ISOLASI DAN EFEKTIVITAS PROMOTER
-AKTIN DALAM
MENGARAHKAN EKSPRESI GEN TARGET PADA
TRANSGENESIS IKAN MAS Cyprinus carpio
ANDI ALIAH HIDAYANI
C151060011
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
I. PENDAHULUAN
1.1 Latar Belakang
Efektivitas ekspresi gen dalam transgenesis ditentukan oleh promoter yang diligasi pada posisi hulu (upstream, terminal 5’) dari gen target dalam suatu
konstruksi. Promoter merupakan suatu sekuen DNA yang menginisiasi, mengatur letak, waktu dan tingkat ekspresi gen (Toha, 2001; Beaumont & Hoare, 2003). Pada awal perkembangan transgenesis ikan, peneliti menggunakan konstruksi gen dengan promoter yang berasal dari mamalia dan virus, tetapi dengan alasan keamanan pangan, maka dikembangkanlah konstruksi gen dengan promoter yang berasal dari ikan, atau disebut sebagai konstruksi gen all-fish. Lebih baik lagi bila
menggunakan promoter dari ikan yang sama (homolog) atau hubungan kekerabatannya dekat dengan ikan transgenik (Hwang et al., 2003).
Beberapa jenis promoter yang telah diisolasi dan diuji pada beberapa spesies ikan oleh para peneliti, yaitu promoter cytomegalovirus (CMV) dari virus manusia, elongation factor-1α (EF-1α) dari ikan medaka, β-aktin dari ikan medaka dan myosin light chain-2 (Mylz-2) dari ikan zebra (Alimuddin, 2003). Berdasarkan penelitian Alimuddin (2003) pada ikan zebra, promoter β-aktin dan Mylz-2 menunjukkan aktivitas paling kuat dibandingkan EF-1α, sedangkan CMV menunjukkan aktivitas paling rendah. Hal ini dikarenakan promoter CMV berasal dari virus manusia dimana ada kemungkinan bahwa tidak semua elemen
cis-acting-nya dikenali oleh faktor trans-acting ikan zebra, sedangkan promoter
lainnya yang berasal dari ikan menunjukkan aktivitas yang tinggi. Selanjutnya β -aktin merupakan promoter yang bersifat house-keeping; selalu aktif sepanjang
hidup organisme. Selain itu, promoter β-aktin juga mempunyai sifat ubiquitous
(Hackett, 1993), yaitu promoter ini akan aktif dimana-mana, dan constitutive
(Volckaert et al., 1994) yang berarti bahwa promoter ini dapat aktif tanpa
diberikan rangsangan dari luar seperti suhu dan hormon. Promoter ini sering digunakan dalam penelitian transgenesis seperti pada ikan mas (Liu et al., 1990),
mud loach (Nam et al., 2001), ikan nila (Hwang et al., 2003) dan ikan kakap merah (Kato et al., 2007).
Berdasarkan hasil penelitian, dikemukakan bahwa promoter heterolog (promoter yang berasal dari ikan yang berbeda dengan ikan uji) memiliki efektivitas yang berbeda dengan promoter homolog (promoter yang berasal dari ikan yang sama dengan ikan uji). Seperti pada ikan zebra menggunakan promoter β-aktin dari ikan kakap merah menunjukkan tingkat kelangsungan hidup hanya 63% dibandingkan dengan penggunaan promoter ini pada spesies yang sama tingkat kelangsungan hidupnya mencapai 95% (Kato et al., 2007). Oleh karena itu
dalam rangka menghasilkan promoter homolog, maka pada penelitian ini dilakukan isolasi promoter -aktin yang berasal dari ikan mas. Ikan mas merupakan salah satu jenis ikan air tawar yang memiliki nilai ekonomis tinggi.
Efektivitas promoter dapat dilihat jika promoter ini disambungkan dengan gen target. Gen target yang digunakan dalam penelitian ini adalah Green Fluorescent Protein (GFP), dimana gen ini juga berperan sebagai gen penanda
(marker) karena memiliki kelebihan dalam pendeteksiannya yaitu tidak membutuhkan penambahan substrat dan ekspresinya dapat divisualisasikan dalam sel hidup di atas pemaparan cahaya UV dengan menggunakan mikroskop fluoresen atau secara kuantitatif dengan menggunakan PCR (Iyengar et al., 1996). Untuk menguji efektivitas promoter yang digunakan dalam penelitian ini maka perlu adanya promoter pembanding. Dengan menggunakan promoter β-aktin dari ikan mas (Cyprinus carpio) sebagai promoter homolog, ikan nila (Oreochromis niloticus), dan ikan medaka (Oryzias latipes) sebagai promoter heterolog yang
dicobakan pada ikan mas, diharapkan model ini dapat diaplikasikan pada ikan-ikan budidaya lainnya sehingga proses transgenesis akan lebih optimal.
1.2 Tujuan Penelitian
Tujuan penelitian ini adalah :
1. Mengisolasi promoter β-aktin dari ikan mas
2. Menguji efektivitas promoter β-aktin homolog dan heterolog pada ikan mas (Cyprinus carpio) dengan menggunakan gen target Green Fluorescent Protein
1.3 Hipotesis
Hipotesis yang diajukan adalah promoter homolog memiliki efektivitas yang berbeda dengan promoter heterolog dalam mengarahkan ekspresi gen target.
1.4 Manfaat Penelitian
Manfaat dari penelitian ini adalah :
1. Menyediakan promoter yang mendukung ekspresi gen target dalam transgenik ikan mas (Cyprinus carpio).
II. TINJAUAN PUSTAKA
2.1 Transgenik
Salah satu pendekatan untuk perbaikan genetik organisme akuatik yang muncul sebagai suatu disiplin ilmu tersendiri baru-baru ini adalah transgenesis, yaitu proses transfer gen-gen asing ke inang yang baru (Lutz, 2001). Teknik ini digunakan untuk mengintroduksikan karakter-karakter genetik yang baru atau over-ekspresi ke suatu individu melalui introduksi gen asing yang disambungkan ke genom inang dan diharapkan dapat diwariskan ke keturunannya melalui sel gonad (Yoshizaki, 2001). Prinsip utama dari teknik transfer gen adalah memasukkan DNA asing ke dalam nukleus suatu sel target dan menggabungkannya ke genom inang. Aplikasi teknologi ini sendiri dapat meningkatkan kualitas ikan, seperti pertumbuhan, resistensi terhadap penyakit dan adaptasi terhadap lingkungan yang ekstrim (Sarmasik, 2002).
Teknik transgenesis pertama kali dilaporkan sukses pada ikan koki (Carassius auratus) di Cina pada pertengahan tahun 1980-an (Hulata, 2001). Sejak saat itu, eksperimen mengenai transgenik telah dilakukan pada berbagai laboratorium di seluruh dunia pada 35 spesies ikan yang berbeda (Anderson, 2004). Selanjutnya akhir abad ke-19 teknik ini berkembang pada strain-strain ikan yang bernilai komersial dan difokuskan pada peningkatan pertumbuhan. Kemudian cDNA hormon pertumbuhan dan sekuen-sekuen genom lebih dari 40 ikan telah diisolasi, dikarakterisasi dan digunakan untuk pembuatan konstruksi gen ”all fish” untuk transformasi pada spesies ikan lain (Teufel et al., 2002).
konstruksi DNA ini ternyata menimbulkan beberapa masalah misalnya hal yang tidak dapat diterima untuk ikan-ikan yang dikonsumsi manusia dan meningkatnya resiko ekologi (Rajesh & Majumdar, 2006). Konstruksi DNA yang terdiri dari sekuen-sekuen ”all-piscine” telah dilaporkan oleh beberapa peneliti. Salah satunya penelitian yang dilakukan oleh Du et al. (1992) dalam Rahman & Maclean (1999), yang menggunakan suatu konstruksi gen (opAFPcsGH) yang mengandung gen GH salmon chinook (Oncorhynchus tschawytscha) yang
dikontrol oleh sekuen promoter protein antibeku dari ikan ocean pout
(Macrozoarces americanus) untuk menghasilkan transgenik salmon Atlantik
(Salmo salar). Peningkatan pertumbuhan ikan salmon transgenik tersebut
dilaporkan 2 sampai 6 kali lipat melebihi kontrol. Contoh lainnya, ikan hias strain baru yaitu ikan zebra memiliki warna tertentu yang dapat terlihat pada kondisi cahaya biasa dengan mengintroduksikan gen GFP (green fluorescent protein),
YFP (yellow fluorescent protein) dan RFP (red fluorescent protein) (Gong et al., 2002).
Ikan dipertimbangkan sebagai kandidat yang paling baik untuk pemasaran hewan transgenik pertama untuk konsumsi manusia (Zbikowska, 2003). Hal ini disebabkan karena ikan memiliki banyak keuntungan dibandingkan dengan hewan lain seperti mamalia, yaitu fekunditas yang tinggi, pembuahannya eksternal dan perkembangan embrio yang dapat diamati dengan mudah. Selain itu proses embriogenesis cepat melalui pembentukan dan tahap morfologi yang jelas karena embrio transparan (Winkler et al., 1990; Iyengar et al., 1996). Penggunaan ikan khususnya dalam penelitian juga dapat menurunkan eksploitasi mamalia secara signifikan, mengurangi biaya dan waktu (Zbikowska, 2003).
2.2 Teknik Mikroinjeksi
studi transfer gen. Contohnya mikroinjeksi dan elektroporasi saat ini digunakan untuk transfer gen pada ikan yang bertelur, sedangkan teknik lainnya seperti transfer gen mediasi vektor retroviral digunakan pada ikan yang melahirkan anak (tanpa bertelur) (Sarmasik, 2002).
Mikroinjeksi merupakan teknik yang dipertimbangkan paling efektif dalam transfer gen pada vertebrata tingkat tinggi seperti tikus dan domba. Dalam teknik ini, transgen secara langsung dimikroinjeksikan ke dalam pronukleus telur-telur yang telah dibuahi (Sarmasik, 2002). Aplikasi mikroinjeksi pada ikan sendiri memiliki beberapa kelemahan seperti nukleus dari telur yang telah dibuahi sangat sulit diidentifikasi di bawah mikroskop karena nukleus telur kecil dan volume sitoplasma yang besar, korion telur yang sangat keras seperti telur pada ikan salmon yang menyebabkan telur sulit ditembus oleh jarum mikroinjeksi (Yoshizaki, 2001). Untuk mengatasi masalah-masalah ini, beberapa peneliti membuat ikan transgenik dengan cara penyuntikan gen dengan jumlah copy yang banyak ke dalam sitoplasma telur yang telah dibuahi sebagai alternatif penyuntikan ke inti telur (Alimuddin et al., 2003). Untuk mengatasi masalah korion telur yang keras, korion dapat dibuang dengan menggunakan proteinase dan selanjutnya telur dapat disuntik dengan mudah (Ueno et al., 1994 dalam
Alimuddin et al., 2003). Selain itu, perkembangan embrio yang cepat setelah pembuahan dan sel tunggal telur yang telah dibuahi dalam beberapa jam berubah menjadi embrio multiseluler. Ini mengimplikasikan bahwa jika injeksi transgen tidak dilakukan secepatnya dalam beberapa jam pertama, walaupun injeksi itu berhasil akan menghasilkan ikan dewasa yang mosaik untuk transgen (Rahman & Maclean, 1991). Umumnya transgen yang diinjeksikan pada tahap 1 sel dapat terintegrasi ke dalam genom semua sel. Akan tetapi integrasi ini seringkali hanya ada setelah beberapa kali pembelahan sel yang menghasilkan embrio yang tidak membawa transgen atau disebut mosaik (Beaumont & Hoare, 2003).
2.3 Promoter
Promoter merupakan sekuen DNA yang terletak upstream (terminal 5’)
2003). Menurut Glick dan Pasternak (2003), suatu promoter yang kuat merupakan promoter yang memiliki aktivitas yang tinggi terhadap RNA polimerase yang mengakibatkan daerah yang berbatasan downstream dicetak
secara teratur. Promoter inilah yang menjadi kekuatan gen untuk mengekspresikan ciri-cirinya pada tingkat yang sangat tinggi dan juga potensial dalam mempengaruhi gen yang lain dalam suatu organisme (Anderson, 2004).
Promoter pertama yang digunakan dalam bioteknologi ikan diisolasi dari genom virus (misalnya Rous Sarcoma Virus (RSV), Simian Virus (SV40) atau Cytomegalovirus (CMV)), mamalia (misalnya mouse metallothionein-1 (mMT-1)), burung (misalnya promoter β-aktin dari ayam) atau katak Xenopus laevis (1α
-enhanced promoter). Tingkat ekspresi gen yang dihasilkan ternyata rendah sehingga pencarian promoter-promoter ikan yang lebih efektif tetap dilakukan secara intensif selama 15 tahun terakhir (Teufel et al., 2002).
Promoter β-aktin dari berbagai spesies ikan telah dilaporkan sebagai promoter ubiquitous yang efisien dalam penelitian ikan transgenik. Promoter β -aktin dari ikan medaka telah diisolasi dan dengan menggunakan gen reporter lacZ menunjukkan ekspresi yang kuat pada ikan medaka, promoter β-aktin dari ikan zebra juga menunjukkan hal yang sama setelah diinjeksikan pada ikan zebra (Hwang et al., 2003). Kedua promoter di atas disebut sebagai promoter homolog yaitu promoter yang menggunakan konstruksi gen homolog spesies yaitu donor dan penerima berasal dari spesies yang sama, sedangkan promoter heterolog adalah promoter yang menggunakan konstruksi gen heterospesies yaitu donor dan penerima donor berasal dari spesies yang berbeda dengan ikan transgenik yang dihasilkan (Rajesh & Majumdar, 2006). Berdasarkan beberapa penelitian penggunaan promoter homolog menghasilkan ekspresi gen lebih baik dibandingkan dengan promoter heterolog. Seperti pada ikan zebra menggunakan promoter β-aktin dari ikan kakap merah menunjukkan tingkat kelangsungan hidup hanya 63% dibandingkan dengan penggunaan promoter ini pada spesies yang sama tingkat kelangsungan hidupnya mencapai 95% (Kato et al., 2007). Menurut
Palmiter et al. (1982) dalam Nam et al. (2008), bahwa suatu promoter yang
promoter heterolog secara potensial mengakibatkan regulasi ”feedback negative”. Namun, beberapa promoter β-aktin heterolog yang telah digunakan dalam penelitian transgenesis dapat menghasilkan ekspresi gen yang baik pada ikan uji seperti penelitian yang dilakukan oleh Yoshizaki (2001) dengan menggunakan β -aktin dari ikan medaka ternyata mampu mengekspresikan gen GFP yang kuat pada ikan rainbow trout.
Faktor-faktor transkripsi yang mempengaruhi aktivitas promoter β-aktin adalah boks TATA, boks CCAAT dan CC(A/T)6GG atau motif CArG. Motif CArG berperan dalam pengaturan ekspresi transgen (Takagi et al., 1994).
Menurut Liu et al. (1990), motif CArG berada pada 2 tempat, yang pertama ada di
antara boks CCAAT dan boks TATA, dan yang kedua berada pada intron 1. Motif CArG ysng terletak pada intron 1 berfungsi sebagai peningkat (enhancer) dalam
aktivitas transkripsi. Boks TATA merupakan elemen yang umum dijumpai pada sekuens promoter yang berperan sebagai tempat melekatnya (binding) RNA polimerase pada saat transkripsi RNA akan berlangsung (Glick & Pasternak, 2003). Secara in vitro penghapusan boks TATA membuat promoter tidak aktif dan pada in vivo aktivitasnya menurun (Quitschke et al., 1989). Selanjutnya dikatakan aktivitas promoter β-aktin tergantung pada keberadaan elemen CCAAT, dengan adanya elemen ini tingkat aktivitas tertinggi promoter β-aktin tercapai (Quitschket et al., 1989).
Pembuatan konstruksi gen memerlukan promoter dengan elemen CCAAT, boks TATA, motif CArG, ekson 1 dan intron 1 (Higashijima et al., 1997). Dalam proses transkripsi mula-mula pra-mRNA yang besar disintesis, disebut heterogenous nuclear RNA (hnRNA) yang di dalamnya terkandung bagian intron yaitu ruas-ruas gen yang akan hilang pada mRNA fungsional (mature) dan ekson
yaitu ruas-ruas gen yang ditranskripsikan ke dalam mRNA fungsional (mature) dan akan ditranslasi menjadi protein (Jusuf, 2001). Berdasarkan penelitian yang dilakukan oleh Hwang et al. (2003), dari penderetan, ukuran dan variasi sekuen
dibandingkan dengan ekson 1 persentase homologi sekuen genom β-aktin ikan nila adalah 86,8% dengan ikan medaka dan 74,1% dengan ikan mas. Hal ini menunjukkan bahwa promoter β-aktin dari ikan nila memiliki homologi yang lebih tinggi pada ikan medaka dibandingkan dengan ikan mas.
2.4 Gen Target GFP (Green Fluorescent Protein)
Promoter dikatakan efektif apabila gen target yang disambungkan ke promoter dapat terekspresi dengan level yang tinggi. Menurut Iyengar et al.
(1996), gen penanda yang biasa digunakan dalam pengujian aktivitas promoter yaitu gen chloramphenicol acetyl transferase (CAT), neomycin phosphotransferase (NEO), -galactosidase (lacZ), luciferase, green fluorescent protein (GFP), tyrosinase, dan melanin concentrating hormone. Gen GFP merupakan gen yang mengkodekan protein yang berpendar hijau dan dapat divisualisasikan ekspresinya pada sel hidup dengan menggunakan sinar ultraviolet (Chalfie, 1994 dalam Iyengar et al., 1996). Selain itu, gen ini tidak memerlukan
substrat tambahan untuk ekspresinya (Chalfie, 1994 dalam Iyengar et al., 1996). Berdasarkan penelitian Yoshizaki et al. (2000) dalam Dunham (2004), GFP pertama kali dapat diamati dengan menggunakan promoter rainbow trout vasa-like gene pada tahap blastula, tetapi ekspresinya pada sel spesifik tidak terdeteksi. Pada tahap pembentukan bintik mata, kira-kira 30% embrio transgenik yang dapat mengekspresikan GFP dalam sel-sel bakal gonad dan meningkat hingga 70% setelah penetasan. Sel-sel yang mengekspresikan GFP terletak pada daerah genital. Selain itu ekspresi GFP dengan menggunakan promoter vasa
medaka dapat juga terdeteksi pada daerah ventrolateral usus halus pada tahap sirkulasi darah, tetapi setelah penetasan pindah ke daerah gonadal (Kinoshita & Tanaka, 2002 dalam Dunham, 2004).
Gen GFP awalnya diisolasi dari ubur-ubur (Aequorea victoria) yang
memancarkan cahaya hijau berpendar dengan kuat dan stabil. Perkembangan saat ini, GFP telah dimutasi dan pendarannya menjadi lebih kuat, yaitu enhanced GFP
III. METODE PENELITIAN
3.1 Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan mulai bulan November 2007 hingga Juli 2009, bertempat di Laboratorium Reproduksi dan Genetika Organisme Akuatik Departemen Budidaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor serta di Balai Besar Pengembangan Budidaya Air Tawar (BBPBAT) Sukabumi.
3.2 Prosedur Kerja
3.2.1 Isolasi Promoter β-Aktin dari Ikan Mas
Proses PCR awal menggunakan cetakan DNA yang berasal dari genom ikan mas, sepasang primer degenerated berdasarkan database yang ada di Bank Gen, yaitu primer forward F-BP1 (5’-GTGWGTGACGCYGGACCAATC-3’) dan primer reverse R-BP1 (5’-TAGAAGGTGTG-RTGCCAGATCTTC-3’), dimana W = A + T, R = A + G, Y = C + T. Hasil PCR awal diamplifikasi kembali dengan primer FBP1 dan RBP2 (5’-TTGCACATRCCRGAKCCGTTGTC-3’), dimanaK = G + T. Produk PCR dielektroforesis menggunakan gel agarosa 0,7%.
Fragmen DNA dari hasil elektroforesis dipotong dan kemudian diisolasi menggunakan Gene Mate Purification Kit (ISC BioExpress). Hasil purifikasi dari gel diligasi dengan vektor kloning pGEM-T Easy (Gambar 1) lalu ditransformasi dengan sel kompeten E.coli DH-5α.
Seleksi koloni bakteri yang membawa plasmid pGEM-Teasy yang mengandung promoter β-aktin (pT-ccBA) dilakukan dengan metode cracking,
kemudian plasmid tersebut diisolasi menggunakan kit EZ-10 Spin Column Plasmid DNA Minipreps (Bio Basic Inc). Fragmen DNA yang ada di dalam
plasmid disekuensing dengan mesin ABI PRISM 3100-Avant Genetic Analyzer
Gambar 1. Peta vektor kloning pGEM-T Easy (Promega, WI, USA)
.
Gambar 2. Mesin ABI PRISM 3100-Avant Genetic Analyzer
3.2.2 Pembuatan Konstruksi pccBA-EGFP
Plasmid pEGFP-N1 (BD Biosciences Clontech) yang mengandung gen pengkode protein yang berpendar hijau hasil mutasi dan dikenal dengan nama EGFP (enhanced green fluorescent protein) dipotong menggunakan enzim restriksi Kpn I dan Apa I. Untuk pengujian aktivitas promoter β-aktin ikan mas,
sekuens DNA yang digunakan mencakup promoter proksimal (sekuens sebelum ekson 1), ekson 1 dan intron 1. Fragmen DNA promoter tersebut diamplifikasi menggunakan PCR dengan primer forward (5’ -TTGGTACCGTGACGCCG-GACCAATC-3’) dan reverse (5’-TTGGGCCCAAGTACCAAAAAGCATG-3’). Nukleotida yang digarisbawahi dalam sekuens primer masing-masing merupakan situs restriksi enzim Kpn I dan Apa I. Hasil amplifikasi diligasi dengan vektor
kloning pGEM-T Easy dan selanjutnya fragmen ccBA tersebut diisolasi kembali dengan cara memotong plasmid menggunakan enzim restriksi Kpn I dan Apa I. Fragmen ccBA hasil restriksi diligasi dengan fragmen EGFP-N1 yang telah dipotong untuk membuat konstruksi gen pccBA-EGFP. Selanjutnya konstruksi gen ini ditransformasi, koloni bakteri diseleksi dan plasmid diisolasi dengan mengikuti metode sebelumnya (metode 3.2.1). Untuk pengecekan kesempurnaan konstruksi pccBA-EGFP yang telah dibuat, sebagian plasmid dielektroforesis, sedangkan plasmid yang lainnya siap digunakan untuk proses mikroinjeksi.
3.2.3 Pengujian Efektivitas Promoter 3.2.3.1 Pengujian Secara Kualitatif
Tiga jenis konstruksi gen digunakan dalam penelitian ini, yaitu pccBA-GFP, ptiBA-GFP (Alimuddin et al., 2007a) dan pmkBA-GFP (Hamada et al.,
1998) yang masing-masing dikontrol oleh promoter ccBA ikan mas (panjang sekuen 1,3 kb), tiBA ikan nila (1,2 kb; Octavera, 2008) dan mkBA ikan medaka (3,7 kb; Takagi et al., 1994). Setiap konstruksi gen tersebut dengan konsentrasi 50
Gambar 3. Injeksi pada blastodisk embrio ikan mas
Sebanyak 8 embrio diambil dari akuarium inkubasi, kemudian diletakkan dan diatur secara hati-hati menggunakan pipet ke dalam cawan agarosa (Gambar 4) yang berfungsi sebagai penahan telur. Posisi embrio diatur sedemikian rupa sehingga blastodisk mengarah ke jarum mikroinjeksi untuk memudahkan proses injeksi.
Gambar 4. Cawan agarosa 0,7% yang mengandung lubang yang berfungsi sebagai penahan embrio saat injeksi
Proses mikroinjeksi dilakukan di bawah mikroskop (Olympus SZX 16) (Gambar 5a) dengan bantuan mikromanipulator (Gambar 5b) dan mikroinjektor (Gambar 5c). Jumlah embrio yang diinjeksi untuk setiap konstruksi gen adalah sebanyak 30 butir dan dilakukan sebanyak 2 kali ulangan untuk masing-masing konstruksi gen.
Jarum Injeksi Blastodisk
Gambar 5. Seperangkat alat mikroinjektor yang terdiri atas mikroskop Olympus SZX 16 (A), mikromanipulator (B) dan mikroinjektor (C).
Embrio-embrio yang telah diinjeksi dimasukkan ke dalam akuarium inkubasi yang telah diberikan methylene blue dan suhu airnya berkisar 28C
(Gambar 6). Selama pengamatan, embrio yang mati dibuang.
Gambar 6. Akuarium inkubasi embrio
Embrio-embrio yang telah diinjeksi, dimasukkan ke dalam cawan petri dan selanjutnya diamati menggunakan mikroskop fluoresen (Olympus SZX 16) untuk melihat ekspresi gen Green Fluorescent Protein (GFP) pada perbesaran 0,8
kamera digital High Speed Compact Color 2 megapiksel (DP20) (Gambar 7c) kemudian ditransfer ke komputer (Gambar 7d) yang memiliki software Olympus
DH2-BW melalui remote controller (Olympus DP-20) (Gambar 7e).
Gambar 7. Seperangkat alat untuk pengamatan ekspresi gen GFP yang terdiri dari mikroskop fluoresen (Olympus SZX 16) (A), burner (Olympus) (B), kamera digital High Speed Compact Color 2 megapiksel (DP20) (C),
komputer (D) dan remote controller (Olympus DP-20) (E).
promoter mkBA dilakukan pada jam ke 10 (puncak ekspresi setiap promoter). Persentase larva yang mengekspresikan gen GFP (PLMG) diperoleh dari perbandingan jumlah larva yang mengekspresikan gen GFP dibandingkan dengan jumlah total embrio diinjeksi yang menetas. Pengamatan tingkat dan pola ekspresi gen GFP dilakukan pada fase embrio yaitu pada saat puncak ekspresi masing-masing promoter. Pengamatan pola ekspresi dilakukan pada jam kedua setelah fertilisasi setiap 2 jam hingga larva menetas. Data hasil pengamatan dan perhitungan dari kedua ulangan dirata-ratakan dan kemudian dianalisis secara deskriptif dan disajikan dalam bentuk tabel, grafik dan gambar.
3.2.3.2 Pengujian Secara Kuantitatif Menggunakan Reverse Transcriptase
RT-PCR
RNA total diekstraksi dari 3 butir embrio ikan mas pada jam ke-30 untuk promoter ccBA dan tiBA serta jam ke 10 untuk promoter mBA yang merupakan waktu puncak aktivitas setiap promoter. Setelah ekstraksi RNA, sintesis cDNA dilakukan menggunakan kit Ready-To-Go You-Prime First-Strand beads (GE Healthcare, UK). Amplifikasi PCR sampel cDNA dilakukan dengan menggunakan primer GFP-F (5’-GGTCGAGCTGGACGGCGACG-3’) dan primer GFP-R (5’-ACGAACTCCAGCAGGACCAT-3’). PCR dilakukan dengan denaturasi awal 2 menit pada suhu 94C, 35 siklus yang terdiri atas 30 detik denaturasi pada suhu 94C, 30 detik annealing pada suhu 62C dan 1 menit extension pada suhu 72C. Dua μl hasil PCR dielektroforesis menggunakan gel
agarosa 0,7%, distaining dengan etidium bromida dan difoto dengan kamera digital dalam kondisi disinari dengan cahaya ultraviolet. Sebagai kontrol internal kesamaan jumlah RNA dalam sintesis cDNA, gen β-aktin diamplifikasi menggunakan primer yang didesain berdasarkan database gen β-aktin dari ikan mas (no. Akses Bank Gen: M24113), yaitu primer forward (5’
-ATGGTTGGTATGGACAGAAGGAC-3’) dan primer reverse (5’
IV. HASIL DAN PEMBAHASAN
4.1. Isolasi Promoter -Aktin Ikan Mas
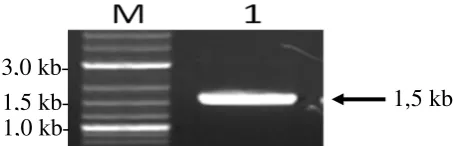
Promoter -Aktin dari ikan mas diisolasi dengan menggunakan metode PCR dengan primer yang dibuat berdasarkan data yang ada di Bank Gen. Panjang fragmen DNA hasil amplifikasi PCR adalah sekitar 1,5 kb (Gambar 8).
Gambar 8. Elektroforesis hasil isolasi promoter -Aktin Ikan Mas (1) dan (M) marker ukuran fragmen DNA 2-log ladder (Biolabs, New England). Fragmen DNA target ditunjukkan dengan tanda panah.
Panjang sekuen hasil sekuensing adalah 1528 bp (Gambar 9). Berdasarkan analisis sekuens diperoleh faktor transkripsi (transcription factor) yang merupakan elemen-elemen yang mempengaruhi aktivitas promoter. Elemen penting bagi promoter -aktin adalah boks TATA, motif CCAAT, motif CArG atau CC(A/T)6GG. Hubungan tingkat aktivitas promoter -aktin dengan sekuens CCAAT telah diteliti oleh Quitschke et al. (1989). Kegunaan motif CArG sebagai elemen responsif terhadap serum terletak antara CCAAT dan boks TATA, telah dijelaskan oleh Liu et al. (1990). Boks TATA merupakan elemen yang umum
dijumpai pada sekuens promoter, sebagai tempat RNA polimerase melekat pada saat transkripsi RNA berlangsung (Glick & Pasternak, 2003). Kerja sama antara ketiga elemen tersebut menyebabkan promoter dapat aktif dan mengendalikan ekspresi transgen pada waktu dan tempat yang tepat.
Hasil analisis menunjukkan bahwa ketiga elemen faktor transkripsi ini telah dilaporkan dijumpai pada promoter -aktin dari berbagai jenis ikan juga terdapat pada sekuens DNA hasil isolasi dari ikan mas. Sekuens CCAAT pada ikan mas terletak pada nukleotida 12-16, motif CArG terletak pada nukleotida 42 - 51, serta boks TATA terletak pada nukleotida 75-80 dihitung dari ujung terminal 5. Kandidat boks TATA lainnya untuk ikan mas terletak pada nukleotida 965-970
1,5 kb 1,0 kb-
dari ujung terminal 5. Selain itu beberapa binding site lain yang potensial dalam transkripsi penting adalah GATA,TFIID, CBF, Sp1, TBP dan sebagainya juga ditemukan dalam sekuen ini. Elemen-elemen ini mungkin berfungsi sebagai regulasi transkripsi gen -aktin.
Analisis pensejajaran (alignment) hasil sekuensing gen β-aktin ikan mas
dengan data -aktin ikan mas dari bank gen (no. Aksesi Bank Gen: M24113) terdapat kemiripan yang tinggi, yaitu 97,52% (Gambar 9). Dari hasil analisis sekuen dan pensejajaran diduga bahwa sekuen hasil isolasi merupakan promoter
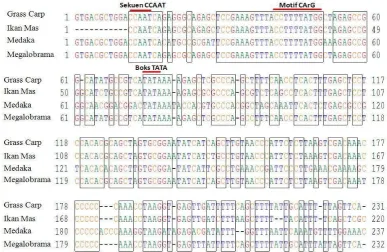
-aktin ikan mas. Alignment hasil sekuensing promoter β-aktin ikan mas dengan ikan grass carp (no. Aksesi Bank Gen: M25013), ikan medaka (no. Aksesi Bank
Gen: S74868) dan ikan megalobrama (no. Aksesi Bank Gen: AY170122) ditunjukkan pada Gambar 10. Berdasarkan hasil alignment diketahui bahwa posisi sekuen CCAAT, motif CArG dan boks TATA promoter β-aktin ikan mas relatif sama dengan ikan grass carp, ikan medaka dan megalobrama yang telah dilaporkan sebelumnya. Hal ini memperkuat dugaan bahwa hasil kloning merupakan promoter β-aktin ikan mas.
4.2 Konstruksi pccBA-EGFP
Untuk mengetahui promoter β-aktin yang diisolasi dari ikan mas dapat aktif atau tidak, maka promoter tersebut disambungkan/ligasi dengan gen penanda GFP. Keberhasilan ligasi dianalisis menggunakan metode “cracking” seperti ditunjukkan pada Gambar 11. Ukuran ccBA-GFP lebih besar dibandingkan dengan plasmid pEGFP-N1 dan plasmid dari kontrol berupa bakteri biru. Hal ini menunjukkan bahwa konstruksi gen pccBA-EGFP berhasil dibuat dan koloni bakteri yang membawa plasmid ini berhasil diidentifikasi (no. 1-6 pada Gambar 11).
4.3 Pengujian Efektivitas Promoter
derajat penetasan (DP), dan persentase embrio (PEMG) dan larva (PMLG) yang mengekspresikan gen GFP.
Gambar 9. Alignment hasil sekuensing promoter -aktin ikan mas dengan gen -aktin ikan mas (no. Akses Bank Gen: M24113). Garis bawah merah
lines 1 dan 2 merupakan sekuen primer forward yang digunakan untuk
isolasi promoter maupun untuk pembuatan konstruksi gen. Lines 22 dan 23 merupakan sekuen primer reverse yang digunakan untuk pembuatan konstruksi, lines 26 dan 27 merupakan sekuen primer
Gambar 10. Alignment hasil sekuensing promoter β-aktin ikan mas, ikan grass carp (no. Akses Bank Gen: M25013), ikan medaka (no. Akses Bank Gen: S74868) dan ikan megalobrama (no. Akses Bank Gen: AY170122). Posisi sekuen CCAAT, motif CArG dan boks TATA ditunjukkan di atas sekuens. Posisi sekuen CCAAT, motif CArG dan boks TATA yang ada dalam sekuens primer F untuk ikan mas sama dengan ikan lainnya dilihat dari ujung terminal 5 (dilihat dari sebelah kiri).
Gambar 11. Elektroforesis hasil cracking bakteri koloni berwarna biru (K) dan yang putih (no. 1-6). No. 1-5: bakteri yang membawa konstruksi gen pccBA-EGFP, no. 6: bakteri yang membawa plasmid pEGFP-N1. Tanda minus (▬) di sebelah kanan gambar menunjukkan ukuran DNA plasmid yang mengandung DNA insersi atau pccBA-EGFP, tanda kepala panah tertutup (◄) untuk plasmid DNA pEGFP-N1, sedangkan tanda kepala panah terbuka (<) untuk plasmid dari bakteri anti koloni biru putih.
K 1 2 3 4 5 6
◄ ▬
K 1 2 3 4 5 6
◄ ▬
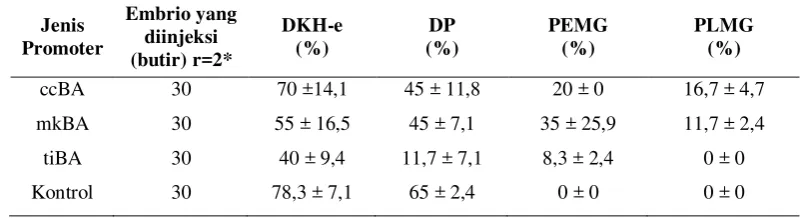
Nilai DKH-e, DP, PEMG dan PLMG diperlihatkan pada Tabel 1. Nilai DKH-e pada ccBA-GFP(70%) lebih tinggi dibandingkan dengan mkBA-GFP (55%) dan tiBA-GFP (40%), tetapi nilai DP pada ccBA-GFP relatif sama dengan mkBA-GFP yaitu 45% dibandingkan dengan tiBA-GFP (11,67%). Hal ini menunjukkan bahwa dengan menggunakan ccBA-GFP ternyata tidak terlalu mempengaruhi perkembangan embrio. Namun, nilai DKH-e dan DP dari perlakuan masih lebih rendah jika dibandingkan kontrol. Hal ini diduga disebabkan adanya kerusakan sel akibat injeksi atau kemungkinan akibat tingginya volume larutan DNA yang diinjeksikan. Menurut Zbikwoska (2003) untuk memastikan material genetik masuk dalam pronukleus, konsentrasi larutan DNA yang tinggi diinjeksikan ke dalam sitoplasma telur-telur yang telah difertilisasi. Namun semakin tinggi konsentrasi larutan DNA yang diinjeksikan juga akan meningkatkan mutagenesis atau meningkatkan jumlah partikel asing yang dapat menyebabkan kematian pada embrio (Hackett, 1993).
Sementara itu PEMG dan PLMG hanya ditemukan pada perlakuan. PEMG pada mkBA-GFP (35%) lebih tinggi dibandingkan dengan ccBA-GFP (20%) dan tiBA-GFP (8,3%). Selanjutnya, PLMG pada ccBA-GFP (16,67%) lebih tinggi dibandingkan dengan mkBA-GFP (11,67%), sedangkan pada tiBA-GFP tidak ada larva yang mengekspresikan GFP. Data keseluruhan disajikan pada Lampiran 1. Dengan jumlah dan jenis faktor transkripsi sama, dapat disimpulkan bahwa promoter homolog (ccBA) lebih efektif dalam mengatur ekspresi gen target dibandingkan dengan promoter heterolog (mkBA dan tiBA).
Tabel 1. Derajat kelangsungan hidup embrio (DKH-e), derajat penetasan (DP), persentase embrio yang mengekspresikan gen GFP (PEMG) dan persentase larva yang mengekspresikan gen GFP pada ikan mas.
4.3.1 Pola Ekspresi secara Kualitatif
Tingkat ekspresi gen GFP yang dihasilkan dari perlakuan dibagi menjadi tiga kelompok yaitu pendaran hijau kurang terang, pendaran hijau terang dan pendaran hijau sangat terang. Perbandingan tingkat ekspresi antar konstruksi DNA diamati pada jam ke-30 untuk promoter dari ikan mas (ccBA) dan promoter dari ikan nila (tiBA), sedangkan promoter dari ikan medaka (mkBA) pada jam ke-10 dan disajikan pada Tabel 2.
Tabel 2. Tingkat ekspresi gen ccBA-GFP, mkBA-GFP dan tiBA-GFP pada
Keterangan : 1. Pendar hijau kurang terang. 2. Pendar hijau terang. 3. Pendar hijau sangat terang
Berdasarkan Tabel 2 terlihat bahwa pada ketiga klasifikasi tingkat ekspresi gen GFP, persentase embrio yang mengekspresikan GFP pada promoter mkBA lebih tinggi dibandingkan dengan promoter ccBA dan promoter tiBA. Selanjutnya, promoter ccBA lebih tinggi dibandingkan dengan promoter tiBA. Hal ini menunjukkan bahwa promoter homolog (ccBA) memiliki aktivitas lebih tinggi dibandingkan dengan yang heterolog (tiBA). Namun demikian, meskipun promoter mkBA heterolog bagi ikan mas, perbedaan panjang sekuen yang sangat signifikan antara promoter mkBA (3,7 kb) dan ccBA (1,3 kb) mungkin mempengaruhi ekspresi GFP.
embrio-embrio ikan yang diinjeksi. Ekspresi pada setiap embrio sebagian atau tidak tersebar dimana ada beberapa jaringan yang terekspresi kuat sementara yang lain tidak ada. Kebanyakan tingkat ekspresi diduga berhubungan kuat dengan jumlah copy transgen pada setiap sel (Hwang et al., 2003). Alimuddin et al. (2007b) menjelaskan bahwa ekspresi gen yang lemah atau bahkan tidak ada sama sekali bila terintegrasi di sentromer atau di telomer dimana DNA tidak aktif mengalami transkripsi dan diposisikan di heterokromatin. Selain itu, kemungkinan integrasi gen terjadi secara acak dalam kromosom, sehingga terjadi perbedaan tingkat ekspresi gen di jaringan yang berbeda.
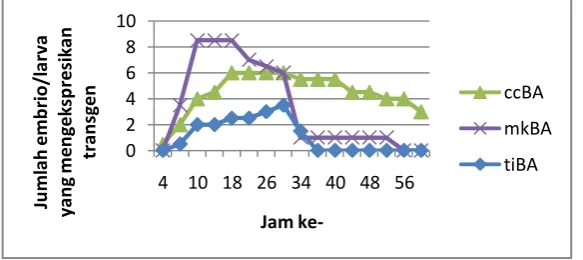
Pola ekspresi gen GFP yang diamati pada embrio hasil mikroinjeksi bersifat sementara (transient expression) dimana ekspresi yang dihasilkan
awalnya rendah, kemudian meningkat dan akhirnya menurun (Gambar 12).
Gambar 12. Pola ekspresi gen GFP pada ccBA-GFP, mKBA-GFP dan tiBA-GFP pada embrio/larva ikan mas Cyprinus carpio
dari ikan mas memiliki efektivitas yang lebih tinggi dibandingkan dengan promoter β-aktin dari ikan medaka dan ikan nila.
Gambar 13. Ekspresi gen GFP menggunakan ccBA-GFP pada embrio dan larva ikan mas. Inisiasi ekspresi pada jam ke-6 (A), Ekspresi gen pada jam ke-10 (B), Puncak ekspresi pada jam ke-30 (C) Ekspresi pada larva jam ke-34 (D), Ekspresi pada larva jam ke-56 (E), dan Ekspresi pada larva jam ke-192 (F).
Gambar 15. Ekspresi gen GFP menggunakan tiBA-GFP pada embrio ikan mas. Inisiasi ekspresi gen pada jam ke-6 (A), Ekspresi gen pada jam ke-10 (B), Puncak ekspresi gen pada jam ke-30 (C), Akhir ekspresi gen pada jam ke-32 (D) dan ekspresi gen tidak nampak lagi pada larva jam ke-34 (E).
Pola ekspresi gen asing yang tinggi terjadi pada fase mid blastula transition (MBT) hingga fase gastrula sebagai hasil dari akumulasi DNA yang diinjeksikan akibat replikasi selama tahap pembelahan dan akumulasi enzim yang menyebabkan dimulainya proses transkripsi pada fase MBT (Iyengar et al., 1996).
Ekspresi gen mulai menurun dengan waktu yang berbeda pada tiap promoter. Hal ini mungkin berhubungan dengan aktivitas enzim eksonuklease dan/atau endonuklease di dalam embrio dan larva. Aktivitas enzim tersebut dalam memotong konstruksi DNA asing diduga berbeda antara yang homolog dan heterolog, sehingga kecepatan turunnya ekspresi GFP menjadi berbeda.
Gambar 16. Posisi ekspresi gen GFP menggunakan ccBA-GFP pada larva ikan mas pada kepala (A), kuning telur (B), dan otot (C).
Gambar 17. Posisi ekspresi gen GFP menggunakan mkBA-GFP pada larva ikan mas pada kepala (A), otot (B), dan ekor (C).
Ekspresi gen GFP yang dikendalikan oleh promoter ccBA-GFP dan promoter mkBA-GFP tidak spesifik pada suatu organ. Hal ini berkenaan dengan sifat ubiquitous (terdapat dimana-mana) dari promoter β-aktin yang artinya
promoter ini dapat aktif pada semua jaringan otot. Menurut Iyengar et al. (1996),
distribusi yang tidak sama dari copy transgen dalam jaringan poliploid seperti sel-sel otot atau sel-sel-sel-sel pada lapisan syncytial telur (YSL) mungkin menjelaskan tingginya variabilitas ekspresi transgen dalam jaringan. Lebih lanjut dijelaskan bahwa perbedaan ekspresi transgen mungkin berhubungan dengan posisi dimana transgen terintegrasi dalam kromosom yang mengaktifkan gen tersebut di otot. Ekspresi gen GFP menggunakan ccBA-GFP dan mkBA-GFP pada larva tergolong kuat. Hal ini terjadi diduga karena ekspresi dapat muncul dan menguat kembali pada saat dimana pertumbuhan berlangsung cepat sehingga banyak sel yang membelah dan kembali terjadi peningkatan replikasi atau dapat pula terjadi pada saat mulai terjadinya pembentukan otot-otot karena promoter β-aktin diisolasi dari otot dan dapat aktif pada semua jaringan otot (Winkler et al., 1991). Kejadian -
kejadian ini mungkin terjadi mengingat promoter β-aktin memiliki sifat
housekeeping (dapat aktif kapan saja) dan constitutive (aktif tanpa faktor pemicu).
Akan tetapi ekspresi ini kelihatan tidak nampak lagi pada waktu tertentu karena telah tertutup oleh warna kulit, daging dan sebagainya bergantung dari kemampuan masing-masing promoter dalam mengekspresikan gen GFP. Lain halnya dengan ekspresi gen menggunakan tiBA-GFP pada larva tidak nampak lagi. Ekspresi gen GFP ditemukan hanya sampai fase gastrula, sedangkan pada embrio yang sudah melewati fase tersebut (older embryos) DNA asing yang
bertahan hanya dalam jumlah yang terbatas akibat adanya degradasi (Winkler et al., 1991).
4.3.2 Pola Ekspresi secara Kuantitatif
Metode semi-kuantitatif Reverse Transcription PCR (RT-PCR) dilakukan
untuk mengetahui tingkat ekspresi transgen GFP yang dikendalikan oleh promoter homolog dan heterolog dari setiap konstruksi gen yang diuji.
Gambar 18. Analisis ekspresi gen GFP menggunakan RT-PCR. (a) Kolom no 1, 2 dan 3, produk PCR dari embrio yang masing-masing telah diinjeksi dengan ccBA-GFP, mkBA-GFP dan tiBA-GFP. K- adalah produk PCR tanpa cetakan DNA, K+ adalah produk PCR dengan cetakan plasmid mengandung GFP, dan M adalah marker molekuler DNA. Panjang fragmen GFP yang diamplifikasi adalah sekitar 600 bp. (b) Produk PCR dengan primer gen β-aktin (200 bp).
V. KESIMPULAN DAN SARAN
5.1 Kesimpulan
Dari hasil penelitian dapat disimpulkan bahwa :
1. Promoter β-aktin ikan mas telah berhasil diisolasi dengan panjang 1,5 kbp dan yang digunakan dalam pembuatan konstruksi adalah 1,3 kbp. Pada sekuens yang diisolasi terdapat faktor transkripsi yang biasa dijumpai pada promoter β -aktin, yaitu boks TATA, motif CCAAT dan motif CArG (CC(A/T)6GG).
2. Promoter homolog memiliki efektivitas yang lebih tinggi daripada promoter heterolog pada transgenesis ikan mas.
5.2 Saran
DAFTAR PUSTAKA
Alimuddin. 2003. Introduction and expression of foreign Δ6 desaturase-like gene in a teleostean fish [Thesis]. Graduate School of Fisheries Science. Tokyo University of Fisheries.
Alimuddin, Yoshizaki G, Carman O dan Sumantadinata K. 2003. Aplikasi transfer gen dalam akuakultur. JurnalAkuakultur Indonesia 2: 41-50.
Alimuddin, Sumantadinata K dan Arifin OZ. 2007a. Teknologi transgenesis dalam peningkatan kecepatan tumbuh ikan nila merah (Oreochromis niloticus). Laporan Riset Insentif KNRT.
Alimuddin, Yoshizaki G, Carman O dan Takeuchi T. 2007b. Efektivitas promoter hCMV, mEF1α dan mAct dalam mengatur ekspresi gen asing pada transgenik ikan zebra. JurnalAkuakultur Indonesia 6: 65-77.
Anderson L. 2004. Genetically engineered fish-new threats to the environment.
Book Article. Greenpeace International. Netherlands.
Arai R, Makita Y, Oda Y and Nagamune T. 2001.Construction of green fluorescent protein reporter gene for genotoxicity test (SOS/umu-test) and improvement of mutagen-sensitivity. Bioscience and Bioengineering 92: 301-304.
Beaumont and Hoare. 2003. Biotechnology and Genetics in Fisheries and Aquaculture. Blackwell Science Ltd. Blackwell Publishing Inc. USA:
128-128.
Chou CY, Horng LS and Tsai HJ. 2001. Uniform GFP-expression in transgenic medaka Oryzias latipes at the F0 generation. Transgenic Research 10:
303-315
Dunham RA. 2004. Aquaculture and Fisheries Biotechnology: Genetic Approaches. CABI Publishing. Cambridge, MA, USA, p.160-164.
Felts K, Rogers B, Chen K, Ji H, Sorge J and Vaillancourt P. 2001. Recombinant
Renilla reniformis GFP displays low toxicity. Stratagene 13: 85-87.
Glick BR and Pasternak JJ. 2003. Molecular Biotechnology: Principles and Applications of Recombinant DNA. 3rd Edition. ASM Press. Washington DC.
Gong Z, Wan H, Ju B, He J, Wang X and Yan T. 2002. Generation of living color transgenic fish. In: Shimizu N, Aoki T, Hirono I and Takashima F. (Eds.)
Aquatic Genomics : Steps Toward a Great Future. Springer-Verlag. New
Hackett PB. 1993. The molecular biology of transgenic fish. In : Hocachka and Mommesen (Eds.). Biochemistry and Molecular Biology of Fishes 2 : 218 – 229.
Hamada K, Tamaki K, Sasado T, Watai Y, Kani S, Wakamtsu Y, Ozato K, Kinoshita M, Kohno R, Takagi S and Kimura M. 1998. Usefulness of the medaka β-actin promoter investigated using a mutant GFP reporter gene in transgenic medaka Oryzias latipes. Molecular Marine Biology and Biotechnology, 7:173-180.
Higashijima S, Okamoto H, Ueno N, Hotta Y and Eguchi G. 1997. High frequency generation of transgenic zebrafish which reliably express GFP in whole muscles or the whole body by using promoter of zebrafish origin.
Developmental Biology 92 : 289-299.
Hulata G. 2001. Genetic manipulations in aquaculture: a review of stock improvement by classical and modern technologies. Genetica 111: 155–173.
Hwang GL, Rahman MA, Razak SA, Sohm F, Farahmand H, Smith A, Brooks C and Maclean N. 2003. Isolation and characterisation of tilapia β-actin promoter and comparison of its activity with carp β-actin promoter.
Biochimica et Biophysica Acta, 1625 : 11-18.
Iyengar A, Muller F and Maclean N. 1996. Regulation and expression of transgenes in fish-a review. Transgenic Research 5: 147-166.
Jusuf M. 2001. Genetika I Struktur dan Ekspresi Gen. CV. Sagung Seto. Jakarta :
288-289.
Kato K, Takagi M, Tamaru Y, Akiyama S-1, Konishi T, Murata O and Kumai H. 2007. Construction of an expression vector containing β-actin promoter region for gene transfer by microinjection in Red sea bream Pagrus major.
Fisheries Science, 73 : 440-445.
Liu Z, Moav B, Faras AJ, Guise KS, Kapucinski AR and Hackett PB. 1990. Functional analysis of elements affecting of the β-actin gene of carp.
Molecular Cell Biology 10: 3432-3440.
Lutz CG. 2001. Practical Genetics for Aquaculture. Fishing New Books.
pp.218-219. Blackwell Science Company. USA.
Nam YK, Maclean N, Hwang G and Kim DS. 2008. Autotransgenic and allotransgenic manipulation of growth traits in fish for aquaculture: a review. Fish Biology 72: 1–26.
Octavera A. 2008. Isolasi promoter β-actin ikan nila Oreochromis niloticus
dengan metode degenerate PCR [skripsi]. Bogor: Fakultas Perikanan dan
Ilmu Kelautan, Departemen Budidaya Perairan, Institut Pertanian Bogor. Quitschke WW, Lin Z-Y, DePoti-Zilli L and Paterson BM. 1989. The β-actin
promoter. Biology and Chemistry 264: 9539-9546.
Rahman MA and Maclean N. 1999. Growth performance of transgenic tilapia containing an exogenous piscine growth hormone gene. Aquaculture 173:
333-346.
Rahman MA and Maclean N. 1991. Production of transgenic tilapia (Oreochromis niloticus) by one-cell-stage microinjection. Aquaculture 105: 219-232.
Rajesh R and Majumdar KC. 2006. Transgene integration- an analysis autotransgenic Labeo rohita Hamilton (Pisces: Cyprinidae). Fish Physiology and Biochemistry 31: 281-287.
Sarmasik A. 2002. Application of gene transfer technology for genetic improvement of fish: review. Turk J. Zool. 27: 1-6.
Takagi, S, Sasado G, Tamiya G, Ozato K, Wakamatsu Y, Takeshita A and Kimura M. 1994. An efficient expression vector for transgenic medaka construction. Molecular Marine Biology and Biotechnology 3:192-199. Teufel J, Pätzold F and Potthof C. 2002. Scientific research on transgenic fish
with special focus on the biology of trout and salmon. Environmental Research of Federal Ministry of Environment, Nature Conservation and Nuclear Safety, Research Report 360 05 023. Öko-Institut e.V., Institut für
angewandte Ökologie, Pätzold Gewässerökologie. Germany.
Tsai HJ. 2003. Transgenic fish: researches and applications. J. Fish. Soc. 30:
263-277.
Toha AHA. 2001. Deoxyribo Nucleac Acid: Keanekaragaman, Ekspresi, Rekayasa dan Efek Pemanfaatannya. Alfabeta. Bandung: 55-56.
Winkler, C, Vielkind JR and Schartl M. 1991. Transient expression of foreign DNA during embryonic and larval development of the medaka fish Oryzias latipes. Molecular General Genetics 226: 129 – 140
Yazawa R, Hirono I and Aoki T. 2006. Transgenic zebrafish expressing chicken lysozyme show resistance againts bacterial disease. Transgenic Research 3:
Yazawa R, Hirono I, and Aoki T. 2005. Characterization of promoter activities of four different Japanese flounder promoters in transgenic zebrafish. Marine Biotechnology 7(6): 625-633.
Yoshizaki G. 2001. Gene transfer in salmonidae: applications to aquaculture,
Suisanzoshoku 49 : 137-142.
Lampiran 1. Derajat kelangsungan hidup embrio (DKH-e), derajat penetasan (DP), persentase embrio yang mengekspresikan gen GFP (PEMG) dan persentase larva yang mengekspresikan gen GFP pada ikan mas untuk tiap ulangan.
Keterangan :
n = jumlah embrio yang diinjeksi (butir) DKH-e = Derajat Kelangsungan Hidup Embrio (%) DP = Derajat Penetasan (%)
PEMG = Persentase Embrio yang Mengekspresikan Gen GFP PLMG = Persentase Larva yang Mengekspresikan Gen GFP
Lampiran 2. Tingkat ekspresi gen green fluorescent protein pada embrio ikan mas untuk tiap ulangan.
Jenis
Promoter Ulangan
Tingkat Ekspresi Gen* Jml embrio yg terekspresi
1 2 3
Butir % Butir % Butir % (Butir)
ccBA
1 3 10 2 6,67 1 3,33 6
2 2 6,67 3 10 1 3,33 6
Rata-rata 2,5 8,33 2,5 8,33 1 3,33 6
Standar Deviasi 0,7 2,4 0,7 2,4 0,0 0,0 0,0
mkBA
1 6 20 6 20 4 13,33 16
2 2 6,67 2 6,67 1 3,33 5
Rata-rata 4,0 13,3 4 13,3 2,5 8,3 10,5
Standar Deviasi 2,8 9,4 2,8 9,4 2,1 7,1 7,8
tiBA
1 2 6,67 1 3,33 0 0 3
2 1 3,33 1 3,33 0 0 2
Rata-rata 1,5 5 1 3,33 0 0 2,5
Standar Deviasi 0,7 2,4 0,7 0 0,0 0,0 0,7