Jurnal Biologi Sumatera, Januari 2007, hlm. 17 – 20 ISSN 1907-5537
Vol. 2, No. 1
ANALISIS MODEL KETAHANAN RUMPUT GAJAH DAN RUMPUT RAJA
AKIBAT CEKAMAN KEKERINGAN BERDASARKAN RESPONS ANATOMI
AKAR DAN DAUN
Riyanto Sinaga
Departemen Biologi, Fakultas MIPA, Universitas Sumatera Utara, Jln. Bioteknologi No. 1, Kampus USU, Padang Bulan, Medan 20155
Abstract
The analysis model of the resistance of Elephant and King Grass on drought based on root and leaf anatomy response had been established by using multiple regression analysis method. Stepwise regression procedure has obtained three regression equation.The best regression equation is: Y = -164 + 0.116 Stele diameter + 0.973 thick of leaf.
Keywords: king grass and elephant grass, multiple regression analysis, stepwise regression
PENDAHULUAN
Masalah yang dihadapi peternak pada musim kemarau adalah kekurangan hijauan. Sebagai negara beriklim tropik, produksi hijauan makanan ternak di Indonesia sangat bervariasi akibat ketersediaan air dari hujan yang tidak menentu, yang berakibat pada fluktuasi status air tanah dan ketersediaan hara untuk tanaman. Peranan air sangat besar dalam menunjang pertumbuhan tanaman, yaitu untuk kelangsungan proses metabolisme. Tanaman yang mengalami kekeringan pada waktu yang lama akan mengalami perubahan-perubahan morfologi, anatomi, fisiologi dan biokimia yang tidak dapat kembali pulih sehingga dapat menyebabkan kematian.
Cekaman kekeringan adalah keadaan lingkungan yang menyebabkan kekurangan air bagi tanaman. Cekaman air pada tanaman dapat disebabkan oleh dua hal yaitu: (1) kekurangan air di daerah perakaran, (2) laju evapotranspirasi lebih tinggi dibandingkan dengan laju absorbsi oleh akar tanaman sehingga kebutuhan air pada daun lebih tinggi. Penyerapan air oleh tanaman dipengaruhi oleh faktor lingkungan dan faktor tanaman. Faktor lingkungaan yang berpengaruh adalah kandungan air tanah, kelembaban udara dan suhu tanah. Faktor tanaman adalah efesiensi perakaran, perbedaan tekanan difusi air tanah ke akar, dan keadaan protoplasma tanaman (Kramer 1969).
Perubahan-perubahan morfologi pada tanaman yang mengalami kekeringan antara lain terhambatnya pertumbuhan akar, tinggi tanaman, diameter batang, luas daun dan jumlah daun. Sedangkan pengaruh fisiologi dan biokimia adalah, penurunann hasil atau bahan kering, perubahan
alokasi asimilat, penurunan laju fotosintesis, penurunan diameter hidraulik xilem akar dan laju pertumbuhan tanaman. Pada penelitian ini diamati juga perubahan-perubahan anatomi pada tanaman yang diakibatkan oleh cekaman air antara lain, tebal epidermis daun, tebal mesofil, tebal daun, diameter akar, kerapatan stomata dan jumlah stomata.
Tanggap fisiologi tanaman yang berkaitan dengan ketahanan terhadap kekeringan sudah lama dipelajari. Gangguan fisiologis akibat cekaman air dapat berupa terhambatnya translokasi hara mineral dan asimilat, transpirasi dan fotosintesis. Secara visual tampak adanya kelayuan atau menggulungnya daun sehingga menghambat fotosintesis. Selanjutnya akan mempengaruhi pertumbuhan dan perkembangan organ
source and sink, seperti berkurangnya luas daun, mempengaruhi kandungan dan organisasi klorofil, perkembangan bunga dan buah atau organ reproduktif. Tanaman yang lebih toleran terhadap cekaman kekeringan mempunyai pertumbuhan yang relatif kurang terhambat dalam kondisi kekeringan dibandingkan pertumbuhan tanaman yang lebih peka (Kirkham 1990).
Tanaman beradaptasi terhadap cekaman lingkungan dengan menghasilkan senyawa-senyawa osmoregulasi yang dapat menurunkan potensial osmotik sehingga menurunkan potensial air dalam sel tanpa membatasi fungsi enzim serta menjaga turgor sel. Beberapa senyawa yang berperan dalam penyesuaian tekanan osmotik sel yang juga termasuk dalam kelompok molekul organik antara lain peningkatan akumulasi prolin dalam daun (Maestri et al. 1995, Hamim et al. 1996), asparagin dan betain (Munns et al. 1979, Maestri et al. 1995), protein dehidrin (Close 1997), gula osmotik (Wang et al.
SINAGA J. Biologi Sumatera
18
1995, Yakushiji et al. 1998), dan asam absisik (ABA) (Dingkuhn et al. 1991).
Rumput Gajah (P. purpureum Schum.) adalah salah satu jenis hijauan unggul untuk makanan ternak karena berproduksi tinggi, kualitasnya baik, dan daya adaptasinya tinggi. Rumput Gajah ini banyak ditanam dan dimanfaatkan pada peternakan penggemukan sapi, persusuan dan pembibitan. Di Indonesia produksi segar rumput Gajah jenis Hawaii berbulu mencapai 277 ton/ha/tahun (36 ton/ha/tahun bahan kering). Umumnya rumput Gajah digunakan sebagai rumput potong.
Rumput Raja adalah hasil persilangan antara
purpureum dan thypoides. Rumput Raja adalah jenis tanaman perenial yang membentuk rumpun, daya adaptasi yang baik di daerah tropis, tumbuh baik pada tanah yang tidak terlalu lembab dan didukung dengan irigasi yang baik. Pertumbuhan awal rumput Raja lebih lambat dan memerlukan perawatan yang lebih intensif dibandingkan dengan rumput Gajah namun memiliki pertumbuhan yang cepat mengalahkan rumput Gajah (BPTHMT Baturaden 1989).
Walaupun potensi rumput Gajah dan rumput Raja sangat besar dalam meningkatkan produktifitas ternak, akan tetapi sangat sedikit pengetahuan tentang mekanisme adaptasinya terhadap cekaman kekeringan. Oleh karena itu pada penelitian ini dilakukan analisis model ketahanan rumput Gajah dan rumput Raja akibat cekaman kekeringan berdasarkan respon anatomi akar dan daun sehingga diketahui peubah-peubah yang berpengaruh terhadap indikator ketahanan kedua jenis rumput terhadap cekaman kekeringan yaitu bobot kering tajuk.
BAHAN DAN METODA
Penyiapan media tanam dan pemeliharaan.
Penelitian ini dilakukan dalam pot plastik berukuran 8 kg menggunakan tanah podsolik yang dikeringkan selama 2 hari dan dicampur dengan pupuk dasar TSP dan KCl dengan dosis 4,05 g/pot. Setiap pot ditanam tiga stek yang panjangnya 30 cm dan setelah tiga minggu dilakukan penjarangan dengan meninggalkan satu stek yang masih hidup. Pemeliharaan dilakukan dengan pemberian pupuk urea pada 0 dan 3 minggu setelah tanam dengan dosis 8,1 g/pot dan pemberian air sampai 100% air tanah tersedia.
Menentukan perlakuan ketersediaan air.
Kapasitas lapang (KL) dihitung dengan cara penimbangan dan oven setelah polibag diberi air yang jenuh dan dibiarkan tertutup selama 24 jam dan diketahui KL adalah 51,16%. Untuk mengukur titik layu permanen (TLP) dilakukan dengan menggunakan alat Pressure Plate Apparatus dengan pF 4.20 pada tekanan 15 bar dan dengan cara gravimetric diketahui nilainya adalah 26,88%. Ketersediaan air dalam tanah (Kat) ditentukan dengan mencari selisih antara kadar air kapasitas lapang dan titik layu permanen (KA =
51,16% – 26,88% = 24,28%). Ditentukan juga kadar air kering pot (KP) dan Bobot kering tanahnya dan diketahui besarnya masing-masing adalah 27,94% dan 6252,93 g. Untuk menentukan tingkat kadar air dari masing-masing perlakuan (KAp) sebagai berikut.
Kadar air perlakuan (KAp) untuk tiap perlakuan adalah: (Perl. x Kat) + TLP
Penyesuaian kadar air untuk tiap-tiap perlakuan dilakukan dengan menimbang bobot tanah dan tanaman pada saat penyiraman.
Peubah yang diukur. Peubah yang diukur
meliputi peubah tak bebas dan peubah bebas. Peubah
tak bebas yaitu bobot kering tajuk (g) yang diperoleh dari pemanenan tajuk dan penimbangan setelah dimasukkan dalam oven 70 OC selama 48 jam.
Peubah bebas meliputi tebal epidermis daun(µm), tebal mesofil daun (µm), tebal daun (µm) (Widjaya 1996); diameter akar (µm), diameter stele (µm), tebal korteks (µm), kerapatan stomata adaksial dan abaksial (n/mm2), jumlah stomata adaksial dan abaksial (n/luas daun), jumlah xilem akar. Khusus untuk peubah diameter hidraulik xilem akar (µm4) dihitung berdasarkan persamaan Lewis dan Boose (1995).
Model pendugaan bobot kering tajuk.
Analisis model ketahanan rumput Gajah dan rumput Raja dinilai berdasarkan model pendugaan bobot kering tajuknya, karena bobot kering tajuk adalah cermin langsung dari respon kedua jenis rumput terhadap perubahan keadaan lingkungan.
Peubah yang dipakai sebagai penyusunan model (fungsi) adalah peubah tidak bebas yaitu Y
(bobot kering tajuk; gram) dan peubah bebas yang terdiri dari X1 – X10 (karakter anatomi akar dan daun). Pemilihan model yang merupakan fungsi linier dilaksanakan dengan melakukan pendugaan parameter, dalam hal ini konstanta pada fungsi regresi, pengujian keandalan dan pengujian keabsahan. Pendugaan parameter dilakukan dengan metode jumlah kuadrat terkecil, kriteria koefesien determinasi (R2), pengujian koefesien regresi, bentuk sebaran sisaan dan koefesien Cp-mallow (Afifi dan Clark 1984).
HASIL DAN PEMBAHASAN
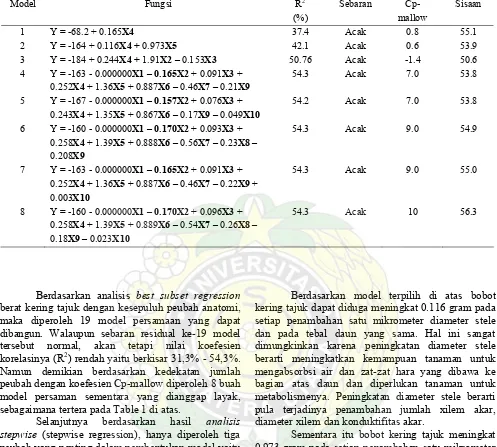
Dengan Best Subset Regression pada program MINITAB dapat dilakukan penyisihan sehingga diperoleh beberapa model untuk sementara yang dapat diandalkan sebagaimana tertera pada tabel 1 di bawah ini.
Vol. 2, 2007 J. Biologi Sumatera 19
Tabel 1. Model pendugaan sementara hasil analisis best subset regression dan analisis stepwise
Model Fungsi R2
(%)
Sebaran Cp- mallow
Sisaan
1 Y = -68.2 + 0.165X4 37.4 Acak 0.8 55.1
2 Y = -164 + 0.116X4 + 0.973X5 42.1 Acak 0.6 53.9
3 Y = -184 + 0.244X4 + 1.91X2 – 0.153X3 50.76 Acak -1.4 50.6 4 Y = -163 - 0.000000X1 – 0.165X2 + 0.091X3 +
0.252X4 + 1.36X5 + 0.887X6 – 0.46X7 – 0.21X9
54.3 Acak 7.0 53.8
5 Y = -167 - 0.000000X1 – 0.157X2 + 0.076X3 + 0.243X4 + 1.35X5 + 0.867X6 – 0.17X9 – 0.049X10
54.2 Acak 7.0 53.8
6 Y = -160 - 0.000000X1 – 0.170X2 + 0.093X3 + 0.258X4 + 1.39X5 + 0.888X6 – 0.56X7 – 0.23X8 –
0.208X9
54.3 Acak 9.0 54.9
7 Y = -163 - 0.000000X1 – 0.165X2 + 0.091X3 + 0.252X4 + 1.36X5 + 0.887X6 – 0.46X7 – 0.22X9 + 0.003X10
54.3 Acak 9.0 55.0
8 Y = -160 - 0.000000X1 – 0.170X2 + 0.096X3 + 0.258X4 + 1.39X5 + 0.889X6 – 0.54X7 – 0.26X8 – 0.18X9 – 0.023X10
54.3 Acak 10 56.3
Berdasarkan analisis best subset regression
berat kering tajuk dengan kesepuluh peubah anatomi, maka diperoleh 19 model persamaan yang dapat dibangun. Walaupun sebaran residual ke-19 model tersebut normal, akan tetapi nilai koefesien korelasinya (R2) rendah yaitu berkisar 31,3% - 54,3%. Namun demikian berdasarkan kedekatan jumlah peubah dengan koefesien Cp-mallow diperoleh 8 buah model persaman sementara yang dianggap layak, sebagaimana tertera pada Table 1 di atas.
Selanjutnya berdasarkan hasil analisis stepwise (stepwise regression), hanya diperoleh tiga peubah yang penting dalam pembentukan model yaitu X4 (diameter stele), X5 (tebal daun) dan X3 (diameter akar). Dengan berasaskan pada kemudahan penggunaan model, kedekatan koefesien Cp-mallow dan jumlah peubah, sisaan yang kecil, koefesien korelasi yang besar serta kemudahan dalam penggunaan model, maka model terpilih adalah model ke-2 yaitu: Bobot kering tajuk = -164 + 0.116 diameter stele + 0.973 tebal daun
Sedangkan model yang lain dianggap sulit dan kurang efisien dalam menduga bobot kering tajuk karena menggunakan lebih banyak peubah sehingga menyulitkan dalam menginterpretasikan dan menyulitkan dalam pengerjaan di lapangan. Kemudahan suatu model adalah kemampuan suatu model untuk menjelaskan peubah tak bebas dengan memakai peubah bebas paling sedikit (Afifi dan Clark 1984).
Berdasarkan model terpilih di atas bobot kering tajuk dapat diduga meningkat 0.116 gram pada setiap penambahan satu mikrometer diameter stele dan pada tebal daun yang sama. Hal ini sangat dimungkinkan karena peningkatan diameter stele berarti meningkatkan kemampuan tanaman untuk mengabsorbsi air dan zat-zat hara yang dibawa ke bagian atas daun dan diperlukan tanaman untuk metabolismenya. Peningkatan diameter stele berarti pula terjadinya penambahan jumlah xilem akar, diameter xilem dan konduktifitas akar.
Sementara itu bobot kering tajuk meningkat 0.973 gram pada setiap penambahan satu mikrometer tebal daun pada keadaan diameter akar yang sama. Kondisi inipun sangat logis dan dimungkinkan karena daun adalah organ tempat berlangsungnya proses fotosintesis penghasil karbohidrat, bahan makanan penyusun dinding sel. Peningkatan tebal daun berarti pula menambah ketebalan mesofil dan semakin besarnya jumlah klorofil yang dapat dikandung oleh daun (Salisbury dan Ros 1995).
DAFTAR PUSTAKA
Afifi A A, Clark V. 1984. Computer Aided Multivariate Analysis. Van Nostrand Reinhold Co. New York
Balai Penelitian Ternak dan Hijauan Makanan Ternak Baturaden. 1989. King Grass. Direktorat Bina Produksi Peternakan. Direktorat Jenderal Peternakan Baturaden. Baturaden.
SINAGA J. Biologi Sumatera
20
Dingkuhn M, Cruz RT, O’Toole, Turner NC, Doerffling. 1991. Responses of seven diverse rice cultivars to water deficits. III. accumulation of abscisic acid and proline in relation to leaf water-potensial and osmotic adjustment. Field Crops Res. 27: 103-117. Hamim, Soepandie D, Yusus M. 1996. Beberapa
karakteristik morfologi dan fisiologi kedelai toleran dan peka terhadap cekaman kekeringan. Hayati 3: 30-40.
Kirkham MB. 1990. Plant response to water deficits. Di dalam Stewart BA, Nielsen DR, editors.
Irrigation of Agricultural Crops. Wisconsin: Madison hlm 323-342.
Kramer PJ. 1969. Plant and Soil Water Relationship. Mc. New York: Graw HJill Book Company. Inc. hlm 347.
Lewis AM, Boose ER. 1995. Estimating volume flow rates through xilem conduits. Di dalam. Eshel A, Ilona Shick, Waisel Y, Stokes A, editors. 2000. The efficiency of the water conducting system of tomato roots. Hydraulic conductivity of tomato roots. Netherlands: Kluwer Academic Publishers hlm 371-375.
Maestri M, Da Matta FM, Regazzi AJ, Barros RS. 1995. Accumulation of praline and quaternary ammonium compounds in mature leaves of water stressed coffe plants (Coffea arabica
and C. canephora). J. Hort. Sci. 70: 229-233. Munns RC, Brady J, Barlow EWR. 1979. Solute
accumulation in apex and leaves of wheat during water stress. Aust. J. Physiol. 6: 379-389.Salisbur dan Ros 1995).
Salisbury FB, Ross CW. 1995. Fisiologi Tumbuhan. Terjemahan Lukman DR dan Sumaryono. Jilid 3. Penerbit ITB Bandung.
Wang Z, Quebedeaux, Stutte GW. 1995. Osmotic adjustment: effect of water stress on carbohydrates in leaves, steams and roots of apple. Aust. J. Plant physiol. 22: 747-754. Yakushiji H, Morinaga K, Nonami H. 1998. Sugar
accumulation and partitioning in Satsuma Mandarin tree tissue and fruit in response to drought stress. J. Amer. Soc. Hort. Sci. 123 (4): 7.