DAN POTENSINYA DALAM PENGENDALIAN
BUSUK BUAH PHYTOPHTHORA
EFI TODING TONDOK
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi ‘Keragaman Cendawan Endofit pada Buah Kakao dan Potensinya dalam Pengendalian Busuk Buah Phytophthora’ adalah karya saya dengan arahan dari komisi pembimbing dan belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir setiap topik disertasi ini.
Bogor, Januari 2012
Efi Toding Tondok
EFI TODING TONDOK. Diversity of Endophytic Fungi on Pods of Cacao and Its Potency as Biocontrol Agents of Black Pod Disease of Cacao. Under direction of MEITY SURADJI SINAGA, WIDODO and MAGGY THENAWIDJAJA SUHARTONO.
Endophytic fungi have been reported capable to suppress development of black pod disease in cacao. In Indonesia, research on endophytic fungi in general has just been started. The aim of this study were to investigate factors related to the incidence of black pod disease in the field, to explore the diversity of endophytic fungi during ENSO simulation period and its correlation to black pod disease, and to examine the potency of selected endophytic fungi in controlling black pod disease. Field observation was conducted in Palolo and Kulawi, cacao plantations located at the border of Lore Lindu National Park, Central Sulawesi, laboratory and greenhouse experiments were performed at Bogor Agricultural University, Bogor, Indonesia and Georg-August University, Goettingen, Germany. Black pod disease incidence was highest in plots that: were located at > 600 m a.s.l.; which were previously occupied by pristine forest or perennial plants; had prior intensive inputs of pesticides and fertilizers; and had a high frequency of weeding. While application of nitrogen fertilizer, distance to pristine forests, plant age, canopy cover, diversity of endophytic fungi and degree of shading by shade trees were not associated with the disease development. A total of 2843 isolates were obtained from 1440 husk segments of healthy pods. By morphological and molecular techniques, the identification of these isolates resulted in 37 species of fungi, six belong to Basidiomycetes, 30 are Ascomycetes and one is unidentified yet. The diversity of endophytic fungi, based on Shannon’s and Simpson’s index, was higher during lower rainfall (January-April 2008) compared to higher rainfall condition (March-April 2007). The dominance of endophytic fungi changed with season, Fusarium lichenicola was the most dominance during wet condition and Phomopsis columnaris during dry condition. Several species, such as Fusarium sp. T47a, Xylaria sp., Polyporaceae DIS 126a, Aschersonia sp., Calocybe gambosa, and Xylariaceae were negatively correlated to the incidence of black pod disease. Among the tested endophytic fungi, Xylariaceae and Callocybe gambosa generated the highest effectiveness control level of black pod disease on detached pods, 38.8% and 33.8%, respectively, followed by Resinicium friabile and Aschersonia treatment, 17.4% dan 12.7% respectively. Growth inhibition of pathogen and induced resistance of plant were partially responsible for disease suppression by Xylariaceae, C. gambosa, R. friabile and Aschersonia sp.
EFI TODING TONDOK. Keragaman Cendawan Endofit pada Buah Kakao dan Potensinya dalam Pengendalian Busuk Buah Phytophthora. Dibimbing oleh MEITY SURADJI SINAGA, WIDODO dan MAGGY THENAWIDJAJA SUHARTONO
Gerakan Nasional Kakao yang dicanangkan pemerintah pada tahun 2006 bertujuan untuk meningkatkan produksi dan mutu kakao Indonesia. Peningkatan produksi dengan cara penambahan areal pertanaman kakao banyak dilakukan petani dengan merambah areal hutan. Pembukaan areal hutan tentu berpengaruh terhadap ekosistem yang telah ada. Untuk melihat pengaruh lingkungan hutan dan praktek budidaya yang diaplikasikan di lapangan terhadap kejadian penyakit busuk buah kakao yang disebabkan oleh Phytophthora palmivora, maka dilakukanlah serangkaian perlakuan dan pengamatan di lapangan di areal sekitar Taman Nasional Lore Lindu, Sulawesi Tengah, yang merupakan hutan primer. Pada salah satu plot dilakukan juga simulasi kekeringan yang berasosiasi dengan ENSO (El-nino Southern Oscillation) untuk melihat fluktuasi keragaman cendawan endofit dalam kondisi kering dibandingkan dengan kondisi basah dan korelasinya dengan kejadian penyakit busuk buah. Selanjutnya, enam cendawan endofit yang berkorelasi negatif dengan kejadian penyakit di lapangan diuji kemampuannya dalam menekan perkembangan penyakit busuk buah kakao.
Faktor yang mempengaruhi epidemik penyakit busuk buah kakao diamati pada 86 plot di dua kecamatan yang berbatasan dengan Taman Nasional Lore Lindu, Sulawesi Tengah. Faktor yang diamati berupa lingkungan (ketinggian tempat, jarak ke hutan primer, praktek budidaya sebelumnya, sejarah penggunaan lahan, keragaman morfospesies cendawan endofit), praktek budidaya (frekuensi penyiangan, tingkat naungan, pemupukan, tutupan kanopi), dan umur tanaman kakao. Analisis korespondensi sederhana (Simple Correspondence Analysis, SCA) dilakukan untuk melihat hubungan antara epidemi busuk buah dengan peubah yang diamati. Frekuensi penyiangan, ketinggian tempat, praktek budidaya sebelumnya, dan sejarah penggunaan lahan sebelum ditanami kakao berasosiasi nyata dengan epidemi busuk buah kakao, sedangkan aplikasi pupuk nitrogen, jarak ke hutan primer, umur tanaman, penutupan oleh tajuk kakao, keragaman morfospesies cendawan endofit dan tingkat naungan oleh pohon peneduh tidak berasosiasi dengan perkembangan penyakit. Kejadian penyakit tertinggi ditemukan pada plot: yang terletak >600 m dpl; sebelum ditanami kakao merupakan hutan primer atau tanaman tahunan; sebelumnya mendapat input pestisida dan pupuk yang intensif, dan dengan frekuensi penyiangan yang tinggi.
selama kondisi basah dan Phomopsis columnaris selama kondisi kering. Beberapa spesies, seperti Fusarium sp. T47a, Xylaria sp, Polyporaceae DIS 126a, Aschersonia sp., Calocybe gambosa, dan Xylariaceae berkorelasi negatif dengan kejadian penyakit busuk buah. Cendawan-cendawan ini berpotensi sebagai agens biokontrol penyakit busuk buah kakao.
Penelitian selanjutnya bertujuan untuk menguji potensi cendawan endofit terpilih sebagai agens biokontrol busuk buah kakao. Enam cendawan endofit terpilih dari total 37 spesies (2843 isolat) yang berhasil diisolasi, diuji keefektifannya sebagai agens pengendali hayati. Buah kakao di lapangan disemprot dengan propagul cendawan, setelah dua minggu buah dipanen dan diinokulasi dengan patogen P. palmivora di laboratorium. Perkembangan gejala penyakit pada buah diamati, dihitung periode laten, keparahan penyakit, laju infeksi (r), nilai AUDPC dan tingkat keefektifan penekanan penyakit busuk buah. Kemampuan cendawan endofit menginduksi ketahanan tanaman dan penghambatan pertumbuhan patogen in-vitro juga dianalisis. Cendawan Xylariaceae dan Calocybe gambosa menghasilkan tingkat penekanan penyakit paling tinggi, berturut-turut sebesar 38.8% dan 33.8%, diikuti oleh Resinicium friabile dan Aschersonia masing-masing sebesar 17.4% dan 12.7%. Cendawan Pestalotiopsis dan Fusarium tidak efektif menekan keparahan penyakit. Periode laten sangat berkaitan dengan keparahan penyakit. Penghambatan pertumbuhan patogen dan induksi ketahanan tanaman berperan dalam penekanan penyakit oleh Xylariaceae, C. gambosa, R. friabile dan Aschersonia.
(c) Hak cipta milik Institut Pertanian Bogor, tahun 2012 Hak cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tersebut tidak merugikan kepentingan yang wajar IPB. 2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya
BUSUK BUAH PHYTOPHTHORA
EFI TODING TONDOK
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor
pada Program Studi Fitopatologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup: 1. Dr Ir Sudradjat, MS 2. Dr Ir A Munif, MSc Agr
Penguji pada Ujian Terbuka: 1. Dr Ir Ade Wachjar, MS
Judul Disertasi : Keragaman Cendawan Endofit pada Buah Kakao dan Potensinya dalam Pengendalian Busuk Buah Phytophthora Nama : Efi Toding Tondok
NIM : A461040011
Disetujui, Komisi Pembimbing
Prof Dr Ir Meity Suradji Sinaga, MSc Ketua
Dr Ir Widodo, MS Prof Dr Ir Maggy Thenawidjaja Suhartono
Anggota Anggota
Diketahui,
Ketua Program Studi Fitopatologi Dekan Sekolah Pascasarjana
Dr Ir Sri Hendrastuti Hidayat, MSc Dr Ir Dahrul Syah, MSc Agr
Puji dan syukur penulis panjatkan ke hadirat Tuhan Yang Maha Kuasa atas segala berkat dan karunia-Nya sehingga penulis dapat menyelesaikan penelitian dan penulisan disertasi ini.
Penelitian ini didukung oleh proyek Stability of Rainforest Margins in Indonesia (STORMA) di bawah pendanaan German Research Foundation (DFG-SFB 552), juga didukung oleh Program Sandwich-like Dikti 2008, PT AGSI, Klinik Tanaman IPB: diucapkan terimakasih kepada para donatur.
Penulis mengucapkan terima kasih kepada Prof Dr Ir Meity S Sinaga MSc, Dr Ir Widodo MS, dan Prof Dr Ir Maggy Thenawidjaja S sebagai komisi pembimbing yang telah memberikan saran, dorongan, kritik dan dukungan kepada penulis sehingga disertasi ini dapat terwujud.
Ucapan terima kasih disampaikan kepada Departemen Proteksi Tanaman, Faperta dan IPB yang telah memberikan kesempatan kepada penulis untuk menempuh studi S3.
Ucapan terima kasih juga disampaikan kepada PTP VIII Saguling, Cianjur atas ijin penggunaan tanaman kakao untuk uji bioassay cendawan endofit.
Terima kasih juga dihaturkan kepada penguji luar komisi pada sidang tertutup, Dr Ir Sudradjat MS dan Dr Ir A Munif MSc Agr dan sidang terbuka Dr Ir Ade Wachjar MS dan Dr Ir Darmono Taniwiryono MSc. Saran dan masukan dari penguji turut memperkaya isi disertasi ini.
Ucapan terima kasih disampaikan kepada Prof Dr Stefan Vidal, Prof Dr Petr Karlovsky, Dr Suryo Wiyono, Dr Yann Clough, Dr Hermanu Triwidodo yang telah turut memberi masukan dan memperkaya isi dari disertasi ini, Dr Endah R Palupi atas ‘pelajaran bahasa Inggris’ dan koreksi beberapa istilah agronomi yang digunakan dalam disertasi ini. Terima kasih juga kepada Stefan Erasmi dan Dominic Seidel untuk berbagi data lebar tajuk kakao, group Profesor Gravenhost untuk data cuaca di Rahmat dan Toro, Manajemen Proyek STORMA, Carsten Schmitd, Hasrul, dan 'Kelompok peneliti kakao cs di Palu'. Ucapan terima kasih juga disampaikan kepada Irene, Nenk Agustina, Ita Juwita yang banyak membantu pekerjaan di laboratorium. Demikian juga disampaikan ucapan terima kasih kepada rekan Ahmad Rizali untuk MDS dan Bonjok Istiaji yang sangat membantu di saat-saat akhir mengedit bahan presentasi. Juga disampaikan ucapan terima kasih kepada rekan-rekan Fitopatologi dan Entomologi atas segala bantuan dan kebersamaan selama ini, serta kepada semua pihak yang tidak dapat disebutkan satu persatu di sini.
Ucapan terima kasih dihaturkan kepada kedua orangtua, kak Ita, kak Uti, adek Pina, adek Harun dan seluruh keluarga yang terus mendukung dalam doa, kesabaran dan segala perhatian yang diberikan. Khusus kepada suami tercinta, Harsman Tandilittin, terima kasih telah hadir dalam hidup saya, terima kasih atas segala dorongan, pengertian dan kesabarannya selama ini.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2012
Penulis dilahirkan di Tana Toraja, 23 Januari 1971, sebagai anak ketiga dari Bapak JP Toding Tondok dan Ibu Charlota Upa’. Pendidikan Sarjana Hama dan Penyakit Tumbuhan diselesaikan di Institut Pertanian Bogor pada tahun 1996. Pendidikan Master of Science in Agriculture diselesaikan penulis di Georg-August University Goettingen, Germany pada tahun 2001. Pada tahun 2004 penulis mendapatkan kesempatan melanjutkan studi S3 di Program Studi Fitopatologi, Sekolah Pascasarjana, Institut Pertanian Bogor melalui program Beasiswa Program Pasca Sarjana (BPPS) Direktorat Jenderal Pendidikan Tinggi. Penulis juga mendapat kesempatan untuk melanjutkan penelitian dengan Sandwich-Like Program dari Direktorat Jenderal Pendidikan Tinggi di Laboratorium Agroentomology dan Laboratorium Molecular Phytopathology and Mycotoxin Research, Department of Crop Science, Georg-August University Goettingen, Germany pada Desember 2008-Juli 2009.
Halaman
DAFTAR ISI ……… xii
DAFTAR TABEL ... xiv
DAFTAR GAMBAR ... xv
DAFTAR LAMPIRAN ... xvii
BAB 1. PENDAHULUAN ……….. 1
BAB 2. TINJAUAN PUSTAKA ………... 9
BAB 3. ASOSIASI FAKTOR LINGKUNGAN DAN PRAKTEK BUDIDAYA DENGAN EPIDEMIK PENYAKIT BUSUK BUAH KAKAO: STUDI KASUS DI TEPI TAMAN NASIONAL LORE LINDU, SULAWESI TENGAH ... 21 Pendahuluan ..……….. 22
Bahan dan Metode ……….. 24
Hasil dan Pembahasan ………. 29
Kesimpulan ……… 39
Daftar Pustaka ……….. 39
BAB 4. KERAGAMAN CENDAWAN ENDOFIT BUAH KAKAO PADA TINGKAT KEKERINGAN YANG BERBEDA SERTA KORELASINYA DENGAN EPIDEMIK PENYAKIT BUSUK BUAH………. 45
Pendahuluan ………... 46
Bahan dan Metode ……….. 47
Hasil dan Pembahasan ………. 50
Kesimpulan ……… 65
Daftar Pustaka ……….. 65
BAB 5. POTENSI CENDAWAN ENDOFIT SEBAGAI PENGINDUKSI KETAHANAN TANAMAN TERHADAP PENYAKIT BUSUK BUAH PHYTOPHTHORA ... 69 Pendahuluan ……….. 70
Bahan dan Metode ……….. 71
Hasil dan Pembahasan ………... 75
Kesimpulan ………. 83
Keadaan Umum Lokasi Penelitian ……… 89
Keragaman Cendawan Endofit dan Pengelolaan Kesehatan Tanaman ……… 90
Kesimpulan ……… 97
Saran ………. 98
DAFTAR PUSTAKA ……… 99
Halaman
3.1 Pengelompokan praktek budidaya dan faktor lingkungan serta
status inang ke dalam kelas kategori …...…………... 26
3.2 Hubungan antara beberapa praktek budidaya dan faktor lingkungan dengan epidemik penyakit busuk buah berdasarkan
analisa Chi-kuadrat (χ2) ... 30
3.3 Nilai mass dan inersia faktor yang berasosiasi nyata dengan epidemik penyakit busuk buah berdasarkan analisis korespondensi sederhana ... 31
4.1 Hasil identifikasi cendawan endofit secara molekuler, nomor
aksesi dan persen kemiripan DNA pada Genbank... 51
4.2 Indeks keragaman tiap plot pada pengambilan awal dan akhir
beserta estimasi populasinya... 57
4.3 Suhu dan kelembaban rata-rata 1 m dan 3 m di atas permukaan
tanah selama pengambilan sampel (data Moser et al. 2010) ... 57
4.4 Indeks kemiripan jenis spesies antar plot ………. 60
4.5 Perubahan dominansi cendawan endofit pada tiap plot pada saat
pengambilan sampel yang berbeda ... 61
4.6 Korelasi frekuensi kemunculan spesies cendawan endofit
dengan kejadian penyakit busuk buah ... 64
5.1 Skor gejala penyakit pada buah yang diinokulasi dengan
Phytophthora palmivora ... 73 5.2 Bioesei cendawan endofit pada buah kakao dilanjutkan
inokulasi dengan patogen P. palmivora ... 75
5.3 Korelasi antara periode laten, laju infeksi, tingkat keefektifan dengan nilai AUDPC pada buah kakao dengan perlakuan cendawan endofit dan kontrol, zona penghambatan uji koloni ganda, dan produksi asam salisilat dan aktivitas peroksidase
Halaman
1.1 Alur Penelitian ... 8
3.1 Lokasi pengambilan sampel. (a) Pulau Sulawesi di tengah-tengah kepulauan Indo-Malay. (b) Hutan Lindung Lore Lindu di Sulawesi Tengah. (c) Lokasi 43 pertanaman (titik putih) di mana penelitian dilakukan, di tepi Hutan Lindung Lore Lindu di Palolo
dan Kulawi ... 25
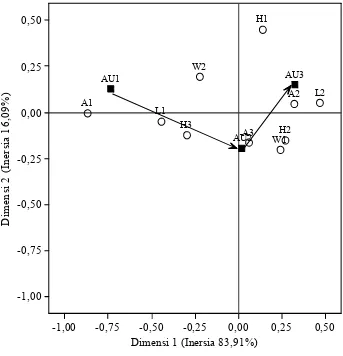
3.2 Pengelompokan kelas faktor yang berasosiasi dengan kelas AUDPC berdasarkan hasil analisis korespondensi sederhana. Segiempat hitam melambangkan kelas AUDPC dan lingkaran putih sebagai kelas faktor (AU1=AUDPC1, AU2=AUDPC2, AU3=AUDPC3, A=ketinggian tempat, H=sejarah penggunaan
lahan, L=praktek budidaya sebelumnya, W=penyiangan) ... 32
4.1 Pengelompokan waktu dan tempat pengambilan sampel ... 47
4.2 Frekuensi keberadaan tiap spesies cendawan endofit yang
berhasil diisolasi, mengikuti pola kurva distribusi normal ... 55
4.3 Pola peningkatan jumlah spesies yang diamati dan diharapkan
ketika diplotkan dengan pola peningkatan jumlah isolat ... 55
4.4 Curah hujan dan kandungan air tanah pada plot selama pengambilan sampel (dalam lingkaran, Maret-April 2007 dan
Januari-April 2008) (Moser et al. 2010) ... 58
4.5 Kemiripan jenis spesies antar plot dari indeks Sorensen dengan Multidimensional Scaling (MDS). Tiap titik mewakili ulangan dari tiap perlakuan (BC: Plot kontrol, pengambilan pertama; BR: Plot perlakuan, pengambilan pertama; AC: Plot kontrol,
pengambilan kedua; AR: Plot perlakuan, pengambilan kedua).... 60
4.6 Perubahan dominansi cendawan endofit pada plot dan waktu pengambilan sampel yang berbeda ( BC: Plot kontrol, pengambilan pertama; BR: Plot perlakuan, pengambilan pertama; AC: Plot kontrol, pengambilan kedua; AR: Plot
perlakuan, pengambilan kedua) ... 62
5.2 (A) produksi asam salisilat ( ppm) dan (B) aktivitas peroksidase (UAE) daun bibit kakao hari ke 9 dan 26 setelah inokulasi CE dan P. palmivora (UAE = Unit Aktivitas Enzim , Xy = Xylariaceae, Cg = Calocybe gambosa, Rf = Resinicium friabile,
TABEL
Halaman
3.1 Frekuensi kemunculan tiap faktor pada tiap kelas kategori
AUDPC ……… 107
4.1 Morfospesies cendawan endofit buah kakao dari Marena, Sulawesi Tengah beserta nama isolat sumber DNA untuk
identifikasi molekulernya ... 111
4.2 Untaian basa DNA penyandi 28S rDNA hasil sekuens isolat cendawan endofit yang diisolasi dari buah kakao di Marena,
Sulawesi Tengah dengan primer LROR dan LR6 ... 113
4.3 Kelas cendawan endofit hasil isolasi dari buah kakao ... 132
GAMBAR Halaman
3.1 Kondisi lahan sebelum ditanami kakao: (A) dengan membuka hutan primer, (B) mengganti tanaman tahunan yang telah ada,
(C) menanam kakao pada bekas pertanaman jagung ... 108
3.2 Fluktuasi suhu dan kelembaban udara harian rata-rata di Kulawi
dan Palolo, Sulawesi Tengah selama penelitian ... 109
3.3 Fluktuasi curah hujan dan radiasi harian rata-rata di di Kulawi
dan Palolo, Sulawesi Tengah selama penelitian ... 110
4.1 Pemasangan atap plastik pada plot di Marena, Sulawesi Tengah untuk memimik kekeringan (ENSO) (A: pemasangan panel plastic; B: plot control tanpa panel plastic; C: tepi plot ditutup
sedemikian rupa agar air tidak masuk ke dalam plot) ... 133
4.2 Fotomikroskopis beberapa cendawan endofit (kd: konidia; kf: konidiofor; mak: makrokonidia; mik: mikrokonidia; pik:
piknidia) ... 134
5.1 Uji patogenisitas patogen Phytophthora palmivora dan cendawan endofit (CE) pada daun pesemaian (A: tanaman utuh; B0: daun dengan agar; B1: Daun dengan agar + patogen; B2: Daun dengan agar + CE; C0: Agar dibuang, tanpa gejala; C1:
Kakao, tanaman penghasil bahan olahan untuk coklat, merupakan tanaman yang sangat penting bagi petani Indonesia. Sejak 1999 produksi kakao Indonesia menempati urutan ketiga di dunia setelah Pantai Gading dan Ghana (International Cocoa Organization, ICCO; www.icco.org/statistics/production.aspx). Prediksi produksi kakao Indonesia oleh ICCO akan mencapai 470 000 ton pada musim panen 2010/2011 dari total 4 195 000 ton produksi dunia atau sekitar 11.2% dari total produksi dunia. Pertanaman kakao Indonesia yang produksinya menempati posisi ketiga dunia tersebut, sebagian besar merupakan pertanaman rakyat. Pada tahun 2009, total pertanaman kakao di Indonesia adalah 1 592 983 hektar, hanya 3.88% yang dikelola oleh negara, 3.41% oleh perkebunan besar swasta, dan sisanya 92.7% dikelola oleh rakyat ( (Dirjen Bina Produksi Perkebunan dalam
Baon dan Wardani 2010). Kenyataan tersebut menunjukkan bahwa sangat banyak petani dan keluarganya yang menggantungkan hidupnya pada pertanaman kakao dengan luas lahan rata-rata 1 ha tiap petani.
Permasalahan utama kakao Indonesia ialah tingginya gangguan hama dan
penyakit, tanaman yang semakin tua, dan adanya automatic detention dari
pemerintah USA. Penggerek buah kakao (disebabkan oleh Conophomorpha
cramerella) dan busuk buah kakao (disebabkan oleh Phytophthora palmivora) merupakan hama dan penyakit utama pertanaman kakao Indonesia, di samping
Pemerintah Indonesia telah mencanangkan Gerakan Nasional Kakao (Gernas Kakao) yang dituangkan di dalam Peraturan Menteri Pertanian Nomor : 33/Permentan/OT.140/7/2006 untuk terus memacu peningkatan produksi dan mutu kakao Indonesia. Gerakan Nasional Kakao ini diaplikasikan misalnya berupa peremajaan tanaman tua dengan penyambungan menggunakan ranting tanaman yang produktivitasnya tinggi ataupun pemanfaatan hasil kultur jaringan somaklonal embriogenesis, melaksanakan intensifikasi pertanian pada lahan yang sudah ada dan rehabilitasi lahan yang sudah lama tidak terurus dengan baik.
Peningkatan produksi dengan cara penambahan areal pertanaman kakao banyak dilakukan petani dengan cara merambah areal hutan. Pembukaan areal hutan tentu berpengaruh terhadap ekosistem alami yang sebenarnya kaya dengan mikroba bermanfaat, terutama pada hutan tropika. Untuk melihat pengaruh lingkungan hutan yang sudah terganggu dan praktek budidaya di pertanaman kakao yang diaplikasikan petani terhadap epidemik penyakit busuk buah kakao yang disebabkan oleh Phytophthora palmivora, maka dilakukanlah serangkaian studi kasus, meliputi faktor-faktor yang mungkin mempercepat terjadinya epidemik penyakit busuk buah di areal pertanaman kakao sekitar Taman Nasional Lore Lindu yang merupakan hutan primer.
Penyakit busuk buah kakao yang disebabkan oleh Phytophthora palmivora
sebagai pengelola pertanaman akan mempengaruhi ketiga faktor tersebut. Metode yang disarankan dengan penggunaan varietas tahan, mengurangi kelembaban di dalam pertanaman dengan cara pemangkasan dan fitosanitari dengan cara mengubur buah dan kulit buah yang sakit. Sayangnya di tingkat petani belum tersedia varietas tahan dengan sifat agronomi dan mutu buah yang diinginkan. Demikian juga dengan praktek fitosanitari (seperti yang dianjurkan oleh pemerintah Indonesia pada Gerakan Nasional Kakao dan dipraktekkan oleh beberapa petani di Sulawesi Tengah), memerlukan waktu yang lama dan tempat yang luas untuk mengubur kulit buah kakao dan buah sakit karena kulit buah kakao sangat lambat terdekomposisi.
Saat ini, alternatif pengendalian yang menjanjikan dan sedang banyak diteliti adalah memanfaatkan hubungan antara mikroorganisme dengan tanaman sebagai inangnya. Pada umumnya tumbuhan dikolonisasi oleh beragam mikroorganisme, berada di atas permukaannya atau masuk ke dalam jaringan inang. Cendawan endofit, yaitu cendawan yang hidup di dalam jaringan inang tetapi tidak menimbulkan gejala penyakit pada inangnya, diduga merupakan mikroorganisme yang paling banyak sebagai pengkoloni inang (Schulz & Boyle 2005). Bahkan cendawan endofit di daerah tropis dikatakan sebagai hyperdiverse,
keragamannya sangat tinggi (Arnold et al. 2000). Cendawan endofit dicirikan dengan mengkolonisasi jaringan inang tanpa menimbulkan gejala penyakit (Wilson 1995). Selain pada Gramineae, cendawan ini juga ditemukan dalam keragaman dan populasi yang tinggi pada tumbuhan pohon/berkayu (Johnston
1998; Arnold et al. 2001; Gamboa et al. 2002). Cendawan endofit dapat
ditransmisikan secara vertikal dari biji ke bagian tanaman lainnya atau secara horizontal dari tanaman satu ke tanaman lainnya. Cendawan endofit dapat meningkatkan ketahanan inangnya terhadap lingkungan yang tidak mendukung dengan cara menginduksi ketahanan inang atau memproduksi metabolit sekunder yang menekan perkembangan patogen. Di lain pihak cendawan endofit mendapatkan makanan dari inang dan terlindung dari pencucian karena hujan atau angin karena berada di dalam jaringan inang.
Arnold et al. (2001) melaporkan bahwa tanaman kakao membawa
diduga dapat berperan sebagai penginduksi ketahanan tanaman terhadap penyakit (Arnold et al. 2003; Rubini et al. 2005).
Pemanfaatan cendawan endofit dapat menjadi alternatif pengendalian penyakit busuk buah kakao yang menjanjikan (Guest 2007). Arnold et al. (2003), Rubini et al. (2005), Mejia et al. (2008) berhasil mengisolasi cendawan ini dan menguji kemampuannya untuk menekan pertumbuhan dan perkembangan patogen secara in vitro dan in vivo. Berdasarkan pengujian tersebut, ditemukan beberapa cendawan endofit yang potensial sebagai agens hayati, di antaranya
Colletotrichum gloeosporioides dan Clonostachys rosea.
Sayangnya, penelitian tentang endofitik pada kakao masih terpusat di daerah Amerika Tengah, daerah di mana tanaman kakao diduga berasal (Sereno et al. 2006). Segala hipotesis tentang interaksi inang dengan cendawan endofit
khususnya kakao (Herre et al. 2005) hanya mewakili daerah tersebut, belum
mewakili kondisi di daerah tropis penanam kakao lainnya di dunia. Oleh karena itu, perlu dilakukan suatu studi yang bertujuan untuk mendapatkan gambaran tentang keragaman cendawan endofit di Indonesia sebagai salah satu produsen kakao tertinggi di dunia.
Studi ini diawali dengan menginventarisir faktor-faktor yang berhubungan dengan kejadian penyakit di lapangan, termasuk keragaman morfospesies cendawan endofitnya. Dari studi pertama ini akan diperoleh beberapa faktor lingkungan dan praktek budidaya yang berkaitan dengan tinggi atau rendahnya kejadian penyakit di lapangan. Studi selanjutnya dimaksudkan untuk mengeksplorasi mikroba bermanfaat yang ada di lapangan. Cendawan endofit dipilih sebagai objek penelitian berikutnya karena studi tentang mikroba ini tergolong masih jarang dilakukan. Keragaman dan kelimpahan cendawan endofit dianalisis, dikorelasikan dengan epidemik penyakit busuk buah di lapangan. Spesies cendawan endofit yang kelimpahannya berkorelasi negatif dengan kejadian penyakit, dalam arti kejadian penyakit rendah ketika kelimpahannya tinggi, diuji lebih lanjut untuk membuktikan kemampuannya dalam menekan keparahan penyakit pada buah kakao dan mekanisme yang berpotensi mendasarinya.
Urgensi (Keutamaan) Penelitian
Penelitian tentang cendawan endofit di Indonesia tergolong sebagai hal yang relatif baru. Studi yang dilakukan ini merupakan pengembangan tentang interaksi mikroba-tanaman terutama cendawan di dalam tanaman tahunan di daerah tropis. Walaupun aspek mengenai cendawan endofit sudah dikemukakan oleh de Barry pada tahun 1866 (dalam Dayle et al. 2001), namun penelitian yg intensif baru dimulai pada akhir tahun 1980-an ( Clay 1988). Penelitian ini diawali dengan adanya ternak yang keracunan setelah memakan rumput yang membawa cendawan endofit yang memproduksi alkaloid. Penelitian pada tumbuhan berkayu berkembang kemudian, khususnya di daerah subtropis. Belum banyak publikasi tentang cendawan endofit di daerah tropis, baru dimulai oleh Arnold dan kawan-kawan pada di awal tahun 2000-an. Mereka melakukan penelitian keragaman cendawan endofit pada daun tanaman kakao di Amerika Selatan.
Selain itu, penelitian ini juga merupakan terobosan teknologi dalam bidang pertanian. Penelitian ini memanfaatkan interaksi tanaman dengan mikroba khususnya pemanfaatan cendawan endofit dalam pengendalian penyakit tanaman.
Teknologi ini juga dapat menunjang peningkatan produksi dan mutu kakao yang sedang dicanangkan oleh pemerintah melalui Gerakan Nasional Kakao. Pemanfaatan somatik embriogenesis yang relatif bebas dari mikroba endofit dalam peremajaan kakao diduga lebih rentan terhadap serangan hama dan penyakit sehingga membutuhkan input mikroba bermanfaat dari luar. Adanya perhatian konsumen coklat terhadap masalah kerusakan lingkungan dengan penggunaan input pestisida sintetis menjadi nilai lebih pemanfaatan cendawan endofit dalam pengendalian penyakit kakao.
Tujuan Penelitian
• Menganalisis interaksi faktor lingkungan, praktek budidaya, inang,
keragaman morfospesies cendawan endofit dengan epidemik penyakit busuk buah kakao.
• Mengkaji keragaman cendawan endofit buah kakao pada kondisi
ekosistem yang relatif basah dan kering serta korelasinya dengan penyakit busuk buah kakao
• Menguji kemampuan beberapa cendawan endofit dalam menekan penyakit
busuk buah kakao dan mekanisme yang mendasarinya.
Dari penelitian ini diharapkan dapat diinventarisir faktor-faktor yang mendukung perkembangan penyakit busuk buah pada buah kakao, didapatkan isolat cendawan endofit yang berpotensi sebagai agensia hayati dalam mengendalikan busuk buah kakao. Juga diharapkan didapatkan informasi dasar tentang cara kerja (mode of action) cendawan endofit dalam mengendalikan busuk buah kakao.
Hypotesis
1. Semakin kecil intervensi pada agroekosistem kakao, maka keseimbangan
ekologi alami akan lebih baik, sehingga kejadian penyakit busuk buah pada kakao semakin rendah
2. Kejadian penyakit busuk buah akan rendah pada pertanaman yang dekat
ke hutan primer, frekuensi penyiangannya lebih jarang, keragaman cendawan endofitnya tinggi, berada pada ketinggian yang lebih rendah, tanaman yang berumur tua, persentase naungan dari tanaman pelindung rendah, sebelumnya adalah areal hutan, diameter tajuk rendah.
3. Penggunaan pestisida kimia pada musim sebelumnya dan penggunaan
pupuk N akan menyebabkan tingginya epidemik penyakit busuk buah kakao.
5. Ada korelasi antara kelimpahan cendawan endofit dengan kejadian penyakit busuk buah Phytophthora.
6. Beberapa cendawan endofit berperan sebagai penginduksi ketahanan
Gambar 1.1 Alur Penelitian
PENELITIAN 2
• Eksplorasi dan identifikasi cendawan endofit, penghitungan kejadian penyakit pada plot ENSO
PENELITIAN 3
Potensi cendawan endofit sebagai penginduksi ketahanan tanaman
• Penghitungan kejadian penyakit
• Pencatatan faktor lingkungan, praktek budidaya, penyiangan dan pemupukan
• Isolasi dan identifikasi morfospecies cendawan endofit
Keluaran:
• Hubungan epidemik penyakit dengan faktor lingkungan dan praktek
budidaya, umur inang serta keragaman morfospesies cendawan endofit
• Isolasi cendawan endofit
• Identifikasi secara morfologi dan molekuler
• Penghitungan keragaman cendawan endofit
• Penghitungan kejadian penyakit
Keluaran:
• Identifikasi spesies yang lebih akurat
• Hasil sekuen LSU rDNA isolat
• Keragaman cendawan endofit pada musim hujan dan kemarau
•Koleksi cendawan endofit
• Korelasi kejadian penyakit dengan kelimpahan cendawan endofit
• Isolasi patogen dan uji patogenisitasnya
• Uji patogenesitas cendawan endofit
• Uji kemampuan sebagai endofit
• Evaluasi aktivitas cendawan endofit terhadap patogen in vitro dan in vivo
• Evaluasi kemampuan endofit menginduksi ketahanan tanaman
Keluaran:
Diperoleh cendawan endofit yang berpotensi sebagai agens biokontrol PENELITIAN 1
Identifikasi faktor-faktor pendukung berkembangnya penyakit di lapangan
• Rekomendasi pengelolaan pertanaman yang sehat, berkelanjutan dan mendukung konservasi cendawan endofit
Busuk Buah Kakao oleh Phytophthora palmivora
Busuk buah kakao yang disebabkan oleh Phytophthora palmivora merupakan penyakit paling penting di pertanaman kakao di seluruh dunia (Guest 2007). Penyakit ini menyebabkan hilangnya produksi dunia 20% hingga 30%. Di daerah dengan curah hujan dan kelembaban yang tinggi kehilangan hasil dapat lebih tinggi.
Selain menyerang buah, P. palmivora juga menyerang daun, pucuk, akar, buah kecil, bunga dan batang (kanker batang). Kematian tanaman karena serangan patogen ini juga cukup tinggi di seluruh dunia, dapat mencapai 10%
dalam setahun (Guest 2007).
Tanaman inang P. palmivora bukan hanya kakao, tetapi juga lada, kelapa, dan karet. Ruzelfin (1989) menguji virulensi patogen yang berasal dari ketiga tanaman tersebut terhadap kakao dan menemukan bahwa isolat dari karet dan kelapa dapat menginfeksi/patogenik terhadap kakao, tetapi isolat dari lada tidak dapat menginfeksi kakao. Dia juga menemukan bahwa isolat dari karet lebih virulen bila dibandingkan dengan isolat dari kelapa.
Di Brazil, ada tiga spesies Phytophthora yang menyerang kakao, yaitu P. capsici, P. palmivora,dan P. citrophthora. Di Afrika, P. megakarya juga menyerang kakao. P. capsici kurang virulen bila dibandingkan dengan yang lainnya walaupun pada awalnya merupakan spesies yang paling dominan (populasi paling tinggi) di Bahia, Brazil. Namun pada saat ini, P. palmivora ditemukan paling dominan di pertanaman kakao di Brazil dengan mengisolasi dari tanah (Pereira 1992).
Cendawan endofit
sebagai asosiasi yang saling menguntungkan (simbiosis mutualisme). Cendawan endofit memiliki kespesifikan inang yang tinggi, simbiosis mutualisme: tidak ada kerusakan pada sel atau jaringan, terjadi siklus nutrisi atau bahan kimia antara endofit dan inangnya, meningkatkan daya bertahan hidup inang, meningkatkan kemampuan berfotosintesis inang, juga meningkatkan kemampuan bertahan hidup cendawan.
Penelitian tentang cendawan endofit awalnya dimulai pada rumput-rumputan di daerah subtropis-temperate. Asosiasi rumput dan endofit terutama didasarkan pada proteksi inang terhadap stress abiotik dan biotik tidak seperti simbiosis tumbuhan dan mikroba lainnya yang didasarkan pada akuisisi sumber mineral (nutrisi) (Clay dan Schardl 2002). Azevedo et al. (2000) mengungkapkan bahwa masih sangat kurang informasi tentang cendawan endofit dari daerah tropik.
Cendawan endofit diduga berasal dari cendawan-cendawan tanah, kemudian masuk ke dalam jaringan inang. Cendawan endofit pada rumput tumbuh dalam interseluler dan sistemik pada bagian tanaman di atas permukaan tanah (Clay dan Schardl 2002). Sebagian besar cendawan endofit ditransmisikan secara horizontal seperti pada kakao (Arnold 2003), sebagian lagi secara vertikal atau melalui biji seperti pada rumput-rumputan (Ernst et al. 2003). Dongyi et al.
Bukti secara molekuker memperlihatkan bahwa banyak endofit yang ditransmisikan lewat biji merupakan interspesific hybrids (Clay & Schardl 2002).
Agar dapat hidup selaras bersama-sama, genom inang-endofit diwariskan bersama-sama. Oleh karena itu simbion ini terkait secara langsung dengan perubahan perlahan-lahan (evolusi) populasi inangnya (Clay et al. 2005).
Sebagian besar endofit menghasilkan senyawa alkaloid yang berperan dalam pertahanan tanaman terhadap gangguan hama dan penyakit. Hibridisasi memungkinkan proliferasi dari gen yang mengontrol produksi alkaloid di antara aseksual endofit. Alkaloid ergot ergovaline, lolitrems, dan lolines diproduksi hanya oleh seksual Epichloe festucae, tetapi senyawa ini juga umum ditemukan pada endofit yang ditransmisikan lewat biji. Hal ini mengindikasikan bahwa gen untuk mensintesisnya berasal dari E. festucae. Aseksual hybrids mungkin juga didapat dari pemulihan akumulasi mutasi delesi (deleterious mutation) (Muller’s rachet) (Clay & Schardl 2002). Cendawan endofit juga berusaha untuk beradaptasi dengan lingkungannya dengan membuat keragaman genetiknya misalnya dengan hibridisasi. Sullivan dan Faeth (2004) menggunakan marker microsatelite untuk melihat laju gene flow di antara empat populasi Neotyphodium dari rumput Arizona fescue (Festuca arizonica). Keragaman haplotipe umumnya rendah; hanya satu populasi yang memiliki lebih dari dua haplotipe. Haplotipe
membawa banyak lokus di mana beberapa atau semua lokus mikrosatelit juga ditemukan, yang mengindikasikan bahwa hibridisasi vegetatif antara Neotyphodium dengan inang rumput menghambat Epichloe, sedangkan produksi alkaloid hanya dikontrol oleh seksual Epichloe. Gene flow di antara populasi Neotyphodium juga sangat rendah, lebih rendah daripada gene flow oleh pollen inang. Perbedaan kecepatan gene flow ini diprediksi menjadi penyebab ketidakcocokan (mismatching) antara endofit dengan inangnya. Hal ini juga diduga menjadi penyebab rendahnya atau malah tidak diproduksinya alkaloid pada rumput Arizona fescue dan rumput liar lainnya (Faeth et al. 2004).
aktivitas peroxidase tinggi pada benih terinfeksi endofit, sedangkan catalase dan superoxide dismutase tidak berbeda nyata pada kedua perlakuan.
Selain untuk kesehatan tanaman, cendawan endofit juga banyak diteliti untuk kesehatan manusia. Wiyakrutta et al. (2004) mengisolasi cendawan endofit dari berbagai tanaman obat di Thailand. Berdasarkan perbedaan morfologi, cendawan tersebut dipisah-pisahkan. Tiap isolat dibiakkan di dalam malt Czapek broth dan yeast extract sucrose broth, ekstraknya diambil untuk diuji aktifitas biologinya. Dari ekstrak tersebut, 92 isolat dapat menghambat Mycobacterium tuberculosis, ekstrak 6 isolat dapat menghambat Plasmodium palsivarum, 40 isolat dapat menghambat virus Herpes, 60 dapat menghamat pertumbuhan kanker mulut, 48 menghambat sel kanker payudara.
Pengaruh kehadiran cendawan endofit pada tingkat individu inang dan dalam komunitas
Rumput-rumputan (famili Poaceae) dan cendawan dari famili Clavicipitaceae telah diketahui hidup bersimbiosis saling menguntungkan (Schardl et al. 2004). Pada simbiosis mutualisme, cendawan Epichloe (bentuk aseksual: Neotyphodium spp.) ditransmisikan secara vertikal lewat biji inang (Pan & Clay 2002). Cendawan endofit akan mendapatkan keuntungan berupa tempat
hidup, nutrisi dan penyebaran melalui perbanyakan inang dan cendawan endofit dapat meningkatkan ketahanan tanaman terhadap hama, nematode akar, meningkatkan ketahanan terhadap kekeringan dan toleran terhadap nutrisi rendah.
Azevedo et al. (2000) dan Faeth & Fagan (2002) mengungkapkan bahwa ekspresi kehadiran cendawan endofit pada inang dipengaruhi oleh genetik endofit dan inangnya dan kondisi lingkungan saat itu. Faeth & Fagan (2002) membuat model produksi alkaloid oleh inang dengan perubahan konsentrasi nitrogen. Mereka menyimpulkan bahwa produksi alkaloid akan tinggi bila nitrogen tinggi dan sebaliknya akan rendah bila N rendah. Namun demikian, di alam, genotype inang dan faktor lingkungan lainnya seperti ketersediaan air juga berpengaruh.
nutrisi dan perlakuan pengairan. Dalam kondisi kering dan rendah nutrisi, pertumbuhan vegetatif dan produksi benih inang akan berkurang bila dibandingkan dengan inang tanpa cendawan endofit. Hal ini merupakan ‘harga yang harus dibayar’ dengan adanya cendawan endofit, tidak seperti dalam berbagai literatur yang mengatakan bahwa cendawan endofit selalu menguntungkan tanaman. Kedua inang yang diuji memiliki tingkat respon yang berbeda juga. Perbedaan itu mungkin disebabkan oleh perbedaan strategi bertahan hidup (life-history strategies) dan kebutuhan lingkungan bagi kedua spesies gramineae dan spesies cendawan yang mungkin berubah sepanjang sejarah hidup masing-masing tumbuhan tersebut.
Penelitian yang dilakukan oleh Lewis (2004) memperlihatkan bahwa pengaruh cendawan endofit Neotyphodium lolii terhadap tiga genotype Lolium perenne tidak konsisten terhadap pertumbuhan inang dalam kondisi stress biotik (infeksi ryegrass mosaic virus) dan stress abiotik (pH rendah, waktu interval pemanenan, tinggi pemanenan, peneduh, N rendah).
Pada interaksi endofit-inang dari rumput liar (native grass) didapatkan bahwa infeksi cendawan endofit Neothypodium pada rumput asli Arizona fescue menurunkan bobot biomas akar dan tajuk, menurunkan fitness inang dengan menurunnya competitive properties, paling tidak dalam jangka waktu pendek. Hal ini berbeda sangat nyata dengan hasil penelitian terhadap rumput introduksi yang telah dibudidayakan yang memperlihatkan sifat mutualistik yang tinggi dengan cendawan endofit (Faeth et al. 2004).
Cendawan sistemik Epichloe glyceriae yang menginfeksi rumput Glyceria striata akan menghambat produksi biji (Schardl et al. 2004). Penelitian yang dilakukan oleh Pan & Clay (2002) memperlihatkan bahwa rumput tersebut akan memproduksi stolon (clonal growth) lebih banyak dibandingkan dengan rumput yang tidak terinfeksi endofit. Cara ini mungkin merupakan mekanisme yang efektif bagi genotype inang tersebut untuk tetap bertahan hidup, yang mungkin berlaku bagi cendawan endofit lainnya yang menghambat pembentukan biji.
mutualist, maka keresistenan tersebut dapat dipatahkan sehingga pendatang dapat bertahan hidup di tempat baru atau dapat masuk ke dalam komunitas tersebut (Rutgers et al.2005).
Penelitian yang dilakukan oleh Lehtonen et al. (2005) membuktikan bahwa kekompleksan (complexity) terjadi pada interaksi tingkat komunitas, dan interaksi multispesies tidak dapat diprediksi dari pemasangan dua spesies saja. Mereka melakukan penelitian terhadap inang rumput (Lolium pretense) dengan cendawan endofitnya (Neotyphodium uncinatum), tumbuhan parasit akar (Rhinanthus serotinus), dan herbivore generalis (aphis Aulacorhum solani). Dari penelitian tersebut didapatkan bahwa parasit akar mengambil mikotoksin yang diproduksi oleh cendawan endofit dari inang rumput. Dengan mikotoksin tersebut, parasit akar dapat bertahan dari serangan herbivora aphid, sehingga parasit akar akan makin menekan inang rumput. Dengan demikian, kehadiran cendawan endofit bermanfaat (mutualist) berubah menjadi merugikan bagi inang dengan kehadiran tumbuhan parasit akar.
Sumber Inokulum Cendawan Endofit
Cendawan endofit lebih banyak ditemukan pada rumput liar yang tumbuh secara alami (10 dari 14 spesies yang diteliti), dibandingkan dengan rumput yang dibudidayakan (hanya pada 2 spesies dari 13 spesies (97 kultivar) yang diamati). Di alam, cendawan endofit memberikan kemampuan bagi inangnya untuk dapat bertahan hidup bila dibandingkan dengan yang tidak terinfeksi (Saikkonen et al. 2000).
Pada tanaman kakao (Theobroma cacao), adanya naungan dan umur daun mempengaruhi keragaman cendawan endofit yang diisolasi dari tempat tersebut, tetapi tidak dipengaruhi oleh letak geografis tanaman dan ada tidaknya infeksi patogen (Arnold & Herre 2003). Cendawan endofit juga tidak spesifik pada jaringan tertentu, isolat yang ditemukan di batang juga ditemukan di daun.
Banyak cendawan endofit yang membentuk struktur reproduksi hanya pada bagian tanaman yang sudah tua/mati. Contoh ekstrim akan hal ini misalnya endofit yang diisolasi dari conifer. Studi histology memperlihatkan bahwa endofit ini hanya terdiri dari beberapa hifa bersel satu hingga tanaman tua (Dix dan Webster 1995).
Ketahanan Tanaman terhadap Patogen
Mekanisme ketahanan tanaman terhadap patogen secara umum merupakan kombinasi dari dua sistim pertahanan yaitu ketahanan struktural dan ketahanan biokimia (Agrios 2005). Kedua ketahanan tersebut dapat berupa ketahanan yang bersifat pasif atau yang bersifat aktif sebagai hasil induksi dari faktor luar (Huang 2001). Ketahanan yang bersifat pasif merupakan ketahanan yang sudah terbentuk di dalam tanaman dan merupakan karakteristik yang normal dalam perkembangan tanaman tersebut, misalnya ketebalan kutikula, pembukaan stomata, jumlah
trikoma, dan adanya senyawa antimikroba (fitoantisipin) di dalam tanaman tersebut. Ketahanan yang bersifat aktif atau ketahanan terinduksi merupakan ketahanan yang terekspresi segera setelah adanya serangan mikroba patogen. Penginduksi ketahanan tanaman dapat berupa bahan kimia, adanya pelukaan pada tanaman, mikroba non patogenik, atau patogen itu sendiri. Hasil dari ketahanan ini juga berupa ketahanan struktural (misalnya fortifikasi atau penebalan dinding sel) dan biokimia (biosintesis fitoaleksin dan akumulasi PR protein) (Huang 200; van Etten et al. 1994 ).
Fitoaleksin didefenisikan sebagai metabolit sekunder yang disintesis dan terakumulasi di dalam tanaman setelah terekspos atau terpapar terhadap mikroorganisme (Paxton 1980). Fitoaleksin merupakan senyawa dengan berat molekul yang rendah (Hammerschmidt 1999). Mert-Tűrk (2002) mengemukakan bahwa bila fitoantisipin tidak berhasil menghambat perkembangan patogen yang menyerang tanaman, maka fitoaleksin akan disintesis dan melanjutkan kerja fitoantisipin dalam menghambat perkembangan patogen.
Ketahanan biokimia selanjutnya adalah akumulasi Pathogenesis Related protein (PR protein). PR protein didefenisikan sebagai protein yang disandikan oleh tanaman inang tetapi terinduksi hanya dalam kondisi sakit atau keadaan yang berhubungan dengan kondisi patologi (Antoniw et al. 1981). Sedikitnya telah diketahui 14 kelas PR protein dengan ciri-ciri protein dengan berat molekul rendah (6-43 kDa), dapat diekstraksi dan stabil dalam kondis pH rendah (< 3),
tahan panas, resisten terhadap protease (van Loon dan van Strein 1999). PR protein ini di antaranya adalah ekstraselular acidic protein, β-1,3-glukanase,
kitinase I-V, kitosanase, sweet tasting protein thaumatin, protease inhibitor, endoproteinase, peroxidase, protein intraselular mirip ribonuclease, tionin dan plant defensins, proteins pentransfer lipid, α- amylase (Huang 2001).
Biotik penginduksi PR protein yang telah dikenal adalah patogen,
serangga, nematoda, herbivora dan gulma Orobanche (tumbuhan parasit); bahan kimia penginduksi seperti asam salisilat, poliakrilik, asam lemak, garam inorganik; stimulasi fisik dapat juga berperan sebagai penginduksi seperti pelukaan, radiasi UV-B, tekanan osmotik tiba-tiba, suhu rendah, kekurangan atau kelebihan air (Edreva 2005).
Ketahanan tanaman yang didapatkan karena induksi seperti dijelaskan di atas disebut Systemic Acquired Resistance (SAR). Induksi ketahanan lainnya adalah Induced Systemic Resistance (ISR), yaitu ketahanan yang didapatkan dari induksi oleh mikroba non patogenik Plant Growth Promoting Rhizobacteria (PGPR) dan mikoriza dengan etilen dan asam jasmonat sebagai senyawa signal transduksinya dan tidak melibatkan gen PRs (Pieterse et al. 1996).
peroksidase sebagai respon ketahanan biokimia juga berperan dalam lignifikasi/penebalan dinding sel tanaman. Oleh karena itu, pemisahan antara ketahanan struktural dan ketahanan biokimia sering kabur dan saling tumpang tindih (Huang 2001).
Mikroba Endofit sebagai Penginduksi Ketahanan Tanaman
Penelitian terbaru tentang mekanisme kerja mikroba endofit dalam meningkatkan ketahanan inang terus berkembang. Dari berbagai penelitian tersebut diketahui bahwa mekanisme utama endofit dalam meningkatkan ketahanan inang berupa induksi ketahanan, dengan cara mengaktifkan gen-gen ketahanan inang yang ada tetapi dalam kondisi inaktif.
Mekanisme induksi ketahanan tanaman yang telah diketahui secara umum adalah dengan cara Induced Systemic Resistance (ISR) atau Systemic Acquired Resistance (SAR). ISR merupakan induksi ketahanan sistemik pada tanaman oleh mikroba tanah bermanfaat dan tidak patogenik seperti plant growth promoting rhizobacteria dan mikoriza, melibatkan jasmonic acid dan etilen sebagai senyawa penginduksi, serta tidak melibatkan ekspresi gen PR (Pathogenesis Related genes) tetapi gen lain yang belum diketahui. Sedangkan SAR merupakan induksi ketahanan sistemik yang dipicu oleh adanya infeksi patogen, adanya
hipersensitive reaction, atau aplikasi bahan kimia, melibatkan senyawa asam salisilat, dan melibatkan ekspresi gen PR (Pieterse et al. 1996).
Hasil penelitian induksi ketahanan oleh cendawan endofit diketahui ternyata melibatkan senyawa asam salisilat, asam jasmonat dan peroksidase (Shirasu et al. 1997; Segarra et al. 2007). Bila melihat defenisi ISR dan SAR (Pieterse et al. 1996), maka mekanisme induksi ketahanan oleh cendawan endofit melibatkan keduanya. Namun karena cendawan endofit tidak patogenik, maka Gao et al. (2010) mengusulkan bahwa mekanisme cendawan endofit adalah ISR tetapi dengan melibatkan gen PR.
tomat dan maculicola dengan 4 isolat PGPR, 2 isolat PGPR tidak membutuhkan asam jasmonat atau etilen sebagai molekul signalnya (Ryu et al. 2003). Penelitian Zhang et al. (2002) mendapatkan bahwa bakteri PGPR Serratia marcescens strain 90-166 yang berperan dalam menekan patogen kapang abu (blue mold) Peronospora tabacina pada tembakau juga mampu menghasilkan asam salisilat dalam media tryptic soy broth (TSB) dan di dalam pesemaian pada minggu pertama setelah inokulasi PGPR.
Cendawan endofit Trichoderma viride diketahui ternyata mampu menekan perkembangan patogen Diplodia corticola, penyebab kanker, nekrosis pada pembuluh dan mati pucuk pada tanaman oak (Campanile et al 2007). Mendoza dan Sikora (2009) mengkombinasikan cendawan endofit Fusarium oxysporum strain 162 dengan bakteri antagonis Bacillus firmus untuk mengendalikan patogen nematode Radopolus similis pada tanaman pisang dan berhasil menurunkan populasi patogen hingga 86.2%, lebih tinggi bila dibandingkan dengan aplikasi tunggal yang hanya sebesar 63.7% bila bakteri sendiri dan 27.8% bila cendawan sendiri. Narisawa et al (2002) memanfaatkan cendawan endofit Heteroconium chaetospira, P. fortinii, dan unidentified species of Fusarium, Penicillium, Trichoderma dan Mycelium radicis atrovirens (MRA) yang diisolasi dari akar terong, stoberi dan kubis, ternyata mampu mengendalikan penyakit layu pada
terong yang disebabkan oleh Verticillium dahliae hampir 100%.
Metode Cepat untuk Evaluasi Ketahanan Kakao
DENGAN EPIDEMIK PENYAKIT BUSUK BUAH KAKAO: STUDI KASUS DI TEPI TAMAN NASIONAL LORE LINDU, SULAWESI
TENGAH
(ENVIRONMENTAL FACTORS AND MANAGEMENT PRACTICES RELATED TO THE EPIDEMICS OF BLACK POD DISEASE OF CACAO
IN SULAWESI) Abstract
Some factors affecting epidemics of black pod disease (BPD) of cacao were investigated in 86 plots in two sub-districts adjacent to Lore Lindu National Park, Sulawesi, Indonesia. The factors investigated included environmental factors (altitude, distance to pristine forest, preceding cropping practices, land use history, the diversity of endophytic fungi morphospecies), management practices (weeding frequency, degree of shading, fertilizer application, canopy cover), and the age of host trees. Simple correspondence analyses were performed to examine the association between the BPD epidemics and the assessed parameters. Weeding frequency, altitude, preceding cropping practices, and land use history before establishment of the cacao plantation were significantly associated with the BPD epidemics of cacao, while application of nitrogen fertilizer, distance to pristine forests, plant age, canopy cover, diversity of endophytic fungi and degree of shading by 40 shade trees were not associated with disease development. BPD incidence was highest in plots that: were located at > 600 m a.s.l. (above sea level); were previously occupied by pristine forest or perennial plants; had prior intensive inputs of pesticides and fertilizers; and had a high frequency of weeding. Key words: pristine forest, Phytophthora palmivora, correspondence analysis, weeding frequency, shade trees
Abstrak
tertinggi ditemukan pada plot yang terletak >600 m dpl; areal yang sebelum ditanami kakao merupakan hutan primer atau tanaman tahunan; sebelumnya mendapat input pestisida dan pupuk yang intensif, dan dengan frekuensi penyiangan yang tinggi.
Kata kunci: hutan primer, Phytophthora palmivora, analisis korespondensi sederhana, frekuensi penyiangan, tanaman pelindung
Pendahuluan
Tanaman kakao (Theobroma cacao L.) diduga berasal dari hutan hujan tropis di Brasil dan Meksiko (Orwa et al. 2009). Produk tanaman berupa biji yang manfaat utamanya sebagai bahan baku untuk pembuatan bubuk kakao dan coklat. Sejak tahun 1999, sekitar 70% dari pasokan kakao dunia diproduksi oleh Pantai Gading, Ghana, dan Indonesia (Gray 2001; www.fao.org; www.icco.org/statis-tics/production.aspx). Pada negara produsen kakao dunia, sebagian besar kakao ditanam oleh petani kecil dan menjadi mata pencaharian utama mereka (Gray, 2001). Demikian juga halnya di Indonesia, pada tahun 2009, 92.7% dari luas total perkebunan kakao di Indonesia dimiliki oleh petani kecil (Baon & Wardani, 2010).
Kendala utama dalam budidaya kakao di seluruh dunia adalah penyakit busuk buah kakao, yang dapat mengurangi produksi hingga 20-30%. Dalam
kasus-kasus tertentu, kehilangan hasil bahkan dapat mencapai 90% (Bowers et al. 2001; Guest, 2007).
ITS rDNA 22 isolat P. palmivora yang terdiri dari dua ras asal kakao dan satu asal kelapa. Epidemik penyakit busuk buah mungkin berbeda bergantung kepada spesies dan ras Phytophthora, namun hal ini masih perlu penelitian lebih lanjut.
Seperti penyakit tanaman lainnya, penyakit busuk buah dipengaruhi oleh berbagai faktor abiotik dan biotik (Mora-Aguilera dan Campbell 1997) termasuk kondisi lingkungan, praktek budidaya, dan status tanaman inangnya (Agrios 2005; Avelino et al. 2006; Hardwick 1998).
Sehubungan dengan tanaman inang, tipe kakao yang paling banyak ditanam di Asia adalah jenis Trinitario, yang merupakan hibrida antara jenis Criollo dan Forastero (www.worldcacao-foundation.org; Motamayor et al. 2003). Criollo dianggap rentan terhadap P. palmivora, sedangkan Trinitario dianggap lebih tahan (Motamayor et al. 2003). Mawardi (1982) melaporkan bahwa jenis Trinitario adalah kultivar yang paling banyak ditanam di Indonesia, tetapi beberapa kultivar lainnya telah diperkenalkan di daerah penanam kakao sejak tahun 1974 (Butarbutar, 1977 dalam Tjasadihardja, 1987), termasuk Amelonado (West African Amelonado, WAA) dan kelompok Amazon (Upper Amazon Hybrid, UAH) (Angkapradipta et al. 1988; Wardojo, 1991). Kedua kultivar kakao ini merupakan jenis Forastero (Motamayor et al. 2003). Sebenarnya, kultivar kakao yang pertama diekspor dari Indonesia adalah jenis Criollo yang ditanam di
Sulawesi dan mungkin diperkenalkan di kepulauan ini sekitar 1750-1778 (Toxopeus dan Giesberger, 1983) atau bahkan pada tahun 1560 dibawa oleh Spanyol dari Filipina (Mawardi, 1982). Pada awal 1900-an, jenis Criollo secara bertahap digantikan oleh jenis Trinitario karena Trinitario lebih tahan terhadap serangan Helopeltis dan penggerek buah kakao (Toxopeus dan Giesberger, 1983). Hibrida UAH memiliki fase produktif yang dimulai lebih awal (hanya 2 tahun dibandingkan dengan 4 tahun untuk Trinitario) dan produktivitas lebih tinggi, menyebabkan peneliti dan petani mulai meliriknya (Angkapradipta et al. 1988).
relatif tinggi (27.5°-30.0°C) untuk pertumbuhan sporangia serta >20°C untuk pembentukan oogonium (Brasier, 1969).
Faktor biotik yang dapat mempengaruhi terjadinya penyakit busuk buah adalah keberadaan mikroorganisme lain dan vegetasi yang tumbuh di pertanaman (Krauss dan Soberanis 2001). Mejia et al. (2008) melaporkan bahwa cendawan endofit mampu menekan perkembangan penyakit busuk buah. Selain itu, praktek budidaya juga mempengaruhi terjadinya epidemik berbagai penyakit tanaman. Praktek budidaya dalam hal ini mencakup aplikasi pupuk, aplikasi pestisida, sanitasi, dan pengelolaan naungan (Agrios 2005; Avelino et al. 2006).
Di Indonesia, pengaruh faktor lingkungan, praktek budidaya serta status tanaman inang terhadap epidemik penyakit busuk buah kakao belum diteliti secara komprehensif. Oleh karena itu, perlu dilakukan studi yang bertujuan untuk menganalisis faktor lingkungan, praktek pengelolaan tanaman dan status inang yang terkait dengan epidemi penyakit busuk buah kakao. Hipotesis yang diajukan bahwa semakin kecil gangguan pada lingkungan alami pertanaman kakao maka keseimbangan ekologi relatif tetap seimbang seperti keseimbangan biologi pada ekosistem alami sehingga kejadian penyakit busuk buah pada kakao semakin rendah. Informasi ini diperlukan untuk pengembangan dan penerapan tindakan pengendalian penyakit busuk buah yang efektif sambil meminimalkan kerusakan
terhadap lingkungan.
Bahan dan Metode Tempat dan Waktu
Penelitian dilakukan di pertanaman kakao, Lembah Kulawi (400-925 m dpl) dan Lembah Palolo (525-950 m dpl), Sulawesi Tengah, Indonesia, sejak November 2007 hingga Agustus 2008.
Gambar 3.1 Lokasi pengambilan sampel. (a) Pulau Sulawesi di tengah-tengah kepulauan Indo-Malay. (b) Taman Nasional Lore Lindu di Sulawesi Tengah. (c) Lokasi 43 pertanaman (titik putih) di mana penelitian dilakukan, di tepi Taman Nasional Lore Lindu di Palolo dan Kulawi.
Pengamatan dilakukan di 22 pertanaman di Kulawi dan 21 di Palolo. Ke 43 pertanaman ini mewakili berbagai kondisi lingkungan, sejarah pertanaman, dan praktek budidaya yang diaplikasikan petani. Setiap pertanaman kemudian dibagi
tanaman kakao, sejarah penggunaan lahan sebelumnya, persentase penutupan oleh tajuk kakao, keragaman morfospesies cendawan endofit pada buah kakao, dan tingkat naungan dari tanaman peneduh yang tumbuh di dalam pertanaman kakao (Tabel 3.1).
Pengaruh Perlakuan
Penyiangan. Plot disiangi setiap 6 bulan di 22 plot di Kulawi dan di 22 plot di Palolo, dan setiap 2 bulan di 22 plot di Kulawi dan di 20 plot di Palolo. Penyiangan dilakukan secara manual dimulai pada Desember 2006 sampai Desember 2008. Tidak ada herbisida yang digunakan dalam penelitian ini.
Pemupukan dengan N. Kandungan nitrogen tanah di lokasi penelitian merupakan faktor pembatas dalam produksi kakao (Dechert et al. 2005), oleh karena itu tiap
Tabel 3.1 Pengelompokan praktek budidaya dan faktor lingkungan serta status inang ke dalam kelas kategori
Faktor Kode Satuan Kelas kategori
Frekuensi penyiangan W1 Bulan Tiap dua bulan
W2 Tiap enam bulan
Aplikasi pupuk F1 - Tanpa pupuk
F2 Aplikasi pupuk N
Praktek budidaya sebelumnya L1 Kec. Tidak intensif, Kulawi
L2 Intensif, Palolo
Sejarah penggunaan lahan H1 Hutan primer
H2 Tanaman tahunan
Keragaman cendawan endofit E1 Morfo- 4-14
E2 species 15-30
E3 31-47
Naungan dari pohon pelindung S1 % x ≤ 20% S2 20 < x ≤ 40%
pertanaman kakao dibagi menjadi dua plot, satu plot (20x40 m/plot dan sekitar 60pohon/plot) diberi perlakuan pupuk (penambahan pupuk N) dan plot lainnya tanpa perlakuan pupuk. Pupuk Urea diaplikasikan ke salah satu plot pada bulan Desember 2006, Juli 2007, Desember 2007, dan Juli 2008 dengan 217 g Urea (kira-kira setara dengan 100 g N2O5) per pohon per aplikasi. Pupuk untuk setiap pohon dibagi menjadi 10 bagian yang dimasukkan ke dalam 10 lubang melingkar di bawah garis tajuk dari pohon. Lubang-lubang tersebut kemudian ditutup dengan tanah. Pada plot tanpa perlakuan pupuk, lubang yang sama digali dan ditutup tetapi tanpa pupuk.
Pengumpulan Data Primer dan Sekunder
Data dikumpulkan dengan pengamatan langsung di lapangan dan dengan mewawancarai petani.
Pengamatan kejadian penyakit Kejadian penyakit dihitung tiap bulan dengan pengamatan langsung pada 86 plot sejak November 2007 hingga Agustus 2008. Kejadian penyakit dihitung sebagai persentase pohon dengan buah terinfeksi P. palmivora dibagi dengan jumlah total pohon kakao yang berbuah. Jumlah buah per pohon terinfeksi tidak digunakan dalam analisis karena patogen menyebar lebih cepat secara vertikal (dari buah terinfeksi ke buah yang sehat dalam satu pohon) daripada secara horisontal (Muller 1974). Jadi, penyebaran patogen secara
horisontal dari satu pohon ke pohon lainnya menyediakan data yang lebih dapat menggambarkan epidemik penyakit busuk buah. Buah matang yang sehat maupun sakit dipanen setiap 2 minggu sekali, kemudian semua buah sakit ditimbun untuk mengurangi salah satu sumber inokulum patogen (Ndoumbe-Nkeng et al. 2004; Soberanis et al. 1999).
Ketinggian tempat.
Persentase penutupan tajuk kakao. Persentase penutupan oleh tajuk kakao terhadap areal pertanaman ditentukan dengan estimasi dari foto-foto yang diambil 5.8 m di atas permukaan tanah pada tahun 2007.
Suhu udara. Suhu udara di dalam pertanaman dicatat dengan menggunakan i-button data loggers (Thermochron DS1921G-F5#, Maxim/Dallas Semiconductor, Sunnyvale, CA, USA). Sebagian besar pertanaman di Palolo dan Kulawi dipantau dari November 2007 hingga April 2008, tetapi satu pertanaman di Palolo dan satu di Kulawi dipantau dari November 2007 hingga Agustus 2008.
Umur dan riwayat penggunaan lahan. Data tentang umur tanaman kakao dan riwayat penggunaan lahan diperoleh dari petani pemilik pertanaman.
Praktek pengelolaan sebelumnya. Dari wawancara tidak terstruktur dengan petani dan pengamatan langsung di lapangan diketahui bahwa pertanaman di Kulawi lebih sedikit bahkan tanpa pupuk atau pestisida sintetis, sedangkan pertanaman di Palolo diberi pupuk dan pestisida sintetis. Dalam penelitian ini, pengelolaan intensif yang dimaksudkan dibatasi pada penggunaan pupuk dan pestisida sintetis. Isolasi cendawan endofit. Cendawan endofit diisolasi dari buah kakao, dari 4 pohon tiap pertanaman, 2 buah per pohon, 3 bagian (basal, pertengahan, apikal) per buah, dan dari 5 titik pada kulit buah kakao per bagian. Setelah sterilisasi permukaan berseri dengan ethanol 70%, natrium hipoklorit 2% selama 1-2 menit,
etanol 95%, lalu ke aquades steril dua kali, 5 mm2 dari 5 titik kulit kakao diinkubasi dalam Cawan Petri diam cm 9 yang berisi MEA 10% (Malt Extract Agar). Miselia yang tumbuh dari kulit kakao diisolasi, dimurnikan, dan diidentifikasi morfospesiesnya berdasarkan 10 karakteristik morfologi dari Arnold et al. (2001).
Analisis data
Untuk menggambarkan besarnya kejadian penyakit selama periode pengamatan, dihitung nilai AUDPC (area under disease progress curve) berdasarkan kejadian penyakit menurut formula Madden et al. (2007):
n-1
AUDPC =
∑
(yi + yi+1) (ti+1 – ti)i=1 2
di mana yi = kejadian penyakit, didapatkan pada waktu ti,
yi = ni x 100% N
ni = pohon dengan buah yang terinfeksi patogen N = total pohon berbuah
yi+1 = kejadian penyakit, didapatkan pada waktu ti+1
t = waktu, 1 hingga 10
Nilai AUDPC kemudian dikelompokkan ke dalam tiga kelas: rendah (x ≤ 175), sedang (175 < x ≤ 350), dan tinggi (x > 350), nilai faktor-faktor juga dikelompokkan ke dalam kelas-kelas (Tabel 3.1). Data disusun dalam matriks dengan kelas AUDPC sebagai kolom dan faktor-faktor sebagai baris. Hubungan antara AUDPC dengan masing-masing faktor diuji dengan uji chi-kuadrat (χ2).
Selanjutnya tabel kontingensi antara AUDPC dan faktor-faktor dengan nilai P≤0.05 pada uji chi-kuadrat (χ2) dianalisis menggunakan analisis korespondensi sederhana (Simple Correspondence Analysis, SCA; Savary et al.1995). Analisis korespondensi adalah metode yang sederhana dan terpercaya untuk mensintesis informasi yang terkandung dalam satu atau beberapa tabel kontingensi, mengingat bahwa setiap korelasi antara variabel dengan faktor-faktor tidak selalu linier, tetapi dengan analisis korespondensi, diintegrasikan ke dalam satu grafik (Savary et al. 1995). Analisis dengan SCA menghasilkan grafik dua dimensi sebagai hasil dari AUDPC yang dibagi dalam 3 kelas kategori. Letak dari setiap faktor pada tiap koordinat x dan y ditentukan dari penghitungan frekuensi tiap faktor dalam tabel kontingensi. Pengolahan data dilakukan dengan menggunakan MINITAB 15 (www.Minitab.com).
Hasil dan Pembahasan
Berdasarkan analisis Chi-kuadrat, faktor yang berasosiasi nyata dengan
Tabel 3.2 Hubungan antara beberapa praktek budidaya dan faktor lingkungan dengan epidemik penyakit busuk buah berdasarkan uji Chi-kuadrat (χ2)
Faktor Uji χ
2 Nilai χ2 P
Frekuensi penyiangan 7.986 0.018*
Aplikasi pupuk 2.059 0.357
Praktek budidaya sebelumnya 18.089 0.000**
Ketinggian tempat 20.813 0.000**
Jarak ke hutan primer 7.463 0.113
Umur tanaman kakao 2.869 0.238
Sejarah penggunaan lahan 11.104 0.025*
Persentase penutupan tajuk kakao 1.356 0.852
Keragaman cendawan endofit 1.607 0.808
Naungan dari pohon pelindung 2.720 0.606
Keterangan : * = berasosiasi nyata (P < 0.05) berdasarkan uji χ2 ** = berasosiasi sangat nyata (P < 0.01) berdasarkan uji χ2 tanpa asteriks = tidak berasosiasi berdasarkan uji χ2
morfospesies cendawan endofit, dan tingkat naungan yang dihasilkan oleh pohon peneduh (P>0.05).
Pada analisis selanjutnya, hanya variabel-variabel yang berasosiasi nyata dengan kejadian penyakit yang dianalisis. Dengan SCA, didapatkan total inersia sebesar 0.1686 (Tabel 3.3). Inersia untuk dimensi 1 sebesar 0.1414 (83.91%) dan 0.0271 (16.09%) untuk dimensi 2 (Gambar 3.2). Inersia (Inertia) adalah (total) nilai Chi-kuadrat (χ2) dibagi (total) jumlah frekuensi, menggambarkan tingkat
Tabel 3.3 Nilai mass dan inersia faktor yang berasosiasi nyata dengan epidemik penyakit busuk buah berdasarkan analisis korespondensi sederhana
Faktor Kode Mass Inersia
Frekuensi penyiangan W1 0.061 0.058
W2 0.064 0.055
Praktek budidaya sebelumnya L1 0.064 0.124 L2 0.061 0.130
Ketinggian tempat A1 0.029 0.212
A2 0.076 0.076 A3 0.020 0.006 Sejarah penggunaan lahan H1 0.029 0.062 H2 0.044 0.041 H3 0.052 0.054
Total 0.1686
Keterangan: mass = frekuensi relatif kemunculan tiap kelas kategori Inersia = (total) nilai χ2 dibagi dengan (total) frekuensi
Epidemik penyakit busuk buah berkembang lebih lambat pada kondisi kebun dengan penyiangan yang kurang intensif (tiap 6 bulan) dibandingkan dengan penyiangan yang lebih intensif (tiap 2 bulan). Praktek budidaya sebelumnya secara nyata berasosiasi dengan kejadian penyakit busuk buah di pertanaman kakao. Pada pertanaman kakao di Palolo (L2, intensif) kejadian penyakit lebih tinggi daripada di Kulawi (L1, non-intensif, Gambar 3.2).
Kejadian penyakit busuk buah berasosiasi nyata dengan ketinggian tempat (Gambar 3.2). Nilai AUDPC yang rendah (AUDPC1) berasosiasi dengan lokasi pertanaman di dataran rendah (<600 m dpl), nilai AUDPC sedang (AUDPC2) berasosiasi dengan ketinggian >800 m dpl, dan nilai AUDPC yang tertinggi (AUDPC3) berasosiasi dengan ketinggian antara 600 dan 800 m dpl. Hal ini menunjukkan bahwa kondisi yang cocok untuk perkembangan penyakit busuk buah yang optimal berada pada ketinggian 600-800 m dpl. Suhu rata-rata pada ketinggian <600 m dpl sebesar 24oC (maksimum 31.0oC, minimum 20.4°C), di pertanaman antara 600 sampai 800 m dpl ( di mana kejadian penyakit busuk buah
0,50
Gambar 3.2 Pengelompokan kelas faktor yang berasosiasi dengan kelas AUDPC berdasarkan hasil analisis korespondensi sederhana. Segiempat hitam melambangkan kelas AUDPC dan lingkaran putih sebagai kelas faktor (AU1=AUDPC1, AU2=AUDPC2, AU3=AUDPC3, A=ketinggian tempat, H=sejarah penggunaan lahan, L=praktek budidaya sebelumnya, W=penyiangan)
Epidemik penyakit busuk buah juga berasosiasi nyata dengan sejarah lahan sebelum ditanami kakao. Nilai AUDPC sedang sampai tinggi berasosiasi dengan plot yang sebelumnya adalah hutan primer dan tanaman tahunan (biasanya kopi,
Beberapa faktor tidak terkait dengan epidemi penyakit busuk buah, ditunjukkan oleh rendahnya nilai χ2 dan nilai P yang tinggi (Tabel 3.2). Jarak pertanaman ke hutan primer (0-2400 m) dan aplikasi pupuk N tidak berasosiasi dengan kejadian penyakit busuk buah. Demikian juga umur tanaman kakao, yang berkisar 8 sampai 22 tahun, tidak berasosiasi dengan kejadian penyakit busuk buah. Keanekaragaman morfospesies cendawan endofit yang diisolasi dari buah kakao sehat (4-47 morfospesies), juga tidak berasosiasi dengan kejadian penyakit busuk buah. Tingkat naungan, yang berkisar dari 0 hingga 60%, dan persentase penutupan areal oleh tajuk kakao (yang berkisar 23-81%) juga tidak berasosiasi dengan kejadian penyakit busuk buah.
Penyebab penyakit busuk buah di Indonesia, P. palmivora, mampu bertahan dan berkembang di dalam tanah, yang menjadi sumber inokulum terus-menerus bagi pertanaman kakao (Ristaino dan Gumpertz, 2000). Biasanya, patogen ditularkan secara pasif dari tanah ke daun, batang, buah, atau bagian tanaman lainnya di atas tanah dengan percikan air atau oleh arthropoda, seperti semut (McGregor dan Moxon 1985). Penyebaran patogen secara horizontal dari pohon sakit ke pohon sehat dapat disebabkan oleh percikan air hujan atau oleh aktivitas manusia dan avertebrata; patogen dapat menyebar di dalam tanah, pada bagian tanaman, atau sebagai propagul patogen yang bebas (Ristaino dan Gumpertz
Ketahanan tanaman kakao terhadap penyakit busuk buah bergantung kepada ketersediaan nutrisi yang cukup. Penghentian aplikasi N sendiri tidak mempengaruhi epidemi penyakit busuk buah seperti hasil dalam penelitian ini, namun penghentian pemupukan P dan K dapat menurunkan resistensi (Huber dan Haneklaus 2007). Hal ini diduga menyebabkan tingginya kejadian penyakit di daerah Palolo yang petaninya sudah terbiasa menggunakan pupuk kimia dibandingkan dengan petani Kulawi.
Pestisida utama yang digunakan di Palolo adalah fungisida mancozeb dan herbisida glifosat. Beberapa peneliti telah melaporkan bahwa aplikasi jangka panjang dari kedua pestisida tersebut memiliki efek merugikan, baik langsung atau tidak langsung, pada populasi mikroba tanah dan tanaman di ekosistem pertanian (Bradley et al. 2002;. Morjan et al. 2002;. Fernandez et al. 2009.; Rasool dan Reshi 2010; Dick et al. 2010). Meskipun dengan data terbatas, diduga bahwa aplikasi pestisida terus menerus di masa lalu telah mengurangi mikroorganisme tanah yang bermanfaat menghambat P. palmivora dan mungkin telah berkontribusi pada tingginya kejadian penyakit di wilayah Palolo (van Dipeningen 2006).
Epidemi penyakit busuk buah di pertanaman kakao lebih tinggi pada lahan yang sebelumnya adalah hutan primer atau yang sebelumnya ditanami tanaman