IgY... ... 43
Uji Aktivitas IgY Setelah Perlakuan pH ... ... 43
Uji Aktivitas IgY Setelah Perlakuan Suhu ... 43
Uji Aktivitas IgY Setelah Perlakuan Enzim Pepsin, Tripsin, dan Protease ... 44
Uji Potensi IgY ... 44
Uji Tantang pada Hewan Coba ... 45
HASIL DAN PEMBAHASAN ... 47
Produksi IgY antitetanus pada Telur Ayam ... 47
Ekstraksi, Purifikasi dan Karakterisasi IgY Antitetanus dari Kuning Telur ... 52
Aktivitas Biologis IgY antitetanus ... 59
Titrasi Toksoid, Sampel IgY Ayam, dan Konjugate Enzim .... 59
Aktivitas IgY Antitetanus Setelah Perlakuan pH ... 59
Aktivitas IgY Antitetanus Setelah Perlakuan Suhu ... 62
Aktivitas IgY Antitetanus Setelah Perlakuan Enzim Pepsin, Tripsin, dan Protease ... 67
Hasil SDS-PAGE IgY Antitetanus Setelah Perlakuan Fisik dan Kimia ... 69
Potensi IgY Antitetanus ... 70
Uji Tantang Pada Hewan Coba ... 72
Berat Badan ... 75
Feed Intake ... 78
Laju Respirasi ... 80
Refleksitas ... 82
Pemberian IgY Antitetanus Secara Oral ... 83
x Halaman
1. Karakter imunoglobulin itik ... 12
2. Kelebihan IgY dibandingkan dengan IgG mamalia ... 16
3. Hasil uji AGP IgY antitetanus pada serum dan telur ayam ... 47
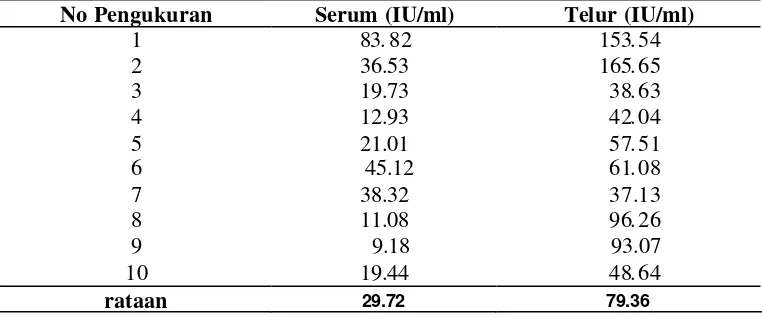
4. Hasil pengukuran titer IgY antitetanus (IU/ml) ... 51
5. Konsentrasi protein hasil ekstraksi dan purifikasi IgY antitetanus ... 54
6. Uji potensi IgY antitetanus ... 71
7. Protective dose-50 ATS standar ……… 71
8. Protective dose-50 ATS uji ……… 71
xi Halaman
1 Perbedaan struktur IgY dan IgG. ... 12
2 Fragmen Hc toksin tetanus ... 25
3 Proses perlekatan toksin bakteri pada sel ... 27
4 Interaksi ganggliosida pada dua permukaan binding site fragmen Hc... 28
5 Patogenesis penyakit tetanus ... 31
6 Proses penghambatan toksin tetanus terhadap reseptor transmiter inhibitor ... 32
7. Rataan konsentrasi IgY antitetanus pada serum dan telur ... 48
8. Hasil uji imunodifusi IgY antitetanus Ayam ... 48
9. Penambahan kloroform pada laruta n kuning telur. B. Pemisahan supernatan yang mengandung IgY antitetanus dari lemak telur yang telah mengendap ... 52 10 . A. Penambahan PEG 6000 pada supernatan. B. Pemisahan pelet dari supernatan setelah disentrifuse ... 53
11 . Profil pita protein dari IgY antitetanus hasil ekstraksi dengan metode PEG – kloroform ... 55
12 Kromatogram hasil FPLC IgY antitetanus ... 56
13 Hasil uji agar gel presipitasi IgY antitetanus setelah pemurnian dengan FPLC. ... ... 57
14 Hasil SDS-PAGE IgY antitetanus setelah pemurnian FPLC. ... 58
15 Pola aktifitas IgY antitetanus setelah perlakuan pH ... 60
16 Penurunan aktivitas IgY antitetanus setelah perlakuan pH ... 61
17 Pola aktifitas IgY antitetanus setelah perlakuan suhu ... 62
18 Penurunan aktivitas IgY antitetanus setelah perlakuan suhu dengan waktu inkubasi berbeda ... 63
19 Penambahan larutan sukrosa pada larutan IgY antitetanus ... 64
20 Penambahan larutan glukosa ke dalam larutan IgY antitetanus ... 65
21 Pola aktifitas IgY antitetanus setelah perlakuan enzim pepsin, tripsin, dan protease ... 68
22 Hasil SDS-PAGE IgY antitetanus setelah perlakua n fisik dan kimia . . 69
23 Frekuensi pemunculan kasus dengan gejala klinis khas tetanus ... 73
24 Jumlah gejala klinis yang muncul harian. ... 74
25 Gejala klinis yang muncul pada dosis IgY berbeda. ... 74
xii
28 Pola kecenderungan garis regresi feed intake mencit ……… 79
29 Pola kecenderungan garis regresi laju respirasi mencit ... 80
30 Konsumsi oksigen mencit ... 81
xiii Halaman 1. Reagensia untuk Fast Protein Liquid Chromatography (FPLC) ... 99 2. Reagensia untuk ELISA ... 100 3. Perhitungan berat molekul protein ... 101 4. Prosedur pengenceran serum antitetanus standar dan toksin
tetanus ...
Sistem Imun
Sistem imun dimiliki oleh semua spesies, digunakan untuk mempertahankan diri melawan benda asing yang masuk ke dalam tubuh. Keberadaan protein asing dan patogen dimonitor secara rutin oleh tubuh melalui pelepasan imunoglobulin. Sistem kekebalan akan mampu mengenali spesifik zat kimia yang membedakan sebuah patogen asing dari yang lainnya, serta mampu mengenali molekul asing dengan sel-sel tubuh beserta protein -proteinnya (Kuby 1997). Antibodi mampu mengenali dan berikatan dengan antigen spesifik sampai ribuan atau jutaan antigen. Interaksi antigen-antibodi merupakan interaksi biologi yang sangat spesifik. Sifat khusus itu yang dimanfaatkan dalam teknik imunologi (Abbas et al. 1997).
Pertahanan tubuh melawan infeksi dapat diba gi atas dua yaitu : kekebalan alamiah (non spesifik) dan kekebalan spesifik (adaptive). Kekebalan non spesifik meliputi empat tipe pertahanan yaitu : pertahanan secara anatomi, fisiologi, fagositik, dan peradangan. Pertahanan secara anatomi merupakan pertahanan tubuh yang pertama mencegah masuknya mikrob patogen ke dalam tubuh. Pertahanan secara anatomi terdiri atas barier fisik kulit, selaput lendir, silia, proses batuk, dan bersin. Barier fisik berperan mencegah penetrasi patogen ke dalam tubuh dengan cara melisiskan dan menghambat kolonisasi kuman. Sebagian besar bakteri gagal untuk hidup lebih lama pada kulit karena pengaruh hambatan langsung dari asam laktat dan asam lemak yang disekresikan kelenjar keringat dan sekresi glandula sebaseus (Roitt 1988) .
Respon imun spesifik terbentuk dari kemampuan tubuh menghasilkan respon yang spesifik untuk melawan patogen yang masuk ke dalam tubuh. Secara klasik respon imun spesifik dikelompokkan menjadi kekebalan humoral dan kekebalan berperantara sel (Roitt 1988). Dua tipe sel yang berperan secara aktif yaitu makrofag dan limfosit. Makrofag menguraikan antigen untuk disajikan pada sistem imum, dan limfosit mengenali fragmen antigen yang disajikan untuk produksi antibodi (Halliwell dan Gorman 1989). Imunitas spesifik selama merespon substansia asing juga membentuk sel memori sehingga mudah mengenali antigen jika terjadi paparan yang berulang (Roitt 1988).
Respon humoral meliputi interaksi sel B (sel plasma) dengan antigen dan selanjutnya proliferasi dan diferensiasi membentuk antibodi dengan atau tanpa bantuan sel T. Limfosit B mengekspresikan imunoglobulin permukaan yang spesifik terhadap epitop dari antigen, dan limfosit T mengenali antigen yang telah diproses pada sel presenting antigen. Antibodi yang disekresikan oleh sel plasma menghasilkan antibodi soluble (terlarut). Respon imun selular meliputi interaksi reseptor sel T dan antigen yang telah diproses. Respon itu melalui dua jalur. Pertama , interaksi sel T dengan antigen dan sekresi limfokin untuk menarik makrofag yang akan memfagositosis antigen. Kedua, interaksi sel T sitotoksik dengan antigen yang dipresentasikan oleh MHC II yang akan menyebabkan lisis sel (Roitt 1988).
Berdasarkan proses terbentuknya kekebalan dalam tubuh, kekebalan dibedakan atas dua tipe, yaitu kekebalan aktif dan kekebalan pasif. Pada proses imunisasi aktif tubuh akan memproduksi antibodi dan memberi kekebalan yang lama. Pada imunisasi pasif antibodi terbentuk segera tetapi memberikan perlindungan dalam waktu singkat (Abbas et al. 1997).
optimal memproduksi antibodi, dibandingkan dengan mamalia yang hanya memproduksi kolustrum saat partus (Da vis and Reeves 2002). Produk imun itu memberikan perlindungan secara alami terhadap infeksi selama perkembangan sistem imun anak belum berfungsi optimal (Anonim 2002).
Imunoglobulin
Imunoglobulin atau antibodi adalah kelompok protein yang mempunyai kema mpuan berikatan secara spesifik pada antigen dan mengeluarkan antigen itu dari tubuh. Antibodi adalah molekul protein yang dihasilkan oleh sel plasma sebagai akibat interaksi antara limfosit B peka antigen dengan antigen khusus (Kuby 1997). Struktur dasar dari antibodi tersusun atas empat rantai polipeptida yaitu dua rantai berat dan dua rantai ringan yang identik (Male et al. 1987).
Rantai berat (H) dan rantai ringan (L) disatukan oleh ikatan kovalen disulfida. Posisi ikatan sulfida bervariasi tergantung dari kelas dan subkelas antibodi. Setiap molekul antibodi terbagi atas bagian yang dapat berubah (variable) dan bagian yang tetap (konstan). Bagian variable merupakan tempat pertautan antigen, sedangkan bagian konstan tempat sifat biologi antibodi. Bagian variabel dihubungkan dengan bagian konstan oleh bagian engsel. Pada bagian variabel terdapat bagian hipervariabel untuk mengenali berbagai variasi antigen. Bagian variabel dan konstan terdapat pada rantai berat dan rantai ringan antibodi (Kuby 1997).
fabricius, bone marrow, limpa, timus, glandula harderian, limponodus, limfosit yang bersirkulasi, dan jaringan limfoid pada saluran cerna (Shimizu et al. 1992; Hatta et al. 1993; Sharma 1997).
Imunoglobulin Y
Terminologi (istilah) IgY telah diperkenalkan sejak tahun 1969 dalam literatur yang diistilahkan dengan 7-S Ig terutama yang terdapat di serum, tetapi juga ditemukan dalam isi duodenum, bilasan trakea, dan plas ma seminal (Hadge dan Ambrosius 1984). Imunoglobulin Y telah diisolasi dari unggas (kalkun, ayam, itik, angsa) , ampibi, reptil (Hadge 1985), dan kura-kura darat (Hadge dan Ambrosius 1986). Pada awalnya, beberapa peneliti menduga bahwa IgY yang dihasilkan bangsa unggas sama dengan IgG mamalia, sedangkan kenyataannya berbeda (Szabo et al. 1998).
Transpor IgY dari serum induk ke anak meliputi dua proses. Pertama , IgY ditransfer melewati epitel folikular dari ovari dan berakumulasi dalam kuning telur selama masa oogenesis, yang mirip dengan proses transfer IgG melalui plasenta pada mamalia. Kedua, pemindahan IgY dari kuning telur ke embrio yang sedang berkembang. Isotipe antibodi yang lain seperti IgA dan IgM ditransfer dalam jumlah terbatas ke putih telur (Sharma 1997). Konsentrasi IgY dalam kuning telur konstan sampai oosit matang. IgY tidak terdapat dalam putih telur, sedangkan IgA dan IgM hanya terdapat dalam putih telur. Tidak terjadi seleksi atau destruksi IgY selama proses transfer itu (Davis and Reeves 2002). IgY dalam kuning telur dipersiapkan untuk memberikan kekebalan pasif pada anak ayam. Kuning telur mengandung 8 sampai 20 mg IgY per ml atau 136 sampai 340 mg per kuning telur. Dalam setahun dapat diisolasi 30 g sampai 40 g IgY (Shimizu et al. 1992), sedangkan pada mamalia hanya 1.3 g (Davis and Reeves 2002). Hal itu menyebabkan ayam sebagai sumber IgY mendapat perhatian serius (Shimizu et al. 1992). Penelitian dan penggunaan Ig dari ayam, terutama IgY untuk terapi, pencegahan, dan diagnostik dalam satu setengah dekade terakhir berkembang dengan pesat.
antigen yang diberikan (protein, bakteri, virus dan parasit) (Hatta et al. 1993). Sedangkan berat molekul, morfologi, dan mobilitas imunoelektroforetik dari IgA dan IgM ayam mirip dengan IgA dan IgM mamalia (Davis and Reeves 2002). IgY tidak bereaksi silang dengan komponen struktural jaringan mamalia (Larsson et al. 1993). Hal ini me mberikan indikasi penggunaan IgY dalam diagnostik imunologis akan menghasilkan reaksi yang lebih spesifik. Hassl et al. (1987) melaporkan spesifisitas antibodi serum IgY ayam yang di imunisasi dengan antigen toxoplasma gondii lebih tinggi dibandingkan dengan serum antibodi IgG kelinci. Lebih lanjut, antibodi spesifik (IgY) yang ada dalam darah induk ayam, secara baik dapat ditransfer ke dalam telur. Titer IgY dalam darah dan dalam telur tidak berbeda secara signifikan (Larsson et al. 1993), dan tidak ada perbedaan kandungan IgY pada dua spesies ayam berbeda (Li et al. 1998). Sehingga telur dapat digunakan sebagai sumber protein hewani dan sebagai pabrik produksi antibodi (Regenmortel 1993; Losch et al. 1986).
Gambar 1 Perbedaan struktur IgY dan IgG (Sumber. Schade et al. 1996).
Pada itik dilaporkan memiliki tiga tipe imunoglobulin serum yaitu IgM dan dua bentuk mirip (isoform) IgY yaitu IgY utuh dan IgY terpotong. IgY utuh memiliki berat molekul 200 kDa dengan koefisien sidementasi 7.8 S dan IgY terpotong memiliki berat molekul 130 kDa dengan koefisien sidementasi 5.7 S. IgY terpotong kehilangan dua domain terminal pada regio konstan dari rantai berat yaitu domain 3 dan 4 (Warr dan Higgins 1995).
Tabel 1 Karakter imunoglobulin itik
Berat Molekul Jenis imuno
globulin
Koefisien
sidementasi Molekul utuh
(kDa)
Rantai Berat (kDa)
Rantai Ringan (kDa)
Ig M 800 –900 86 23 – 25
Ig Y utuh 7.8 S 178 – 200 62 – 67 22 – 25
Ig Y terpotong 5.7 S 118 - 130 35 – 42 22 – 25
Dikutip dari :Warr dan Higgins, (1995)
al. 1994). Struktur dan antigenitas 5.7S IgY mirip dengan fragmen F(ab’)2 dari 7.8S IgY (Warr dan Higgins 1995). Itik membentuk dalam jumlah besar IgY()Fc). Bentuk ini cacat karena kehilangan dua domain C-terminal pada rantai H (υ). Struktur abnormal dari IgY()Fc) menyebabkan penurunan fungsi biologis Ig seperti aglutinasi, presipitasi, fiksasi komplemen, opsonisasi (Chan et al. 1999; Lundqvist et al. 2001), walaupun level serum dari boster meningkat (Warr dan Higgins 1995). Faktor lain yang berpengaruh yaitu pembentukan sterik dari lengan Fab (berfungsi monovalensi), regio engsel (hinge) yang kaku, keragaman yang sempit atau terbatas, kegagalan dalam pematangan ikatan antigen pada antibodi (Magor et al. 1994). Respon imun mukosa dependen-IgA itik perkembangannya terlambat selama penetasan dibandingkan dengan ayam (Lundqvist et al. 2001). Pada itik IgA mulai dideteksi pada umur 14 hari setelah menetas dan berfungsi optimal setelah umur 35 hari, sedangkan pada ayam telah berfungsi optimal pada umur 5 hari setelah menetas (Magor et al. 1998; Chan et al. 1999).
Berbagai metode ekstraksi dan purifikasi telah dilaporkan oleh beberapa ahli. Ekstraksi IgY me lalui water dilusi (pelarutan dalam air) kuning telur (Akita dan Nakai 1992); presipitasi lemak dengan dektran sulfat yang mengandung CaCl2 (Szabo et al. 1998). Hasil ekstraksi dilakukan purifikasi dengan
kromatografi menggunakan ion exchange (DEAE-Sephacel) dan filtrasi gel (Szabo et al.1998), mencampur serum dengan asam caprylat, diendapkan dengan amonium sulfat dan didialisis dengan PBS. Teknik ini sangat cepat, murah, sederhana dibandingkan dengan menggunakan metode ion exchange atau gel filtrasi kromatografi (Bhanushali et al. 1994). Purifikasi IgY dari telur dengan
thiophilic interaction chromatography merupakan prosedur purifikasi untuk homogenitas IgY dalam langkah kromatogra fi tunggal setelah fraksinasi amonium sulfat. Recoveri dengan prosedur ini mampu sampai 100% (Hansen et al. 1998).
Metode pelarutan dalam air dilakukan untuk memisahkan plasma protein terlarut dari granul kuning telur. IgY aktif dengan tingkat kemurnian yang tinggi didapat dari kombinasi beberapa teknik seperti presipitasi garam, filtrasi gel dan
lain untuk isolasi adalah metode dua langkah purifikasi yaitu presipitasi dengan PEG diikuti de ngan perlakuan alkohol. Uji spesifisitas dilakukan dengan cara hemaglutinasi indirek, uji imunodifusi, dan imunoelektroporesis (Hassl et al. 1987). Jumlah Ig spesifik yang terdapat dalam telur dari ayam yang diimunisasi adalah 1% dari total IgY (Hansen et al. 1998).
Keuntungan Penggunaan Imunoglobulin Y
Sistem imun ayam dilaporkan telah dipelajari lebih dari satu abad yang lalu, di awali dengan pengamatan pada ayam yang diimunisasi menunjukkan adanya transfer imunoglobulin dari serum ke kuning telur (Ca menisch et al. 1999). Transfer ini diperlukan embrio aves dan anak untuk melawan berbagai penyakit. Penelitian pada sistem imun ayam berkontribusi secara substansial untuk memahami konsep mendasar dari imunologi dan perkembangan kelas Ig yang berbeda. Perkembangan penelitian pada imunoglobulin unggas terutama ayam juga di dukung oleh perkembangan ilmu pengetahuan dan semakin tingginya kesadaran akan animal welfare. Perkembangan penelitian itu melaporkan ayam sebagai alternatif terbaik untuk produksi antibodi poliklonal (Akita dan Nakai 1992; Shimizu et al.1992; Hatta et al. 1993; Schade dan Hlinak 1996; Camenisch et al. 1999).
Penggunaan ayam sebagai sumber imunoglobulin mempunyai beberapa keuntungan antara lain : pemeliharaan ayam tidak mahal, koleksi te lur tida k invasif, isolasi dan afinitas purifikasi IgY cepat dan sederhana, aplikasi IgY sangat luas (Camenisch et al. 1999) . Ekstraksi IgY dari telur lebih menguntungkan dibandingkan dengan ekstraksi Ig mamalia. Keuntungan yang nyata adalah : lebih mudah mengkoleksi telur dari ayam dibandingkan koleksi serum dari mamalia, ketika mengambil darah dari mamalia memerlukan keahlian khusus sedangkan telur dapat dikoleksi oleh tenaga yang tidak dilatih secara khusus, harga pakan dan kandang ayam lebih murah diba ndingkan dengan hewan laboratorium (Camenisch
tidak mengalami cekaman meskipun dalam periode bertelur. Sehingga penggunaan ayam menjadi metode alternatif untuk mengurangi penderitaan hewan. Jumlah hewan yang diperlukan untuk produksi antibodi lebih sedikit, karena ayam mampu memproduksi antibodi lebih tinggi dibandingkan kelinci (Gross dan Speck 1996).
Antibodi ayam memiliki lebih banyak epitop terhadap antigen mamalia (Carlander et al. 1999) , dapat digunakan untuk menghindari kesalahan
immunoassay akibat sistem komplemen (Fryer et al. 1999), faktor rheumatoid, dan reseptor Fc bakteri (Carlander et al. 1999). Pada pengukuran High-sensitivity C-reaktive protein (hs-CRP) yang merupakan salah satu marker untuk pengukuran resiko jantung, penggunaan antibodi mamalia seperti kelinci, tikus, dan kambing memberikan hasil kurang memuaskan, hal ini akibat faktor rheumatoid dalam sampel meyebabkan reaksi positif palsu. Permasalaha n itu dapat ditanggulangi dengan penggunaan IgY (Tsen et al. 2003).
Perbedaan jarak pilogenetik antara mamalia dan avian menyebabkan protein mamalia yang conserved (sulit isolasi juga unik) lebih imunogenik pada ayam dibandingkan dengan mamalia dan respon antibodi spesifik yang dihasilkan sangat tinggi (Akita dan Nakai 1992; Lee et al. 1997; Halper et al. 1999; Orsini et al. 2001). Sehingga ayam sebagai pilihan terbaik untuk produksi antibodi dibandingkan dengan mamalia jika antigen berasal dari manusia atau mamalia.
Isolasi dan metode purifikasi IgY sederhana dan mudah (Gassmann, 2002). Kuning telur mengandung lemak yang tinggi (lipoprotein, granul phospitin yang bercampur dengan livetin dan low density lipoprotein), yang bermasalah jika digunakan secara langsung (Makvandhi dan Fiuzi 2002). IgY yang telah dimurnikan dapat bertahan satu tahun pada suhu 4 oC dengan ditambahkan anti pertumbuhan bakteri seperti Na-azide. Pada suhu kamar stabil selama sebulan. Untuk freeze antibodi hendaknya dibuat aliquot dan hindari freeze dan thawing.
Freeze dan thawing lebih merusak antibodi dibandingkan disimpan pada suhu 4
oC selama satu minggu atau sebulan (Polson 1990) .
Beberapa dari metode itu telah dimodifikasi untuk menyesuaikan dengan sifat antibodi ayam. Hasil penelitian menunjukkan IgY ayam mampu menggantikan IgG yang dihasilkan dengan metode tradisional dari mamalia. Penggunaan IgY sangat memperhatikan keamanan hewan, produktivitas tetap tinggi, dan kekhususan tertentu yang dimiliki IgY untuk tujuan diagnosis (Gross dan Speck 1996), dan modifikasi diagnostik (Higgins et al.1995; Doellgast et al. 1997; Kummer dan Li-Chan 1998; Kim et al. 1999). Penggunaan IgY pada uji ELISA tidak berkompetitor dibandingkan dengan menggunakan antibodi mamalia (Benkirane et al. 1998). Aplikasi potensial penggunaan IgY terus meningkat untuk pencegahan penyakit, agen diagnostik dan biologis, suplemen pakan, dan pemberian secara oral untuk propilaksis (Akita dan Nakai 1992).
Tabel 2 Kelebihan IgY dibandingkan dengan IgG mamalia
No IgY Unggas IgG (Mamalia)
1 Cara Pengambilan sampel Tidak menyakiti hewan 4. Reaksi dengan faktor
rheumatoid
kelinci (Schade dan Hlinak 1996). Secara ringkas beberapa kelebihan lain dari IgY dibandingkan dengan IgG mamalia dipaparkan pada Tabel 2.
Penggunaan Teknologi IgY
Teknologi IgY telah digunakan untuk berbagai hal sehubungan dengan imunoterapi dan imunodiagnostik (Sugita-Konishi et al. 1996). Telur (IgY) ayam sebagai makanan mempunyai peran ganda yaitu peran fungsional dan
neutraceutical. Secara fungsional sebagai sumber protein, dan secara
neutraceutical mampu meningkatkan fungsi imun. Peningkatan kekebalan dengan pemberian secara oral Ig telah dilakukan oleh sejumlah peneliti. Pemberian IgY dilakukan melalui produk makanan, terutama untuk formula anak-anak, karena anak-anak merupakan kelompok rentan terhadap penularan patogen melalui makanan (Akita dan Nakai 1992; Makoto et al. 1998). Dilaporkan `pencegahan E coli pada pedet sapi dengan pemberian kolustrum dicampur IgY, pencegaha n rotavirus berhasil dengan baik pada mencit, serta pencegahan diare perjalanan (wisata) (Davis dan Reeves 2002).
Penggunaan IgG mamalia untuk diagnostik pada uji ELISA sering menghasilkan reaksi positif palsu. Hal itu akibat reaksi silang dari IgG suatu spesies dengan spesies lain. Masalah itu dapat ditanggulangi dengan pemakaian IgY ayam. Davis dan Reeves (2002) melaporkan IgY tidak bereaksi silang pada pemeriksaan laktoferin dan proteoglikan manusia dan sapi pada uji ELISA. Spesifitas IgY dari ayam dapat dimanfaatkan sebagai reagen standar untuk alat diagnostik dan mampu meningkatkan akurasi dalam penelitia n.
Stabilitas IgY
Pengetahuan terhadap stabilitas molekul IgY sangat penting, jika IgY digunakan sebagai reagen dalam berbagai kondisi. Stabilitas dari molekul IgY dapat dipengaruhi oleh berbagai perubahan fisik maupun kimia seperti suhu, asam, dan enzim pencernaan. Stabilitas IgY menjadi sangat penting jika dipakai untuk terapi imunisasi pasif yang diberikan secara oral. Aplikasi yang praktis pemberian suatu antibodi pasif dilakukan dengan mencampur antibodi dengan makanan atau material farmaceutikal, sehingga pertimbangan stabilitas antibodi terhadap panas, pH atau enzim digesti harus diketahui dengan baik (Hatta et al. 1993).
Valensi dari IgY adalah dua, sama dengan antibodi mamalia. Regio engsel pada IgY tidak ada menyebabkan IgY kurang fleksibel. Mobilitas yang terbatas akibat kakunya regio engsel berpengaruh terhadap kemampuan antibodi dalam presipitasi atau aglutinasi antigen. Stabilitas IgY dibawah kondisi asam dan digesti pepsin lebih rendah dibandingkan dengan IgG sapi. Tetapi IgY lebih stabil terhadap digesti enzim protease internal seperti tripsin dan kemotripsin, dan terlihat ada subpopulasi IgY tahan terhadap digesti papain (Hatta et al. 1993).
Para peneliti melaporkan, stabilitas IgG kelinci terhadap panas dan asam lebih tinggi dibandingkan dengan IgY. Bentuk dari molekul IgY sering berubah karena pengaruh asam, yang berakibat penurunan aktivitas antibodi (Shimizu et al. 1992). Stabilitas IgY anti HRV pada temperatur di atas 70 oC dan pH 2 sampai 3 lebih rendah diba ndingkan dengan IgG anti HRV kelinci. Temperatur maksimum untuk denaturasi IgG kelinci adalah 77 oC (Hatta et al. 1993). Aktivitas IgY pada kuning telur dan ekstrak kasar menurun dengan meningkatnya suhu dari 70 oC sampai 80 oC, tetapi denaturasi panas antara kedua sampel tidak berbeda. (Chang et al. 1999).
Digesti pepsin sangat cocok untuk preparasi dan purifikasi Fab. Isolasi Fab optimum didapat setelah digesti pepsin terhadap IgY pada pH 4 selama 9 jam dalam konsentrasi NaCl rendah. Kondisi itu mendigesti secara lengkap fragmen Fc antibodi dan hanya menyisakan fragmen Fab (Akita dan Nakai 1993a). Liofilisasi kuning telur dengan 5% gum arabic serbuk sangat baik untuk menjaga stabilitas terhadap protease (Chang et al. 1999).
Tetanus
Tetanus adalah salah satu penyakit yang lazim terjadi pada manusia dan hewan vertebrata. Tetanus telah dikenal oleh manusia sejak sejarah kesehatan mulai dikenal, tetapi sampai saat ini masih merupakan masalah besar pada kesehatan masyarakat di beberapa negara berkembang, terutama pada negara miskin dengan kondisi kesehatan buruk, beriklim panas dan lembab (Bizzini 1993).
Agen penyebab tetanus adalah Clostridium tetani, tumbuh dalam kondisi anaerob, spora berbentuk batang (Kiefer 2004) , di bawah mikroskop terlihat seperti stik drum dengan gelembung di kedua ujungnya, dengan pewarnaan gram sel bakteri menyerap warna sedangkan spora tidak terwarnai. C. tetani tumbuh optimum pada media agar darah yang diinkubasikan pada suhu tubuh manusia. Bakteri akan berada dalam bentuk inaktif dengan menghasilkan spora ketika lingkungan tempat tumbuhnya tertekan. Dalam kondisi seperti itu, bakteri sangat toleran dengan kondisi lingkungan yang ekstrim, sedangkan dalam bentuk aktif mensekresikan eksotoksin yang sangat poten menyebabkan penyakit tetanus (Anonim 2003).
Habitat alami kuman tetanus adalah tanah, debu, saluran cerna beberapa hewan, dan kadang-kadang pada feses manusia (Ray 2004). Beberapa varian dari kuman ini telah dipetakan secara genomik. C. tetani E88 merupakan varian dari
gen terlihat tidak berfungsi karena terjadi penurunan ORF akibat insertion, delesi, dan poin mutasi. Variasi G+C pada genom sangat rendah, hanya pada region yang kandungan G+C nyata tinggi (sebanding 50%) memiliki 6 gen cluster rRNA dan gen penyanding protein ribosom. Rendahnya fluktuasi dari G+C menyebabkan genom dari C. tetani lebih stabil dibandingkan enteropatogen lain.
C. tetani terus dapat tumbuh apabila tersedia ion sodium secara ekstensif sebagai bioenergetik (Bruggemann e t al. 2003).
Semua spesies hewan rentan terhadap toksin tetanus, tetapi tingkat kerentanannya berbeda -beda. Manusia dan kuda paling rentan sedangkan kucing dan burung lebih tahan. Berdasarkan LD50/kgBB, pada burung diperlukan 10 000 sampai 300 000 kali dosis toksin lebih tinggi dibandingkan dengan kuda untuk menimbulkan sakit (Bizzini 1993).
Manifestasi penyakit tetanus dikelompokkan menjadi empat yaitu : paralisis spastis umum (general), cephalic, lokal, dan neonatal. Tetanolisin dan tetanospasmin merupakan toksin yang dihasilkan oleh kuman tetanus dan sangat berbahaya (Ray 2004). Pada manusia dosis letal dari toksin adalah 1 ng/kg BB. (Bruggemann et al. 2003). Tetanus lokal ditandai dengan kekakuan otot disekitar luka. Gejala ini akibat kegagalan inhibisi dari inervasi syaraf spinal dan medula pada otot yang terserang. Tetanus lokal serangan ringan dan mortalitas kurang dari 1%. Tetanus cephalic kejadiannya jarang. Masa inkubasi sangat pendek yaitu satu sampai dua hari, dan penyakit muncul apabila te rjadi luka di daerah wajah dan kepala. Kelumpuhan daerah wajah dan okulomotoris merupakan gejala utama dan sering diikuti dengan dispagia. Prognosisnya jelek karena penyakit sering ke arah tetanus general. Tetanus neonatal terjadi karena kontaminasi pada daerah umbilikalis. Dengan masa inkubasi yang sangat pendek sehingga bayi tak bisa terawat. Kegagalan terjadi karena gangguan pernafasan dengan kematian empat sampai 14 hari (Bizzini 1993).
ditemukan pada luka nekrosis yang tertutupi oleh keropeng, tanah, debu dan terbungkus kain (Siegmund 1979; Mims 1982). Infeksi dari spora C. tetani juga dapat melalui luka saat kastrasi, potong ekor, pencukuran bulu, dan tali pusar (Lewis 1998). Luka dengan kondisi oksidasi-reduksi yang rendah mempercepat spora germinasi dan berubah menjadi sel bakteri yang aktif (Kiefer 2004).
Keparahan kejadian tetanus tergantung atas jumlah toksin yang mampu mencapai CNS, dan masa inkubasi yang pendek. Periode inkubasi (waktu yang diperlukan dari saat inokulasi sampai muncul gejala pertama) dapat dibedakan atas periode onset atau waktu yang diperlukan dari gejala pertama muncul sampai reflek spasmus yang pertama. Bakteri ini bersifat noninvasif dan gejala yang timbul karena pengaruh toksin yang akan terbentuk setelah periode inkubasi. Lamanya periode inkubasi dan keparahan penyakit dipengaruhi oleh jumlah toksin yang terbentuk pada awal infeksi dan toksigenitas strain yang menginfeksi, jumlah dan kecepatan toksin mencapai neural pathways dan sirkulasi darah, kemampuan perpindahan toksin pada neural pathways, panjang dari neural pathways, dan kerentanan reseptor pada CNS (tergantung spesies). Pada manusia masa inkubasi 14 hari sedangkan pada hewan 24 jam sampai dua minggu atau lebih (Bizzini 1993).
Angka kematian akibat C. tetani menjadi sangat tinggi jika penanganan luka tidak baik dan terjadi dekat organ vital (Ray 2004). Awal infeksi, gejala pada lesi tidak teramati. Apabila kondisi lingkungan tidak mendukung spora akan dorman dalam beberapa hari sampai minggu (Kiefer 2004), jika kondisi mendukung spora mengalami germinasi, dan sel yang baru terbentuk melepaskan toksin yaitu tetanolisin dan tetanospasmin. Angka mortalitas lebih tinggi pada kuman dengan masa inkubasi pendek. C. tetani memerlukan adanya infeksi bakteri lain untuk berkembang di tempat infeksi awal, terutama untuk menimbulkan reaksi peradangan (Ray 2004), sehingga pemberian antitetanus sering dikombinasikan dengan obat untuk bakteri lain (Guidolin et al. 1998).
dengan kekakuan seluruh tubuh, kemudian spasmus otot. Kematian terjadi karena kegagalan pernafasan (Kiefer 2004). Kejadian penyakit lebih sering terjadi di daerah pedesaan di negara dengan iklim panas terutama pada petani yang mendapat luka saat pengolahan tanah pertanian. Pada negara dengan program imunisasi tidak teratur, kejadian tetanus lebih banyak bersifat neonatal (Ray 2004). Di negara maju seperti Amerika, kejadian tetanus masih ditemukan meskipun rendah (50 orang pertahun) terutama pada orang yang tidak diimunisasi, imunisasi tidak lengkap atau teratur, dan telah diimunisasi dengan lengkap tetapi dosis boster tidak ma mpu memberikan perlindungan yang protektif (Ray 2004).
Toksin Tetanus
Toksigenesis adalah kemampuan bakteri patogen memproduksi toksin untuk menimbulkan penyakit. Ditinjau dari sifat biokimia, ada dua tipe toksin bakteri, yaitu toksin yang tersusun atas lipopolisakarida dan protein. Sedangkan berdasarkan atas proses pembentukan dibedakan atas endotoksin dan eksotoksin. Endotoksin adalah toksin yang dihasilkan bakteri gram negatif dari komponen struktural membran luar sel bakteri, dilepaskan dari sel bakteri yang lisis akibat pertahanan inang (enzim lisosim). Komponen penyusun endotoksin adalah lipopolisakarida (LPS) (Emsley 2002) .
Eksotoksin merupakan tipe toksin protein terlarut (soluble), disekresikan oleh bakteri hidup selama masa pertumbuhan eksponensial. Produksi eksotoksin spesifik dari masing-masing spesies bakteri karena memiliki aktivitas sitotoksik pada sel yang khusus seperti tetanus dan botulinum hanya menyerang sel syaraf sedangkan pada tipe sel yang lain kerusakan yang ditimbulkan tidak khas. Dalam aksi sitotoksiknya memerlukan substrat khusus. Substrat itu merupakan komponen dari sel, organ atau cairan tubuh inang. Terminologi terhadap toksin protein bakteri disesuaikan dengan tempat kerja toksin itu seperti enterotoksin, neurotoksin, leukosidin, atau hemolisin. Toksin merupakan faktor virulensi dan hanya diproduksi oleh strain bakteri yang virulen (Todar 2002).
dengan berat molekul 50 kDa, dan ujung carboxyl (C-terminal atau fragmen B) rantai berat (H) dengan berat molekul 100 kDa. Kedua komponen tersebut dihubungkan oleh ikatan disulf ida (Emsley 2000). Toksin dari kuman tetanus merupakan protein yang sangat poten apabila berikatan dengan axon neural syaraf perifer. Toksin yang terbentuk akan mencapai neuron motor dan menyebar secara lokal untuk mencapai sistem syaraf pusat (Mims 1982).
Pada sekuen tingkat DNA, toksin tetanus memiliki homolog dengan neurotoksin botulinum. Memiliki dua rantai disulfida yang berlokasi antara rantai berat dan rantai ringan (cys 438 sampai cys 466), dan dalam fragmen C (cys 1076 sampai cys 1092). Toksin yang dihasilkan oleh kuman tetanus yaitu : tetanospasmin (zink metalloprotease) yang sangat poten menyerang jaringan syaraf (neurotoksin) , dan tetanolisin dengan sifat seperti hemolisin. Produksi toksin tergantung atas kondisi luka dan kultur yang tersedia. Tetanolisin dihasilkan dalam jumlah sedikit oleh strain patogenik dan tidak memainkan peran penting dalam proses penyakit (Bizzini 1993).
Toksin tidak stabil terhadap panas, cahaya, asam, dan enzim proteolitik, sehingga harus disimpan dalam ruang gelap dan dingin. Toksin dapat dipecah oleh enzim proteolitik seperti tripsin, kemotripsin, elastase, clostripain (Habermann 1988), dan papain (Rowe et al. 2000). Digesti toksin dengan enzim papain akan memecah molekul toksin menjadi dua fragmen, yaitu rantai ringan dan rantai berat (Marvaud et al. 1998). Fragmen tunggal toksin kurang toksik dibandingkan dengan toksin secara utuh, untuk penghambatan neuromuskular. Rantai berat dan ringan dari toksin dapat dipisahkan secara isoelektrik. Rantai berat berperan dalam pelepasan noradrenalin dari otak dan K+ dari eritrosit. Pemberian toksin tidak efektif lewat mulut. Toksin dapat diendapkan dengan amonium sulfat dan dalam kondisi kering sangat poten dalam jangka lama (Bizzini 1993).
178 asam amino, dengan gambaran pada DNA-binding protein bermotif helix-turn-helix. Mekanisme pengaturan oleh gen TetR merupakan mekamisme regulasi
conserved untuk gen neurotoksin. Selain mekanisme pengaturan melalui gen, produksi toksin pada C. tetani dipengaruhi oleh suatu peptida rantai pendek pada
casein hydrolysate, dan faktor lain yang penting adalah signal lingkungan di tempat kuman yaitu keberadaan ion Zn (Marvaud et al. 1998). Plasmid pE88 juga menyandi faktor virulen yang lain seperti kolagenase (114 kDa) yang ditandai dengan ColT, tetanolisin O, hemolisin, protein binding-fibronektin. Enzim kolagenase memainkan peranan penting pada patogenesis C. tetani, karena fungsi dari enzim ini untuk merusak integritas jaringan dari inang yang terinfeksi. ColT mirip dengan ColB yang dihasilkan oleh C. botulinum sedangkan dengan spesies clostridium yang lain berbeda (segmen 2 pada ColT tidak ada) (Bruggemann et al. 2003).
Sifat toksisitas toksin protein dapat dihilangkan tetapi sifat antigeniknya tetap dipertahanka n, yang disebut dengan toksoid. Toksoid dibuat dengan cara memberikan perlakuan pada toksin dengan berbagai reagen seperti formalin, iodine, pepsin, asam askorbat, dan keton. Larutan diinkubasikan pada suhu 37 oC dengan pH 6 sampai 9 selama beberapa minggu. Toksoid dapat digunakan dalam imunisasi buatan dan mampu menimbulkan titer antitoksin yang tinggi dalam serum (Todar 2002).
Fragmen HC yang juga disebut fragmen C rantai berat adalah fragmen
terminal karboksil (COOH-terminal) dari toksin tetanus dengan berat molekul 50 kDa, diperlukan pada stadium awal proses intoksikasi untuk aktivitas perlekatan pada gangliosida (Halpern dan Loftus 1993). Topologi dari fragmen HC, terdiri
Gambar 2 Fragmen Hc toksin tetanus (Emsley et al. 2000).
Beberapa peneliti menyebutkan toksin protein khususnya yang bereaksi intraseluler terdiri atas dua komponen yaitu : subunit A (rantai ringan) berespon untuk aktivitas enzimatik dari toksin; subunit B (rantai berat) untuk berikatan dengan reseptor spesifik pada sel membran inang dan tempat transfer enzim untuk melewati membran sel. Toksin tetanus disintesis sebagai polipeptida tunggal, dibagi menjadi domain A dan B yang dapat dipisahkan dengan enzim proteolitik. Komponen enzimatik (subunit A) ini tidak aktif sampai dilepaskan dari toksin natif (A+B). Isolasi subunit A secara enzimatik aktif tetapi kurang mampu berikatan dan masuk ke dalam sel. Isolasi subunit B mampu berikatan dengan sel target tetapi tidak toksik (Todar 2002).
Mekanisme Kerja Toksin Tetanus
Ada dua mekanisme toksin masuk ke sel target (sel syaraf). Pertama, disebut mekanisme langs ung, yaitu subunit B pada toksin natif berikatan dengan reseptor spesifik pada sel target (ujung syaraf motorik) dan menyebabkan terbentuknya lubang pada membran sel sebagai tempat masuknya subunit A ke dalam sitoplasma sel. Proses terikatnya toksin pada syaraf diawali terikatnya toksin dengan afinitas rendah pada gangliosida, selanjutnya kompleks membran-gangliosida-toksin bergerak ke arah lateral sampai berikatan dengan afinitas tinggi pada reseptor protein spesifik toksin. Proses ini diikuti dengan langkah internalisasi toksin setelah itu bergerak retrograde pada axon ke ventral spinal cord dan batang otak (Bizzini 1993). Toksin yang terbentuk menyebar ke otot disekitarnya melalui ikatan dengan terminal presinaptik pada axon motor. Selanjutnya toksin masuk ke sistem limpatik dan vascular darah untuk menyebar ke seluruh otot dan ujung syaraf. Cara penyebaran toksin seperti ini menyebabkan tetanus general atau juga disebut descending tetanus. Blood brain barier dan
blood barier nerve perifer merupakan jalan masuk langsung toksin ke sistem syaraf (Bizzini 1993).
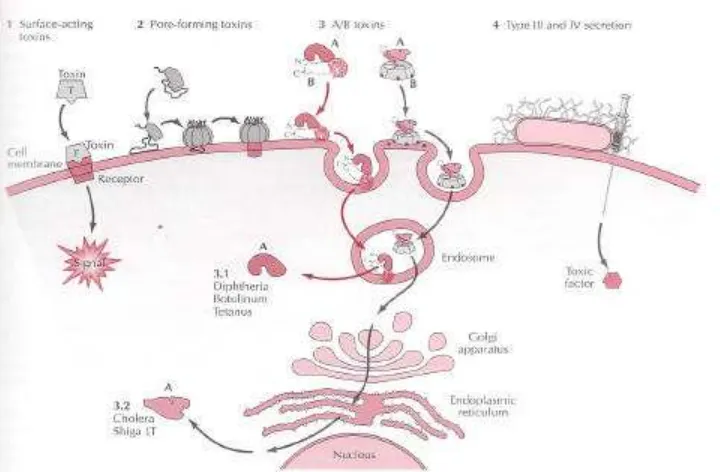
Gambar 3 Proses perlekatan toksin bakteri pada sel (Rappuoli dan Montecucco 1997).
Mekanisme kerja toksin tetanus meliputi empat proses yaitu : perlekatan pada sel ganglion; internalisasi (masuk) ke dalam vesicular; translokasi sitoplasmik; dan pelepasan proteolitik pada subs trat neuron. Perlekatan toksin tetanus pada sel ganglion diawali dengan terikatnya toksin pada gangliosida permukaan sel syaraf. Gangliosida sel syaraf mengandung substansia sialogangliosida yang sering disebut protein-G pada membran sel. Hal ini dimungkinkan karena ujung karbonil (HC) fragmen rantai berat dari toksin tetanus
mengandung ganglioside-binding site (Gambar 4) . Masing-masing kuman menggunakan protein-G yang berbeda, misalnya toksin kolera menggunakan ganglioside GM1 (Bruggemann et al. 2003), toksin botulinum menggunakan ganglioside GT1b (Yowler et al. 2002), dan toksin tetanus menggunakan ganglioside GT1 dan atau GD1b; dan N-glikosilat p15 (Miana-Mena et al. 2002).
raft. Lipid raft (anyaman lipid) adalah suatu kompleks yang terbentuk dari protein glycosylphospatidylinositol (GPI), gangliosida, kolesterol dan spingolipid yang terletak dipermukan neuron. Komponen tersebut membentuk suatu anyaman yang lebih dominan kandungan lipidnya (Herreross et al. 2001). Perlekatan toksin tetanus pada lipid raft juga karena pengaruh marker rafts yang terkandung didalamnya. Marker rafts tersusun atas glikoprotein yang mempunyai berat molekul 15 kDa, sehingga sering disebut p15 (Herreros et al. 2000). Ketahanan dan keutuhan dari lipid rafts itu dipengaruhi oleh kandungan kolesterol pada permukaan neuron. Beberapa toksin yang dikenal sebagai pore-forming toxin
berikatan pada komponen lipid rafts dipermukaan sel (Herreros et al. 2001). Anyaman lipid (lipid raft) adalah mikrodomain pada membran plasma. Komponen ini berfungsi untuk menyeleksi zat yang akan masuk ke dalam vesikel, lalu lintas menuju puncak membran, dan penerima tanda (signaling) (Brown dan London 2000).
Molekul gangliosida merupakan kelas glikospingolipid, ditemukan dalam konsentrasi tinggi pada membran sel neuron (Fotinou et al. 2001). Gangliosida tersusun atas asam sialat (N-acetylneuranimic) yang terikat pada oligosakarida (galaktose dan N-acetylgalaktosamin) dan linked dengan ceramide. Bentuk dasar gangliosida adalah Galβ3GalNAcβ4 (NeuAcα3) Galβ4GlcβCer, dengan satu atau
lebih asam sialat. GM1 dan GD1b memiliki residu asam monosialik dan disialik
yang berikatan dengan residu internal galaktosa, sedangkan GT1b dan 6Q1b berikatan dengan residu terminal galaktosa (Emsley 2000). Fragmen HC toksin
tetanus akan mengenali reseptor gangliosida dan satu gangliosida akan berikatan secara simultan terhadap lebih dari satu molekul toksin tetanus (Lalli et al. 1999; Williamson et al. 1999; Knight et al. 1999). Toksin tetanus berikatan pada bagian Gal4-GalNac3 dan Sia7-Sia6 (Gambar 4)(Fotinou et al. 2001). Perlekatan toksin tetanus pada gangliosida, dan patogen lain seperti toksin E coli heat-labil tipe I, toksin kolera dan simian virus untuk menghindari degradasi oleh lisosim (Miana-Mena et al. 2002).
Terikatnya fragmen HC dari subunit B menyebabkan terbentuknya lubang
pada membran sel syaraf, melalui lubang ini toksin masuk ke dalam vesikular sel. Fragmen Hc mempunyai kemampuan melekat pada neuron dan diperlukan sebagai alat transporasi intraseluler oleh toksin tetanus (Lalli et al. 1999). Toksin tetanus mengalami proses pemecahan proteolitik menjadi ujung amino rantai L dan rantai H. Rantai H dapat dipecah menjadi fragmen HC dan HN.
Masing-masing fragmen memiliki fungsi yang berbeda. Fragmen HC untuk berikatan pada
sel yang disensitisasi kemudian internalisasi ke vesikel. Sedangkan fragmen HN
metaloprotease untuk melawan sinaptobrevin dan merangsang terjadinya keracunan (Emsley et al. 2000; Fotinou et al. 2001). Aktivitas proteolitik dari rantai L terjadi secara selektif pada protein sinap, yaitu hanya pada sinaptobrevin (Herreros et al. 2001). Sinaptobrevin yaitu suatu komponen esensial untuk eksositosis sel neuron. Keberadaan bioenergetika ion sodium yang ekstensif dipercaya sebagai faktor tambahan keberhasilan invasi kuman tetanus ke jaringan (Bruggemann et al. 2003).
Toksin untuk sampai pada sistem syaraf pusat (CNS) melalui syaraf perifer mene mbus sawar darah pada sinap (Mims 1982). Toksin berkumpul pada ujung syaraf (presinaptik) dan translokasi pada alpa motor neuron (Habermann 1988; Lewis 1998) kemudian berjalan sepanjang axis silinder syaraf motorik. Rantai berat dari toksin akan berikatan dengan reseptor gangliosida neuron dan rantai ringan (sangat toksik) akan mengganggu kontrol horn anterior. Motor syaraf pada batang otak sangat pendek sehingga toksin akan cepat sampai di nervus cranialis I yang berakibat spasmus otot mata dan rahang (Mims 1982). Toksin yang terlepas awalnya terserap oleh motor syaraf yang ada disekitar luka dan melalui traktus syaraf mencapai spinal cord, proses ini disebut ascenden tetanus. Peristiwa ini menyebabkan gejala spasmodik, kontraksi tonik. Jika toksin yang dilepaskan pada tempat infeksi, menyebar melalui jaringan limpe kemudian buluh darah kemudian sampai pada CNS, proses ini disebut descenden tetanus dengan gejala berupa tetanus general (Siegmund 1979).
Pergerakan toksin pada organel axonal diperlukan mikrofilamen dan setelah axonal retrograde diperlukan peran myosin Va dan motor mikrotubuli (Lalli et al. 2003). Waktu yang diperlukan untuk perjalanan toksin menuju otak berhubungan dengan masa inkubasi penyakit (Gambar 5). Pemutusan syaraf itu akan memperlambat perjalanan toksin. Pemotongan spinal cord akan mencegah toksin sampai di otak. Sedangkan gangglion spinal dari syaraf sensoris sebagai barier penyebaran toksin (Lewis 1998).
sampai empat hari setelah muncul gejala klinis pertama (Lewis 1998). Tempat berikatan toksin tetanus dan toksin botulinum sama pada ujung terminal
neuromuscular motor junction, tetapi mekanisme intraseluler pada sistem syaraf berbeda sehingga kedua toksin menunjukan gejala yang berbeda. Toksin botulinum (BoNTs) kerjanya bersifat lokal pada sistem syaraf perifer dengan mengganggu perlepasan neurotransmiter yaitu menghambat pelepasan a setilkolin
Gambar 5 Patogenesis penyakit tetanus (Anonim 2003).
dan menyebabkan paralisis lemah. Sebaliknya toksin tetanus (TeNT) bekerja pada sistem syaraf pusat, berjalan secara retrograde pada axon neuron inhibitor dalam spinal cord dan memecah sinaptobrevin. Pelepasan rantai ringan juga menghalangi pelepasan neurotransmiter dengan cara mencegah pembentukan komplek SNARE sinaptik (Fotinou et al. 2001; Herreros et al. 2001). Miana-Mena et al. (2002) melaporkan toksin tetanus mengambat aktivitas neuromuskular presinaptik melalui penghambatan (menutupi) reseptor tempat berikatan ion sodium pada membran sel. Toksin tetanus hanya menghambat transmisi sinaptik tanpa merusak integritas dari syaraf. Sedangkan toksin botulinum dilaporkan
1. Luka terkontaminasi spora C. tetani
2. Penyebaran Toksin 3. Gejala klinis
Kekakuan,dan paralisis pada nervus motor cranialis dan perifer . Contoh lockjaw
menghambat transmisi pada postsinaptik, yang khusus berikatan dan menghambat reseptor asetilkolin tanpa mempengaruhi fungsionalitas syaraf seperti aliran ion ke dalam sel syaraf.
Gambar 6 Proses penghambatan toksin tetanus terhadap reseptor transmiter inhibitor (Emsley et al. 2000).
Toksin tetanus menghambat pelepasan neurotransmiter dari membran presinaptik pada neuron inhibitor nervus terminal (Gambar 6) (Emsley 2000), kemudian berjalan melalui transpor retrograde dari neuromuskular junction ke sistem syaraf pusat dengan target penghambatan pada neuron di dalam spinal cord dan batang otak mamalia (Bruggemann et al. 2003). Transpor toksin ke tempat aksi di CNS tergantung atas te rikatnya toksin pada reseptor membran presinaptik (keterlimpahan disialo-dan trisialogangliosida atau sialoglikoprotein pada vehikel sinaptik) dan internalisasi ke dalam membran transpor retroaxonal dalam sistem
Waktu yang diperlukan oleh toksin untuk sampai di batang otak tergantung dari lokasi lesi, dari percobaan injeksi pada otot lidah diperlukan waktu kurang dari dua jam mencapai batang otak, sedangkan injeksi melalui otot gastrocnemius diperlukan waktu lebih dari enam jam (Miana -Mena et al. 2002). Toksin mengkatalisis vesikel protein sinaptobrevin pada sinap melalui pelepasan proteolitik. Hal ini akan menyebabkan konstraksi otot secara kontinyu yang pertama kali dilihat pada otot dagu dan leher (lockjaw) (Bruggemann et al. 2003). Tetanospasmin merupakan Zink metaloprotease, dilepaskan dalam luka dan berikatan dengan motor neuron terminal daerah perifer, masuk ke dalam akson dan melalui transpor retr ograde intraneural mencapai nervus cell body di batang otak dan spinal cord. Toksin mengalami migrasi dari daerah sinap ke terminal presinap. Di daerah presinap toksin memblok pelepasan inhibitor neurotransmiter glisin dan gamma-aminobutyric acid (GABA) dengan cara memecah protein yang penting untuk fungsi pelepasan vesikel sinaptik. Salah satu protein penting itu yaitu sinaptobrevin. Protein itu menyebabkan penurunan efek penghambatan dan meningkatkan resting firing rate pada neuron motor sehingga terjadi kekakuan otot (Ray 2004).
Berkurangnya waktu aktivitas reflek, berakibat polisinaptik menyebar ke impul (aktivitas glicinergik). Frekuensi agonis dan antagonis lebih sering dibandingkan dengan inhibisi sehingga terjadi spasmus. Hilangnya inhibisi juga mempengaruhi neuron preganglion simpatetik di daerah lateral substansia abu-abu spinal cord dan menyebabkan hiperaktivitas simpatik dan kadar katekolamin yang bersirkulasi tinggi (Ray 2004).