PENGARUH STRUKTUR LANSKAP
TERHADAP KEANEKARAGAMAN HYMENOPTERA
PARASITIKA PADA LAHAN MENTIMUN
SUMEINIKA FITRIA LIZMAH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa Tesis berjudul Pengaruh Struktur Lanskap terhadap Keanekaragaman Hymenoptera Parasitika pada Lahan Mentimun adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
SUMEINIKA FITRIA LIZMAH. Pengaruh Struktur Lanskap terhadap Keanekaragaman Hymenoptera Parasitika pada Lahan Mentimun. Dibimbing oleh DAMAYANTI BUCHORI, PUDJIANTO, dan AKHMAD RIZALI.
Struktur lanskap memengaruhi keanekaragaman hayati pada suatu ekosistem. Secara umum, lanskap yang kompleks cenderung memiliki keanekaragaman dan komposisi spesies yang lebih tinggi dibandingkan dengan lanskap yang sederhana. Lanskap kompleks dicirikan dengan banyaknya tanaman-tanaman non pertanian di sekitar ekosistem pertanian, sehingga selalu tersedia sumber pakan dan habitat hidup yang sesuai. Sementara itu, lanskap sederhana umumnya didominasi oleh tanaman-tanaman pertanian yang cenderung tidak bervariatif, ditambah dengan sedikitnya tanaman non pertanian. Tujuan dari penelitian ini adalah untuk memelajari keanekaragaman Hymenoptera parasitika pada dua lanskap yang berbeda, yaitu lanskap kompleks dan lanskap sederhana.
Penelitian dilakukan pada empat lahan pertanaman mentimun di tiga kecamatan di Kabupaten Bogor yang dibagi menjadi dua lanskap yaitu lanskap kompleks (K1 dan K2) dan sederhana (S1 dan S2). Di setiap lanskap dilakukan pengambilan contoh Hymenoptera parasitika pada lahan mentimun seluas 25 m x 50 m. Penelitian ini dilakukan pada dua kali musim tanam mentimun. Pengkoleksian Hymenoptera parasitika dilakukan dengan tiga metode, yaitu pemasangan perangkap nampan kuning, perangkap malais, dan pengambilan inang dengan metode transek. Untuk mengetahui hubungan keanekaragaman Hymenoptera parasitika dengan struktur lanskap, dilakukan pengukuran struktur lanskap melalui pemetaan vegetasi dan penggunaan lahan pada masing-masing lanskap dengan radius 50 m, 100 m, 150 m, 200 m, dan 250 m dari lahan mentimun.
Sebanyak 233 spesies Hymenoptera parasitika diperoleh dari empat lahan mentimun di dua musim tanam berbeda. Hasil penelitian menunjukkan bahwa kompleksitas lanskap tidak memengaruhi keanekaragaman dan komposisi Hymenopetara parasitika, tapi berpengaruh terhadap kelimpahannya. Berdasarkan hasil analisis korelasi, parameter lanskap yaitu class area (CA) dan jumlah patch (NumP) dari pepohonan dan lahan kosong memiliki hubungan dengan keberadaan Hymenoptera parasitika di ekosistem pertanian.
SUMMARY
SUMEINIKA FITRIA LIZMAH. Effects of Landscape Structure on Diversity of Parasitic Hymenoptera in Cucumber Field. Supervised by DAMAYANTI BUCHORI, PUDJIANTO, and AKHMAD RIZALI.
Landscape structure has been known to affect biodiversity in an ecosystem. In general, biodiversity and species composition tends to be higher in complex landscape compared to simple landscape. Complex landscape is characterized by the presence of non crop plants that provides food source and alternative habitats for insects, whilst simple landscapes are usually dominated by monoculture crop plants that does not provide many variation of plants as food sources and alternative habitats. The aim of this research is to study the diversity of parasitic Hymenoptera under two types of landscapes, i.e. complex and simple landscape
Fields research was conducted at four agricultural landscape of cucumber fields in Bogor that were grouped into complex landscape (K1 andK2) and simple landscape(S1 andS2). In each landscape, parasitic Hymenoptera were sampled from cucumber field (25 m x 50 m) at two different planting seasons. Parasitic Hymenoptera were collected using three methods i.e. yellow pan trap, malaise trap, and host collecting using transect method. To study the relationship between parasitic Hymenoptera and landscape structures, each landscape was quantified through mapping the vegetation and land-uses within a radius of 50 m, 100 m, 150 m, 200 m, and 250 m from each of the cucumber fields.
In total, 233 species of parasitic Hymenoptera were found from four cucumber fields at two planting seasons. The result showed that landscape complexity did not affect the diversity of parasitic Hymenoptera, but it did affect the species composition. Based on correlation analysis, landscape parameters i.e. class area (Ca) and number of patch (NumP) of trees and open areas are correlated with the presence of parasitic Hymenoptera in agricultural ecosystem.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Entomologi
PENGARUH STRUKTUR LANSKAP
TERHADAP KEANEKARAGAMAN HYMENOPTERA
PARASITIKA PADA LAHAN MENTIMUN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Alhamdulillahi Rabbil’alamin, Puji dan syukur penulis panjatkan kepada Allah SWT yang telah memberikan karunia dan hidayah-Nya sehingga penulis dapat menyelesaikan tesis yang berjudul “Pengaruh Struktur Lanskap terhadap Keanekaragaman Hymenoptera Parasitika pada Lahan Mentimun”, sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada Program Studi Entomologi, Sekolah Pascasarjana, Institut Pertanian Bogor. Penelitian ini telah dilaksanakan di Laboratorium Pengendalian Hayati dan beberapa lahan Pertanaman mentimun di Kabupatn Bogor pada bulan Juli 2013 hingga Agustus 2014.
Pnulis mnghanturkan trima kasih yang sbsar-bsarnya kpada:
1. Ibu Prof Dr Ir Damayanti Buchori MSc, Bapak Dr Ir Pudjianto MSi dan Bapak Dr Akhmad Rizali SP MSi selaku komisi pembimbing yang tlah banyak memberikan masukan, pelajaran, dan motivasi kepada penulis selama penelitian maupun penulisan tesis ini
2. Direktorat Jenderal Pendidikan Tinggi, Kementerian Pendidikan dan Kebudayaan atas Beasiswa Unggulan tahun 2012-2014 dan Hibah Kompetensi tahun 2013 yang telah membantu mendanai penelitian ini 3. FAO (Food and Agriculture Organization) melalui proyek kerjasama
IPB-FAO, Pollinator assessment in Indonesia tahun 2013
4. Ayahanda Sulaiman dan Ibunda Nur Aini Amin atas segala nasihat, pelajaran hidup, semangat dan doa dalam setiap langkah penulis sehingga sampai di titik ini
5. Adik-adik tercinta, Mulya Afrianti, Qhusnul Qhatimah, Muhammad Nuzul Fajar, dan Aulia Suci Ramadhani atas dorongan semangat yang telah diberikan
6. Adik dan sahabat tersayang Arum “Ayumi” Marlina atas segala motivasi, semangat, dan bantuan yang diberikan di masa-masa penyelesaian tesis ini 7. Rekan-rekan seperjuangan Entomologi angkatan 2012, Bang Agus, Bang
Topan, Bang Gilang, Bang Mus, Rion, Ihsan, Nadzir, Bu Wid, Mba Dika, Ita, Yenni, Lutfi, dan Wiwik atas kerjasama, semangat dan bantuannya 8. Rekan-rekan di Laboratorium Pengendalian Hayati, Ibu Adha Sari SP,
Mba Nita, Pak Ucup, Mba Ratna, Ibu Evawati, Kak Laras, Kak Amanda, Bayu, Cici, Rizky, Susi, Ihsan, Badrus, Arfiani, dan Novi atas segala bantuan dan masukannya
9. Teman-teman tersayang, SHINee Shawol, Tohoshinki Cassiopea, EXO EXO-L untuk selalu memberikan semangat kepada penulis dalam menyelesaikan tesis ini.
Semoga hasil penelitian ini bermanfaat dan menambah wawasan bagi kita semua.
DAFTAR ISI
2.1 Hymenoptera Parasitika 4
2.2 Pengertian dan Teori Ekologi Lanskap 5
2.3 Struktur dan Kompleksitas Lanskap 5
2.4 Pengaruh Lanskap terhadap Keanekaragaman Parasitoid dan
Pengendalian Hayati 6
3 METODE PENELITIAN 9
3.1 Tempat dan Waktu Penelitian 9
3.2 Pelaksanaan Penelitian 9
3.3 Analisis Data 14
4 HASIL 16
4.1 Keanekaragaman dan Kelimpahan Hymenoptera Parasitika
pada Pertanaman Mentimun 16
4.2 Perbedaan Komposisi Hymenoptera Parasitika pada Lanskap
yang Berbeda 19
4.3 Hama, Parasitoid dan Tingkat Parasitisasi pada Pertanaman
Mentimun 20
4.4 Pengaruh Struktur Lanskap terhadap Hymenoptera Parasitika 24
DAFTAR TABEL
3.1 Lokasi, pemilik serta luasan lahan penelitian 9
3.2 Kriteria pengelompokan lanskap 10
4.1 Kekayaan spesies dan keanekaragaman Hymenoptera parasitika yang ditemukan pada lanskap kompleks dan sederhana di
Kabupaten Bogor 16
4.2 Jumlah individu, spesies, dan famili Hymenoptera parasitika pada
lanskap kompleks dan sederhana 17
4.3 Jumlah individu masing-masing famili Hymenoptera parasitika pada lanskap kompleks dan sederhana dari penggunaan
perangkap berbeda 19
4.4 Persentase parasitisasi pada larva inang yang ditemukan pada
pertanaman mentimun 22
4.5 Persentase parasitisasi terhadap D. indica dan S. litura pada
pertanaman mentimun di lanskap berbeda dan umur tanaman berbeda 23 4.6 Hiperparasitisasi Stictopisthus sp. terhadap A.Taragamae pada inang
D. indica 23
DAFTAR GAMBAR
3.1 Peta lokasi penelitian di Kabupaten Bogor 9
3.2 Contoh kondisi lahan di sekitar pertanaman mentimun 10 3.3 Karakterisasi lanskap disekitar lahan penelitian 11 3.4 Digitasi struktur lanskap berdasarkan penggunaan lahan (landuse) 11
3.5 Petak contoh lahan pertanaman mentimun 12
3.6 Desain plot pengambilan contoh parasitika pada lahan mentimun 13 3.7 Perangkap nampan kuning (yellow pan trap) 13
3.8 Perangkap malais 14
4.1 Jumlah individu yang diperoleh berdasarkan perangkap 18 4.2 NMDS dari komposisi parasitika berdasarkan indeks
ketidakmiripan Bray-Curtis 19
4.3 Persentase parasitisasi pada lanskap kompleks dan sederhana 21
4.4 Imago hiperparasitoid Stictopisthus sp. 22
4.5 Hubungan parameter lanskap dengan jumlah spesies parasitika 23
DAFTAR LAMPIRAN
1 Keanekaragaman dan kelimpahan Hymenoptera parasitika yang ditemukan pada pertanaman mentimun pada lanskap kompleks (K1
dan K2) dan lanskap sederhana (S1 dan S2) 36
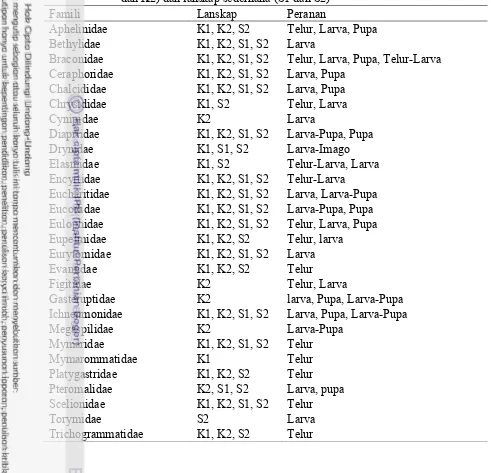
2 Parasitoid dan peranannya yang ditemukan pada lanskap kompleks
1
PENDAHULUAN
1.1 Latar Belakang
Hymenoptera parasitika merupakan bagian dari ordo Hymenoptera yang berperan sebagai parasitoid (Mason dan Huber 1993). Serangga parasitoid hidup pada atau di dalam tubuh inangnya dengan memakan cairan inang sehingga menyebabkan kematian pada inang. Berbeda dengan jenis parasit pada umumnya, serangga parasitoid hanya fase pradewasanya yang menyerang inang sementara dewasa (imago) hidup bebas di luar inang (Godfray 1994; Driesche dan Bellows 1996). Jenis inang dari parasitoid umumnya berasal dari kelas serangga ataupun Arthropoda lainnya (Mason dan Huber 1993; Godfray 1994). Serangga-serangga parasitoid sudah sangat banyak dimanfaatkan sebagai salah satu agen pengendali hayati hama seperti hama pertanian dan perkebunan.
Pengendalian hayati termasuk salah satu bentuk jasa ekosistem yang tersedia di alam selain penyerbukan, kesuburan tanah, dan kesehatan lingkungan (Power 2010, Tscharntke et al. 2012). Jasa ekosistem tersebut tidak hanya bermanfaat untuk kehidupan manusia tetapi juga seluruh komponen ekosistem termasuk tumbuhan, hewan, dan mikroorganisme. Keberhasilan suatu pengendalian hayati tentu saja tidak terlepas dari keanekaragaman dan kelimpahan parasitoid di lapangan. Terdapat beberapa faktor yang memengaruhi keberadaan parasitoid di alam, diantaranya adalah ketersediaan inang, makanan untuk imago, kesesuaian mikroklimat dan keadaan habitat (Debach dan Rosen 1991; Dyer dan Landis 1996, 1997; Godfray 1994). Faktor-faktor tersebut saling terhubung dan memengaruhi serta membentuk suatu fungsi lanskap di dalam ekosistem pertanian yang tentu saja tidak terlepas dari struktur lanskap pertanian itu sendiri (Forman 1995).
Konservasi, dalam hal ini konservasi parasitoid, merupakan salah satu cara untuk menjaga parasitoid agar tetap berada di lapangan (Tscharntke et al. 2007). Konservasi parasitoid dapat dilakukan dengan menerapkan atau menjalankan manajemen lanskap untuk menjaga sumber daya (resource) seperti inang dan makanan, terus tersedia di lapangan (Landis et al. 2000). Menurut Bianchi (2006), lanskap habitat alami memiliki peranan penting dalam pengendalian hama. Lanskap yang didukung oleh habitat alami atau habitat semi-alami berperan sebagai sumber inang alternatif dan mangsa, sumber nektar, refugium bagi musuh alami sehingga mencegah kerusakan tanaman (Thies et al. 2003; Bianchi et al. 2006). Sementara lanskap pertanian memiliki keanekaragaman spesies tumbuhan yang rendah dan kompleksitas tumbuhan yang sederhana (Lawton dalam Menalled et al. 1999) sehingga dengan menerapkan manajemen lanskap yang baik dapat menciptakan keadaan seperti habitat alami atau semi-alami.
Manajemen dan kompleksitas lanskap pertanian dapat memengaruhi keanekaragaman tumbuhan, hewan, dan mikroorganisme di dalamnya (Altieri 1999; Tscharntke et al. 2005), dimana kedua hal tersebut memengaruhi aliran energi, pengaturan air, dan keberadaan musuh alami termasuk parasitoid (Altieri 1999). Menurut Bianchi et al. (2006), kompleksitas lanskap juga memengaruhi kekayaan spesies di lanskap tersebut. Manajemen lanskap dapat dilakukan dengan manipulasi habitat yaitu pemanfaatan tanaman-tanaman non pertanian di sekitar areal pertanian sehingga memberikan fungsi seperti habitat semi-alami (Gámez-Virués et al. 2012). Keberadaan habitat semi-alami berguna dalam menjaga keberadaan parasitoid, termasuk meningkatkan keanekaragaman dan kelimpahannya (Landis et al. 2000). Habitat semi-alami berupa tanaman-tanaman non pertanian yang ditanam di sekitar ekosistem pertanian juga berdampak positif terhadap tanaman pertanian itu sendiri. Tanaman non pertanian ini berguna sebagai penyedia sumber makanan, sumber inang alternatif, refugia, dan habitat yang sesuai bagi parasitoid (Thies et al. 2003; Bianchi et al. 2006).
Struktur lanskap umumnya dikelompokkan berdasarkan komposisi vegetasi pada lanskap tersebut. Pertanian dengan lanskap kompleks dicirikan dengan dominannya tanaman non pertanian di sekitarnya seperti pepohonan, semak, tanaman pagar dan gulma. Sementara itu, lanskap sederhana memiliki proporsi tanaman non pertanian lebih sedikit dan cenderung homogen (Menalled et al. 1999; Plećaš et al. 2014). Struktur lanskap suatu pertanian dapat memengaruhi keanekaragaman, kelimpahan dan keefektifan parasitoid di dalamnya (Marino dan Landis 1996; Menalled et al. 1999). Lanskap kompleks cenderung memiliki keanekaragaman musuh alami lebih tinggi dibandingkan dengan lanskap sederhana (Purtauf et al. 2005; Schmidt et al. 2005). Hasil penelitian Marino dan Landis (1996) menunjukkan bahwa persentase parasitisasi Meteoris communis (Hymenoptera: Braconidae) terhadap Pseudaletia unipuncta (Lepidoptera: Noctuidae) lebih tinggi (13.1%) pada lanskap kompleks dari pada lanskap sederhana (2.4%). Sementara itu Plećaš et al. (2014) menemukan bahwa kepadatan kutudaun diikuti dengan peningkatan rata-rata parasitisasi, kekayaan spesies parasitoid serta hiperparasitisasinya pada lanskap yang dikelola secara ekstensif dan ditanami berbagai jenis tanaman non-pertanian.
Pengaruh kondisi lanskap terhadap keberadaan parasitoid penting untuk diplajari lebih dalam karena dapat memberikan informasi penting mengenai hubungan penyusun lanskap (struktur lanskap) dengan keanekaragaman, kelimpahan dan komposisi parasitoid. Informasi yang diperoleh dapat dimanfaatkan sebagai data penunjang dalam melakukan perancangan strategi konservasi musuh alami, dalam hal ini parasitoid, khususnya di wilayah kabupaten Bogor.
1.2 Tujuan Penelitian
Penelitian ini bertujuan untuk (1) mempelajari keanekaragaman Hymenoptera parasitika di lahan mentimun pada lanskap yang berbeda, (2) membandingkan keanekaragaman dan komposisi Hymenoptera parasitika di lahan mentimun pada kondisi lanskap berbeda, dan (3) mempelajari pengaruh struktur lanskap terhadap keanekaragaman dan komposisi Hymenoptera parasitika di lahan mentimun.
1.3 Manfaat Penelitian
3 mentimun, (2) pengaruh kondisi lanskap terhadap keanekaragaman Hymenoptera parasitika pada pertanaman mentimun, dan (3) data ilmiah sebagai bahan pertimbangan atau rekomendasi dalam melakukan perencanaan kawasan pertanian dalam upaya mewujudkan pertanian yang berkelanjutan.
1.4 Hipotesis
Hipotesis yang diajukan dalam penelitian ini adalah:
H0: struktur lanskap tidak memengaruhi keanekaragaman Hymenoptera parasitika pada pertanaman mentimun.
2
TINJAUAN PUSTAKA
2.1 Hymenoptera Parasitika
Keanekaragaman hayati (biological diversity) didefinisikan sebagai keanekaragaman makhluk hidup (tumbuhan, hewan, mikroorganisme) dari berbagai habitat termasuk daratan, lautan dan ekosistem akuatik, dan kompleks-kompleks ekologi lainnya, mencakup keanekaragaman di dalam spesies, antar spesies dan ekosistem yang saling berinteraksi (Heywood 1995, Altieri dan Nicholls 2004). Harper dan Hawksworth (1995) pertama kali membagi keanekaragaman hayati kedalam tiga komponen, yaitu keanekaragaman genetik (di dalam spesies), keanekaragaman spesies (jumlah spesies), dan keanekaragaman ekologi (di dalam komunitas).
Serangga sebagai salah satu penyusun keanekaragaman hayati bumi memiliki lebih dari jutaan spesies yang sudah ditemukan. Hymenoptera termasuk salah satu ordo dengan jumlah spesies terbesar dalam kelas Insecta selain Coleoptera, Lepidoptera dan Diptera (LaSalle dan Gauld 1993, CSIRO 2001). Ordo ini mewakili sekitar 7.7% dari keseluruhan spesies serangga di dunia dengan lebih dari 115.000 spesies telah teridentifikasi (LaSalle dan Gauld 1993).
Secara fungsional, ordo Hymenoptera dikelompokkan kedalam dua subordo yaitu Symphyta dan Apocrita (Goulet dan Huber 1993). Subordo Symphyta disebut sebagai Hymenoptera primitif, dengan pertulangan sayap (venasi) yang lengkap dan abdomen tanpa penggentingan (CSIRO 2001). Spesies Symphyta umumnya bersifat fitofag dan memiliki bentuk serta perilaku atau biologi yang sama dengan larva lepidoptera (LaSalle dan Gauld 1993). Subordo ini terdiri dari 14 famili dan mempunyai tidak lebih dari 15% spesies dari keseluruhan Hymenoptera (LaSalle dan Gauld 1993). Subordo Apocrita diketahui memiliki peranan yang beragam terhadap ekosistem, diantaranya sebagai parasitoid, predator, pemakan madu (mellivorous), pemakan jamur (fungivorous) atau pemakan bangkai (scavengers); serta berperan sebagai fitofag pada fase larvanya (CSIRO 2001). Berdasarkan peranannya, subordo Apocrita dikelompokkan menjadi dua yaitu aculata dan parasitika. Apocrita aculata memiliki modifikasi pada bagian ovipositor menjadi alat penyengat (stinger) dan umumnya berperan sebagai predator dan polinator (LaSalle dan Gauld 1993, Mason dan Huber 1993). Kelompok parasitika dicirikan dengan ovipositor tidak berkembang sebagai penyengat melainkan sebagai alat untuk meletakkan telur (LaSalle dan Gauld 1993, Mason dan Huber 1993). Kelompok parasitika lebih dikenal dengan sebutan parasitoid. Hampir 75% dari keseluruhan spesies subordo Apocrita adalah parasitoid (Goulet dan Huber 1993) dan berjumlah sekitar 39 famili (LaSalle dan Gauld 1993).
5
2.2 Pengertian dan Teori Ekologi Lanskap
Pembahasan mengenai lanskap di dalam ekologi pertanian tidak terlepas dari dua teori dasar yang dikemukakan oleh Price dan Waldbauer (1975) yaitu teori yang fokus pada ruang atau space (teori biogeografi kepulauan oleh MacArthur dan Wilson 1967) dan teori yang fokus pada waktu atau time (suksesi komunitas oleh Odum 1969). Kedua teori tersebut saling berkaitan dimana ruang dan waktu dapat menentukan kestabilan suatu populasi di dalam ekosistem termasuk populasi hama dan musuh alaminya. Suatu lahan pertanian dapat dianalogikan sebagai sebuah pulau, jarak dan ukuran pulau dari sumber daya menentukan proses terbentuknya kolonisasi ataupun kestabilan populasi serangga, terutama setelah kestabilan tumbuhan di areal tersebut terbentuk (Gámez-Virués et al. 2012).
Lanskap adalah suatu bentang alam atau areal heterogen yang tersusun dari berbagai ekosistem yang saling berinteraksi dan memengaruhi serta memiliki pola sama yang berulang-ulang (Forman dan Godron 1986). Secara lebih sederhana, Turner et al. (2001) mendefinisikan lanskap sebagai suatu daerah atau areal heterogen yang terdiri dari habitat atau jenis penutup areal yang berbeda dan mempunyai ukuran yang bervariasi.
Teori skala spatio-temporal dan perkembangan teknologi memiliki konstribusi dalam munculnya konsep dan prinsip ekologi lanskap yang penting untuk dipelajari. Forman dan Godron (1986) mengidentifikasi beberapa prinsip umum dalam membedakan lanskap berdasarkan ekologi ekosistem, biogeografi, dan bentuk geografi yaitu struktur dan fungsi lanskap, keragaman biotik, aliran spesies, redistribusi nutrisi, aliran energi, perubahan lanskap, dan stabilitas lanskap. Menurut Forman (1995), di dalam sebuah lanskap terjadi tiga proses dasar yang saling berintegrasi, yaitu struktur lanskap, fungsi lanskap, dan dinamika lanskap. Sebuah pengembangan teori yang dilakukan Forman (1995) dalam menyelesaikan berbagai masalah terkait penggunaan lahan dan lingkungan sosial, yaitu lanskap dan region, patches dan koridor, mosaik, dan penerapan lanskap.
Turner et al. (2001) mengemukanan bahwa terdapat beberapa konsep mendasar terkait dengan manajemen hama pertanian, yaitu pertama, komposisi yang merupakan perbedaaan proporsi habitat atau jenis penutup yang ditemukan di areal, kedua, konfigurasi yang berarti susunan spasial yang spesifik dari jenis penutup di atas lanskap, dan ketiga adalah konektivitas dimana terdapat ruang spasial yang berkelanjutan untuk jenis tanaman penutup bentangan lanskap. Sebuah konfigurasi spasial sangat penting dalam proses ekologi, terutama ekologi lanskap. Di sisi lain, luasan spasial dan pengaruh aktivitas manusia juga menjadi komponen penting dalam membentuk pola lanskap (Turner et al. 2001).
2.3 Struktur dan Kompleksitas Lanskap
matriks yang mengelilinginya (Forman dan Godron 1986, Forman 1995). Koridor adalah lahan sempit yang diapit oleh dua sisi yang linear (Forman dan Godron 1986, Forman 1995), dapat berfungsi sebagai habitat, pembatas (barrier) maupun lahan perpindahan serangga dari satu lahan ke lahan yang lainnya (Forman 1995). Masing-masing elemen penyusun lanskap ini mempunyai sifat tertentu yang dapat diukur seperti luasan, jarak, bentuk, jumlah habitat dan hubungan satu bidang lahan terhadap bidang lahan lainnya.
Kompleksitas lanskap adalah istilah yang umum digunakan dalam ekologi lanskap. Istilah ini terkait dengan keanekaragaman elemen lanskap, kelompok spasial, dan bentuknya. Beberapa matriks lanskap mendeskripsikan dimensi yang berbeda dari kompleksitas lanskap (Gámez-Virués et al. 2012). Adapun beberapa istilah umum terkait kompleksitas lanskap seperti yang dikemukakan oleh Turner et al. (2001), antara lain, Shannon diversity index, Shannon evenness index, mean shape index, edge density, dan contagion. Shannon diversity index menggambarkan keanekaragaman pada tipe habitat dengan memperhatikan jumlah kelas tutupan lahan dan distribusi proporsionalnya. Indeks ini mirip dengan indeks keanekaragaman Simpson. Sedangkan Shannon evenness index menggambarkan kemerataan areal antar kelas tutupan lahan dimana nilai tinggi menggambarkan kemerataan daerah antar kelas. Mean shape index menggambarkan hubungan antara perimeter dan luas patch sebagai rataan dari semua patch yang ditemukan. Nilai tertinggi dari rataan menunjukkan bentuk lanskap yang lebih kompleks. Edge density menyediakan pengukuran panjang segmen tepi per hektar dan tergantung antara kedua patch dan bentuk patch. Contagion menggambarkan tingkat kedekatan sel (patch yang berdekatan dari jenis patch yang sama). Contagion dipengaruhi oleh dispersi dan interspersi kelas tutupan lahan.
2.4 Pengaruh Lanskap terhadap Keanekaragaman Parasitoid dan Pengendalian Hayati
Struktur lanskap dan manajemen habitat merupakan dua hal yang memengaruhi konservasi parasitoid dan musuh alami lainnya di lapangan (Altieri 1999, Tscharntke et al. 2005). Struktur lanskap pertanian dalam skala spasial tersusun atas berbagai komponen diantaranya isolasi fragmentasi habitat, area patch, kualitas patch, diversitas patch, edge, derajat monokultur, dan mikroklimat yang memengaruhi kelimpahan dan kekayaan spesies serangga (Marino dan Landis 1999, Hunter 2002). Lanskap pertanian jika diibaratkan sebagai kepulauan, menyebabkan terjadinya isolasi-isolasi habitat sehingga keberadaan parasitoid juga tersebar tidak merata. Hal ini dikarenakan suatu lanskap yang terisolasi belum tentu menyediakan sumber daya yang konstan. Isolasi habitat akibat dari fragmentasi habitat diketahui memengaruhi keanekaragaman dan kelimpahan suatu organisme dalam ekosistem, terutama organisme-organisme penting seperti parasitoid dan spesies lokal. Menurut Kruess dan Tscharntke (1994), isolasi yang terjadi akibat fragmentasi habitat menyebabkan berkurangnya kemunculan jumlah spesies sehingga berdampak pada penurunan keefektifan parasitoid. Sisa-sisa fragmentasi habitat yang kurang terisolasi meningkatkan keanekaragaman spesies dan parasitisme serangga hama potensial dalam membentuk stabilitas fungsi ekosistem (Kruess dan Tscharntke 2000).
7 dan saling memengaruhi, begitupun terhadap parasitoid. Pertukaran ini dapat terjadi pada skala global atau regional (Marino dan Landis 1999). Lanskap pertanian yang didukung oleh keberadaan berbagai tanaman dan tumbuhan liar memungkinkan parasitoid hidup lebih banyak. Tumbuhan liar atau refugium merupakan tanaman non pertanian yang biasanya tumbuh atau ditanam di sekitar lanskap pertanian. Pemanfaatan tanaman liar termasuk salah-satu upaya manajemen lanskap yang disebut dengan manipulasi habitat yang bertujuan untuk meningkatkan efektivitas pengendalian hayati, terutama pada lanskap sederhana (Gámez-Virués et al. 2012). Selain manipulasi habitat, sistem budi daya pertanian, kompleksitas, dan efek tanaman tepi juga memengaruhi keanekaragaman parasitoid pada lanskap (Marino dan Landis 1999). Tanaman non-pertanian yang digunakan untuk manipulasi lokal biasanya menggunakan flower strips, sementara untuk manipulasi lanskap dengan menanami pepohonan non-pertanian (Gámez-Virués et al. 2012).
Di pertanian konvensional, flower strips yang biasanya digunakan adalah yang dapat mengurangi pengaruh hama (Fiedler et al. 2008). Penanaman flower strips biasanya dilakukan dengan memilih tanaman eksotik atau tanaman lokal yang ditanam disepanjang tepi tanaman atau antara baris tanaman. Flower strips berguna sebagai pemasok sumber nektar, polen, dan tempat tinggal bagi parasitoid (Landis et al. 2000). Flower strips memberikan manfaat yang besar dalam mendukung keberhasilan penendalian hama, seperti peningkatan tingkat parasitisasi, lama hidup dan fekunditas. Kartosuwondo (1994) menemukan bahwa parasitisasi Diadegma semiclausum (Hymenoptera: Ichneumonidae) parasitoid larva Plutella xylostella (Lepidoptera: Pyralidae) pada tanaman kubis, tiga kali lebih tinggi pada petakan yang ditanami flower strips. Penanaman flower strips juga meningkatkan lama hidup dan fekunditas Diadegma insulare (Cresson) (Hymenoptera: Ichneumonidae) yang merupakan parasitoid P. xylostella (Idris dan Grafius 1995). Hal yang sama juga berpengaruh pada lama hidup dan parasitisasi parasitoid Copidosoma koehleri (Hymenoptera: Encyrtidae) pada hama kentang Phtorimaea operculella (Lepidoptera: Gelechiidae) (Baggen dan Gurr 1998).
Selain berguna sebagai penyedia sumber daya, flower strips juga berperan dalam penyediaan inang alternatif bagi parasitoid sehingga populasi parasitoid pada ekosistem pertanian pun terpelihara (Scherber et al. 2012). Flower strips haruslah ditanam dengan pertimbangan jarak tertentu dengan harapan disinggahi musuh alami saat melakukan perjalanannya mencari inang (Gámez-Virués et al. 2012). Lavandero et al. (2005) mengamati bahwa D. semiclausum dewasa dapat berpindah sejauh 80 km, padahal jarak yang umum ditempuhnya dalam pencarian inang adalah 60 km. Hal ini dikarenakan pada skala jarak tertentu terdapat tanaman liar yang dapat digunakan sebagai sumber makanan oleh parasitoid.
parasitoid dan hiperparasitoid kutudaun lebih tinggi pada lanskap dengan habitat tanaman non-pertanian yang lebih luas dan beragam, positif memengaruhi pengendalian biologis.
9
3
METODE
3.1 Tempat dan Waktu Penelitian
Pengambilan contoh serangga dilakukan pada pertanaman mentimun di empat lokasi yang tersebar di wilayah Kabupaten Bogor (Gambar 3.1). Penyortiran dan identifikasi serangga dilakukan di Laboratorium Pengendalian Hayati, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Penelitian lapangan dilakukan mulai dari Juni 2013 sampai November 2013 dilanjutkan dengan identifikasi di laboratorium pada bulan Desember 2013 sampai Agustus 2014.
Gambar 3.1 Peta lokasi penelitian di Kabupaten Bogor 3.2 Pelaksanaan Penelitian
Penentuan Lokasi dan Lanskap
Lokasi penelitian yang digunakan merupakan lanskap pertanian dataran rendah di tiga kecamatan di Kabupaten Bogor, yaitu di Kecamatan Dramaga (Desa Cangkurawuk), Kecamatan Ciampea (Desa Cihideung Udik dan Desa Banteng), dan Kecamatan Cibungbulang (Desa Cibatok). Tabel 3.1 menjelaskan keterangan untuk masing-masing lokasi penelitian.
Tabel 3.1 Lokasi, pemilik serta luasan lahan penelitian
Lokasi* Kecamatan Desa Pemilik lahan Luas lahan
K1 Cibungbulang Cibatok Pak Ages 25 m x 75 m
K2 Ciampea Benteng Pak Suhanda 50 m x 57 m
S1 Ciampea Cihideng Udik Pak Kosasih 26 m x 52 m
S2 Dramaga Cangkurawuk Pak Jaya 26 m x 53 m
* K (kompleks), S (sederhana)
kualitatif (keanekaragaman vegetasi, komposisi (heterogen) vegetasi, dan penggunaan lahan (land use)). Masing-masing kriteria kemudian dilakukan pembobotan berdasarkan hasil pemetaan lanskap dengan nilai persentase untuk keseluruhan adalah 100%. Hasil pengklasifikasian diperoleh dua kelompok lanskap yaitu lanskap kompleks (K1 dan K2) dan lanskap sederhana (S1 dan S2) (Tabel 3.2). Lanskap kompleks memiliki lebih dari 50% non pertanian (lahan hijau, pepohonan) dengan komposisi lahan yang heterogen, sedangkan lanskap sederhana terdiri dari kurang 30% tanaman non pertanian dengan komposisi lahan homogen serta vegetasi yang tidak beragam (Gambar 3.2).
Tabel 3.2 Kriteria pengelompokan lanskap Lanskap1 Lahan
Hijau Tanaman berbunga Kondisi lahan Komposisi lahan Keanekagaman vegetasi Penggunaan Lahan2
K1 84.76% 19.84% 40.00% Heterogen Beragam 1, 2, 3
K2 73.00% 38.59% 31.42% Heterogen Beragam 1, 2, 3
S1 79.22% 29.58% 21.16% Homogen Beragam 1, 2, 3
S2 61.15% 13.52% 31.34% Homogen Tidak beragam 1, 2, 3 1 K (kompleks), S (sederhana), 2)1: Pertanian, 2: hutan, 3: perumahan
Gambar 3.2 Contoh kondisi lahan di sekitar pertanaman mentimun, a) lanskap kompleks b) lanskap sederhana
Pemetaan dan Pengukuran Struktur Lanskap
Pemetaan diawali dengan menandai titik lokasi penelitian menggunakan global positioning system (GPS), kemudian memberikan kode pada masing-masing titik sesuai dengan kelompok lanskapnya. Metode pemetaan lanskap yang digunakan mengadopsi metode pemetaan yang dilakukan oleh Scherber et al. (2012) yaitu melakukan pemetaan vegetasi dan penggunaan lahan pada setiap gradien jarak meliputi 50 m, 100 m, 150 m, 200 m, dan 250 m (Gambar 3.3). Di setiap radius dilakukan ground check untuk mencatat komoditas pertanian yang ditanam dan vegetasi tanaman lain seperti pepohonan di sekitar lahan pada setiap lanskap. Data yang diperoleh dari lapangan, selanjutnya dipetakan secara digital dengan menggunakan perangkap lunak Q-GIS (Quantum GIS Development Team 2014).
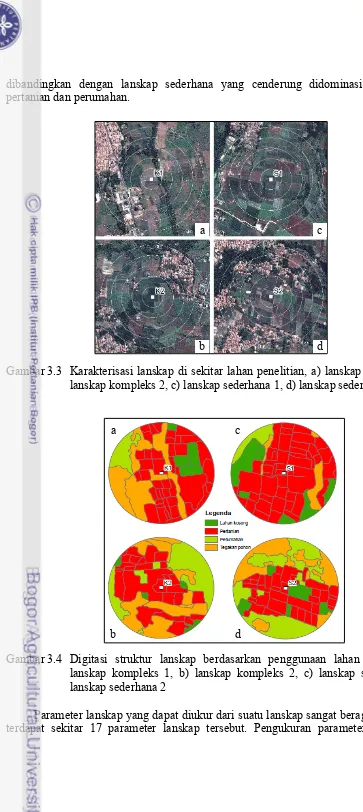
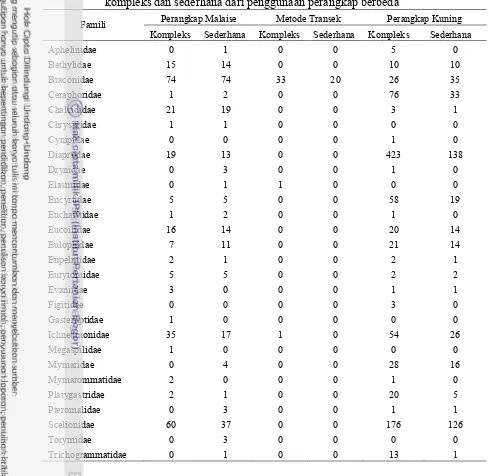
Hasil pemetaan lanskap menggambarkan tipe penggunaan lanskap yang terdapat pada masing-masing pertanaman mentimun pada radius 250 m di masing-masing lanskap penelitian (Gambar 3.4). Lanskap kompleks memiliki proporsi penggunaan lahan (land use) yang lebih merata dibandingkan dengan lanskap sederhana. Lanskap kompleks mempunyai jumlah pepohonan dan tanaman non pertanian lain yang lebih dominan
11 dibandingkan dengan lanskap sederhana yang cenderung didominasi oleh tanaman pertanian dan perumahan.
Gambar 3.3 Karakterisasi lanskap di sekitar lahan penelitian, a) lanskap kompleks 1, b) lanskap kompleks 2, c) lanskap sederhana 1, d) lanskap sederhana 2
Gambar 3.4 Digitasi struktur lanskap berdasarkan penggunaan lahan (land use), a) lanskap kompleks 1, b) lanskap kompleks 2, c) lanskap sederhana 1, d) lanskap sederhana 2
Parameter lanskap yang dapat diukur dari suatu lanskap sangat beragam, setidaknya terdapat sekitar 17 parameter lanskap tersebut. Pengukuran parameter lanskap yang
a
b d
c
a c
Panjang 50 m
digunakan dalam penelitian ini mengikuti metode dari McGarigal et al. (2014). Adapun parameter lanskap yang diukur dalam penelitian ini adalah:
- Class Area (CA): jumlah keseluruhan tipe patch pada tingkatan kelas yang sama. - Number of Patch (NumP): jumlah keseluruhan patch, baik pada tingkatan kelas
maupun tingkatan lanskap.
- Shannon’s Diversity Index (SDI): indeks keanekaragaman Shannon untuk mengukur tingkat keanekaragaman patch dalam lanskap.
- Shannon’s Evenness Index (SEI): untuk mengukur tingkat distribusi patch.
Penyiapan plot pengamatan dan Pengambilan Contoh Serangga
Penyiapan Lahan Tanaman Mentimun untuk Pengamatan. Penanaman
mentimun dilakukan sebanyak dua kali. Penanaman pertama dilakukan dari bulan Juni sampai Agustus 2013, sementara penanaman kedua dilakukan pada bulan September sampai November 2013 pada lokasi yang sama dengan penanaman pertama. Di setiap lanskap dilakukan penanaman mentimun pada lahan seluas 25 m x 50 m (Gambar 3.5). Varietas mentimun yang ditanam yaitu Mutiara Bumi, Alicia F1. Jarak tanaman mentimun yang diterapkan pada lokasi pengamatan adalah 0.6 m x 0.6 m. Benih mentimun ditanam di setiap bedengan dengan lebar 1,0 m – 1.2 m dan tinggi 0.4 m.
Gambar 3.5 Petak contoh lahan pertanaman mentimun
Budidaya Tanaman Mentimun. Budidaya tanaman mentimun pada penanaman
pertama dan penanaman kedua dibedakan oleh adanya pengaplikasian pestisida pada penanaman pertama. Pada pertanaman pertama, 3 lahan pengamatan yaitu K1, K2, dan S1 diaplikasikan pestisida sementara lahan S2 tidak dilakukan penyemprotan pestisida. Adapun pemakaian pestisida ini dilakukan pada minggu keempat atau kelima setelah tanam. Pemakaian pupuk baik sintetis dan organik dilakukan dengan jumlah yang sama dikedua penanaman.
Pengambilan Contoh Serangga. Pengambilan contoh serangga dilakukan dengan
15 Persentase kejadian parasitisasi = x 100%
4
HASIL
4.1 Keanekaragaman dan Kelimpahan Hymenoptera Parasitika pada Pertanaman Mentimun
Keanekaragaman Hymenoptera parasitika yang diperoleh pada penelitian ini berjumlah 233 spesies yang terdiri dari 28 famili dan 2 077 individu. Di lanskap kompleks ditemukan 233 spesies dan 1 382 individu dari 28 famili, sedangkan pada lanskap sederhana ditemukan 23 famili dari 157 spesies, dan 695 individu (Tabel 4.1; Lampiran 1). Kekayaan spesies Hymenoptera parasitika yang diperoleh lebih tinggi di lahan mentimun dengan lanskap kompleks dibandingkan dengan lanskap sederhana. Namun hasil analisis menunjukkan bahwa keanekaragaman Hymenoptera parasitika dari kedua lanskap (kompleks dan sederhana) tidak memiliki perbedaan. Sementara itu, kelimpahan Hymenoptera parasitika yang ditemukan dipengaruhi oleh struktur lanskap. Berdasarkan uji ANOVA diketahui bahwa perbedaan struktur lanskap pada keempat lokasi tidak memengaruhi jumlah spesies (F1.2= 1.657, P= 0.327), keanekaragaman/H’
(F1.2= 1.657, P= 0.327) dan kemerataan/E (F1.2= 6.36, P= 0.128) dari Hymenoptera
parasitika yang diperoleh (Tabel 4.1).
Tabel 4.1 Kekayaan spesies dan keanekaragaman Hymenoptera parasitika yang ditemukan pada lanskap kompleks (K1 dan K2) dan lanskap sederhana (S1 dan S2) di Kabupaten Bogor
* H’: Indeks keanekaragaman Shannon-Winner; E: Indeks kemerataan Shannon
17 Tabel 4.2 Jumlah individu, spesies dan famili Hymenoptera parasitika pada lanskap
kompleks dan sederhana
1K1-K2: lanskap kompleks; 2S1-S2: lanskap sederhana; 3S: jumlah spesies; 4N: jumlah individu
Gambar 4.1 Jumlah individu yang diperoleh berdasarkan perangkap
19 Tabel 4.3 Jumlah individu masing-masing famili Hymenoptera parasitika pada lanskap
kompleks dan sederhana dari penggunaan perangkap berbeda
Famili Perangkap Malaise Metode Transek Perangkap Kuning
Kompleks Sederhana Kompleks Sederhana Kompleks Sederhana
Aphelinidae 0 1 0 0 5 0
4.2 Perbedaan Komposisi Hymenoptera Parasitika pada Lanskap yang Berbeda
Komposisi dan kekayaan spesies Hymenoptera parasitika pada setiap lanskap tidak memiliki perbedaan (Tabel 4.2). Komposisi dan kekayaan spesies Hymenoptera parasitika (beta-diversity) dinilai lebih dapat menggambarkan pengaruh perbedaan struktur lanskap dibandingkan hanya berdasarkan jumlah spesies Hymenoptera parasitika (alpha-diversity). Salah satu analisis beta-diversity yang sering digunakan adalah analisis NMDS, yaitu untuk mengetahui hubungan keanekaragaman Hymnoptera parasitika yang terdapat pada lanskap yang berbeda di sekitar pertanaman mentimun. NMDS diperoleh dari analisis berdasarkan indeks Bray-Curtis. Untuk mengetahui tingkat perbedaan antar lanskap dilakukan uji lanjut dengan menggunakan analisis kemiripan (ANOSIM).
kelompok yang terlihat dalam analisis NMDS (Gambar 4.2). Perbedaan tersebut dikarenakan kompleksitas lanskap di sekitar pertanaman mentimun yang berbeda.
Gambar 4.2 NMDS dari komposisi parasitika berdasarkan indeks Bray-Curtis. Kode yang terdapat di dalam gambar menunjukkan area studi: K1 dan K2= Kompleks, S1 dan S2= Sederhana; angka 1 dan 2 setelah titik menunjukkan musim tanam
4.3 Hama, Parasitoid dan Tingkat Parasitisasi pada Pertanaman Mentimun
Berdasarkan hasil koleksi inang secara langsung (metode transek), hama yang menyerang langsung pada pertanaman mentimun baik menyerang organ daun, bunga, batang, dan buah adalah larva serangga dari Ordo Lepidoptera, yaitu Diaphania indica (Crambidae) dan Spodoptera litura (Noctuidae). Hama lainnya yang terlihat selama pengamatan dilapangan antara lain imago dari kumbang Chrysomelidae dan kutudaun (Aphid). Kumbang Chrysomelidae atau oleh petani disebut cacantal (bahasa Sunda) menyerang bagian daun sehingga membuat daun berlubang, sementara kutudaun terlihat menutupi permukaan daun dan batang secara bergerombol.
Semua serangga yang diperoleh di lapang kemudian dikelompokkan peranannya berdasarkan informasi yang diperoleh dari pustaka (CSIRO 2001, Goulet dan Huber 1993). Dari hasil tersebut tampak bahwa sebagian parasitoid adalah parasitoid pupa (29% dari total parasitoid yang ditemukan) diikuti parasitoid telur sbesar 25%. Selain itu ditemukan juga parasitoid larva (22%) dan parasitoid larva-pupa (22%) (Lampiran 2).
Secara keseluruhan tipe parasitoid yang ditemukan pada kedua lanskap tidak terlalu berbeda. Persentase tipe parasitoid tertinggi ditemukan pada parasitoid pupa (31% pada lanskap kompleks dan 26% pada lanskap sederhana) dan parasitoid telur (25% pada lanskap kompleks dan sederhana). Akan tetapi, pada lanskap kompleks tipe parasitoid larva-pupa lebih banyak (24%) ditemukan dari pada parasitoid larva (19%). Sebaliknya, pada lanskap sederhana parasitoid larva memiliki persentase ditemukan lebih tinggi (28%) dibandingkan parasitoid larva pupa (19%). Sementara itu, tipe parasitoid imago
21 dan parasitoid telur-larva sama-sama hanya ditemukan sekitar 1% pada kedua struktur lanskap yang berbeda dengan jumlah individu kurang dari 20 individu.
Struktur lanskap memengaruhi persentase parasitisasi parasitoid pada pertanaman mentimun meskipun dengan nilai yang tidak terlalu besar (Gambar 4.3). Di lanskap kompleks, kejadian parasitisasi lebih tinggi dengan persentase hampir 43% dimana sebanyak 27 individu terparasit dari 63 individu larva yang ditemukan. Sebanyak 22 individu larva terparasit dari 54 larva yang ditemukan pada lanskap sederhana dengan persentase parasitisasi sekitar 41%.
Gambar 4.3 Persentase parasitisasi pada lanskap kompleks dan sederhana
Selama pengambilan contoh serangga, larva inang yang dijumpai adalah D. indica dan S. litura. Dari 117 larva D. indica yang ditemukan, 45 individu diantaranya terparasit dan sisanya tidak terparasit (menjadi imago inang) atau mati. Terdapat 3 jenis parasitoid yang diketahui berasosiasi dengan D. indica pada penelitian ini, yaitu Apanteles taragamae, Apanteles claviatus, dan Elasmus sp.. Sementara itu, S. litura dijumpai dalam jumlah kecil dan berasosiasi dengan Microplitis sp..
Tabel 4.4 Persentase parasitisasi pada larva inang yang ditemukan pada pertanaman mentimun
Larva Parasitoid
A. taragamae A. claviatus Elasmus sp. Microplitis sp.
Diaphania indica 35.04 2.56 0.85 -
(n: 41; N: 117) (n: 3; N:117) (n:1; N:117) -
Spodoptera litura - - - 100
(n: 5; N: 5)
*n:jumlah inang terparasit, N: jumlah inang yang ditemukan
23 Tabel 4.6 Hiperparasitisasi Stictopisthus sp. terhadap A.taragamae pada inang D. indica
Lanskap tanaman Umur Jumlah larva
D. indica
Jumlah kokon
A. Taragamae
A. taragamae
yang muncul Stictopisthusyang muncul sp. parasitisasi Persentase
S1 2 MST 1 16 8 8 50 %
S2 2 MST 1 11 4 6 54.5 %
K1 2 MST 1 27 10 10 37 %
4.4 Pengaruh Struktur Lanskap terhadap Hymenoptera Parasitika
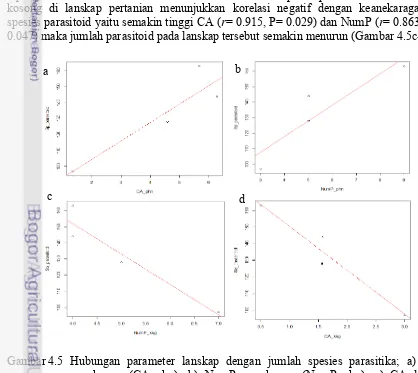
Hasil analisis hubungan parameter lanskap dengan keanekaragaman parasitoid menunjukkan bahwa parameter lanskap tertentu memengaruhi keanekaragaman parasitoid pada suatu lahan pertanian. Parameter lanskap yaitu class area (CA) dan jumlah patch (NumP) menunjukkan korelasi signifikan dengan keanekaragaman parasitoid (Gambar 4.5 a-b). CA dan NumP pepohonan di lanskap pertanian menunjukkan korelasi positif dengan keanekaragaman spesies parasitoid yaitu semakin tinggi CA (r= 0.744, P= 0.089) dan NumP (r= 0.743, P= 0.090) maka jumlah parasitoid pada lanskap tersebut juga semakin meningkat (Gambar 4.5a-b).
Lahan kosong adalah suatu areal yang tidak dimanfaatkan lagi sebagai lahan pertanian atau tidak ditanami tanaman pertanian, misalnya lahan sisa pertanian palawija, lapangan bola dan bekas tambak ikan. Parameter lanskap berupa CA dan NumP lahan kosong di lanskap pertanian menunjukkan korelasi negatif dengan keanekaragaman spesies parasitoid yaitu semakin tinggi CA (r= 0.915, P= 0.029) dan NumP (r= 0.863,P= 0.047) maka jumlah parasitoid pada lanskap tersebut semakin menurun (Gambar 4.5c-d).
Gambar 4.5 Hubungan parameter lanskap dengan jumlah spesies parasitika; a) CA pepohonan (CA_phn), b) NumP pepohonan (NumP_phn), c) CA lahan kosong (CA_ksg), dan d) NumP lahan kosong (NumP_ksg)
c d
5
PEMBAHASAN
Hasil dari penelitian ini menunjukkan bahwa struktur lanskap yang berbeda antara lanskap kompleks dan sederhana di sekitar pertanaman mentimun tidak memengaruhi kekayaan dan indeks keanekaragaman Hymenoptera parasitika yang diperoleh, namun memengaruhi kelimpahannya. Indeks keanekaragaman alpha dalam penelitian ini tidak menunjukkan perbedaaan, baik di lanskap kompleks maupun lanskap sederhana. Bahkan, keanekaragaman Hymenoptera parasitika yang diduga lebih tinggi di lanskap kompleks karena memiliki variasi vegetasi yang beragam juga tidak terlihat berbeda. Keanekaragaman disusun oleh beragamnya spesies yang ditemukan baik spesies yang menetap pada suatu lanskap ataupun spesies yang hanya melewati atau singgah di lanskap tersebut. Nilai kelimpahan spesies yang tinggi tidak selalu diikuti dengan keanekaragaman yang tinggi pula. Kelimpahan Hymenoptera parasitika tertinggi ditemukan pada lanskap kompleks dibandingkan lanskap sederhana. Hal ini dapat disebabkan oleh adanya vegetasi yang lebih beragam di sekitar tanaman mentimun pada lanskap kompleks. Vegetasi yang dimaksud disini adalah tanaman-tanaman non pertanian seperti pepohonan, semak, dan rerumputan yang terdapat di sekitar tanaman pertanian. Semakin beragamnya vegetasi disuatu lanskap kelimpahan Hymnoptera juga semakin tinggi dikarenakan sumber makanan yang dibutuhkan oleh Hymenoptera tersebut untuk berkembangbiak tercukupi. Hochberg dan Ives (2000) menyatakan bahwa keberadaan vegetasi di sekitar tanaman dapat meningkatkan populasi parasitoid, karena tersedia sumber makanan imago dan inang alternatif untuk parasitoid. Meskipun dalam penelitian ini kompleksitas lanskap tidak memengaruhi keanekaragaman Hymenoptera parasitika, namun beberapa penelitian lain, menunjukkan keanekaragaman cenderung tinggi pada lanskap yang kompleks (Marino dan Landis 1996, Plećaš et al. 2014).
Selain variasi vegetasi, lanskap kompleks juga dicirikan dengan penggunaan lahan yang beragam. Di lanskap sederhana, tipe penggunaan lahan cenderung didominasi oleh satu atau dua jenis penggunaan lahan seperti pertanian dan perumahan, sedangkan pada lanskap kompleks, selain pertanian tipe penggunaan lahan disertai dengan adanya habitat semi-alami atau hutan. Tanaman-tanaman non pertanian dan habitat semi alami tersebut berguna sebagai sumber daya seperti ketersediaan makanan, inang alternatif, tempat tinggal bahkan refugia bagi parasitoid yang aktif terbang dalam jarak yang cukup jauh seperti yang dipaparkan oleh Plećaš et al. (2014).
25 Tingginya perolehan individu dari famili Scelionidae, Diapriidae, dan Braconidae dikarenakan penyebaran spesies dari famili ini yang cukup merata dan menempati beragam tipe habitat. Di sekitar tanaman mentimun sendiri terdapat habitat yang cukup mendukung kehidupan ketiga famili ini. Sebagai contoh, famili Diapriidae yang memiliki variasi spesies yang banyak dan mempunyai kisaran inang yang luas, terutama memarasit lalat (Goulet dan Huber 1993). Famili ini ditemukan lebih banyak pada lanskap kompleks dibandingkan pada lanskap sederhana. Tingginya perolehan famili ini dapat dikarenakan adanya peternakan disekitar pertanaman mentimun baik di lanskap kompleks 1 (K1) maupun lanskap kompleks 2 (K2). Sementara famili Scelionidae diketahui memiliki jumlah spesies yang banyak dan memarasit telur dari banyak tipe seranggga dan laba-laba (Goulet dan Huber 1993). Famili Braconidae yang diketahui memarasit serangga hama dari ordo Lepidoptera seperti famili Pyralidae yang merupakan hama pada pertanaman mentimun. Ketiga famili ditemukan hampir merata baik dari lanskap kompleks maupun lanskap sederhana.
Pengaruh struktur lanskap juga terjadi pada famili atau spesies Hymenoptera parasitika yang ditemukan, dimana selain ketiga famili sebelumnya yang ditemukan lebih dominan, terdapat famili-famili yang ditemukan dalam jumlah kecil dan hanya pada tipe lanskap tertentu saja. Sebagai contoh famili Cynipidae, Gasteruptidae, Megaspilidae, dan Mymarommatidae hanya ditemukan pada lanskap kompleks. Hal ini dikarenakan lanskap kompleks memiliki variasi pepohonan dan tanaman lainnya seperti rerumputan yang berguna sebagai sumber makanan ataupun inang alternatif bagi parasitoid-parasitoid tersebut.
Dimensi pada NMDS menunjukkan bahwa semakin berdekatan titik maka komposisinya semakin mirip. Kelompok kemiripan komposisi spesies Hymenoptera parasitika antara lanskap kompleks terpisah dengan lanskap sederhana, yang berarti kemiripan diantara kedua lanskap ini cenderung berbeda, meskipun secara statistik (analisis ANOSIM) perbedaan tersebut tidak berbeda nyata. Perbedaan ini kemungkinan dikarenakan jumlah sampel serangga yang diperoleh terlalu sedikit sehingga tidak mewakili keadaan yang sebenarnya di lapangan. Selain itu, adanya kemungkinan perbedaan penyusun masing-masing lanskap yang berbeda sehingga memberikan pengaruh pada komposisi Hymenoptera parasitika di dalamnya. Menurut Tscharntke et al. (2007), keanekaragaman beta contohnya, komposisi spesies, cenderung tinggi pada lanskap kompleks. Meskipun demikian, pada suatu lanskap yang sama terjadi perbedaan kemiripan yang disebabkan perbedaan mikroklimat atau keadaan lingkungan yang dapat saja terjadi pada waktu yang berbeda meskipun berada pada satu lanskap yang sama. Hal ini dikarenakan serangga mengikuti atau beradaptasi pada kondisi lingkungan yang berbeda (Bianchi et al. 2006). Pada dasarnya tidak pernah ada lanskap yang benar-benar mirip atau sama secara keseluruhan.
kumbang daun Aulacophora similis (Coleoptera: Chrysomelidae), dan ulat daun D. indica.
Banyaknya jenis hama yang ditemukan pada tanaman mentimun diikuti dengan ditemukannya parasitoid yang berasosiasi dengan hama-hama pada pertanaman mentimun tersebut. Secara keseluruhan parasitoid yang dominan ditemukan adalah jenis parasitoid pupa, parasitoid telur, parasitoid larva dan parasitoid larva-pupa (Lampiran 2). Hal ini dapat dikarenakan tanaman-tanaman pertanian terutama tanaman sayuran dan palawija cenderung rentan terserang hama sehingga kelimpahan dari parasitoid jenis ini akan mengikuti kelimpahan hama di lahan tersebut. Semakin tinggi populasi hama, maka populasi musuh alami juga akan meningkat (Landis et al.2000).
Keefektifan parasitoid di lapangan tergantung pada keberhasilan parasitisasinya. Suatu parasitoid baru dapat dikatakan efektif dalam mengendalikan atau menekan populasi serangga hama apabila parasitisasinya berlangsung dengan baik. Hasil parasitisasi pada penelitian ini menunjukkan persentase parasitisasi sedikit lebih tinggi pada lanskap kompleks (43%) dibandingkan dengan lanskap sederhana (41%). Penentu tingginya parasitisasi pada lanskap kompleks dikarenakan kebutuhan parasitoid untuk memarasit seperti sumber nektar atau embun madu yang tersedia lebih banyak sebagai akibat dari beragamnya vegetasi di lanskap kompleks tersebut. Nektar dan embun madu dibutuhkan parasitoid dewasa untuk mematangkan sel-sel telur di dalam ovarinya dan juga untuk mendapatkan energi yang diperlukan pada saat mencari atau memarasit inang (Godfray 1994).
Asosiasi antara parasitoid dengan hama pada pertanaman mentimun dapat dilihat pada hubungan Apanteles taragamae dengan D. indica. Selain A. taragamae, terdapat juga parasitoid A. claviatus dan Elasmus sp. yang juga berasosiasi dengan D. indica. Namun dilihat dari persentase parasitisasi A. taragamae memiliki keberhasilan memarasit yang lebih tinggi (35.04%) dibandingkan dengan dua parasitoid lainnya. Sementara itu, S. litura yang ditemukan juga memakan daun-daun mentimun diketahui berasosiasi dengan Microplitis sp. dengan tingkat keberhasilan parasitisasi sebanyak 100%. Dengan demikian, keberadaan S. litura pada tanaman mentimun terkontrol dengan sangat baik.
Tingginya parasitisasi di pertanaman mentimun dapat disebabkan oleh pengaruh struktur lanskap dan umur tanaman mentimun itu sendiri. Seperti yang sering dipaparkan dalam beberapa penelitian, lanskap kompleks cenderung menaikkan tingkat parasitisasi, selain berkaitan dengan adanya sumber daya makanan yang tercukupi, lanskap kompleks juga menyediakan inang alternatif bagi parasitoid. Tanaman mentimun pada lanskap yang sama dengan umur tanaman berbeda mempunyai hasil parasitisasi yang berbeda pula. Semakin bertambahnya umur tanaman terjadi peningkatan jumlah dan volume bagian tanaman, maka serangan hama seperti D. indica juga ikut meningkat. Peningkatan populasi D. indica menarik parasitoidnya yaitu A. taragamae untuk memarasit dan meletakkan telur dalam jumlah banyak juga. Hal ini berdampak pada terkontrolnya populasi D. indica pada pertanaman mentimun sehingga sampai sejauh ini status D. indica belum menjadi hama utama pada pertanaman mentimun di Indonesia terutama di Kabupaten Bogor. Berbeda halnya di beberapa negara di Asia, Afrika dan Australia, D. indica dianggap sangat mengganggu dan menjadi hama penting pada pertanaman mentimun dan famili Cucurbitacae lainnya seprti melon dan semangka (Morgan dan Midmore 2002).
27 mencapai 50% dimana dari 16 kokon A. taragamae yang muncul, 8 kokon diantaranya terparasit. Hiperparasitisasi tertinggi terjadi pada lanskap sederhana (54.5%) sedangkan pada lanskap kompleks sebesar 37%. Hal ini mengindikasikan bahwa pada lanskap sederhana yang cenderung didominasi oleh tanaman-tanaman pertanian dengan kurangnya habitat alami dapat meningkatkan kejadian hiperparasitisasi. Dengan adanya hiperparasitisasi terhadap A. taragamae, maka pengendalian terhadap D. indica juga dapat berkurang. Dampaknya populasi ngengat ini di lapangan dapat meningkat apabila keadaan hiperparasitisasi ini terus berlangsung.
Manajemen lanskap menjadi lanskap kompleks cenderung lebih baik dibandingkan dengan lanskap sederhana. Ekosistem pertanian dengan lanskap sederhana berupa pertanian monokultur seperti pertanian padi berpotensi mengalami kerugian hasil yang lebih tinggi apabila diserang hama pada waktu yang sama. Pada lanskap pertanian monokultur, kelimpahan dan keanekaragaman parasitoid terbatas baik dikarenakan tidak adanya variasi inang ataupun ketersediaan sumber nektar bagi parasitoid dewasa. Sementara itu lanskap kompleks terbentuk dari pertanian polikultur yang terdiri dari berbagai habitat seperti habitat padi, sayur, tumbuhan liar, dan tanaman non pertanian akan terus menyuplai pasokan nutrisi dan inang yang berkelanjutan bagi musuh alami, dalam hal ini adalah parasitoid (Yaherwandi et al. 2008).
Tanaman-tanaman non-pertanian ini menyediakan sumber makanan berupa nektar bagi imago parasitoid, sehingga kemampuan imago parasitoid meningkat. Pepohonan atau tanaman non-pertanian yang terdapat pada suatu lanskap juga dapat berguna sebagai habitat bagi serangga herbivor, yang dapat dijadikan sebagai inang alternatif oleh parasitoid. Patch pepohonan di dalam lanskap menyediakan berbagai sumber daya seperti tempat tinggal, nektar, polen, embun madu, mangsa non-hama dan inang (Thies et al. 2003, Dauber et al. 2003, Rand et al. 2006). Lanskap dengan jumlah pepohonan yang banyak semakin menyerupai habitat alami sehingga semakin menggambarkan kompleksitas lanskap tersebut. Selain itu pepohonan dapat tumbuh dalam waktu yang lebih lama dibandingkan tanaman pertanian, semak ataupun rerumputan, sehingga ketersediaan sumber daya makanan dan inang dari pepohonan berkelanjutan bagi parasitoid. Altieri dan Nicholls (2004) menyatakan bahwa sumber nutrisi dan refugia bahkan inang alternatif pada pertanian yang terdapat patch pepohonan di sekitarnya dapat terus meningkat sepanjang waktu.
Manipulasi lanskap dengan menyediakan habitat pepohonan non-pertanian diketahui sebagai strategi efektif dalam konservasi biodiversitas dan juga efektif mengurangi efek buruk dari fragmentasi habitat atau akibat aktivitas manusia lainnya seperti pembukaan areal baru menjadi perkampungan. Woltz et al. (2014) mendapatkan hasil dimana kelimpahan Coccinellidae tinggi pada lahan kacang kedelai yang ditanami gandum, Fagopyrum esculentum, sebagai flower strips dibandingkan dengan kontrol, dan kelimpahan Coccinellidae di kacang kedelai berkorelasi positif dengan jumlah vegetasi semi-alami di lanskap tersebut. Hal yang sama juga dilaporkan Plećaš et al. (2014) dimana kelimpahan Aphid, rata-rata parasitisasi dan kekayaan spesies parasitoid dan hyperparasitoid lebih tinggi pada lanskap dengan habitat tanaman non-pertanian lebih luas dan beragam, positif mempengaruhi pengendalian biologis.
29
6
SIMPULAN DAN SARAN
6.1 Simpulan
Struktur lanskap tidak memengaruhi kekayaan dan indeks keanekaragaman Hymenoptera parasitika pada pertanaman mentimun, akan tetapi memengaruhi kelimpahannya. Meskipun secara statistik komposisi Hymnoptera parasitika pada pertanaman mentimun tidak dipengaruhi oleh struktur lanskap di sekitarnya, namun komposisi spesies memiliki kecenderungan berbeda berdasarkan analisis NMDS. Spesies Hymenoptera parasitika yang ditemukan paling banyak adalah Famili Diapriidae, Scelionidae, dan Braconidae. Struktur lanskap dan umur tanaman mentimun memengaruhi parasitisasi parasitoid, semakin kompleks dan bertambahnya umur tanaman mentimun maka semakin tinggi keberhasilan parasitisasi A. taragamae terhadap D. indica. Parasitoid Stictopisthus sp. adalah hiperparasitoid yang menyerang A. taragamae dengan tingkat parasitisasi lebih tinggi pada lanskap sederhana dibandingkan lanskap kompleks. Parameter struktur lanskap seperti keberadaan area pepohonan memiliki konstribusi dalam mendukung keberadaan parasitoid pada lahan pertanian. Semakin banyak area pepohonan di suatu lanskap, jumlah spesies parasitoid juga semakin meningkat. Sebaliknya lahan yang dibiarkan kosong menjadikan parasitoid di lahan pertanian berkurang.
6.2 Saran
DAFTAR PUSTAKA
[CSIRO] Commonwealth Scientific and Industrial Research Organisation. 2001. The insect of Australia. Australia: Melbourne University Press.
Altieri MA. 1999. The ecological role of biodiversity in agroecosystems. Agric. Ecosyst. Environ. 74:19-31.
Altieri MA, Nicholls CI. 2004. Biodiversity and Pest Management in Agroecosystems. Binghamton (US): Imprint Haworth Pr.
Baggen LR, Gurr Gm. 1998. The influence of food on Capidosoma koehleri (Hymenoptera: Encyrtidae), and the use of flowering plants as a habitat management tool to enhance biological control potato moth, Phthorimaea operculella (Lepidoptera: Gelechiidae). Biological Control. 11:9-17.
Bianchi FJJA, Booij CJH, Tscharntke T. 2006. Sustainable pest regulation inagricultural landscapes: a review on landscape composition, biodiversity and natural pest control. P Roy Soc Bio. 273:1715–1727. Doi: 10.1098/rspb.2006.3530
Clarke KR. 1993. Non-parametric multivariate analyses of change in community structure. Australian Journal of Ecology. 18: 117-143.
Clausen CP. 1940. Entomophagous Insects. New York (US): McGraw-Hill.
Corbett A, Rosenheim JA. 1996. Impact of a natural enemy overwintering refuge and its interaction with the surrounding landscape. Ecol Entomol. 21:155-164.
Costamagna AC, Menalled FD, Landis DA. 2004. Host density influences parasitism of the seminatural landscapes. Ecol. Lett. 14:101-112.
Dauber J, Hirsch M, Simmering D, Waldhardt R, Otte A, Wolters V. 2000. Landscape structure as an indicator of biodiversity: matrix effects on species richness. Agriculture Ecosystems & Environment. 98:321–329. doi:10.1016/S0167-8809(03)00092-6.
Debach P, Rosen P. 1991. Biological Control by Natural Enemies. Ney Work (US): Cambridge Univ Pr.
Driesche RG, Bellows TS. 1996. Biological Control. New York (US): Chapman & Hall. Dyer LE, Landis DA. 1996. Effects of habitat, temperature, and sugar availability on
longevity of Eriborus terebrans (Hymenoptera: Ichneumonidae). Environ Entomol. 25:1192–1201.
Dyer LE, Landis DA. 1997. Influence of noncrop habitats on the distribution of Eriborus terebrans (Hymenoptera: Ichneumonidae) in cornfields. Environ Entomol. 26:924–932.
Fiedler AK, Landis DA, Wratten SD. 2008. Maximizing ecosystem services from conservation biological control: the role of habitat management. Biological Control. 45:254-271.
Forman RTT. 1995. Some general principles of landscape and regional ecology. Landscape Ecology. 10:133–142.
Forman RTT, Godron M. 1986. Landscape ecology. Singapore (SG): J Willey.
Gámez-Virués S, Jonsson M, Ekbom B. 2012. Biodiversity and Insect Pest: Key issues for sustainable management. Gurr. GM, Wratten SD, Snyder WE, Read DMY, editor. West Sussex (UK): Wiley-Blackwell: J Wiley.
Godfray HCJ. 1994. Parasitoids: Behavioral and Evolutionary Ecology. Princetown, New Jersey (US): Princeton Univ Pr.
31 Griffiths GJK, Holland JM Bailey A, Thomas MB. 2008. Efficiacy and economics of shelter habitat for conservation biological control. Biological Control. 45:200-209.
Harper JL, Hawksworth DL. 1995. Bodiversity: Measurement and Estimation. Hawksworth DL, editor. London (UK): Chapman & Hall.
Heong KI, Aquino GB, dan Barrion AQ. 1991. Arthtopod community structure of rice ecosystem in Philiphines. Bull Entomol Research. 81:407-416.
Herlina N, Rizali A, Moerfiah, sahari B, Buchori D. 2011. Pengaruh habitat sekitar lahan persawahan dan umur tanaman padi terhadap keanekaragaman Hymenoptera parasitika. J. Entomol. Indon.8:17-26.
Heywood VH. 1995. Global Biodiverasity Assessment. Cambride (UK): Cambridge Univ Pr.
Hochberg ME & Ives AR. 2000. Parasitoid Population Biology. New Jersey (UK): Princeton University Press Princeton and Oxford.
Hunter MD. 2002. Landscape structure, habitat fragmentation, and the ecology of insects. Agricultural and Forest Entomology. 4:159–166.
Idrris AB, Grafius E. 1997. Nectar-collecting behavior of Deadegma insulare (Hymenoptera : Ichneumonidae), a parasitoid of diamondback moth (Lepidoptera: Plutellidae). Environmental Entomology. 26:114-120.
Kartosuwondo U. 1994. Populasi Pluetella xylostella (L.) (Lepidoptera: Yponomeutidae) dan parasitoid Diadegma semiclausum Hellen (Hymenoptera: Ichneumonidae) pada kubis dan dua jenis Brasicaceae liar. Bull. HPT. 7:39-49.
Kruess A, Tscharntke T. 1994. Habitat fragmentation, species loss, and biological control. Science. 264:1581–1584.
Kruess A, Tscharntke T. 2000. Species richness and parasitism in a fragmented landscape: experiments and field studies with insects on Vicia sepium. Oecologia. 122:129–137.
Landis DA, Wratten SD, Gurr GM. 2000. Habitat management to conserve natural enemies of Arthropod pests in agriculture. Annu. Rev. Entomol. 2000. 45:175-201. LaSalle J, Gauld ID. 1993. Hymenoptera and Biodiversity. LaSalle J, Gauld ID, editor.
London (UK): C.A.B. International.
Lavandero B, Wratten S, Shishehbor P, Worner S. 2005. Enhaching the effectiveness of the parasitoid Diadegma semiclausum (Helen): movement after use of nectar in the field. Biological Control. 34:152-158.
MacArthur RH, Wilson EO. 1967. The Theory of Island Biogeography. Princeton, New Jersey (US). Princeton Univ Pr.
Magurran AE. 2004. Measuring Biological Diversity. Malden (US). Blackwell Science. Marino PC, Landis DA. 1996. Effect of landscape structure on parasitoid diversity and
parasitism in agroecosystems. Ecol Society America. 6:276-284.
McGarigal K, Cushman SA, Ene E. 2012. FRAGSTATS v4: Spatial pattern analysis program for categorical and continuous maps. Dapat diakses pada web: http://www.umass.edu/landeco/research/fragstats/fragstats.html.
Menalled FD, Costamagna AC, Marino PC, Landis DA. 2003. Temporal variation in the response of parasitoids to agricultural landscape structure. Agri Ecosyst & Environ. 96:29-35.
Menalled FD, Marino PC, Gage SH, Landis DA. 1999. Does agricultural lanskap structure affect parasitism and parasitoid diversity? Ecol applications. 9:634-641.
Nixon GEJ. 1967. The Indo-Australian species of the ultorgroup of Apanteles Forster (Hymenoptera: Braconidae). Bull British Museum Entomology. 21:1-34.
Noyes JS. 1994. A study of five methods of sampling Hymenoptera (insecta) in a tropical rainforest, with special reference to the parasitica. J Nat Hist. 23:285-298.
Östman Ö, Ekbom B, Bengtsson J. 2001. Landscape heterogeneity and farming practice influence biological control. Basic and Applied Ecology. 2:365-371.
Odum EP. 1969. The strategy of ecosystem development. Science. 164, 267-270.
Oliveros JC. 2007. VENNY. An interactive tool for comparing lists with Venn Diagrams. http://bioinfogp.cnb.csic.es/tools/venny/index.html.
Peter C, David BV. 1990. Biology of Apanteles taragamae Viereck (Hymenoptera : Braconidae) a parasitoid of Diaphania indica (Saunders) (Lepidoptera: Pyralidae). Insect Sci. Applic. Vol. 13:7-17.
Plećaš M, Gagić V, Janković M, Petrović-Obradović O, Kavallieratos NG, Tomanović Ž, Thies C, Tscharntke T, Ćetković A. 2014. Landscape composition and configuration influence cerealaphid–parasitika–hyperparasitika interactions and biological control differentially across years. Agric Ecosyst & Environ. 183:1-10. Power AG. 2010. Ecosystem services and agriculture: tradeoffs and synergies. Phil Trans
R Soc. 365: 2959-2971.
Price PW, Waldbauer GP. 1975. Ecological Aspects of Pest Management: in Introduction to Insect Pest Management. New York (US): Wiley Interscience.
Purtauf T, Roschewitz I, Dauber J, Thies C, Tscharntke T, Wolters V. 2005. Landscape context of organic and conventional farms: influences on carabid beetle diversity. Agric Ecosyst & Environ. 108: 165-174.
Quantum GIS Development Team. 2014. Quantum GIS Geographic Information System. Open Source Geospatial Foundation Project.
R Development Core Team. 2012. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna.
Rand T, Tylianakis JM, Tscharntke T. 2006. Spillover edge effect: the dispersal of agriculturally subsidized insect natural enemies into adjacent natural habitat. Ecol letters. 9:603-614.
Roschewitz I, Hücker M, Tscharntke T, Thies C. 2005. The influence of landscapecontext and farming practices on parasitism of cereal aphids. Agric Ecosyst & Environ. 108:218–227.
Rubiana R. 2014. Pengaruh transformasi habitat terhadap keanekaragaman dan struktur komunitas semut di Jambi [tesis]. Bogor (ID): Institut Pertanian Bogor.
Scherber C, Lavandero B, Meyer KM, Perovic D, Visser U, Wiegand K, Tscharntke T. 2012. Biodiversity and Insect Pests: Key Issues for Sustainable Management. Gurr. GM, Wratten SD, Snyder WE, Read DMY, editor. West Sussex (UK): Wiley-Blackwell: J Wiley.
Schmidt MH, Thies C, Tscharntke T. 2005. Differential effects of landscape and management on diversity and density of ground-dwelling farmland spiders. J Appl Ecol. 42:281-287.
Thies C, Roschewitz I, Tscharntke T, 2005. The landscape context of cereal aphid– parasitoid interactions. P RoySoc Bio. 272:203–210.
Thies C, Steffan-Dewenter I, Tscharntke T. 2003. Effect of landscape context on herbivory and parasitism at different spatial scales. Oikos. 101:18-25.