STATISTIK DEMOGRAFI
Bemisia tabaci
Gennadius
(HEMIPTERA: ALEYRODIDAE) PADA TANAMAN KEDELAI
VARIETAS EDAMAME DAN WILIS
SITI RIZKAH SAGALA
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Statistik Demografi
Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) pada Tanaman Kedelai Varietas Edamame dan Wilis adalah benar karya saya dengan arahan dari dosen pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2015
Siti Rizkah Sagala
ABSTRAK
SITI RIZKAH SAGALA. Statistik Demografi Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) pada Tanaman Kedelai Varietas Edamame dan Wilis. Dibimbing oleh HERMANU TRIWIDODO.
B. tabaci adalah serangga polifag yang menyerang lebih dari 600 spesies tanaman. B. tabaci merupakan hama penting pada tanaman kedelai. Penelitian ini bertujuan untuk meneliti biologi dan statistik demografi B. tabaci pada tanaman kedelai varietas Edamame dan Wilis. Penelitian dilakukan di Laboratorium WiSH Bogor Indonesia pada bulan Januari sampai April 2015. Penelitian menunjukkan adanya perbedaan biologi B. tabaci yang dipelihara pada varietas Edamame dan Wilis. Masa inkubasi telur, stadia nimfa instar 4 dan pupa serta siklus hidup B. tabaci pada varietas Edamame lebih singkat dibandingkan dengan varietas Wilis. Keperidian B. tabaci pada varietas Edamame lebih kecil dibandingkan dengan varietas Wilis. Laju reproduksi kotor (GRR), laju reproduksi bersih (Ro), laju pertumbuhan intrinsik (r) dan waktu berlipat ganda B. tabaci pada varietas Edamame masing-masing adalah 28.365 ± 0.530, 9.925 ± 0.182, 0.095 ± 0.001 dan 7.330 ± 0.069. Laju reproduksi kotor (GRR), laju reproduksi bersih (Ro), laju pertumbuhan intrinsik (r) dan waktu berlipat ganda B. tabaci pada varietas Wilis masing-masing adalah 46.509 ± 0.725, 16.428 ± 0.252, 0.102 ± 0.001 dan 6.808 ± 0.042. Pertumbuhan populasi B. tabaci pada varietas Edamame lebih lambat dibandingkan dengan varietas Wilis.
ABSTRACT
SITI RIZKAH SAGALA. The Statistic Demographic of Bemisia tabaci
Gennadius (Hemiptera: Aleyrodidae) on Soybean Varieties, Edamame and Wilis. Supervised by HERMANU TRIWODODO.
B. tabaci is polyphagous insect and has approximately 600 host plants. B. tabaci is an important pest on soybean. These researches were to study the biology and statistic demographic of B. tabaci on two soybean varieties, Edamame and Wilis. These researches were conducted at Wish Laboratory Bogor Indonesia, from January to April 2015. It is shown there are differences of the biology and statistic demographic of B. tabaci reared on Edamame and Wilis. Egg incubation period, fourth instar and pupae stadia, and life cycle of B. tabaci reared on Edamame are shorter than that reared on Wilis. The fecundity of B. tabaci
reared on Edamame are lower than that reared on Wilis. The gross reproduction rate (GRR), net reproduction rate (Ro), intrinsic growth rate (r) and doubling time of B. tabaci on Edamame respectively are 28.365 ± 0.530, 9.925 ± 0.182, 0.095 ± 0.001 and 7.330 ± 0.069. The gross reproduction rate (GRR), net reproduction rate (Ro), intrinsic growth rate (r) and doubling time of B. tabaci on Wilis respectively are 46.509 ± 0.725, 16.428 ± 0.252, 0.102 ± 0.001 and 6.808 ± 0.042. The population growth of B. tabaci reared on Edamame are slower than that reared on Wilis.
©
Hak Cipta milik IPB, tahun 2015 Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
STATISTIK DEMOGRAFI
Bemisia tabaci
Gennadius
(HEMIPTERA: ALEYRODIDAE) PADA TANAMAN KEDELAI
VARIETAS EDAMAME DAN WILIS
SITI RIZKAH SAGALA
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
PRAKATA
Segala puji dan syukur penulis panjatkan kehadirat Allah SWT yang telah melimpahkan segala rahmat dan karunia-Nya sehingga penulis dapat menyelesaikan tugas akhir yang berjudul “Statistik Demografi Bemisia tabaci
Gennadius (Hemiptera: Aleyrodidae) pada Tanaman Kedelai Varietas Edamame dan Wilis”. Penulisan tugas akhir penelitian ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Pertanian di Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
Penulis mengucapkan terima kasih kepada Ayahanda Erwin Sagala, Ibunda Halimah Tussaddiah dan adik penulis Syifa Fahrani Sagala serta keluarga besar yang telah memberikan semangat, dukungan dan motivasi serta mendoakan penulis sehingga dapat menyelesaikan tugas akhir ini. Penulis juga mengucapkan terima kasih yang sebesar-besarnya kepada Dr. Ir. Hermanu Triwidodo, M.Sc. selaku dosen pembimbing skripsi yang selalu memberikan bimbingan, ilmu pengetahuan, saran, arahan, dan masukan kepada penulis. Ucapan terima kasih juga disampaikan kepada Prof. Dr. Ir. Dadang, M.Sc. selaku dosen pembimbing akademik dan Dr. Ir. Supramana, M.Si. selaku dosen penguji yang telah memberikan kritik dan saran dalam penulisan tugas akhir ini. Terima kasih juga diucapkan kepada Pak Adi Toxs, Pak Sodik, Asep, Mba Dila, Bang Rudi, Kak Ali, Mas Wildan, Mbak Saksak, Agustina Diprianti, Iyun, Selvia Wulan Hajijah, Lina F. Jannah, Phor Bho Ayuwati, Listihani, serta teman-teman lainnya di Departemen Proteksi Tanaman yang tidak bisa disebutkan satu per satu yang telah membantu penulis dalam penelitian dan penulisan tugas akhir ini.
Penulis menyadari bahwa masih banyak kekurangan dalam skripsi ini. Oleh karena itu, penulis mengharapkan saran, masukan dan kritik yang dapat membangun dan memotivasi penulis agar dapat menulis karya tulis yang lebih baik. Semoga Skripsi ini dapat bermanfaat bagi penulis maupun pembaca.
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
BAHAN DAN METODE 3
Tempat dan Waktu 3
Bahan dan Alat 3
Metode Penelitian 3
Persiapan Tanaman Uji 3
Perbanyakan B. tabaci 4
Infestasi B. tabaci 4
Pembuatan Preparat B. tabaci 5
Pengamatan Biologi B. tabaci 5
Neraca Kehidupan dan Statistik Demografi B. tabaci 5 Pendugaan Keragaman Data Menggunakan Metode Jackknife 6 Rancangan Percobaan 6
HASIL DAN PEMBAHASAN 7
Pengaruh Varietas terhadap Biologi B. tabaci 7
Pengaruh Varietas terhadap Statistik Demografi B. tabaci 11
Pembahasan Umum 15
SIMPULAN DAN SARAN 17
Simpulan 17
Saran 17
DAFTAR PUSTAKA 18
LAMPIRAN 23
DAFTAR TABEL
.1 Biologi B. tabaci pada tanaman kedelai varietas Edamame dan kedelai
varietas Wilis 8
.2 Statistik demografi B.tabaci pada tanaman kedelai Edamame dan
tanaman kedelai Wilis 13 3 Lampiran 1 siklus hidup B. tabaci pada tanaman kedelai Edamame 24 4 Lampiran 2 siklus hidu B. tabaci pada tanaman kedelai Wilis 25 5 Lampiran 3 lama hidup, praoviposisi, oviposisi dan keperidian
B. tabaci pada tanaman kedelai Edamame 26 6 Lampiran 4 lama hidup, praoviposisi, oviposisi dan keperidian
B. tabaci pada tanaman kedelai Wilis 27
.7 Lampiran 5 neraca kehidupan B. tabaci tanaman kedelai Edamame 28
8 Lampiran 6 neraca kehidupan B. tabaci tanaman kedelai Wilis 29
DAFTAR GAMBAR
.1 Tanaman kedelai Edamame dan kedelai Wilis untuk perlakuan 3
.2 Tanaman cabai keriting untuk perbanyakan B. tabaci 4
.3 Pemeliharaan kohort B. tabaci 4
.4 Morfologi antena (a), panjang tubuh abdomen (b), seta kauda B. tabaci 7
5 Peluang hidup dan keperidian harian B. tabaci pada tanaman kedelai
PENDAHULUAN
Latar Belakang
Kedelai (Glycine max L.) merupakan komoditas pertanian yang sangat dibutuhkan di Indonesia baik sebagai bahan pangan manusia, pakan ternak, maupun bahan baku industri (Cahyono 2007). Kebutuhan kedelai dalam negeri meningkat setiap tahun seiring pesatnya perkembangan industri pangan dan pakan olahan berbahan baku kedelai. Produksi kedelai dalam negeri mengalami penurunan sebesar 62 990 ton (7.47%) dengan jumlah produksi kedelai nasional sebesar 780 160 ton biji kering pada tahun 2013 (BPS 2014). Salah satu faktor yang menyebabkan produktivitas kedelai menurun adalah serangan hama. Puslitbang Tanaman Pangan (2006) melaporkan bahwa ada 15 spesies hama utama pada pertanaman kedelai, salah satunya adalah Bemisia tabaci. Kehilangan hasil akibat serangan hama ini dapat mencapai 80% hingga gagal panen (Marwoto
et al. 2011).
B. tabaci merupakan salah satu hama yang bersifat polifag dan dapat menyerang lebih dari 600 spesies tanaman baik tanaman sayuran, hias, buah-buahan, dan kacang-kacangan (McKenzie et al. 2004; Oliviera et al. 2001). Sentra pertanaman kedelai Edamame di kabupaten Jember melaporkan bahwa serangan
B. tabaci sangat tinggi pada pertanaman kedelai Edamame. Serangannya dapat mengakibatkan kerusakan langsung maupun tidak langsung pada tanaman. Kerusakan langsung menimbulkan gejala berupa bercak klorosis yang mengakibatkan berkurangnya jumlah klorofil pada daun sehingga mengganggu proses fotosintesis tanaman inang (Van den Ven et al. 2000). Kerusakan tidak langsung berkaitan dengan kemampuannya sebagai vektor berbagai virus yang menyebabkan penyakit pada berbagai tanaman di antaranya tanaman kedelai, tomat dan cabai (Borror et al. 1996; Hidayat et al. 2008; Legg et al. 2002). Tingkat kerusakan dan serangan B. tabaci ditentukan oleh tinggi rendahnya populasi, fase pertumbuhan tanaman, bagian tanaman yang terserang dan ketahanan varietas.
Populasi B. tabaci sering mengalami fluktuasi yang dipengaruhi tanaman inang dan suhu (Subagyo 2010). Menurut Naranjo dan Ellsworth (2005) tanaman inang merupakan salah satu faktor biotik yang dapat mempengaruhi aspek biologi dan kelangsungan hidup suatu organisme. Perbedaan varietas tanaman juga berpengaruh terhadap biologi B. tabaci seperti perkembangan stadia, peluang hidup, lamanya hidup dan laju reproduksi (Kim dan Lee 2002). Penelitian Silva (2012) menyatakan bahwa penggunaan beberapa varietas kedelai berpengaruh terhadap siklus hidup B. tabaci. Kualitas dan kuantitas tanaman inang juga mempengaruhi perkembangan B. tabaci (Du et al. 2004; Razmjou et al. 2006). Selain itu, efek dari tanaman inang sebelumnya dapat mempengaruhi perkembangan B. tabaci pada tanaman inang yang baru. Hal ini disebabkan oleh adanya nutrisi atau senyawa-senyawa kimia pada tanaman inang, yang memiliki peranan penting dalam perkembangan B. tabaci.
2
akan memberikan pengaruh lebih besar daripada tanaman inang sebelumnya. Hal ini disebabkan adanya perbedaan nutrisi atau senyawa kimia pada tanaman cabai keriting dengan tanaman kedelai Edamame dan Wilis. Nutrisi atau senyawa-senyawa kimia yang dimiliki varietas Edamame dan Wilis seperti protein, karbohidrat, lemak nabati, isoflavon dapat mengganggu proses makan dan kehidupan B. tabaci. Tinggi rendahnya protein yang dikonsumsi berpengaruh terhadap reproduksi serangga (Chapman 1982). Selain aktivitas makan yang terganggu, nutrisi atau senyawa kimia tanaman juga akan mempengaruhi pertumbuhan, perkembangan, keperidian, dan mortalitas. Hal ini dapat dijadikan sebagai salah satu faktor pembatas perkembangan populasi B. tabaci di lapang.
Pengaruh dari varietas Edamame dan Wilis terhadap peluang hidup dan keperidian B. tabaci dapat diketahui dengan merancang neraca kehidupan. Naranjo dan Ellsworth (2005) menyatakan bahwa bentuk neraca kehidupan merupakan metode untuk menggambarkan dan mengukur kematian dalam suatu populasi. Informasi yang telah diperoleh dari neraca kehidupan dapat dilanjutkan untuk menentukan peubah statistik demografi lainnya (Price 1997; Wilson dan Bossert 1971).
Statistik demografi diartikan sebagai analisis secara kuantitatif pertumbuhan populasi serangga dalam hubungannya dengan kelangsungan hidup, keperidian dan pola pertumbuhan populasi (Zeng et al. 1983). Penelitian mengenai pengaruh varietas kedelai terhadap biologi B. tabaci telah banyak dilakukan di beberapa negara. Namun, informasi mengenai biologi dan statistik B. tabaci pada varietas kedelai Edamame dan Wilis di Indonesia masih terbatas. Penelitian neraca kehidupan dan statistik demografi dapat menjadi informasi dasar dalam pengambilan keputusan untuk mengendalikan populasi B. tabaci di lapang.
Tujuan Penelitian
Penelitian ini bertujuan mengetahui pengaruh kedelai varietas Edamame dan varietas Wilis terhadap biologi dan statistik demografi B. tabaci.
Manfaat Penelitian
BAHAN DAN METODE
Tempat dan Waktu Penelitian
Penelitian dilaksanakan di Laboratorium WiSH Indonesia, Bogor. Identifikasi B. tabaci dilakukan di Laboratorium Biosistematika Serangga, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. B. tabaci diperoleh dari tanaman cabai keriting di Laboratorium WiSH. Penelitian ini dilaksanakan pada bulan Januari sampai April 2015.
Bahan dan Alat
Bahan yang digunakan dalam penelitian ini adalah benih kedelai Edamame dan Wilis masing-masing sebanyak 100 biji, benih cabai keriting varietas Lado F1, imago B. tabaci, media tanam dari campuran tanah dan kompos 2:1, pupuk NPK dan pupuk KCL. Bahan yang digunakan dalam identifikasi B. tabaci adalah alkohol 50%, 80%, 95%, aquades, larutan KOH 10%, acid alkohol 50%, acid fuchsin, carbol xylene, minyak cengkeh dan balsam kanada sebagai media perekat dalam pembuatan preparat slide.
Alat yang digunakan sebagai berikut: cangkul, gunting, aspirator, loop, ajir, karton hitam, polybag ukuran 25 cm x 25 cm dan ukuran 30 cm x 30 cm, tali rafia, lem, alat tulis, kurungan serangga, ajir bambu, termometer. Alat yang digunakan untuk identifikasi adalah buku kunci identifikasi serangga karangan Borror et al.
(1996) dan Martin (1987), kantung plastik, tabung reaksi, cawan sirakus, kaca objek, penutup preparat, kompor listrik, mikroskop stereo, mikroskop cahaya, kamera, dan alat tulis.
Metode Penelitian Persiapan Tanaman Uji
Benih kedelai Edamame dan Wilis dipilih dengan tampilan permukaan benih tidak keriput, masing-masing varietas sebanyak 100 biji. Benih tersebut ditanam pada 100 buah polybag ukuran 30 cm x 30 cm yang diisi media tanam tanah dan kompos 2:1 sebanyak 4 kg/polybag serta ditambahkan pupuk NPK dengan dosis 0.5 g/polybag. Setiap polybag ditanam masing-masing 2 benih kedelai. Tanaman dipelihara, disiram setiap hari dengan air bersih dan diberi pupuk. Pemupukan dilakukan 3 kali yaitu saat awal penanaman, saat pembungaan yaitu 3 minggu setelah tanam (3 MST) dan saat pengisian polong (5 MST). Penyiangan gulma dilakukan pada umur tanaman berumur 2 MST dan diberi ajir sebagai penunjang pertumbuhan tanaman.
4
Perbanyakan B. tabaci
Benih cabai keriting varietas Lado F1 untuk perbanyakan B. tabaci ditanam pada 10 polybag ukuran 25 cm x 25 cm yang diisi dengan media tanam tanah dan kompos 2:1. Setiap polybag ditanam 3 benih cabai kemudian dipelihara di dekat laboratorium WiSH. Imago B. tabaci didapat dari pertanaman cabai di Laboratorium WiSH. B. tabaci diinfestasi, dipelihara, dan diperbanyak pada tanaman cabai keriting yang telah berumur 4 MST. Tanaman cabai keriting tersebut dimasukkan ke dalam kurungan serangga yang berukuran 100 cm x 50 cm x 150 cm sampai jumlahnya cukup untuk digunakan pada perlakuan.
Gambar 2 Tanaman cabai keriting dalam kurungan serangga untuk perbanyakan B. tabaci
Infestasi B. tabaci
Populasi kohort merupakan sejumlah individu yang memiliki umur seragam (Begon et al. 2008). Imago B. tabaci dalam populasi kohort diinfestasikan pada daun trifoliet muda tanaman kedelai varietas Edamame dan varietas Wilis yang berumur 4 MST sebanyak 5 ekor imago B. tabaci/polybag (2 ekor betina dan 3 ekor jantan). Daun tersebut diberi karton hitam yang mengelilingi bagian tangkainya dan dikurung dengan mika plastik berbentuk silindris (d= 15 cm, t= 15 cm) dengan bagian atas dan bawahnya menggunakan kain kasa. Tujuan pemberian kertas karton hitam yaitu untuk memudahkan pengamatan saat B. tabaci ganti kulit. Setelah serangga imago betina menghasilkan telur, serangga tersebut dikeluarkan. Telur diamati setiap hari sampai menjadi imago dan imago tersebut mati.
5
Pembuatan Preparat B. tabaci
Imago B. tabaci diambil beberapa ekor dan diawetkan ke dalam alkohol 70% untuk dibuat preparat dan diidentifikasi. Imago tersebut dibuat preparat slide dengan cara imago B. tabaci dimasukkan ke dalam alkohol 95% lalu dipindahkan ke dalam KOH 10%, direbus hingga transparan dan dikeluarkan isi abdomen B. tabaci dengan ditusuk-tusuk bagian abdomennya di bawah mikroskop stereo. Setelah itu B. tabaci dicuci dengan akuades sebanyak dua kali dan dimasukkan ke dalam acid alkohol 50% selama 10 menit. Selanjutnya B. tabaci tersebut dimasukkan ke dalam acid fuchsin selama 20 menit dan direndam dalam alkohol 80%, 95% masing-masing selama 10 menit. Kemudian B. tabaci dimasukkan ke dalam carbol xylene, alkohol 95% dan minyak cengkeh masing-masing selama selama 10 menit. B. tabaci tersebut diletakkan di atas kaca objek dan diberi media
canada balsam. Preparat mikroskop yang telah dibuat dipanaskan ke dalam hot plate dan diidentifikasi dengan kunci identifikasi Martin (1987).
Pengamatan Biologi B. tabaci
Pengamatan biologi B. tabaci dilakukan setiap hari dengan mengamati individu yang masih hidup atau sudah mati, ganti kulit dengan adanya eksuvia, dan jumlah telur yang dihasilkakan. Siklus hidup B. tabaci dihitung sejak telur diletakkan oleh imago betina pada tanaman kedelai Edamame dan Wilis sampai menjadi imago dan meletakkan telur untuk pertama kalinya. Selama perkembangan menjadi imago, B. tabaci mengalami 4 stadia instar yang ditandai dengan pergantian kulit dan morfologi instar. Praoviposisi B. tabaci dihitung sejak hari pertama menjadi imago hingga imago tersebut meletakkan telur untuk pertama kalinya. Pengamatan lama hidup B. tabaci dihitung sejak hari pertama menjadi imago hingga imago tersebut mati. Keperidian B. tabaci diperoleh dari jumlah telur yang dihasilkan oleh setiap imago selama hidupnya. Data yang dipperoleh disusun dalam bentuk tabel biologi B. tabaci (life table).
Neraca Kehidupan dan Statistik Demografi B. tabaci
Peubah pertumbuhan populasi B. tabaci sangat ditentukan oleh jumlah individu yang hidup setiap hari sehingga diperoleh data peluang hidup (lx) pada berbagai umur. Pengamatan keperidian harian (mx) adalah rata-rata jumlah telur yang dihasilkan oleh setiap imago setiap harinya pada berbagai umur (x). Data yang didapat dari pengamatan peluang hidup dan keperidian harian digambarkan dalam bentuk kurva dan diperoleh neraca kehidupan.
6
Laju reproduksi bersih (Ro) merupakan rata-rata jumlah keturunan (dari fase pertama dalam suatu siklus hidup) yang dihasilkan oleh setiap imago (Begon et al.
2008). Laju pertumbuhan intrinsik (r) menggambarkan laju peningkatan populasi dengan sumberdaya yang tidak terbatas (Price 1997). Rataan lama generasi (T) merupakan rataan waktu yang dibutuhkan sejak telur diletakkan sampai menjadi imago dan meletakkan telur untuk pertama kalinya (Price 1997; Begon et al.
2008). Doubling time (DT) yaitu waktu yang dibutuhkan untuk populasi B. tabaci
menjadi berlipat ganda.
Pendugaan Keragaman Data Menggunakan Metode Jacknife
Keragaman data diolah dengan menggunakan metode jacknife. Menurut Iskandar et al. (2013) metode jackknife diperkenalkan Tukey untuk melakukan uji hipotesis dan menghitung selang kepercayaan. Jackknife digunakan dalam statistika inferensia untuk menduga bias dan galat baku suatu statistik. Prinsip pendugaan metode jackknife adalah melakukan perhitungan dengan menghilangkan satu data pengamatan kemudian berdasarkan gugus data baru tersebut dihitung bias dan ragam statistiknya. Data kohort B.tabaci pada tanaman kedelai varietas Edamame dan varietas Wilis diolah dengan menggunakan metode
jackknife. Menurut Marlena (2014) langkah-langkah untuk menghitung statistik demografi B. tabaci menggunakan metode jackknife sebagai berikut:
1. Membuang baris ke-i data kohort asli sehingga terbentuk data baru yang berukuran k = b-1, dengan i = 1, 2, ..., b;
2. a. Menghitung nilai mx, lx, dan lxmx; b. Menyusun nilai Fx = mx dan px;
3. a. Menyusun tabel kehidupan dari nilai yang diperoleh pada langkah 2a; b. Menyusun matriks Leslie dari nilai yang diperoleh pada langkah 2b;
4. Menghitung nilai dugaan GRR, Ro, r, T, dan DT. Statistik demografi r dan T dihitung dua kali. Pertama berdasarkan tabel kehidupan (3a) dan yang kedua dihitung berdasarkan matriks Leslie (3b);
5. Mengulang langkah 1, 2, 3, dan 4 sampai baris yang dibuang adalah baris ke-n data kohort contoh asli;
6. Menghitung dugaan GRR, Ro, r, T, dan DT hasil resampling jackknife; 7. Menghitung dugaan galat baku untuk setiap statistik demografi tersebut; 8. Membuat selang kepercayaan (SK) 95% bagi GRR, Ro, r, T, dan DT.
Rancangan Percobaan
Penelitian ini menggunakan 2 perlakuan yaitu kedelai varietas Edamame dan varietas Wilis dengan 50 ulangan pada masing-masing perlakuan. Data hasil pengamatan diolah dengan menggunakan Microsoft Excel 2007, program
HASIL DAN PEMBAHASAN
Pengaruh Varietas terhadap Biologi B. tabaci
Dinamika populasi merupakan fluktuasi populasi serangga dari waktu ke waktu. Pertumbuhan dan perkembangan populasi serangga ditentukan oleh faktor luar (faktor eksternal) dan faktor dalam dirinya sendiri (faktor internal). Faktor eksternal yang berpengaruh adalah lingkungan dan makanan. Faktor lingkungan yang berpengaruh salah satunya adalah suhu, yang sangat berkaitan terhadap iklim mikro perkembangan populasi serangga (Speight et al. 2008). Berdasarkan hasil pengamatan harian penelitian, suhu rata-rata lingkungan untuk pertumbuhan B. tabaci adalah 25.2oC, suhu minimal 22.5oC dan suhu maksimal 29oC. Bonaro et al. (2007) menyatakan bahwa suhu optimum untuk pertumbuhan populasi B. tabaci adalah 32.5oC.
Imago B. tabaci hasil perbanyakan diambil dan dibuat preparat untuk diidentifikasi. Identifikasi morfologi berdasarkan buku kunci identifikasi Martin
et al. (1987). Hasil identifikasi menunjukkan bahwa imago kutu kebul tersebut merupakan spesies dari Bemisia tabaci Gennadius, ordo Hemiptera, subordo Sternorrhyncha, famili Aleyrodidae, superfamili Aleyrodoidea dengan melihat ciri-ciri morfologinya (Gambar 4).
a b c
Gambar 4 Morfologi antena (a), panjang tubuh (b), seta kauda (c) B. tabaci
Panjang tubuh B. tabaci berkisar 1-1.5 mm dan tubuhnya berwarna kuning. Antena terdiri dari 7 ruas dan antena pada ruas ketiga ukurannya lebih panjang dari ruas lainnya. Sayap imago transparan ditutupi tepung berwarna putih (lilin) dan matanya berwarna merah (Borror et al. 1996). Telur berwarna kekuning-kuningan, ditutupi oleh lilin dan diletakkan di bawah permukaan daun. Setelah 24 jam warna telur akan berubah menjadi coklat. Nimfa instar 1 berbentuk oval, pipih, berwarna putih kehijauan, panjang tubuh 0.22 mm, lebar 0.13 mm dan memiliki tungkai. Nimfa instar 2 berwarna hijau gelap, panjang tubuh 0.28 mm, lebar 0.17 mm, memiliki antena sangat pendek dan tungkainya tereduksi. Pupa berbentuk oval, bagian toraks agak melebar, cembung, abdomennya tampak jelas dan terdapat satu pasang seta kauda pada ujung anal (Kalshoven 1981).
Perkembangan B. tabaci terdiri dari telur, nimfa, pupa dan imago. B. tabaci
mengalami 4 instar, perubahan tiap instar ditandai dengan adanya eksuvia (pergantian kulit). Perkembangan dan pertumbuhan imago B. tabaci berbeda-beda tergantung pada tanaman inang dan suhu.
1 mm
8
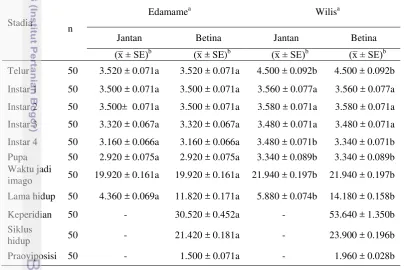
Siklus hidup adalah waktu yang diperlukan mulai dari telur diletakkan sampai menjadi imago dan meletakkan telur pertama. Perbedaan varietas tanaman mempengaruhi siklus hidup B. tabaci. Silva (2012) menyatakan bahwa siklus hidup B. tabaci pada tanaman kedelai varietas IAC-PLI, varietas BRS-242 RR dan TMG-103 RR secara berurutan adalah 21.3, 20.1 dan 19.3 hari pada suhu 25oC. Hasil tersebut menunjukkan bahwa lamanya siklus hidup hasil penelitian Silva berbeda dengan hasil penelitian ini dikarenakan adanya perbedaan lingkungan dan varietas tanaman yang digunakan pada penelitian. Sunjaya (1970) menyatakan bahwa kualitas makanan yang ada dalam tanaman inang dapat menentukan pertumbuhan, perkembangan, kesuburan, mortalitas dan keperidian serangga. Perbedaan varietas juga berpengaruh nyata terhadap lama inkubasi telur, lama stadia instar dan pupa pada tanaman kedelai varietas Edamame dan varietas Wilis (Tabel 1).
Tabel 1 Biologi B. tabaci pada tanaman kedelai varietas Edamame dan varietas Wilis
Angka pada baris dan jenis kelamin yang sama yang diikuti dengan huruf yang sama tidak berbeda nyata berdasarkan uji t pada taraf nyata 5%. b x̅: rata-rata, SE: standar error.
9 (Agustini 2013). Tertundanya waktu reproduksi akan berdampak terhadap waktu generasi menjadi lebih lama dan peningkatan mortalitas sebelum reproduksi (Kozlowski 1992). Laju perkembangan yang terhambat akan menurunkan kebugaran serangga dan memperlambat inisiasi reproduksi serangga (Begon et al. 2008). Inisiasi reproduksi yang lambat akan memperpanjang siklus hidup suatu generasi. Siklus hidup yang panjang akan memiliki lama generasi yang panjang pula.
Masa inkubasi telur pada tanaman kedelai varietas Edamame lebih pendek dibandingkan dengan varietas Wilis. Masa inkubasi telur pada varietas Edamame adalah 3.520 ± 0.071 hari, sedangkan pada varietas Wilis adalah 4.500 ± 0.092 hari. Lama masa inkubasi telur berkorelasi dengan lama perkembangan stadia nimfa dan pupa B. tabaci. Hal ini disebabkan oleh adanya pengaruh terhadap imago betina B. tabaci yang berasal dari tanaman cabai keriting, dimana nutrisi atau senyawa kimia pada tanaman cabai keriting berbeda dengan tanaman kedelai sehingga berpengaruh terhadap masa inkubasi telur yang dihasilkan.
Waktu perkembangan stadia nimfa instar 1, instar 2 dan instar 3 B. tabaci
pada tanaman kedelai Edamame tidak berbeda dengan tanaman kedelai Wilis. Menurut Marwoto dan Inayati (2011) nimfa instar 1 yang baru menetas aktif bergerak mencari tempat tinggal dan nutrisi makanan yang sesuai sehingga nimfa hanya makan sedikit. Selanjutnya nimfa instar 2 dan instar 3 akan diam dan menetap setelah menemukan tempat dan makanan yang sesuai karena tungkainya tereduksi. Nimfa tersebut berada dalam masa pencocokan dan penghisapan cairan tanaman sehingga perbedaan varietas kedelai terhadap nimfa instar 1, 2 dan 3 tidak berpengaruh. Namun saat nimfa instar 4, perbedaan varietas kedelai berpengaruh terhadap lamanya waktu stadia nimfa instar 4 dan pupa. Nimfa instar 4 telah beradaptasi dengan lingkungannya dan menghisap banyak cairan tanaman sebelum stadia pupa sehingga nutrisi yang dikonsumsinya berpengaruh terhadap perkembangannya. Hal ini disebabkan oleh pengaruh nutrisi yang berbeda pada tanaman inang dan faktor nutrisi tersebut memerlukan waktu untuk memberikan pengaruh terhadap B. tabaci pada tanaman kedelai varietas Edamame dan varietas Wilis. Menurut Shah dan Liu (2013) tanaman inang memiliki pengaruh yang penting terhadap kelangsungan hidup nimfa B. tabaci.
Nutrisi atau senyawa-senyawa kimia pada tanaman sangat berpengaruh terhadap perkembangan B. tabaci. Namun, nutrisi atau senyawa kimia tersebut memerlukan waktu untuk memberikan efek terhadap pertumbuhan dan perkembangan B. tabaci. Hasil ini sejalan dengan penelitian Shah dan Liu (2013) yang menyatakan bahwa tanaman inang sebelumnya memberikan pengaruh terhadap B. tabaci, kemudian seiring berjalannya waktu tanaman inang yang baru yang memiliki peranan penting dan memberikan pengaruh yang lebih besar daripada tanaman inang sebelumnya. Penundaan waktu perkembangan nimfa dalam dinamika populasi memberikan dampak positif terhadap pengendalian hama dan berpengaruh terhadap praoviposisi dan oviposisi (Wirianti 2006). Praoviposisi yang terhambat akan berpengaruh terhadap keperidian hama.
10
lebih menyukai meletakkan telur pada tanaman kedelai Wilis. Hal ini sesuai dengan McAuslane et al. (1996) yang menyatakan bahwa jumlah trikoma daun berkorelasi positif dengan oviposisi B. tabaci pada tanaman kedelai. Pendapat tersebut sejalan dengan Tama (2011) yang menyatakan bahwa imago B. tabaci
lebih menyukai hidup pada trikoma yang berjumlah banyak daripada trikoma yang berjumlah sedikit pada tanaman kedelai. Trikoma yang berjumlah banyak dapat berfungsi sebagai pegangan atau tempat berjangkar imago B. tabaci pada saat angin kencang dan hujan. Selain itu, trikoma yang rapat juga befungsi sebagai penahan telur agar tidak terbawa air pada saat hujan dan tidak terjatuh atau terbang saat angin kencang. Butler dan Wilson (1984) menyatakan bahwa imago
B. tabaci lebih menyukai meletakkan telur pada varietas kapas yang memiliki trikoma lebih banyak daripada varietas kapas yang memiliki trikoma sedikit karena adanya rangsangan nutrisi atau senyawa kimia pada daun. Ketertarikan imago B. tabaci untuk hidup dan meletakkan telur pada tanaman kedelai juga dipengaruhi oleh adanya senyawa kimia pada daun, gula, tanin, konsentrasi kelenjar gosipol, O-dihidroksifenol dan nutrisi tanaman (Bentz et al. 1995; Butter
et al. 1990). Komposisi zat kimia seperti lupeol (triterpenol) pada tanaman kedelai dapat mempengaruhi perilaku makan kutukebul (Lambert et al. 1995). Varietas kedelai dengan kandungan lupeol rendah cenderung disukai kutukebul.
Lama hidup B. tabaci pada tanaman kedelai Edamame lebih singkat dibandingkan pada tanaman kedelai Wilis. Lama hidup imago betina B. tabaci
pada varietas Edamame 11.820 hari sedangkan pada varietas Wilis 14.180 hari. Lama hidup B. tabaci dipengaruhi oleh nutrisi dari tanaman inang yang dikonsumsi untuk kelangsungan hidupnya. Menurut lembaga penelitian Soyfoods Association of North America (2005), kandungan nutrisi pada tanaman kedelai Edamame lebih tinggi dibandingkan tanaman kedelai kuning biasa. Namun kadar protein kedelai Edamame lebih sedikit daripada kedelai Wilis. Protein merupakan nutrisi yang sangat dibutuhkan oleh serangga dalam reproduksi (Chapman 1982). Semakin sedikit protein yang dikonsumsi B. tabaci maka akan menurunkan reproduksi serangga sehingga mempengaruhi keperidian B. tabaci. Syahputra et al. (2002) menyatakan bahwa protein yang diserap oleh Croccidolomia pavonana
dalam jumlah yang rendah mampu menurunkan keperidian dan mempersingkat lama hidup. Rendahnya kualitas nutrisi yang dimiliki tanaman inang berpengaruh terhadap biologi serangga sehingga serangga kekurangan nutrisi yang dibutuhkan untuk perkembangannya (Panizzi dan Parra 2009). Nutrisi atau senyawa kimia yang dimiliki masing-masing varietas tanaman, juga dapat meningkatkan ketahanan tanaman terhadap seranga hama (Agustini 2013). Adanya senyawa kimia pada tanaman inang menyebabkan B. tabaci tidak mampu hidup lebih lama pada tanaman kedelai varietas Edamame. Hal ini disebabkan oleh adanya sistem ketahanan tanaman yang diinduksi oleh senyawa kimia tertentu pada kedelai varietas Edamame.
11 yang dihasilkan seekor imago betina B. tabaci mencapai 28-300 telur tergantung pada tanaman inang dan suhu lingkungan (Hirano et al. 2002). Rata-rata jumlah telur yang dihasilkan oleh imago betina B. tabaci pada tanaman kedelai sebanyak 70 telur, pada tanaman mentimun sebanyak 66 telur, pada tanaman tomat sebanyak 167.6 telur dan sebanyak 77.5 telur pada tanaman ubi kayu dengan kondisi suhu 25oC (Tsai dan Wang 1996). Reproduksi serangga dipengaruhi oleh kandungan protein yang diperolehnya. Sedikitnya jumlah telur yang dihasilkan pada tanaman kedelai varietas Edamame disebabkan oleh protein yang diserap imago betina B. tabaci belum mampu memenuhi kebutuhan nutrisinya. Selain itu, nutrisi yang tidak terpenuhi juga akan berpengaruh terhadap waktu keperidian.
Waktu keperidian yang singkat berhubungan dengan mortalitas imago betina B. tabaci yang cepat. Natawigena (1990) menyatakan bahwa semakin lama umur imago betina maka akan semakin lama pula kesempatan bertelur atau melahirkan. Menurut Begon et al. (2008) laju perkembangan yang cepat dapat meningkatkan kebugaran serangga dan mempercepat inisiasi reproduksi. Inisiasi reproduksi lebih awal dapat diartikan sebagai mempercepat siklus hidup suatu generasi. Lambatnya siklus hidup B. tabaci pada tanaman kedelai varietas Wilis berbanding lurus dengan terhambatnya waktu reproduksi. Waktu reproduksi yang terhambat dapat menurunkan laju reproduksi. Li et al. (2005) menyatakan bahwa terhambatnya waktu reproduksi dapat meningkatkan mortalitas serangga. Laju reproduksi merupakan salah satu faktor yang menentukan B. tabaci untuk memperbanyak populasinya. Populasi B. tabaci yang tinggi dapat meningkatkan kerusakan yang tinggi pula pada tanaman kedelai di lapang.
Tanaman kedelai varietas Edamame menyebabkan B. tabaci kekurangan nutrisi terutama protein sehingga kemampuan reproduksinya menjadi berkurang. Kemampuan reproduksi yang menurun dapat menyebabkan populasi B. tabaci
menjadi rendah. Hasil penelitian ini menunjukkan bahwa biologi B. tabaci pada tanaman kedelai varietas Edamame berbeda dengan varietas Wilis. Hal ini diduga adanya nutrisi yang berbeda dan senyawa kimia tertentu pada tanaman inang yang berpengaruh terhadap biologi B. tabaci pada tanaman kedelai varietas Edamame dan varietas Wilis.
Pengaruh Varietas terhadap Statistik Demografi B. tabaci
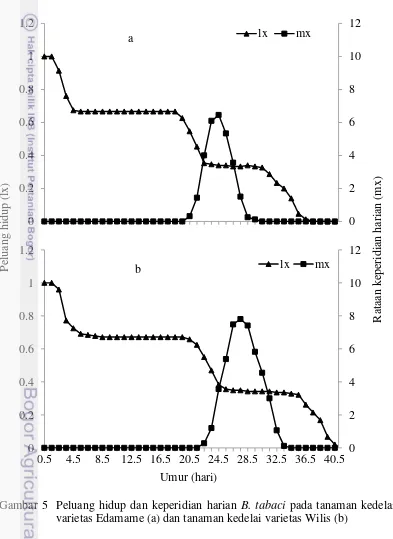
Setiap organisme memiliki variasi jangka hidup terbatas, yang dapat menentukan karakteristik kelangsungan hidupnya dalam suatu populasi. Statistik demografi merupakan suatu analisis secara kuantitatif untuk menduga pertumbuhan populasi suatu organisme. Pertumbuhan populasi dapat dihitung berdasarkan peluang hidup (lx) dan rata-rata jumlah keturunan yang dihasilkan imago betina (mx). Peluang hidup B. tabaci pada tanaman kedelai varietas Wilis lebih besar daripada varietas Edamame. B. tabaci pada kedelai Wilis mengalami mortalitas saat umur 40.5 hari, sedangkan pada kedelai Edamame mortalitasnya saat umur 37.5 hari. Hal ini disebabkan oleh adanya nutrisi atau senyawa kimia tertentu dalam tanaman yang menginduksi ketahanan tanaman sehingga mempercepat mortalitas B. tabaci.
12
kurva tipe II menunjukkan laju kematian populasi suatu individu yang konstan, dan tipe III adalah kematian populasi yang besar yang terjadi saat waktu muda. Populasi yang memiliki angka kematian individu yang konstan akan memiliki populasi yang hampir sama pada generasi berikutnya. Lamanya waktu proses kematian memiliki peranan penting dalam perkembangan suatu populasi. Semakin lama umur imago betina maka akan semakin lama pula pertumbuhan dan perkembangan populasinya di lapang.
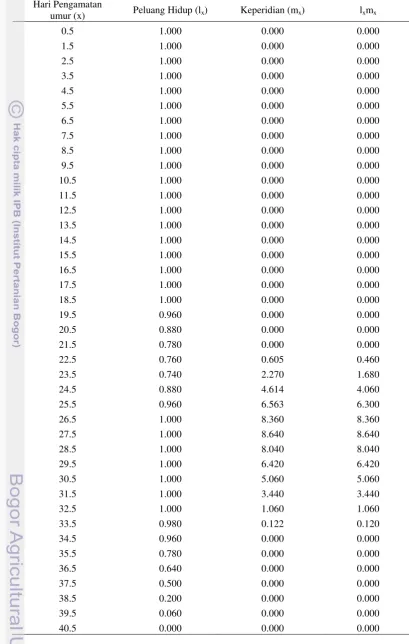
Gambar 5 Peluang hidup dan keperidian harian B. tabaci pada tanaman kedelai varietas Edamame (a) dan tanaman kedelai varietas Wilis (b)
Banyaknya jumlah telur yang dihasilkan oleh setiap imago betina B. tabaci
13 Wilis (Gambar 5). Keperidian harian tertinggi yang dihasilkan oleh imago betina
B. tabaci dapat mencapai 8 telur pada tanaman kedelai Wilis, sedangkan pada tanaman kedelai Edamame hanya 6 telur. Imago betina B. tabaci tidak menghasilkan telur pada saat umur 29.5-30.5 hari pada varietas Edamame, sedangkan pada varietas Wilis saat umur 34.5-35.5 hari. Hal ini menunjukkan adanya perbedaan nutrisi pada masing-masing inang sehingga waktu reproduksinya berbeda. Selain faktor nutrisi, senyawa kimia pada tanaman seperti alkaloid dapat menghambat proses fisiologi dalam tubuh serangga (Schoonhoven
et al. 2005). Perilaku peneluran B. tabaci juga ditentukan oleh faktor fisik dan kimia pada daun. Butler dan Wilson (1984) menyatakan bahwa imago B. tabaci
lebih menyukai meletakkan telur pada varietas kapas yang memiliki trikoma lebih banyak daripada varietas kapas yang memiliki trikoma sedikit karena adanya rangsangan nutrisi atau senyawa kimia pada daun. Ketertarikan imago B. tabaci
untuk hidup dan meletakkan telur juga dipengaruhi oleh adanya senyawa kimia pada daun, gula, tanin, konsentrasi kelenjar gosipol, O-dihidroksifenol dan nutrisi tanaman (Bentz et al. 1995; Butter et al. 1990). Bentuk kurva keperidian B. tabaci
pada tanaman kedelai varietas Edamame lebih rendah dibandingkan dengan varietas Wilis. Tanaman kedelai varietas Edamame dapat menurunkan keperidian harian imago betina B. tabaci.
Data peluang hidup (lx) dan keperidian harian (mx) dari imago betina B. tabaci dapat digunakan untuk menghitung statistik demografi. Nilai laju reproduksi bersih (Ro) B. tabaci pada tanaman kedelai Edamame lebih rendah daripada tanaman kedelai Wilis (Tabel 2). Nilai Ro menunjukkan bahwa generasi
14
Nilai r diperoleh dan ditentukan oleh berbagai aspek yang berhubungan dengan kehidupan organisme seperti kematian, kelahiran, dan waktu perkembangan B. tabaci. Gill et al. (1989) menyatakan bahwa nilai r dapat memberikan pengetahuan yang lebih mengenai karakteristik pola kehidupan spesies yang diamati. Nilai r yang tinggi dapat diartikan bahwa populasi suatu organisme memiliki kemungkinan untuk terus tumbuh. Laju pertambahan intrinsik (r) dapat digunakan untuk memprediksi pertumbuhan populasi serangga dalam jangka waktu yang panjang. Nilai r pada tanaman kedelai Edamame sebesar 0.095 ± 0.001 nimfa per hari, sedangkan nilai r pada tanaman kedelai Wilis sebesar 0.102 ± 0.001 nimfa per hari. Hal ini menunjukkan bahwa nilai r B. tabaci pada tanaman kedelai Edamame lebih rendah daripada tanaman kedelai Wilis, dikarenakan adanya pengaruh dari masing-masing varietas terhadap proses fisiologi B. tabaci. Tingginya nilai r disebabkan oleh tingginya keperidian, rendahnya mortalitas pradewasa dan masa dewasa (Kurniawan 2007). Andrewartha dan Birch (1954) menyatakan bahwa serangga yang tinggal pada inang yang sesuai laju pertumbuhannya cepat, sedangkan pada inang yang kurang sesuai lambat karena proses fisiologinya terganggu. Selain pertumbuhan serangga terhambat, ukuran dan bobot tubuhnya menjadi rendah (Schoonhoven et al. 2005). Pertumbuhan B. tabaci yang terhambat memiliki tingkat infestasi dan kerusakan yang ditimbulkannya berbeda-beda pada setiap tanaman inang. El-Lakwah et al. (2010) menyatakan bahwa pengujian beberapa varietas kacang-kacangan menunjukkan tingkat infestasi B. tabaci yang berbeda-beda pada masing-masing inang. Menurut Amro (2008) pengujian beberapa varietas mentimun menunjukkan tingkat kerentanan yang berbeda-beda terhadap B. tabaci.
B. tabaci yang dipelihara pada tanaman kedelai Edamame membutuhkan waktu perkembangan generasi yang lebih cepat dibandingkan dengan tanaman kedelai Wilis. Lama waktu generasi dipengaruhi oleh lamanya siklus hidup suatu organisme. Individu dalam suatu populasi yang mempunyai nilai T (lama generasi) yang rendah akan tumbuh lebih cepat dibandingkan dengan individu yang mempunyai nilai T yang tinggi (Mawan dan Herma 2011). Siklus hidup B. tabaci pada tanaman kedelai Edamame lebih singkat sehingga perkembangannya menjadi imago dewasa menjadi lebih cepat pula dibandingkan dengan tanaman kedelai Wilis. Semakin cepat lama generasi (T) maka akan semakin cepat pula laju pertumbuhan intrinsik B. tabaci. Menurut Southwood dan Henderson (2000) pertumbuhan populasi tergantung dengan lama generasi yang berkaitan dengan laju pertumbuhan. Semakin lama waktu yang dibutuhkan satu generasi untuk berkembang maka akan semakin lambat pula pertumbuhan populasi tersebut. Hal ini disebabkan oleh perbedaan nutrisi atau senyawa kimia tertentu pada varietas Edamame dan varietas Wilis. Kesesuaian inang tidak hanya digambarkan dari nilai T, tetapi juga nilai GRR, Ro, r, dan DT juga sangat berpengaruh.
15 1997; Southwood dan Henderson 2000). Serangga yang memiliki waktu berlipat ganda (DT) yang cepat, akan mempercepat penurunan sumberdaya makanan (Birch 1948). Waktu berlipat ganda yang lama akan mengurangi populasi B. tabaci sehingga mengurangi infestasi hama pada tanaman kedelai. Penggunaan kedelai Edamame memberikan kehidupan yang tidak sesuai untuk B. tabaci.
Pembahasan Umum
Penggunaan kedelai varietas Edamame dan varietas Wilis berpengaruh terhadap biologi B. tabaci. Pertumbuhan dan perkembangan B. tabaci menjadi terganggu karena adanya perbedaan nutrisi dan mekanisme ketahanan dari tanaman. Beberapa mekanisme ketahanan tanaman terhadap serangan hama yaitu toleran, antisenosis dan antibiosis (Teetes 2004). Mekanisme ketahanan kedelai terhadap B. tabaci dipengaruhi oleh struktur, kualitas dan sifat agronomis dari tanaman kedelai (Xu 2009). Kerapatan trikoma daun merupakan salah satu mekanisme ketahanan kedelai terhadap hama yang berupa antisenosis. Menurut Indrayani dan Siwi (2012) semakin tinggi kerapatan bulu daun semakin rendah populasi serangga dengan alat mulut menusuk menghisap karena menghalangi stilet serangga menembus lamina daun sehingga aktifitas makannya menjadi terganggu. Namun, hasil tersebut berbeda dengan Tama (2011) yang menyatakan bahwa imago B. tabaci lebih menyukai hidup pada trikoma yang berjumlah banyak daripada trikoma yang berjumlah sedikit pada tanaman kedelai. Trikoma yang berjumlah banyak dapat berfungsi sebagai penahan telur dan sebagai pegangan atau tempat berjangkar imago B. tabaci pada saat angin kencang dan saat hujan. Selain kerapatan trikoma, ketertarikan imago B. tabaci untuk hidup dan meletakkan telur juga dipengaruhi oleh adanya senyawa kimia pada daun, gula, tanin, konsentrasi kelenjar gosipol, O-dihidroksifenol dan nutrisi tanaman (Bentz et al. 1995; Butter et al. 1990). Kerapatan trikoma berbeda-beda tiap tanaman inang dan tiap varietas tanaman inang. Menurut Junior et al. (2007) jumlah trikoma tidak dapat digunakan untuk menentukan ketidaksesuaian imago
B.tabaci dalam meletakkan telur.
Varietas kedelai Edamame berpengaruh terhadap B. tabaci yaitu menurunkan keperidian, laju pertumbuhan intrinsik yang rendah, lama hidup yang singkat dan lamanya waktu berlipat ganda. Hal ini disebabkan oleh faktor nutrisi pada tanaman inang. Sunjaya (1970) menyatakan bahwa kualitas makanan yang ada dalam tanaman inang dapat menentukan pertumbuhan, perkembangan, kesuburan, mortalitas dan keperidian serangga. Senyawa kimia tertentu pada tanaman juga berpengaruh terhadap B. tabaci dan meningkatkan ketahanan tanaman. Ketahanan tanaman dengan adanya senyawa kimia tersebut secara tidak langsung menghasilkan antibiotik (metabolik sekunder). Tanaman kedelai memiliki mekanisme antibiosis terhadap serangan B. tabaci. Menurut Vieira et al.
16
Menurut Naranjo dan Ellsworth (2005) tanaman inang merupakan salah satu faktor biotik yang dapat mempengaruhi aspek biologi dan kelangsungan hidup suatu organisme. Isaacs et al. (1998) menyatakan bahwa perbedaan varietas tanaman melon dapat memberikan pengaruh terhadap mekanisme fisiologi dan perilaku B. tabaci. Mekanisme fisiologi diperlihatkan dengan penurunan berat tubuh yang sejalan dengan penurunan konsentrasi karbohidrat. Menurut Kogan (1982) faktor fisik dan kimia tanaman memegang peranan penting dalam pemilihan dan penentuan inang. Lembaga penelitian Soyfoods Association of North America (2005) menyatakan bahwa Edamame yang telah matang mengandung 11 g/80 g protein, 142 mg/80 g unsur P, 130 mg/80 g kalsium, 13 mg/80 g natrium, 10 g/80 g karbohidrat, 485 mg/80 g kalium dan 49 mg/80 g isoflavon. Sedangkan kedelai Wilis mengandung 14 g/80 g protein, 210 mg/80 g unsur P, 88 mg/80 g kalsium, 0 mg/80 g natrium, 8 g/80 g karbohidrat, 442 mg/80 g kalium, 24 mg/80 g isoflavon. Menurut Soepardi (1983) menyatakan bahwa peranan unsur P pada tanaman sangat penting, salah satunya memperkuat daya tahan terhadap hama dan penyakit tanaman. Nutrisi merupakan salah satu faktor penting yang mempengaruhi kehidupan serangga. Ketersediaan nutrisi dipengaruhi oleh faktor fisik dan kimia tanaman. Faktor-faktor tersebut tidak bekerja secara tunggal tapi bekerja secara bersama-sama membentuk suatu sistem pertahanan tanaman, sehingga kebutuhan nutrisi bagi serangga menjadi terhambat (Wiseman 1984).
Dinamika populasi B. tabaci pada tanaman kedelai varietas Edamame memiliki siklus hidup yang cepat dan keperidian yang lebih rendah daripada varietas Wilis. Nilai r yang tinggi dapat diartikan bahwa populasi suatu organisme memiliki kemungkinan untuk terus tumbuh. Nilai r yang tinggi pada kedelai Wilis menunjukkan B. tabaci memiliki kesempatan yang lebih besar untuk terus tumbuh pada generasi berikutnya di bandingkan pada kedelai Edamame. Pertumbuhan populasi B. tabaci yang tinggi akan mempercepat terjadinya ledakan B. tabaci di pertanaman kedelai. Selain itu, nilai Ro dan GRR yang tinggi pada tanaman kedelai Wilis memperlihatkan tingkat kesesuian hidup pada tanaman inang.
17
SIMPULAN DAN SARAN
Simpulan
Perbedaan varietas kedelai memberikan pengaruh terhadap biologi B. tabaci. Masa inkubasi telur, nimfa instar 4, pupa dan siklus hidup B. tabaci lebih singkat pada tanaman kedelai varietas Edamame daripada varietas Wilis. Hasil pengamatan harian memperlihatkan dengan penggunaan varietas Edamame dapat mengurangi peluang hidup, menurunnya keperidian harian dan memperpendek lama waktu keperidian B. tabaci. Selain itu, penggunaan varietas Edamame dapat menurunkan laju reproduksi kotor (GRR), laju reproduksi bersih (Ro), laju pertumbuhan intrinsik (r), mempercepat lama generasi (T) dan memperpanjang
doubling time (DT) B. tabaci sehingga mempengaruhi pertumbuhan populasinya di pertanaman kedelai.
Saran
DAFTAR PUSTAKA
Agustini A. 2013. Pengaruh plant growth promoting rhizobacteria terhadap biologi dan statistik demografi Aphis glycines (Hemiptera: Aphididae) pada tanaman kedelai [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Amro, MARM. 2008. Population fluctuation of certain arthropod pests inhabiting selected cucurbit varieties and their resistance status to the main sap sucking pests. Egypt Journal Agriculture Research. 86(2): 697–709.
Andrewartha HG, Birch LC. 1954. The Distribution and Abundance of Animals. Chicago (US): University of Chicago Press.
Begon M, Townsend CR, Herper JL. 2008. Ecology: From Individuals to Ecosystems. 4th edition. Oxford (GB): Blackwell Puslishing.
Bentz J, Reeves III J, Barbosa P, Francis B. 1995. Within plant variation in nitrogen and sugar content of poinsettia and its effects on the oviposition pattern, survival and development of Bemisia argentifolii (Homoptera: Aleyrodidae). Environmental Entomology. 24(1): 271-277.
Birch LC. 1948. The intrinsic rate of natural increase of an insect population.
Journal Animal Ecology. 17(1):15-28.
Bonaro O, Lurette A, Vidal C, Fargues J. 2007. Modelling temperature-dependent bionomics of Bemisia tabaci (Q-biotype). Physiological Entomology. 32(1): 50-55.
Borror DJ, Johnson NF, Triplehorn CA.1996. Pengenalan Pelajaran Serangga.
Ed ke-6. Partosoedjono S, penerjemah. Yogyakarta (ID): Gadjah Mada University Press. Terjemahan dari: An Introduction to the Study of Insect.
[BPS] Badan Pusat Statistik. 2014. Produksi padi, jagung dan kedelai. Berita resmi statistik. No 22.03.13.XVII Maret 2014 [Internet]. [diunduh 2014 Mei 6]. Tersedia pada: http://www.badanpusatstatistik.go.id.
Butler GD, Wilson FD. 1984. Activity of adult whiteflies (Homoptera: Aleyrodidae) within plantings of different cotton strains and cultivars as determined by sticky-trap catches. Journal of Economic Entomolgy. 77(1): 1137-1140.
Butter NS, Vir BK, Kaur G, Singh TH, Raheja RK. 1990. Biochemical basis on resistance to whitefly Bemisia tabaci Gennadius (Aleyrodidae: Homoptera) in cotton. Tropical Agriculture. 69(1): 119-122.
Cahyono B. 2007. Kedelai, Teknik Budidaya dan Analisis Usaha Tani. Semarang (ID): CV Aneka Ilmu.
Chapman RF. 1982. The Insects Structure and Function. 3th ed. Cambridge (US): Harvard University Press.
Du L, Ge F, Zhu S, Parajulee MN. 2004. Effect of cotton cultivar on development and reproduction of Aphis gossypii (Homoptera: Aphididae) and its predator
Propylaea japonica (Coleoptera: Coccinellidae). Journal of Economic Entomology. 97(1): 1278–1283.
19 Gill JS, Sidhu AS, Sigh J. 1989. A study to determine innate capacity for increase in numbers of Earias insulana (Boisd) on cotton. Journal of Insect Science. 2(1): 289-295.
Hanafy ARI. 2004. Studied on the most important cucumber pests in the open field and suitable control programs [thesis]. Moshtohor (MT): Zagazig University.
Hidayat P, Aidawati N, Hidayat SH, Sartiami D. 2008. Tanaman indikator dan teknik RAPD-PCR untuk penentuan biotipe Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae). Jurnal Hama dan Penyakit Tumbuhan Tropika
19(1): 44-53.
Hirano K, Budiyanto E, Winarni S. 2002. Biological characteristic and forecasting outbreaks of the whitefly Bemisia tabaci, a vector of virus diseases in soybean fields [Internet]. [diunduh 2015 Februari 10]. Tersedia pada: http://www.agnet.org.
Indrayani I, Siwi S. 2012. Pengaruh kerapatan bulu daun dan kelenjar gosipol terhadap Amrasca biguttula dan Helicoverpa armigera pada kapas. Jurnal Littri. 18(3): 95-101.
Iskandar R, Muhlasah N, Neva S. 2013. Perbandingan metode bootstrap dan
jackknife dalam menaksir parameter regresi untuk mengatasi multikolinearitas. Buletin Ilmiah Matematika Statistik dan Terapannya. 2(2): 137-146.
Junior ALB, Zeneide RC, Andre LL, Alcebiades RC. 2007. Adult attractiveness and oviposition preference of Bemisia tabaci (Genn.) (Homoptera: Aleyrodidae) B-biotype in cotton genotypes. Science Agriculture. 64(2): 147-151.
Kalshoven LGE. 1981. The Pests of Crops in Indonesia. Laan PA van der, penerjemah. Jakarta (ID): Ichtihar Baru-van Hoeve. Terjemahan dari: De Plagen van de Cultuurgewassen in Indonesie.
Kim DS, Lee JH. 2002. Egg and Larval Survivorship of Carposina sasakii
(Lepidoptera: Carposinidae) in Apple and Peach and Their Effects on Adult Population Dynamics in Orchards. Environment Entomology. 31(1): 686-692.
Kozlowski J. 1992. Optimal allocation of resources to growth and reproduction implications for age and size at maturity. Trends in Ecology & Evolution. 7(1):15-19.
Kurniawan HA. 2007. Neraca kehidupan kutukebul Bemisia tabaci Gennadius (Hemiptera: Aleyrodidae) biotipe-B dan non-B pada tanaman mentimun (Cucumis sativus L.) dan cabai (Capsicum annum L.) [tesis]. Bogor (ID): Fakultas Pertanian, Institut Pertanian Bogor.
Lambert AL, McPherson RM, Espeliei KE. 1995. Soybean host plant resistance mechanisms that alter abundance of whiteflies (Homoptera: Aleyrodidae).
Environment Entomology. 24(6): 1381–1386.
20
Li B, Guan-lin X, Soad A, Coosemans J. 2005. Suppression of Meloidogyne javanica by antagonistic and plant growth promoting rhizobacteria. Journal of Zhejiang University Science. 6(6): 496-501.
Marlena L. 2014. Optimasi ukuran subcontoh melalui bootstrap dan jackknife untuk pendugaan statistik demografi hama Aphis glycines [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Martin JH. 1987. An identification guide to common whitefly pest species of the world (Homoptera: Aleyrodidae). Tropical Pest Management. 33(4): 298-322.
Marwoto, Inayati. 2011. Pengendalian kutu kebul pada kedelai. Iptek Tanaman Pangan. 6(1): 87-98.
Marwoto, Indriani FC, Sulistyo A, Hapsari RT. 2011. Diagnosis Ledakan Populasi Hama Kutu kebul (Bemisia tabaci) pada Tanaman Kedelai (Glycine max). Di dalam: Indriyani, editor. Prosiding Seminar Nasional Hasil Penelitian Aneka Kacang dan Ubi tahun 2009; 2009 Juni 17-21; Malang, Indonesia. Malang (ID): Balai Penelitian Tanaman Kacang-kacangan dan Umbi-umbian. hlm 255-262.
Mawan A, Herma A. 2011. Statistik demografi Riptortus linearis (Hemiptera: Alydidae) pada tanaman kacang panjang. Jurnal Entomologi Indonesia. 8(1): 8-16.
McAuslane HJ. 1996. Influence of leaf pubescence on ovipositional preference of
Bemisia argentifolii (Homoptera: Aleyrodidae) on soybean. Environmental Entomology. 25(4): 834-841.
McKenzie CL, Anderson PK, Villarreal N. 2004. An extensive survey of Bemisia tabaci (Homoptera: Aleyrodidae) in agricultural ecosystems in Florida.
Florida Entomologist. 87(3): 403-407.
Naranjo SE, Ellsworty PC. 2005. Mortality dinamic and population regulation in
Bemisia tabaci. Entomologia exsperimentalalis et aplicata. 116(2): 93-108. Natawigena H. 1990. Entomologi Pertanian. Bandung (ID): PT Orba Sakti
Bandung.
Oleivera MRV, Henneberry TJ, Anderson P. 2001. History, current status and collaborative projects for Bemisia tabaci. Spesies issue: Challenges and oppurtunities for pest management of Bemisia tabaci in the new century.
Crop Protection. 20(9): 725-737.
Panizzi AR, Parra JRP. 2009. Bioekologi dan gizi serangga sebagai dasar untuk pengelolaan hama terpadu. Neotropical Entomology. 68(2): 1107-1140. Price PW. 1997. Insect Ecology. 3rd ed. New York (US): John Wiley & Sons. Pusat Penelitian dan Pengembangan Tanaman Pangan. 2006. Hama, Penyakit dan
Masalah Hara pada Tanaman Kedelai. Bogor (ID): hlm 53-67.
Razmjou J, Moharramipour S, Fathipour Y, Mirhoseini SZ. 2006. Effect of cotton cultivar on performance of Aphis gossypii (Homoptera: Aphididae) in Iran.
Journal of Economic Entomology. 99(1): 1820–1825.
Schoonhoven LM, Van loon JJA, Dicke M. 2005. Insect Plant Biology. Oxford (GB): University Press.
21 Silva JPGFd, Edson LLB, Efrain SdS, Andre LL. 2012. Assessing Bemisia tabaci
(Genn) biotype B resistance in soybean genotypes: antixenosis and antibiosis. Chilean Journal Of Agricultural Research. 72(4): 516-522. Soepardi G. 1983. Sifat dan Ciri Tanah. Bogor (ID): Institut Pertanian Bogor. Southwood TRE, Henderson PA. 2000. Ecological Method. 3thed. Oxford (GB):
Blackwell Science.
Soyfoods Association of North America. 2005. Whole Soybean [Internet]. [diunduh 2015 Mei 15]. Tersedia pada: http://www.soyfoods.org.
Speight MR, Hunter MD, Watt AD. 2008. Ecology of Insect: Concept and Application. 2nd ed. Oxford (GB): John Wiley & Sons.
Subagyo VNO. 2010. Neraca kehidupan kutukebul Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) pada tanaman tomat (Lycopersicon esculentum
Mill.), tanaman cabai (Capsicum annuum L.), dan gulma babadotan (Ageratum conyzoides L.) pada suhu 250C dan 290C [skripsi]. Bogor (ID): Institut Pertanian Bogor.
Sulistyadi FW, Serafinah I, Suharsono. 2012. Hubungan kerapatan panjang trikoma daun kacang tanah (Arachis hypogaea) terhadap preferensi peletakan telur Bemisia tabaci. Jurnal Littri. 4(1): 10-13.
Sunjaya PI. 1970. Dasar-dasar Ekologi Serangga. Diktat tidak dipublikasikan. Bogor (ID): Institut Pertanian Bogor.
Syahputra E, Djoko P, Partomuan S. 2002. Pengaruh fraksi aktif kulit batang
Dysoxylum acutangulum (Meliacea) terhadap reproduksi Crocidolomia pavonana (Lepidoptera: Pyralidae). Jurnal Hama dan Penyakit Tumbuhan Tropika. 2(1): 1-7.
Tama OH. 2011. Analisis Kerapatan Trikoma dan Preferensi Bemisia tabaci terhadap Ketahanan Kedelai Tahan CPMMV Berdaya Hasil Tinggi dan Pemanfaatannya sebagai Bahan Ajar Pengelolaan Hama Terpadu [tesis]. Malang (ID): Universitas Negeri Malang.
Teetes GL. 2004. Plant resistance to insects: a fundamental component of IPM [Internet]. [diunduh 2015 Februari 4]. Tersedia pada: http://ipmworld. umn.edu/chapters/teetes.htm.
Tsai JH, Wang K.1996. Development and reproduction of Bemisia argentifolii
(Homoptera: Aleyrodidae) on five host plants. Environtmental Entomology.
25(2): 810-816.
Van den Ven WTG, Levesque CS, Perring TM, Walling LL. 2000. Local and sistemic chages in squash gene exspession in response to silverleaf whitefly feeding. the Plant Cell. 12(1): 1409-1423.
Vieira SS, Bueno AF, Boff MIC, Bueno RCOF, Hoffman-Campo CB. 2011. Resistance of soybean genotypes to Bemisia tabaci (Genn.) biotype B (Hemiptera: Aleyrodidae). Neotropical Entomology. 40(1): 117-122.
Wilson EO, Bossert WH. 1971. A Primer of Population Biology. Sunderland (US): Sinauer Associates.
Wirianti DA. 2006. Pengaruh penggunaan rhizobacteri pemacu pertumbuhan tanaman terhadap perkembangan populasi kutu kebul Bemisia tabaci
(Gennadius) (Hemiptera: Aleyrodidae) pada tanaman cabai [skripsi]. Bogor (ID): Institut Pertanian Bogor.
22
Xu R. 2009. Evaluation and inheritance of resistance to whitefly Bemisia tabaci
Gennadius in soybean [dissertation]. China (CN): Nanjing Agriculture College.
Zeng F, Pederson G, Ellsbury M, Davis F. 1983. Demographic statistics for pea aphids (Homoptera: Aphididae) on resistant and susceptible red clovers.
Journal of Economic Entomology. 86(1): 1852-1856.
23
24
26
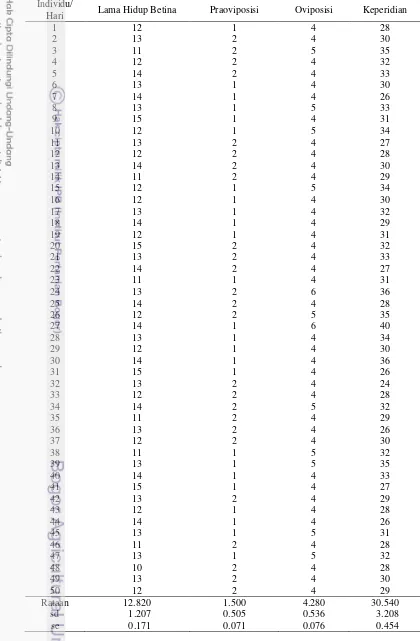
Tabel Lampiran 3 Lama hidup betina, praoviposisi, oviposisi dan keperidian B. tabaci pada tanaman kedelai Edamame
Individu/
Hari Lama Hidup Betina Praoviposisi Oviposisi Keperidian
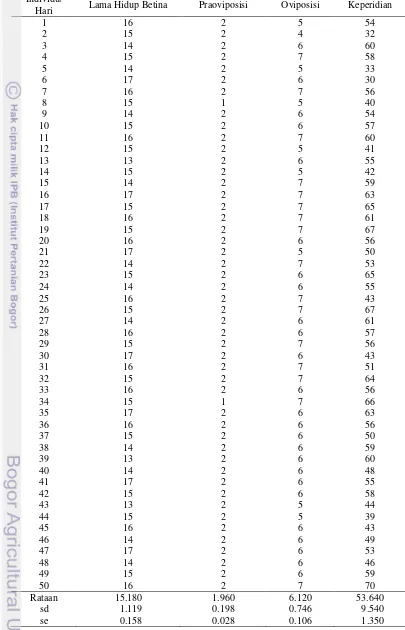
27 Tabel Lampiran 4 Lama hidup betina, praoviposisi, oviposisi, dan keperidian B.
tabaci pada tanaman kedelai Wilis Individu/
Hari Lama Hidup Betina Praoviposisi Oviposisi Keperidian
28
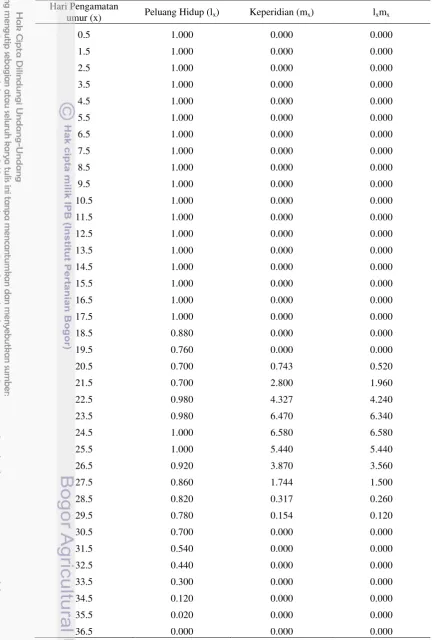
29 Tabel Lampiran 6 Neraca kehidupan B. tabaci pada tanaman kedelai Wilis
30
RIWAYAT HIDUP
Penulis dilahirkan di Suka Mulia, Pertanian, Kec. Kualuh Selatan, Kab. Labuhanbatu Utara, Aek Kanopan, Medan pada tanggal 20 Oktober 1993. Penulis merupakan anak sulung dari dua bersaudara, dari pasangan Erwin Sagala dan Halimah Tussaddiah. Penulis menyelesaikan pendidikan Sekolah Menengah Atas di SMAN 1 Kualuh Hulu Aek Kanopan program IPA pada tahun 2011. Pada tahun yang sama, penulis diterima di Institut Pertanian Bogor, Fakultas Pertanian, Program Studi Proteksi Tanaman, melalui jalur Undangan Seleksi Masuk IPB. 2015. Penulis mendapatkan beasiswa Bidikmisi dari pemerintah selama S1.