KAITANNYA DENGAN UNSUR HARA DAN
CAHAYA DI PERAIRAN MUARA JAYA
TELUK JAKARTA
USMAN MADUBUN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa tesis Produktivitas Primer Fitoplankton dan Kaitannya dengan Unsur Hara dan Cahaya adalah karya saya sendiri dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada pihak mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Juli
2008
Usman Madubun
USMAN MADUBUN. Phytoplankton Primary Productivity and it’s relation with Nutrient and Light in Muara Jaya Waters, Jakarta Bay. Under direction of ARIO DAMAR and RICHARDUS F. KASWADJI.
The research was conducted in Muara Jaya waters, Jakarta Bay, from July-August 2007. The research was aimed to study the phytoplankton primary productivity in relation to nutrient and light distribution. Survey of post facto approach was implemented in three stations arranged from estuary towards the sea. Parameters studied were primary productivity, nutrients, light intensity, chlorophyll a, and the abundance and composition of phytoplankton. Results showed that primary productivity ranged from 39,57-96,89 (67,36±25), 21,31-107,01 (63,38±29), and 18,07-108,12 (62,61±30) mgCm-3h-1 for station 1, 2, and 3, respectively. There was a significant difference among incubation depths, but not among stations. Nutrients had varied role to variability of primary productivity, and there was no significant difference among incubation depths and stations, except for silicate. Light intensity had significant influence on primary productivity, implied significant differences among stations and incubation depths. Nutrient concentrations and primary productivity in estuary were higher compared to those in other stations, while light intensity showed the opposite figure. This indicates the role of incoming loads from the river. Chlorophyll a
concentration in estuary station was higher than in other stations, showing significant role of nutrients and light intensity. Phytoplankton composition were predominated by Bacillariophyceae (29 species), Cyanophyceae (1 species), and Dinophyceae (8 species).
USMAN MADUBUN. Produktivitas Primer Fitoplankton dan kaitannya dengan Unsur Hara dan Cahaya di perairan Muara Jaya Teluk Jakarta. Dibawah bimbingan ARIO DAMAR and RICHARDUS F. KASWADJI.
Penelitian ini telah dilakukan di perairan Muara Jaya Teluk Jakarta dari bulan Juli – Agustus 2007. Tujuan penelitian adalah untuk mengkaji produktivitas primer fitoplankton sehubungan dengan distribusi unsur hara dan cahaya. Survey post facto dilakukan di tiga stasiun yang ditetapkan dari muara ke arah laut. Parameter yang diukur adalah produktivitas primer, unsur hara, cahaya, klorofil-a, kelimpahan dan komposisi fitoplankton. Hasil penelitin memperlihatkan bahwa kisaran nilai produktivitas primer berturut-turut untuk stasiun 1, 2, dan 3 adalah 39,57-96,89 (67,36±25), 21,31-107,01 (63,38±29), dan 18,07-108,12 (62,61±30) mgCm-3jam-1, terdapat perbedaan nyata antar kedalaman tetapi tidak antar stasiun. Unsur hara memperlihatkan peranan yang bervariasi terhadap keragaman nilai produktivitas primer, dan tidak terdapat perbedaan nyata antara kedalaman inkubasi maupun stasiun, kecuali untuk silikat. Intensitas cahaya berperan nyata pada produktivitas primer, dan menunjukkan perbedaan nyata antar stasiun dan kedalaman inkubasi. Konsentrasi nutrien dan produktivitas lebih tinggi di stasiun muara dibandingkan dengan stasiun lainnya, sedangkan cahaya sebaliknya. Hal ini menggambarkan peranan beban masukan dari sungai. Tingkat klorofil-a lebih tinggi pada stasiun di muara, menggambarkan pentingnya peranan unsur hara dan cahaya. Komposisi jenis fitoplankton terdiri dari kelas Bacillariophyceae (29 jenis), Cyanophyceae (1 jenis), dan Dinophyceae (8 jenis).
© Hak Cipta milik Institut Pertanian Bogor, tahun 2008
Hak Cipta dilindungi Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau
CAHAYA DI PERAIRAN MUARA JAYA
TELUK JAKARTA
USMAN MADUBUN
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Perairan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul : Produktivitas Primer Fitoplankton dan Kaitannya dengan Unsur Hara dan Cahaya di Perairan Muara Jaya Teluk Jakarta
Nama : Usman Madubun NRP : C151050061
Disetujui
Komisi Pembimbing
Dr. rer.nat. Ir. Ario Damar, M.Si. Dr. Ir. Richardus Kaswadji, M.Sc.
Ketua Angggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana
Prof. Dr. Ir. Enang Harris, M.S. Prof. Dr. Ir. Khairil A. Notodiputro, M.Sc.
Penguji Luar Komisi pada Ujian Tesis: Dr. Ir. Niken Tunjung Murti Pratiwi, M.S.
Alhamdulillah penulis panjatkan kehadirat Allah SWT, atas limpahan kasih sayang-Nya sehingga penulisan tesis ini dapat diselesaikan. Tema penelitian ini adalah fitoplankton dengan judul Produktivitas primer fitoplankton dan kaitananya dengan unsur hara dan cahaya yang dilaksanakan di perairan Muara Jaya Teluk Jakarta dari bulan Juli hingga Agustus 2007.
Terima kasih dan penghargaan penulis ucapkan kepada Dr. Ario Damar dan Dr. Richardus F. Kaswadji selaku komisi pembimbing, Dr. Niken TM Pratiwi selaku penguji luar komisi serta Dr. Kardiyo Praptokardiyo yang telah banyak memberikan arahan dan saran. Di samping itu, penghargaan penulis sampaikan juga kepada Bapak Direktur Politeknik Perikanan Negeri Tual dan Bapak Direktur Jenderal Pendidikan Tinggi Departemen Pendidikan Nasional atas kesempatan yang diberikan untuk melanjutkan studi. Terima kasih penulis sampaikan pula kepada bapak Deden atas bantuannya selama pengambilan contoh di lapangan serta Ibu Anna beserta staf di Laboratorium Produktivitas dan Lingkungan Perairan, MSP-IPB atas bantuan analisis contoh air. Demikian pula terima kasih atas bantuan dari teman-teman seangkatan di PS Ilmu Perairan, dan teman-teman seperjuangan dari Tual, Is, Dani, Cecu, Beni, dan Yula.
Ungkapan terima kasih dan penghargaan yang sama penulis sampaikan pula
kepada (alm & almh) Ayah dan Ibu, Drs. Jusuf Madubun, M.Si. sekeluarga, Ir. Idrus Madubun sekeluarga, dan Khadijah Madubun, S.Sos., serta semua
keluarga atas bantuan materil dan dukungan moral selama ini. Terakhir, terima kasih dan penghargaan yang sangat mendalam penulis sampaikan kepada istri tercinta Ratna Alwiyah Rahajaan, S.T. atas pengorbanan, kesabaran, dorongan, doa, dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2008
Penulis dilahirkan di Tual, Kabupaten Maluku Tenggara pada tanggal 25 Agustus 1970 dari pasangan ayah (alm) Mihidin Madubun dan ibu (almh) Aminah Madubun. Penulis merupakan putra ke delapan dari 12 bersaudara.
DAFTAR ISI
Halaman
DAFTAR TABEL ... xi
DAFTAR GAMBAR ... xii
DAFTAR LAMPIRAN ... xiv
I. PENDAHULUAN ... 1
1.1 Latar Belakang ... 1
1.2 Perumusan Masalah ... 2
1.3 Tujuan dan Manfaat ... 4
1.4 Hipotesis ... 4
II. TINJAUAN PUSTAKA ... 5
2.1 Fitoplankton ... 5
2.2 Produktivitas Primer ... 8
2.3 Cahaya ... 10
2.4 Unsur Hara ... 12
2.5 Suhu ... 15
2.6 Kekeruhan ... 16
2.7 Salinitas ... 16
III. METODE ... 18
3.1 Lokasi dan Waktu Penelitian ... 18
3.2 Metode dan Desain Penelitian ... 18
3.3 Teknik/Metode Analisis Data ... 23
IV. HASIL DAN PEMBAHASAN ... 25
4.1 Parameter Penunjang ... 25
4.2 Unsur Hara ... 30
4.3 Intensitas Cahaya Matahari ... 37
4.4 Fitoplankton ... 41
4.5 Klorofil-a ... 47
4.6 Produktivitas Primer Fitoplankton ... 50
4.5 Hubungan Cahaya dengan Produktivitas Primer ... 52
4.7 Hubungan Unsur Hara dengan Produktivitas Primer ... 56
V. SIMPULAN ... 62
DAFTAR PUSTAKA ... 63
DAFTAR TABEL
Halaman
1 Parameter utama dan penunjang yang diamati... 20 2 Kedalaman Secchi (Sechhi depth (Sd)) yang terukur pada ketiga
stasiun selama penelitian ... 27 3 Hasil perhitungan nilai koefisien peredupan (k’m-1) menurut
stasiun dan waktu pengamatan (Perhitungan mengacu kepada
Tillman et al. 2000) ... 39 4 Hasil analisis regresi linier sederhana, koefisien determinasi (R2)
dan koefisien korelasi Pearson (r) antara NPP (Y) dengan
DAFTAR GAMBAR
Halaman
1 Peta Lokasi Penelitian ... 19 2 Rataan nilai salinitas terdistribusi menurut stasiun dan
kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari
tiga pengamatan, KI adalah kedalaman Inkubasi) ... 25 3 Rataan nilai suhu terdistribusi menurut stasiun dan kedalaman
inkubasi (Bar menunjukkan Standard Deviasi dari tiga
pengamatan, KI adalah Kedalaman Inkubasi). ... 26 4 Rataan nilai kekeruhan terdistribusi menurut stasiun dan
kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari
tiga pengamatan, KI adalah Kedalaman Inkubasi) ... 28 5 Rataan TSS (Total Suspended Solid) atau total padatan terlarut
terdistribusi menurut stasiun dan kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari tiga pengamatan, KI adalah
Kedalaman Inkubasi) ... 29 6 Rataan ammonium-nitrogen (NH4-N) terdistribusi menurut
stasiun dan kedalaman inkubasi (Bar menunjukkan Standard
Deviasi dari tiga pengamatan, KI adalah Kedalaman Inkubasi) ... 30 7 Rataan nitrit-nitrogen (NO2-N) terdistribusi menurut stasiun dan
kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari
tiga pengamatan, KI adalah kedalaman Inkubasi) ... 31 8 Rataan nitrat-nitrogen (NO3-N) terdistribusi menurut stasiun dan
kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari
tiga pengamatan, KI adalah Kedalaman Inkubasi) ... 33 9 Rataan ortofosfat (PO4) terdistribusi menurut stasiun dan
kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari
tiga pengamatan, KI adalah Kedalaman Inkubasi) ... 34 10 Rataan silikat (SiO2) terdistribusi menurut stasiun dan
kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari
tiga pengamatan, KI adalah Kedalaman Inkubasi) ... 35 11 Distribusi intensitas cahaya matahari (Klux) di atas permukaan
air (udara) dan tepat di permukaan perairan... ... 38 12 Hasil perhitungan intensitas (Klux) dan persentase (%) cahaya
matahari pada kedalaman inkubasi serta pada dasar perairan
(Bar menunjukkan standar deviasi) ... 40 13 Jumlah genera fitoplankton menurut stasiun dan kedalaman
14 Rataan kelimpahan fitoplankton (sel l-1) menurut stasiun dan kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari
tiga pengamatan, KI adalah Kedalaman Inkubasi) . ... 45 15 Rataan nilai konsentrasi klorofil-a (mg m-3) terdistribusi
menurut stasiun dan kedalaman inkubasi (Bar menunjukkan SD
dari tiga pengamatan, KI adalah Kedalaman Inkubasi) ... 48 16 Nilai produktivitas primer bersih pada kedalaman inkubasi di
ketiga stasiun penelitian (Bar menunjukkan standar deviasi). ... 51 17 Pola hubungan antara produktivitas primer dengan intensitas
cahaya matahari pada ketiga stasiun ... 53 18 Pola hubungan antara produktivitas primer dengan kedalaman
perairan di ketiga stasiun yang diteliti ... 55 19 Plot data antara produktivitas primer dengan konsentrasi unsur
hara di stasiun 1 ... 58 20 Plot data antara produktivitas primer dengan konsentrasi unsur
hara di stasiun 2 ... 59 21 Plot data antara produktivitas primer dengan konsentrasi unsur
DAFTAR LAMPIRAN
Halaman
1 Berbagai nilai parameter penunjang dan utama yang diukur
selama penelitian ... 69 2 Intensitas cahaya matahari (lux) di permukaan (udara), tercatat
setiap 10 menit ... 71 3 Intensitas cahaya matahari (lux) di permukaan perairan, tercatat
setiap 10 menit ... 72 4 Intensitas cahaya matahari (Klux) pada berbagai kedalaman dan
waktu inkubasi selama tiga kali pengamatan di lokasi penelitian ... 73 5 Kelimpahan fitoplankton (sel l-1) pada periode I (12 Juli 2007) di
perairan Muara Jaya Teluk Jakarta ... 75 6 Kelimpahan fitoplankton (sel l-1) pada periode II (29 Juli 2007)
di perairan Muara Jaya Teluk Jakarta ... 77 7 Kelimpahan fitoplankton (sel l-1) pada periode III (12 Agustus
2007) di perairan Muara Jaya Teluk Jakarta ... 79 8 Nilai produktivitas primer bersih pada setiap lapisan kolom air
stasiun 1 selama waktu inkubasi ... 81 9 Nilai produktivitas primer bersih pada setiap lapisan kolom air
stasiun 2 selama waktu inkubasi ... 82 10 Nilai produktivitas primer bersih pada setiap lapisan kolom air
stasiun 3 selama waktu inkubasi ... 84 11 Hasil analisis ANOVA antar stasiun untuk masing-masing
kedalaman ... 87 12 Hasil analisis ANOVA antar kedalaman di masing-masing
1. Latar belakang
Peningkatan beban masukan limbah organik akan terkait dengan pengayaan nutrien di perairan pantai. Pengayaan nutrien yang berlebihan atau eutrofikasi dapat berpengaruh negatif terhadap ekosistem perairan. Pengaruh negatif tersebut di antaranya adalah menstimulasi produktivitas primer yang mendorong ke arah akumulasi biomassa fitoplankton, terjadinya ledakan alga termasuk yang beracun, deplesi oksigen terlarut saat proses pembusukan biomassa alga yang telah mati tersebut serta kematian ikan dan kerang-kerangan (Cloern 1996).
Masukan limbah organik juga berdampak terhadap ketersediaan cahaya di perairan. Limbah organik baik terlarut maupun tersuspensi akan meningkatkan kekeruhan perairan. Semakin tinggi kekeruhan maka penetrasi cahaya ke perairan yang lebih dalam menjadi terbatas. Hal ini menyebabkan berkurangnya zona eufotik, sehingga menurunkan lapisan produktif di perairan terkait dengan proses fotosintesis.
Untuk memahami dampak pengayaan nutrien secara lengkap terhadap ekosistem perairan pantai, maka pemahaman tentang ekologi fitoplankton merupakan salah satu kunci utama. Hecky dan Kilham (1988) menyatakan bahwa eutrofikasi adalah suatu proses ekologi, jadi harus dipelajari dari suatu perspektif ekosistem yang mempertimbangkan semua proses-proses interaksi fisika, kimia, trofik, dan sejarah hidup dari fluktuasi populasi fitoplankton. Menurut Nontji (1984) untuk memahami ekologi fitoplankton diperlukan pengetahuan tentang bagaimana kondisi biomassa, produktivitas, dan struktur komunitas fitoplankton serta kaitannya dengan kondisi lingkungan.
Perairan Muara Jaya Teluk Jakarta menerima beban limbah yang masuk melalui sungai, di antaranya melalui sungai Citarum dan Bekasi. Beban limbah tersebut berasal dari limbah permukiman, pertanian, perkotaan, dan industri yang berada di catchment area. Limbah tersebut kemudian berdampak terhadap pengayaan nutrien di perairan ini. Damar (2003) mencatat bahwa beban fosfat, DIN (Dissolved Inorganik Nitrogen) dan silikat yang masuk perairan Teluk Jakarta melalui sungai berturut-turut adalah 6.741 ton th-1, 21.260 ton th-1 dan 52.417 ton th-1 yang kemudian secara nyata meningkatkan konsentrasi nutrien di perairan ini. Kondisi bio-fisika-kimia perairan ini dipengaruhi pula oleh kondisi musim, terutama musim hujan dan musim kemarau. Karenanya, setiap musim akan dicirikan oleh sifat-sifat bio-fisika-kimia perairan yang berbeda-beda dan berdampak pada keberadaan, penyebaran, dan aktivitas fitoplankton baik horizontal maupun vertikal.
Meskipun penelitian serupa telah banyak dilakukan untuk perairan Teluk Jakarta, namun informasi tentang perairan di sekitar muara-muara sungai pada pantai Bekasi, wilayah perairan Teluk Jakarta sebalah Timur ini masih sedikit. Penelitian dengan tema yang sejenis baru dilakukan oleh Kaswadji et al. (1993) namun hanya terbatas pada musim peralihan Barat - Timur. Penelitian lain oleh Zudiana (1997) dan Abdunnur (1997) yang keduanya dilakukan pada perairan muara Jaya pantai Bekasi, lebih banyak mengungkapkan parameter fisik-kimia, namun tidak dalam konteks respon fitoplankton. Dengan demikian penelitian untuk mengkaji kaitan antara faktor lingkungan dengan respon fitoplankton pada musim Timur masih perlu dilakukan, baik dalam kerangka mengungkapkan informasi ekologinya maupun sebagai kegiatan monitoring, sehubungan dengan adanya proses eutrofikasi di perairan ini.
2. Perumusan masalah
Pada musim Barat atau musim hujan, curah hujan tinggi, sehingga limpasan air tawar yang masuk ke perairan pantai juga besar. Pada musim Timur atau musim kemarau curah hujan rendah, sehingga aliran air tawar yang masuk juga kecil. Besar kecilnya debit sungai mempengaruhi beban limbah organik yang masuk ke perairan pantai.
Akibat perubahan musim tersebut, pola penyebaran serta ketersediaan intensitas cahaya dan unsur hara sebagai dua faktor utama yang sangat mempengaruhi fitoplankton juga berubah, baik secara spasial maupun temporal. Di perairan Teluk Jakarta, Damar (2003), Syam (2002), dan Nontji (1984) menunjukkan bahwa, dengan meningkatnya salinitas atau semakin jauh dari garis pantai, nutrien, biomassa, dan produksi fitopankton menunjukkan penurunan, sedangkan intensitas cahaya meningkat. Dikatakan pula bahwa, konsentrasi nutrien, biomassa, dan produktivitas fitoplankton lebih tinggi selama musim hujan dibanding musim kemarau.
Di perairan Bekasi, Kaswadji et al. (1993) menunjukkan bahwa konsentrasi nutrien tertinggi terdapat di lokasi pertemuan antara air tawar dan laut, yang berjarak sekitar 500 m dari pantai pada musim peralihan Barat – Timur. Selanjutnya, kelimpahan, dan produktivitas primer fitoplankton lebih tinggi di lokasi sungai daripada lokasi yang jauh dari pantai, tetapi tidak untuk kandungan klorofil-a. Zudiana (1997) dan Abdunnur (1997) mencatat bahwa di sedimen muara Jaya Bekasi, kandungan bahan organik (C-organik) serta konsentrasi unsur hara N dan P, menunjukkan nilai yang lebih tinggi baik di sedimen maupun perairannya daripada yang ditemui di perairan yang jauh dari muara sungai. Lebih lanjut dikatakan bahwa, faktor yang menyebabkan tingginya nilai tersebut adalah masukan limbah dari daratan yang terbawa melalui aliran sungai.
Sehubungan dengan pengaruh musim Timur terhadap cahaya dan nutrien yang mengatur dinamika fitoplankton di perairan Muara Jaya Teluk Jakarta, maka muncul berbagai pertanyaan berikut:
2) Bagaimana gambaran karakter fitoplankton terutama tentang produktivitas primernya disamping kelimpahan, biomassa, dominansi maupun komposisi jenis.
3) Adakah hubungan nyata antara respon karakter fitoplankton tersebut dengan karakteristik perairan.
4) Jika terdapat hubungan nyata, faktor mana yang lebih dominan mempengaruhi dinamika respon fitoplankton tersebut.
Secara singkat, yang menjadi rumusan masalah pokok adalah, bagaimana hubungan antara produktivitas primer fitoplankton dengan cahaya dan unsur hara dalam skala spasial di perairan ini pada musim Timur.
3. Tujuan dan Manfaat
Secara umum tujuan penelitian yang dilakukan di perairan Bekasi pada musim Timur ini, adalah mengkaji hubungan antara respon fitoplankton dengan penyebaran unsur hara dan cahaya. Secara lebih terperinci penelitian ini bertujuan untuk mempelajari :
1. Distribusi konsentrasi unsur hara dan intensitas cahaya secara spasial. 2. Nilai produktivitas primer dan biomassa fitoplankton.
3. Faktor lingkungan (cahaya atau unsur hara) yang secara nyata mempengaruhi produktivitas primer fitoplankton.
Hasil penelitian ini diharapkan dapat memberikan informasi tentang respon fitoplankton sehubungan dengan konsentrasi unsur hara dan intesitas cahaya di perairan ini selama musim Timur. Informasi selama musim Timur ini bersama-sama dengan musim lainnya dapat dipakai sebagai salah satu dasar untuk menentukan produktivitas perairan Muara Jaya Teluk Jakarta serta untuk kepentingan pengelolaan perairan ini selanjutnya.
4. Hipotesis
1. Fitoplankton
Plankton merupakan organisme yang berukuran sangat renik yang hidup melayang-layang dalam air dan memiliki kemampuan gerak yang sangat lemah sehingga perpindahannya sangat dipengaruhi oleh pergerakan massa air. Plankton yang berukuran mikroskopis meliputi tumbuhan dan hewan. Golongan dari tumbuhan disebut fitoplankton dan dari hewan disebut zooplankton (Odum 1971; Sverdrup et al. 1972; Nybakken 1992; Parsons et al. 1984).
Pengelompokan plankton biasanya didasarkan pada ukuran (net dan non-net plankton), habitat (haliplankton dan limnoplankton) dan daur hidup (holoplankton dan meroplankton). Berdasarkan ukurannya, Levinton (1982) dan Nybakken (1992) mengelompokkan plankton atas ultraplankton (< 2 μm), nanoplankton (2-20 μm), mikroplankton (20-200 μm), makroplankton (0,2- 2 mm), dan megaplankton (>2 mm).
Kilham dan Hecky (1988) menyatakan bahwa fitoplankton lautan didominasi oleh sejumlah jenis Chrysophyta yaitu diatom, cocolithophore, dan silicoflagelata, serta Pyrrhophyta (dinoflagellata). Selanjutnya dinyatakan pula bahwa, beberapa kelompok lain dari fitoplankton ada yang kadang-kadang melimpah, tetapi mereka diwakili oleh jenis yang sangat sedikit. Ini meliputi Cyanophyta (cyanobacteria; sebagai contoh jenis-jenis dengan ukuran sel yang sangat kecil dari Synechococcus atau berkas-berkas besar dari filamen
Oscillatoria [Trichodesmium]). Anggota lain dari fitoplankton lautan adalah
Halosphaera (divisi Chlorophyta, class Prasinophyceae) yang memiliki sel-sel
hijau berbentuk bola.
menyumbangkan komposisi yang bervariasi pula terhadap struktur komunitas fitoplankton, serta kehadirannya dapat mencirikan kondisi lingkungan tertentu.
Sebaran fitoplankton di laut dipengaruhi oleh banyak faktor baik fisika, kimia, maupun biologi. Penelitian dan tulisan mengenai faktor-faktor yang mempengaruhi sebaran fitoplankton di laut telah banyak dilakukan oleh para ahli. Parsons et al. (1984) menjelaskan bahwa distribusi biogeografis plankton sangat ditentukan oleh faktor lingkungan, seperti cahaya, suhu, salinitas, nutrien, dan faktor-faktor lainnya. Faktor tersebut sangat menentukan keberadaan dan kesuksesan jenis plankton di suatu lingkungan perairan.
Cebrian dan Valiela (1999) menunjukkan pada ekosistem pantai tertutup dan terbuka di daerah temperate bagian utara, terdapat dua pola puncak biomassa fitoplankton, yaitu pada akhir musim dingin dan gugur di perairan ekosistem pantai tertutup, sedangkan pada perairan ekosistem pantai terbuka umumnya mencapai puncak dalam musim semi dan akhir musim panas. Interaksi antara kedalaman terbatas atau pencampuran pasang surut dengan variasi musiman cahaya di ekosistem pantai tertutup cenderung bertanggunjawab terhadap perbedaan tersebut. Selain itu ditunjang pula oleh tingginya konsentrasi nutrien di kolom air.
Penelitian yang dilakukan oleh Chang dan Gall (1998) di daerah
Subtropical Convergence (STC) pada tiga lokasi mendapatkan bahwa, komposisi
jenis dan standing stok/biomassa fitoplankton berbeda secara regional dan musiman (musim semi dan musim dingin). Pada kedua musim tersebut, diatom berukuran besar mendominasi pada lokasi frontal dari STC baik musim dingin maupun semi, dan terutama pada musim semi menyebabkan pengurangan
dissolved reactive silicate (DRSi) dan NO3-. Hal yang sama ditemukan pula pada
Laju pertumbuhan fitoplankton di perairan estuari maupun perairan pantai menunjukan respon yang berbeda-beda terhadap kondisi bio-fisika-kimia. Kondisi biogeokimia dimaksud antara lain berupa; flushing (Ferreira et al. 2005), toleransi salinitas (Ferreira et al. 2005; Caraco et al. 1987), cahaya, unsur hara, (Ferreira et al. 2005; Smith 1984; Hecky & Kilham 1988; Howarth 1988; Culvin-Aralar et al. 2004) maupun pemangsaan (Ferreira et al. 2005; Levinton 1982). Dengan begitu penyebaran fitoplankton di peraiaran estuari dan pantai, akan mengikuti distribusi dari kondisi bio-fisika-kimia tersebut.
Zat hara anorganik utama yang diperlukan fitoplankton untuk tumbuh dan berkembang biak ialah nitrogen dan fosfor. Disamping itu, silikat juga merupakan salah satu hara yang diperlukan dan mempunyai pengaruh terhadap proses pertumbuhan dan perkembangan organisme laut (Nybakken 1988). Kelimpahan komunitas fitoplankton di laut sangat berhubungan dengan kandungan nutrien seperti fosfat, nitrat, silikat, dan hara lainnya. Kandungan nutrien dapat mempengaruhi kelimpahan fitoplankton dan sebaliknya fitoplankton yang padat dapat menurunkan kandungan nutrien dalam air. Perubahan komposisi fitoplankton selanjutnya dapat mempengaruhi komposisi zooplankton dan komunitas plankton secara keseluruhan dalam suatu ekosistem (Prescott 1963).
Komunitas fitoplankton akan mengalami suatu suksesi dominasi jenis secara terus menerus. Faktor lingkungan yang dapat mempengaruhi dominasi dan suksesi jenis meliputi cahaya, konsentrasi dan rasio unsur hara, dan bentuk-bentuk kimia unsur hara (Goldman & Carpenter 1974). Peningkatan unsur hara yang terus menerus dapat mempengaruhi pertumbuhan dan struktur komunitas fitoplankton, bahkan pada estuari yang kaya nutrien. Karena jenis fitoplankton memperlihatkan perbedaan kebutuhan untuk berbagai nutrien, perubahan pada struktur komunitas terjadi karena perubahan fluks dan konsentrasi relatif unsur hara (Sanders at al. 1987).
2. Produktivitas Primer
Selain berperan dalam perikanan, produktivitas primer juga penting dalam kaitan dengan lingkungan. Produktivitas perairan dapat digunakan untuk menduga produksi ikan atau potensi sumberdaya perikanan dengan mengetahui faktor efisiensi ekologi dalam rantai makanan (Kaswadji et al. 1993).
Pada pendekatan tropo dinamik dalam ekosistem, diusahakan untuk diukur produksi pada setiap tingkat. Produksi adalah penyatuan materi organik baru ke dalam jaringan hidup, yang berarti pertambahan biomassa. Untuk tumbuhan dilakukan melalui proses fotosintesis, disebut juga produktivitas primer (Miller 2004; Siege 2004; Valiela 1995; Lederman 1988). Secara umum dianggap bahwa produktivitas primer di laut adalah fotosintesis dari senyawa-senyawa karbon organik oleh fitoplankton, walaupun sejumlah kecil produktivitas primer dapat dihasilkan oleh bakteri kemosintetis (Lederman 1988; Nybakken 1988). Unsur-unsur lingkungan yang penting untuk reaksi fotokimia ini adalah energi matahari, CO2, air, dan nutrien, sedangkan laju fotosintesis di laut biasanya dikontrol oleh
ketersediaan cahaya matahari dan nutrien.
Kirk (1994) mengemukakan bahwa proses fotosintesis dapat dibagi atas dua bagian, reaksi terang dan reaksi gelap. Secara ringkas dijelaskan bahwa, pada reaksi terang akan dibebaskan oksigen yang bersumber dari air serta dihasilkan energi bebas yang bersumber dari serangkaian perubahan ADP (Adenosine
diphosphate) dan fosfat inorganik menjadi ATP (Adenosine triphosphate).
Sedangkan pada reaksi gelap, akan dihasilkan karbohidrat yang direduksi dari karbondioksida dan menghasilkan sejumlah energi bebas, yang sumbernya berasal dari degradasi (decay) ATP yang telah dibentuk selama reaksi terang.
Energi yang terikat oleh fotosintesis tersedia untuk sintesis biomassa baru, namun pada saat yang sama beberapa dari energi itu digunakan untuk pengaturan
(maintanance) biomassa yang ada. Proses pengaturan ini disebut respirasi dan
secara efektif berlawanan dengan proses fotosintesis, respirasi pada akhirnya merubah energi kimia yang terikat menjadi energi radiasi yang dibaurkan sebagai panas.
primer bersih adalah yang tersedia untuk herbivora (Miller 2004; Siege 2004; Valiela 1995; Lederman 1988). Jadi besarnya energi dari biomassa tumbuhan adalah hasil dari keseimbangan antara fotosintesis dan respirasi, dan laju fotosintesis yang teramati adalah hasil bersih dari dua proses tersebut, sementara respirasi biasanya dianggap : (1) terkait dengan biomassa dan konstant sepanjang waktu percobaan, dan (2) sebanding dengan kehilangan energi yang terikat yang akan berlangsung pada reaksi gelap pada suhu yang sama (Kirk 1994).
Faktor-faktor yang membatasi produktivitas primer fitoplankton di perairan di antaranya adalah intensitas cahaya matahari, suhu, unsur hara, dan biomassa fitoplankton (Siege 2004; Valiela 1995; Parsons et al. 1984). Penyebaran produktivitas primer fitoplankton bervariasi secara luas. Variasi tersebut berkaitan dengan lintang geografis dan musim. Di daerah temperate pada musim dingin, cahaya seringkali membatasi nilai produktivitas primer (Holm-Hansen et al. 2004; Diaz at el. 2002; Malone et al. 1988; Gibss & Vant 1997), sedangkan di daerah tropis ketersediaan nutrien sering menjadi faktor pembatas produktivitas primer fitoplankton (Miller 2004; Burnes & Hughes 1999; Valilela 1995).
Selain musim fraksi ukuran dari fitoplankton juga memperlihatkan laju produktivitas primer dan biomassa fitoplankton yang berbeda (Vant & Safi, 1996). Pada daerah di sekitar Kutub Utara produktivitas dan biomassa fitoplankton juga dipengaruhi oleh konsentrasi Fe, sementara konsentrasi Fe dipengaruhi oleh pencampuran massa air yang berbeda (Holm-Hansen et al.
2004). Menurut Raymont (1980) ada suatu hubungan yang positif antara kelimpahan fitoplankton dengan produktivitas primer, yaitu jika kelimpahan fitoplankton di suatu perairan tinggi, maka perairan tersebut cederung mempunyai produktivitas primer yang tinggi pula.
cahaya nilai fotosintesis akan meningkat dengan cepat (secara proporsional) hingga mencapai jenuh atau mencapai titik plateu. Pada titik ini peningkatan cahaya tidak diikuti oleh peningkatan fotosintesis (nilai fotosintesis cenderung tetap) dengan kata lain intensitas cahaya menghambat fotosintesis (photoinhibiton). Di atas intensitas cahaya jenuh, peningkatan intensitas cahaya akan mengakibatkan penurunan dalam nilai fotosintesis (Barnes & Huges 1999; Kirk 1994; Valiela 1995; Miller 2004; Levinton 1982; Sigee 2004).
Distribusi fotosintesis dengan kedalaman perairan juga memperlihatkan fenomena yang umum. Pada permukaan perairan niliainya cenderung rendah, kemudian meningkat secara perlahan dengan bertambahnya kedalaman sampai mencapai maksimum, kemudian menurun lagi hingga mencapai nilai nol. Hal ini terjadi, berkaitan dengan penyebaran intensitas cahaya matahari di perairan secara vertikal.
Ada dua teknik standar untuk mengukur fotosintesis yaitu metode penyerapan 14C dan metode perubahan oksigen. Kedua metode mengukur secara langsung perubahan terus menerus dari substrat atau hasil fotosintesis dan masing-masing memiliki kekurangan dan kelebihan.
3. Cahaya
Menurut Kirk (1994), intensitas dan distribusi spektrum radiasi matahari yang diterima oleh bumi adalah fungsi dari karakteristik emisi dan jarak dari matahari. Flux radiasi matahari yang menimpa permukaan bumi adalah sekitar 1,775 x 1014W, dan total energi radiasi yang diterima bumi dari matahari setiap tahun adalah sekitar 5,53 x 1024 J. Selanjutnya dikatakan pula, meskipun kondisi langit jernih, intenitas seberkas cahaya matahari secara nyata berkurang selama melewati seluruh atmosfir. Pengurangan intensitas ini, sebagian karena scatering
atas atmosfer. Ketinggian cahaya matahari di atmosfer juga berpengaruh terhadap radiasi cahaya matahari yang mencapai bumi.
Menurut Levinton (1982), energi dari sumber cahaya matahari dapat disimbolkan dalam bentuk-bentuk unit energi, seperti Langley per menit (gcalm-2min-1). Sudut datang sinar matahari yang berbeda-beda dalam sehari,
lintang, dan faktor-faktor lainnya berperan terhadap distribusi spektrum cahaya yang menimpa permukaan laut dan sejumlah tertentu akan dibaurkan balik. Distribusi spektrum cahaya yang menimpa permukaan laut tersebut, meliputi suatu bagian besar dari spektrum infra merah hingga ultraviolet, tetapi hanya bagian spektrum cahaya tampak yang berpenetrasi ke bagian yang lebih dalam. Cahaya berkurang dalam kolom air melalui penyerapan dan pembauran. Pembauran dapat dilakukan oleh molekul-molekul air, materi organik terlarut, materi inorganik dan organik pertikulat, dan plankton hidup itu sendiri.
Cahaya matahari yang menembus permukaan laut akan mengalami dua perubahan penting. Pertama, energinya akan semakin berkurang secara eksponensial dan kedua, lebar spektrumnya semakin menyempit. Di perairan samudra, gelombang cahaya biru (475 nm) yang paling dalam dapat menembus laut, sedangkan di perairan pantai bergeser ke gelombang yang lebih panjang (hijau sampai kuning). Secara esensial semua serapan cahaya yang berlangsung di perairan alami disebabkan karena empat komponen dalam ekosistem perairan tersebut: air itu sendiri, pigmen-pigmen kuning (substansi humic) yang terlarut, biota fotosintesis, dan materi-materi partikulat yang mati (Kirk 1994).
Kelompok-kelompok fitoplankton akan berespon secara berbeda terhadap jumlah intensitas cahaya matahari yang tiba. Respon ini kemudian menghasilkan fitoplankton yang senang cahaya sun type dan yang kurang senang dengan cahaya
shade type. Tipe sun akan memiliki nilai fotosintesis yang tinggi pada intensitas cahaya yang juga tinggi. Yang tergolong tipe shade, akan beradaptasi dengan baik pada intensitas cahaya rendah, dan menghasilkan nilai fotosintesis yang tinggi pada intensitas cahaya rendah (Parsons et al. 1984).
4. Unsur hara
Unsur hara merupakan faktor penting dalam proses produksi fitoplankton. Unsur hara ini ada yang dibutuhkan dalam jumlah banyak, ada pula yang sedikit. Parsons et al. (1984) membagi nutrien yang dibutuhkan oleh tumbuhan termasuk fitoplankton menjadi dua bagian yaitu : (1) nutrien makro, dibutuhkan dalam jumlah yang banyak, dan (2) nutrien mikro, dibutuhkan dalam jumlah yang sedikit. Elemen-elemen yang termasuk nutrien makro antara lain C, H, N, P, Mg, dan Ca, sedangkan yang dibutuhkan dalam konsentrasi sangat kecil termasuk nutrien mikro antara lain Fe, Mn, Cu, Si, Zn, Na, Mo, Cl, V, dan Co
Menurut Hecky dan Kilham (1988) tiga unsur nutrien utama yang dibutuhkan fitoplankton adalah P (fosfat), N (nitrogen), dan Si (silikat). Kebutuhan akan nutrien sangat berbeda antara fitoplankton yang hidup di perairan tawar maupun perairan laut. Howarth (1988) menyatakan bahwa umumnya komposisi unsur-unsur C:N:P pada fitoplankton laut mengikuti raio Redfield yaitu 106:16:1, atau sedikit di bawah ratio tersebut.
Tumbuhan memanfaatkan nitrogen dalam bentuk senyawa anorganik. seperti NO3-N dan NH3-N. Nitrogen diserap oleh fitoplankton dalam bentuk nitrat
(NO3-N) dan ammonia (NH3-N). Fitoplankton lebih menyukai menyerap
berubah menjadi amonia dan saat kandungan oksigen tinggi nitrogen berubah menjadi nitrat.
Fosfor tersedia di laut dalam bentuk fosfat (PO43-) terikat dalam berbagai
molekul biologi seperti asam-asam amino, ADP, dan ATP (Miller 2004). Menurut Goldman dan Horne (1983) fitoplankton hanya dapat menggunakan fosfor dalam bentuk fosfat (PO4) untuk pertumbuhan. Parsons et al. (1984) menyatakan bahwa
fosfor di air laut berada dalam tiga bentuk utama yaitu fosfor anorganik terlarut, fosfor organik terlarut, dan fosfor partikulat. Fosfor merupakan salah satu unsur penting dalam pertumbuhan dan metabolisme tubuh diatom. Konsentrasi fosfor di perairan umumnya berkisar dari 0.001 sampai 0.005 ppm (Boyd 1981).
Silikon merupakan unsur kedua setelah oksigen yang cukup melimpah di alam (25,75%), yang terutama bersumber dari batuan granit. Batuan ini menyebar di sekitar teluk dan mencapai pula sungai. Secara alami, pencucian silikon batuan akan menjadi sumber penting silikon di laut, yang dibawah oleh aliran sungai ke dalam teluk, dan menyebabkan konsentrasi silikat meningkat di perairan pantai dan estuari (Yang et al. 2005). Ditambahakan bahwa, hanya diatom dan beberapa flagellata dari Chrysophyceae yang membutuhkan sejumlah besar silikon. Silikat diserap oleh fitoplankton dalam bentuk SiO2.nH2O. Kebanyakan dari senyawa ini
digunakan untuk membangun dinding sel fitoplankton dan sisanya untuk mengatur biosintesis dalam fitoplankton.
Asam silika adalah suatu unsur air laut penting untuk dinding sel diatom. Berkurangnya silika menghasilkan terhambatnya pembelahan sel dan dapat menekan aktivitas metabolisme sel. Di perairan alami, pengurangan silika dapat membatasi populasi fitoplankton dan dapat pula menyebabkan suksesi fitoplankton dari diatom ke fitoplankton yang tidak membutuhkan silika (Levinton 1982).
Respon komunitas fitoplankton terhadap peningkatan konsentrasi unsur hara di ekosistem perairan pantai memiliki hubungan penting dalam ekologi laut dan memiliki implikasi yang luas dalam bentuk kualitas air maupun perikanan. Siklus tahunan produktivitas fitoplankton diatur oleh PAR (Photosynthetically Available
Radiation), suhu, dan regenerasi nitrogen. Variasi antar tahunan dan variasi
terhadap variasi dalam masukan air tawar dan dengan variasi masukan nitrat (Malone et al.1988).
Umumnya unsur hara yang paling membatasi pertumbuhan fitoplankton di air tawar adalah fosfor dan di laut adalah nitrogen. Pada zona eufotik dari kebanyakan perairan, penyerapan fitoplankton mengurangi unsur-unsur tersebut ke konsentrasi yang sangat rendah. Rasio atom dari ketersediaan N:P dalam sistem air tawar secara umum lebih besar dari 15:1, rasio yang dibutuhkan untuk pertumbuhan fitoplankton. Di perairan laut, rasio ketersediaan N:P cenderung sama atau lebih rendah dari rasio Redfield 16:1. Rasio N:P yang lebih tinggi di perairan tawar mengindikasikan bahwa P merupakan unsur pembatas di perairan tawar. Di sisi lain, pada perairan pantai N biasanya merupakan nutrien pembatas. Hasil ini sesuai dengan rasio N:P yang relatif rendah yang dijumpai pada sistem perairan laut (Caraco et al. 1987).
Bukti yang mendukung nitrogen sebagai nutrien pembatas di perairan laut dikemukkan pula oleh Gibbs dan Vant (1997) bahwa di perairan laut terdapat kekurangan NH4 dan NO3, sedangkan PO4 terdapat dalam jumlah yang melimpah
dan tidak pernah berkurang sepanjang periode studi tersebut. Hasil ini didukung pula oleh percobaan pengayaan nutriennya, yang menunjukkan bahwa penambahan NH4 meningkatkan laju pertumbuhan fitoplankton hingga 15 kali
lipat, sedangkan penambahan PO4 memperlihatkan perbedaan yang kecil dengan
kontrol.
5. Suhu
Suhu air adalah salah satu sifat fisika perairan yang secara langsung dipengaruhi oleh adanya radiasi atau perambatan cahaya ke dalam perairan. Suhu air merupakan salah satu faktor penting bagi kehidupan organisme perairan. Di perairan suhu air dapat mempengaruhi produktivitas primer baik secara langsung maupun tidak langsung. Pengaruh suhu air secara langsung yaitu dengan meningkatnya suhu yang masih pada kisaran toleransi organisme nabati, akan meningkatkan laju metabolisme dan aktivitas fitoplankton yang ada di dalamnya.
Suhu air juga mempunyai pengaruh kuat terhadap perkembangan maupun penurunan populasi fitoplankton. Setiap jenis fitoplankton mempunyai suhu optimal untuk pertumbuhannya, tetapi dengan berbagai jenis yang bercampur di dalam perairan alami dengan suhu dari 0 sampai 30 °C terjadi kenaikan laju pertumbuhan pada seluruh komunitas yang sesuai dengan kenaikan suhu (Welch 1980).
Reaksi biokimia dalam sel fitoplankton umumnya dipengaruhi suhu. Peningkatan suhu terjadi secara eksponensial sampai pada batas maksimum. Peningkatan ini biasanya bervariasi untuk masing-masing reaksi, yaitu antara 25 sampai 40 °C. Kisaran suhu tersebut mempengaruhi laju fotosintesis maksimal untuk komunitas fitoplankton (Harper 1992).
Suhu dan salinitas mempengaruhi densitas air. Semakin dalam perairan, suhunya semakin rendah dan salinitas semakin meningkat, hingga rapat air juga meningkat yang selanjutnya membentuk stratifikasi yang kuat, dengan lapisan pegat (discontinuity) yang tajam yang akan sukar ditembus oleh fitoplankton (Raymont 1980).
6. Kekeruhan
Kekeruhan (turbiditas) adalah gambaran sifat optik air dari suatu perairan yang ditentukan berdasarkan banyaknya cahaya yang dipancarkan dan diserap oleh partikel-partikel yang ada dalam air tersebut (APHA 1989). Padatan terlarut dan tersuspensi mempengaruhi kekeruhan dan kekeruhan sangat berpengaruh terhadap proses fotosintesis.
Kekeruhan di perairan disebabkan oleh bahan organik tersuspensi, seperti liat, lempung, partikel karbonat, partikel organik halus, plankton, dan organisme renik lainnya. Bahan tersuspensi menyebabkan cahaya menjadi lebih tersebar dan diserap daripada ditransmisi. Ukuran dan karakteristik refraksi bahan partikel, secara optik penting untuk menunjang hubungan langsung dengan gravitasi
spesifik dan konsentrasi suspensi. Pengukuran parameter ini sangat berguna
untuk mengevaluasi stratifikasi secara mikro dari organisme-organisme antar lapisan perairan (Stewart et al. 1965, diacu dalam Wetzel dan Linkens 1979).
Perairan yang mempunyai kekeruhan yang tinggi akan mengurangi penetrasi cahaya ke dalam kolom air, sehingga membatasi proses fotosintesis. Produktivitas perairan dapat berkurang apabila dalam perairan terjadi kekeruhan tinggi yang disebabkan oleh partikel-partikel tersuspensi.
7. Salinitas
Salinitas merupakan salah satu parameter perairan yang berpengaruh pada fitoplankton. Variasi salinitas mempengaruhi laju fotosintesis, terutama di daerah estuari khususnya pada fitoplankton yang hanya bisa bertahan pada batas-batas salinitas yang kecil atau stenohaline (Kaswadji et al. 1993).
suhu menentukan densitas air, maka salinitas ikut pula mempengaruhi pengambangan atau penenggelaman fitoplankton.
1. Lokasi dan waktu penelitian
Penelitian ini berlokasi di perairan Muara Jaya Teluk Jakarta yang berada di wilayah administratif Kabupaten Bekasi (Gambar 1). Analisis laboratorium dilakukan di Laboratorium Produktivitas dan Lingkungan Perairan, Manajemen Sumberdaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor. Waktu pengukuran dan pengambilan contoh selama musim Timur dari bulan Juli sampai Agustus 2007. Pengukuran kualitas air dan pengambilan contoh air laut dilakukan tiga kali, yaitu tanggal 12 Juli, 29 Juli, dan 12 Agustus 2007.
2. Metode dan Desain Penelitian
Penentuan titik pengambilan contoh
Pada lokasi penelitian, dipilih tiga titik sebagai stasiun tempat dilakukan pengambilan contoh, yang ditentukan dari muara sungai ke arah laut. Stasiun 1 terletak tepat di muara sungai dengan posisi geografis pada 06o01’37,3’’ LS dan 106o59’32,2’’ BT, kedalaman rata-rata 1,3 m. Stasiun 2 dengan posisi geografis pada 06o01’44,4’’ LS dan 106o58’41.6’’ BT, berjarak 2.142 km dari stasiun 1, kedalaman rata-rata 5 m. Serta stasiun 3 dengan jarak 2.022 km dari stasiun 2, terletak pada posisi geografis 06o01’29,0’’ LS dan 106o57’36,1’’ BT, kedalaman 10 m. Pembagian stasiun sedemikian dimaksudkan agar dapat diperoleh gambaran tentang seberapa jauh pengaruh masukan dari daratan berperan di wilayah perairan ini. Dengan demikian, ketiga stasiun ini diharapkan akan mewakili pengaruh dari daratan, lepas pantai, dan kondisi peralihan dari keduanya.
Sumber : Damar 2003
Sumber : http://www.googlemaps.com Stasiun pengambilan sampel (Tgl 28 April 2007)
Gambar 1 Peta lokasi penelitian.
6000’ S
107o00’ E 106050’ E
106040’ E
St.1 St.2
Teknik pengukuran dan pengambilan contoh air laut
Pengukuran produktivitas primer dan kualitas air serta pengambilan contoh air dilakukan setiap dua minggu dengan ulangan sebanyak tiga kali. Waktu pengukuran dan pengambilan contoh air disesuaikan dengan waktu pengukuran produktivitas primer fitoplankton. Lama waktu inkubasi produktivitas primer adalah lima jam, dimulai dari jam 09:00–14:00 WIB.
Contoh air diambil dengan menggunakan botol Van Dorn kapasitas 5 liter dari setiap kedalaman inkubasi. Contoh air didistribusikan pada wadah yang telah disediakan yaitu masing-masing untuk analisis unsur hara (0,25 l), klorofil-a (1 l), kekeruhan (0,1 l), dan produktivitas primer (1 l), serta fitoplankton (25 l). Khusus untuk analisis fitoplankton, contoh air diambil sebanyak lima kali dari masing-masing kedalaman inkubasi. Selain produktivitas primer, semua contoh air yang lain diawetkan dan disimpan dalam cool box hingga dianalisis di laboratorium. Metode yang digunakan untuk mengukur masing-masing parameter dimaksud disajikan dalam Tabel 1.
Tabel 1 Parameter utama dan penunjang yang diamati
No Parameter Satuan Metode dan Alat Uji Tempat
A. Utama
1 Intensitas Cahaya Lux Luxmeter in-situ 2 Ortofosfat μM Asam molibdate, spektrofotometer Lab 3 Nitrat-Nitrogen μM Brusin sulfat, spektrofotometer Lab 4 Nitrit-Nitrogen μM Sulfanilamid, spektrofotometer Lab 5 Amonia-Nitrogen μM Phenate, spektrofotometer Lab 6 Silikat μM Molibdosilicate, spektrofotometer Lab 7 Klorofil-a mg m3 Aseton 90%, spektrofotometer Lab 8 Produktivitas Primer mgCm-3jam-1 Teknik Oksigen, titrasi In-situ 9 Fitoplankton Sel l-1 Pencacahan, mikroskop Lab
B. Penunjang
Unsur hara
Contoh air dimasukkan ke dalam botol polyetilene 250 ml, kemudian disimpan dalam pendingin sebelum dianalisis. Analisis unsur hara dimulai dengan menyaring 250 ml air tersebut melalui saringan nucleopore diameter 47 mm dan ukuran porositas 0,2 μm yang dibantu dengan memasang pompa vakum yang melewati suatu saringan gelas microfibre, guna mempercepat proses penyaringan.
Klorofil-a
Air laut diambil sebanyak 1 l, dimasukkan ke dalam botol polyetilene
berwarna hitam dan disimpan dalam pendingan hingga dianalisis di laboratorium. Contoh air lalu disaring melalui gelas microfibre filter (Whatman GF/C, diameter 47 mm porositas 1,2 μm) dengan bantuan vakum pump. Hasil penyaringan diekstrak dalam 10 ml etanol 90% kemudian disentrifuge, supernatan hasil
sentrifuge diukur absorbannya dengan spektrofotometer. Persamaan untuk
menghitung kandungan klorofil-a merujuk pada APHA (1998) yaitu:
Pengukuran intensitas cahaya dilakukan dengan Luxmeter tipe Lutron LX-101 (Digital Luxmeter Takemura Elektric Work. Ltd). Pengukuran intensitas cahaya dilakukan di atas permukaan perairan dengan interval waktu setiap 10 menit. Pengukuran dimulai dari jam 06:00 sampai 18:00 WIB. Dengan demikian akan diperoleh pola distribusi cahaya dalam sehari.
Untuk mengetahui nilai intensitas cahaya di masing-masing kedalaman inkubasi digunakan hukum Lambert-Beer dalam Valiela (1995), yaitu :
Iz = Ioe-kz
Keterangan :
Iz = Intensitas cahaya pada kedalaman z
Io = Intensitas cahaya pada permukaan perairan
k = Koefisien peredupan cahaya z = Kedalaman
Koefisien peredupan cahaya dalam kolom air dihitung dari pembacaan kedalaman keping Secchi (Sd (m)), dengan menggunakan hubungan empiris k = 0,191 + 1,242/Sd (r2 = 0,853) (Tillman et al. 2000).
Produktivitas primer
Produktivitas primer diukur dengan metode botol terang-gelap. Produktivitas primer yang diukur dalam penelitian ini adalah produktivitas primer bersih. Prinsip kerjanya adalah mengukur perubahan kandungan oksigen dalam botol terang pada waktu awal dan akhir inkubasi, yang berisi air contoh setelah diinkubasi pada kedalaman perairan yang telah ditentukan. Konsentrasi oksigen terlarut diukur dengan cara titrasi Winkler. Produktivitas primer bersih dengan nilai oksigen terlarut dari metode ini kemudian dikonversi ke dalam satuan mgC/m3/jam. Prosedur serta rumus untuk perhitungan produktivitas primer selengkapnya merujuk pada Umaly dan Cuvin (1988) sebagai berikut:
Keterangan :
NPP = Fotosintesis bersih (mgC/m3/jam)
O2BT = Oksigen terlarut Botol terang akhir inkubasi (mg/l)
O2BA = Oksigen terlarut Botol terang awal inkubasi (mg/l)
1000 = Konversi liter menjadi m3
PQ = Photosynthetic Quotient = 1,2.
t = Lama inkubasi (jam)
0,375 = Koefisien konversi oksigen menjadi karbon (12/32)
Fitoplankton
25 liter air laut diambil dengan botol Van Dorn kapasitas 5 liter dari setiap kedalaman inkubasi dan disaring pada plankton net ukuran mata 20 μm.
NPP= 0,375
(t) (PQ)
BA)(1000) 2
(O BT) 2 (O
x
Untuk memperoleh 25 liter contoh air tersebut, pengambilan air laut diulang sebanyak 5 kali dari setiap kedalaman inkubasi. Air yang telah disaring dimasukkan ke dalam botol contoh 100 ml kemudian diawetkan dengan larutan Lugol 1%, 1 ml lugol tiap 100 ml contoh air. Identifikasi jenis fitoplankton dilakukan dengan menggunakan literatur di antaranya dari Allen and Cupp (1998), Tomas (1997), Yamaji (1979), Smith (1977), dan Davis (1955). Kelimpahan sel fitoplankton dihitung dengan metode strip berdasarkan APHA (1998) sebagai berikut :
N = n x (1/Vd) x (Vt/Vcg) x (Oi/Op)
Keterangan :
N = Kelimpahan fitoplankton (sel/l) n = Jumlah sel yang tercacah (sel) Vd = Volume air contoh yang disaring (l)
Vt = Volume air contoh yang tersaring (ml)
Vcg = Volume air di bawah cover glass (Sedwick Rafter Cell)(ml)
Oi = Luas gelas penutup preparat (Sedwick Rafter Cell) (mm2)
Op = Luas strip yang teramati (mm2)
3. Teknik/Model Analisis Data
Analisis ragam (ANOVA)
Analisis statistika ini digunakan untuk mengetahui ada tidaknya perbedaan nyata penyebaran parameter-parameter yang diukur antar stasiun dan kedalaman. Apabila terdapat perbedaan maka dilakukan uji lanjut untuk mengetahui lokasi yang saling berbeda nyata.
Analisis korelasi dan regresi
Analisis ini digunakan untuk mengukur derajat hubungan serta besarnya peranan antara variabel bebas (cahaya dan unsur hara) dengan variabel tak bebas (produktivitas primer).
Produktivitas Primer dengan Cahaya
Y = a (1-e-bX) e-cX Keterangan : Y = Produktivitas primer
X = Cahaya a,b,c = Konstanta
Produktivitas Primer dengan Unsur Hara
Untuk mengetahui peranan masing-masing unsur hara terhadap produktivitas primer di setiap stasiun, dilakukan analisis regresi linier sederhana (Steel and Torrie 1989) sebagai berikut
Y = a + bX
Keterangan : Y = Produktivitas primer,
X= Peubah bebas masing-masing nutrien (NH4-N, NO2-N,
NO3-N, dan PO4)
a = Koefisien Regresi b = Interseps
Nilai koefisien korelasi Pearson digunakan untuk mengetahui keeratan dan arah hubungan antara variabel bebas dan tak bebas, yang nilainya berkisar antara 0 dan ± 1 (-1≤ r ≤1). Arah hubungan dapat berpola searah (positif), berkebalikan (negatif) atau tidak ada pola arah hubungan (r = 0).
1. Parameter Penunjang
Salinitas
Variasi salinitas yang terjadi pada stasiun 1-3 disajikan pada Gambar 2, dan Lampiran 1. Pengukuran salinitas yang dilakukan pada tiap stasiun menunjukkan adanya fluktuasi. Kisaran nilai salinitas yang teramati berkisar antara 20–34 promil. Salinitas antar kedalaman inkubasi di masing-masing stasiun dan salinitas antar stasiun pada masing-masing kedalaman inkubasi tidak menunjukkan perbedaan nyata (ANOVA, p>0,05). Salinitas rata-rata terendah terukur pada stasiun 1 yang terletak tepat di muara sungai, sedangkan nilai tertinggi tercatat pada stasiun 3 yang terletak paling jauh dari muara sungai. Hal ini mengindikasikan terjadinya pengenceran air laut bersalinitas tinggi di muara sungai oleh masukan air tawar dari daratan sehingga menurunkan salinitas di stasiun 1 tersebut. Sesuai dengan pendapat Sachlan (1982), nilai salinitas yang terukur ini berada dalam kisaran yang mendukung perkembangan fitoplankton perairan laut.
20 22 24 26 28 30 32 34 36
1 2 3
Stasiun
Sali
nit
as (
p
ro
mil)
KI 0,2 m KI 1 m KI 2 m KI 3 m KI 4 m
Pola penyebaran salinitas yang demikian juga sama seperti yang dilaporkan oleh Damar (2003), Zudiana (1997), Kaswadji et al. (1993), dan Nontji (1984), dengan perbedaan yang hanya terletak pada besarnya kisaran nilai yang diperoleh. Pola demikian mencerminkan adanya masukan air tawar dari daratan – terutama melalui sungai - (Damar 2003).
Suhu
Suhu perairan selama penelitian ditampilkan dalam Gambar 3 dan Lampiran 1. Fluktuasi suhu yang teramati selama penelitian tidak menunjukkan variasi yang besar, baik antar stasiun maupun antar kedalaman inkubasi. Berdasarkan analisis ragam antar stasiun di setiap kedalaman serta antar kedalaman pada masing-masing stasiun, tidak memperlihatkan adanya perbedaan nyata (ANOVA, p>0,05). Hal ini dimungkinkan karena kondisi cuaca selama pengamatan relatif sama. Nontji (1984) mendapatkan adanya korelasi yang nyata antara suhu air dengan kecepatan angin dan kelembaban di perairan ini. Namun demikian dari Gambar 3, terlihat bahwa semakin dalam perairan dan semakin ke arah laut suhu cenderung mengalami sedikit penurunan.
27 28 29 30
1 2 3
Stasiun
Su
hu
(
o C)
KI 0,2 m KI 1 m KI 2 m KI 3 m KI 4 m
Gambar 3 Rataan nilai suhu terdistribusi menurut stasiun dan kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari tiga pengamatan, KI adalah Kedalaman Inkubasi).
dibandingkan dengan hasil pengamatan peneliti lain di perairan ini dan wilayah Teluk Jakarta lainnya, yang berkisar 27,5–31 oC (Damar 2003, Zudiana 1997, Syam 2002, Kaswadji et al. 1993; Nontji 1984).
Kedalaman Secchi, Kekeruhan, dan Padatan Tersuspensi Total
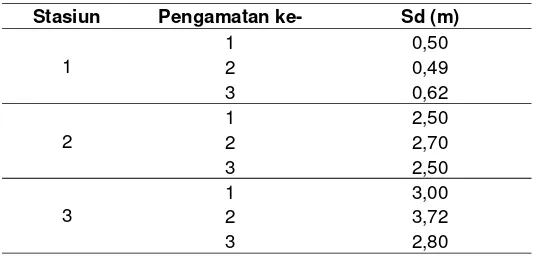
Kedalaman pembacaan cakram Secchi pada stasiun 1-3 selama penelitian ini memberikan hasil seperti tercantum dalam Tabel 2 . Kisaran nilai yang terukur berkisar dari 0,49–3,72 m. Kedalaman Secchi yang relatif tinggi ditemui pada stasiun yang letaknya cenderung jauh dari muara sungai. Nilai ini hampir sama dengan yang diperoleh Zudiana (1997) yang berkisar dari 0,1 m di mulut muara sampai 5 m pada stasiun terjauh dari muara.
Tabel 2 Kedalaman Secchi (Secchi depth (Sd)) yang terukur pada ketiga stasiun selama penelitian
Stasiun Pengamatan ke- Sd (m)
1
1 0,50 2 0,49 3 0,62
2
1 2,50
2 2,70 3 2,50
3
1 3,00 2 3,72 3 2,80
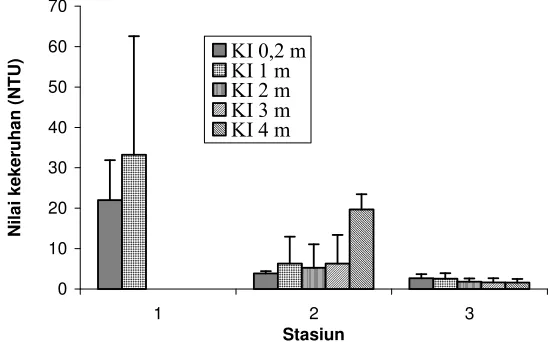
kekeruhan antar kedalaman tidak berbeda nyata (ANOVA, p<0,05), sedangkan pada stasiun 2 kekeruhan pada kedalaman 4 m berbeda nyata (ANOVA, p<0,05) dengan kedalaman lainnya. Kekeruhan menunjukkan hubungan terbalik dengan kedalaman Secchi (Tabel 2 dan Gambar 4). Hal ini berarti bahwa, semakin tinggi kekeruhan maka kedalaman Secchi akan berkurang. Dengan kata lain secara horizontal, semakin jauh dari muara sungai kedalaman Secchi meningkat sedangkan kekeruhan menurun.
0
Gambar 4 Rataan nilai kekeruhan terdistribusi menurut stasiun dan kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari tiga pengamatan, KI adalah Kedalaman Inkubasi).
tidak terdapat perbedaan nyata (ANOVA, p>0,05). TSS maksimum yang diperoleh ini sedikit lebih kecil dari hasil yang diperoleh Zudiana (1997) yakni 144-323 mg/l.
0 50 100 150 200 250
1 2 3
Stasiun
K
onse
n
tr
a
s
i TSS (mg/l)
KI 0,2 m KI 1 m KI 2 m KI 3 m KI 4 m
Gambar 5 Rataan TSS (Total Suspended Solid) atau total padatan terlarut terdistribusi menurut stasiun dan kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari tiga pengamatan, KI adalah Kedalaman Inkubasi).
2. Unsur Hara
Ammonium-N (NH4-N), Nitrit-N (NO3-N), dan Nitrat-N (NO3-N)
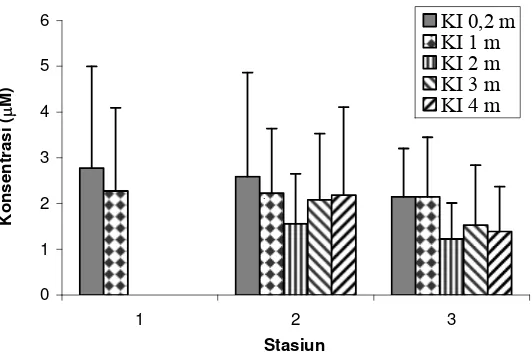
Kisaran konsentrasi ammonium hasil pengukuran selama periode pengamatan pada stasiun 1, 2, dan 3 berturut-turut adalah 0,241-4,753 μM, 0,306-4,951 μM, dan 0,192-3,444 μM (Lampiran 1). Hasil ini hampir sama dengan yang diperoleh Zudiana (1997) dan Kaswadji et al. (1993) yakni berturut-turut 0,335-4,170 μM dan 1.00-2,30 μM, namun lebih tinggi dari nilai yang diperoleh Kononen et al. (1996) di laut Baltik yaitu berkisar antara 0,14–0,39 μM. Pola distribusi vertikal ammonium untuk ketiga stasiun cenderung sama serta tidak berbeda nyata (ANOVA, p>0,05) (Gambar 6). Konsentrasi ammonium memperlihatkan nilai maksimum pada permukaan perairan kemudian menurun sampai mencapai nilai terendah di lapisan kedalaman 2 m dan meningkat lagi secara perlahan seiring dengan penambahan kedalaman perairan. Sebaran konsentrasi ammonium antar stasiun pada masing-masing kedalaman inkubasi juga memperlihatkan sedikit variasi. Konsentrasi ammonium memperlihatkan nilai yang menurun ke arah laut namun tidak berbeda nyata (ANOVA, p>0,05).
0 1 2 3 4 5 6
1 2 3
Stasiun
Kon
sentrasi (
μ
M)
KI 0,2 m KI 1 m KI 2 m KI 3 m KI 4 m
.
Gambar 6 Rataan ammonium-nitrogen (NH4-N) terdistribusi menurut stasiun
Diduga sumber utama unsur ini berasal dari daratan terutama dari limbah rumahtangga dan pertanian yang masuk melalui aliran sungai disamping sumber-sumber non-point source lainnya (Damar 2003). Lokasi yang dekat dengan muara sungai cenderung menerima ammonium dengan konsentrasi yang lebih tinggi dibandingkan dengan stasiun yang letaknya agak jauh dari muara sungai, sehingga memperlihatkan perbedaan konsentrasi ke arah laut (jauh dari muara). Meskipun terdapat variasi konsentrasi antar kedalaman dan lokasi, akan tetapi secara statistik tidak terdapat perbedaan nyata.
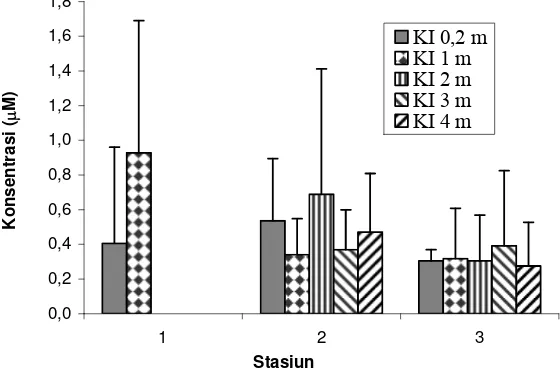
Kisaran hasil pengukuran nitrit selama periode pengamatan adalah 0,043-1,652 μM di stasiun 1, 0,130-1,522 μM di stasiun 2, dan 0,130-0,891 μM di stasiun 3 (Lampiran 1). Nilai ini hampir sama dengan hasil Zudiana (1997) yaitu 0,043-1,739 μM. Di antara ketiga bentuk nitrogen yang diukur, konsentrasi nitrit selalu ditemukan lebih rendah dari kedua bentuk nitrogen lainnya. Hal ini karena nitrit merupakan bentuk nitrogen yang tidak stabil, sehingga senyawa ini mudah mengalami perubahan menjadi ammonium atau nitrat.
0,0
Gambar 7 Rataan nitrit-nitrogen (NO2-N) terdistribusi menurut stasiun dan
kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari tiga pengamatan, KI adalah kedalaman Inkubasi).
konsentrasinya lebih kecil dari stasiun 2 serta pada kedalaman 3 m di stasiun 2 yang juga lebih kecil dari stasiun 3 (Gambar 7). Di stasiun 1 pada kedalaman 1 m, kadar nitrit dua kali lebih besar dari permukaan. Pada stasiun 2 dan 3, sebaran menegak cenderung memperlihatkan variasi nilai yang tidak besar, kecuali pada kedalaman 2 m di stasiun 2 yang konsentrasinya hampir dua kali lebih besar dari kedalaman yang lain, dan merupakan nilai maksimum di stasiun ini. Hasil analisis ragam menunjukkan tiadanya perbedaan nyata antar stasiun maupun kedalaman (ANOVA, p>0,05).
Konsentrasi nitrat di stasiun 1 berkisar antara 1,016-19,274 μM, stasiun 2 sebesar 0,742-7,916 μM, dan di stasiun 3 berkisar 0,081-9,219 μM (Lampiran 1), kisaran nilai maksimum yang diperoleh dalam penelitian ini lebih tinggi dari hasil yang diperoleh Zudiana (1997) yang berkisar 1,274-6,871 μM, masih berada dalam kisaran yang diperoleh Liu & Dagg (2003) di plume Sungai Missisipi yaitu antara 0,075-37,01 μM, namun lebih besar dari konsentrasi yang diperoleh Kononen et al. (1996) di laut Baltik sebesar 0,03-0,04 μM. Konsentrasi nitrat cenderung memperlihatkan nilai yang lebih besar pada kedalaman perairan yang lebih dalam (Gambar 8). Sampai dengan kedalaman 1 m, stasiun yang berada dekat muara memiliki konsentrasi yang lebih tinggi berturut-turut sampai stasiun yang terjauh, sedangkan dari kedalaman 2 sampai 4 m konsentrasi nitrat meningkat ke arah laut. Di masing-masing stasiun, variasi perbedaan penyebaran antar kedalaman tidak besar. Secara statistik tidak terdapat perbedaan nyata (ANOVA, p>0,05) penyebaran nitrat antar stasiun pada setiap kedalaman maupun antar kedalaman di masing-masing stasiun.
0
Gambar 8 Rataan nitrat-nitrogen (NO3-N) terdistribusi menurut stasiun dan
kedalaman inkubasi (Bar menunjukkan Standard Deviasi dari tiga pengamatan, KI adalah Kedalaman Inkubasi).
Ortofosfat (PO4-P)
Berdasarkan hasil pengukuran selama pengamatan, konsentrasi ortofosfat berkisar 0,021-1,295 μM, 0,053-1,191 μM, dan 0,032-1,243 μM berturut-turut pada stasiun 1, 2, dan 3 (Lampiran 1). Nilai-nilai tersebut berada dalam kisaran yang diperoleh Kaswadji et al, (1993) yaitu 0,045-0,265 μM, dan Zudiana (1997) berkisar 0,084-0,526 μM di lokasi yang sama, serta Liu & Dagg (2003) di plume
Sungai Missisipi berkisar 0,07-0,62 μM, namun lebih besar dari hasil yang dicatat Diaz et al. (2001) di perairan pantai Mediterania oligotrof yaitu kurang dari 0,04 μM. Sebaran ortofosfat baik secara horizontal maupun vertikal memperlihatkan pola yang bervariasi (Gambar 9). Tidak ada perbedaan nyata penyebaran fosfat antar stasiun dari masing-masing kedalaman mulai permukaan sampai 4 m, demikian pula antar kedalaman dalam stasiun yang sama tidak memperlihatkan perbedaan nyata (ANOVA, p>0,05).
Untuk pertumbuhan optimum fitoplankton, konsentrasi ortofosfat yang dibutuhkan berkisar 0,27-5,51 mg/L atau setara dengan 2,843-58,017 μM (Bruno
et al. dalam Widjaja et al. 1994).
0,0 0,2 0,4 0,6 0,8 1,0 1,2
1 2 3
Stasiun
Ko
nse
n
trasi
(
μ
M)
KI 0,2 m KI 1 m KI 2 m KI 3 m KI 4 m
Gambar 9 Rataan ortofosfat (PO4) terdistribusi menurut stasiun dan kedalaman
inkubasi (Bar menunjukkan Standard Deviasi dari tiga pengamatan, KI adalah Kedalaman Inkubasi).
Silikat (SiO2)
0 2 4 6 8 10
1 2 3
Stasiun
Ko
nsentrasi (
μ
M)
KI 0,2 m KI 1 m KI 2 m KI 3 m KI 4 m
Gambar 10 Rataan silikat (SiO2) terdistribusi menurut stasiun dan kedalaman
inkubasi (Bar menunjukkan Standard Deviasi dari tiga pengamatan, KI adalah Kedalaman Inkubasi).
Tingginya kandungan unsur hara di suatu perairan pantai antara lain disebabkan oleh masukan dari daratan melalui sungai atau sumber-sumber lain yang berada di sepanjang perairan pantai, di samping pula oleh adanya pengayaan dari lapisan lebih dalam karena penaikan massa air (upwelling) maupun pengadukan dasar laut (turbulensi atau mixing). Dikemukakan oleh Valiela (1995) bahwa masukan sungai dan air tawar lainnya sering merupakan sumber utama nutrien untuk beberapa perairan pantai. Ditambahkan oleh Alongi (1997) bahwa ketersediaan nutrien di estuari dan perairan pantai ditentukan di antaranya oleh proses fisika seperti laju flushing dan pencampuran atau pengenceran air, reaksi kesetimbangan geokimia, dan proses biologi karena aktivitas fitoplankton dan bakteri. Liu et al. (2003) memperkuat argumen tersebut dengan menyatakan bahwa transport air sungai adalah cara utama unsur hara terlarut dari darat ke laut.
penting terhadap beban masukan tersebut. Sesampai di perairan pantai, semua konsentrasi unsur hara memperlihatkan penurunan ke arah laut (Gambar 6-10), hal ini terjadi sebagai akibat dari pengenceran air sungai oleh air laut. Fenomena ini merupakan hal yang umumnya terjadi pada perairan pantai atau teluk yang memperoleh masukan air sungai, seperti yang teramati juga di Teluk Meksiko Utara (Liu & Dagg 2003), di Teluk Chesapeake (Malone et al. 1988), dan di Teluk Jiaozhou China (Yang et al. 2005).
Distribusi konsentrasi semua unsur hara secara vertikal pada masing-masing stasiun yang teramati dalam penelitian ini memperlihatkan variasi yang kecil. Dengan kata lain, rata-rata konsentrasi semua unsur hara menyebar hampir seragam dari permukaan ke arah dasar laut di setiap stasiun. Hal ini menggambarkan tidak terdapatnya stratifikasi unsur hara pada lapisan yang lebih dalam secara permanen. Keadaan ini terjadi akibat adanya pencampuran massa air secara vertikal sehingga mengaduk dan menyebarkan unsur hara antar lapisan dalam kolom air, di samping juga berkaitan dengan proses penyerapan unsur hara akibat aktivitas fotosintesis fitoplankton. Hasil penelitian oleh Seuront et al.
(2002) di perairan pantai yang teraduk oleh pasang surut menyimpulkan bahwa sebaran dan pengelompokan unsur hara dalam skala kecil dipengaruhi oleh interaksi kompleks antara kondisi-kondisi hidrodinamika, proses biologi yang terkait dengan populasi fitoplankton, dan efisiensi produksi populasi bakteri. Valiela (1995) menyatakan, meskipun di perairan pantai yang biasanya kaya nutrien, unsur hara dapat berkurang di zona eufotik karena penggunaan oleh fitoplankton akan mengurangi bagian terbesar dari ketersediaan nutrien.
Rasio molar DIN (penjumlahan semua spesies nitrogen) terhadap PO4
Kilham 1988; Smith 1984). Rasio ini sering digunakan untuk mengetahui unsur hara mana yang berpotensi membatasi pertumbuhan dan produktivitas fitoplankton (Damar 2003).
Menurut Howarth (1988) untuk berbagai ekosistem perairan, ada tiga faktor kontrol utama terhadap apakah nitrogen atau fosfor yang menjadi pembatas yang meliputi : a). Rasio N:P dalam input nutrien eksternal. b). Apakah N atau P yang lebih dulu hilang dari zona eufotik karena proses-proses biogeokimia seperti denitrifikasi, sedimentasi, dan adsorpsi, serta c). Tingkat dari berbagai kekurangan ketersediaan nitrogen dipenuhi melalui fiksasi nitrogen. Sebagai hasil dari ketiga proses tersebut, ekosistem estuari dan perairan pantai lebih dibatasi nitrogen dari pada di danau. Tetapi ada pula kejadian bahwa beberapa estuari dibatasi oleh fosfor atau bergantian secara musiman antara pembatasan fosfor dan pembatasan nitrogen.
3. Intensitas Cahaya Matahari
Intensitas cahaya matahari di udara bervariasi dari waktu ke waktu. Variasi nilai-nilai ini terjadi akibat adanya berbagai zat di udara yang menyerap maupun membaurkan seberkas cahaya yang melewatinya, letak lintang, posisi matahari di atas cakrawala, dan penutupan awan (Valiela 1995). Selain itu, perbedaan intensitas dalam sehari juga dipengaruhi atau mengikuti siklus pencahayaan harian. Pada pagi hingga tengah hari, intensitas akan bergerak meningkat sampai mencapai maksimum kemudian menurun lagi sampai terbenamnya matahari (Barnes & Huges 1982). Dalam siklus harian tersebut, nilai pencahayaan maksimum diperoleh pada tengah hari sedangkan minimum dijumpai pada pagi hari saat matahari baru terbit dan menjelang malam ketika matahari akan tenggelam.
sebesar 65,97 Klux terekam pada pukul 12,00 WIB. Besarnya intensitas minimum yang tercatat adalah 0,025 Klux (pengamatan I jam 18,00 WIB), 0,039 Klux (pengamatan II jam 06,00 WIB), dan 0,038 Klux (pengamatan III jam 18,00 WIB), sedangkan nilai rata-rata terkecil untuk ketiga waktu pengamatan tercatat sebesar 0,035 Klux pada jam 18,00 WIB.
Intensitas cahaya yang mencapai permukaan laut, juga menunjukkan pola yang sama dengan intensitas yang tiba di atas permukaan laut (Gambar 11). Perbedaannya hanya terletak pada besarnya nilai intensitas cahaya matahari yang telah berkurang sekitar 10% (Lampiran 3) atau dengan kata lain, hanya 90% dari seberkas cahaya yang tiba di atas permukaan laut akan sampai tepat di permukaan laut (Kirk 1994; Iwasaka et al. 2000). Intensitas yang tiba di permukaan laut ini akan berpenetrasi ke kedalaman yang lebih dalam. Penetrasi cahaya ini penting guna menyediakan cahaya di berbagai kedalaman, sehingga mendukung proses-proses fisika maupun kimia yang berlangsung di kolom air maupun organisme yang menghuni kolom air dimaksud.
0
Gambar 11 Distribusi intensitas cahaya matahari (Klux) di atas permukaan air (udara) dan tepat di permukaan perairan.
(Parsons et al. 1984; Kirk 1994). Besarnya bagian cahaya yang berkurang dengan bertambahnya kedalaman dapat didekati, dengan lebih dulu menghitung koefisien peredupan. Nilai koefisien ini mencerminkan integrasi kehilangan cahaya oleh faktor-faktor seperti, molekul air, substansi-substansi humus (pigmen-pigmen kuning) yang terlarut, biota fotosintesis, dan material pertikulat yang mati dalam perairan tersebut (Kirk 1994).
Koefisien peredupan selama penelitian ini berbanding lurus dengan tingkat kekeruhan tetapi berbanding terbalik dengan kedalaman Secchi. Nilainya berturut-turut menunjukkan bahwa stasiun yang dekat dengan muara sungai memiliki nilai koefisien peredupan yang terbesar dibandingkan dengan stasiun yang semakin jauh ke tengah laut (Tabel 3). Pola ini mendukung hasil yang dijumpai oleh Damar (2003) pada perairan Teluk Jakarta lainnya terutama di muara sungai Angke, Priok, dan Marunda. Ditambahkan pula bahwa, aliran air sungai yang membawa serta partikel organik dan inorganik terlarut dan tersuspensi serta pencampuran vertikal massa air memegang peranan penting dalam meningkatkan nilai koefisien peredupan di stasiun dekat muara.
Tabel 3 Hasil perhitungan nilai koefisien peredupan (k’m-1) menurut stasiun dan waktu pengamatan (Perhitungan mengacu kepada Tillman et al. 2000)
Stasiun Pengamatan ke- k'
1
1 2,68 2 2,73 3 2,19
2
1 0,69 2 0,65 3 0,69
3
1 0,61 2 0,52 3 0,63
bervariasi (Lampiran 4). Variasi intensitas tersebut memperlihatkan penurunan secara eksponensial dengan bertambahnya kedalaman (Gambar 12).
Stasiun 1
Gambar 12 Hasil perhitungan intensitas (Klux) dan persentase (%) cahaya matahari pada kedalaman inkubasi serta pada dasar perairan (Bar menunjukkan standar deviasi).