PRODUKSI BAKTERIOSIN ASAL Lactobacillus plantarum
SEBAGAI ANTIMIKROB DAN PENGUJIAN
KETAHANANNYA TERHADAP PANAS
SKRIPSI
TRI SANTI MARSELLY
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
RINGKASAN
TRI SANTI MARSELLY. D14070060. 2012. Produksi Bakteriosin Asal
Lactobacillus plantarum sebagai Antimikrob dan Pengujian Ketahanannya
terhadap Panas. Skripsi. Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Dr. Irma Isnafia Arief, S.Pt., M.Si. Pembimbing Anggota : Tuti Suryati S.Pt., M.Si.
Penggunaan bakteriosin sebagai biopreservatif dalam makanan saat ini telah berkembang di negara-negara maju. Bakteriosin merupakan salah satu substrat antimikrob yang dihasilkan oleh bakteri Gram positif maupun Gram negatif dan berpotensi sebagai bahan pengawet makanan yang alami. Pemanasan merupakan metode yang umum digunakan dalam pengolahan makanan termasuk daging dan produk olahan asal daging untuk memusnahkan mikrob pembusuk dan patogen. Penelitian sebelumnya melaporkan bahwa isolat indigenousLactobacillus plantarum
yang diisolasi dari daging sapi lokal Indonesia menghasilkan suatu senyawa anti-mikrob yang diidentifikasi awal sebagai plantaricin.
Tujuan penelitian ini adalah mengkarakterisasi ketahanan bakteriosin dari
Lactobacillus plantarum terhadap pemanasan pada suhu pasteurisasi dan sterilisasi. Penelitian dilakukan di Laboratorium Mikrobiologi dan Laboratorium Terpadu, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian dilaksanakan dari bulan April 2011 hingga September 2011. Pengujian stabilitas aktivitas plantaricin yang telah mengalami pemanasan dilakukan terhadap bakteri uji Escherichia coli ATCC 25922, Salmonella enteriditis
ser. Typhimurium ATCC 14028, Staphylococcus aureus ATCC 25923, Pseudomo-nasaeruginosa ATCC 27853 dan Bacillus cereus.
Hasil penelitian menunjukkan bahwa plantaricin dari keempat galur
Lactobacillus plantarum yaitu L. plantarum 2C12, 1B1, 2B2 dan 1A5 tidak berbeda nyata terhadap perlakuan pemanasan. Hal ini menunjukkan bahwa plantaricin dari keempat galur Lactobacillus plantarum stabil terhadap perlakuan pemanasan (80 °C selama 30 menit dan 121 °C selama 15 menit). Galur L. plantarum yang berbeda menghasilkan plantaricin yang mempunyai aktivitas antimikrob yang berbeda pula dalam menghambat bakteri Pseudomonas aeruginosa ATCC 27853, Staphylococcus aureus ATCC 25923, Escherichia coli ATCC 25922 dan Salmonella enteriditis ser. Typhimurium ATCC 14028 namun aktivitas antimikrobnya tidak berbeda dalam menghambat Bacillus cereus. Kemampuan plantaricin yang mempunyai stabilitas aktivitas antimikrob terhadap bakteri indikator setelah pemanasan membuktikan potensi plantaricin sebagai biopreservatif pangan.
ABSTRACT
Production of Bacteriocins of Lactobacillus plantarum as Antimicrobial and Their Heat Stability Test
Marselly, T.S., I.I. Arief and T. Suryati
Bacteriocin is commonly used to biopreservative agent in food. Bacteriocin is an antimicrobial substance that is capable to inhibit toward pathogenic and food borne bacteria. It was reported that indigenous isolate of Lactobacillus plantarum isolated from Indonesian local beef producing an antimicrobial substance (Arief et al., 2008). The objective of this research was to characterize the heat stability of plantaricin at pasteurization and sterilization temperature. The treatments were heat temperature (80 °C for 30 minutes and 121 °C for 15 minutes) and strains of Lactobacillus plantarum (2C12, 1A5, 1B1 and 2B2) usage. The variable was diameter of inhibition zone of antagonistic activity. Antimicrobial activity of plantaricin of Lactobacillus plantarum 2C12, 1B1, 2B2, and 1A5 did not lost after treated at 80 °C for 30 minutes or at 121 °C for 15 minutes. That result showed that plantaricin was stable at pasteurization and sterilization temperature. Plantaricin produced by strains of L. plantarum against indicator bacteria such as Pseudomonas ATCC 27853,
Staphylococcus aureus ATCC 25923, Escherichia coli ATCC 25922 and Salmonella enteriditis ser. Typhimurium ATCC 14028 had different activity each other. However, inhibition activity of plantaricin did not lead against Bacillus cereus. Stability of antimicrobial activity of plantaricin to inhibit indicator bacteria after heat treatment proved that plantaricin has a potential as food biopreservative.
PRODUKSI BAKTERIOSIN ASAL Lactobacillus plantarum
SEBAGAI ANTIMIKROB DAN PENGUJIAN
KETAHANANNYA TERHADAP PANAS
TRI SANTI MARSELLY
D14070060
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
Judul : Produksi Bakteriosin Asal Lactobacillus plantarum sebagai Antimikrob dan Pengujian Ketahanannya terhadap Panas
Nama : Tri Santi Marselly
NIM : D14070060
Menyetujui,
Pembimbing Utama,
(Dr. Irma Isnafia Arief, S.Pt., M.Si.) NIP.19750304 199903 2 001
Pembimbing Anggota,
(Tuti Suryati S.Pt., M.Si.) NIP.19720516 199702 2 001
Mengetahui: Ketua Departemen
Ilmu Produksi dan Teknologi Peternakan
(Prof. Dr. Ir. Cece Sumantri, M.Agr.Sc.) NIP: 19591212 198603 1 004
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 23 Maret 1990 di Tegal, Jawa Tengah. Penulis merupakan anak ketiga dari empat bersaudara dari pasangan Bapak Untung Masyugi dan Ibu Ratini. Penulis mengenal pendidikan formal di Taman Kanak-kanak Pertiwi pada tahun 1993-1995. Tahun 2001 penulis menyelesaikan pendidikan dasar di SD Negeri Kalibakung 02, Balapulang, Tegal. Pendidikan dilanjutkan di SLTP Negeri 14 Tegal, hingga tahun 2004 dan pendidikan lanjutan menengah atas diselesaikan pada tahun 2007 di SMA Negeri 1 Tegal.
Penulis diterima di Fakultas Peternakan IPB melalui jalur Undangan Seleksi Masuk IPB (USMI) pada tahun 2007. Selama kuliah, penulis pernah menjadi asisten mata kuliah Teknik Pengolahan Daging pada tahun ajaran 2010/2011. Penulis aktif dalam Organisasi Mahasiswa Daerah Ikatan Mahasiswa Tegal di IPB sebagai sekretaris, periode 2009/2010. Pelatihan yang pernah diikuti penulis yaitu pelatihan
Hazard Analysis and Critical Control Points (HACCP) yang diselenggarakan olah Departemen Ilmu Produksi dan Teknologi Peternakan pada tahun 2010. Penulis menjadi penerima beasiswa PPA (Peningkatan Prestasi Akademik) pada tahun 2008-2010. Prestasi yang dicapai oleh penulis adalah penulis lolos untuk mengikuti Program Kreativitas Mahasiswa (PKM) bidang penelitian tahun 2011.
Penulis melakukan penelitian dan penulisan skripsi yang berjudul “Produksi Bakteriosin Asal Lactobacillus plantarum sebagai Antimikrob dan Pengujian
KATA PENGANTAR
Segala puji bagi Allah SWT yang telah memberikan rahmat serta karunia-Nya sehingga penulisan skripsi ini dapat diselesaikan dengan baik. Semoga shalawat dan salam tercurah kepada Nabi Muhammad shallallahu’alaihi wa salam.
Penulis juga mengucapkan terima kasih kepada semua pihak yang telah mendukung baik secara moril maupun materil sehingga penulis dapat menyelesaikan tugas akhir penelitian dan skripsi dengan judul “Produksi Bakteriosin Asal
Lactobacillus plantarum sebagai Antimikrob dan Pengujian Ketahanannya
terhadap Panas” inisebagai salah satu syarat memperoleh gelar Sarjana Peternakan pada Fakultas Peternakan IPB.
Bakteriosin merupakan salah satu substrat antimikrob yang dihasilkan oleh bakteri asam laktat dan berpotensi sebagai bahan pengawet makanan yang alami.
Plantaricin merupakan salah satu contoh bakteriosin yang dihasilkan oleh
Lactobacillus plantarum. Perkembangan teknologi yang diterapkan pada makanan saat ini mengacu pada kombinasi pengolahan dan penambahan bahan pengawet alami. Metode yang umum digunakan dalam pengolahan makanan termasuk daging dan produk olahan asal daging untuk memusnahkan mikrob pembusuk dan patogen yakni dengan pemanasan. Keamanan produk pangan asal daging tidak dapat dijamin jika metode pemanasan yang digunakan tidak dilakukan dengan benar sehingga perlu adanya agen preservatif alami yang tahan terhadap pemanasan. Banyak penelitian telah berhasil mengkarakterisasi kestabilan bakteriosin terhadap pemanasan. Penelitian ini berupaya memberikan informasi tentang karakteristik kestabilan
plantaricin terhadap pemanasan dalam menghambat dan menekan pertumbuhan bakteri patogen dan pembusuk makanan.
Penulis berharap semoga skripsi ini dapat memberikan manfaat bagi para pembaca pada umumnya dan terutama bagi penulis sendiri pada khususnya. Amien.
Bogor, Pebruari 2012
DAFTAR ISI
Mekanisme Aktivitas Baktesiosin ... 4
Perlakuan Pemanasan terhadap Bakteriosin ... 5
Penggunaan Bakteriosin ... 6
Bakteri Patogen dan Pembusuk Makanan ... 7
Escherichia coli ... 7
Staphylococcus aureus ... 8
Salmonella enteriditis ser.Typhimurium ... 9
Bacillus cereus ... 10
Pseudomonas ... 11
Pemurnian Bakteriosin ... 11
Purifikasi Parsial dengan Presipitasi Amonium Sulfat ... 11
Kromatografi Pertukaran ion ... 12
METODE ... 14
Lokasi dan Waktu ... 14
Materi ... 14
Prosedur ... 14
Penyegaran Kultur L. plantarum dan Bakteri Indikator ... 15
Uji Kemurnian Kultur dengan Menggunakan Uji Pewarnaan Gram ... 15
Pembiakan Kultur dan Identifikasi Keberadaan Bakteriosin ... 16
viii Produksi dan Purifikasi Parsial dengan Menggunakan
Presipitasi Amonium Sulfat ... 16
Dialisis ... 17
Purifikasi Plantaricin dengan Kromatografi Kolom ... 18
Stabilitas Protein Plantaricin Selama Pemanasan ... 18
Uji Kemampuan Antagonistik Plantaricin ... 19
Rancangan dan Analisis Data ... 20
Identifikasi Keberadaan Bakteriosin ... 20
Produksi Plantaricin ... 20
Stabilitas Aktivitas Plantaricin terhadap Pemanasan ... 21
HASIL DAN PEMBAHASAN ... 23
Uji Kemurnian Isolat Lactobacillus plantarum dan Bakteri Indikator .. 23
Identifikasi Keberadaan Bakteriosin ... 26
Produksi dan Purifikasi Plantaricin dengan Purifikasi Parsial ... 27
Stabilitas Protein Plantaricin Setelah Pemanasan ... 29
Uji Kemampuan Antagonistik Plantaricin Setelah Pemanasan pada Berbagai Bakteri Indikator ... 30
Staphylococcus aureus ATCC 25923 ... 30
Pseudomonasaeruginosa ATCC 27853 ... 31
Escherichia coli ATCC 25922 ... 32
Salmonella enteriditis ser. Typhimurium ATCC 14028 ... 33
Bacillus cereus ... 34
KESIMPULAN DAN SARAN ... 36
Kesimpulan ... 36
Saran ... 36
UCAPAN TERIMAKASIH ... 37
DAFTAR PUSTAKA ... 38
DAFTAR TABEL
1. Penggunaan Padatan Amonium Sulfat (% Penjenuhan) ... 17 2. Karakteristik Isolat Bakteri Asam Laktat ... 23 3. Karakteristik Isolat Bakteri Indikator ... 25 4. Diameter Zona Hambat Supernatan Bebas Sel (pH netral) L. plantarum
terhadap Bakteri Indikator ... 26 5. Kondisi pH Supernatan Lactobacillus plantarum ... 27 6. Diameter Zona Hambat Plantaricin Asal Empat Galur L. plantarum
dengan Perlakuan Pemanasan terhadap S. aureus ATCC 25923 ... 30 7. Diameter Zona Hambat Plantaricin Asal Empat Galur L. plantarum
dengan Perlakuan Pemanasan terhadap P. aeruginosa ATCC 27853 ... 31 8. Diameter Zona Hambat Plantaricin Asal Empat Galur L. plantarum
terhadap Escherichia coli ATCC 25922 ... 32 9. Diameter Zona Hambat Plantaricin Asal Empat Galur L. plantarum
dengan Perlakuan Pemanasan terhadap Salmonella enteriditis ser.
Typhimurium ATCC 14028 ... 33 10.Diameter Zona Hambat Plantaricin Asal Empat Galur L. plantarum
DAFTAR GAMBAR
1. Pengelompokan Bakteriosin yang Telah Dimodifikasi ... 3 2. Metode Pengukuran Zona Hambat ... 19 3. Konsentrasi Protein Presipitat Bakteriosin Hasil Presipitasi Parsial dan
Plantaricin Kasar Hasil Dialisis Asal Empat Galur Lactobacillus
plantarum (mg/ml) ... 28 4. Konsentrasi Protein Plantaricin Asal Empat Galur L. plantarum
Sebelum dan Setelah Perlakuan Pemanasan (mg/ml) ... 29 5. Zona Hambat Plantaricin Asal L. plantarum 2B2 Setelah Pemanasan
DAFTAR LAMPIRAN
1. Uji Kruskal-Wallis Diameter Zona Hambat Supernatan Bebas Sel (pH
netral) L. plantarum terhadap Bakteri Indikator ... 43 2. Uji Pembandingan Berganda Kruskal-Wallis Diameter Zona Hambat
SupernatanBebas Sel (pH netral) Lactobacillus plantarum terhadap
Bakteri Indikator ... 43 3. Uji Kruskal-Wallis Diameter Zona Hambat Supernatan Bebas Sel (pH
Netral) Asal Empat Galur Lactobacillus plantarum ... 43 4. Uji Pembandingan Berganda Kruskal-Wallis Diameter Zona Hambat
Supernatan Bebas Sel (pH netral) Asal Empat Galur L. plantarum ... 44 5. Uji Kruskal-Wallis pH Supernatan Bebas Sel Asal Empat Galur L.
plantarum ... 44 6. Uji Kruskal-Wallis pH Supernatan Netral Asal Empat Galur L. plantarum 44 7. Data Konsentrasi Protein Presipitat Bakteriosin dan Plantaricin Kasar
Asal Empat Galur Lactobacillus plantarum (µg/ml) ... 45 8. Data Konsentrasi Protein Plantaricin Kasar Asal Empat Galur L.
plantarum Sebelum dan Setelah Perlakuan Pemanasan (µg/ml) ... 45 9. Analisis Ragam Diameter Zona Hambat Plantaricin Asal Empat Galur
L. plantarum dengan Perlakuan Pemanasan terhadap S. aureus ATCC
25923 ... 46 10.Uji Tukey Diameter Zona Hambat Plantaricin Asal Empat Galur
L. plantarum terhadap Staphylococcus aureus ATCC 25923 ... 46 11.Analisis Ragam Diameter Zona Hambat Plantaricin Asal Empat Galur
L. plantarum dengan Perlakuan Pemanasan terhadap P. aeruginosa
ATCC 27853 ... 46 12.Uji Tukey Diameter Zona Hambat Plantaricin Asal Empat Galur L.
plantarum terhadap P. aeruginosa ATCC 27853 ... 47 13.Uji Kruskal-Wallis Diameter Zona Hambat Plantaricin dengan Perlakuan
Pemanasan terhadap E. coli ATCC 25922 ... 47 14.Uji Kruskal-Wallis Diameter Zona Hambat Plantaricin Asal Empat Galur
L. plantarum terhadap E. coli ATCC 25922 ... 47 15.Uji Pembandingan Berganda Kruskal-Wallis Diameter Zona Hambat
Plantaricin Asal Empat Galur L. plantarum terhadap Escherichia Coli
ATCC 25922 ... 48 16.Analisis Ragam Diameter Zona Hambat Plantaricin Asal Empat Galur
Lactobacillus plantarum dengan Perlakuan Pemanasan terhadap
xii 17.Uji Tukey Diameter Zona Hambat Plantaricin Asal Empat Galur L.
plantarum terhadap Salmonella enteriditis ser. Typhimurium ATCC
14028 ... 48
18.Uji Kruskal-Wallis Diameter Zona Hambat Plantaricin dengan Perlakuan Pemanasan terhadap Bacillus cereus ... 49
19.Uji Kruskal-Wallis Diameter Zona Hambat Plantaricin Asal Empat Galur L. plantarum terhadap Bacillus cereus ... 49
20.Persiapan Membran Dialisis ... 49
21.Gambar Alat Centrifuge untuk Produksi Plantaricin ... 50
22.Purifikasi Parsial dengan Menggunakan Presipitasi Amonium Sulfat ... 50
23.Persiapan dan Proses Dialisis ... 50
PENDAHULUAN
Latar Belakang
Penggunaan bakteriosin dalam makanan saat ini telah berkembang di negara-negara maju. Bakteriosin merupakan salah satu substrat antimikrob yang dihasilkan oleh bakteri Gram positif maupun Gram negatif yang memiliki aktivitas antagonistik, baik bakteriostatik (menghambat pertumbuhan bakteri lain) maupun bakterisidal (membunuh bakteri lain). Beberapa bakteriosin asal bakteri asam laktat (BAL) ber-potensi sebagai agen biopreservatif pangan dan penggunaan bakteriosin pada industri pangan dapat mengurangi penggunaan pengawet kimia. Bakteriosin yang baru dimanfaatkan secara komersial adalah nisin yang diproduksi oleh Lactococcus lactis.
Pemanasan merupakan metode yang umum digunakan dalam pengolahan makanan termasuk daging dan produk olahan asal daging untuk memusnahkan mikrob pembusuk dan patogen yang terdapat pada bahan makanan. Proses pemanas-an ypemanas-ang digunakpemanas-an pada pengolahpemanas-an daging yakni: a) pasteurisasi, pempemanas-anaspemanas-an suhu sedang atau moderat dengan suhu produk berkisar 58-80 ºC untuk memusnahkan mikrob patogen atau pembusuk, misalnya melalui proses perebusan, dan b) sterilisasi, yakni pemanasan suhu tinggi di atas 100 ºC untuk menghasilkan produk daging yang bebas dari mikrob, misalnya untuk produk olahan daging komersial seperti daging kaleng dan kornet daging sapi dalam kaleng.
Proses pemanasan merupakan titik kritis pada pengolahan pangan, untuk menjamin keamanan pangan. Namun demikian keamanan produk pangan asal daging tidak dapat dijamin jika metode penyimpanan setelah pemanasan tidak dilakukan dengan benar. Penambahan bakteriosin sebagai biopreservatif alami dapat menjadi alternatif untuk melindungi produk pangan selama penyimpanan. Bakteriosin dapat ditambah-kan langsung atau disemprotkan ke permukaan ke makanan dalam bentuk kultur ter-konsentrasi maupun dikombinasikan dengan film selulosa menjadi bahan kemasan aktif. Bakteriosin ini diharapkan dapat menghambat pertumbuhan bakteri pembusuk makanan dan patogen pada produk olahan daging setelah proses pemanasan.
diidenti-2 fikasi awal sebagai plantaricin (Arief et al., 2008). Penelitian tentang stabilitas
plantaricin asal empat galur Lactobacillus plantarum terhadap panas belum pernah dilaporkan. Oleh karena itu, plantaricin asal empat galur Lactobacillus plantarum
perlu diteliti lebih lanjut ketahanannya terhadap perlakuan pemanasan. Tujuan
3
TINJAUANPUSTAKA
Bakteriosin
Bakteriosin merupakan senyawa protein (umumnya berupa peptida kecil) yang bersifat bakterisidal terhadap galur Gram positif dan mikroorganisme (bakteri) yang ditinjau dari segi filogeniknya (genetiknya) berdekatan dengan mikroorganisme penghasil bakteriosin tersebut (Jay, 2000). Bakteriosin yang diproduksi oleh Lacto-bacillus plantarum dikenal dengan nama plantaricin (Omar et al., 2008). Plantaricin
ST31 terdiri atas 20 asam amino dengan massa total 2.755,63 Da (Todorov et al., 2004). Bakteriosin adalah metabolit sekunder yang dapat menghambat pertumbuhan bakteri lain dalam konsentrasi yang relatif kecil (Jack et al., 1995). Menurut Ray dan Bhunia (2008), bakteriosin adalah kelompok peptida-peptida bioaktif yang dipro-duksi oleh banyak strain bakteria baik yang termasuk bakteri Gram positif maupun Gram negatif.
Klaenhammer (1993) mengelompokkan bakteriosin berdasarkan genetik dan komponen penyusun biokimianya menjadi: (a) kelas I, mencakup lantibiotik seperti nisin, (b) kelas II adalah peptida kecil (<10 kDa) yang stabil panas seperti lactacin F, (c) kelas III adalah protein besar (>30 kDa) yang tidak tahan terhadap panas seperti
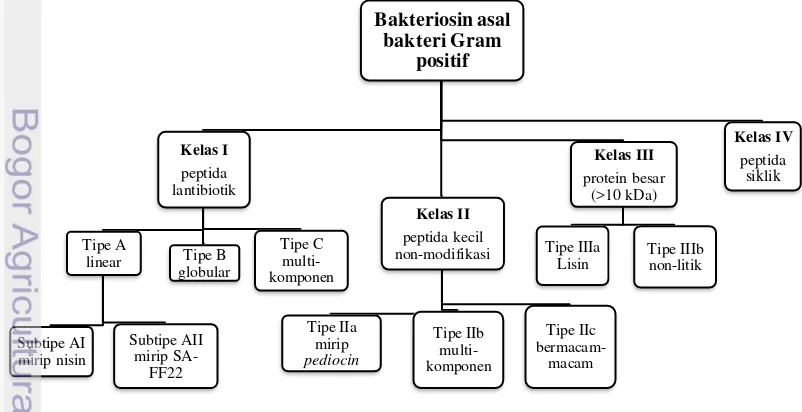
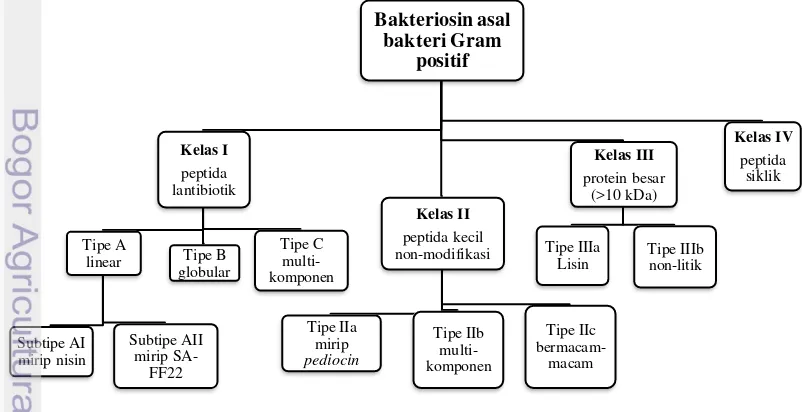
helveticin J, dan (d) kelas IV adalah protein yang terdiri atas sebuah kompleks dengan faktor-faktor lain seperti lipida esensial atau karbohidrat. Heng et al. (2007) telah memodifikasi pengelompokkan bakteriosin asal bakteri Gram positif secara lebih kompleks.
Gambar 1. Pengelompokan Bakteriosin Menurut Heng et al. (2007).
4 Beberapa bakteriosin dari bakteri asam laktat antara lain: plantaricin A dari
Lactobacillus plantarum (Nissen-Meyer et al., 1993), gassericin A dari Lacto-bacillus LA39 gasseri (Muriana dan Klaenhammer, 1991) dan plantaricin-149 dari
Lactobacillus plantarum KTP 149 (Kato et al., 1994) yang telah terdeteksi, dimurni-kan dan dikarakterisasi. Lactocin LC-09 yang diproduksi oleh Lactobacillus strain LC-09 memiliki daya hambat terhadap banyak spesies Lactobacilli dan bakteri Gram positif lain termasuk Listeria ivanovii, Streptococcus agalactii dan Streptococcus pyogenes (Khalid et al., 1999). Bakteriosin yang baru dimanfaatkan secara komersial adalan nisin yang diproduksi oleh Lactococcus lactis (Kato et al., 1993).
Mekanisme Aktivitas Bakteriosin
Kemampuan suatu senyawa antimikrob dalam menghambat pertumbuhan mikrob merupakan salah satu kriteria yang penting dalam pemilihan suatu senyawa antimikrob yang berfungsi sebagai bahan pengawet. Antimikrob menurut Gan dan Setiabudi (1987), adalah zat yang dapat menghambat pertumbuhan mikrob dan digunakan untuk pengobatan infeksi mikrob pada hewan dan manusia. Antimikrob harus mempunyai toksisitas setinggi mungkin terhadap bakteri target, tetapi relatif tidak toksik terhadap induk semangnya (Gan dan Setiabudi, 1987).
Berdasarkan sifat toksisitas selektifnya antimikrob dibedakan menjadi dua bagian, yaitu: (i) antimikrob yang bersifat bakteriostatik, yaitu antimikrob yang menghalangi pertumbuhan mikroorganisme, tetapi tidak mematikan organisme tersebut, dan (ii) antimikrob yang bersifat bakterisidal, yaitu antimikrob yang menyebabkan kematian dan lisisnya mikroorganisme. Beberapa mikrob yang bersifat bakteriostatik dapat berubah menjadi bakterisidal bila digunakan digunakan dalam dosis tinggi (Gan dan Setiabudi, 1987). Dwiajiseputro (1990) membedakan anti-mikrob berdasarkan efektivitas kerjanya terhadap berbagai mikroorganisme, yaitu: (i) antimikrob yang berspektrum luas, yaitu antimikrob yang efektif terhadap berbagai jenis mikroorganisme, dan (ii) antimikrob yang berspektrum sempit, yaitu antimikrob yang efektif terhadap mikroorganisme tertentu.
Banyak penelitian yang melaporkan jenis bakteri dari famili Lactobacillaceae
anti-5 mikrob bakteri asam laktat berkaitan dengan adanya asam organik (asam laktat, asam asetat dan asam format), hidrogen peroksida dan bakteriosin yang dihasilkannya (Schved et al.,1992).
Menurut Gonzalez et al. (1996), bakteriosin berpengaruh nyata pada proses gangguan potensial membran yaitu destabilisasi membran sitoplasmik. Bakteriosin yang terabsorpsi akan masuk ke dalam lapisan fosfolipida membran sel membentuk agregat yang melintang dari satu sisi membran ke sisi lain (Gonzalez et al., 1996). Moll et al. (1999) menambahkan bahwa kebanyakan bakteriosin berinteraksi dengan lipida anionik yang tersedia banyak dalam membran bakteri Gram positif. Gangguan potensial membran ini merupakan awal pembentukan lubang yang menyebabkan dibebaskannya molekul-molekul kecil dan menghilangnya substrat intraseluler (Gonzalez et al., 1996).
Ray dan Bhunia (2008) memaparkan bahwa secara kimia, peptida bakteriosin disintesis secara ribosomal, amfipatik, kationik dan mempunyai struktur α-helical
atau β-sheet, atau kedua-duanya, dan dapat juga mempunyai tioeter, jembatan disulfida, atau kelompok tiol bebas. Terdapatnya sebuah struktur amfipatik α-helical
dengan sisi polar dan nonpolar yang berlawanan membuat bakteriosin dapat berinteraksi dengan kedua fase air dan lipida ketika terikat dengan permukaan membran dari sel sensitif bakteri, sehingga sel mengalami destabilisasi fungsional dan sel tersebut mati (Ray dan Bhunia, 2008; Walker, 2000).
Perlakuan Pemanasan terhadap Bakteriosin
6 Proses pemanasan ini setara dengan pemanasan pada suhu 89 ºC selama 1 detik, 90 ºC selama 0,5 detik, dan 94 ºC selama 0,1 detik (Fardiaz, 1996).
Beberapa bakteriosin yang tahan terhadap pemanasan telah dilaporkan seperti Joshi et al. (2006) yang melaporkan bahwa aktivitas bakteriosin yang diproduksi dari isolat asal sayur fermentasi dapat stabil sampai dengan pemanasan pada suhu 100 °C tetapi aktivitas antimikrobnya lebih rendah jika dibandingkan dengan pemanasan pada suhu 68 °C dan aktivitas antimikrobnya benar-benar hilang setelah diberi pemanasan pada suhu 121 °C. Gonzalez et al. (1994) juga melaporkan bahwa
plantaricin C yang diproduksi Lactobacillus plantarum yang diisolasi dari susu menunjukkan aktivitas yang sangat stabil pada pemanasan pada suhu 100 °C selama 60 menit dan 121 °C selama 10 menit. Khalid et al. (1999) yang telah mendeteksi dan mengkarakterisasi bakteriosin asal isolat Lactobacilli (lactocin) melaporkan bahwa lactocin dapat stabil terhadap pemanasan pada suhu 100 °C selama 4 jam dan masih dapat mempertahankan aktivitasnya setelah pemanasan pada suhu 121 °C selama 15 menit. Kriteria kestabilan panas pada bakteriosin tergantung pada tahap pemurnian, pH, keadaan medium (Joshi et al., 2006).
Hata et al. (2010) melaporkan bahwa plantaricin ASM1 yang diproduksi oleh
Lactobacillus plantarum A-1 yang diisolasi dari tortila (roti jagung tradisional asal Meksiko) menunjukkan aktivitas yang stabil terhadap panas pada suhu 90 °C selama 15 menit dan zona hambat yang ditunjukkan hampir sama jika dibandingkan dengan protein yang tidak dipanaskan. Plantaricin ASM1 juga menunjukkan stabilitas untuk pemanasan dan berbagai kondisi pH, terutama di pH netral dan basa jika dibanding-kan dengan nisin A. Secara umum, ada banyak madibanding-kanan dengan pH 7,0-8,5 sehingga diharapkan akan mendapatkan makanan dengan efek yang sama (Hata et al., 2010).
Penggunaan Bakteriosin
anti-7 mikrob dengan spektrum relatif luas, terhadap bakteri patogen dan pembusuk makanan, dan (vi) memiliki aktivitas yang bersifat bakterisidal, bekerja pada membran sitoplasma bakteri: tidak ada resistensi silang dengan antibiotik (Gálvez et al., 2007; Jay et al., 2000).
Gálvez et al. (2007) menyatakan bahwa bakteriosin dapat ditambahkan langsung ke makanan dalam bentuk kultur terkonsentrasi yang dapat berperan sebagai pengawet makanan, bahan yang memperpanjang umur simpan, bahan tambahan makanan maupun sebagai bahan peramu. Bakteriosin dalam bentuk imobil juga dapat digunakan sebagai aplikasi untuk pengembangan kemasan bioaktif pada makanan seperti sosis. Nguyen et al. (2008) menambahkan bahwa kombinasi nisin dan film selulosa asal bakteri dapat menjadi bahan kemasan aktif. Film selulosa asal bakteri yang mengandung nisin menunjukkan efektivitas dalam pengendalian
Listeria monocytogenes dan mengurangi total mikrob pada permukaan frankfurter yang dikemas secara vakum (Nguyen et al., 2008). Hal ini menunjukkan bahwa penggunaan film selulosa aktif asal bakteri akan menjadi metode yang menjanjikan untuk meningkatkan keamanan dan memperpanjang umur simpan dari daging olahan (Nguyen et al., 2008).
Bakteri Patogen dan Pembusuk Makanan
Frazier dan Westhoff (1978) menyatakan bahwa terdapat dua cara bakteri dapat menularkan penyakit pada manusia yaitu 1) intoksikasi; melalui toksin yang dihasilkan bakteri yang tumbuh di dalam makanan tersebut, dan 2) infeksi; melalui bakteri yang masuk ke dalam tubuh melalui makanan yang telah terkontaminasi dan adanya reaksi dari tubuh terhadap keberadaan atau metabolit-metabolit yang dihasil-kan bakteri selama tumbuh di dalam tubuh. Keseimbangan flora usus harus dijaga dengan baik untuk menjaga tidak terkena infeksi atau intoksikasi (Frazier dan Westhoff, 1978).
Escherichia coli
Bakteri ini termasuk dalam famili Enterobacteriaceae yang termasuk Gram negatif dan berbentuk batang yang fermentatif (Blackburn, 2002). Escherichia coli
8 sedang dan penyebaran yang cepat dalam kasus keracunan makanan (Gaman dan Sherrington, 1992). Pelczar dan Chan (2005) menyatakan bahwa Escherichia coli
0157:H7 banyak dijumpai pada daging giling dan sejenisnya. Bakteri ini dapat tumbuh dalam kisaran suhu yang luas yaitu 1-45 °C, sehingga kemungkinan pangan Indonesia tercemar bakteri ini sangat besar mengingat penanganan bahan pangan yang kurang memadai (Pelczar dan Chan, 2005). Escherichia coli juga hidup dalam jumlah besar di dalam usus manusia, yaitu membantu sistem pencernaan manusia dan melindunginya dari bakteri patogen (Blackburn, 2002; Pelczar dan Chan, 2005).
Staphylococcus aureus
Bakteri ini merupakan bakteri Gram positif berbentuk bulat umumnya bergerombol seperti buah anggur, non motil, tidak berkapsul dan tidak membentuk spora (Ray dan Bhunia, 2008). Bakteri ini ditemukan individual, berpasangan, berantai pendek. Koloni pada media agar berbentuk bundar, licin, berwarna jingga hingga putih, berkilauan, menonjol dan menyebar (Pelczar dan Chan, 2005).
Gaman dan Sherrington (1992) menyatakan bahwa Staphylococcus aureus
menyebabkan infeksi jaringan organ atau tubuh dengan tanda-tanda peradangan dan pembentukan abses serta menginfeksi luka-luka dan bisul. Staphylococcus aureus
dapat menghasilkan toksin dengan periode inkubasi akibat intoksikasi dalam tubuh sekitar 2-6 jam (Gaman dan Sherrington, 1992). Gejala yang timbul ialah muntah, kram perut (Jay et al., 2000), diare bahkan pingsan (Gaman dan Sherrington, 1992).
9
Salmonella enteriditis ser. Typhimurium
Bakteri ini merupakan bakteri berbentuk batang Gram negatif, motil, anaerobik dan tidak membentuk spora (Frazier dan Westhoff, 1978). Bakteri ini dapat tumbuh pada pH netral 6,6-8,2 dan memiliki ketahanan panas yang tinggi pada pH 5,5 dan aw rendah. Suhu optimum untuk pertumbuhan 37 °C, suhu maksimum 45,6 °C dengan nilai aw umumnya berkisar 0,93-0,95 (Frazier dan Westhoff, 1978). Sel bakteri ini dapat bertahan dalam suhu beku dan keadaan kering dalam waktu yang lama (Ray dan Bhunia, 2008).
Penamaan Salmonella tidak mengikuti aturan nomenklatur yang biasanya.
Salmonella typhimurium menyebabkan typhoid pada tikus dan Salmonella typhi
menyebabkan typhoid pada manusia. Istilah serovar digunakan untuk membedakan galur berdasarkan perbedaan pelengkap antigenik (Frazier dan Westhoff, 1978) yang memiliki perbedaan reaksi aglutinasi antara antisera homolog dan kombinasi antigen yang terkandung setiap galur Salmonella (Blackburn, 2002).
Infeksi oleh bakteri genus Salmonella menyerang saluran gastrointestin yang mencakup perut, usus halus, dan usus besar atau kolon (Pelczar dan Chan, 2005). Delapan sampai empat puluh delapan jam setelah makan makanan yang tercemar dengan Salmonella, timbul rasa sakit perut yang mendadak dengan diare encer atau berair, kadang-kadang dengan lendir atau darah. Sering kali mual atau muntah, demam dengan suhu 38-39 oC umum terjadi. Gejala-gejala ini ada hubungannya dengan endotoksin tahan panas yang dihasilkan oleh Salmonella. Gejala-gejala tersebut biasanya hilang dalam waktu 2-5 hari dan berakhir dengan kesembuhan si penderita (Pelczar dan Chan, 2005). Infeksi yang disebabkan oleh Salmonella dapat berakibat fatal, terutama jika menyerang bayi. Oleh sebab itu infeksi Salmonella
pada makanan mendapat perhatian yang serius, karena bakteri ini seringkali menim-bulkan penyakit terutama di negara-negara industri (Ray dan Bhunia, 2008).
10 minuman yang terkontaminasi. Makanan tersebut termasuk daging sapi, daging ayam, daging kalkun, daging babi, telur, susu, dan produk olahannya (Pelczar dan Chan, 2005; Ray dan Bhunia, 2008). Penggunaan antibiotik pada pakan hewan dan burung menyebabkan tingkat resisten (multidrug resistant) galur Salmonella
terhadap banyak obat meningkat termasuk serotip dari Salmonella enterica. Galur ini resistan terhadap beberapa antibiotik seperti ampisilin, kloramfenikol, streptomisin, sulfonamida, florfenikol, dan tetrasiklin (Ray dan Bhunia, 2008).
Bacillus cereus
Bakteri iniberbentuk batang berukuran 0,3-2,2 x 1,27-7,0 µm, sebagian besar motil dengan flagela khas lateral, membentuk endospora (tidak lebih dari satu sel sporangium), sel sensitif terhadap pasteurisasi. Spora dapat bertahan pada perlakuan panas tinggi yang digunakan pada proses pemanasan pada umumnya (Pelczar dan Chan, 2005; Ray dan Bhunia, 2008). Bacillus cereus merupakan salah satu bakteri patogen dan perusak pangan yang menghasilkan enterotoksin yang menyebabkan keracunan makanan pada manusia (Jay, 2000). Kemampuan untuk memproduksi berbagai enzim pencerna makanan seperti protease, amilase, lesitinase membuat
Bacillus cereus dapat bertahan dan tumbuh baik dalam kondisi yang berbeda-beda.
Bacillus cereus dapat memproduksi sedikitnya tiga enterotoksin yang berbeda, dua dari enterotoksin tersebut adalah toksin tiga pihak (tripartite) dan keduanya berhubungan dengan penyakit asal makanan (Blackburn, 2002).
11
Pseudomonas aeruginosa
Bakteri ini dapat ditemukan di tanah, air dan saluran urinasi yang terinfeksi dan juga telah diketahui sebagai penyebab mastitis. Pseudomonas aeruginosa
merupakan bakteri yang berbentuk batang lurus atau melengkung, psikotrof dan tergolong bakteri Gram negatif (Ray dan Bhunia, 2008). Pada umumnya berukuran 0,5-1,0 x 1,5-4,0 µm. Bakteri ini dapat bergerak dengan flagela polar; monotrikus atau multitrikus, tidak menghasilkan selongsong prosteka, tidak dikenal adanya stadium istirahat, kemoorganotrof, metabolisme dengan respirasi, tidak pernah fermentatif (Pelczar dan Chan, 2005). Pseudomonasaeruginosa mudah tumbuh dan menyebabkan kerusakan pada produk pangan disebabkan kemampuannya untuk menggunakan berbagai sumber karbon bukan karbohidrat dan tumbuh baik pada suhu dingin (dalam lemari pendingin) serta menghasilkan senyawa-senyawa penyebab bau busuk pada pangan (Frazier dan Westhoff, 1978).
Pemurnian Bakteriosin
Purifikasi Parsial dengan Presipitasi Amonium Sulfat
12 Ukuran partikel endapan saat terjadi pengendapan ditentukan oleh laju relatif dua proses berikut: (i) pembentukan inti (nuclei), yang disebut nukleiasi, dan (ii) pertumbuhan inti-inti untuk membentuk partikel-partikel yang cukup besar untuk mengendap. Jika laju pembentukan inti lebih rendah daripada pertumbuhan inti maka hanya dihasilkan partikel dengan jumlah yang sedikit namun ukurannya relatif besar sehingga lebih murni dan mudah disaring (Day dan Underwood, 2002). Pengotor pada permukaan partikel-partikel protein yang sangat kecil dapat dicuci secara normal, karena pertikel-partikel tersebut tidak terikat kuat satu sama lain dan cairan pencuci dapat memasuki semua bagian partikel tersebut. Proses pencucian ini disebut proses dialisis.
Kromatografi Pertukaran Ion
Resin penukar ion terdiri atas matriks tak larut dengan gugus bermuatan yang terikat secara kovalen. Menurut Day dan Underwood (2002), resin pertukaran ion diperoleh dengan memasukkan gugus yang dapat diionisasi ke dalam matriks polimer organik, yang paling umum adalah polistirena terhubung silang yang selanjutnya disebut adsorben. Secara komersial resin penukar ion ada dua yaitu penukaran kation (bermuatan negatif) akan mengikat ion bermuatan positif (kation), dan resin penukar anion (bermuatan positif) akan mengikat ion bermuatan negatif (anion). Dua resin yang sering digunakan dalam kromatografi penukar ion adalah karboksimetil selulosa (KM selulosa) dan dietilamino etil selulosa (DEAE selulosa). SP SepharoseTMFast Flow merupakan salah satu penukar kation kuat yang memiliki
kapasitas tinggi untuk semua nilai pI (titik isoelektrik) protein. Kelompok penukar ion SP SepharoseTM adalah kelompok sulfopropil yang stabil baik secara fisik
maupun kimia (Wikströms, 2002).
13 Day dan Underwood (2002) menyatakan bahwa protein merupakan polimer asam amino terkondensasi. Melalui gugus α-amino dan karboksil yang mudah terdisosiasi dan pada beberapa kasus, penambahan ionisasi dari fungsionalitas gugus R yang terionisasi, muatan pada sebuah molekul asam amino dapat dimanipulasi melalui pH, sehingga pertukaran ion dapat digunakan dalam fraksinasi. Protein tertentu memiliki titik isoelektrik (pI). Titik isoelektrik (pI) adalah nilai pH yang jumlah muatan negatif sama dengan jumlah muatan positif atau tidak memiliki muatan. Protein tidak akan terikat pada resin penukar ion dan akan mengalir keluar kolom pada pH isoelektrik (pI). Protein akan bermuatan positif dan akan berikatan dengan penukar kation (SP Sepharose) ketika pH di bawah pH isoelektrik (pI). Cakupan pH efektif SP Sepharose berkisar pada pH 4-13 (Wikströms, 2002).
Molekul-molekul netral tidak dapat menemukan jalannya menuju pori-pori suatu resin pertukaran ion, dan pada umumnya molekul tersebut dapat dibilas keluar dengan air atau dengan pelarut yang lain. Buffer yang digunakan pada kromatografi penukar kation adalah buffer anion seperti asetat, barbiturat, dan fosfat (Wilson, 2000). Zat terlarut yang ada dalam resin yang tidak secara kuat sebagai ion dikatakan “tersorpsi” (sorbed), pemilihan zat terlarut seperti itu oleh resin disebut “sorpsi”.
Sebuah kation tidak dapat berpartisipasi dalam resin pertukaran anion, kation ini mungkin bisa menuju pori-pori resin melalui semacam sorpsi elektrolit, tetapi kation ini tidak akan terikat secara kuat dan mungkin dapat dicuci keluar oleh pelarut (Day dan Underwood, 2002).
METODE
Lokasi dan Waktu
Penelitian dilakukan di Laboratorium Mikrobiologi dan Laboratorium Terpadu, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian dilakukan mulai bulan April 2011 hingga September 2011.
Materi
Bahan utama yang digunakan adalah kultur Lactobacillus plantarum 2C12, 1A5, 1B1 dan 2B2 koleksi Laboratorium Teknologi Hasil Ternak, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, IPB. Bakteri indikator yang digunakan untuk uji antagonis adalah bakteri patogen dan pembusuk makanan (Escherichia coli ATCC 25922, Salmonella enteriditis ser. Typhimurium ATCC 14028, Staphylococcus aureus ATCC 25923, Pseudomonas aeruginosa ATCC 27853 dan Bacillus cereus). Media yang digunakan yaitu media Mueller Hinton medium agaruntuk media konfrontasi plantaricin dengan bakteri indikator, media de Man Rogosa and sharp broth (MRSB), de Man Rogosa and Sharpe agar (MRSA),
nutrient broth (NB), nutrient agar (NA), yeast ekstrak, membran saring Sartosius, NaOH 1 N, serbuk amonium sulfat, buffer kalium fosfat pH 6,8 (campuran KH2PO4 dan K2HPO4) dan resin SP SepharoseTM fast flow, kristal violet, safranin, etanol 95%
dan larutan Mc. Farland no. 0,5. Larutan pengencer yang digunakan adalah NaCl 0,85% dan akuades.
Alat-alat yang digunakan adalah autoclave, waterbath, inkubator, mikroskop OPMIAS En Ver1.0, centrifuge, mikropipet, pipet Pasteur, tip, cawan petri, tabung reaksi, labu Erlenmeyer, gelas objek, refrigerator, spektrofotometer UV-visible,ose, pH meter, jangka sorong digital, bunsen, timbangan, oven, vortex, dan cork borer.
Prosedur
Penelitian ini diawali dengan penyegaran kultur Lactobacillus plantarum dan bakteri indikator, kemudian diuji kemurnian kultur melalui pewarnaan Gram. Tahap selanjutnya adalah pembiakan kultur Lactobacillus plantarum dan identifikasi keberadaan substrat antimikrob dari supernatan bebas sel netral asal empat galur
planta-15
ricin, stabilitas aktivitas plantaricin selama pemanasan dan uji kemampuan antago-nistik plantaricin pada berbagai bakteri indikator dengan metode difusi sumur.
Penyegaran Kultur Lactobacillus plantarum dan Bakteri Indikator (Pelczar dan Chan, 2005)
Kultur Lactobacillus plantarum dan bakteri indikator masing-masing dilaku-kan penyegaran pada media de Man Ragosa sharp broth (MRSB) dan nutrient broth
(NB). Kultur induk sebanyak 250 µ l diinokulasikan secara duplo pada media sehingga menghasilkan kultur sebanyak 5 ml. Kultur kemudian diinkubasi pada suhu 37 °C selama 24 jam yang hasilnya disebut kultur antara. Sebanyak 1 ml kultur antara diinokulasikan secara duplo pada media sehingga menghasilkan kultur sebanyak 10 ml. Kultur diinkubasikan kembali yang hasilnya disebut kultur kerja. Kultur kerja ditumbuhkan pada media de Man Ragosa sharp agar (MRSA) dan
nutrient agar (NA) untuk dihitung populasinya dan digunakan untuk pewarnaan Gram.
Uji Kemurnian Kultur dengan Menggunakan Uji Pewarnaan Gram (Pelczar dan Chan, 2005)
16 Pembiakan Kultur dan Identifikasi Keberadaan Bakteriosin
Sebanyak 5 kultur Lactobacillus plantarum masing-masing diinokulasikan ke media de Man Ragosa Sharp Broth (MRSB) dan diinkubasikan pada suhu 37 °C selama 24 jam. Supernatan bebas sel diperoleh melalui sentrifugasi yang selanjutnya disebut supernatan. Supernatan disaring dengan membran Sartorius 0,22 µm dan dikondisikan pada pH 5,8-6,2 dengan penambahan NaOH 1 N. Selanjutnya super-natan yang merupakan ekstrak kasar bakteriosin tersebut siap untuk diuji aktivitasnya dengan metode difusi sumur.
Produksi dan Purifikasi Plantaricin
Purifikasi bertujuan untuk mengetahui komponen-komponen senyawa kimia yang dapat terpisah dan kandungan senyawa aktifnya. Purifikasi plantaricin terdiri atas purifikasi parsial dengan menggunakan presipitasi amonium sulfat dan kromato-grafi pertukaran kation.
Produksi dan Purifikasi Parsial Plantaricin dengan Menggunakan Presipitasi
Amonium Sulfat. Sebanyak 1 liter media MRS broth ditambah yeast ekstrak 3% dan NaCl 1% diinokulasikan dengan 10% (v/v) kultur Lactobacillus plantarum. Terdapat empat galur Lactobacillus plantarum yang digunakan untuk menghasilkan senyawa antimikrob yaitu Lactobacillus plantarum 2C12, 1A5, 1B1 dan 2B2 yang telah disegarkan. Selanjutnya media yang mengandung kultur diinkubasi pada suhu 37 oC selama 20 jam. Setelah itu biakan kultur disimpan dalam refrigerator dengan suhu 4 oC selama dua jam. Tahap selanjutnya dilakukan sentrifugasi pada kecepatan 10000 rpm selama 20 menit pada suhu 4 oC. Setelah selesai, penyaringan dilakukan dengan menggunakan membran saring Sartorius diameter 0,22 µ m dan selanjutnya supernatan bebas sel dari setiap galur Lactobacillus plantarum dinetralkan pH-nya menjadi pH 5,8-6,2 dengan menggunakan 1 N NaOH. Semua tahapan proses ini dilakukan pada suhu dingin (4 oC) (Todorov et al., 2008).
Setiap supernatan antimikrob yang telah disaring steril ditambahkan serbuk amonium sulfat sebanyak 80% secara bertahap (20%, 40%, 60% dan 80%) ke dalam supernatan antimikrob yang telah disaring steril, kemudian dihomogenkan dengan
17 ditambahkan ke 1000 ml supernatan didasarkan pada Tabel 1. Pengecekan protein presipitat bakteriosin diamati dengan menggunakan spektrofotometer UV-visible
pada panjang gelombang 280 nm.
Tabel 1. Penggunaan Padatan Amonium Sulfat (% Penjenuhan) Awal
% 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90 95 100
Konsentrasi Akhir dari Padatan Amonium Sulfat (gram)
0 106 134 164 194 226 258 291 326 361 398 436 476 516 559 603 650 697
selanjut-18 nya disebut plantaricin kasar. Pengecekan protein plantaricin hasil dialisis diamati dengan menggunakan spektrofotometer UV-visible pada panjang gelombang 280 nm.
Purifikasi Plantaricin dengan Menggunakan Kromatografi Kolom. Kolom di-pasang penjepit buret kemudian buffer kalium fosfat pH 6,8 dituangkan ke dalam kolom. Buffer dibuang secara perlahan. Resin SP SepharoseTM fast flow dimasukkan
ke dalam kolom secara perlahan dengan menggunakan pipet Pasteur dan dijaga agar tidak ada udara (gas) yang masuk ke dalam kolom. Resin akan menjadi gel. Di atas resin diberikan buffer dan kolom disimpan pada suhu dingin sampai siap untuk digunakan.
Tabung penampung ditempatkan di bawah kolom untuk menampung eluat yang keluar dari kolom. Plantaricin kasar hasil dialisis dimasukkan ke dalam kolom secara perlahan. Eluat pertama adalah buffer. Eluatberikutnya adalah sampel planta-ricin murni. Kecepatan alir eluat yang keluar dari kolom adalah 0,8 ml/menit. Setelah selesai, dilakukan pencucian kembali dengan buffer dan ditampung untuk mengambil eluat yang terikat pada gel (resin). Semua dilakukan di ruang dingin (4 °C). Setelah selesai dalam beberapa tabung koleksi didapatkan eluat yang berisikan plantaricin
murni. Plantaricin murni disimpan pada suhu dingin (4 °C) dan selanjutnya siap untuk dianalisis karakteristiknya. Pengecekan protein plantaricin diamati dengan menggunakan spektrofotometer UV-visible pada panjang gelombang 280 nm (Hata
et al., 2010).
Stabilitas Protein Plantaricin Selama Pemanasan
19
Plantaricin yang diuji kemampuan antagonistiknya terhadap bakteri indikator diperoleh dari plantaricin dengan nilai protein tertinggi yang dilihat dari spektrum tertinggi.
Uji Kemampuan Antagonistik Plantaricin Setelah Pemanasan pada Berbagai Bakteri Indikator dengan Metode Difusi Sumur (Savadogo et al., 2006)
Plantaricin murni hasil kromatografi kolom diuji dengan menggunakan metode difusi sumur (Savadogo et al., 2006). Kultur bakteri indikator (patogen dan pembusuk makanan) sebanyak 107 cfu/ml yang berumur 24 jam dipipet ke dalam cawan petri dan ditambahkan media konfrontasi Muller Hinton medium agar sebanyak 20 ml.
Setelah agar dalam cawan mengeras, ditengah-tengah agar dibuat lubang sumur dengan menggunakan cork borer berdiameter 5 mm. Plantaricin murni kemu-dian dipipet ke dalam lubang sumur sebanyak 50 µ l kemukemu-dian disimpan dalam
refrigerator (suhu 7 °C) untuk memberikan kesempatan plantaricin meresap ke dalam agar selama dua jam. Selanjutnya agar diinkubasi pada suhu 37 °C selama 24 jam. Zona hambat diukur dengan mengukur zona bening atau semu di sekeliling sumur. Zona bening yang terbentuk di sekitar area sumur menandakan bahwa
plantaricin mempunyai aktivitas penghambatan total, sedangkan zona semu yang terbentuk menandakan plantaricin mempunyai aktivitas penghambatan parsial terhadap bakteri indikator. Pengukuran diameter zona penghambatan menggunakan jangka sorong. Tiap area zona bening diukur diameternya sebanyak tiga kali di tempat yang berbeda dan hasilnya dirata-ratakan.
Gambar 2. Metode Pengukuran Zona Hambat. A= Lubang Sumur untuk Plantaricin
(5 mm); B= Zona Hambat (Zona Bening/Semu); C= Cawan Petri (Media MHA); Garis = Pengukuran Diameter Zona Bening.
20 Rancangan dan Analisis Data
Penelitian ini dikelompokkan berdasarkan tahap penelitian yakni identifikasi keberadaan bakteriosin, produksi plantaricin, serta uji stabilitas aktivitas plantaricin
akibat pemanasan. Rancangan dan analisis data yang digunakan pada setiap tahap tersebut berbeda-beda seperti pada uraian berikut.
Identifikasi Keberadaan Bakteriosin
Rancangan yang digunakan adalah rancangan acak lengkap (RAL) faktorial dengan dua faktor perlakuan yaitu galur Lactobacillus plantarum yang berbeda (L. plantarum 2C12, 1B1, 2B2 dan 1A5) dan jenis bakteri indikator (Pseudomonas aeruginosa ATCC 27853, Staphylococcus aureus ATCC 25923, Bacillus cereus, Salmonella enteriditis ser. Typhimurium ATCC 14028, dan Escherichia coli ATCC 25922) dengan 3 ulangan. Peubah yang diamati yaitu diameter zona hambat supernatan netral. Model analisis yang digunakan sebagai berikut:
Yijk = µ + αi + βj + αβij + εijk Keterangan:
Yijk = Peubah respon karena pengaruh bersama taraf ke-i faktor α (galur L. plantarum) dan taraf ke-j faktor β (jenis bakteri indikator) pada ulangan ke-k (k = 1, 2, 3)
µ = Pengaruh rata-rata galur L. plantarum dan jenis bakteri indikator αi = Pengaruh perlakuan galur L. plantarum ke-i (i = 1, 2, 3, 4) βj = Pengaruh perlakuan jenis bakteri indikator ke-j (j = 1, 2, 3, 4, 5)
αβij = Pengaruh interaksi antara galur L. plantarum ke-i dengan jenis bakteri indikator ke-j
εijk = Galat percobaan pada ulangan ke-k pada kombinasi αi dengan βj dan interaksi αβij
Produksi Plantaricin
21 Yijk = µ + αi + εij
Keterangan:
Yijk = Peubah respon ke-k karena pengaruh bersama taraf ke-i faktor galur L. plantarum
pada ulangan ke-k (k = 1, 2, 3)
µ = Pengaruh rata-rata galur L. plantarum
αi = Pengaruh perlakuan galur L. plantarum ke-i (i = 1, 2, 3, 4)
εij = Galat percobaan dari nilai respon ke-j dari perlakuan pada taraf ke-i
Uji Stabilitas Aktivitas Plantaricin akibat Pemanasan
Faktor perlakuan pertama (α) adalah suhu pemanasan yang digunakan, terdiri atas dua taraf yaitu pemanasan pada suhu 80 °C (waterbath) selama 30 menit sebagai representasi suhu pasteurisasi dan pemanasan pada suhu 121 °C (pemanas ber-tekanan) selama 15 menit sebagai representasi suhu sterilisasi. Kontrol perlakuan adalah plantaricin yang belum diberikan perlakuan pemanasan. Faktor perlakuan yang kedua (β) adalah bakteriosin asal Lactobacillus plantarum dengan galur yang berbeda, yang terdiri atas empat galur yaitu Lactobacillus plantarum 2C12, 1B1, 2B2 dan 1A5.
Rancangan yang digunakan untuk menentukan ketahanan plantaricin akibat pemanasan terhadap bakteri indikator adalah rancangan acak lengkap faktorial dengan pola 3x5 dan tiga kali ulangan. Peubah yang diamati adalah diameter zona hambat hasil uji antagonistik dari plantaricin dengan perlakuan pemanasan yang berbeda (kontrol, suhu pasteurisasi 80 oC selama 30 menit dan suhu sterilisasi 121 oC selama 15 menit) dan bakteriosin asal Lactobacillus plantarum dengan galur yang berbeda. Rancangan tersebut dapat ditulis sebagai berikut:
Yijk = µ + Pi + Yj + PYij + εijk Keterangan:
Yijk = Peubah respon karena pengaruh bersama taraf ke-i faktor α (suhu pemanasan) dan taraf ke-j faktor β (jenis bakteriosin asal L. plantarum) pada ulangan ke-k (k = 1, 2, 3)
µ = Pengaruh rata-rata suhu pemanasan dan jenis bakteriosin asal L. plantarum
terhadap stabilitas plantaricin
αi = Pengaruh perlakuan suhu pemanasan ke-i (i = 1, 2, 3)
22 αβij = Pengaruh interaksi antara suhu pemanasan ke-i dengan jenis bakteriosin asal
L. plantarum ke-j
εijk = Galat percobaan pada ulangan ke-k pada kombinasi αi dengan βj dan interaksi αβij
HASIL DAN PEMBAHASAN
Uji Kemurnian Isolat Lactobacillus plantarum
dan Bakteri Indikator
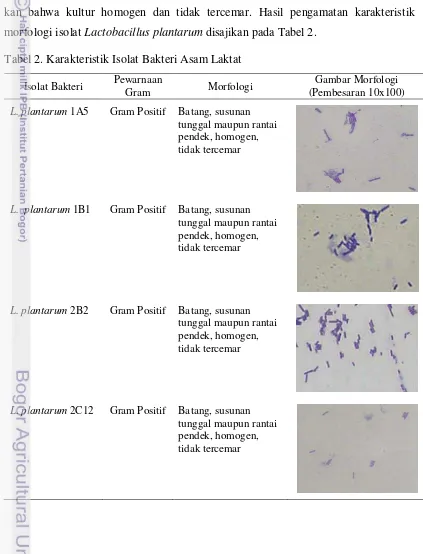
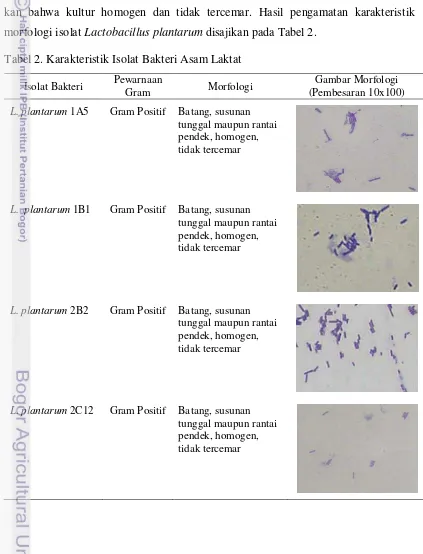
Hasil pengujian menunjukkan bahwa isolat Lactobacillus plantarum 1A5, 1B1, 2B2 dan 2C12 tergolong dalam Gram positif yang berbentuk batang dengan susunan tunggal maupun rantai pendek. Karakteristik morfologis tersebut menunjuk-kan bahwa kultur homogen dan tidak tercemar. Hasil pengamatan karakteristik morfologi isolat Lactobacillus plantarum disajikan pada Tabel 2.
Tabel 2. Karakteristik Isolat Bakteri Asam Laktat
Isolat Bakteri Pewarnaan
Gram Morfologi
Gambar Morfologi (Pembesaran 10x100)
L. plantarum 1A5 Gram Positif Batang, susunan tunggal maupun rantai pendek, homogen, tidak tercemar
L. plantarum 1B1 Gram Positif Batang, susunan tunggal maupun rantai pendek, homogen, tidak tercemar
L. plantarum 2B2 Gram Positif Batang, susunan tunggal maupun rantai pendek, homogen, tidak tercemar
24 Pewarnaan Gram dimaksudkan untuk mengetahui karakteristik morfologis dan kemurnian kultur bakteri yang digunakan. Hasil pengujian menunjukkan hasil yang tidak berbeda dengan hasil yang diperoleh pada penelitian Hidayati (2006), Permanasari (2008), dan Syahniar (2009) yang menyatakan bahwa isolat L. plantarum 1A5, 1B1, 2B2 dan 2C12 tergolong dalam kelompok bakteri genus
Lactobacillus sp. Karakteristik morfologis Lactobacillus sp. tergolong bakteri Gram positif yang mempunyai bentuk batang bervariasi dari panjang dan ramping sampai kokobasilus pendek. Pembentukan rantai umum dijumpai, terutama pada fase pertumbuhan logaritma lanjut (Frazier dan Westhoff, 1978; Pelczar dan Chan, 2005).
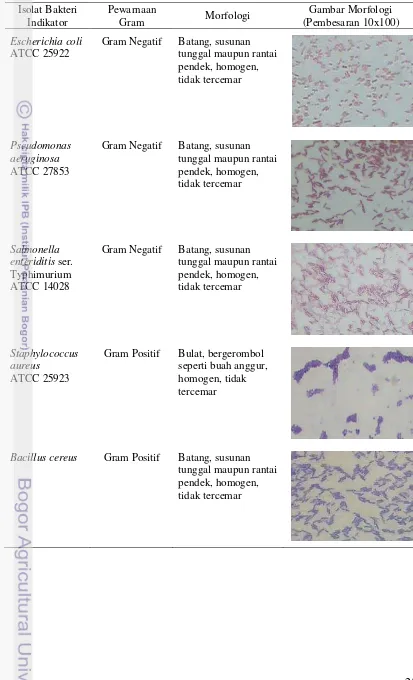
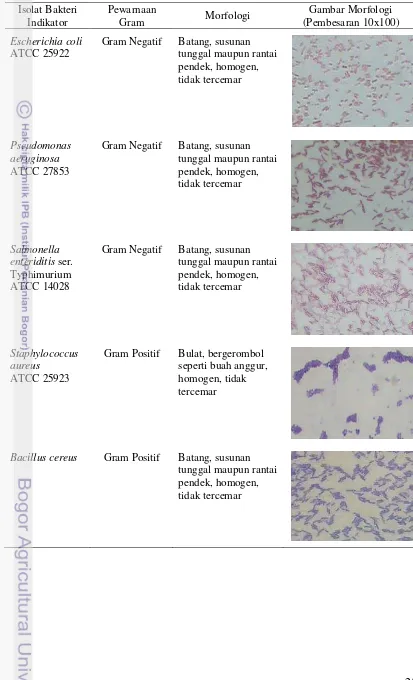
Karakteristik morfologis bakteri indikator yang digunakan, antara lain
Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853 dan
Salmonella enteriditis ser. Typhimurium ATCC 14028 yang berbentuk batang dengan susunan sel tunggal maupun rantai pendek dan tergolong bakteri Gram negatif. Hasil ini sesuai pernyataan Gaman dan Sherrington (1992) yakni Escherichia coli termasuk dalam famili Enterobacteriaceae yang termasuk gram negatif dan berbentuk batang yang fermentatif. Hasil ini juga sesuai Pelczar dan Chan (2005) yang menyatakan bahwa Pseudomonas merupakan bakteri yang berbentuk sel tunggal, batang lurus atau melengkung, namun tidak berbentuk heliks, pada umum-nya berukuran 0,5-1,0 x 1,5-4,0 µm. Perumum-nyataan Frazier dan Westhoff (1978) juga sesuai dengan hasil yang didapat yakni Salmonella merupakan bakteri berbentuk batang, Gram negatif, motil, anaerobik dan tidak membentuk spora.
Staphylococcus aureus ATCC 25923 mempunyai bentuk bulat bergerombol yang menyerupai buah anggursedangkan Bacillus cereus memiliki morfologi dengan bentuk batang dengan susunan tunggal maupun rantai pendek. Staphylococcus aureus ATCC 25923 dan Bacillus cereus tergolong bakteri Gram positif. Hal ini sesuai dengan penyataan Ray dan Bhunia (2008) yakni Staphylococcus aureus
25
Bacillus cereus Gram Positif Batang, susunan tunggal maupun rantai pendek, homogen, tidak tercemar
26 Identifikasi Keberadaan Bakteriosin
Hasil identifikasi keberadaan bakteriosin dengan uji antagonistik supernatan asal galur Lactobacillus plantarum menunjukkan bahwa galur-galur tersebut memproduksi bakteriosin yang memiliki daya hambat terhadap kelima bakteri indikator. Diameter zona hambat supernatan yang terbentuk pada uji antagonistik dapat dilihat pada Tabel 4.
Tabel 4. Diameter Zona Hambat Supernatan Bebas Sel (pH Netral) Lactobacillus
Keterangan: superskrip huruf kecil berbeda pada kolom yang sama menunjukkan berbeda nyata (P<0,05), superskrip huruf kapital berbeda pada baris yang sama menunjukkan berbeda sangat nyata (P<0,01), besarnya diameter zona hambat tidak termasuk diameter lubang sumur (5 mm).
Galur Lactobacillus plantarum memiliki pengaruh sangat nyata (P<0,01) terhadap diameter zona hambat. Pada hasil uji lanjut ditunjukkan bahwa supernatan asal galur Lactobacillus plantarum 2C12 menghasilkan diameter yang berbeda jika dibandingkan dengan supernatan asal galur lain. Diameter zona hambat supernatan juga dipengaruhi oleh jenis bakteri indikator (P<0,05). Aktivitas hambatan
Escherichia coli ATCC 25922 berbeda nyata dengan Bacillus cereus (P<0,05). Aktivitas hambatan Escherichia coli ATCC 25922 tidak berbeda nyata (P>0,05) dengan Staphylococcus aureus ATCC 25923, Salmonella enteriditis ser. Typhimurium ATCC 14028, dan Pseudomonas aeruginosa ATCC 27853.
Pengamatan ini menunjukkan bahwa supernatan asal empat galur
27 pernyataan Smaoui et al. (2009) bahwa bakteriosin yang dihasilkan oleh
Lactobacillus plantarum sp. TN635 dapat menghambat pertumbuhan bakteri Gram negatif (Salmonella enterica ATCC43972, Pseudomonas aeruginosa ATCC 49189,
Hafnia sp. and Serratia sp.) dan Candida tropicalis R2 CIP203 yang termasuk jamur
(fungi) patogen.
Produksi dan Purifikasi Plantaricin dengan Menggunakan Presipitasi Parsial
Supernatan yang telah dihasilkan berada pada kondisi asam. Kondisi asam tersebut disebabkan adanya asam-asam organik yang terbentuk sebagai metabolit primer dari bakteri asam laktat dan mempunyai aktivitas antimikrob (Schved et al., 1992). Keberadaan asam organik dalam supernatan dapat menutupi aktivitas bakteriosin yang terbentuk. Penetralan pH supernatan diperlukan untuk meng-optimalkan bakteriosin dalam menghambat bakteri indikator pada uji antagonistik. Kondisi pH supernatan yang diberi kultur Lactobacillus plantarum yang berbeda dapat dilihat pada Tabel 5.
Tabel 5. Kondisi pH Supernatan Lactobacillus plantarum
Galur L. plantarum pH Supernatan Bebas Sel pH Supernatan Netral
1A5 4,01 ± 0,04 6,11 ± 0,34
1B1 3,94 ± 0,11 5,87 ± 0,12
2B2 4,00 ± 0,02 6,17 ± 0,31
2C12 3,98 ± 0,01 6,04 ± 0,16
28 Hasil kuantitatif protein presipitat bakteriosin hasil presipitasi parsial dan
plantaricin kasar hasil dialisis dapat dilihat pada Gambar 2. Secara deskriptif, hasil kuantitatif ini menunjukkan bahwa rataan konsentrasi protein yang dihasilkan oleh galur Lactobacillus plantarum 1A5, 1B1 dan 1A5 merupakan nilai yang tinggi jika dibandingkan dengan galur Lactobacillus plantarum 2C12. Rataan konsentrasi protein presipitat bakteriosin dan plantaricin kasar dari yang terbesar berturut-turut adalah galur Lactobacillus plantarum 1B1, 1A5, 2B2 dan 2C12.
Gambar 3. Konsentrasi Protein Presipitat Bakteriosin Hasil Presipitasi Parsial dan
Plantaricin Kasar Hasil Dialisis Asal Empat Galur L. plantarum (mg/ml). = Presipitat Bakteriosin; = Plantaricin Kasar.
Nilai rataan konsentrasi protein plantaricin kasar lebih tinggi daripada nilai konsentrasi protein presipitat bakteriosin pada semua galur Lactobacillus plantarum
kecuali galur Lactobacillus plantarum 2C12. Presipitat bakteriosin merupakan hasil dari purifikasi parsial dengan konsentrasi yang tinggi namun masih mengandung garam mineral yang digunakan untuk mengendapkan protein. Proses dialisis atau proses pencucian yang bertujuan untuk menghilangkan pengotor pada permukaan partikel-partikel protein sehingga dapat menghasilkan plantaricin kasar dengan konsentrasi yang lebih tinggi dari presipitat bakteriosin (Day dan Underwood, 2002). Parente dan Ricciardi (1999) menambahkan bahwa beberapa faktor yang mempenga-ruhi produksi bakteriosin antara lain: (i) galur yang memproduksi, (ii) media (karbo-hidrat dan sumber nitrogen dan kation) dan (iii) kondisi fermentasi (pH dan suhu).
29 Stabilitas Protein Plantaricin Setelah Pemanasan
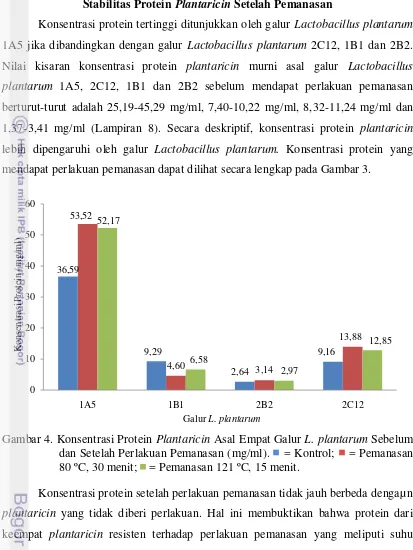
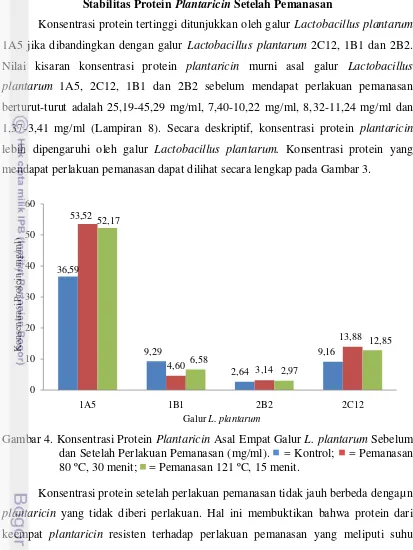
Konsentrasi protein tertinggi ditunjukkan oleh galur Lactobacillus plantarum
1A5 jika dibandingkan dengan galur Lactobacillus plantarum 2C12, 1B1 dan 2B2. Nilai kisaran konsentrasi protein plantaricin murni asal galur Lactobacillus plantarum 1A5, 2C12, 1B1 dan 2B2 sebelum mendapat perlakuan pemanasan berturut-turut adalah 25,19-45,29 mg/ml, 7,40-10,22 mg/ml, 8,32-11,24 mg/ml dan 1,37-3,41 mg/ml (Lampiran 8). Secara deskriptif, konsentrasi protein plantaricin
lebih dipengaruhi oleh galur Lactobacillus plantarum. Konsentrasi protein yang mendapat perlakuan pemanasan dapat dilihat secara lengkap pada Gambar 3.
Gambar 4. Konsentrasi Protein Plantaricin Asal Empat Galur L. plantarum Sebelum dan Setelah Perlakuan Pemanasan (mg/ml). = Kontrol; = Pemanasan 80 ºC, 30 menit; = Pemanasan 121 ºC, 15 menit.
Konsentrasi protein setelah perlakuan pemanasan tidak jauh berbeda dengaµ n
plantaricin yang tidak diberi perlakuan. Hal ini membuktikan bahwa protein dari keempat plantaricin resisten terhadap perlakuan pemanasan yang meliputi suhu pasteurisasi dan suhu sterilisasi. Walker (2000) menambahkan bahwa struktur tersier protein khususnya interaksi hidrofobik, ikatan hidrogen dan jembatan disulfida dapat melindungi protein terdenaturasi akibat perlakuan seperti pemanasan. Hal ini diperkuat dengan penelitian sebelumnya yang dilakukan oleh Hata et al. (2010) yang melaporkan bahwa plantaricin ASM1 menunjukkan stabilitas pada pemanasan jika
30 dibandingkan dengan nisin A. Joshi et al. (2006) menambahkan bahwa kriteria kesta-bilan panas pada bakteriosin tergantung pada tahap pemurnian, pH, keadaan medium.
Uji Kemampuan Antagonistik Plantaricin Setelah Pemanasan pada Berbagai Bakteri Indikator
Staphylococcus aureusATCC 25923
Stabilitas aktivitas penghambatan plantaricin yang mendapatkan perlakuan pemanasan berbeda terhadap bakteri indikator dapat dilihat pada Tabel 6. Hasil pengamatan pada penelitian ini menunjukkan bahwa tidak terlihat interaksi antara perlakuan pemanasan dan galur Lactobacillus plantarum (P>0,05) terhadap diameter zona hambat pada Staphylococcus aureus ATCC 25923. Galur Lactobacillus plantarum memiliki pengaruh nyata (P<0,05) terhadap diameter zona hambat. Zona penghambatan terbesar dihasilkan oleh galur 2C12 sebesar 5,34 mm, sedangkan zona penghambatan terkecil dihasilkan oleh galur 1B1 yaitu 3,39 mm.
Tabel 6. Diameter Zona Hambat Plantaricin Asal Empat Galur Lactobacillus plantarum dengan Perlakuan Pemanasan terhadap Staphylococcus aureus
ATCC 25923
Keterangan: superskrip berbeda pada kolom yang sama menunjukkan berbeda nyata (P<0,05), besarnya diameter zona hambat tidak termasuk diameter lubang sumur (5 mm).
Plantaricin yang dihasilkan dari keempat galur Lactobacillus plantarum
mempunyai sifat stabil pada pemanasan ditunjukkan oleh kemampuannya memper-tahankan aktivitas penghambatan terhadap bakteri Staphylococcus aureus ATCC 25923 setelah dipanaskan baik pada suhu pasteurisasi maupun sterilisasi. Hal ini sesuai dengan pernyataan Hata et al. (2010) menyatakan bahwa stabilitas plantaricin
menunjuk-31 kan zona hambat hampir sama dibandingkan dengan bakteriosin yang tidak dipanas-kan. Namun hasil ini berbeda dengan Villani et al. (2001) yang melaporkan bahwa aktivitas antimikrob benar-benar hilang setelah inkubasi pada 121 °C selama 15 menit.
PseudomonasaeruginosaATCC 27853
Stabilitas aktivitas penghambatan plantaricin akibat pemanasan terhadap
Pseudomonas aeruginosa ATCC 27853 dapat dilihat secara lengkap pada Tabel 7. Analisis ragam menunjukkan bahwa aktivitas penghambatan plantaricin terhadap
Pseudomonas aeruginosa ATCC 27853 tidak dipengaruhi oleh interaksi (P>0,05) antara galur Lactobacillus plantarum dan perlakuan pemanasan yang berbeda. Tabel 7. Diameter Zona Hambat Plantaricin Asal Empat Galur Lactobacillus
plantarum dengan Perlakuan Pemanasan terhadap Pseudomonas aeruginosa ATCC 27853
Keterangan: superskrip berbeda pada kolom yang sama menunjukkan berbeda sangat nyata (P<0,01), besarnya diameter zona hambat tidak termasuk diameter lubang sumur (5 mm).
Stabilitas aktivitas plantaricin terhadap Pseudomonas aeruginosa ATCC 27853 hanya dipengaruhi oleh galur Lactobacillus plantarum secara sangat nyata (P<0,01) yang dapat ditunjukkan oleh rataan diameter zona hambat terhadap
Pseudomonas aeruginosa ATCC 27853. Pada hasil uji lanjut dapat dilihat bahwa galur Lactobacillus plantarum 2C12 memberikan hasil yang berbeda sangat nyata pada aktivitas hambatan terhadap Pseudomonas aeruginosa ATCC 27853 dengan galur Lactobacillus plantarum 1B1, 2B2 dan 1A5 (P<0,01). Namun, galur
32 galur Lactobacillus plantarum dan tidak dipengaruhi oleh perlakuan pemanasan. Hal ini ditunjukkan oleh tidak adanya interaksi antara kedua perlakuan tersebut.
Escherichia coliATCC 25922
Stabilitas aktivitas penghambatan plantaricin yang mendapat perlakuan pemanasan berbeda terhadap bakteri indikator Escherichia coli ATCC 25922 dapat dilihat pada Tabel 8. Stabilitas aktivitas plantaricin terhadap Escherichia coli ATCC 25922 tidak dipengaruhi oleh interaksi (P>0,05) antara galur Lactobacillus plantarum dan perlakuan pemanasan yang berbeda.
Tabel 8. Diameter Zona Hambat Plantaricin Asal Empat Galur Lactobacillus plantarum terhadap Escherichia coli ATCC 25922
Galur
Keterangan: superskrip berbeda pada kolom yang sama menunjukkan berbeda nyata (P<0,05), besarnya diameter zona hambat tidak termasuk diameter lubang sumur (5 mm).
Stabilitas aktivitas plantaricin dipengaruhi secara nyata oleh galur
Lactobacillus plantarum terhadap diameter zona hambat pada Escherichia coli
ATCC 25922 (P<0,05). Perlakuan panas tidak berpengaruh terhadap diameter zona hambat (P>0,05). Aktivitas hambatan galur Lactobacillus plantarum 2C12 berbeda nyata dengan galur Lactobacillus plantarum 1B1 (P<0,05). Galur Lactobacillus plantarum 2C12 menunjukkan aktivitas hambatan yang tidak berbeda nyata dengan galur Lactobacillus plantarum 2B2 dan 1A5 (P>0,05). Aktivitas hambatan galur
Lactobacillus plantarum 2B2 dan 1A5 juga tidak berbeda nyata dengan galur
Lactobacillus plantarum 1B1.
33 kasus keracunan makanan (Gaman dan Sherrington, 1992). Aktivitas penghambatan
plantaricin terhadap Escherichia coli ATCC 25922 dapat dilihat pada Gambar 4.
Gambar 5. Zona Hambat Plantaricin Asal L. plantarum 2B2 Setelah Pemanasan pada Suhu 121 °C selama 15 menit terhadap E. coli ATCC 25922. Lingkaran pada Gambar Menunjukkan Diameter Zona Hambat. A= Kontrol (Plantaricin yang Tidak Diberi Perlakuan Pemanasan); B=
Plantaricin Setelah Perlakuan Pemanasan.
Salmonella enteriditis ser. Typhimurium ATCC 14028
Hasil pengamatan stabilitas aktivitas plantaricin setelah mendapat perlakuan pemanasan terhadap bakteri indikator Salmonella enteriditis ser. Typhimurium ATCC 14028 dapat dilihat secara lengkap pada Tabel 9. Stabilitas aktivitas
plantaricin terhadap Salmonella enteriditis ser. Typhimurium ATCC 14028 tidak dipengaruhi oleh interaksi (P>0,05) antara galur Lactobacillus plantarum dan perlakuan pemanasan yang berbeda. Hasil analisis ragam menunjukkan bahwa galur
Lactobacillus plantarum yang berbeda mempengaruhi stabilitas aktivitas plantaricin
terhadap Salmonella enteriditis ser. Typhimurium ATCC 14028.
Tabel 9. Diameter Zona Hambat Plantaricin Asal Empat Galur Lactobacillus plantarum dengan Perlakuan Pemanasan terhadap Salmonella enteriditis
ser. Typhimurium ATCC 14028
Keterangan: superskrip berbeda pada kolom yang sama menunjukkan berbeda nyata (P<0,05), besarnya diameter zona hambat tidak termasuk diameter lubang sumur (5 mm).