EVALUASI Saccharomyces cerevisiae TERADAPTASI HIDROLISAT
Kappaphycus alvarezii DALAM KEMAMPUAN MENGONVERSI
HIDROLISAT MENJADI BIOETANOL
HENG RAKA ABIMANYU
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
HENG RAKA ABIMANYU. Evaluasi Saccharomyces cerevisiae Teradaptasi Hidrolisat Kappaphycus alvarezii dalam Kemampuan Mengonversi Hidrolisat Menjadi Bioetanol. Dibimbing oleh GAYUH RAHAYU dan DWI SETYANINGSIH.
Bioetanol merupakan salah satu sumber energi hasil fermentasi yang dapat diperbaharui dan ramah lingkungan. Produksi bioetanol memerlukan dua komponen penting yaitu substrat dan agen fermentasi. Kappaphycus alvarezii merupakan substrat potensial untuk difermentasi oleh Saccharomyces cerevisiae, mikroorganisme yang paling umum digunakan dalam proses fermentasi. Penelitian ini bertujuan untuk mengetahui kemampuan S. cerevisiae strain (IPBCC 05.548 AL IX) yang telah teradaptasi lambat selama 9 generasi untuk mengubah hidrolisat E. cottonii menjadi bioetanol. Pertumbuhan khamir maksimum pada hidrolisat maupun Yeast Malt Glucose Pepton (YMGP) terjadi pada 42 jam inkubasi. Pada hidrolisat, ukuran sel khamir relatif lebih panjang dengan jumlah sel besarnya lebih banyak dibandingkan pada YMGP. Sekitar 90 mL hidrolisat K. alvarezii yang difermentasikan dengan 10 mL inokulan pada konsentrasi 106 mL-1, menghasilkan bioetanol maksimum sebesar 1,94% (v/v) pada 168 jam inkubasi. Efisiensi fermentasi, dan efisiensi substratnya berturut-turut sebesar 36,57%, dan 59,85%. Pada periode inkubasi yang sama dan konsentrasi inokulan 106 sel mL-1 menghasilkan bioetanol maksimum sebesar 1,02% (v/v) dengan efisiensi fermentasi dan efisiensi substratnya berturut-turut sebesar 21,29%, dan 70,07%.
ABSTRACT
HENG RAKA ABIMANYU. Evaluation of Saccharomyces cerevisiae Adapted Hydrolysates Kappaphycus alvarezii in Converting Capabilities Hydrolysates to Become Bioetanol. Supervised by GAYUH RAHAYU and DWI SETYANINGSIH.
EVALUASI Saccharomyces cerevisiae TERADAPTASI HIDROLISAT
Kappaphycus alvarezii DALAM KEMAMPUAN MENGONVERSI
HIDROLISAT MENJADI BIOETANOL
HENG RAKA ABIMANYU
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul
: Evaluasi
Saccharomyces cerevisiae
Teradaptasi Hidrolisat
Kappaphycus alvarezii
dalam Kemampuan Mengonversi Hidrolisat
Menjadi Bioetanol
Nama
: Heng Raka Abimanyu
NIM
: G34080017
Menyetujui,
Tanggal lulus :
Pembimbing 1
Dr. Ir. Gayuh Rahayu
NIP. 19580105 198303 2 002
Pembimbing II
Dr. Ir. Dwi Setyaningsih, M.Si.
NIP. 19700103 199412 2 001
Mengetahui,
Ketua Departemen Biologi
Dr. Ir. Ence Darmo Jaya Supena, M.Si.
PRAKATA
Alhamdulillah puji syukur penulis panjatkan kehadirat Allah SWT yang telah memberikan rahmat-Nya kepada penulis sehingga karya ilmiah ini berhasil diselesaikan. Adapun Tema yang dipilih dalam karya ilmiah yang dilaksanakan sejak bulan Maret 2012 sampai dengan September 2012 berjudul Evaluasi Saccharomyces cerevisiae Teradaptasi Hidrolisat Kappaphycus alvarezii dalam Kemampuan Mengonversi Hidrolisat Menjadi Bioetanol. Pelaksanaan karya ilmiah bertempat di laboratorium Surfactant and Bioenergy Research Center (SBRC) dan laboratorium Mikologi IPB.
Penulis mengucapkan terima kasih kepada Ibu Dr. Ir. Gayuh Rahayu selaku pembimbing pertama dan Dr. Ir. Dwi Setyaningsih, M.Si. selaku pembimbing kedua yang telah sepenuh hati memberikan bimbingan, arahan, dan nasihat yang berarti sehingga karya ilmiah ini dapat selesai. Ucapan terima kasih pula penulis ucapkan kepada Prof. Dr. Anja Meryandini, M.S. sebagai dosen penguji yang telah memberikan sarannya. Terima kasih kepada Dr. Nunik Sri Ariyanti, M.Si. yang telah memberikan masukan dan sarannya. Terima kasih kepada Proyek atas nama Ibu Dr. Ir. Dwi Setyaningsih, M.Si. yang telah membiayai penelitian ini. Terima kasih kepada direktur dan karyawan SBRC yang telah menyediakan tempat dan fasilitas selama penulis melakukan karya ilmiah. Tidak lupa juga penulis ucapkan terima kasih kepada teman-teman Biologi angkatan 45 atas kebersamaan serta rasa kekeluargaan yang diberikan. Terima kasih yang tak terhingga kepada orang tua tercinta yang telah menyemangati dan memberikan do’a yang tak kenal lelah kepada penulis.
Semoga karya ilmiah ini dapat bermanfaat.
Bogor, Februari 2013
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 6 Mei 1990 sebagai anak pertama dari dua bersaudara pasangan Bapak Rahmadi dan Ibu Suwarni. Penulis menyelesaikan pendidikan dasar di SDN Cipete Utara 15 Pagi Kebayoran Baru pada tahun 2002. Penulis menyelesaikan pendidikan menengah pertama di SLTP N 56 Jeruk Purut tahun 2005. Pendidikan Sekolah Menengah Atas diselesaikan pada tahun 2008 di SMA N 97 Brigif Jakarta Selatan. Penulis diterima di Institut Pertanian Bogor di tahun yang sama melalui jalur Undangan Seleksi Masuk IPB (USMI).
DAFTAR ISI
Halaman
DAFTAR TABEL ... viii
DAFTAR LAMPIRAN ... viii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 1
BAHAN DAN METODE ... 1
Waktu dan Tempat ... 1
Bahan ... 2
Persiapan Kultur Kerja ... 2
Pertumbuhan S. cerevisiae pada Media YMGP dan Hidrolisat K. alvarezii ... 2
Produksi Bioetanol pada Beberapa Waktu Inkubasi ... 2
Produksi Bioetanol pada Beberapa Konsentrasi Inokulan ... 2
HASIL ... 2
Pertumbuhan S. cerevisiae pada Media YMGP dan Hidrolisat K. alvarezii ... 2
Produksi Bioetanol pada Beberapa Waktu Inkubasi ... 4
Produksi Bioetanol pada Beberapa Konsentrasi Inokulan ... 4
PEMBAHASAN ... 5
SIMPULAN ... 6
SARAN ... 6
DAFTAR PUSTAKA ... 6
DAFTAR TABEL
Halaman
1 Ukuran sel khamir selama 192 jam inkubasi pada media YMGP dan hidrolisat K. alvarezii .. 3 2 Rasio sel khamir selama 192 jam inkubasi pada media YMGP dan hidrolisat K. alvarezii ... 4 3 Volume destilat, produksi bioetanol, efisiensi fermentasi, dan efisiensi substrat pada
fermentasi hidrolisat K. alvarezii oleh S. cerevisiae selama 192 jam inkubasi ... 4 4 Volume destilat, produksi bioetanol, efisiensi fermentasi, dan efisiensi substrat hasil
fermentasi hidrolisat K. alvarezii oleh S. cerevisiae dengan jumlah inokulum tertentu pada waktu inkubasi 168 jam ... 5
DAFTAR LAMPIRAN
Halaman
1 Komposisi berbagai media dan larutan yang digunakan dalam penelitian ... 9 2 Perhitungan produksi bioetanol, efisiensi fermentasi, efisiensi substrat, dan kadar
gula pereduksi ... 9 3 Diagram alir pengukuran gula pereduksi ... 10 4 Konsentrasi gula pereduksi selama pertumbuhan S. cerevisiae dan selama produksi
1
PENDAHULUAN
Latar BelakangBioetanol mulai dikembangkan pada tahun 1970-an dan merupakan salah satu sumber energi yang dapat diperbaharui dan ramah lingkungan (Jeffries 2005). Berdasarkan peraturan Kementerian Energi dan Sumber Daya Mineral No 32 Tahun 2008, pada tahun 2008 sampai 2010 etanol harus mensubstitusi premium sebanyak 3% untuk transportasi dan akan meningkat menjadi 5% di tahun 2015, 10% pada tahun 2020 dan 15% di tahun 2025 (Sutarto 2009). Oleh sebab itu, bioetanol sangat dibutuhkan.
Bioetanol dapat dibuat dari bahan baku berbasis gula, pati atau serat. Sakarifikasi pati dan serat (selulosa, hemiselulosa, dan lignoselulosa) dapat diperoleh melalui hidrolisis asam encer, hidrolisis enzimatis atau menggabungkan keduanya. Hidrolisis polisakarida merupakan reaksi kimia menggunakan air untuk memecah atau memutus rantai panjang polisakarida menjadi rantai-rantai pendek atau gula sederhana (Yuniwati et al. 2011). Hidrolisis asam encer biasanya menggunakan asam seperti H2SO4
dan HCl. Hidrolisis asam encer memiliki beberapa kelebihan yaitu harganya lebih murah, lebih cepat, mudah didapat, dan rendemen gula lebih tinggi jika dibandingkan dengan hidrolisis enzim. Akan tetapi, hidrolisis asam encer juga memiliki kekurangan yaitu cenderung korosif terhadap alat, toksik terhadap lingkungan dan dapat menghasilkan produk sampingan yang bersifat toksik (Taherzadeh & Karimi 2007).
Salah satu sumber polisakarida yang tersedia secara berlimpah adalah rumput laut. Kappaphycus alvarezii merupakan salah satu jenis rumput laut yang dibudidayakan dan dapat menghasilkan karaginan (Distantina et al. 2010). Adapun komposisi dari K. alvarezii terdiri atas karbohidrat (41,43%), protein (1,86%), lemak (0,35%), air (11,28%), abu (36,05%), dan serat kasar (8,96%) (Devis 2008). Karaginan merupakan polisakarida galaktosa. Sebagian besar karaginan mengandung natrium, magnesium, dan kalsium yang dapat terikat pada gugus ester sulfat dari galaktosa dan kopolimer 3,6- anhydro-galaktosa yang sulit digunakan oleh mikroba dibandingkan glukosa (Bawa et al. 2007). Salah satu mikroba yang dapat menggunakan galaktosa sebagai sumber karbon adalah khamir.
Saccharomyces cerevisiae merupakan khamir yang paling banyak digunakan dalam
proses fermentasi (Jeffries & Shi 1999) dan khamir tersebut juga sangat dikenal oleh masyarakat luas sebagai khamir roti. Khamir roti ini digunakan dalam pembuatan makanan, minuman, dan juga dalam industri etanol (Ostergaard et al. 2000). Khamir ini digunakan dalam proses fermentasi alkohol karena mampu memproduksi bioetanol dalam jumlah besar ± 15% (v/v) (Martini 2003), toleran terhadap etanol yang cukup tinggi 9-12% (v/v), dan toleran terhadap gula tinggi ± 25% (b/v) (Osho 2005).
Adanya inhibitor seperti 5-hidroksimetilfurfural (HMF), furfural, asam asetat dan asam levulinat yang terbentuk dalam hidrolisis asam encer menghambat metabolisme S. cerevisiae sehingga mengakibatkan penurunan biomassa dan produksi bioetanol. S. cerevisiae yang tahan terhadap inhibitor bisa didapatkan dengan proses adaptasi. Beberapa penelitian telah menunjukkan bahwa dengan proses adaptasi S. cerevisiae pada media hidrolisat asam akan meningkatkan kemampuan S. cerevisiae dalam memfermentasi substrat yang mengandung inhibitor (Meinita et al. 2011; Hawkins & Peterson 2011).
Penelitian sebelumnya menunjukkan bahwa S. cerevisiae yang telah mengalami masa adaptasi lambat sebanyak 9 generasi melalui media perantara PSA (Potato Sucrose Agar) dan YMGP (Yeast Malt Glucose Pepton) pada hidrolisat K. alvarezii dapat menghasilkan bioetanol optimum sebesar 2% (v/v) pada waktu fermentasi selama 144 jam dengan konsentrasi inokulum 107 sel mL-1 (Setyaningsih et al. 2012). Padahal kondisi ini belum tentu merupakan kondisi optimum S. cerevisiae dalam produksi bioetanol dari hidrolisat K. alvarezii. Oleh sebab itu, kondisi fermentasi (waktu fermentasi dan konsentrasi inokulan) untuk produksi bioetanol dengan substrat hidrolisat K. alvarezii perlu diketahui.
Tujuan
Mengetahui waktu fermentasi dan konsentrasi inokulan yang dibutuhkan S. cerevisiae yang telah teradaptasi (IPBCC 05.548 AL IX) hidrolisat K. alvarezii untuk mengubah hidrolisat tersebut menjadi bioetanol.
BAHAN DAN METODE
Waktu dan Tempat
2
Laboratorium SBRC (Surfactant and Bioenergy Research Center) dan Laboratorium Mikologi, Departemen Biologi, FMIPA, IPB.
Bahan
Bahan yang digunakan adalah isolat S. cerevisiae koleksi IPB Culture Collection (IPBCC 05.548) yang telah mengalami adaptasi lambat dihidrolisat K. alvarezii selama 9 kali (IPBCC 05.548 AL IX), hidrolisat K. alvarezii, aquades, alkohol teknis 70 %, NPK, spirtus, H2SO4 3%, urea, kapur
tohor (Ca(OH)2), media PSA (Potato Sucrose
Agar) (Lampiran 1A), media YMGP (Yeast Malt Glucose Pepton) (Lampiran 1B), dan larutan DNS (Lampiran 1C).
Persiapan Kultur Kerja
Sebanyak 2 ose kultur S. cerevisiae IPBCC 05.548 AL IX dari PSA miring (stock) yang telah diremajakan di media PSA cawan petri, diambil dan dimasukkan ke dalam tabung reaksi yang berisi 10 mL media YMGP yang telah disterilkan di dalam autoklaf pada suhu 121oC, 1 atm selama 15 menit. Kultur diinkubasi di atas mesin pengocok pada kondisi ruang selama 48 jam. Biakan berumur 48 jam dijadikan kultur kerja.
Pertumbuhan S. cerevisiae pada Media YMGP dan Hidrolisat K. alvarezii
Hidrolisat K. alvarezii dipersiapkan menurut metode (Setyaningsih et al. 2012). Sebanyak 10 mL inokulan (kultur kerja) masing-masing dimasukkan ke 90 mL hidrolisat dan YMGP. Kemudian suspensi tersebut digoyangkan dengan kecepatan 120 rpm selama 192 jam. Pertumbuhan sel S. cerevisiae ditetapkan berdasarkan kerapatan sel dan diukur dengan hemasitometer setiap 6 jam selama 60 jam. Sel diukur dengan mikrometer pada mikroskop cahaya setiap 24 jam selama 192 jam. Jumlah sel besar dan sel kecilnya dihitung. Sel besar adalah sel dengan ukuran >5 m dan sel kecil adalah sel dengan ukuran ≤5 m (Volk & Thomas 2000).
Produksi Bioetanol pada Beberapa Waktu Inkubasi
Sebanyak 10 mL inokulan dengan konsentrasi sel 107 mL-1 dimasukkan ke Erlenmeyer yang berisi 90 mL 100% hidrolisat yang sebelumnya telah
dipasteurisasi suhu 70-80 °C selama 15 menit. Suspensi ini ditambahkan 0,06% NPK dan 0,5% urea dan diinkubasi selama 48, 72, 96, 120, 144, 168, dan 192 jam. Produksi bioetanol dinyatakan dalam volume etanol yang terbentuk. Hidrolisat terfermentasi didestilasi pada suhu 78°C dan volume destilatnya diukur, kemudian kadar bioetanolnya ditetapkan dengan bantuan densitometer (% v/v) (Lampiran 2A). Efisiensi fermentasi dan efisiensi substrat juga dihitung (Lampiran 2B dan 2C). Setiap perlakuan dilakukan dengan 3 ulangan dan setiap pengukuran diulang 2 kali.
Produksi Bioetanol pada Beberapa Konsentrasi Inokulan
Sebanyak 105, 106, dan 107 sel mL-1 dipersiapkan dengan metode pengenceran dan dijadikan inokulan. 1 mL dan 10 mL inokulan masing-masing dimasukkan ke dalam 99 mL dan 90 mL sehingga dihasilkan jumlah sel 105 dan 106 sel mL-1. Untuk didapatkan konsentrasi sel 107, yaitu dengan sentrifuse 10 mL inokulan dengan kecepatan 4000 rpm, sehingga didapatkan pelet yang kemudian dimasukkan ke dalam 100 mL hidrolisat. Kultur diinkubasi pada masa inkubasi yang menghasilkan bioetanol maksimum dari parameter waktu inkubasi. Produksi bioetanol diukur seperti cara sebelumnya. Efisiensi fermentasi dan efisiensi substrat juga dihitung. Setiap perlakuan dilakukan dengan 3 kali ulangan dan setiap pengukuran diulang 2 kali.
HASIL
Pertumbuhan S. cerevisiae pada Media YMGP dan Hidrolisat K. alvarezii
Pada saat inokulasi, populasi S. cerevisiae pada YMGP sebesar 2,0 x 107 sel mL-1 sedangkan pada hidrolisat, populasinya sebesar 1,3 x 107 sel mL-1. Perbedaan konsentrasi inokulan secara konsisten berhubungan dengan pertambahan populasinya. Pada YMGP dan hidrolisat K. alvarezii, populasi S. cerevisiae langsung meningkat dan mencapai populasi maksimum pada jam ke- 42 inkubasi. Setelah jam ke-42, populasi S. cerevisiae menurun (Gambar 1).
3
awalnya berukuran 2,5-6,3 x 2,5-5 m. Selama 48 jam pertama, ukuran sel pada YMGP maupun hidrolisat relatif sama. Namun setelah 48 jam inkubasi, sel-sel S. cerevisiae pada YMGP relatif lebih pendek daripada sel-sel S. cerevisiae pada hidrolisat (Tabel 1). Jumlah sel-sel besar S. cerevisiae
[image:11.595.111.496.192.404.2]pada YMGP juga lebih sedikit daripada hidrolisat, sedangkan sel kecil (sel tunas/anakan) pada YMGP lebih banyak dibandingkan sel kecil pada media hidrolisat (Tabel 2). Pertambahan sel S. cerevisiae yang pesat ditunjukkan dengan semakin besar jumlah sel kecil yang dihasilkan.
Gambar 1 Kurva pertumbuhan S. cerevisiae (IPBCC 05.548 AL IX) pada media YMGP (Yeast Malt Glucose Pepton) dan hidrolisat K. alvarezii.
Tabel 1 Ukuran sel S. cerevisiae selama 192 jam inkubasi pada media YMGP dan hidrolisat K. alvarezii
Waktu Inkubasi Media YMGP Media Hidrolisat (Jam) Panjang ( m) Lebar ( m) Panjang ( m) Lebar ( m)
0 2,5-6,3 2,5-5,0 2,5-6,3 2,5-5,0 48 2,5-7,5 2,5-7,5 1,3-6,3 1,3-6,3 72 2,5-7,5 2,5-6,3 2,5-8,8 2,5-8,8 96 2,5-6,3 2,5-6,3 1,3-7,5 1,3-6,3 120 1,3-7,5 1,3-7,5 2,5-10 2,5-7,5 144 1,3-7,5 1,3-7,5 1,3-7,5 1,3-7,5 168 2,5-7,5 2,5-7,5 2,5-10 2,5-6,3 192 2,5-7,5 2,5-6,3 2,5-8,8 2,5-7,5 7,00
7,20 7,40 7,60 7,80 8,00 8,20 8,40 8,60
0 6 12 18 24 30 36 42 48 54 60
Lo
g
se
l
m
L
-1
Waktu inkubasi (jam)
[image:11.595.105.511.475.691.2]4
Tabel 2 Rasio sel S. cerevisiae selama 192 jam inkubasi pada media YMGP dan hidrolisat K. alvarezii
Waktu
Inkubasi Media YMGP Media Hidrolisat
(Jam) Ʃ sel besar Ʃ sel kecil Rasio sel Ʃ sel besar Ʃ sel kecil Rasio sel 0 3 87 1 : 29 3 87 1 : 29 48 29 74 1 : 2 7 125 1 : 18 72 17 102 1 : 6 52 127 1 : 2 96 16 110 1 : 7 64 128 1 : 2 120 17 56 1 : 3 20 78 1 : 4 144 18 80 1 : 5 33 209 1 : 6 168 6 85 1 : 14 17 135 1 : 8 192 13 90 1 : 7 64 137 1 : 2
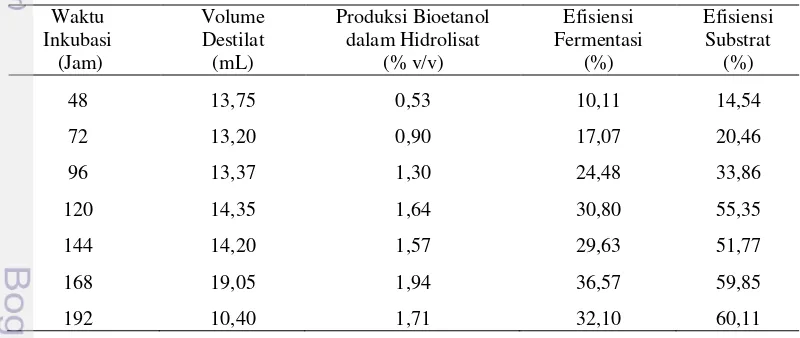
Produksi Bioetanol pada Beberapa Waktu Inkubasi
Jumlah sel inokulum sebesar 107 sel mL-1 dimasukkan ke dalam media hidrolisat K. alvarezii, dan diinkubasi selama 192 jam. Inokulan ini mampu hidup dan menghasilkan bioetanol dengan persentase volume terhadap hidrolisat berbeda-beda bergantung pada waktu inkubasinya. Produksi bioetanol selama
dengan 168 jam dan setelah itu menurun. Sampai dengan 168 jam inkubasi, produksi etanol berbanding lurus dengan efisiensi fermentasi dan efisiensi substratnya (Tabel 3). Produksi etanol terbesar terjadi pada jam ke-168 sebesar 1,94% (v/v), dengan volume destilat, efisiensi fermentasi, dan efisiensi substratnya berturut-turut sebesar 19,05 mL, 36,57%, dan 59,85% (Tabel 3).
masa inkubasi meningkat terus sampai
Tabel 3 Volume destilat, produksi bioetanol, efisiensi fermentasi, dan efisiensi substrat pada fermentasi hidrolisat K. alvarezii oleh S. cerevisiae selama 192 jam inkubasi
Waktu Inkubasi
(Jam)
Volume Destilat
(mL)
Produksi Bioetanol dalam Hidrolisat
(% v/v)
Efisiensi Fermentasi
(%)
Efisiensi Substrat
(%)
48 13,75 0,53 10,11 14,54
72 13,20 0,90 17,07 20,46
96 13,37 1,30 24,48 33,86
120 14,35 1,64 30,80 55,35
144 14,20 1,57 29,63 51,77
168 19,05 1,94 36,57 59,85
192 10,40 1,71 32,10 60,11
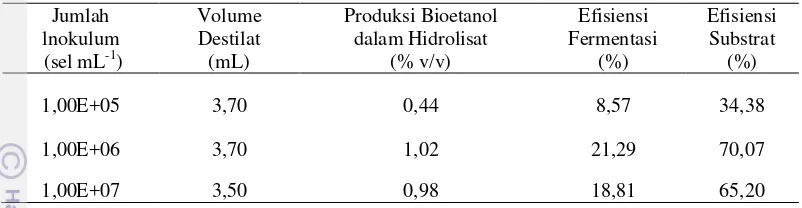
Produksi Bioetanol pada Beberapa Konsentrasi Inokulan
Diantara konsentrasi inokulan S. cerevisiae (105, 106, dan 107 sel mL-1) yang dipergunakan dalam fermentasi hidrolisat K. alvarezii dalam masa inkubasi 168 jam, produksi bioetanol maksimum yaitu sebesar 1,02% (v/v) dihasilkan oleh inokulan pada konsentrasi 106 sel mL-1 (Tabel 4). Volume destilat, efisiensi fermentasi, dan efisiensi
[image:12.595.106.506.110.270.2] [image:12.595.109.509.448.617.2]3
Tabel 4 Volume destilat, produksi bioetanol, efisiensi fermentasi, dan efisiensi substrat hasil fermentasi hidrolisat K. alvarezii oleh S. cerevisiae dengan jumlah inokulum tertentu pada waktu inkubasi 168 jam
Jumlah lnokulum (sel mL-1)
Volume Destilat (mL) Produksi Bioetanol dalam Hidrolisat (% v/v) Efisiensi Fermentasi (%) Efisiensi Substrat (%)
1,00E+05 3,70 0,44 8,57 34,38
1,00E+06 3,70 1,02 21,29 70,07 1,00E+07 3,50 0,98 18,81 65,20
PEMBAHASAN
Pada penelitian ini, pertumbuhan S. cerevisiae sangat dinamik. Pada saat jam ke-6 inkubasi, populasi sel S. cerevisiae pada YMGP sudah mencapai lebih dari tiga kali populasi awalnya, dan pada hidrolisat populasinya kurang lebih dua kali populasi awalnya, sehingga diduga fase lag terjadi pada periode sebelum 6 jam inkubasi. Menurut Singh et al. (2006), populasi S. cerevisiae akan meningkat sebanyak dua kali, 2 jam setelah inkubasi, sehingga fase lag terjadi selama 2 jam pertama periode inkubasi. Penelitian lain juga membuktikan bahwa fase lag sangat dipengaruhi oleh kondisi media. Media yang mengandung senyawa toksik akan memperpanjang fase lag (Hawkins & Peterson 2011).
Pola pertumbuhan S. cerevisiae pada YMGP mirip dengan pola pertumbuhannya pada hidrolisat K. alvarezii. Pertumbuhan S. cerevisiae dipengaruhi oleh ketersediaan sumber karbon dan menurut Kurtzman dan Fell (2000), glukosa menjadi sumber karbon utama bagi S. cerevisiae. Glukosa (4% b/v) merupakan komponen utama gula pada YMGP sedangkan pada hidrolisat K. alvarezii, galaktosa (2,24% b/v) merupakan gula utama disamping glukosa (0,01% b/v) (Meinita et al. 2011). Dalam penelitian ini, gula pereduksi awal (sebelum fermentasi) pada YMGP (4,73% b/v) kurang dari hidrolisat K. alvarezii (7,58% b/v) (Lampiran 4A). Pada jam ke-42 inkubasi, kandungan gula pereduksi yang tersisa pada YMGP dan hidrolisat berturut-turut sebesar 0,19% (b/v) dan 2,50% (b/v) (Lampiran 4A). Penurunan gula pereduksi ini disebabkan oleh adanya pemanfaatan gula untuk pertumbuhan populasi S. cerevisiae. Pertambahan populasi sel S. cerevisiae sampai dengan jam ke-42 inkubasi pada hidrolisat
menunjukkan bahwa S. cerevisiae dapat menggunakan glukosa dan galaktosa yang terdapat dalam hidrolisat. Sel S. cerevisiae harus mulai menggunakan galaktosa ketika glukosa dalam hidrolisat habis. Sel S. cerevisiae hasil adaptasi diduga mampu menggunakan galaktosa pada hidrolisat. Hong et al. (2011), juga menemukan dua galur S. cerevisiae yang mampu menggunakan galaktosa. Ketika galaktosa menjadi satu-satunya sumber karbon yang tersedia, S. cerevisiae akan menghasilkan enzim-enzim tertentu dalam metabolismenya untuk mengubah galaktosa menjadi etanol atau untuk perbanyakan selnya. Menurut Timson (2007), ada 5 jenis enzim terlibat dalam metabolisme galaktosa yang dihasilkan sel S. cerevisiae yaitu galaktosa mutarotase, galaktokinase, galaktosa 1-fosfat uridil-transferase, UDP-galaktosa 4-epimerase dan fosfoglukomutase.
Setelah populasi S. cerevisiae mencapai pertumbuhan maksimumnya pada jam ke-42 inkubasi, ukuran dan bentuk sel S. cerevisiae pada hidrolisat berubah. Sel-sel khamir menjadi lebih panjang dan jumlah sel-sel yang berukuran besar relatif lebih banyak. Menurut Adam et al. (1985), ukuran dan bentuk sel S. cerevisiae dipengaruhi oleh proses adaptasi. Ukuran sel juga dipengaruhi oleh fase hidupnya. Hermansyah (2010) menyatakan bahwa umumnya sel diploid memiliki diameter 6 m sedangkan sel haploid memiliki diameter sel 4 m. Sedangkan Volk dan Thomas (2000), menyatakan bahwa umumnya sel besar (sel induk) memiliki diameter >5 m sedangkan sel kecil (sel tunas) memiliki diameter ≤5 m.
6
perbanyakan sel khamir pada hidrolisat lebih lambat dibandingkan perbanyakan sel khamir pada media YMGP. Menurut Cooper dan Strich (2011), periode yang dibutuhkan untuk menghasilkan sel besar umumnya lebih lama dibandingkan periode untuk menghasilkan sel kecil.
Produksi bioetanol merupakan parameter yang dapat menunjukkan keberhasilan dari proses fermentasi alkohol. Produksi bioetanol terbesar terjadi pada jam ke-168 pada konsentrasi 107 sel mL-1 yaitu sebesar 1,94 % (v/v). Produksi bioetanol ini masih kurang dari produksi bioetanol menurut Setyaningsih et al. (2012) yaitu sebesar 2 % pada jam ke 144 inkubasi dengan konsentrasi inokulum 107 sel mL-1. Pada uji selanjutnya, konsentrasi inokulum yang sama menghasilkan bioetanol dengan konsentrasi yang rendah (0,98 % v/v) pada 168 jam inkubasi. Hal ini menunjukkan bahwa kemampuan S. cerevisiae teradaptasi dalam produksi bioetanol belum stabil.
Menurunnya produksi bioetanol kemungkinan besar disebabkan oleh kemampuan detoksifikasi zat toksin pada medium fermentasi oleh S. cerevisiae berubah-ubah. Pada awal fermentasi, sel khamir berusaha mendetoksifikasi senyawa penghambat (Maharani 2011). Menurut Meinita et al. (2011), beberapa senyawa toksik penghambat fermentasi yang terdapat pada hidrolisat K. alvarezii yaitu 5-hydroxy methyl furfural (HMF) dan asam levulinat. Hawkins dan Peterson (2011) juga menyatakan bahwa senyawa HMF, furfural, dan asam asetat menghambat pertumbuhan populasi S. cerevisiae.
Rendahnya produksi bioetanol juga dapat disebabkan oleh ketidakmampuan sel S. cerevisiae untuk toleran terhadap etanol. Prescott dan Dunn (1981) menyatakan bahwa S. cerevisiae sangat peka etanol, konsentrasi 1-2% (b/v) cukup menghambat pertumbuhan dan pada konsentrasi etanol 13% (b/v) laju pertumbuhan khamir hampir berhenti.
Pada saat produksi bioetanol mencapai maksimum pada jam ke-168, efisiensi fermentasinya paling besar. Menurut Devis (2008), produksi bioetanol yang dihasilkan dalam proses fermentasi tergantung pada efisiensi fermentasi.
Diantara 3 konsentrasi inokulum (105, 106, dan 107 sel mL-1) yang digunakan dengan waktu inkubasi 168 jam, konsentrasi sel 106 memproduksi bioetanol terbesar. Konsentrasi inokulan sebesar 107 sel mL menghasilkan destilat dan bioetanol kurang dari destilat dan bioetanol pada saat penetapan waktu inkubasi.
Hal tersebut diduga karena pada perlakuan tersebut tidak digunakan media starter (YMGP) dalam persiapan inokulan. Media YMGP berperan penting untuk persiapan konsentrasi inokulum sebelum fermentasi di media hidrolisat (Maharani 2011), media YMGP digunakan oleh khamir untuk tumbuh (Bergman 2001).
SIMPULAN
Pertumbuhan khamir S. cerevisiae IPBCC 05.548 AL IX pada hidrolisat maupun YMGP mengalami pertumbuhan maksimum pada jam ke-42. Pada hidrolisat, ukuran sel khamir lebih panjang dengan jumlah sel besarnya lebih banyak dibandingkan pada media YMGP. Selama 192 jam waktu inkubasi fermentasi, khamir dapat menghasilkan etanol maksimum pada jam ke-168 yaitu sebesar 1,94% (v/v) dengan volume destilat, efisiensi fermentasi dan efisiensi substratnya berturut-turut 19,05 mL, 36,57%, dan 59,85%. Jumlah inokulum 106 sel mL-1 merupakan konsentrasi sel khamir yang optimum untuk produksi etanol terbanyak yaitu sebesar 1,02% (v/v) dengan volume destilat, efisiensi fermentasi, dan efisiensi substratnya berturut-turut 3,70 mL, 21,29%, dan 70,07%.
SARAN
Adaptasi S. cerevisiae lebih lanjut sampai diperoleh galur yang stabil dalam kemampuan fermentasi hidrolisat K. alvarezii menjadi bioetanol perlu dilakukan.
DAFTAR PUSTAKA
Adam J, Paquin C, Oeller WP, Lee WL. 1985. Physiological characterization of adaptive clones in evolving populations of the yeast, Saccharomyces cerevisiae. Journal genetics 110: 173-185.
Bawa GAGI, Putra BAA, Laila RI. 2007. Penentuan pH optimum isolasi karaginan dari rumput laut jenis Eucheuma cottonii. Jurnal Kimia 1: 15-20.
Bergman WL. 2001. Growth and maintenance of yeast. Molecular Biology 177: 9-14. Cooper FK, Strich R. 2011. Meiotic control of
7
Devis FH. 2008. Bioetanol berbahan dasar ampas rumput laut Kappaphycus alvarezii. [skripsi]. Bogor: Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor. Distantina S, Fadilah, Rochmadi, Fahrurrozi
M, Wiratni. 2010. Proses ekstraksi karagenan dari Eucheuma cottonii. Naskah Seminar Nasional Rekayasa Kimia dan Proses, 4-5 Agustus 2010, Semarang. Hawkins MG, Peterson DJ. 2011. A strain of
Saccharomyces cerevisiae evolved for fermentation of lignocellulosic biomass displays improved growth and fermentative ability in high solids concentration and in the presence of inhibitory compound. Biotechnology for Biofuels 4: 49-62.
Hermansyah. 2010. Penentuan tipe mating ragi Saccharomyces cerevisiae. Sains 13: 13102.
Hong KK, Vongsangnak W, Vemuri GN, Nielsen J. 2011. Unrevelling evolutionary strategies of yeast for improving galactose utilization through integrated systems level analysis. Proceedings of the National Academy of Sciences. China: Soochow University. hlm 12179-12184.
Jeffries WT. 2005. Ethanol fermentation on the move. Nature Biotechnology 23(1): 40-41.
Jeffries WT, Shi NQ. 1999. Genetic engineering for improved xylose fermentation by yeast. Advances In Biochemical Engineering Biotechnology 65: 117-161.
Kurtzman CW, Fell JW. 2000. The Yeast, A Taxonomy Study Fourth Edition. Amsterdam: Elsevier Science B.V. Maharani MD. 2011. Adaptasi
Saccharomyces cerevisiae terhadap hidrolisat asam ubi kayu untuk produksi bioetanol. [thesis]. Bogor: Fakultas Teknologi Industri Pertanian, Institut Pertanian Bogor.
Martini A. 2003. Biotechnology of natural and winery associated strains of Saccharomyces cerevisiae. International Microbiology 6: 207-209.
Meinita NDM, Kang YJ, Jeong TG, Koo MH, Park MS, Hong KY. 2011. Bioethanol production from the acid hydrolisate of the carrageenophyte Kappaphycus alvarezii (cottonii). Journal Applied Phycology 24: 857-862.
Oestergaard S, Olsson L, Nielsen J. 2000. Metabolic engineering of Saccharomyces cerevisiae. Microbiology and Molecular Biology Review 64: 34-50.
Osho A. 2005. Ethanol and sugar tolerance of wine yeasts isolated from fermenting cashew apple juice. African Journal of Biotechnology 4: 660-662.
Prescott SC, Dunns CG. 1981. Industrial Microbiology. Connecticut: The AVI. Publ. Co. Inc. West Port.
Setyaningsih D, Windarwati S, Khayati I, Muna N, Hernowo P. 2012. Acid hydrolisis technique and yeast adaptation to increase red macroalgae bioethanol production. International Journal of Environment and Bioenergy 3(2): 98-110. Singh PG, Volpe G, Creely MC, Grotsch H,
Geli MI, Petrov D. 2006. The lag phase and G1 phase of a single yeast cell monitored by raman microspectroscopy. Journal Raman Spectroscopy 37: 858-864. Sutarto I. 2009. Challenges of biofuel industry in indonesian. Naskah Seminar Nasional pada Dies Natalis Fakultas Teknologi Pertanian IPB, 16 Oktober 2009, Bogor. Taherzadeh MJ, Karimi K. 2007. Acid-based
hydrolysis process for ethanol from lignocellulose material: A review. Journal Bioresources 2(3): 472-499.
Timson DJ. 2007. Galactose metabolism in Saccharomyces cerevisiae: A review. Dynamic Biochemistry 3: 63-73.
Volk, Thomas J. 2000. Tom Volk’s Fungus of The Month. Madison: University of Wisconsin.
8
LAMPIRAN
111
Lampiran 1 Komposisi berbagai media dan larutan yang digunakan dalam penelitian A. Komposisi media PSA (Potato Sucrose Agar) (dalam 1 L)
Kentang 200 g Gula Sukrosa 20 g Agar-agar (swallow) 20 g Akuades 1000 mL
B. Komposisi media YMGP (Yeast Malt Glucose Pepton) (dalam 1 L) Yeast extract 5 g/L
Malt 5 g/L Glukosa 40 g/L Pepton 5 g/L Akuades 1000 mL
C. Komposisi larutan DNS (dalam 1,4 L) Asam 3,5 dinitrolisilat 10,6 g NaOH 19,8 g Na-K Tartat 306 g Na-Metabisulfit 8,3 g Akuades 1416 mL
Lampiran 2 Perhitungan produksi bioetanol, efisiensi fermentasi, efisiensi substrat, dan kadar gula pereduksi
A. Produksi bioetanol
Produksi bioetanol dalam hidrolisat (% v/v) = Volume destilat (mL) x (Kadar etanol/100) x 100% Volume hidrolisat setelah fermentasi (mL)
B. Efisiensi fermentasi
Efisiensi fermentasi (%) = Konsentrasi etanol yang diperoleh aktual (% b/v) x 100% Konsentrasi etanol teoritis
Konsentrasi etanol teoritis = So x 0,51*
Keterangan: *Nilai etanol yang terbentuk pada persamaan glikolisis C. Efisiensi Substrat
Efisiensi substrat (%) = So – S x 100% So
Keterangan: So = gula pereduksi awal/sebelum fermentasi (% b/v) S = gula pereduksi setelah fermentasi (% b/v) D. Kadar Gula Pereduksi (GP)
Asumsi : menggunakan kurva standar glukosa (DNS 7) dengan persamaan Y= 0,003x – 0,166
GP setelah pengenceran (ppm) = (absorbansi + 0,166) 0,003
GP (ppm) = GP setelah pengenceran x faktor pengenceran (fp) GP (% b/v) = GP (ppm)
10.000
11
Kurva standar glukosa (DNS 7) Kurva standar glukosa (DNS 8)
Kurva standar glukosa (DNS 9) Kurva standar glukosa (DNS 11)
Lampiran 3 Diagram alir pengukuran gula pereduksi
Diambil 1 mL sampel cairan hidrolisat dan dimasukkan kedalam tabung reaksi kecil dilakukan pengenceran (FP) dengan akuades
1 mL sampel hidrolisat yang telah dilakukan pengenceran, kemudian ditambahkan 3 mL larutan DNS
Dipanaskan pada gelas kimia yang berisi akuades selama ± 5 menit, kemudian didinginkan
pengukuran sampel dilakukan dengan spektrofotometer dengan 550 nm, dan didapatkan hasil nilai absorbansi sampel
Blanko : 1 mL akuades + 3 mL DNS y = 0,003x - 0,1666
R² = 0,9968
-0,2 0 0,2 0,4 0,6 0,8
0 100 200 300 400
A b so rb a n ppm
y = 0,0029x - 0,1614 R² = 0,9972
-0,2 0 0,2 0,4 0,6 0,8
0 100 200 300 400
A b so rb a n ppm
y = 0,0031x - 0,1947 R² = 0,988
-0,2 0 0,2 0,4 0,6 0,8
0 100 200 300 400
A b so rb a n ppm
y = 0,0031x - 0,1777 R² = 0,9939
-0,2 0 0,2 0,4 0,6 0,8 1
0 100 200 300 400
111
Lampiran 4 Konsentrasi gula pereduksi selama pertumbuhan S. cerevisiae dan selama produksi bioetanol
A. Konsentrasi gula pereduksi selama pertumbuhan S. cerevisiae pada media YMGP dan hidrolisat K. alvarezii
Waktu inkubasi (jam)
Gula Pereduksi (GP) (%b/v)
YMGP Hidrolisat
0 4,73 7,58
6 3,37 6,79
12 1,45 6,64
18 0,32 5,67
24 0,24 4,72
30 0,22 2,89
36 0,21 2,74
42 0,19 2,5
48 0,19 1,98
54 0,19 2,59
60 0,18 2,41
B. Konsentrasi gula pereduksi pada beberapa waktu inkubasi dan konsentrasi inokulan
Perlakuan Gula Pereduksi (%b/v)
Waktu inkubasi (jam) (pada konsentrasi inokulan 107 sel mL-1)
0 8,79
48 7,51
72 6,99
96 5,82
120 3,93
144 4,24
168 3,53
192 3,51
Konsentrasi inokulan (sel mL-1) (pada 168 jam inkubasi)
0 7,93
1,00E+05 5,20
1,00E+06 2,38
1,00E+07 2,76