I.
INTRODUCTION
I n t e n s i v e aquacul t u r e r e q u i r e s adequate suppl y o f seed
t h e whole year round. R e c e n t l y , some experiments have demonstrated t h a t t h e q u a l i t y o f d i e t s c o u l d a f f e c t t h e f e c u n d i t y and t h e h a t c h i n g r a t e of eggs o f r e d sea bream
( Pagrus m a j o r ) , and rainbow t r o u t ( Oncorhynchus myk i s s ) (Takeuchi, Watanabe, Ogino, S a i t o , Nishimura and Nose, 1981; Watanabe, I t o h , Murakami, Tsukashima, K i t a j i m a , and F u j i t a , 1984a ; Watanabe, Ohhashi, I t o h , K i t a j i m a and F u j i t a , 1984b; Watanabe, Arakawa, K i t a j i m a and F u j i t a , 1984c; Watanabe, Takeuchi, S a i t o and Nishimura, 1984d; Watanabe, I t o h , S a i t o , Satoh, K i t a j i m a and F u j i t a , 1985).
H a t c h i n g r a t e o f eggs produced by r e d sea bream and rainbow t r o u t f e d l i n o l e n i c a c i d d e f i c i e n t d i e t i s v e r y low (Watanabe e t a7. 1984a, 1984b, 19844). M o r p h o l o g i c a l study o f d e v e l o p i n g embryo showed some cleavage d i s o r d e r a t t h e 1 6 t o 32 c e l l s t a g e , a b l o c k i n g e f f e c t b e f o r e g a s t r u l a t i o n , and 1 a t e r v a r i o u s a1 t e r a t i ons i n t h e phase o f organogenesi s ( L e r a y , Nonnote, Roubaud and Leger, 1985).
a c i d t o o . Presumably rainbow t r o u t s t i l l r e q u i r e s l i n o l e i c a c i d f o r normal egg development, eventhough i n a s m a l l amount. Yu e t a ? . ( 1 9 7 9 ) concluded t h a t l i n o l e i c a c i d i n t h e eggs was e i t h e r f r o m i n c o m p l e t e l y e x t r a c t e d d i e t i n g r e - d i e n t s such as d e x t r i n , c a s e i n and g e l a t i n , o r was mobi- 1 i z e d f r o m t h e body o f t h e b r o o d f i s h . Another experiment from Rahn, Sand and Schlenk ( 1 9 7 7 ) showed t h a t gourami embryos a p p a r e n t l y r e q u i r e d b o t h l i n o l e n i c and l i n o l e i c a c i d , whereas o n l y l i n o l e n i c a c i d i s e s s e n t i a l t o o l d e r f i s h .
To g a i n t h e maximum growth, t h e j u v e n i l e o f c a t f i s h
( C 7 a r i a s b a t r a c h u s L i n n . ) r e q u i r e s 1 . 5 % o f 1 in o l e i c a c i d and 0 . 6 % l i n o l e n i c a c i d (Mokoginta, 1 9 8 6 ) . B u t f o r broodstock development o f t h i s s p e c i e s t h e l i n o l e i c and l i n o l e n i c a c i d
r e q u i r e m e n t s have n o t y e t been determined. S o t h e o b j e c t i v e s o f t h i s experiment a r e :
1 . t o determine t h e l i n o l e i c and l i n o l e n i c a c i d r e q u i r e - ments o f b r o o d f i s h , t h a t i s s u i t a b l e t o become an a d u l t d i e t , t o develop gonad m a t u r a t i o n , and p r o d u c t i o n o f good q u a l i t y eggs.
2 . t o s t u d y t h e e s s e n t i a l f a t t y a c i d metabolism d u r i n g embryonic development o f c a t f i s h ( C 7 a r i a s b a t r a c h u s
I
I.
LITERATURE REVIEW
Although p r e c i s e i n f o r m a t i o n on t h e n u t r i t i o n a l requirements f o r gonadal m a t u r a t i o n o f t h e broodstock i s l a c k i n g , i t has been agreed t h a t t h e q u a n t i t y , qual i t y and f e e d i n g regiment would a f f e c t f i s h spawning and egg q u a l i t y . Some experiments showed t h a t t h e q u a l i t y o f t h e broodstock d i e t c o u l d a f f e c t t h e qual i t y o f t h e eggs produced by t h e broodstock.
A group o f r e d sea bream broodstock f e d -upon t h e low p r o t e i n d i e t (33.0%), o r f e d upon t h e d i e t w i t h o u t supple- mental phosphorus has a lower number o f eggs compared t o t h e broodstock fed upon t h e c o n t r o l d i e t (45.0% p r o t e i n l e v e l w i t h supplemental of phosphorus, Watanabe
et
a 7 .,
1984a). Growth r a t e and feed e f f i c i e n c y o f t h e rainbow t r o u t brood- s t o c k f e d upon t h e d i e t c o n t a i n i n g 36.0% p r o t e i n , and 18.0% 1 i p i d l e v e l was h i g h e r compared t o t h e growth r a t e and feed e f f i c i e n c y o f t h e broodstock f e d upon 28.0% p r o t e i n and 20.0% l i p i d l e v e l (Watanabe e t a l . , 1984d). Another e x p e r i - ment by Oahl green ( 1980) u s i n g guppy ( Poeci 7 i a r e t i c u l a t a ) broudstock has found o u t t h a t i f t h e f i s h were f e d upon t h e d i e t c o n t a i n i n g 47.0% p r o t e i n l e v e l w i l l produce average gonad0 somatic r a t i o h i g h e r than i f i t was f e d upon a d i e t c o n t a i n i n g 31.0% and 15.0% p r o t e i n l e v e l .centage o f eyed eggs as w e l l as t h e h a t c h a b i l i t y . The broodstock r e c e i v i n g e s s e n t i a l f a t t y a c i d (EFA) d e f i c i e n t d i e t a l s o produced eggs w i t h s i g n i f i c a n t l y low h a t c h a b i l i t y , and most o f t h e hatched l a r v a e showed d e f o r m i t y o f t h e body as w e l l (Watanabe
et
a l . , 1984a, d ) .N u t r i t i o n a l q u a l i t y o f t h e d i e t s g i v e n t o t h e brood- s t o c k would i n f l u e n c e t h e q u a l i t y o f r e d sea bream eggs eventhough on t h e verge o f spawning (Watanabe
et
a l . ,1984a). The f a t t y a c i d o f t h e eggs were g r e a t l y a f f e c t e d by d i e t a r y f a t t y a c i d s u p p l i e d t o t h e broodstock. The propor- t i o n o f a 3 h i g h l y unsaturated f a t t y a c i d s ((#J3 HUFA) was h i g h
i n t h e eggs o f t h e broodstock f e d upon t h e d i e t c o n t a i n i n g a h i g h l e v e l o f cJ3 HUFA, and low i n t h e eggs o f t h e EFA-def i- c i e n t f i s h (Watanabe
et
a l . , 1984b; 1985). The f a t t y a c i d composition i n t h e organs l i k e l i v e r , h e a r t , kidney and b r a i n were a l s o i n f l u e n c e d by f a t t y a c i d composition o f t h e d i e t (Lee, Roehm, Yu and Sinnhuber, 1967; Stickney and Andrews, 1971; C a s t e l l , Lee and Sinnhuber, 1972; C a s t l e d i n e and Buckley, 1980; R e i n i t z and Yu, 1984). F a t t y a c i d compo- s i t i o n o f d i e t a r y l i p i d had a g r e a t e r e f f e c t on t h e f a t t y a c i d composition o f p h o s p h o l i p i d than on t h e f a t t y a c i d composition o f n e u t r a l 1 ip i d .s t u d i e d t h e i n c i d e n c e o f G 3 - f a t t y a c i d d e f i c i e n c y on t h e t r o u t r e p r o d u c t i v e processes. Eggs from broodstock f e d upon E F A - d e f i c i e n t d i e t , has t h e same f e r t i l i z a t i o n e f f i c i e n c y i n two days development compared t o t h e eggs from broodstock f e d uQon a commercial d i e t . A f t e r t h a t , two peaks o f mor- t a l i t y , one a t day 8 and t h e o t h e r a t day 22 a f t e r f e r t i l i - z a t i o n were observed from t h e eggs produced by t h e d e f i c i e n t f i s h . I n a d d i t i o n , v i t e l l u s r e s o r p t i o n was s h o r t e r i n t h e a l e v i n s from t h e EFA-deficient broodstock ( 5 0 days) than t h e a l e v i n s from t h e broodstock f e d upon a commercial c o n t r o l d i e t ( 6 5 days). Morphological study o f t h e development o f t h e c33 d e f i c i e n t embryo showed some cleavage d i s o r d e r a t 16 t o 3 2 c e l l stage, a b l o c k i n g e f f e c t b e f o r e g a s t r u l a t i o n , and l a t e r v a r i o u s a l t e r a t i o n i n organogenesis.
It
was concluded t h a t t h e G=3 f a t t y acids, and e s p e c i a l l y 22:6 G3, p l a y a c r u c i a l and s p e c i f i c r o l e i n t h e t r o u t embryo development.Former experiments demonstrated, t h a t some d i e t a r y p h o s p h o l i p i d were necessary f o r good growth and s u r v i v a l o f t h e l a r v a l f i s h , such as t h e r e d sea bream (Pagrus major), t h e ayu ( PLecog 7ossus a 7 t i v e 1 i s ) , t h e s t a r y f l o u n d e r (Para- 7ychthys o 1 ivaccus) and t h e k n i f e jaw ( Op legnathus f a s c i a -
as l i n o l e i c a c i d (18:Z U 6 ) , docosahexaenoic a c i d (22:6 G3), e c t . as f a t t y a c i d s m o i e t i e s were e f f e c t i v e i n enhancing growth and s u r v i v a l r a t e s of t h e l a r v a l ayu.
The f u n c t i o n o f EFA i n t h e embryogenesis might be r e 1 a t e d t o t h e membrane f 1 u i d i t y and permeabi 1 i t y . The f l u i d mosaic model o f membrane s t r u c t u r e p o s t u l a t e t h a t membranes c o n s i s t s o f p h o s p h o l i p i d arranged i n a f l u i d b i l a y e r w i t h g l o b u l a r p r o t e i n p e n e t r a t i n g i n t o a l i p i d b i l a y e r ( C a p a l d i , 1980). Length and degree o f u n s a t u r a t i o n o f hydrocarbon s i d e c h a i n c o u l d i n f l u e n c e membrane proper- t i e s , l o w e r i n g t h e m e l t i n g temperature o f complex l i p i d ( a r e t s c h e r and R a f t , i n Morson and C l a n d i n i n , 1986). C r i t i - c a l temperature o r t r a n s i ' t i o n 1 i p i d phase ( T t ) and Arrhenius a c t i v a t i o n energy (Ea) o f t h e enzyme have been used t o determine a temperature-induced change i n t h e p h y s i c a l p r o p e r t i e s o f t h e l i p i d membrane ( I n n i s and C l a n d i n i n , 1981;
McMurchie, Gibson, Abeywardena and Charnock, 1983a;
McMurchi, Abeywardena, Charnock and Gibson, 1983b). Divaka- r a n and Venkataraman (1977); Morson and C l a n d i n i n (1986) demonstrated t h a t t h e membrane enzyme a c t i v i t y w i l l be i n f l u e n c e d by t h e p h y s i c a l p r o p e r t i e s o f t h e i r h o s t mem- brane. Recently, i t was shown a l s o t h a t t h e r a t i o o f phos-
hat
i d y l c h o li
ne : phosphatidyl ethanol ami ne i n t h e m i tochon- d r i a l membrane o f cold-aclimated p o i k i l o t h e r m s changes as a f u n c t i o n o f temperature ( T h i l l a r t and Brum i n Robblee and C l a n d i n i n , 1984). I t i s n o t known whether these changes i nt h e membrane. The s u r f a c e charge o f phosphatidylchol i n e ( z e r o a t n e u t r a l pH) i s d i f f e r e n t than t h a t o f phosphati- dylethanolamine, which has a n e g a t i v e charge a t n e u t r a l pH. Therefore, i t was concievable t h a t a change i n t h e r a t i o o f p h o s p h a t i d y l c h o l i n e : phosphatidylethanolamine may a l t e r membrane s u r f a c e p o t e n t i a l and t h e i r membrane p r o p e r t i e s (Robbiee and C l a n d i n i n , 1984). Change i n t h e p h y s i c a l p r o p e r t i e s o f b i o l o g i c a l membrane have been shown t o i n f lu - ence t h e behavior o f c e r t a i n membrane-associated enzymes,
-
and t h i s might a l t e r p h y s i o l o g i c a l processes.
F i s h eggs c o n t a i n most e s s e n t i a l enzymes o f g l y c o l y s i s , t r i c a r b o x y l i c a c i d c y c l e , and r e s p i r a t i o n c h a i n ( M i lman and Yurovi t s k y i n Neyfakh and Abramova, 1974). A1 t e r a t i o n o f some o f these a c t i v i t i e s might a f f e c t o t h e r c e l l metabolism. I n h i b i t i o n o f a l l metabolic processes as i n d i c a t e d by t h e decrease o f r e s p i r a t i o n may cause t h e i n a b i 1 i t y o f c e l l s to aggregate ( F l o r k i n and S t o t z , 1967).
Other f u n c t i o n o f EFA i n t h e embryogenesis might be re1a.ted t o t h e s y n t h e s i s o f oxygenated c y c l i c and non- c y c l i c d e r i v a t i v e s . Precursors o f oxygenated hormone-like compounds belong t o t h e 0 3 and c36 s e r i e s . These compounds might a c t as a modulator o r mediator i n v a r i o u s p h y s i o l o g i - c a l events, e s p e c i a l l y i n t h e processes o f t h e c e l l u l a r
r e c o g n i t i o n o c c u r r i n g d u r i n g embryonic development (Leray e t
a l . , 1985).
111. MATERIALS AND METHODS
The experiment was conducted a t t h e Research Pond Complex o f t h e F a c u l t y o f F i s h e r i e s , I n s t i t u t P e r t a n i a n Bogor ( I P B ) , Bogor, f r o m March t o December 1990. F a t t y a c i d anal y s i s was conducted a t t h e l a b o r a t o r y o f Departement Technology and Human N u t r i t i o n , F a c u l t y o f A g r i c u l t u r a l E n g i n e e r i n g and Technology, IPB.
3.1 M a t e r i a l s
3.1.1 E x p e r i m e n t a l f i s h
F i v e months o l d f i s h , weighing 91.7
-
1 2 2 . 3 g/female, and 9 0 . 0 - 110.0 g/male, were randomly stocked i n 15 t a n k s . S i x females and t h r e e males were p l a c e d i n each t a n k .3 . 1 . 2 E x p e r i m e n t a l d i e t s
F i v e e x p e r i m e n t a l d i e t s were used i n t h i s experiment. T a b l e 1 shows t h e composition o f t h e d i e t s . A l l t h e i n g r e - d i e n t s were mixed t o g e t h e r . A t f i r s t , c h o l i n e c h l o r i d e was mixed w i t h v i t a m i n mix i n a bowl. Then, o t h e r i n g r e d i e n t , one by one, was added t o t h e former m i x t u r e . I t was s t a r t e d
f r o m . m i n e r a l m i x , and was f o l l o w e d by CMC, a - c e l l u l o s e , d e x t r i n , c a s e i n and l i p i d . I n another bowl, 400 m l ( f o r 1 kg d i e t ) o f b o i l e d water was added t o g e l a t i n
.
A f t e r t h e g e l a t i n was c o l d , i t was added t o t h e o t h e r i n g r e d i e n t , and was mixed t o g e t h e r f o r f i v e minutes. The e x p e r i m e n t a l d i e t sFuther, sample
of each experimental diet was analyzed
for proximate composition. Crude protein content of diets
was analyzed by using Kjeldahl method; and crude
l i p i d .by
Table
1 .Composition of the experimental diets
*
Component
( s / l O O s )Casei
n
3 5 . 0 3 5 . 0 35.0 35.0 3 5 . 0Gelatin
9 . 0 9 . 0 9 . 0 9 . 0 9 . 0Dextrin
3 5 . 0 3 5 . 0 35.0 35.0 3 5 . 0CMC
3 . 0 3 . 0 3 . 0 3 . 0 3 . 0a-cellulose
5 . 0 5 . 0 5 . 0 5 . 0 5 . 0Mineral mixa
5 . 0 5 . 0 5.0 5 . 0 5.0Vitamin mix
b
1 . 5 1.5 1.5 1.5 1 . 5Choline chloride
0 . 5 0 . 5 0.5 0.5 0.5Beef tallow
6.0 2.0 2.0 2.0 0.0Corn oil
0 . 0 0 . 0 4.0 3 . 0 0.0Linseed oil
0 . 0 4 . 0 0 . 0 1 .O 0.0Cuttle fish liver oil
0 . 0 0 . 0 0 . 0 0 . 0 6.0 ( g / 1 0 0 gd r y
basic)
1
u6**
0.2 0.6 2.0 1.5 0.2--- -
( a )
See
Appendix
1for the composition of mineral mix.
( b )
See
Appendix
2for
the
composition of vitamin mix.
*
Takeuchi, personal communication; Mokoginta,
1 9 8 6 .**
1
d 6 contain
1 6 : 3w6,
18:2 U S , 20:2t36,
20:3d6
20:4d6,
22:5W6
e t h e r e x t r a c t i o n method. F a t t y a c i d composition o f t h e d i e t was analyzed u s i n g gas l i q u i d chromatography.
Table 2 shows t h e proximate composition o f t h e e x p e r i - mental d i e t s , and t h e W6 and d 3 - f a t t y a c i d c o n t e n t s o f t h e d i e t s . The composition o f f a t t y a c i d s i n t h e d i e t s can be seen i n Appendix 3.
Table 2. Proximate composition o f t h e experimental d i e t s , t h e G;6 and d 3 - f a t t y a c i d c o n t e n t o f t h e d i e t .
P r o x i mate Treatments/diets
composition
(g/100 g d r y A B
c
D Eb a s i c d i e t )
Crude p r o t e i n 47.47 48.29 46.51 47.72 47.58
Crude l i p i d 5.83 5.89 5.96 5.85 5.81
Ash 5.02 5.69 5.16 6.08 6.15
Crude f i b e r 3.19 4.86 5.10 4.79 5.83
1 u 6 0.21 0.67 2.24 1.85 0.26
i: W 3 0.03 2.09 0.07 0.56 1.68
E a3
HUFA 0.00 0.00 0.00 0.00 1.453 . 1 . 3 Experimental t a n k s
(see ~ p p e n d i x 4 ) . The water temperature was r e g u l a t e d a t a range o f 25Oc
-
27Oc by u s i n g a thermostat.quar ria, 60 x 50
x
40 cm, were used f o r egg incubation. F e r t i l i z e d eggs were spread on g l a s s surface 20 x 10 cm, andt h e n they were p l a c e d i n aquaria. Ouring incubation, t h e water was we1 1 aerated, and 1.0 Qpm o f methylene b l u e was
added
%o
t h e water t o g r o t e c t t h e eggs from fungus. The water temperature was 2 6 . 5 ' ~-
2 8 . ~ ~ ~ .3.2.1 Feeding and spawning of
the b r o o d f i s h
The f i s h were t r a i n e d t o feed and acclimated t o t h e l a b o r a t o r y c o n d i t i o n and f e d w i t h c o n t r o l d i e t f o r t w o weeks ~ r i o r
t.o
t h e beginning of t h e experiment. Ouring t h e exper-iment, t h e f i s h were f e d experimental d i e t s , a t s a t i a t i o n ,
once a
day, a t 5.00 o ' c l o c k i n the aflternoon.t h e water i n t h e tank was changed 100% volume,
once
i nt w o days. The tank were scrubbed and cleaned thoroughly once a month. Ouring cleaning, t h e f i s h were observed w h e C h s r t h e y ware mature b y sexing them macroscopically
.
used f o r t h e t r e a t m e n t s were p l a c e d i n d i f f e r e n t tanks. The mature females were removed t o a f i b e r g l a s s t a n k b e f o r e t h e f i s h were i n j e c t e d w i t h c a r p p i t u i t a r y suspension i n o r d e r t o s t i m u l a t e o v u l a t i o n . Females o f c a t f i s h ( C l a r -
i a s batrachus L i n n . ) r e c e i v e d one dosage (body w e i g h t o f donor : body w e i g h t of r e c e p i e n t
=
1 : 1 ) of c a r p p i t u i t a r y suspension f o r every i n j e c t i o n . The females r e c e i v e d twoi n j e c t i o n s , one a t 4:00 o ' c l o c k i n t h e a f t e r n o o n , and t h e o t h e r a t 12:OO o ' c l o c k a t n i g h t . A t 7:00/8:00 o ' c l o c k i n
t h e morning, t h e f i s h were s t r i p p e d . The eggs produced by t h e females were f e r t i l i z e d w i t h sperms o f t h e males f r o m a n o t h e r pond. I n o r d e r t o g e t v i a b l e spermatozoa, t h e males were i n j e c t e d w i t h one dosage o f c a r p p i t u i t a r y suspension, and were s a c r i f i c e d f o r t h a t purpose.
About 20% of u n f e r t i l i z e d eggs were c o l l e c t e d f o r p o l a r and n o n p o l a r l i p i d f r a c t i o n a n a l y s i s , and t h e f a t t y a c i d c o m p o s i t i o n a n a l y s i s . The r e s t o f eggs were f e r t i l i z e d by spermatozoa, and t h a n wqre spread on g l a s s s u r f a c e ( 2 0 x 10 cm) which was p l a c e d i n a f i b e r g l a s s t a n k c o n t a i n i n g w a t e r .
Futhermore, t h r e e o r f o u r p i e c e s o f t h o s e g l a s s e s were removed f r o m f i b e r g l a s s t a n k and were p l a c e d i n an aquarium c o n t a i n i n g water f o r eggs i n c u b a t i o n . The number o f eggs on t h o s e g l a s s e s and t h e l a r v a e produced were counted and were used t o determine t h e h a t c h i n g r a t e o f eggs. The r e s t o f f e r t i l i z e d eggs were d i v i d e d i n t o t h r e e batches and were
l e c t e d one hour a f t e r f e r t i l i z a t i o n ; another b a t c h e l e v e n h o u r s a f t e r f e r t i l i z a t i o n ; and t h e r e s t were c o l l e c t e d a f t e r t h e eggs hatched ( l a r v a e ) . Those samples o f eggs and l a r v a e were used f o r l i p i d c o n t e n t , p o l a r and nonpolar l i p ~ d f r a c - t i o n a n a l y s i s , and f a t t y a c i d c o m p o s i t i o n a n a l y s i s .
The female f i s h were weighed b e f o r e and a f t e r spawning, so t h e w e i g h t o f eggs produced by each female c o u l d be c a l c u l a t e d . Futhermore, t h e number o f eggs f r o m 0 . 5 g ( w i t h t w o r e p l i c a t e s ) sample were counted i n o r d e r t o e v a l u - a t e d i f f e r e n c e s o f egg s i z e between t r e a t m e n t s .
Seven l o t s o f abnormal l a r v a e were c o l l e c t e d f o r h i s t o l o g i c a l e v a l u a t i o n (see Appendix 5 f o r procedure o f h i s t o l o g i c a l p r e p a r a t i o n ) .
I n o r d e r t o e v a l u a t e t h e v i a b i 1 i t y of t h e spermatozoa, t h e eggs produced by one female i n t r e a t m e n t D were d i v i d e d
i n t o f i v e batches, and were f e r t i l i z e d by m i l t o f males f r o m t r e a t m e n t s A, B, C, D and E.
Furthermore, eggs produced by one female f r o m t r e a t - ment E were d e v i d e d i n t o t h r e e batches, and were f e r t i l i z e d
by m i l t o f males f r o m t r e a t m e n t s A, C, and E. Seven h o u r s a f t e r f e r t i l i z a t i ~ n ~ t h e eggs were removed f r o m t h e i n c u b a t i o n a q u a r i u a . The number o f u n f e r t i l i z e d and f e r t i l i z e d eggs were c a l c u l a t e d .
3.2.2
Chemical analysis
F o l c h , Lee and Stanley method (1957, i n Takeuchi, 1988), and then nonpolar l i p i d ( n e u t r a l l i p i d ) was separated from t h e p o l a r l i p i d ( p h o s p h o l i p i d ) f r a c t i o n by u s i n g s i l i c i c a c i d column (Sep-Pak C a r t r i d g e ; Water Associates, USA; Takeuchi,
1988).
F u r t h e r , t h e f a t t y a c i d composition o f nonpolar and p o l a r 1 i p i d f r a c t i o n were determined by u s i n g gas chromato- graphy
3.2.2.1 L i p i d e x t r a c t i o n
Sample o f eggs, l a r v a e o r body t i s s u e ( A g) was placed i n a waring blender. Chloroform/methanol 2 : v o l . / v o l . ) 20 x A m l was added i n t o t h e waring blender,and then t h e sample was homogenized f o r 5 minutes. The homogeni zed sample was f i l t e r e d , and t h e lower l a y e r s o l u t i o n was placed
i n an erlenmeyer (250 m l ) . The upper l a y e r was discarded. The lower l a y e r s o l u t i o n i n t h a t erlenmeyer was t r a n s f e r e d
T o t a l weight o f l i p i d can be c a l c u l a t e d by s u b t r a c t i n g b-a
( D g ) .
Crude l i p i d
=
D I A x 100%3.2.2.2 P o l a r and nonpolar l i p i d f r a c t i o n a n a l y s i s
About 100.0 mg of crude l i p i d was d i s s o l v e d i n 1.0 m l chloroform. That- sample was i n j e c t e d a t t h e
top
o f c a r - t r i d g e . Another s y r i n g e c o n t a i n i n g 20.0 m l o f c h l o r o f o r m was connected i n t o t h etop
of c a r t r i d g e , and then t h e c h l o -r o f o r m was pushed down Shrough t h e c a r t r i d g e w i t h t h e f l o w r a b 25 ml/minute. Nonpolar l i ~ i d f r a c t i o n was c o l l e c t e d i n a p r e v i o u s l y weighed round bottom f l a s k ( a g). Then, a ch1oroform/methano1 (49:1, v o l . / v o l . ) m i x t u r e was used to e l u t e she remaining monog1 ycerides i n t h e c a r t r i d g e . Monoglycerides was a l s o added to t h e nonpolar l i p i d frac- t i o n . F u r t h e r , 30.0 m l methanol was used t o e l u t e t h e w l a r l i p i d , and t h a t p o l a r l i p i d f r a c t i o n was c o l l e c t e d i n
another p r e v i o u s l y wei.ghed round bottom f l a s k
( x
g ) . Both p o l a r and nonpolar l i g i d s o l u t i o n i n t h e f l a s k were evapo-r a t e d fo dryness u s i n g a r o t a r y vacuum evaporator. Then ,the f l a s k was weighed again. I f t h e weight o f f i r s t f l a s k c o n t a i n i n g nonpolar l i p i d was .b g, and t h e second f l a s k p o l a r l i p i d was y g, t h a n t h e weight o f nonpolar l i p i d can be c a l c u l a t e d
by
s u b r a c t i n g b-a g (C g), and p o l a r l i p i d b yC
Nonpolar l i p i d f r a c t i o n
=
x 100%. (C+
Z )-
z
P o l a r l i p i d f r a c t i o n
-
x 100%. (C +Z )
3.2.2.3 F a t t y a c i d a n a l y s i s
M e t h y l e s t e r i f i c a t i o n o f f a t t y a c i d i n p o l a r and nonpo- l a r l i p i d has t o be done b e f o r e those sample can be i n j e c t e d t o t h e gas chromatography.
a. M e t h y l e s t e r i f i c a t i o n ( m o d i f i c a t i o n o f Supelco I n c . method, 1 9 7 5 )
About 5.0
-
15.0 mg l i p i d was p l a c e d i n a small v i a l . Then 0 . 4-
0.8 m l 8F3 methanol (14.5%, b / v ) was added i n t o t h e v i a l , and t h e cap o f v i a l was r e p l a c e d again. The v i a l was h e a t e d a t 6 0 O ~ f o r 60 minutes by p l a c i n g i t i n a h e a t e r b l o c k which was p l a c e d on a s m a l l h o t p l a t e . F u t h e r , 0.4-
0.8 m l s a t u r a t e d NaCl was added i n t o t h e v i a l , and i t was shaken v i g o r o u s l y . F i n a l l y 0.4 m l petroleum e t h e r was added i n t o t h e v i a l . The upper l a y e r s o l u t i o n c o u n t a i n methyl e s t e r o f f a t t y a c i d .b. F a t t y a c i d a n a l y s i s
F a t t y a c i d c o m p o s i t i o n analyses were done by u s i n g Shimadzu Gas Chromatography GC-SAM, w i t h Shimadzu Chromato- pac C-RGA. S t a t i o n a r y phase used i n t h i s a n a l y s i s was SP- 2330 on 100/120 chromosorb, 68% cyanoprophyl, wi%h maximum
were from Sigma Chemical Comp. USA. D e t e c t o r temperature was 260°c, i n j e c t o r temperature was 230°c, t h e column tem- p e r a t u r e was 150-21 OOC ( 5 O ~ / m i n u t e )
.
About 0.5 yl s t a n d a r d f a t t y a c i d s o l u t i o n was i n j e c t e d t o t h e GC. The chromatogram o f t h i s s t a n d a r d f a t t y a c i d would be used f o r sample f a t t y a c i d d e t e r m i n a t i o n . About 0.5 p1 methyl e s t e r o f sample f a t t y a c i d was i n j e c t e d t o GC.
I t had t h e same temperature c o n d i t i o n as above. The chro- matogram was compared t o t h e s t a n d a r d f a t t y a c i d chromato- gram. Each peak o f f a t t y a c i d from t h e sample chromatogram which has t h e same r e t e n t i o n t i m e as t h a t f r o m s t a n d a r d f a t t y a c i d chromatogram was c o n s i d e r e d t o be t h e same f a t t y a c i d as t h a t s t a n d a r d f a t t y a c i d .
3.2.3 Experimental d e s i g n and s t a t i s t i c a l methods
T h i s experiment used t h e c o m p l e t e l y randomized d e s i g n , w i t h f i v e t r e a t m e n t s and t h r e e r e p l i c a t e s f o r each t r e a t - ment. The w e i g h t o f egg produced compared t o t h e body w e i g h t , t h e s i z e o f eggs, t h e h a t c h i n g r a t e o f eggs, p e r - centage o f abnormal l a r v a e and f e e d c o n v e r s i o n were s u b j e c t - ed t o one-way a n a l y s i s of v a r i a n c e and Tuckey's procedure t e s t t o determine s i g n i f i c a n t d i f f e r e n c e s among t r e a t m e n t s ( S t e e l and T o r r i e
,
1980). Percentage o f f e r t i 1 iz e d eggs were n o t s u b j e c t e d t o s t a t i s t i c a l a n a l y s i s , because t h e r e were n o t enough samples o f eggs.ment:
a. The weight o f egg produced compared t o t h e body weight
I t was c a l c u l a t e d as:
t h e weight o f egg produced
x 100% t h e 4ody weight
.
Hatching r a t e :Hatching r a t e was c a l c u l a t e d as:
the
number o f h a t c h i n g eggsx 100 % t h e number o f incubated eggs
c . Percentage o f abnormal l a r v a e
Percentage o f abnormal l a r v a e was c a l c u l a t e d as: t h e number of abnormal l a r v a e
x 100 % t h e number o f t o t a l l a r v a e
d. V i a b i 1
i
t y o f t h e spermatozoaV i a b i l i t y o f t h e spermatozoa was c a l c u l a t e d as: t h e number o f f e r t i l i z e d eggs
x 100 % t h e number o f incubated eggs
e.
Feed conversionFeed conversion was c a l c u l a t e d
as:
t o t a l feed consumed by f i s h ( d r y b a s i c d i e t ) ( g ) f i n a l body w e i g h t - i n i t i a l body weight+dead f i s h body weight ( g )
3. Water
quality analysis
I V . RESULTS
4.1 The percentage o f egg weight/body weight, t h e s i z e o f eggs, t h e hatching r a t e o f eggs and t h e percentage o f &normal l a r v a e
N a t u r a l l y , c a t f i s h ( C t a r i a s batrachus Linn.) w i l l spawn
at
t h e age o f 10-
11 months, but i n t h i s experiment, f i s h s p a w n e dat
t h e age o f 14-
15 months. Presumably some groblems arosed t o t h e f i s h i n a d j u s t i n g t o the r e a r i n gsyswm d u r i n g t h i s experiment. However, a t t h a t age, a l l female f i s h were maturle, and they could then be induced t o
spawn.
The body weight o f f i s h before and a f t e r spawning, and the weight o f eggs produced
by
each female are shown i nf a b l e 3.
It
was found t h a t one female f i s h from treatment A ptuduced non v i a b l e eggs, which was seen t h a t the colour o f .eggs were opague, and a f t e r incubation,
those eggs d i d n o t h a e h a t a l l . Besides, three f i s h from treatment B and two f i s h o f treatment C were macroscopically determined as famaleat t h e beginning of t h i s experiment, but unfortunate-
l y they turned o u t t o be males l a t e r on (Table 3 ) .The weight o f egg produced by each f i s h varied between i n d i v i d u a l s , and bigger f i s h d i d n o t always produce more eggs compared
to
smaller ones. For example, one f i s h from treatment A weighing 260.0 g produced 7 . 0 g eggs (about 2 . 7 %t h e body weight). This was a l s o found i n t h e o t h e r f o u r Table 3. The b r o o d f i s h body weight before and a f t e r spawni ng, and the weight o f eggs produced by each female.
Treatments/ Body 8ody Weight T o t a l
' r e p l i c a t i o n s weight o f weight o f o f eggs weight
f i s h f i s h ( 9 ) compared
before a f t e r t o body
spawn i ng spawn i ng weight
( 9 ) f s )
( % I
0
I
I I
I11
I VD i e t A 1 2
D i e t 8 1 2 3
D i e t C 1
2
Table 3. Continued
Treatments/ Body Body Weight Total
r e p l i c a t i o n s weight o f * weight of o f eggs weight
f i s h f i s h ( 9 ) compared
before a f t e r t o body
spawning spawni ng weight
( 9 ( 9 )
( % I
0 I
I1
I11
IV
1: non v i a b l e eggs
a* t h e f i s h i s male, eventhough macroscopically determined as female
***
a l l eggs were used f o r t e s t o f the v i a b i l i t y o f wermatozoatreatments as we1 1. S t a t i s t i c a l l y , t h e r e were no d i f f e r - ences o f t o t a l egg w e i g h t / t o t a l body weight between t r e a t -
From t h i s r e s u l t i t was shown t h a t t h e d 6 and w 3 - f a t t y a c i d l e v e l i n t h e d i e t d i d n o t a f f e c t t h e weight of egg produced by those broodfish, b u t i t a f f e c t e d t h e q u a l i t y o f eggs. This was shown by the hatching r a t e o f eggs.
Table 4. The number o f eggs per 0.5 g egg from each t r e a t - ment
Rep1 i c a t i o n Treatments/diets
.
A
a
c
DE
Mean
T h e broodfish, fed on low *3 and G 6 - f a t t y a c i d l e v e l i n t h e i r d i e t ( d i e t A, contained 0.21% * - f a t t y a c i d and 0 . 0 3 % d 3 - f a t t y a c i d ) produced eggs w i t h low hatchabi 1
i
ty (see Table 5 and Appendix 8 and 9 ) . The hatching r a t e o f eggs improved by a d d i t i o n o f G36 and (33-fatty a c i d i n t h e d i e t , as was shown by treatments D and E ( d i e t D contained 1 -85% iw.6-fatty a c i d and 0.56% Q3-fatty a c i d l e v e l , whereas d i e tE
contained 0.26% a 6 - f a t t y a c i d and 1.68% a 3 - f a t t y a c i d l e v e l ) . The hatching r a t e o f eggs decreased again, when t h e a d d i t i o n o f a3 o r - - f a t t y a c i d i n t o t h e d i e t was t o o high, as was shown by treatments B and C ( d i e t BTable 5. The h a t c h i n g r a t e o f eggs produced by t h e brood- f i s h r e c e i v i n g experimental d i e t f o r n i n e months.
Rep1 i c a t i o n Treatments/diets
A
B
C D EMean 46.0 74.0 74.4 82.3 80.7
and 0.07% d 3 - f a t t y a c i d ) .
It
was i n t e r e s t i n g t o n o t e t h a t t h e b r o o d f i s h f e d on*
d i e t
E
which contained very low f a t t y a c i d (0.26%) and h i g h c 3 -HUFA (1.68%), and most o f i t were 20:5t;3 and 22;6 G33, c o u l d produce t h e same h a t c h i n g r a t e o f eggs as thoseb r o o d f i s h f e d on d i e t D, which contained 1.85Xc36-fatty a c i d and 0.56% U 3 - f a t t y acid, most o f i t was 18:3 C3.
The € F A - d e f i c i e n t eggs i n t r e a t m e n t A produced low h a t c h a b i l i t y and h i g h percentage o f abnormal l a r v a e as w e l l . Table 6 and Appendix 10 show t h e percentage o f 'abnormal l a r v a from each treatment. The percentage o f abnormal l a r v a ~ r o d u c e d
by
t h e b r o o d f i s h which r e c e i v e d lowG)6 and w 3 - f a t t ya c i d ( d i e t A) was h i g h e r than by those which r e c e i v e d h i g h ~ 2 6 and 4>3-fatty a c i d d i e t s ( d i e t s B, C, 0 , and E, P<0.05,
see Appendix 11 f o r s t a t i s t i c a l a n a l y s i s ) .
Table 6. The percentage o f abnormal l a r v a e produced by t h e b r o o d f i s h f e d experimental d i e t s f o r n i n e months.
R e p l i c a t i o n Treatments/diets
A B C D
Mean
f o r m i t y . I n F i g u r e 1, i t was seen t h a t t h e abnormal l a r v a e were from treatments E and A. F i s h l a r v a e , w i t h c u r l e d t a i l , c o u l d n o t swim s t r a i g h t b u t i n a c i r c l e . Eggs from t r e a t m e n t A (EFA-deficient eggs) would produce e i t h e r abnor- mal l a r v a e ( l i k e i n F i g u r e l e ) , o r normal l a r v a e , which most o f them had slow swimming movement.
H i s t o l o g i c a l l y , i t can be seen t h a t one o f seven sam- p l e s o f abnormal l a r v a e from t r e a t m e n t A showed d e f o r m i t y .of t h e head, e s p e c i a l l y i n t h e mouth region. F i g u r e 2 shows h i s t o l o g i c a l view of abnormal l a r v a ( a ) from t r e a t m e n t A , and normal l a r v a ( b ) from t r e a t m e n t A.
4.2 L i p i d content, p o l a r and nonpol8r l i p i d f r a c t i o n s , and f a t t y a c i d composition
-e
1 . Sample o f normal l a r v a from treatment E ( a ) ; and sampleo f abn
ma1 l a r v a from treatment E( b , c ) ,
and treatmen* ( d , e , f
1.
F i g u mple o f abnormal l a r v a ( a ) from
t egg (treatment A )
and l a r v a e o f treatments A, 8, C, D and E. The l i p i d content o f UF-eggs, 1 HAF-eggs, 11 HAF-eggs and l a r v a e vary between treatments. I n general, t h e l i p i d c o n t e n t o f eggs decreased s i n c e t h e f i r s t hour a f t e r f e r t i l i z a t i o n u n t i l t h e eggs hatched and became larvae. Besides, t h e l i p i d content o f t r e a t m e n t A l a r v a e was t h e lowest.
t nrr 11 nrr
F i g u r e 3. Histogram of l i p i d contenrt o f W-eggs, 1 HAF-
eggs,
1 1 HAF-eggs and l a r v a e produced byb r o o d f i s h fed on ex9erimental d i e t s f o r n i n e months.
P/NP l i p i d r a t i o s o f t h e eggs changed a l s o s i n c e the f i r s t hour a f t e r f e r t i l i z a t i o n . The P/NP l i p i d r a t i o increased s i n c e t h e f i r s t hour a f t e r f e r t i l i z a t i o n , except i n t r e a t - ment A. Besides, t h e P/NP l i p i d r a t i o o f l a r v a e was h i g h e r Shan t h a t o f t h e Uf-eggs, and t h e P/MP r a t i o o f l a r v a e from tmaernent A was t h e h i g h e s t .
T efFe&s o f 0 3 a n d a 6 f a t t y a c i d l e v e l s i n t h e
f i g u r e 4. P o l a r / w n p o l a r 1 i p i d r a t i o s d u r i n g embryogenesis from treatments A, 8 , C , 0, and
E.
b r d f i s h d i e t t o t h e embryogenesis c o u l d be seen c l e a r l y
1 4 , 1 5 , 16, 17, 18). F i s h c o u l d n o t s y n t h e s i z e c 3 6 a n d G l 3 -
f a t t y a c i d . I n general, f i s h would synthesize h i g h W9-fatty a c i d
if
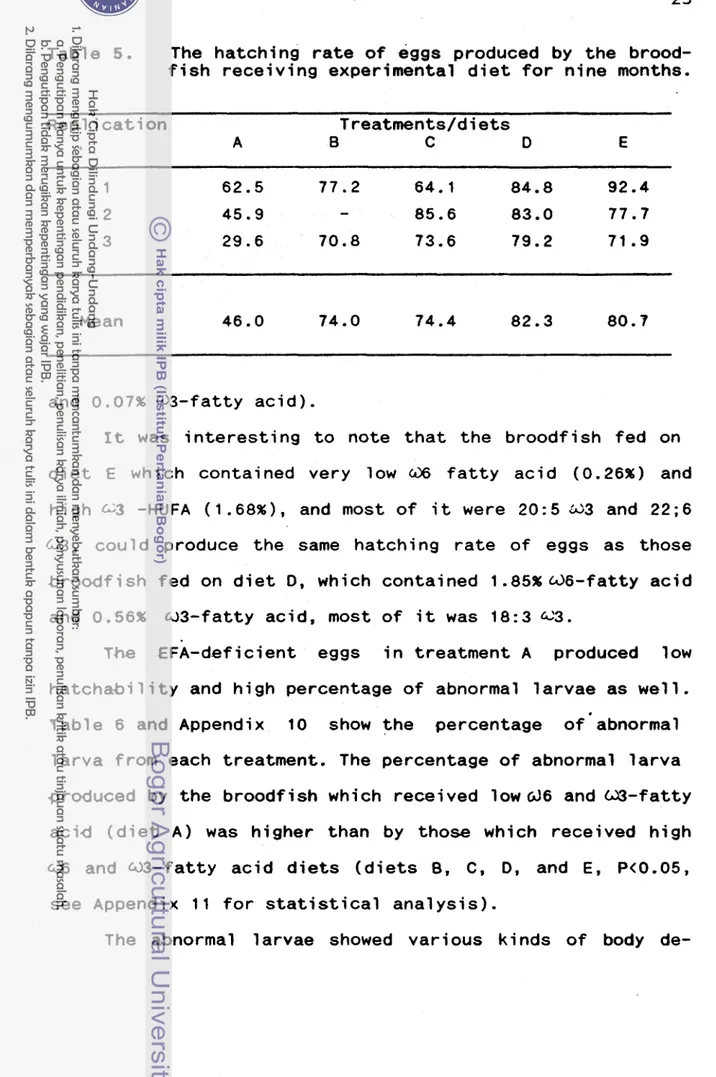
t h e f i s h r e c e i v e d lowW6 and W 3 - f a t t y a c i d i n t h e i r d i e t . So, Show f a t t y a c i d s would be t h e focus o f f a t t y acid e v a l u a t i o n d u r i n g embryogenesis.From F i g u r e 5, low G;6 and d 3 - f a t t y a c i d l e v e l s o f UF-
eggs
were a f f e c t e d by t h e low 6.8 and w 3 - f a t t y a c i d s i n the*b r w d f i s h d i e t . t h e d 9 - f a t t y a c i d l e v e l o f t h e UF-eggs was h i g h , e s p e c i a l l y i n t h e NP l i p i d f r a c t i o n . The unsaturated
f a t t y a c i d l e v e l o f eggs changed s i n c e t h e f i r s t hour a f t e r F e r t i l i z a t i o n . K t was i n t e r e s t i n g t o note, thatc36,W3 and W 9 - f a t t y a c i d l e v e l of 1 HAF-eggs, 1 1 HAF-eggs, and t h e l a r v a s were h i g h e r than t h a t from t h e UF-eggs. I t seemed
&hat
a c e r t a i n l e v e l of d 6 ,0 3
and &.%-fatty a c i d s wereneeded
for embryogenes i s.
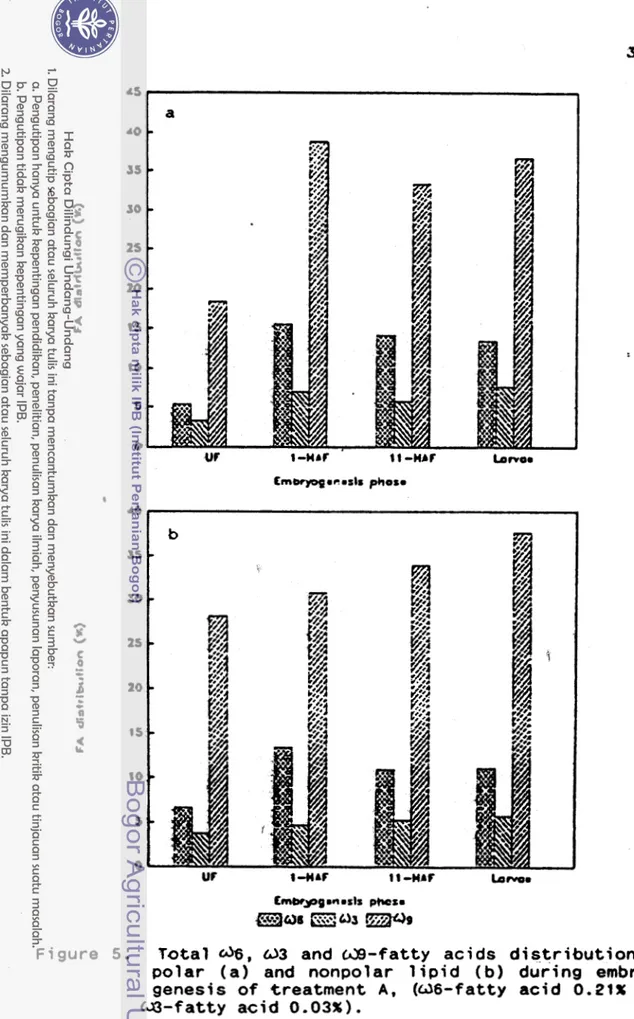
F i g u r e 6 shows t h e fatty a c i d d i s t r i b u t i o n o f UF-eggs, 1 HAF-eggs, 1 1 HAF-eggs, and t h e l a r v a e from treatment 8
X T a t t y a c i d d i s t r i b u t i o n of UF-eggs p o l a r 1 ip i d was missing
uc
1-nrr tc-nbr
E m w p p o e o s ~ s phosm
40
'Of
1
a g a i n i n t h e l a r v a e . On t h e o t h e r hand, t h e d 9 - f a t t y a c i d l e v e l increased i n 1 HAF-eegs, b u t a f t e r t h a t i t decreased a g a i n i n 11 HAF-eggs and l a r v a e . F i n a l l y t h e a 6 and
a3
- f a t t y a c i d l e v e l o f l a r v a e was t h e same as t h a t o f UF-eggs,-
b u t t h e a 9 - f a t t y a c i d l e v e l was h i g h e r t h a t t h a t o f UF-eggs. D i e t C contained very h i g h d 6 - f a t t y a c i d ( 2 . 2 4 % ) and low u 3 - f a t t y a c i d (0.07%). T h i s h i g h d 6 - f a t t y a c i d and low U 3 - f a t t y a c i d l e v e l i n t h e b r o o d f i s h d i e t would a f f e c t t h e and 0 3 - f a t t y a c i d l e v e l o f t h e eggs produced, as i t c o u l d be seen from F i g u r e 7. UF-eggs a l s o contained very h i g h d 6 - f a t t y a c i d , and very low W 3 - f a t t y a c i d l e v e l . F u r t h e r , t h e U 6 - f a t t y a c i d l e v e l o f nonpolar l i p i d from 1 HAF-eggs, 11 HAF-eggs, and l a r v a e were h i g h e r than t h a t from t h e UF-eggs. But t h e a 6 - f a t t y a c i d l e v e l o f p o l a r l i p i d f r o m 1 HAF-eggs, 11 HAF-eggs, and l a r v a e were constant. The G)3 and d 9 - f a t t y a c i d l e v e l o f p o l a r and nonpolar l i p i d a l s o changed s i n c e t h e f i r s t hour a f t e r f e r t i l i z a t i o n as w e l l . Although d i e t C contained h i g h l e v e l o f d 6 - f a t t y a c i d , t h e eggs produced by t h e b r o o d f i s h which r e c e i v e d t h i s d i e t ,
s t i l l contained h i g h l e v e l o f d 9 - f a t t y a c i d .
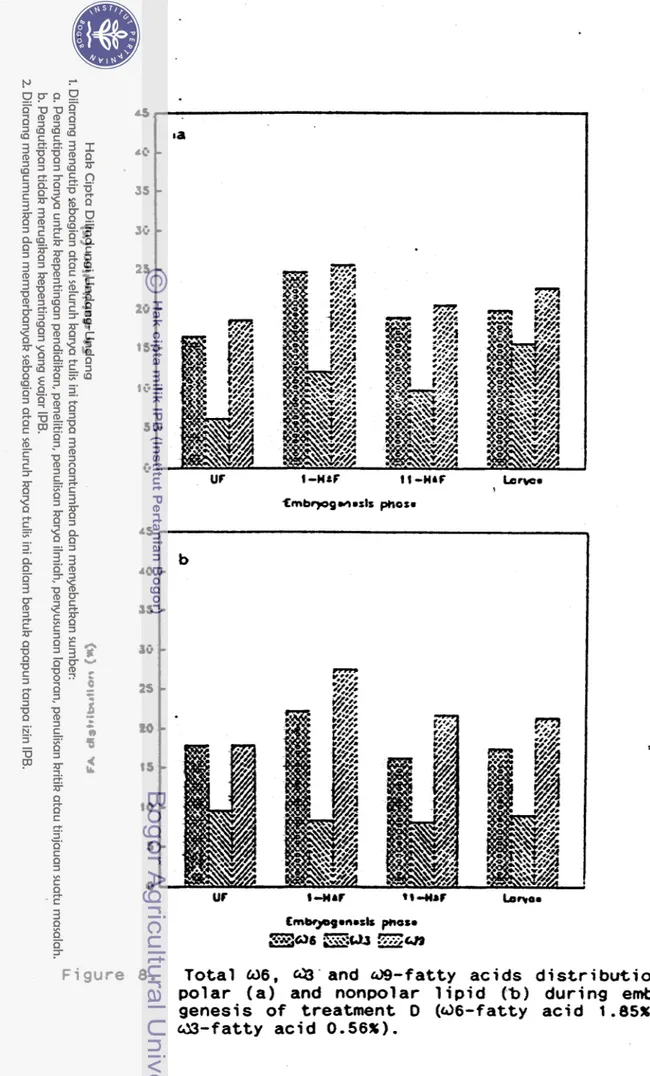
Figure 8. T o t a l U 6 , and @ - f a t t y acids d i s t r i b u t i o n od
p o l a r ( a ) and nonpolar l i p i d ("b) during embryo- genesis o f treatment 0 ( d 6 - f a t t y a c i d 1 .BSX and
by t h e (dl a n d Q 6 - f a t t y a c i d l e v e l i n t h e b r o o d f i s h d i e t , t h e same symptom as i n treatments A, 8 , and C. Both d i e t s C
and*
D.have h i g h h . 6 - f a t t y a c i d l e v e l , b u t d i e t C has much h i g h e r w 6 - f a t t y a c i d l e v e l than d i e t D. So, i t c o u l d be seen from F i w r e 7 and 8, t h a tthe
fJ6-fatty a c i d l e v e l o f U F - e ~ g s i n treatment C was s l s o h i g h e r t h a n t h a t f r o m treatment 0. Qnt h e o t h e r hand, d i e t C contained lower ( 3 3 - f a t t y a c i d l e v e l than d i e t D, 'and t h i s s l s o a f f e c t e d t h e a 3 f a t t y a c i d l e v e l o f W-eggs from treatments C and D. The d 9 - f a t t y acid l e v e l o f eggs from treatment O was lower than treatments A and C, i n d i c a t i n g t h a t t h e f i s h r e c e i v e d enough a3
an&
0 6 - f a t t y a c i d f r o m d i e t D.En treatment D, i t was shown t h a t t h e f a t t y a c i d compor s i t i o n of eggs a l s o changed s i n c e t h e f i r s t hour a f t e r f e r t i l i z a t i o n , t h e same as treatments A , B,
and
C. The w6f a t t y a c i d l e v e l o f non p o l a r l i p i d from 1 HAF-eggs was h i g h e r than from UF-eggs, b u t a f t e r t h a t , t h e d 6 - f a t t y a c i d l e v e l of 11 HAF-ews and t h e l a r v a were almost t h e same as
from UF-eggs. I n p o l a r 1 i p i d , 0 6 - f a t t y a c i d l e v e l o f 1 HAf- eggs, 11 HAF-e9gs and l a r v a were h i g h e r than t h a t from t h e UF-eggs. F u r t h e r
,
t h e d 3 - f a t t y a c i d l e v e l o f p o l a r lipid., t h e @ - f a t t y a c i d l e v e l o f nonpolar and p o l a r l i p i d from 1the same as from UF-eggs.
D i e t E contained low c 6 - f a t t y a c i d l e v e l , and high
a3
- f a t t y a c i d l e v e l , which most o f them were d3-HUFA (seeAppendix 3). The & - f a t t y a c i d l e v e l o f UF-eggs from t r e a t - ment € was lower than t h a t from treatments A, and C, i t was t h e same as i n treaement 0 .
The f a t t y a c i d composition also changed since the f i r s t
hour a f a r f e r t i l i z a t i o n . The G6 andc23-fatty a c i d l e v e l o f
nonpolar l i p i d and the a 9 - f a t t y acid o f p o l a r and nonpolar
l i p i d
f r o m
1 HAF-eggs, 11 HAF-eggs, and from larvae were higher than from t h e UF-eggs. On the other hand, the 06- f a t t y a c i d and c U - f a t t y acid l e v e l o f p o l a r l i p i d from 1HAF-eggs, 11 HAF-eggs, and from the larvae was lower than
From t h e UF-eggs.
From
these
a l l f a t t y a c i d d i s t r i b u t i o n analysis o f p o l a r and nonpolar 1 i p i d o f eggs and larvae, i t seemed t h a tthe f a M y a c i d composition changed since the e a r l y embryo-
genesis. Besides t h a t , i n general, &%-fatty acid, U3-
f a t 4 y acid
and
U S - f a t t y acid l e v e l from 1 HAF-eggs, 1 t HAF-eggs, and fromlarva
were higher than t h a t from UF-egss.
3% seemed t h a t the balance o f .those f a t t y acide, espe- c i a l l y i n t h e p o l a r l i p i d of eggs and larvae was important.
The
f a t t y = i d dJstri-butionof
1 iv e r , and muscle weres
a l s o a f f e c t e d by the f a t t y acid l e v e l o f the broodfish d i e t .
F i g u r e 9 . T o t a l (36, 0 3 and - - f a t t y acids d i s t r i b u t i o n o f
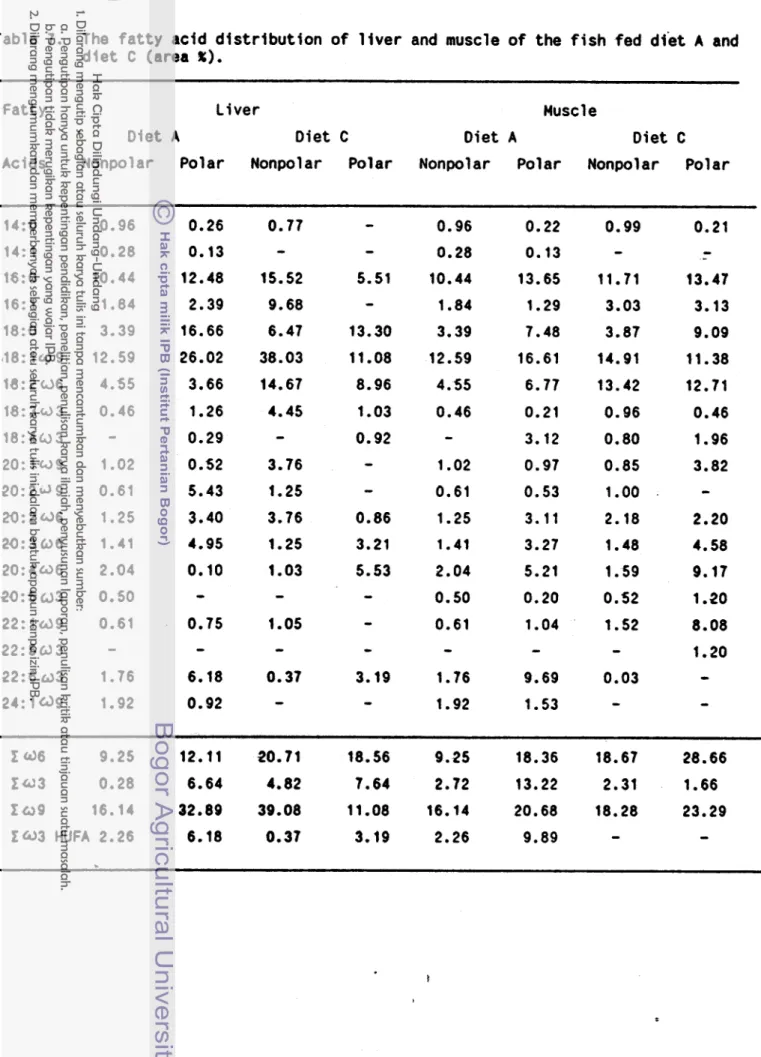
Table
7.
The f a t t y acid d i s t r i b u t i o n o f l i v e r and muscle o f the f i s h fed d i e t A and d i e t C (area %).Fatty Liver Husc 1 e
Diet A Diet C Diet A D i e t C
Acids Nonpolar Polar Nonpolar Polar Nonpolar Polar Nonpolar Polar
2
<36
9.25
12.11
20.71
18.56
9.25
18.36
18.67
28.66
f a 3
0.28
6.64
4.82
7.64
2.72
13.22
2.31
1.66
f
a916.14
32.89
39.08
11.08
16.14
20.68
18.28
23.29
treatment A and C s t i l l contained h i g h 0 9 - f a t t y a c i d l e v e l , t h e same as i n t h e eggs.
4.3 The v i a b i l i t y o f spermatozoa
8esides t h e h a t c h a b i l i t y o f eggs, t h i s experiment
showed t h a t t h e €FA-deficient d i e t d i d n o t produce low q u a l i t y o f spermatozoa. The spermatozoa from t h e male which
r e c e i v e d d i e t A, 6 , C, D , o r E co"ld f e r t i l i z e t h e eggs more than 90.0% (see Table 8 and Appendix 1.6). There. were no d i f f e r e n c e s between t h e percentage o f f e r t i l i z e d - e g g s from
a1 1 treatments.
Table 8. The percentage of eggs which were f e r t i l i z e d by t h e spermatozoa from each treaament
T h e percentage Spermatozoa from treatments
o f f e r t i l i z e d eggs f r o m t r e a t -
ment A
a
c
o
E4.4
Seed
convers4onTable 9 and Appendix 20 showed t h e feed conversion o f d i e t s A, B, C , 0 and €. The feed conversion o f EFA-
d e f i c i e n t d i e t ( d i e t A )
was
t h e same as those o f o t h e r EFA-Table 9. Feed conversion ( d r y basic d i e t )
R e p l i c a t i o n Treatments/diets
8 A 6
c
D EMean 2.95 2.41 2.03 2 . 1 1
2.38
4.5 Water q u a l i t y
A t t h e beginning o f t h i s experiment, water a l k a l i n i t y was a t a range o f 5 2
-
60 ppm CaC03 eq; dissolved oxygen was7 . 2
-
7.4 ppm; t o t a l ammonia was 0.10-
0.19 ppm; and pH was 7.0-
7.4 (see Appendix 22).It
could be c l a s s i f i e d as good water q u a l i t y f o r the f i s h .On t h e s i x t h month o f t h i s experiment, t h e t o t a l ammo-
n i a was a t
a
range o f 0.32 -3.231 ppm, and dissolved oxygen was 2.9-
5.8 ppm (see Appendix 23).Some o f t h e f i s h died,
and
some d i d n o t grow a t a l l . I pressume t h a t t h e stocking r a t e of t h e f i s h was t o o high.ESSENTIAL FATTY ACID REQUIREMENTS
OF
CATFISH
(Clariss
bstrachus
Linn.)
FOR
BROODSTOCK DEVELOPMENT
by
:
ING
MOKOGINTA
PROGRAM
PASCASARJANAlNSTlTUT
PERTANIAN BOGOR
ABSTRACT '
Mokoginta, I n g . E s s e n t i a l F a t t y Acid Requirements o f c a t f i s h ( C l a r i a s batrachus L i n n . ) f o r Broodstock Development
1
Supervised by P r o f . Dr. 0. 5 . Moeljohardjo, P r o f . O r . T. Takeuchi, Dr. I r . Kusman Sumawidjaja, Dr. I r . Dedi F a r d i a z , and P r o f . Dr. 'Sumardi Sastrakusumah (deceased)].T h i s experiment was conducted t o determine e s s e n t i a l f a t t y a c i d requirements o f c a t f i s h ( C l a r i a s batrachus L i n n . ) for broodstock development. The b a s i c d i e t s u p p l i e d t o t h e t e s t f i s h had p r o t e i n and f a t c o n t e n t 46.51
-
48.29% and 5.8-
5.96% r e s p e c t i v e l y . The f i v e t e s t d i e t s d i f f e r e d i n t h e e s s e n t i a l f a t t y a c i d (EFA) l e v e l . D i e t A contained 0 . 2 1 % ( 3 6 - f a t t y a c i d and 0.03% S 3 - f a t t y a c i d ; d i e t B 0.67% d 6 - f a t t y a c i d and 2 . 0 9 % & 3 - f a t t y a c i d ; d i e t C 2 . 2 4 K U 6 - f a t t ya c i d and 0 . 0 7 % & 3 - f a t t y a c i d ; d i e t
0 1.85%(;36-fatty a c i d and
0.56% a 3 - f a t t y a c i d ; d i e tE
0.26% a 6 f a t t y a c i d and 1.68% d 3 - f a t t y a c i d . Most o f U S - f a t t y a c i d i n d i e t B and D were1$:3 (33, on t h e o t h e r hand those i n d i e t E were 20:s c33 and 22:6 W 3 .
The f i s h were f e d t h e experimental d i e t s , a t s a t i a t i o n , once a day a t 5:00 p.m. S i x females and t h r e e males were k e p t i n each tanks ( s i z e 80 x 80 x 100 cm)
.
One hundred p e r c e n t o f t h e water i n t h e tanks was changed, once i n twodays. The water b m p e r a t u r e was k e p t constant a t 25O
-
27%.weight/body weight, t h e eggs s i z e , t h e h a t c h i n g r a t e o f eggs and t h e percentage of abnormal l a r v a e produced by each b r o o d f i s h and t h e feed conversion were used as parameters t o e v a l u a t e t h e d i f f e r e n c e s between treatments.
Samples o f u n f e r t i 1 i zed-eggs (UF-eggs
1 ,
one h o u r a f t e r f e r t i l i z a t i o n - e g g s ( 1 HAF), eleven hours a f t e r f e r t i l i r a -tion-eggs ( 1 1 HAF), and sample of l a r v a e were taken f o r l i p i d c o n t e n t , p o l a r and nonpolar l i p i d f r a c t i o n a n a l y s i s , a d t h e f a t t y a c i d composition a n a l y s i s . Also, samples o f l i v e r and muscle o f t h e f i s h wore taken f o r f a t t y a c i d composition a n a l y s i s . These chemical analyses were done i n o r d e r t o study t h e metabolism of e s s e n t i a l f a t t y a c i d s i n
t h e embryogenesis processes.
The spermatozoa of male f i s h from treatments A, 8 ,
6 ,
0, and E f e r t i l i z e d t h e eggs from treatment 0 , o r E. Thepercentage o f f e r t i l i z e d - e g g s was used as an i n d i c a t o r t o e v a l u a t e t h e v i a b i l i t y o f spermatozoa.
The mean percentage of egg weight/body weight and eggs s i z e were n o t d i f f e r e n t between treatments
(P
>
0.05). The€ F A i n -the b r o o d f i s h d i e t a f f e c t e d t h e q u a l i t y r a t h e r than
t h e q u a n t i t y o f eggs.
The b r o o d f i s h , which r e c e i v e d low a - f a t t y a c i d and w3-
f a t t y a c i d l e v e l i n t h e i r d i e t ( d i e t A ) , produced eggs w i t h l o w h a c h a b i l i t y (46.0%). The h a t c h i n g r a t e o f eggs improved b y a d d i t i o n more a 6 - f a t t y a c i d and 0 3 - f a t t y a c i d i n the bmdf i s h d i e t , as was shown
by treatments
D (82.3%) and E X80.7X) (QC0.05). T h e h a t c h i n g r a t e of eggs decreased again, when t h e a d d i t i o n of U 6 and d 3 - f a t t y a c i d i n t o t h e b r o o d f i s h d i e t w a s Coo h i g h , as was shown by treatments 8( 7 4 . 9 % ) and C (74.4%).
s i o n o f t r e a t m e n t A was n o t d i f f e r e n t from those o f o t h e r t r e a t m e n t s ( P , > 0.05).
I t c o u l d be concluded t h a t 1.85% d 6 - f a t t y a c i d and 0 . 5 6 % d 3 - f a t t y a c i d (most o f ( 2 3 - f a t t y a c i d was 18:3 (33) was r e q u i r e d by c a t f i s h ( C l a r i a s batrachus L i n n . ) broodf i s h i n o r d e r to produced h i g h qual i t y o f eggs. On t h e o t h e r hand 0.26% d 6 - f a t t y a c i d t o g e t h e r w i t h 1.68% G3-HUFA i n t h e b r o o d f i s h d i e t c o u l d produced h i g h qual i t y of eggs. The (a6
o r a 3 - f a t t y a c i d s c o n t e n t i n t h e b r o o d f i s h d i e t should not be more than 2.0%, a 6 - f a t t y a c i d n o t l e s s than 0.3% and d3-
f a t t y a c i d n o t l e s s than 0.6% o f a l e v e l a t 6.0% l i p i d .
From the f a t t y a c i d composition a n a l y s i s o f eggs and l a r v a e , i t was found t h a t t h e a 6 and G 3 - f a t t y a c i d c o n t e n t o f t h e broodf i s h d i e t would a f f e c t those o f t h e eggs. The l i p i d c o n t e n t , t h e p o l a r (P)/nonpolar (NP) l i p i d r a t i o , and t h e f a t t y a c i d composition i n t h e eggs changed s i n c e t h e f i r s t hour a f t e r f e r t i l i z a t i o n . I n general, t h e p o l a r / n o n p o l a r r a t i o from 1 HAF-eggs, 1 1 HAF-eggs, and l a r v a were h i g h e r than t h a t from t h e UF-eggs. Besides t h a t , h i g h l e v e l of c36 and d 3 - f a t t y a c i d were found i n t h e eggs s i n c e t h e f i r s t hour a f t e r f e r t i l i z a t i o n u n t i l t h e eggs hatched and became l a r v a e .
It
was concluded t h a t t h e ~ 3 6 and d 3 - f a t t y a c i d p l a y an i m p o r t a n t r o l e i n embryogenesis.RINGKASAN
Mokoginta, I n g . Kebutuhan I k a n l e l e ( C l a r i a s batrachus L i n n . ) Akan Asam Lemak Esensial Bagi Perkembangan Induk CDi bawah bimbingan P r o f . O r . O.S. M o e l j o h a r d j o sebagai ketua; P r o f . D r . T. Takeuchi sebagai waki1 ketua; D r . I r . Kusman Sumawidjaja, D r .
Ir.
Dedi F a r d i a z , dan P r o f . D r . Sumardi Sastrakusumah (almarhum) sebagai anggota].P e n e l i t i a n i n i b e r t u j u a n untuk menentukan kebutuhan asam lemak e s e n s i a l o l e h i k a n l e l e ( C l a r i a s batrachus L i n n . ) untuk perkembangan induk. Lima macam makanan percobaan yang digunakan berkadar p r o t e i n 46.51
-
48.29%, lemak 5.81-
5.96% ( d a r i bobot k e r i ng makanan),
t e t a p i berbeda dalam kandungan asam lemak L;6 dan asam lemak d3-nya. Makanan A mengandung 0.21% asam lemak d 6 dan 0.03% asam lemak ~ 2 3 ; makanan B 0.67% asam lemak d 6 dan 2.09% asam lemak u 3 ; makanan C 2.24% asam lemak 0 6 dan 0.07% asam lemak d 3 ; makanan D 1.85% asam l e m a k d 6 dan 0.56% asam lemakc33; maka- nanE
0.26% asam lemak Mi dan 1.68% asam lemakd3. Sebagian besar asam .lemak W3 dalam makananB
dan D adalah 18:3 a 3 ,sedangkan makanan E a