RUMEN SEBAGAI BASIS FORMULASI RANSUM TERNAK RUMINANSIA
DENGAN BAHAN LOKAL
H E R M O N
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Indeks Sinkronisasi Pelepasan
N-Protein dan Energi sebagai Basis Formulasi Ransum Ternak Ruminansia dengan
Bahan Lokal adalah karya saya dengan arahan dari komisi pembimbing dan belum
diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi
yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari
penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di
bagian akhir disertasi ini
Bogor,
Januari
2009
Hermon
ABSTRACT
HERMON. Synchrony Index of Releasing N-Protein and Energy in the Rumen as a
Basis of Ruminant Diet Formulation with Local Feedstuffs.
Under direction of SURYAHADI, KOMANG G. WIRYAWAN, and
SOEDARMADI H.
The experiments were conducted to prove the diet formulation technique which
was based on the synchronization of releasing N-protein and energy in addition to
energy and protein requrement in diet for efficiency of rumen microbial N synthesis
and animal production.
Exp. 1. Nylon bag technique was adopted to determine ruminal characteristic of
protein and organic matter (OM) degradation of feedstuffs (forage and concentrate
diet) for which the synchrony index of N-protein and OM fermented in the rumen
might be determined based on synchronization ratio of 20 (I
20), 25 (I
25), and 30 g
N/kg OM (I30) fermented in the rumen. Exp. 2. By using randomized block design,
twelve local cattle were arranged into four groups according to average body weight
of the animals. Each group was fed three types of diet that was different in
synchronization ratio of supplying N-protein and fermented OM in rumen, namely
20 g N/kg OM (R20); 25 g N /kg OM (R25); and 30 g N /kg MO (R30). The diets had
iso-energy and iso-protein contents and had the same synchrony index. Exp 3. By
using randomized block design with a factorial of 3 x 2. The first factor was three
diferent protein levels i.e. 10, 12, and 14 %; the second factor was two deferent levels
of TDN (
total digestible nutrients
) (65 and 70 %). Eighteen local cattles were
arranged into three groups according to average body weight of the animals. Each
group was fed six types of diet that was different in level of CP or TDN. The diets
had the same synchrony index, namely 0,560.
The results of exp.1; Feedstuff of grass and concentrate of energy sources had
high synchrony index in I
20whereas legumes and concentrate of protein sources had
high synchrony index in I
30. Exp. 2 showed that the type of diet had no effect
(P>0.05) on intake and digestibility of nutrients, N retention, average daily gain
(ADG), and feed efficiency. But R20
had higher allantoin concentration in the urine
than that of R25 and R30 (P<0.01). Exp. 3 showed that except the crude fat digestion,
there was no interaction between protein and energy on the other parameters
(P>0.05). Compared to diet with 70% TDN, diet with 65% TDN produced higher
rumen microbial N, consumption and digestion of nutrients, and N retention
(P<0.05). Diet with protein level of 12% tend to have better allantoin concentration
in the urine, consumption and digestion of nutrients, N retention and
blood urea
nitrogen
(BUN) than that of either 10% or 14%. It can be concluded that the diet
having 65% TDN and 12% protein with synchrony index of 0.560 in the
synchronization ratio of 20 g N/kg OM fermented in the rumen will generate more
efficient N synthesis of rumen microbes and average daily gain of local cattle.
RINGKASAN
HERMON. Indeks Sinkronisasi Pelepasan N-Protein dan Energi dalam Rumen
sebagai Basis FormulasiRansum Ternak Ruminansia dengan Bahan Lokal.
Dibimbing oleh SURYAHADI, KOMANG G. WIRYAWAN,dan SOEDARMADI H
Berdasarkan asumsi para ahli dari negara subtropis bahwa nisbah yang optimal
sinkronisasi degradasi protein dan bahan organik (BO) sebesar 25 g N/kg BO
terfermentasi dalam rumen akan dihasilkan efisiensi sintesis N mikroba rumen, atau
dikategorikan mempunyai nilai indeks sinkronisasi 1. Perbedaan jenis dan komposisi
kimia bahan pakan dengan di Indonesia dan di daerah tropis lainnya diduga akan
berbeda pula tingkat dan laju degradasi protein dan BO selanjutnya akan berbeda pula
nisbah sinkronisasi kedua nutrien tersebut. Penggunaan indeks sinkronisasi selain
kebutuhan energi dan protein dalam memformulasi ransum ternak ruminasia
diharapkan akan dihasilkan efisiensi sintesis N mikroba dan produksi ternak yang
optimal.
Penelitian ini dilakukan tiga tahap. Tahap I, Melalui tehnik
in sacco
ditentukan
karakteristik degradasi protein dan BO bahan pakan kelompok hijauan rumput,
leguminosa, konsentrat sumber energi, dan konsentrat sumber protein. Selanjutnya
dapat dihitung “ indeks sinkronisasi” bahan pakan tersebut pada nisbah 20 g N/kg BO
(I20), 25 g N/kg BO (I25), dan pada nisbah 30 g N/kg BO (I30). Tahap II, Sesuai
dengan rancangan acak kelompok (RAK), tiga macam ransum yang berbeda nisbah
sinkronisasinya yakni 20 g N/kg BO (R20), 25 g N/kg BO (R25), dan 30 g N/kg BO
(R
30) yang disusun dari bahan pakan yang sama tetapi berbeda indeks sinkronisasinya
pada I
20, I
25, dan I
30.Ketiga ransum tersebut diberikan kepada empat kelompok sapi
lokal dan dilihat pengaruhnya. Tahap III; Menggunakan RAK pola faktorial 3 x 2.
Faktor pertama adalah protein yang terdiri dari tiga level protein yakni 10, 12, dan 14
%, dan faktor kedua adalah
total didestible nutrients
(TDN) dengan dua level TDN
yakni 65 dan 70%. Dengan demikian diperoleh enam macam ransum yang berbeda
level protein atau TDN satu sama lainnya. Keenam ransum diberikan kepada tiga
kelompok sapi lokal dan dilihat pengaruhnya. Indeks sinkronisasi ransum tersebut
relatif sama (0,560) pada nisbah sinkronisasi 20 g N/BO (nisbah optimal hasil
penelitian tahap II).
ransum (10, 12, dan 14%) berpengaruh tidak nyata (P>0.05) terhadap parameter
tersebut, tetapi ransum dengan 12% protein cenderung menghasilkan kadar allantoin
dalam urin, kon-sumsi dan kecernaan nutrien, retensi N, dan
blood urea nitrogen
(BUN)
terbaik.
Ransum dengan 12 % protein dan 65 % TDN serta indek sinkronisasinya 0.560
pada nisbah sinkronisasi 20 g N/kg BO menghasilkan efisiensi sintesis N mikroba
dan pertambahan bobot badan sapi lokal yang optimal.
© Hak Cipta milik IPB, tahun 2008
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan
pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan,
penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak
merugikan kepentingan yang wajar IPB
INDEKS SINKRONISASI PELEPASAN N-PROTEIN DAN ENERGI DALAM
RUMEN SEBAGAI BASIS FORMULASI RANSUM TERNAK RUMINASIA
DENGAN BAHAN LOKAL
H E R M O N
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Ilmu Ternak
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji diluar komisi pembimbing :
1.
Penguji pada ujian tertutup : Prof.Dr.Ir. Toto Toharmat, MAgr.Sc
2.
Penguji pada ujian terbuka : Dr.Ir. Kartiarso, M.Sc
Judul Disertasi : Indeks Sinkronisasi Pelepasan N-Protein dan Energi dalam
Rumen sebagai Basis Formulasi Ransum Ternak Ruminansia
Dengan Bahan Lokal
Nama : Hermon
NIM : D061050061
Disetujui
Komisi Pembimbing
Dr.Ir. Suryahadi, DEA.
Ketua
Dr.Ir. Komang G. Wiryawan Prof. Dr.Ir. Soedarmadi H, MSc.
Anggota Anggota
Mengetahui
Ketua Departemen
Ilmu Nutrisi dan Teknologi Pakan
Dekan Sekolah Pascasarjana
Dr.Ir. Idat G. Permana, MSc.Agr Prof. Dr.Ir. Khairil A. Notodiputro, M.S.
PRAKATA
Puji syukur penulis panjatkan kehadirat Allah SWT atas segala rahmat dan
karuniaNya sehingga penelitian dan karya ilmiah ini berhasil diselesaikan sebagai
persyaratan meraih gelar doktor pada Program Studi Ilmu Ternak di Sekolah
Pas-casarjana Institut Pertanian Bogor. Tema yang dipilih dalam penelitian ini adalah
Indeks Sinkronisasi Pelepasan N-Protein dan Energi dalam Rumen sebagai Basis
Formulasi Ransum Ternak Ruminansia dengan Bahan Lokal; yang dilaksanakan
sejak bulan Desember 2006 sampai dengan 28 Januari 2008.
Terimakasih disampaikan kepada Bapak Rektor Universitas Andalas dan Bapak
Dekan Fakultas Peternakan Universitas Andalas yang telah memberikan izin tugas
belajar kepada penulis. Terima kasih dan penghargaan yang sebesar-besarnya penulis
sampaikan kepada Bapak Dr. Ir. Suryahadi, DEA. selaku ketua komisi pembimbing,
Bapak Dr. Ir. Komang G. Wiryawan dan Bapak Prof. Dr. Soedarmadi H, MSc. selaku
anggota komisi pembimbing yang dengan tulus ikhlas dan penuh keakraban dalam
membimbing dan memberi petunjuk kepada penulis.
Terimakasih dan penghargaan penulis sampaikan kepada yang terhormat Rektor
Institut Pertanian Bogor, Dekan Sekolah Pascasarjana dan Ketua Program Studi Ilmu
Ternak yang telah berkenan menerima penulis untuk mengikuti pro-gram S3 di
Institut Pertanian Bogor. Demikian pula kepada Pengelola Beasiswa Program
Pascasarjana di Institut Pertanian Bogor dan Direktorat Jenderal Pendi-dikan Tinggi,
Departemen Pendidikan Nasional yang telah berkenan memberikan beasiswa.
Terimakasih penulis sampaikan kepada para Dosen yang telah membe-rikan ilmu
yang berarti dalam memperluas wawasan keilmuan, kepada teknisi dan karyawan di
lingkungan IPB yang telah membantu sehingga proses belajar dan penelitian berjalan
lancar. Demikian pula kepada para mahasiswa dan tehnisi La-boratorium Nutrisi
Ternak Ruminansia Fakultas Peternakan Universitas Andalas yang banyak membantu
penulis dilapangan.
Tak lupa penulis ucapkan terimakasih kepada istriku tercinta Afriyullatiza dan
anaku tersayang Wanda Primaria SSi, Runi Rahmawati dan Hana Raissya yang telah
banyak berkorbandan selalu memberikan dukungan doa, semoga pe-ngorbanan
kalian menjadi pemacu untuk mencapai kebahagiaan kita bersama dunia dan akhirat.
Kepada yang terhormat ayahanda H.Yoebhar St. Rajo Endah (alm) dan ibunda
tercinta Hj.Lainar, dengan tetesan air mata penulis haturkan banyak terimakasih atas
didikan dan doa yang selalu engkau panjatkan serta selalu membukakan pintu maaf.
Penulis menyadari betapa selama ini belum banyak memberikan balas budi yang telah
engkau berikan selama ini, hanya ucapan doa yang tulus yang bisa penulis berikan,
semoga amal dan budi baik ayahanda dan ibunda tercinta diterima oleh Alloh SWT.
Penulis berharap semoga amal dan budi baik dari semua pihak yang telah
diberikan kepada penulis mendapat balasan dan pahala yang berlimbah dari Alloh
SWT, amiin.
Akhir kata semoga karya ilmiah ini dapat bermanfaat bagi pemerhati dan
praktisi peternakan.
Bogor, Januari 2009
RIWAYAT HIDUP
Penulis dilahirkan di Kota Bandung Jawa Barat pada tanggal 24 Juli 1957.
Merupakan anak ke enam dari sepuluh bersaudara. Ayah bernama H. Yoebhar St.
Rajo Endah (alm) dan Ibu bernama Hj. Lainar.
Penulis menyelesaikan pendidikan dasar di SD Negeri Banjarsari Bandung pada
tahun 1970, pendidikan menengah pertama di SMP Negeri II Bandung pada tahun
1973, dan sekolah menengah atas di SMA Negeri IV Bandung pada tahun 1976.
Penulis mendapatkan gelar Sarjana Peternakan (Ir) pada tahun 1983 di Fakultas
Peternakan Universitas Padjadjaran. Pada tahun 1988 penulis mengikuti pendidikan
pascasarjana di Obihiro University of Agriculture and Veterinary Medicine – Obihiro
Japan, dan memperoleh gelar
Master of Agiculture
(M.Agr) pada tahun 1990.
Melalui ikatan dinas khusus (TIDK) sejak tahun 1984 penulis diterima bekerja
sebagai dosen pada Fakultas Peternakan Universitas Andalas Padang, dan terhitung
semenjak tahun 2005 penulis mengikuti program Doktor pada Program Studi Ilmu
Ternak – Sekolah Pascasarjana Institut Pertanian Bogor.
Penulis menikah dengan Afriyullatiza pada tahun 1984 dan dikaruniai tiga
orang putri Wanda Primaria SSi, Runi Rahmawati, dan Hana Raissya.
Bogor, Januari 2009
Penulis,
DAFTAR ISI
Halaman
DAFTAR TABEL ... xiii
DAFTAR GAMBAR... xiv
DAFTAR LAMPIRAN... xv
PENDAHULUAN
Latar Belakang ... 1
Tujuan Penelitian ... 4
Manfaat Penelitian ... 4
TINJAUAN PUSTAKA
Sintesis N Mikroba dalam Rumen ... 5
Metabolisme Protein dalam Rumen ... 10
Nitrogen yang Dibutuhkan Mikroba Rumen ... 13
Kebutuhan Protein dan Energi Mikroba Rumen ... 15
Efisiensi Sintesis N Mikroba Rumen ... 18
Sinkronisasi Pelepasan N-Protein dan Energi Makanan dalam rumen.. 20
Retensi Nitrogen ... 23
Pertambahan Bobot Badan (PBB) ... 25
Kebutuhan Energi dan Protein Ternak Sapi ... 25
Sapi Pesisir di Sumatera Barat ... 28
BAHAN DAN METODE
Tahap I : Penentuan Koefisien dan Indeks Sinkronisasi Degradasi Protein
dan BO Bahan Pakan dalam Rumen ... 30
Bahan Pakan ... 30
Hewan Percobaan ... 31
Prosedur Inkubasi ... 31
Tahap II : Penentuan Nisbah Optimum Sinkronisasi Degradasi protein
dan BO dalam Rumen ... 33
Tahap III : Kandungan Energi dan Protein dalam Ransum Berbasis Sinkronisasi
Degradasi Protein dan BO Ransum dalam Rumen Sapi Lokal ... 35
Waktu dan Tempat Penelitian ... 41
HASIL DAN PEMBAHASAN
Penentuan Koefisien dan Indeks Sinkronisasi Degradasi Protein
dan BO Bahan Pakan dalam Rumen
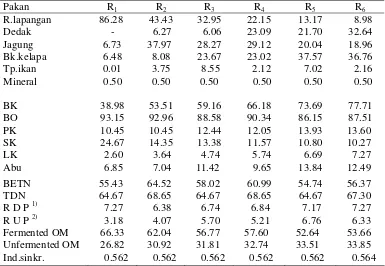
Koefisien Degradasi Protein dan BO Bahan Pakan dalam Rumen ... 42
Penentuan Nisbah Optimum Sinkronisasi Degradasi protein
dan BO dalam Rumen
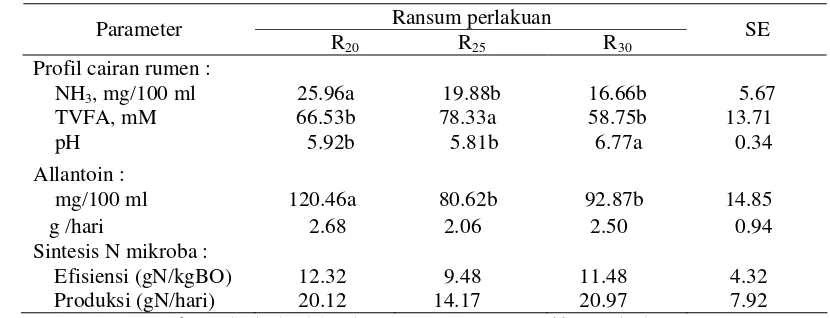
Profil Cairan Rumen dan Sintesis N Mikroba Rumen ... 49
Konsumsi dan Kecernaan serta Pertambahan Bobot Badan ... 53
Kandungan Energi dan Protein dalam Ransum Berbasis Sinkronisasi
Degradasi Protein dan BO Ransum dalam Rumen Sapi Lokal
Profil Cairan Rumen ... 55
Sintesis N Mikroba Rumen ... 59
Konsumsi Nutrien ... 61
Kecernaan Nutrien ... 63
Pertumbuhan Sapi ... 65
PEMBAHASAN UMUM ... 69
SIMPULAN DAN SARAN ... 73
DAFTAR PUSTAKA ... 74
DAFTAR TABEL
Halaman
1
Pengaruh
dilution rate
terhadap produksi N dan efisiensi mikroba ... 9
2
Rataan bobot badan sapi Pesisir Sumatera Barat (kg) ... 28
3
Komposisi kimia (%) bahan pakan hijauan dan konsentrat yang
dievaluasi secara
in sacco
... 31
4
Komposisi kimia (%) dan indeks sinkronisasi bahan pakan
ransum penelitian ... 34
5
Komposisi pakan dan kimia (%) serta indeks sinkronisasi
ransum penelitian ... 34
6
Komposisi pakan dan kimia (%) serta indeks sinkronisasi
ransum penelitian ... 36
7
Rataan koefisien degradasi protein dan bahan organik
pakan dalam rumen ... 42
8
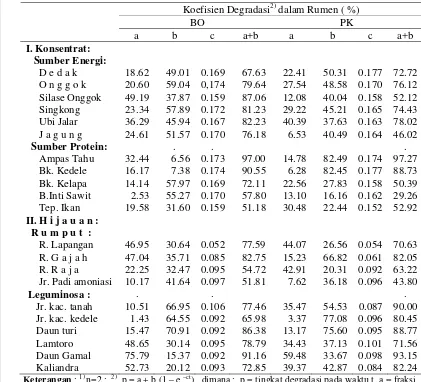
Rataan indeks sinkronisasi dan penyediaan g N/kg BO terfermentasi
per jam bahan pakan dalam rumen ... 46
9
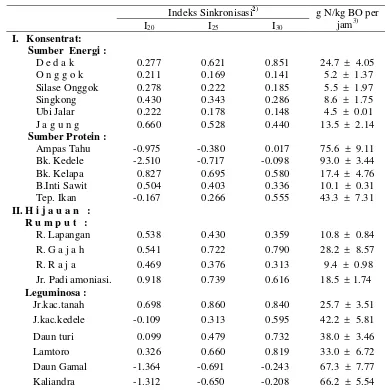
Profil cairan rumen dan allantoin urin sapi yang diberi ransum dengan
berbagai nisbah sinkronisasi degradasi protein dan BO dalam rumen ... 49
10
Konsumsi dan kecernaan nutrien serta pertumbuhan sapi yang diberi
ransum dengan berbagai nisbah sinkronisasi degradasi protein dan
BO dalam rumen ... 53
11
Profil cairan rumen sapi yang diberi ransum yang sinkron dengan
berbagai kandungan energi dan protein ... 56
12
Kadar allantoin dan sintesis N mikroba rumen sapi yang diberi ransum
yang sinkron dengan berbagai kandungan energi dan protein ... 60
13
Konsumsi nutrien ransum yang sinkron dengan berbagai kandungan
energi dan protein ... 61
14
Kecernaan nutrien ransum yang sinkron dengan berbagai kandungan
energi dan protein ... 64
15
Retensi N, PBB, efisiensi ransum, dan PER ransum yang sinkron
DAFTAR GAMBAR
Halaman
1
Ilustrasi populer mengenai saling ketergantungan fermentasi dan
produksi protein mikroba rumen ... 8
2 Proteolisis dalam rumen dan proses produk akhir fermentasi ... 12
3 Sapi Pesisir betina berfistula rumen ... 28
4 Bagan alir tahap penelitian ... 30
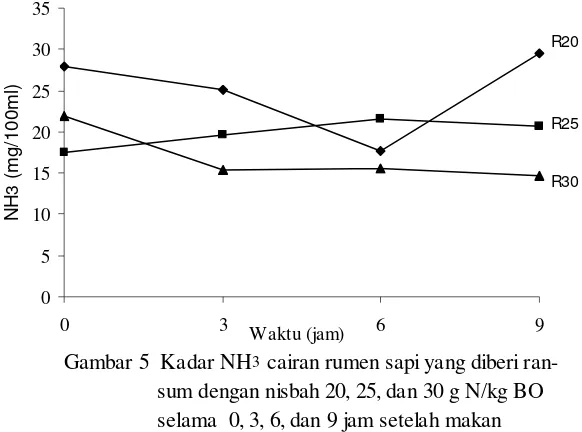
5 Kadar NH
3cairan rumen sapi yang diberi ransum dengan nisbah
20, 25, dan 30 g N/kg BO selama 0, 3, 6, dan 9 jam setelah makan ... 50
6 Kadar VFA cairan rumen sapi yang diberi ransum dengan nisbah
20, 25, dan 30 g N/kg BO selama 0, 3, 6, dan 9 jam setelah makan ... 51
7 pH cairan rumen sapi yang diberi ransum dengan nisbah
20, 25, dan 30 g N/kg BO selama 0, 3, 6, dan 9 jam setelah makan ... 52
8 NH
3cairan rumen sapi dengan ransum R1 = 10% PK, 65% TDN;
R2 = 10%PK, 70%TDN; R3 = 12%PK, 65%TDN; R4 = 12%PK,
70%TDN; R5 = 14%PK, 65%TDN; R6 = 14%PK, 70%TDN ... 56
9 VFA cairan rumen sapi dengan ransum R1 = 10% PK, 65% TDN;
R2 = 10%PK, 70%TDN; R3 = 12%PK, 65%TDN; R4 = 12%PK,
70%TDN; R5 = 14%PK, 65%TDN; R6 = 14%PK, 70%TDN ... 58
DAFTAR LAMPIRAN
Halaman
1 Pemasangan canula pada rumen sapi dan memasukan sampel pakan... 82
2 Korelasi antara sintesis N mikroba rumen dan konsumsi/kecernaan
PENDAHULUAN
Latar Belakang
Berbeda dengan ternak berlambung tunggal (hewan monogastrik), sumber energi
utama dan protein bagi ternak ruminansia berturut-turut berasal dari asam-asam
lemak terbang (
volatile fatty acids
atau disingkat VFA) dan protein yang berasal dari
protein mikroba rumen dan protein pakan yang lolos degradasi mikroba rumen
(
bypass protein
) atau RUP (
rumen undegraded protein
). Peran mikroba rumen
sangat penting yakni selain sebagai penyedia protein (protein mikroba) juga sebagai
penye-dia energi utama bagi ternak ruminansia berupa VFA terutama hasil
fermentasi kar-bohidrat, khususnya serat kasar yang tidak dapat dicerna ternak
ruminansia. Pada fer-mentasi karbohidrat atau bahan organik (BO) pakan akan
dilepaskan energi dalam bentuk ATP dan ATP ini oleh mikroba rumen digunakan
untuk membentuk protein mikroba dengan menggunakan N-protein yang tersedia
terutama dalam bentuk N-protein
hasil degradasi protein pakan (
rumen degradable
protein/
RDP) dan atau dari urea saliva di dalam rumen. Sebagai kerangka karbonnya
berupa asam alfa keto hasil fermentasi karbohidrat, sementara kerangka karbon
bercabang adalah hasil degradasi protein pakan.
Penyediaan N-protein yang lebih cepat daripada penyediaan energi menyebabkan
kelebihan N-protein
akan diangkut via pembuluh darah portal ke hati dibentuk urea
sebagian sebagai bahan siklus ke dalam saluran pencernaan dan sebagian lagi
dibu-ang melalui urin. Hal ini juga akan menurunkan konsumsi pakan ataupun merugikan
terutama bila protein yang terdegradasi tersebut berasal dari bahan pakan yang
ber-kualitas dan mahal harganya. Demikian pula bila energi yang dilepaskan lebih
ba-nyak daripada N-protein yang tersedia akan menyebabkan penurunan efisiensi
per-tumbuhan mikroba dan pencernaan serat kasar yang selanjutnya menurunkan
kon-sumsi.
laju penyediaan N-protein dan energi yang dihasilkan pakan untuk mikroba rumen
ditujukan untuk memaksimalkan pemanfaatan protein yang terdegradasi dalam
rumen, untuk pertumbuhan mikroba rumen yang optimal dan efisiensi pakan.
Sedikit perhatian dalam memformulasi ransum berdasarkan kepada laju
penyediaan N-protein dan energi yang diekspresikan sebagai BO atau karbohidrat
terfermentasi di dalam rumen (Sinclair
et al.
1993). Pada umumnya dalam
penyusunan ransum untuk ternak ruminansia didasarkan kebutuhan akan protein
kasar (PK)/protein dapat dicerna (Prdd) dan energi dalam bentuk TDN (total
digestible nutrients), tanpa mempertimbangkan kebutuhan yang efektif bagi
per-tumbuhan mikroba dalam rumen. Sehingga kadang kala dijumpai perbedaan yang sig-
nifikan penampilan produksi diantara ternak ruminansia percobaan, walaupun
ran-sum masing-masing ternak tersebut disusun relatif sama kandungan protein dan
energi (iso-protein dan iso-energi).
Nilai yang tersedia dalam rumen adalah gabungan dan integrasi daripada
rate of
degradation
(laju degradasi) ,
lag time
dan
rate of passage
(laju makanan) (Orskov
dan McDonald 1979). Keseimbangan laju penyediaan N-protein
hasil degradasi
protein dan laju penyediaan energi hasil degradasi BO atau karbohidrat suatu
makanan dalam rumen, ditujukan agar jumlah gram N-protein yang tersedia setiap
kg BO yang terdegradasi dalam rumen setiap jam (hourly degradation) adalah
sinkron dengan efisiensi sintesis N mikroba yang optimal, yakni 25 g N-protein /kg
BO terdegradasi dalam rumen (Sinclair
et al.
1993). Penentuan indeks sinkronisasi
suatu pakan yakni dengan membandingkan penyediaan per jam N-protein dan energi
pakan tersebut dengan nilai efisiensi sintesis N mikroba tersebut. Bila penyediaan
N-protein/kg BO terfermentasi dalam rumen suatu pakan adalah sama dengan nilai
efisiensi protein mikroba (25g N-protein/BO terfermentasi dalam rumen) maka
indeks sinkronisasi pakan tersebut adalah 1 (Sinclair
et al.
1993).
terfermentasi dalam rumen. Dengan demikian kemungkinan besar nisbah sinkronisasi
penyediaan N-protein dan energi untuk efisiensi sintesis N mikroba rumen akan
berbeda antara daerah tropis dan sub tropis, terutama karena berbedanya sumber
protein makanan yang selanjutnya akan berbeda pula tingkat dan laju degradasi
protein. Blumel
et al
. (2003) melaporkan, bahwa sumber dan karakteristik fermentasi
protein dan karbohidrat dapat mempengaruhi efisiensi sintesis N mikroba rumen,
dengan demikian menolak anggapan bahwa efisiensi sintesis N mikroba rumen
biasanya konstan.
Dengan memasukkan variabel sinkronisasi ini disamping variabel kebutuhan zat
pakan lain misalnya protein kasar (PK) dan
total digestible nutriens
(TDN) dalam
menyusun ransum, diharapkan dicapai pemanfaatan protein pakan yang optimal
khususnya untuk sintesis N mikroba rumen sehingga produksi ternak ruminansia
dapat ditingkatkan.
Nilai indeks sinkronisasi tersebut di atas didapat setelah diketahui nilai koefisien
(karakteristik) degradasi protein dan BO bahan pakan dalam rumen. Penentuan
karakteristik degradasi protein dan BO bahan pakan ini diperoleh melalui evaluasi
bahan pakan tersebut dengan metode
in sacco
(Orskov dan McDonald 1979), yaitu
menggunakan hewan yang berfistula rumen. Dengan demikian hasil metode evaluasi
pakan secara
in sacco
ini dapat diaplikasikan dalam cara memformulasi ransum yang
sinkron penyediaan N-protein dan energi untuk efisiensi sintesis N mikroba rumen
dan efisiensi ransum.
Sumber protein pada ternak ruminansia tidak semata berasal dari protein mikroba
saja tetapi juga berasal dari protein makanan yang lolos degradasi dalam rumen (
by-
pass protein
). Ternak yang mempunyai pertumbuhan yang cepat atau berproduksi
tinggi sangat memerlukan
bypass protein
selain protein mikroba rumen yakni untuk
menunjang produksi ternak yang tinggi tersebut. Agar dicapai pertumbuhan protein
mikroba maupun produksi ternak yang optimal perlu tersedia protein dan energi
makanan yang optimal pula untuk menunjang kebutuhan kedua hal tersebut.
produksi dari pakan dan gilirannya akan menguntungkan peternak. Pakan
non-konvensional ini adalah pakan yang berasal dari limbah/hasil ikutan (
waste/by-product
) agro-industri. Misalnya jerami padi, limbah onggok dan ikan asin afkir di
Sumatra Barat telah dimanfaatkan menjadi pakan ternak sapi walaupun belum
memasyarakat.
Tujuan Penelitian
1.
Mengklarifikasi cara penyusunan ransum dengan memperhitungkan sinkronisasi
penyediaan N-protein dan energi ransum untuk mikroba di dalam rumen selain
memperhitungkan kandungan protein dan energi yang dibutuhkan dalam ransum,
sehingga diperoleh efisiensi pertumbuhan mikroba rumen dan produksi ternak
ruminansia yang optimal serta menguntungkan.
2.
Mengetahui nilai indeks sinkronisasi bahan pakan hijauan dan konsentrat yang
yang dihitung berdasarkan koefisien degradasi protein dan BO bahan pakan
tersebut dalam rumen, kemudian nilai itu dijadikan basis penyusunan ransum
yang sinkron penyediaan N-protein
dan energi untuk efisiensi ransum dan
efisiensi sintesis mikroba dalam rumen.
Manfaat Penelitian
TINJAUAN PUSTAKA
Kajian terakhir di Eropa menunjukkan bahwa biaya tidak tetap (
variable cost
)
yang terbesar untuk produksi susu atau produksi daging ternak ruminansia adalah
biaya pakan yakni lebih dari 50% dan bahwa menyempitnya nisbah harga komoditi
tersebut dengan harga pakan sebagai akibat meningkatnya permintaan bahan pakan
ternak, maka hendaknya peternak perlu meningkatkan sistem penyusunan ransum
yang akurat sehingga akan sangat efisien penggunaan bahan pakan yang tersedia
(Cottrill 1998). Khampa dan Wanapat (2007) menyatakan, bahwa di daerah tropis
umumnya ternak ruminansia diberi makanan berupa hijauan berkualitas rendah,
jerami tanaman pertanian, hasil ikutan (
by-products
) industri yang pada dasarnya
mengandung materi ligno-selulosa yang tinggi, karbohidrat terfermentasi yang rendah
atau kualitas protein yang rendah. Disamping itu pada musim kemarau, tanah yang
tidak subur, dan sedikitnya makanan yang tersedia akan mempengaruhi kuantitas dan
kualitas fermentasi rumen. Atas dasar tersebut dapat dikatakan bahwa untuk
meningkatkan pendapatan peternak, yaitu selain perbaikan dalam penyusunan ransum
dari bahan pakan konvensional juga ditingkatkan lagi pemanfaatan
waste/by-product
agro-industri sebagai bahan pakan, misalnya onggok, limbah darah rumah potong
hewan (RPH) dan ikan asin afkir yang sampai saat ini di Sumatra Barat belum
dimanfaatkan secara luas.
Sintesis N Mikroba dalam Rumen
protein untuk ternak ruminan berdasarkan PK ransum adalah tidak cukup karena
ruminally degradable protein
(RDP) akan menentukan sintesis N mikroba yang mana
sebagai sumber protein utama bagi ternak ruminansia (Karsli dan Russell 2002).
Rumen telah dikenal sebagai tempat terjadinya proses fermentasi yang esensial yaitu
berkemampuan menyediakan hasil ahir berupa
volatile fatty acids
(VFA) dan protein
mikroba sebagai sumber energi dan protein ternak ruminansia itu sendiri (Fellner
2005; Khampa dan Wanapat 2007)
Sel mikroba dan protein makanan yang lolos dari degradasi dalam rumen
(
bypass protein
) adalah sumber utama protein dan asam amino yang dibutuhkan
ternak ruminansia. Protein mikroba menyumbang protein dalam jumlah besar masuk
ke dalam usus halus, karena protein mikroba dapat mensuplai 50% lebih dari
kebutuhan sapi perah, perlu memaksimalkan produksi mikroba (Fellner 2005).
Dengan demikian kondisi dalam rumen harus optimal untuk pertumbuhan mikroba
rumen apabila asam amino yang masuk ke dalam usus halus dan produksi susu serta
komponen air susunya maksimal (Hoover dan Stoke 1991). Sasaran daripada nutrien
dalam rumen adalah untuk memaksimalkan pertumbuhan mikroba rumen dan
pengikatan RDP kedalam sel mikroba rumen, memaksimalkan pengikatan RDP
tersebut tidak hanya memperbaiki penyediaan asam amino ke dalam usus halus tetapi
juga menurunkan N terbuang (Stern
et al.
2006). Dengan demikian pertimbangan
nutrien yang dibutuhkan mikroorganisma rumen adalah sangat penting untuk
dipa-hami metabolisma N dalam rumen demikian juga faktor-faktor yang
memodifika-sinya (Bach
et al.
2005).
Defisiensi suatu nutrien dapat menurunkan sintesis N mikroba dalam rumen,
aliran asam amino ke usus halus, dan menurunkan produksi ternak ruminansia, tetapi
dua faktor nutrisi yang sangat memungkinkan membatasi sintesis N mikroba rumen
adalah energi dan protein (Clark
et al.
1992).
utama untuk memanfaatkan N-protein dalam rumen (Shabi
et al.
1998). Karsli dan
Russel (2002) melaporkan bahwa sintesis N mikroba dan pertumbuhan mikroba
rumen bergantung padakecukupan energi (ATP) hasil fermentasi BO serta kecukupan
N-protein hasil degradasi NPN (non-protein nitrogen) dan protein pakan dalam
rumen, juga membutuhkan zat pakan lain seperti sulfur, phosphor dan mineral lain
serta vitamin.
Di negara berkembang makanan untuk produksi ternak ruminansia berasal dari
padang penggembalaan, jerami tanaman dan biomasa selulosa lainnya, makanan
tersebut umumnya rendah total protein dan N yang terfermentasi serta sering kali
defisien mineralnya (Leng 1997). Di daerah iklim tropis dan subtropis umumnya
dikaitkan dengan rendahmya kandungan mineral yakni P, S, K, Ca, Mg, Cu, Mo, Co,
Se (Kerridge
et al.
1986) dan tinggi Al atau keasamannya yang mana membatasi
pertumbuhan akar (Chen
et al
. 1992). Selanjutnya Leng (1997) menyatakan, bahwa
tanah daerah tropis biasanya defisiensi mineral P. Sementara hasil penelitian Scott
et
al.
(1994) menunjukkan, bahwa kekurangan mineral selain menurunkan sintesis N
mikroba adalah faktor utama penyebab penurunan laju pembentukan tulang pada
kambing yang diberi makan rendah P.
Defisiensi setiap nutrien dapat menurunkan sintesis N mikroba dalam rumen,
laju asam-asam amino (AA) ke dalam usus halus dan menurunkan produksi susu sapi
perah (Clark
et al.
1992). Stern dan Hoover (1979) melaporkan, bahwa sulfur
dibutuhkan mikroba rumen untuk sintesis metionin dan cytein dan konsumsi sulfur
dapat membatasi sintesis N mikroba bila memakai NPN dalam jumlah banyak, dan
diperkirakan nisbah yang optimum nitrogen : sulfur adalah 10 : 1 untuk pertumbuhan
mikroba yang maksimal.
Gambar 1 Ilustrasi populer mengenai saling ketergantungan
fermentasi dan produksi protein mikroba (Orskov 1982)
daripada protein. Energi berasal dari lemak makanan dapat dikonsumsi bakteri
rumen, tetapi lemak tidak cukup mepenyediaan energi untuk mesintesis N mikroba
(Van Soest 1982). Karbohidrat adalah komponen utama dalam makanan ternak
ruminasia dan sangat bervariasi tingkat dan laju fermentabilitasnya dalam rumen
(Fellner 2005). Selulosa dan hemiselulosa yang jumlahnya berkisar dari 15 – 66 %
dari makanan ternak ruminansia adalah
insoluble
dan sebagian yang tidak tersedia
(tidak terfermentasi), pektin walaupun bagian dari
structural carbohydrate
(dinding
sel) bersama dengan karbohidrat yang
soluble
(fruktosan dan sukrosa) difermentasi
secara keseluruhan dan cepat, sementara pati sebagai cadangan karbohidrat yang
insoluble
dan dapat lolos dari degradasi dalam rumen (NRC 1985).
Dewasa ini kajian mengenai keadaan N dan carbon dari AA dalam rumen
menunjukkan bahwa beberapa AA disintesis oleh mikroorganisma rumen dengan
sangat sulit dibandingkan dengan AA yang lain (Atasoglu
et al.
2004). Selanjutnya
mengkonfirmasi teori bahwa bakteri rumen sulit mesintesis Phe, Leu, dan Ile, dan
dinyatakan bahwa Lys adalah suatu AA yang potensial yang membatasi
pertum-buhan bakteri rumen.
Bach
et al
. (2005) melaporkan bahwa selain kecukupan penyediaan sumber
karbohidrat (CHO) dan N, faktor nutrisi lain maupun faktor non-nutrisi yakni pH
rumen dan
dilution rate,
termasuk
juga
mepunyai peranan penting dalam sintesis N
mikroba rumen. Bila jumlah bahan organik (BO) yang difermentasi meningkat,
sintesis N mikroba meningkat pula dan hasilnya adalah adanya korelasi yang negatif
antara pH dan sintesis N mikroba yaitu konsekuensi penyediaan energi yang
meningkat pada ransum yang tinggi terfermentasi (Hoover dan Stokes 1991). Selama
ini dianggap komponen penghasil energi merupakan hal sangat penting untuk
produksi mikroba rumen, tetapi hasil penelitian Gosselink
et al.
(2003) menunjukkan
bahwa meskipun dalam makanan terbatas jumlah karbohidrat yang tersedia untuk
difermentasi mikroba, nitrogen yang tersedia dalam rumen adalah sangat penting
dalam memprediksi produksi mikroba. Sesuai dengan yang dinyatakan Maeng
et al.
(1999) bahwa penyediaan nitrogen PK meningkatkan produksi dan efisiensi mikroba
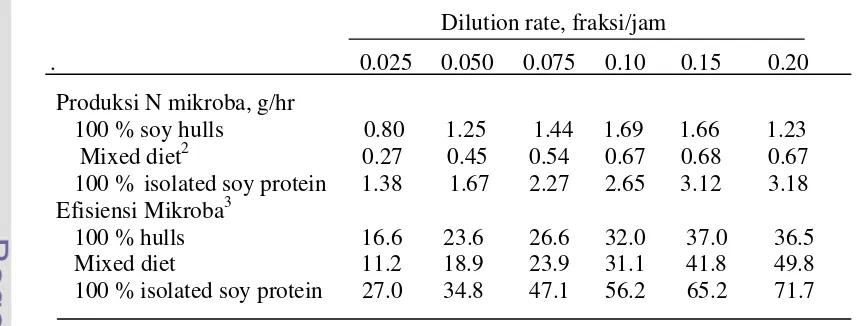
lebih tinggi dibandingkan penyediaan baik serat maupun pati (Tabel 1) dan kejadian
ini konsisten hasilnya pada peningkatan
dilution rate
rumen.
Pertumbuhan dan produksi mikroba biasanya meningkat dengan meningkatnya
dilution rate
,bila
dilution rate
rendah akan lebih banyak jumlah mikroba tinggal
da-lam rumen dan selanjutnya akan mekonsumsi energi yang tersedia untuk hidup pokok
Tabel 1 Pengaruh dilution rate terhadap produksi N dan efisiensi mikroba
1Dilution rate, fraksi/jam
. 0.025 0.050 0.075 0.10 0.15 0.20
Produksi N mikroba, g/hr
100 % soy hulls 0.80 1.25 1.44 1.69 1.66 1.23
Mixed diet
20.27
0.45
0.54 0.67 0.68 0.67
100 %
isolated soy protein 1.38 1.67
2.27 2.65
3.12
3.18
Efisiensi Mikroba
3100 % hulls
16.6 23.6
26.6 32.0 37.0
36.5
Mixed diet
11.2 18.9
23.9 31.1 41.8
49.8
100 % isolated soy protein
27.0 34.8 47.1 56.2 65.2 71.7
1)
Maeng
et al.
1990
2)
78% corn, 14% soy hulls, and 8% isolated soy protei
jam. Faktor yang mempengaruhinya adalah makanan, infus buffer intraruminal,
tingkat konsumsi, dan kondisi lingkungan (Stern dan Hoover 1979).
Perubahan makanan pada sapi jantan muda dari seluruhnya konsentrat menjadi
terkandung 14% hijauan di dalamnya terjadi peningkatan
dilution rate
(0.03 menjadi
0.05/jam) disertai peningkatan sintesis N mikroba rumen (7.5 menjadi 11.8 g/100 g
bahan kering terfermentasi dalam rumen) (Cole
et al.
1976). Stern dan Hoover
(1979) melaporkan, bahwa pemberian monensin kepada ternak domba terjadi
penurunan
dilution rate
dari 0.07 menjadi 0.04/jam disertai penurunan sintesis
mikroba dari 24.5 menjadi 20.2 g N/kg BO terfermentasi. Infus saliva buatan
kedalam rumen telah meningkatkan
dilution rate
dari 0.03 menjadi 0.08/jam,
bersamaan terjadi peningkatan sintesis total AA per mol heksosa yang difermentasi
yakni 25.4 menjadi 29.8 gram. Selanjutnya dilaporkan, bahwa pemeliharaan ternak
domba dari teperatur udara 18 menjadi 21
oC adalah kurang efisien dibandingkan
dengan yang dipelihara dari temperatur -1 menjadi 1
oC, yaitu dalam sintesis N
mikroba (47.9 vs 54.9 g N/kg BO terfermentasi).
Faktor adanya hubungan yang positif antara
dilution rate
dan sintesis N
mikroba, adalah menurunnya
autolysis
bakteria berkurang, menurunnya konsumsi
bakteria oleh protozoa, perubahan struktur populasi mikroba yang dipicu oleh
perubahan substrat atau kemungkinan karena pencucian oleh organisma yang waktu
perkembangannya lambat.
Metabolisme Protein dalam Rumen
Williams 1998; Gustafsson
et al.
2006). RUP dibutuhkan dalam jumlah besar dari
protein tambahan (feed supplement) pada makanan ternak ruminansia berproduksi
tinggi (Stern
et al.
2006).
Sementara sistem evaluasi protein dengan protein kasar (total crude protein) dan
digestible crude protein
(DCP) keduanya mempunyai kelemahan terutama terhadap
perhitungan transaksi nitrogen dalam rumen (Cottrill 1998). Dibandingkan dengan
sistem DCP, sistem MP mepunyai korelasi lebih erat dengan produksi dan juga
adanya kemungkinan yang lebih besar untuk mengkombinasikan bahan pakan agar
meminimalis surplus protein terdegradsi dalam rumen (Bertilsson
et al
. 1991).
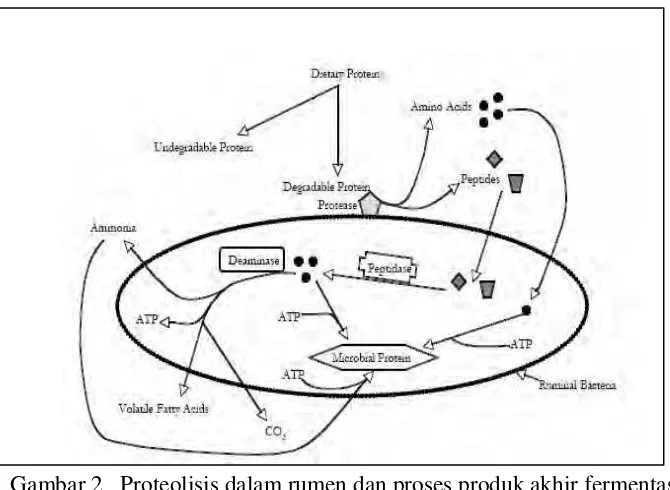
Metabolisma protein dalam rumen dapat dibagi menjadi 2 peristiwa yang
berbeda yaitu degradasi protein yang menyediakan sumber N untuk bakteria, dan
sintesis N mikroba (Baach
et al
. 2005). Selanjutnya dinyatakan metabolisme protein
dalam rumen ini adalah hasil aktifitas metabolisma daripada microorganisma rumen.
Tamminga (1979) menyatakan bahwa mekanisme pemecahan protein makanan
adalah sangat komplek dan belum dipahami secara menyeluruh. Degradasi protein
dalam rumen diawali menempelnya bakteria pada partikel makanan, diikuti oleh
aktifitas enzim protease mikroba (Brock
et al
. 1982). Sekitar 70-80% mikroba rumen
menempel pada partikel makanan dalam rumen (Craig
et al
. 1987), dan 30-50% nya
mempunyai aktivitas proteolitik (Prins
et al.
1983). Sejumlah besar spesies mikroba
yang berbeda berkongsi menempel pada partikel makanan untuk bekerja secara
simbiosis mendegradasi dan memfermentasi nutrien termasuk protein yang
menghasilkan AA dan peptida, karena sejumlah ikatan pada protein menyebabkan
aktivitas yang sinergis dari pada protease yang berbeda diperlukan untuk medegradasi
protein yang lengkap (Wallace
et al.
1997).
Gambar 2 Proteolisis dalam rumen dan proses produk akhir fermentasi
(Bach
et al.
2005)
terbatas, AA akan dideaminasi dan kerangka karbonnya difermentasi menjadi VFA
(Gambar 2) (Bach
et al.
2005) . Sebagian mikroba kurang baik mekanisma
pengang-kutan AA dari sitoplasma ke lingkungan ekstra-selular dan AA yang berlebihan ini
harus diekresi dari sitoplasma berupa ammonia (Tamminga 1979).
dari Lys-Pro dihidrolisis dalam rumen 5x lebih lambat daripada peptida Lys-Ala, dan
peptida yang tersusun dari Pro-Met didegradasi 2.5x lebih lambat daripada peptida
Met-Ala (Yang dan Russell 1992).
Lana
et al.
(1998) melaporkan, bahwa bakteri rumen yang mendapat makanan
100% hijauan dan pH rumen diturunkan dari 6.5 menjadi 5.7 menyebabkan
penurunan konsentrasi ammonia, sementara bakteri rumen pada sapi potong dengan
pakan 90% kosentrat mempunyai konsentrasi ammonia rendah dengan mengabaikan
pH. Assoumani
et al.
(1992) menyatakan, bahwa pati berperan terhadap degradasi
protein yakni dengan penambahan amilase telah meningkatkan degradasi protein.
Bach
et al
. (2005) menyatakan, bahwa protein tanaman banyak yang terikat pada
matrik serat yang perlu didegradasi terlebih dahulu sebelum protease mengakses
protein untuk didegradsi. Lebih lanjut menyatakan suatu hipotesis bahwa penurunan
bakteri selulolitik sebagai konsekuensi pH yang rendah menyebabkan penurunan
degradasi serat, menurunkan akses bakteri proteolitik terhadap protein yang tidak
langsung penurunan degradasi protein. Degradasi protein berhubungan terbalik
dengan laju makanan melalui rumen (Orskov dan McDonald 1979).
Penyebab rendahnya energi yang tersedia didaerah tropis menurut Khampa dan
Wanapat (2007), di daerah tropis umumnya ternak ruminansia diberi makanan
berupa hijauan berkualitas rendah, jerami tanaman pertanian, hasil ikutan (
by-products
) industri yang pada dasarnya mengandung materi ligno-selulosa yang tinggi,
karbohidrat terfermentasi yang rendah atau kualitas protein yang rendah.
Nitrogen yang Dibutuhkan Mikroba Rumen
diperoleh efisiensi sintesis N mikroba yang maksimal dalam rumen, penyediaan AA,
peptide dan nucleotida (DNA dan RNA) diperlukan juga (Jones
et al
. 1998;
Sanchez-Pozo dan Gil 2002; Block 2006).
Mikroba yang mendegradasi
structural carbohydrates
(dinding sel) (selulolitik)
mempunyai kebutuhan hidup pokoknya rendah, tumbuh lambat, dan menggunakan
N-protein
sebagai sumber N utamanya, sedangkan mikroorganisma yang
mendegradasi
non-structural carbohydrates
(karbohidrat isi sel) (amilolitik)
mempunyai kebutuhan hidup pokok yang tinggi, tumbuh cepat, dan menggunakan
N-protein, peptida, dan AA sebagai sumber N (Russell
et al.
1992). Sesuai menurut
Fellner (2005) bakteria yang memfermentasi
non-structural carbohydrates
(NSC)
menyukai asam amino dan peptida dan bakteri yang memfermentasi serat terutama
menyukai ammonia sebagai sumber nitrogen. Tetapi penambahan AA atau peptida
menunjukkan peningkatan pertumbuhan bakteri selulolitik dan amilolitik (Maeng dan
Baldwin 1976; Argyle dan Baldwin 1989). Hal yang sama bahwa kecernaan serat
oleh bakteri selulolitik meningkat dengan penambahan AA (Griswold
et al.
1996;
Carro dan Miller 1999), dan penambahan peptida (Crus Soto
et al
. 1994). Atasoglu
et al.
( 2001) melaporkan bahwa bakteri selulolitik lebih suka mengikat N-AA
daripada N-peptida ke dalam selnya, tetapi pada konsentrasi AA dan peptida tertentu
dalam rumen, sekitar 80% N selnya berasal dari N-protein.
Peningkatan pertumbuhan mikroba dengan penambahan AA dan atau peptida
dapat disebabkan bergabungnya secara langsung AA kedalam protein mikroba dan
atau disebabkan penyediaan kerangka carbon yang meningkat (hasil deaminasi AA)
yang mana dapat digunakan untuk produksi energi atau sebagai kerangka karbon
untuk AA mikroba yang baru (Bryant 1973). Atasoglu
et al
. (2004) menduga, bahwa
beberapa AA diantaranya lysin dapat membatasi pertumbuhan bakteri rumen.
Demeyer dan Fieves (2004) menduga bahwa konsentrasi AA dan peptida yang rendah
berpotensi membatasi pertumbuhan mikroba bila pemberian makan yang kaya pati
dengan partikel yang halus, yaitu menyebabkan pH rumen yang rendah.
Sementara pada penelitian lain melaporkan bahwa adanya hubungan yang negatif
antara konsentrasi N-protein dan persentase protein mikroba yang berasal dari NPN
(Siddon
et al
. 1985; Firkins
et al
. 1987). Bach
et al.
(2005) memperkirakan bahwa
akumulasi N-protein dalam rumen adalah hasil penggunaan pilihan peptida atau AA
oleh mikroba, baik sebagai sumber N atau sebagai sumber energi. Selanjutnya
dinyatakan bahwa proposi N bakteria yang berasal dari N-protein adalah bukan nilai
tetap. Pertumbuhan sel mikroba rumen memerlukan nucleotida baik yang disintesis
atau yang didapat (Block 2006). Nucleotida disintesis dari AA dapat digunakan
untuk pertumbuhan mikroba dengan mengonsumsi pada porsi yang cukup energi dan
nitrogen yang tersedia (Sanchez-Pozo dan Gil 2002).
Pemberian urea sebagai sumber N pada makanan yang semi bebas protein pada
domba meningkatkan konsentrasi N dari 0.95 menjadi 1.82% dan meningkatkan
produksi protein mikroba, tetapi tidak ada peningkatan sintesis N mikroba bila
konsentrasi N makanan meningkat sampai 3.2% (Hume
et al.
1970). Kemungkinan
mikroba rumen memperoleh N melalui pengikatan N dalam bentuk gas.
Pendikteksian fiksasi N secara
in vivo
dapat dilakukan dengan menggunakan isotop
15N (Moisio
et al
. 1969). Dengan menggunakan
acetylene-reduction screening test
,
Hardy
et al.
(1968) mengestimasi bahwa seekor sapi (steer) dapat mengikat 10 mg N
per hari.
Kebutuhan Protein dan Energi
Protein kasar,
nonstructural
dan
structural carbohydrates
adalah komponen
makanan yang potensial dapat dimanipulasi untuk mengoptimalkan fermentasi dalam
rumen dan untuk meningkatkan aliran AA ke usus halus (Clark
et al
. 1992).
Selanjutnya dinyatakan, ketersediaan energi dan N adalah menentukan sekali
terhadap jumlah protein mikroba yang disintesis dalam rumen. Manipulasi protein
yang tersedia dalam rumen kelihatannya sangat menguntungkan apabila sapi dalam
keadaan keseimbangan energi dan cukup lemak tubuh yang tersedia untuk kebutuhan
energi (Nocek dan Russell 1988).
mengindikasikan, bahwa konsentrasi PK makanan dimana pertumbuhan mikroba
yang maksimum terjadi pada ternak sapi sekitar dua persen lebih tinggi dari ternak
domba yang diberi makan yang hampir sama. Diperkirakan bahwa sintesis N
mikroba akan mencapai puncak bila makan sapi mengandung sekitar 12-13% PK,
kisaran ini tidak baku tetapi akan bervariasi menurut kandungan energi terfermentasi
makanan, jumlah NPN makanan, tingkat degradasi protein makanan, efisiensi
pertumbuhan mikroba rumen, dan input N saliva ke dalam rumen (Satter
et al
. 1977).
Selanjutnya dinyatakan bahwa penurunan protein yang tersedia dalam rumen
meningkatkan produksi susu bila konsumsi PK adalah marginal (<14% CP), tetapi
tidak ada respon bila PK lebih dari 16%.
Setelah dicapai kandungan PK optimal dalam ransum, peningkatan lebih lanjut
konsentrasi PK ransum tidak akan meningkatkan sintesis N mikroba tetapi dapat
meningkatkan total asam amino ke usus halus karena lolos dari degradasi protein
dalam rumen (Stern
et al
. 1978). Gustafsson
et al
. (2006) melaporkan, bahwa surplus
PK dalam ransum sapi perah dapat membebani lingkungan dan menurunkan
penampilan reproduksi (fertilitas), dan kerusakan akibat kelebihan N ini dapat
dikurangi melalui peningkatan pengikatan N yang terdegradasi dalam rumen.
Selanjutnya dinyatakan, bahwa protein yang berlebihan dalam ransum
mengakibatkan meningkatnya biaya energi secara khusus yakni kelebihan N
dikonversi menjadi urea dan diekresi dalam urin keduanya membutuhkan energi.
Pelepasan energi (ATP) hasil fermentasi terutama karbohidrat oleh mikroba
digunakan untuk memproduksi sel baru, enzim dsb. Tingkat energi yang dihasilkan
fermentasi tersebut dipengaruhi oleh komposisi makanan, karbohidrat yang berbeda
akan berbeda pula tingkkat dan laju fermentasinya (Gustafsson
et al
. 2006). Clark
et
al.
(1992) melaporkan, bahwa campuran
structural
dan
nonstructural carbohydrates
secara normal adalah sumber energi yang terbaik untuk pertumbuhan bakteri karena
pada proses fermentasi karbohidrat menghasilkan lebih banyak energi per satuan
berat daripada protein. Sedangkan energi yang tersedia dalam makanan berupa lemak
dapat dikonsumsi oleh bakteri rumen, tetapi tidak menyediaan energi untuk sintesis
protein (Van Soest 1982). Asam lemak bebas yang tinggi yang dihasilkan ketika
hidrolisis lemak akan menghambat pencernaan serat, kemungkinan melalui
coating
(menyelimuti) partikel makanan dan preventing baterial kontak (Orskov dan Ryle
1990).
Mikroba rumen yang memfermentasi
non-fiber carbohydrate
(NFC) (terutama
gula-gula yang mudah larut dan pektin) dilaporkan mempunyai kontribusi yang besar
terhadap sintesis N mikroba per satuan karbohidrat terfermentasi (Russell
et al.
1992). Chamberlain
et al.
(1993) mendapatkan, bahwa suplemen gula lebih efektif
dibandingkan pati pada ransum silase rumput untuk meningkatkan ekskresi derivat
purin dalam urin ternak domba, efekifitas karbohidrat secara berurutan adalah
sukrosa > laktosa > fruktosa > xylosa > pati gandum. Trevaskis
et al.
(2001)
melaporkan, bahwa infus sukrosa ke dalam rumen adalah efektif untuk mestimulir
pembentukan protein mikroba yaitu disinkronkan dengan konsentrasi amonia yang
mencapai puncak pada waktu 1-2 jam setelah makan. Penurunan ukuran partikel
makanan butiran meningkatkan pembentukan protein mikroba, bersamaan dengan itu
pH rumen tidak menurun (Broderick 2006).
karbo-hidrat yang mudah terfermentasi (
readily fermentable carbohydrates
) ke dalam
rumen akan meningkatkan pertumbuhan mikroba, tetapi penambahan energi melebihi
tingkat optimal tidak lagi meningkatkan pertumbuhan mikroba. Penurunan aliran
protein mikroba rumen ke dalam usus halus akibat ransum mengandung konsentrat
lebih dari 70% dan hal ini dapat terjadi karena laju degradasi yang cepat dari pada
karbohidrat nonstruktural, selanjutnya terjadi fermentasi
uncoupled.
Uncoupled fermentation
terjadi karena laju pelepasan energi lebih cepat
dibandingkan dengan laju pengikatan dan pemanfaatan oleh bakteri rumen (Karsli
dan Russell 2001). Leng (1997) menyatakan, bahwa
uncoupling
adalah reaksi
pelepasan energi dalam sel mikroba yang dapat juga menyebabkan pelepasan energi
ATP berupa panas tanpa adanya pembelahan sel dan pertumbuhan, hal ini terjadi bila
defisiensi mineral, amonia atau nutrien lain yang esensial untuk pertumbuhan
mikroba dan juga bila sel mikroba lisis dalam rumen.
Makanan yang tinggi
total digestible nutrients
(TDN) pada umumnya lebih
terfermentasi daripada yang rendah TDN, oleh karena itu banyak ammonia (NPN)
dapat dimanfaatkan dengan makanan yang tinggi TDNnya (Satter dan Roffler 1975).
Dalam hal ini dinyatakan bahwa peningkatan TDN merupakan “
opens the gates
” dan
memudahkan bayak ammonia yang digunakan untuk menunjang perkembangan
mikroba lebih tinggi. Tetapi yang tidak menguntungkan penggunaan TDN sebagai
indikator fermentabilitas yang jelek adalah bila ransum mengandung lemak atau
minyak yang tinggi .
Efisiensi Sintesis N Mikroba Rumen
menggunakan BO untuk membuat protein. Penentuan efisiensi mikroba penting
karena efisiensi ini adalah bagian daripada kalkulasi produksi mikroba (
mirobial
yield
) dan penentuan produksi mikroba ini penting karena merupakan suatu indeks
sejumlah protein mikroba yang tersedia bagi sapi setiap hari (Fellner 2005)
Faktor yang mempengaruhi sintesis N mikroba rumen adalah sumber dan
konsentarsi nitrogen dan karbohidrat,
dilution rate
rumen, sulfur makanan, dan
frekuensi pemberian makan (Stern dan Hoover 1979); konsumsi bahan kering (BK),
nisbah hijauan : konsentrat dalam ransum, laju degradasi protein dan karbohidrat,
sinkronisasi penyediaan N-protein dan energi, laju pakan, dan faktor lain yakni
vitamin dan mineral (Karsli dan Russell 2001). Konsumsi,
rumen turnover
dan
efisiensi mikroba adalah berkorelasi positif. Peningkatan konsumsi akan
meningkatkan
rumen turnover
selanjutnya menyebabkan peningkatan pertumbuhan
mikroba (Fellner 2005).
Peningkatan efisiensi sintesis N mikroba dicapai dengan peningkatan konsumsi
BK dan laju pakan, laju degradasi sumber protein dan karbohidrat yang sama-sama
lambat dan sebaliknya, sinkronisasi laju penyediaan N-protein dan karbohidrat yang
sama-sama lambat atau yang sama-sama cepat, campuran hijauan dan konsentrat
(Karsli dan Russell 2001). Lebih lanjut dinyatakan bahwa efisiensi sintesis N
mikroba yang maksimal diperoleh untuk sumber protein yang RDP nya tinggi
dibandingkan yang rendah, adanya sumber asam amino atau peptida selain pemakaian
NPN (urea) dalam ransum, dan pencampuran sumber karbohidrat yang struktural dan
non-struktural karbohidrat. Blumel
et al
. (2003) melaporkan, bahwa sumber dan
karakteristik fermentasi protein dan karbohidrat dapat mempengaruhi efisiensi
sintesis N mikroba rumen, dengan demikian menolak anggapan bahwa efisiensi
sintesis N mikroba rumen biasanya konstan.
Karsli dan Russell (2001) melaporkan, bahwa rata-rata efisiensi sintesis N mikroba
adalah 13.0 dengan kisaran 7.5 – 24.3 untuk makanan dasar hijauan (hasil 34 studi),
17.6 dengan kisaran 9.1 – 27.9 untuk makanan campuran hijauan dan konsentrat
(hasil 36 studi), dan 13.2 denga kisaran 7.0 -23.7 g protein mikroba/100 g BO
terfermentasi dalam rumen untuk makanan konsentrat (hasil 14 studi). Nocek dan
Russell (1988) melaporkan, bahwa makanan hijauan (
roughage
) menghasilkan
sintesis 45 g mikroba/kg karbohidrat tercerna dan hasilnya lebih rendah (29 g
mikroba/kg karbohidrat tercerna) bila digunakan makanan konsentrat.
Semakin tinggi efisiensi sintesis N mikroba semakin besar kontribusi protein
mikroba tersebut terhadap kebutuhan protein total ternak sapi, kontribusinya lebih
tinggi pada sapi potong dibandingkan sapi perah karena kemungkinan kurang
meningkatnya konsumsi BK pada sapi potong sebagai akibat ADG (average daily
gain) yang meningkat (Stern
et al
. 2006). Selanjutnya dinyatakan, bahwa efisiensi
sintesis N mikroba rumen pada sapi potong lebih rendah daripada sapi perah
kemungkinan besar disebabkan populasi bakteria amilolitik yang tinggi.
Penambahan AA
branced-chain
(rantai cabang) akan difermentasi menjadi FVA
rantai cabang, dan penambahan peptida ke dalam rumen telah meningkatkan
kecernaan serat , produksi protein mikroba , dan efisiensi pertumbuhan mikroba
(Russell dan Sniffen 1984; Thomsen 1985).
Sinkronisasi Pelepasan N-protein dan Energi Makanan dalam Rumen
Sinclair
et al.
(1993) menyatakan bahwa formulasi ransum menggunakan
koefisien degradasi protein dan BO bahan pakan yaitu membuat ransum yang sinkron
dalam hal pelepasan N-protein
dan energi per jam (hourly ) untuk fraksi mikroba
dalam rumen, dimana nisbah yang optimal degradasi protein dan BO yang
terfermentasi dalam rumen adalah 25 g N-protein/kg BO (Czerkawski 1986 diacu
Sinclair
et al
. (1993); atau 32 g N-protein/kg karbohidrat (Sinclair
et al
. 1991).
Selanjutnya dinyatakan, bahwa ransum yang sinkron tersebut bertujuan untuk
memaksimalkan sintesis N mikroba dari RDP yang tersedia, menurunkan kebutuhan
akan sumber RUP yang mahal harganya dan juga menurunkan ekskresi N-protein
urin. Ekresi N-protein
melalui urin sebagai akibat kelebihan protein ransum
memer-lukan energi (energy cost) untuk pembentukan urea sebelum dibuang melalui urin
(Gustafsson
et al
. 2006).
Untuk memperdalam konsep sinkronisasi, mikroba rumen membutuhkan sumber
N-protein, energi, mineral, vitamin dan faktor-faktor pertumbuhan untuk
pertumbuh-annya. Tetapi N-protein dan energi dibutuhkan dalam jumlah yang besar dan harus
tersedia secara simultan untuk merangsang pertumbuhan yang cepat (Huber dan
Herrera-Saldana 1994). Kebutuhan terhadap kedua komponen (energi dan N-protein)
tersebut pada aktifitas mikroba rumen tercermin adanya korelasi parsial yang erat (r
= 0.95, P < 0.01) antara degradasi protein dan BO ransum dalam rumen (Hermon
dan Warly, 2001); demikian juga pada silase rumput, yaitu antara degradasi protein
dan bahan kering serta degradasi serat kasar dalam rumen (0.69 vs 0.42 ; P < 0.01)
(Hermon 1999).
dengan kebutuhan untuk pertumbuhan mikroba, sinkronisasi tidak terjadi dan
merupakan suatu
assosiative effect
yang negatif.
Richardson
et al.
(2003) melaporkan bahwa dewasa ini sistem formulasi ransum
berdasarkan sinkronisasi penyediaan N-protein
dan energi dalam rumen per jam
menunjukan adanya keuntungan terhadap peningkatan produksi dan efisiensi
mikro-ba rumen serta terhadap produksi ternak, tetapi hasil peneliti lain menunjukkan
pe-ngaruh yang tidak signifikan terhadap pertumbuhan mikroba atau sangat dipepe-ngaruhi
oleh energi ransum daripada sinkronisasi. Hasil penelitian Richardson
et al.
(2003)
menunjukkan bahwa baik ransum sinkron maupun sumber energi berpengaruh tidak
nyata terhadap laju pertumbuhan kambing. Walaupun demikian ransum tidak sinkron
menyebabkan efisiensi penggunaan energi ransum rendah, dan upaya formulasi
ransum yang sinkron penyediaan zat pakan dalam rumen dapat meningkatkan
efisiensi energi pada kambing muda.
Pengaruh ransum yang sinkron penyediaan N-protein dan energi untuk mikroba
rumen terhadap penampilan ternak adalah tidak konsisten (Bach
et al
. 2005).
Beberapa kajian
in vivo
(Casper dan Schingoethe 1989; Herrera-Saldana
et al.
1990;
Matras
et al.
1991) menunjukkan respon yang positif terhadap penampilan ternak
pada ransum yang sinkron, sementara penelitian
batch culture
(Henning
et al.
1991;
Newbold dan Rust 1992) menunjukkan tidak ada pengaruhnya. Interpretasi kedua
macam kajian tersebut sulit dilakukan karena laju penyediaan energi dan protein
sering kali dikacaukan dengan total penyediaan energi dan protein (Bach
et al.
2005).
Lebih lanjut dinyatakan bahwa konsep sinkronisasi protein dan energi berbasis
teoritis semata, hal ini kemungkinan pada ekosistim yang komplek daripada
campuran mikroba rumen ketika disinkronkan penyediaan nutrien untuk
sub-populasi yang spesifik tetapi kemungkinan tidak sinkron untuk sub-populasi lainnya.
selanjutnya mengakibatkan pH lebih rendah (Satter dan Baumgardt 1962) dan sintesis
mikroba menurun (Robinson 1989). Hasil penelitian Biricik
et al.
(2006), sinkronisasi
degradasi pati dan protein dalam rumen tidak berpengaruh terhadap konsumsi dan
daya cerna nutrien dalam rumen dan seluruh saluran pencernaan pada ternak domba.
Broderick (2006) melaporkan hasil penelitian yang menunjukkan sedikit atau tidak
ada kenutungan produksi hasil manipulasi sinkronisasi degradasi protein dan
fer-mentasi energi dalam rumen, tetapi penelitian tersebut tidak mencoba untuk
memi-nimalkkan konsumsi PK. Selanjutnya dinyatakan, bahwa pemberian makan yang
rendah PK akan membatasi RDP untuk pembentukan protein mikroba dan dapat
diupayakan melalui sinkronisasi fermentasi energi dengan penyediaan N dalam
rumen.
Retensi Nitrogen
Retensi nitrogen merupakan salah satu metode untuk menilai suatu kualitas
protein ransum dengan jalan mengukur konsumsi nitrogen dan pengeluaran nitrogen
ekskreta, sehingga dapat diketahui banyaknya nitrogen yang tertinggal dalam tubuh
(Lioyd
et al.
1978). Pengukuran neraca nitrogen menurut Tillman dkk. (1991)
dilakukan dengan menghitung selisih antara jumlah nitrogen yang dikonsumsi dengan
jumlah nitrogen yang keluar melalui feses dan urin, sehingga dapat diketahui jumlah
nitrogen yang dapat tertinggal dalam tubuh. Selanjutnya dinyatakan, bahwa neraca
nitrogen dapat dirumuskan dengan persamaan :
B = I – (U + F) , dimana :
B = neraca nitrogen
I = konsumsi nitrogen
U = nitrogen urin
F = nitrogen feses
artinya nitrogen yang dikonsumsi hanya cukup untuk kebutuhan hidup pokok saja.
Tetapi bila neraca nitrogen bernilai negatif, ini berarti terjadi suatu kehilangan
nitrogen jaringan melalui metabolisme sebagai akibat nitrogen makanan yang
dikonsumsi tidak mencukupi kebutuhan hidup pokok ternak yang bersangkutan.
Menurut Orskov (1982) faktor-faktor yang mempengaruhi deposit protein pada
bobot hidup yang relatif sama, adalah 1) Genotype; yaitu laju deposit protein pada
sapi dengan bobot hidup relatif sama akan berbeda antara bangsa sapi ukuran kecil
dan ukuran besar; 2) Jenis kelamin; pertambahan bobot hidup pada ternak
ruminansia dengan bobot dan genotype yang relatif sama, pada sapi betina lebih
banyak lemak dan sedikit protein dibandingkan sapi jantan; 3) Pengaruh nutrisi
sebelumnya; pada bangsa dan jenis kelamin yang sama kebutuhan protein dikaitkan
dengan bobot hidup dapat bervariasi sekali akibat nutrisi sebelumnya apakah cukup
atau kekurangan protein.; 4) Level nutrisi; peningkatan konsumsi energi akan
meningkatkan retensi protein sampai suplai protein menjadi pembatasnya.
Peningkatan retensi protein ini bergantung padafase dewasa ternak, jenis kelamin,
dan genotype.
Bani
et al.
(1991) melaporkan bahwa level urea darah (
blood urea
) pada
ternak ruminansia umumnya diduga sebagai ekspresi dari amonia rumen, dengan
demikian urea darah yang sangat tinggi dapat merupakan suatu indeks kelebihan
protein dan resiko yang potensial terhadap kesuburan yang rendah (
low fertility
).
Selanjutnya berdasarkan hasil penelitiannya dinyatakan bahwa penyerapan amonia
rumen dan glukoneogenesis dari absorbsi asam-asam amino menyebabkan
kan-dungan urea darah bervariasi, tetapi glukoneogenesis nampaknya sangat berperan
terutama ketika pemberian protein relatif tinggi. Kandungan
Blood urea nitrogen
(BUN) atau amonia plasma yang tinggi kemungkinan bertalian dengan perubahan
fisiologi ovary dan uterus yang mengakibatkan tidak cukupnya
luteal
dan kematian
embrio (Sinclair
et al
. 2000).
Pertambahan Bobot Badan (PBB)
Ukuran komersil untuk pertumbuhan ternak adalah pertambahan bobot badan,
yakni sebagai penimbunan energi dalam karkas. Kuantitas penimbunan energi
dalam satuan berat ini bergantung pada komposisi kimia pertambahan bobot badan,
yakni air, tulang, protein (daging), dan lemak. Energi yang terkandung pada
pertambahan bobot badan hampir sepenuhnya diwujudkan oleh protein dan lemak,
dengan demikian komposisi pertambahan bobot badan tidak konstan sepanjang fase
pertumbuhan (Davies 1982).
Kenaikan berat badan dengan mudah dilakukan dengan penimbangan yang
berulang-ulang untuk menentukan pertambahan bobot badan tiap hari, tiap minggu,
atau tiap waktu tertentu lainnya (Tillman dkk. 1991). Maynard
et al.
(1969)
menyatakan bahwa semakin baik kualitas makanan, semakin efisien pembentukan
energi dan semakin cepat proses penambahan bobot badan. Selanjutnya dinyatakan
bahwa nutrien utama yang dibutuhkan ternak adalah energi dan protein, kelebihan
energi dan protein dari kebutuhan hidup pokok digunakan untuk pertumbuhan atau
pertambahan bobot badan. Demikian pula menurut Van Soest (1982) bahwa
meningkatnya konsumsi protein dan energi akan meningkatkan pertambahan bobot
badan, dan tidak seluruh energi yang tersedia dalam makanan dapat dimanfaatkan
oleh ternak karena sangat erat hubungannya dengan kapasitas dan kemampuan alat
pencernaan terhadap kualitas bahan makanan.
Tillman dkk. (1991) menyatakan bahwa faktor yang mempengaruhi
pertumbuhan adalah makanan, konsumsi pakan, total protein yang dikonsumsi tiap
hari, jenis atau bangsa ternak dan manajemen ternak. Menurut Davies (1982)
umumnya pertambahan bobot badan pada ternak muda mempunyai kandungan
energi yang rendah karena mengandung air dan tulang yang tinggi dengan sedikit
lemak, sedangkan pertambahan bobot badan ternak dewasa sangat tinggi kandungan
energi yang direfleksikan dengan proporsi lemak yang tinggi.
Kebutuhan Energi dan Protein Ternak Sapi
mempertahankan kesehatan tubuh dan untuk memelihara temperatur tubuhnya tanpa
penambahan atau kehilangan bobot badan, sementara kebutuhan untuk produksi
adalah nutrien yang dibutuhkan ternak selain untuk hidup pokok juga untuk proses
produksi diantaranya pertumbuhan, penggemukan, dan pembentukan air susu (Nelson
1979). Selanjutnya dinyatakan bahwa dua nutrien utama untuk kedua hal tersebut,
yaitu suplai energi dan prortein. Nutrien yang dibutukan ternak adalah energi, protein,
mineral, dan vitamin (Davies 1982).
Kebutuhan energi untuk hidup pokok bergantung pada ukuran atau bobot
tubuh, bangsa sapi, temperatur udara dan aktifitas. Kebutuhan ini erat hubungannya
dengan fungsi bobot tubuh atau dikenal bobot metabolis, yakni W
0,73. Kebutuhan
energi untuk hidup pokok di daerah tropis (temperatur udara panas) dan yang
dikandangkan adalah lebih rendah dibandingkan di daerah
temperate
(udara dingin)
dan yang tidak dikandangkan (Davies 1982). Selanjutnya dinyatakan bahwa jumlah
protein yang dibutuhkan bergantung padakecernaannya dan efisiensi penggunaannya
dalam tubuh; pada ternak ruminansia, proporsi rumen degradable protein (RDP) dan
rumen undegradable protein (RUP) adalah penting ketika laju pertambahan bobot
badan yang cepat.
Kebutuhan energi didasarkan kepada bobot tubuh, pertambahan bobot badan
(PBB), jenis kelamin dan kondisi tubuh. Sapi tipe besar atau mempunyai PBB yang
tinggi atau sapi jantan kebutuhannya lebih tinggi dibandingkan dengan sapi tipe kecil,
PBB yang rendah, sapi betina pada periode yang sama (NRC 1984). Laju
pertumbuhan terutama ditentukan oleh konsumsi energi, pada laju pertumbuhan
tersebut dibutuhkan berbagai asam amino untuk berlangsungnya sintesis protein
jaringan yang baru (Davies 1982).
keambaan (
bulk density
yaitu berat/satuan volume) yang rendah, lemak yang tinggi.
Lebih lanjut dinyatakan bahwa konsentrasi energi yang tinggi menurunkan konsumsi
makanan dan konsumsi energi, kemungkinan disebabkan fermentasi dalam rumen
tidak normal atau hambatan cita rasa (
appetite
) oleh faktor kimiawi yang
dihasilkannya atau yang ada dalam saluran pencernaan.
Peningkatan konsumsi ransum sebagai resultan dari kecernaan dan
passage
rate
akan meningkatkan pula ketersediaan nutrien (terutama protein dan energi)
untuk proses produksi ternak. Faktor yang sangat penting mempengaruhi produksi
ternak adalah total nutrien diserap. Dengan demikian konsumsi dan daya cerna adalah
parameter kunci dalam system efaluasi makanan, diantara keduanya konsumsi sangat
penting karena sangat berbeda antara type makanan (Poppi
et al.
2000). Selanjutnya
dinyatakan, bahwa ada hubungan yang erat antara konsumsi dan daya cerna walaupun
secara keseluruhan tidak selalu ada hubungan. Hubungan antara konsumsi dan daya
cerna ini dapat dijelaskan sebagai berikut : bila kecernaan zat makanan tinggi akan
cepat laju makan selanjutnya akan cepat pula pengosongan perut dan gilirannya akan
cepat mengkonsumsi makanan yang mengandung nutrien tersebut. Konsumsi
dire-gulasi oleh laju pengosongan digesta dari rumen oleh proses pencernaan dan proses
pengaliran (
passage
).
Protein digunakan dalam tubuh ternak terutama untuk unsur pembangun dan
pengatur tubuh (prekursor enzim dan hormon), sehingga kebutuhannya lebih besar
untuk sapi muda dari pada sapi yang telah mencapai dewasa tubuh, untuk sapi yang
mepunyai pertambahan bobot badan yang tinggi daripada yang rendah, untuk sapi
type besar daripada type kecil (NRC 1984)
Sapi Pesisir di Sumatera Barat
Di Sumatera Barat terutama di Kabupaten Pesisir Selatan terdapat sapi lokal
yang disebut masyarakat sebagai sapi Pesisir (Sarbaini 2004). Menurut Merkens
(1926) yang diacu Sarbaini (2004), bahwa di Padang dan dataran tinggi sekitarnya
tedapat sapi dengan ciri-ciri : pejantannya memiliki kepala pendek; bertanduk pendek
dan mengarah ke luar; lehernya lebar, kokoh dan pendek. Punuknya cukup
berkem-bang, daerah pinggulnya pendek dan oval. Bagian depan badannya lebih ringan
Gambar 3 Sapi Pesisir betina berfistula rumen
[image:45.595.192.398.296.454.2]dibandingkan bagian belakangnya, dan kakinya relatif pendek. Pada sapi betina
kepalanya lebih panjang dan kecil, pundak dan dadanya kurang berkembang,
selangkangnya miring ke belakang, pendek dan kecil. Warnanya coklat muda sampai
coklat tua, atau sampai hitam. Sekitar mata, mulut dan sebelah dalam kaki-kakinya,
Tabel 2 Rataan bobot badan (kg) sapi Pesisir Sumatera Barat
perut bagian bawah berwarna lebih muda. Tinggi pundak pada sapi jantan berkisar 1
– 1.26 m (rata-rata 1.15 m) dan pada sapi betina sekitar 1.05 m.
Sejak umur muda sampai dewasa, sapi jantan mempunyai bobot badan yang
BAHAN DAN METODE
Penelitian ini dilakukan dalam tiga tahap ;
Penelitian Tahap I
Penelitian Tahap II
Penelitian Tahap III
Gambar 4 Bagan alir tahap penelitian.
Tahap I
:
Penentuan Koefisien dan Indeks Sinkronisasi Degradasi Protein dan
BO Bahan Pakan dalam Rumen.
Tujuan percobaan ini adalah untuk menentukan koefisien degradasi protein dan
BO bahan pakan secara in sacco (Orskov adan McDonald 1979) yang digunakan
untuk menentukan indeks sinkronisasi penyediaan N dan BO terfermentasi
berdasarkan nisbah degradasi protein dan BO per jam (hour